chapter 25 amino acids, peptides, and proteins copyright © the mcgraw-hill companies, inc....
TRANSCRIPT

Chapter 25Chapter 25Amino Acids, Peptides, Amino Acids, Peptides,
and Proteinsand Proteins
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

25.125.1Classification of Amino Classification of Amino
AcidsAcids

Fundamentals
Amino acids are compounds that contain both an —NH2 group and a —CO2H group, but these groups are actually present in each amino acid as —NH3
+ and —CO2– respectively.
Amino acids are classified as , , , etc. amino acids according the carbon that bears the amino nitrogen.

Examples: Amino Acids NH3
+
CO2–
An -amino acid that is anintermediate in the biosynthesis of ethylene.
+H3NCH2CH2CO2
– A -amino acid that is one ofthe structural units present incoenzyme A.
+H3NCH2CH2CH2CO2
– A -amino acid involved inthe transmission of nerveimpulses.

The 20 Key Amino Acids
More than 700 amino acids occur naturally, but 20 of them are especially important.
These 20 common amino acids are the ones used by organisms to synthesize proteins. All of them are -amino acids.
They differ from one another with respect to the group (side chain) attached to the carbon.
These 20 are listed in Table 25.1.

Table 25.1
The 20 amino acids obtained from hydrolysis of proteins vary according to R (the side chain).
The properties of the amino acids also vary as the structure of R varies.
Major differences among side chains: Size, shape and electronic characteristics.
C C
O
O–
R
H
H3N+

Table 25.1
General ways of classifying -amino acid side chains:
nonpolar side chainspolar uncharged side chainsacidic side chainsbasic side chainsaromatic side chainsside chains containing an OHside chains containing an S

Table 25.1
C C
O
O–
H
H
H3N+
Glycine is the simplest amino acid. It is the only one in the table that is achiral.
In all of the other amino acids in the table the carbon is a chiral center.
Glycine
(Gly or G)

C C
O
O–
CH3
H
H3N+
Table 25.1
Alanine
(Ala or A)
Alanine and valine have alkyl groups as side chains, which are nonpolar and hydrophobic.
C C
O
O–
CH(CH3)2
H
H3N+
Valine
(Val or V)

C C
O
O–
CH2CH(CH3)2
H
H3N+
Leucine
(Leu or L)
Leucine, and isoleucine also have alkyl groups as side chains, which are nonpolar and hydrophobic.
C C
O
O–
CH3CHCH2CH3
H
H3N+
Isoleucine
(Ile or I)
Table 25.1

C C
O
O–
CH3SCH2CH2
H
H3N+
Methionine
(Met or M)
The side chain in methionine is nonpolar, but the presence of sulfur makes it somewhat polarizable.
Table 25.1

Proline
C C
O
O–
CH2
H
H2N+
H2CCH2
(Pro or P)
Proline is the only amino acid that contains a secondary amine function. Its side chain is nonpolar and cyclic.
Table 25.1
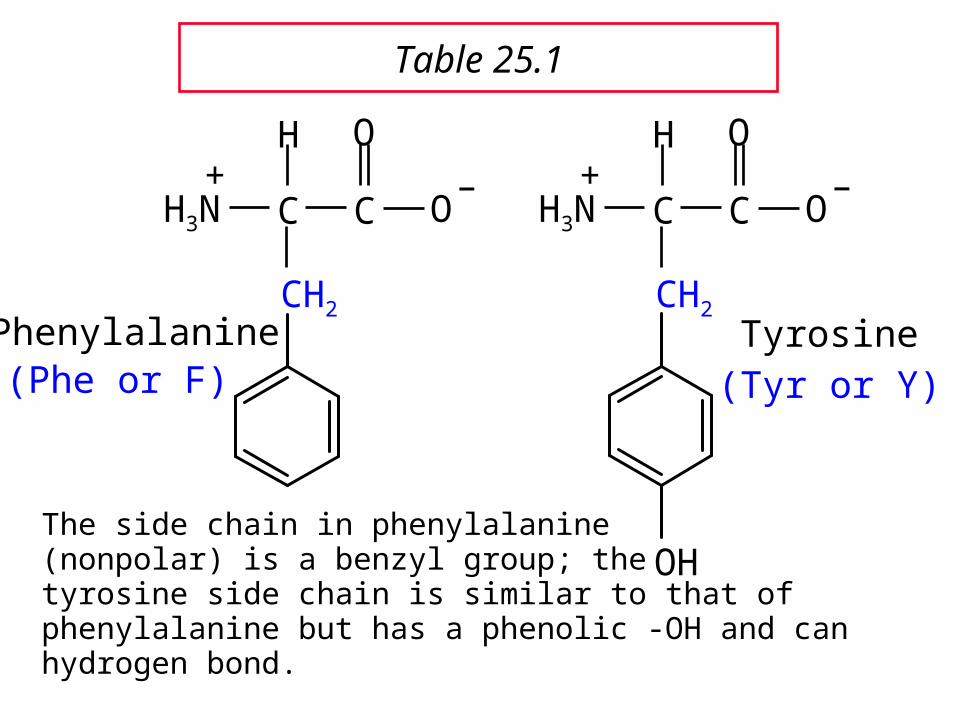
Phenylalanine
C C
O
O–
CH2
H
H3N+
(Phe or F)
The side chain in phenylalanine (nonpolar) is a benzyl group; thetyrosine side chain is similar to that of phenylalanine but has a phenolic -OH and can hydrogen bond.
C C
O
O–
CH2
H
H3N+
OH
Tyrosine(Tyr or Y)
Table 25.1

C C
O
O–
CH2
H
H3N+
N
H
Tryptophan
(Trp or W) The side chain in tryptophan (a nonpolar amino acid) is larger and more polarizable than the benzyl group of phenylalanine.
Table 25.1
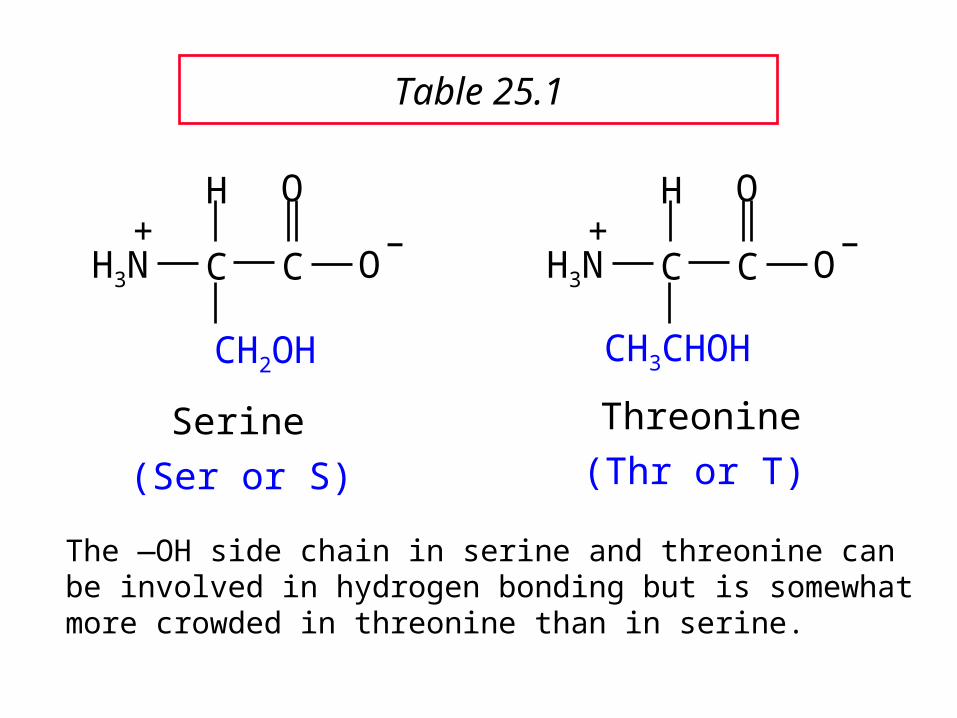
C C
O
O–
CH2OH
H
H3N+
Serine
(Ser or S)
The —OH side chain in serine and threonine can be involved in hydrogen bonding but is somewhat more crowded in threonine than in serine.
C C
O
O–
CH3CHOH
H
H3N+
Threonine
(Thr or T)
Table 25.1
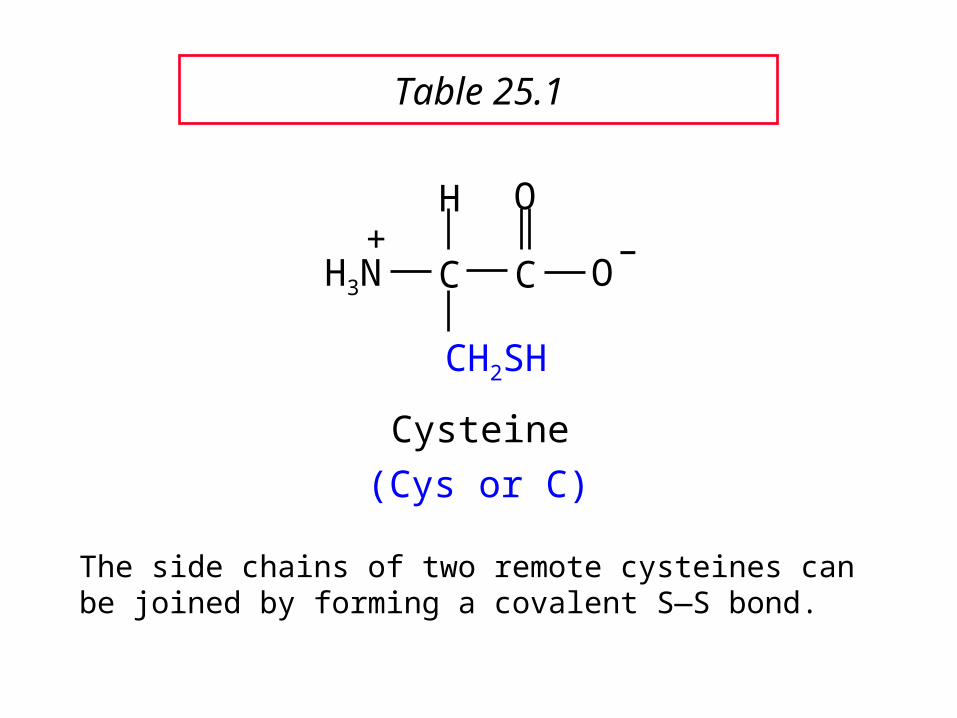
C C
O
O–
CH2SH
H
H3N+
Cysteine
(Cys or C)
The side chains of two remote cysteines can be joined by forming a covalent S—S bond.
Table 25.1
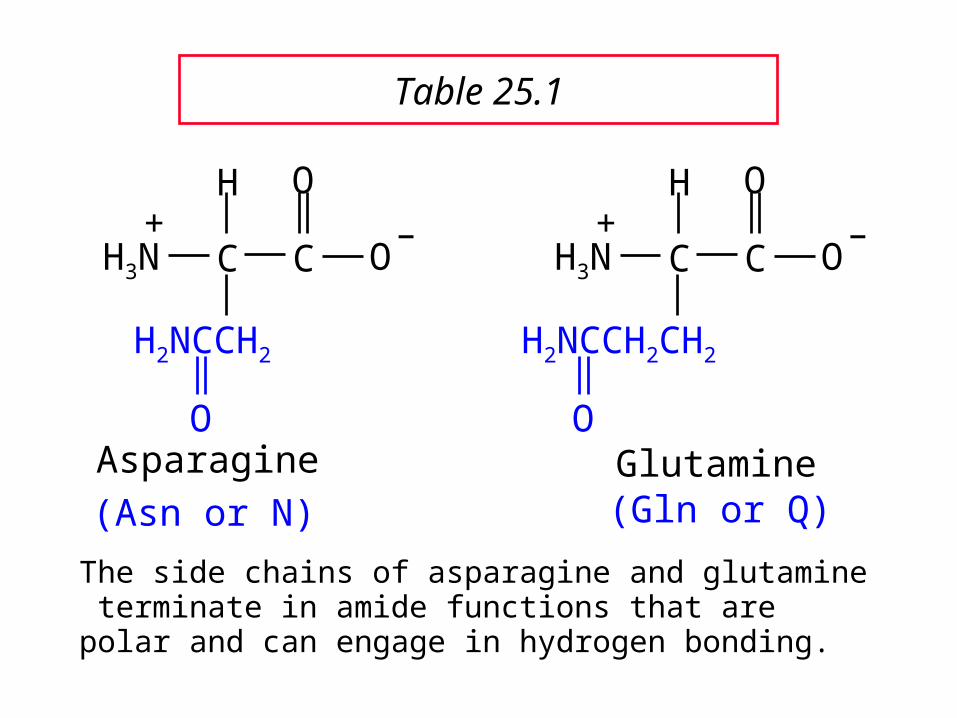
Asparagine
C C
O
O–
H
H3N+
H2NCCH2
O
(Asn or N)
The side chains of asparagine and glutamine terminate in amide functions that are polar and can engage in hydrogen bonding.
Glutamine
C C
O
O–
H
H3N+
H2NCCH2CH2
O
(Gln or Q)
Table 25.1

Aspartic Acid
C C
O
O–
H
H3N+
OCCH2
O
–
(Asp or D)
Aspartic acid and glutamic acid side chains exist as their conjugate bases at biological pH. They are negatively charged and can form ionic bonds with positively charged species.
C C
O
O–
H
H3N+
OCCH2CH2
O
–
Glutamic Acid
(Glu or E)
Table 25.1
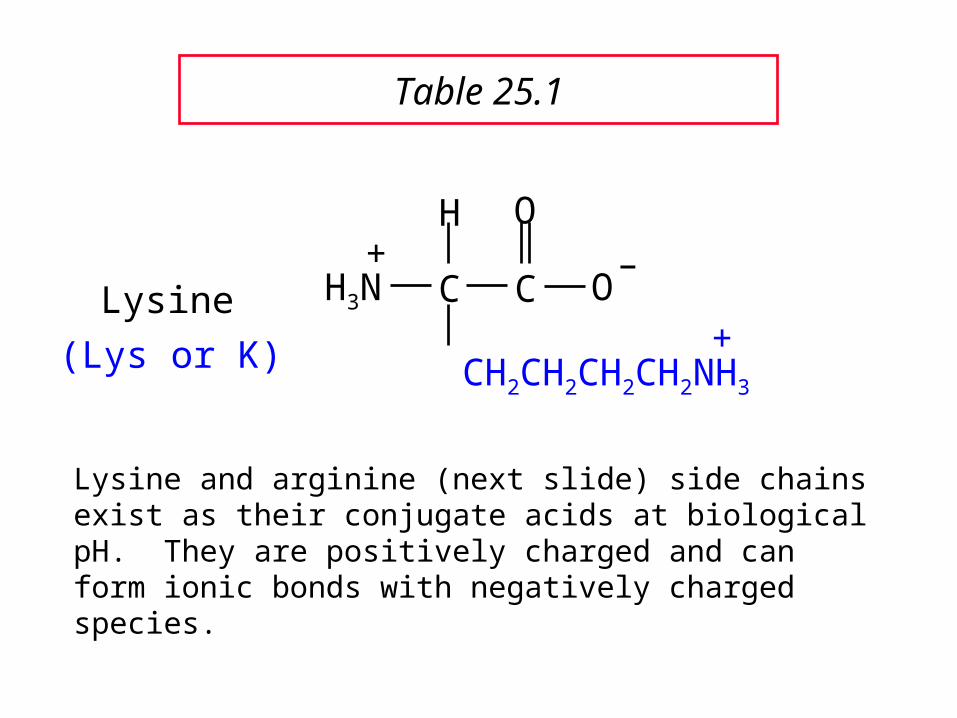
C C
O
O–
CH2CH2CH2CH2NH3
H
H3N+
Lysine+(Lys or K)
Lysine and arginine (next slide) side chains exist as their conjugate acids at biological pH. They are positively charged and can form ionic bonds with negatively charged species.
Table 25.1

C C
O
O–
CH2CH2CH2NHCNH2
H
H3N+
Arginine
+NH2
(Arg or R)
Table 25.1
Arginine has the most basic side chain of the 20 common amino acids (pKa ~12.5).

Histidine C C
O
O–
H
H3N+
CH2 NHN
(His or H)
Histidine is a basic amino acid, but less basic than lysine and arginine. Histidine can interact with metal ions and can help move protons from one site to another.
Table 25.1

25.225.2Stereochemistry of Amino Stereochemistry of Amino
AcidsAcids

Configuration of -Amino Acids
Glycine is achiral. All of the other amino acids in proteins have the L-configuration at the carbon.
H3N+
H
R
CO2–
The convnention for drawing Fischer projections of the amino acids is as shown above.

25.325.3Acid-Base Behavior of Acid-Base Behavior of
Amino AcidsAmino Acids

Recall
While their name implies that amino acids are compounds that contain an —NH2 group and a —CO2H group, these groups are actually present as —NH3
+ and —CO2– respectively.
How do we know this ?
Logic, of course, says that acids protonate bases.
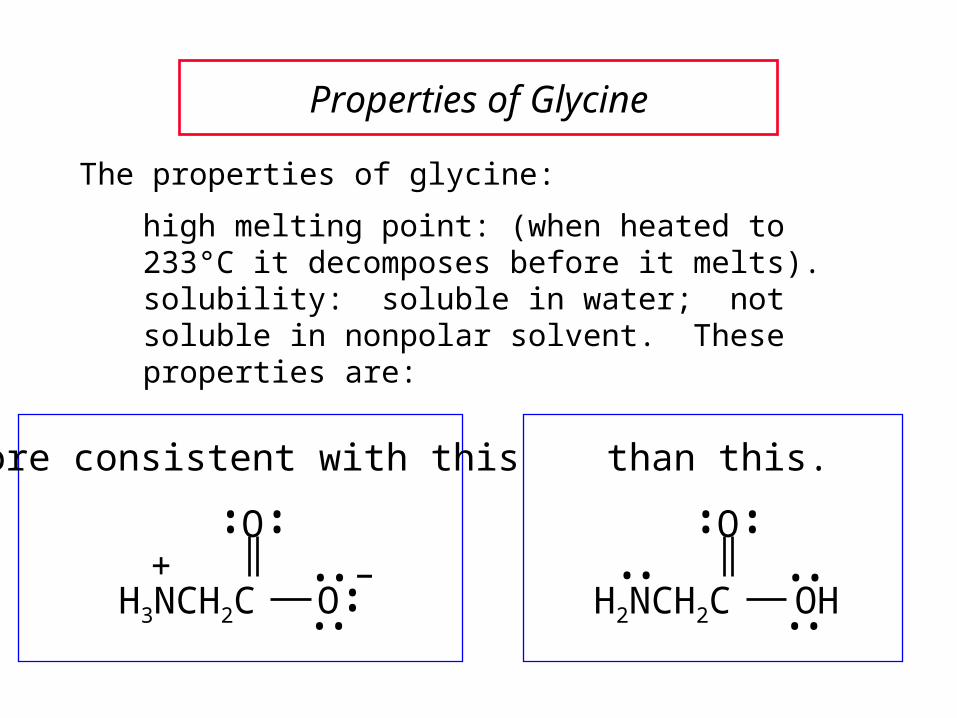
Properties of Glycine
The properties of glycine:
high melting point: (when heated to 233°C it decomposes before it melts).solubility: soluble in water; not soluble in nonpolar solvent. These properties are:
O
OHH2NCH2C••
••
••
•• ••
–••
O
OH3NCH2C ••
••
•• ••+
more consistent with this than this.

Properties of Glycine
This is true for all of the -amino acids.
–••
O
OH3NCH2C ••
••
•• ••+
This ionized form is called a zwitterion or dipolar ion

Acid-Base Properties of Glycine
The zwitterionic structure of glycine also follows from considering its acid-base properties.
A good way to think about this is to start with the structure of glycine in strongly acidic solution, say pH = 1.
At pH = 1, glycine exists in its protonated form (a monocation).
O
OHH3NCH2C+
••
••
•• ••

Now ask yourself "As the pH is raised, which is the first proton to be removed ? Is it the proton attached to the positively charged nitrogen, or is it the proton of the carboxyl group ?"
You can choose between them by estimating their respective pKas.
O
OHH3NCH2C+
••
••
•• ••
typical ammonium ion: pKa ~9
typical carboxylic acid: pKa ~5
Acid-Base Properties of Glycine
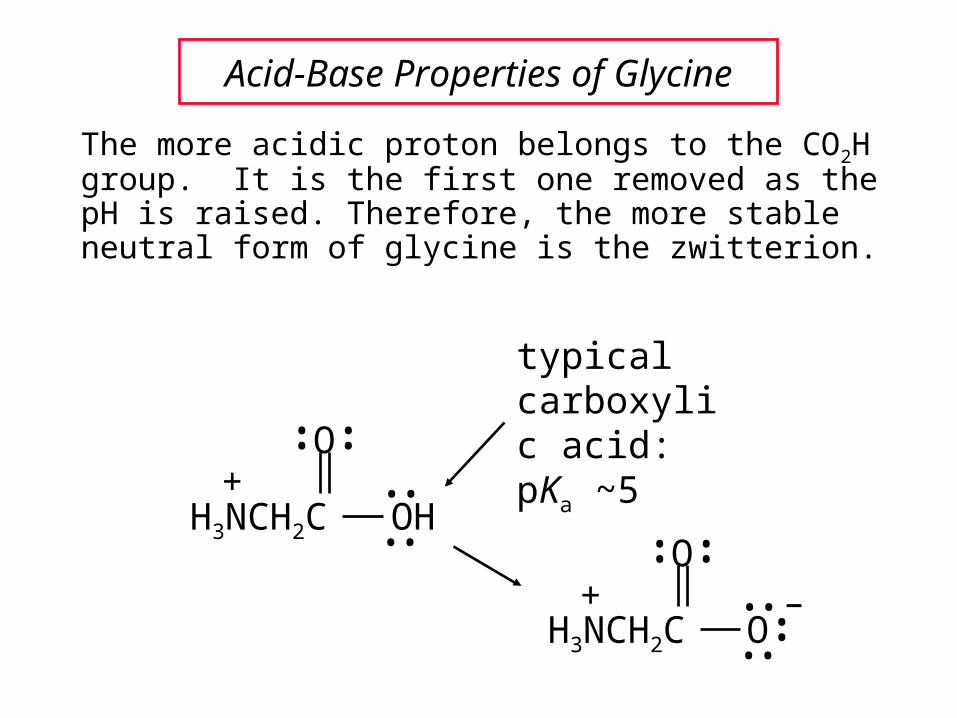
The more acidic proton belongs to the CO2H group. It is the first one removed as the pH is raised. Therefore, the more stable neutral form of glycine is the zwitterion.
typical carboxylic acid: pKa ~5O
OHH3NCH2C+
••
••
•• ••
Acid-Base Properties of Glycine
–••
O
OH3NCH2C ••
••
•• ••+

The measured pKa of glycine is 2.34.
Glycine is stronger than a typical carboxylic acid because the positively charged N acts as an electron-withdrawing, acid-strengthening substituent on the carbon.
typical carboxylic acid: pKa ~5
O
OHH3NCH2C+
••
••
•• ••Glycine pKa is actually 2.34.
Acid-Base Properties of Glycine

–••
O
OH3NCH2C ••
••
•• ••+
The pKa for removal of this proton is 9.60.This value is about the same as that for NH4
+ (9.3) but less than that of a primary amine.
HO ––••
O
OH2NCH2C ••
••
•• ••••
A proton attached to N in the zwitterionic form of nitrogen can be removed as the pH is increased further.
Acid-Base Properties of Glycine
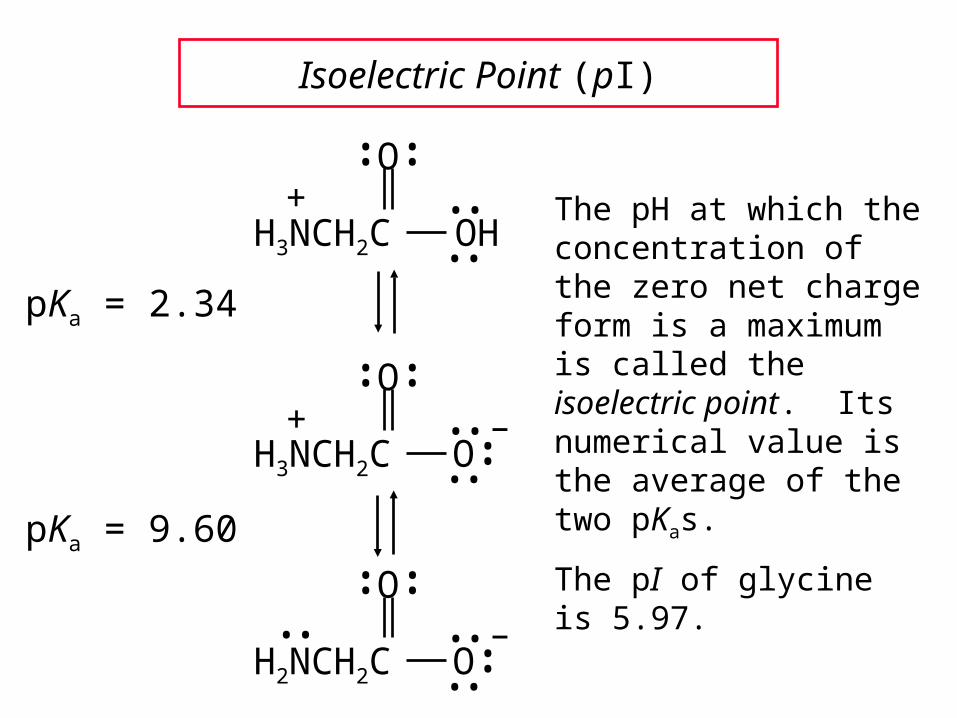
Isoelectric Point (pI)
pKa = 2.34
pKa = 9.60
The pH at which the concentration of the zero net charge form is a maximum is called the isoelectric point. Its numerical value is the average of the two pKas.
The pI of glycine is 5.97.
O
OHH3NCH2C+
••
••
•• ••
–••
O
OH3NCH2C ••
••
•• ••+
–••
O
OH2NCH2C ••
••
•• ••••

One way in which amino acids differ is in respect to their acid-base properties. This is the basis for certain experimental methods for separating and identifying them.
Just as important, the difference in acid-base properties among various side chains affects the properties of the proteins that contain them.
Table 25.2 gives pKa and pI values for amino acids with neutral side chains.
Acid-Base Properties of Amino Acids

Table 25.2 Amino Acids with Neutral Side Chains
C C
O
O–
H
H
H3N+
GlycinepKa1 = 2.34pKa2 = 9.60pI = 5.97

Table 25.2 Amino Acids with Neutral Side Chains
AlaninepKa1 = 2.34pKa2 = 9.69pI = 6.00
C C
O
O–
CH3
H
H3N+

Table 25.2 Amino Acids with Neutral Side Chains
ValinepKa1 = 2.32pKa2 = 9.62pI = 5.96
C C
O
O–
CH(CH3)2
H
H3N+
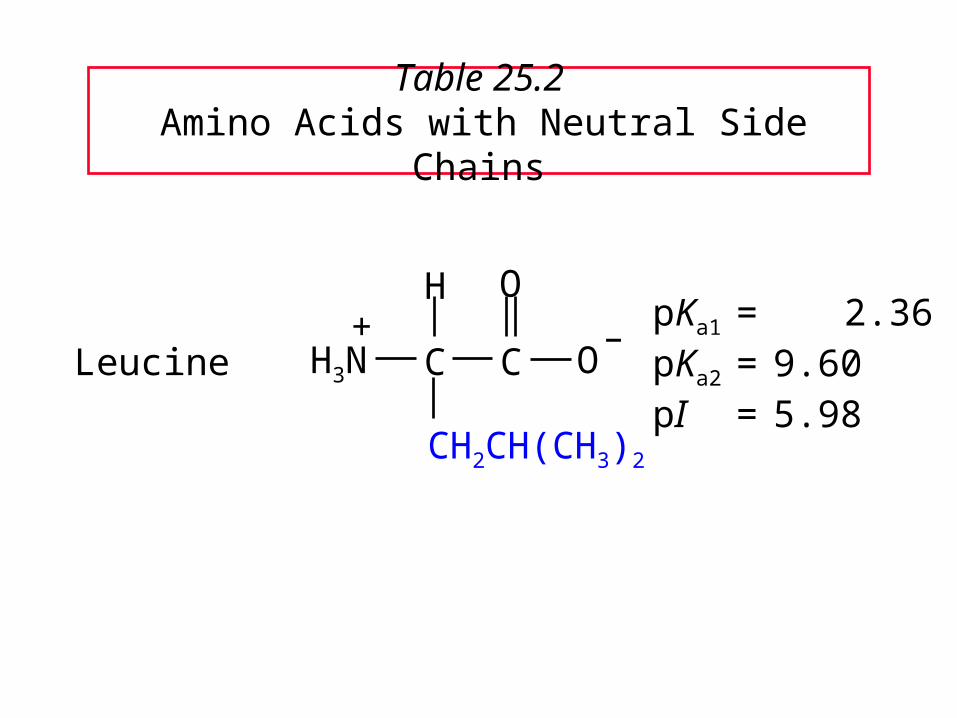
Table 25.2 Amino Acids with Neutral Side Chains
LeucinepKa1 = 2.36pKa2 = 9.60pI = 5.98
C C
O
O–
CH2CH(CH3)2
H
H3N+

Table 25.2 Amino Acids with Neutral Side Chains
IsoleucinepKa1 = 2.36pKa2 = 9.60pI = 6.02
C C
O
O–
CH3CHCH2CH3
H
H3N+

Table 25.2 Amino Acids with Neutral Side Chains
MethioninepKa1 = 2.28pKa2 = 9.21pI = 5.74
C C
O
O–
CH3SCH2CH2
H
H3N+

Table 25.2 Amino Acids with Neutral Side Chains
ProlinepKa1 = 1.99pKa2 = 10.60pI = 6.30
C C
O
O–
CH2
H
H2N+
H2CCH2
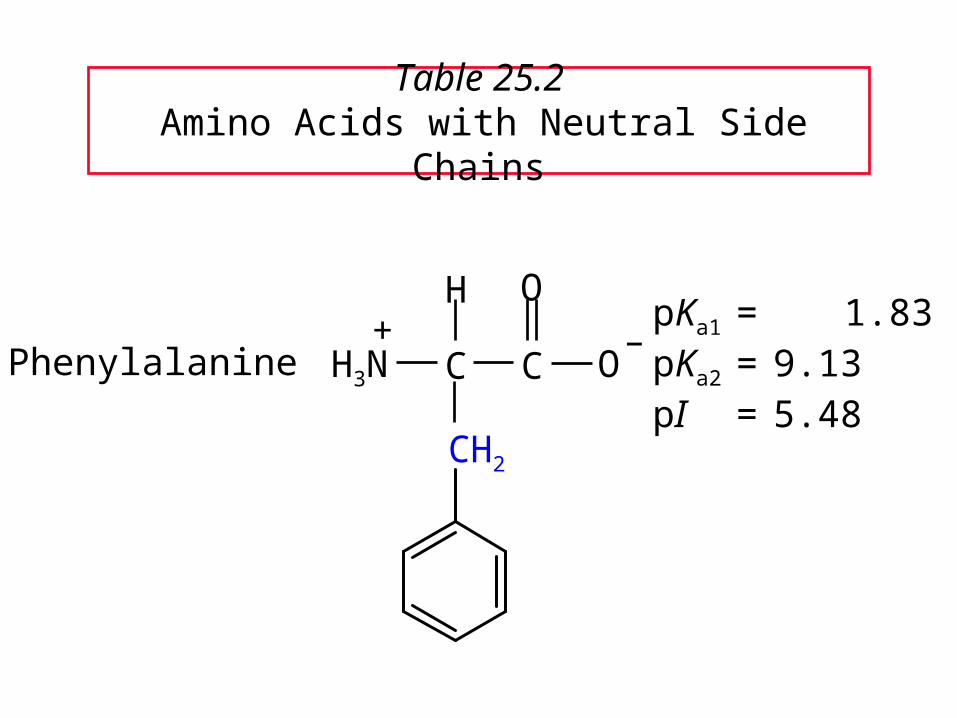
Table 25.2 Amino Acids with Neutral Side Chains
PhenylalaninepKa1 = 1.83pKa2 = 9.13pI = 5.48
C C
O
O–
CH2
H
H3N+

Table 25.2 Amino Acids with Neutral Side Chains
TryptophanpKa1 = 2.83pKa2 = 9.39pI = 5.89
C C
O
O–
CH2
H
H3N+
N
H

Table 25.2 Amino Acids with Neutral Side Chains
AsparaginepKa1 = 2.02pKa2 = 8.80pI = 5.41
C C
O
O–
H
H3N+
H2NCCH2
O
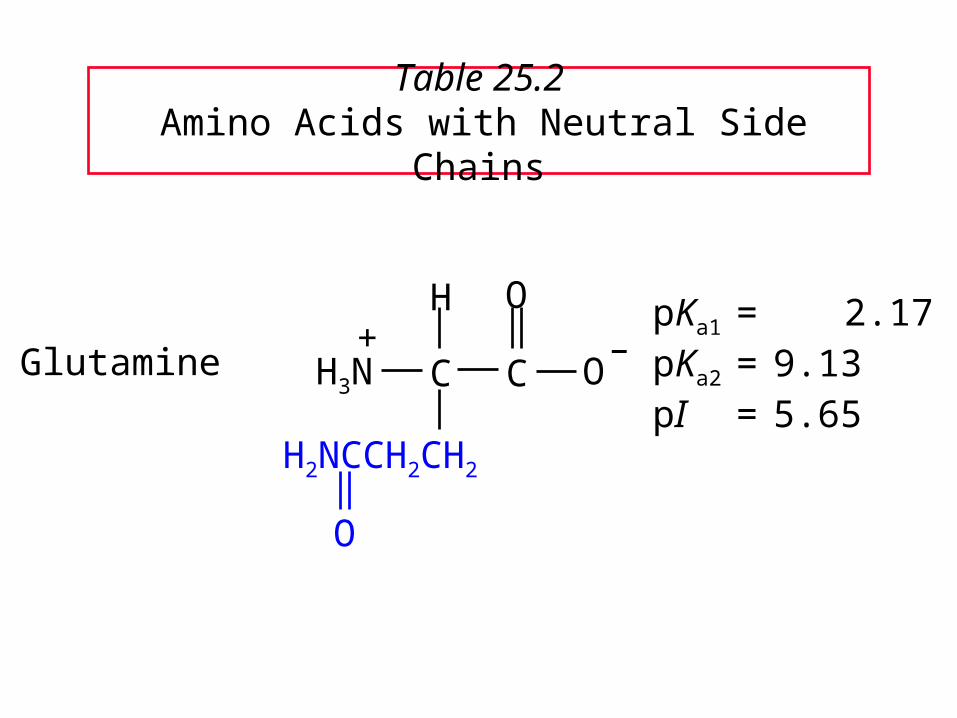
Table 25.2 Amino Acids with Neutral Side Chains
GlutaminepKa1 = 2.17pKa2 = 9.13pI = 5.65
C C
O
O–
H
H3N+
H2NCCH2CH2
O

Table 25.2 Amino Acids with Neutral Side Chains
SerinepKa1 = 2.21pKa2 = 9.15pI = 5.68
C C
O
O–
CH2OH
H
H3N+

Table 25.2 Amino Acids with Neutral Side Chains
ThreoninepKa1 = 2.09pKa2 = 9.10pI = 5.60
C C
O
O–
CH3CHOH
H
H3N+
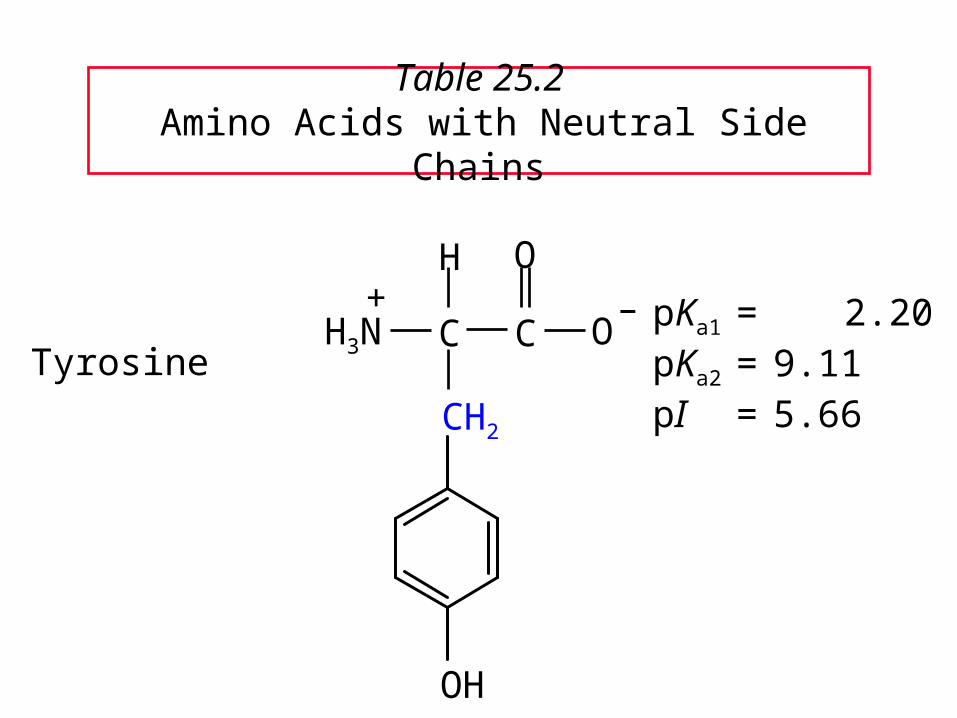
Table 25.2 Amino Acids with Neutral Side Chains
TyrosinepKa1 = 2.20pKa2 = 9.11pI = 5.66
C C
O
O–
CH2
H
H3N+
OH

Aspartic acidpKa1 = 1.88pKa2 = 9.60 pKa* = 3.65 pI = 2.77
For amino acids with acidic side chains, pI is the average of pKa1 and pKa*.
C C
O
O–
H
H3N+
OCCH2
O
–
Table 25.2 Amino Acids with Ionizable Side Chains

Glutamic acidpKa1 = 2.19pKa2 = 9.67 pKa* = 4.25pI = 3.22
C C
O
O–
H
H3N+
OCCH2CH2
O
–
Table 25.2 Amino Acids with Ionizable Side Chains

Lysine
pKa1 = 2.18pKa2 = 8.95 pKa* = 10.53pI = 9.74
For amino acids with basic side chains, pI is the average of pKa2 and pKa*.
C C
O
O–
CH2CH2CH2CH2NH3
H
H3N+
+
Table 25.2 Amino Acids with Ionizable Side Chains

Arginine
pKa1 = 2.17pKa2 = 9.04pKa* = 12.48pI = 10.76
C C
O
O–
CH2CH2CH2NHCNH2
H
H3N+
+NH2
Table 25.2 Amino Acids with Ionizable Side Chains

Histidine
pKa1 = 1.82pKa2 = 6.00pKa* = 9.17 pI = 7.59
C C
O
O–
H
H3N+
CH2 NHN
Table 25.2 Amino Acids with Ionizable Side Chains

25.425.4Synthesis of Amino AcidsSynthesis of Amino Acids

1. From -Halo Carboxylic Acids
CH3CHCOH
Br
O
2NH3+H2O
CH3CHCO
NH3
O
+
–
(65-70%)
+ NH4Br

2. The Strecker Synthesis
NH4Cl
NaCNCH3CH
O
CH3CHC
NH2
N
CH3CHCO
NH3
O
+
– (52-60%)
1. H2O, HCl, heat
2. HO–Hydrolysis of the nitrile
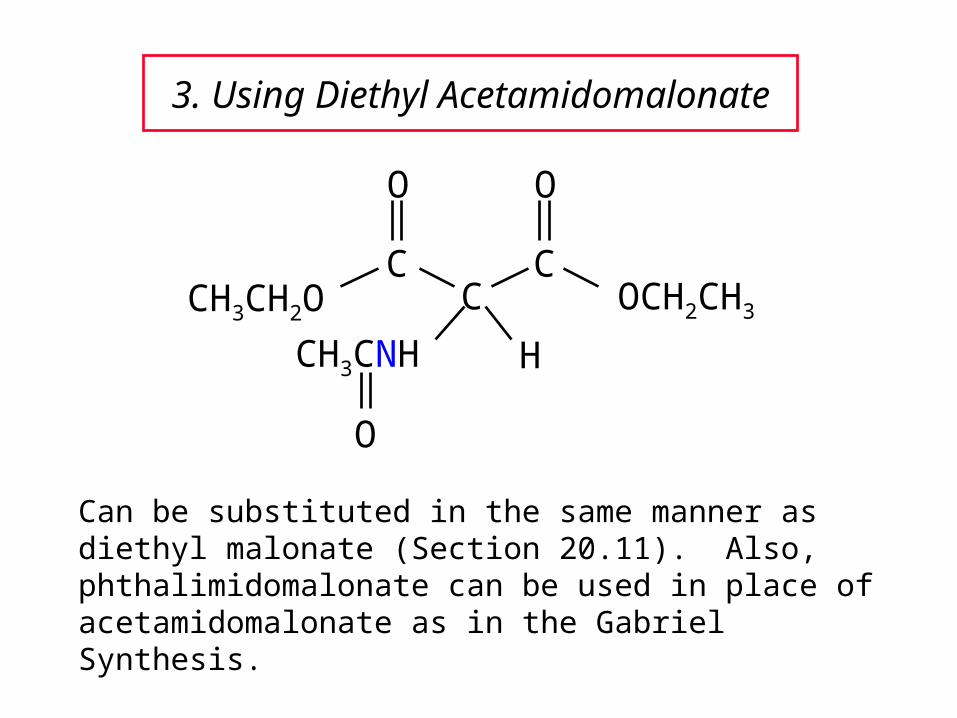
3. Using Diethyl Acetamidomalonate
CC
COCH2CH3
H
O O
CH3CH2O
CH3CNH
O
Can be substituted in the same manner as diethyl malonate (Section 20.11). Also, phthalimidomalonate can be used in place of acetamidomalonate as in the Gabriel Synthesis.

Example
1. NaOCH2CH3
2. C6H5CH2Cl
O O
CH3CH2OCCCOCH2CH3
HCH3CNH
O
O O
CH3CH2OCCCOCH2CH3
CH2C6H5CH3CNH
O
(90%)
Alkylation of malonate

Example
HBr, H2O, heatHydrolysis of ester and amide
O O
HOCCCOH
CH2C6H5H3N+
O
HCCOH
CH2C6H5H3N+(65%)
–CO2
O O
CH3CH2OCCCOCH2CH3
CH2C6H5CH3CNH
O
An amino acid

25.525.5Reactions of Amino AcidsReactions of Amino Acids
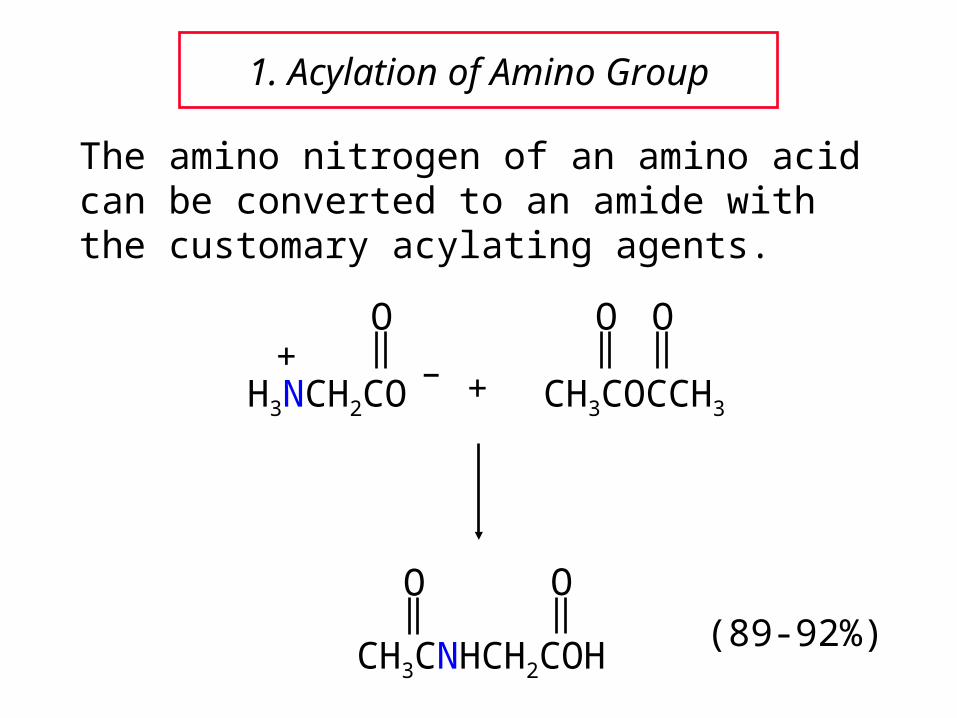
1. Acylation of Amino Group
The amino nitrogen of an amino acid can be converted to an amide with the customary acylating agents.
O
H3NCH2CO–+
+ CH3COCCH3
O O
CH3CNHCH2COH
O O(89-92%)
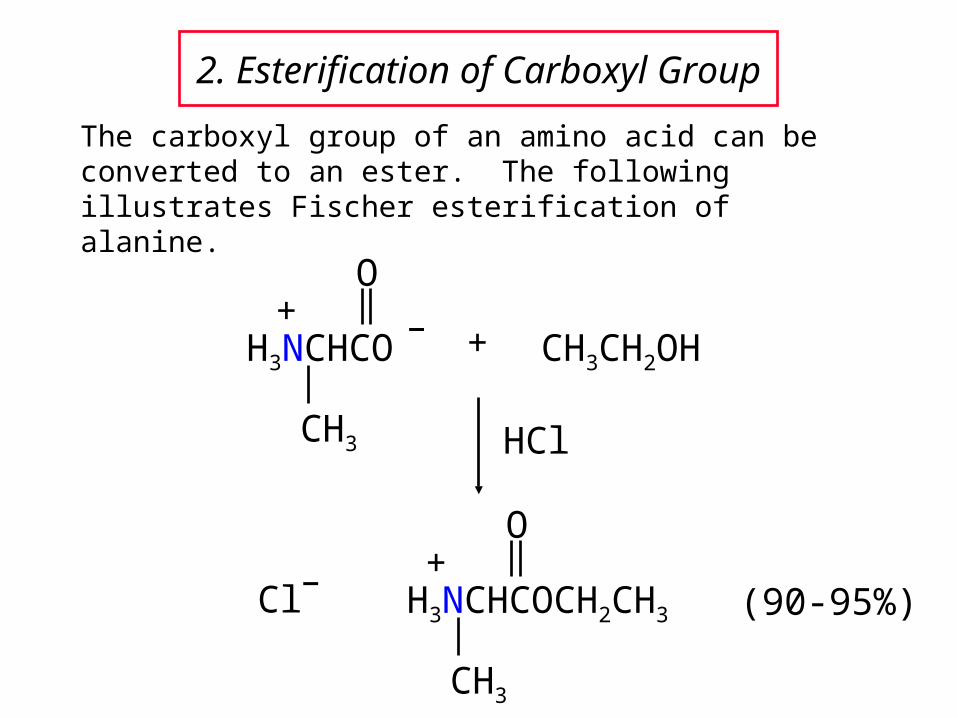
2. Esterification of Carboxyl Group
The carboxyl group of an amino acid can be converted to an ester. The following illustrates Fischer esterification of alanine.
+ CH3CH2OH
HCl
O
H3NCHCO–+
CH3
(90-95%)
O
H3NCHCOCH2CH3
+
CH3
–Cl

3. Ninhydrin Reaction
Amino acids are detected by the formation of Rhueman’s purple on treatment with ninhydrin.
OH
O
O
OH+
O
H3NCHCO–+
R
O O
O
N
O
–O
RCH + CO2 + H2O +

25.625.6Some Biochemical ReactionsSome Biochemical Reactions
of Amino Acidsof Amino Acids

Decarboxylation
–CO2, enzymes
+H3NCH2CH2CH2CO2
–+H3NCHCH2CH2CO2
–
CO2
–
–CO2, enzymes
Decarboxylation is a common reaction of -amino acids. Two examples are given below.
CH2CHCO2–
NH3+
NH
N
L-histidine
CH2CH2 NH2
NH
N
histamine
L-glutamic acid -aminobutyric acid (GABA)

Neurotransmitters
The chemistry of the brain and central nervous system is affected by neurotransmitters.
Several important neurotransmitters are biosynthesized from L-tyrosine.
OH
CO2–
HHH
H3N+
L-Tyrosine
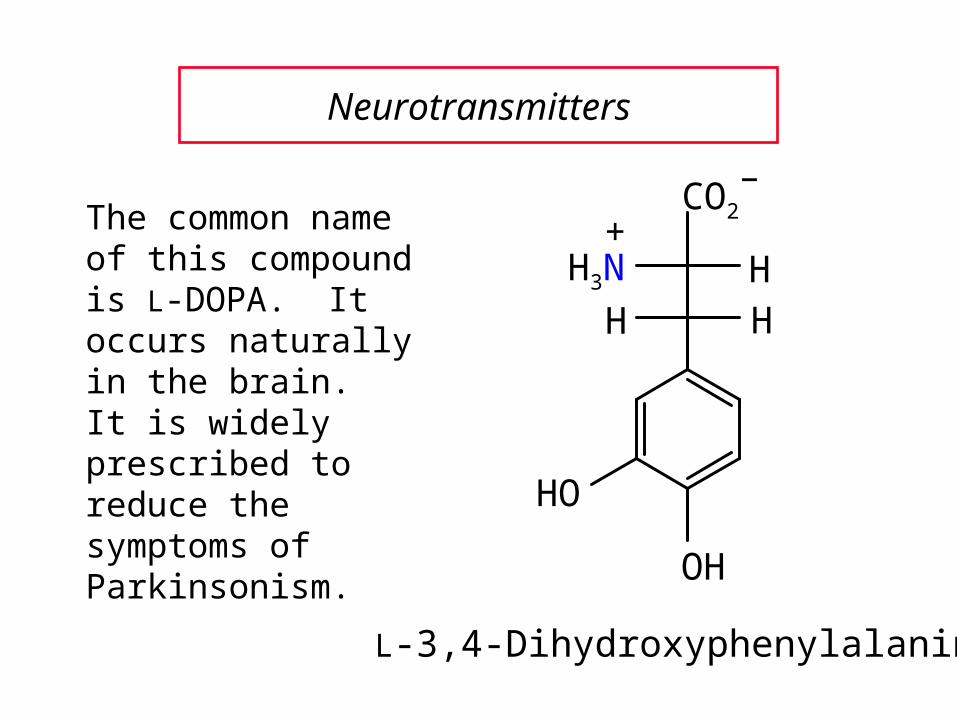
H3N
Neurotransmitters
The common name of this compound is L-DOPA. It occurs naturally in the brain. It is widely prescribed to reduce the symptoms of Parkinsonism.
OH
CO2–
HHH
+
L-3,4-Dihydroxyphenylalanine
HO

Neurotransmitters
Dopamine is formed by decarboxylation of L-DOPA.
OH
H
HHH
H2N
HO
Dopamine

Neurotransmitters
OH
H
HHOH
CH3NH
HO
Epinephrine
OH
H
HHOH
H2N
HO
Norepinephrine

25.725.7PeptidesPeptides

Peptides
Peptides are compounds in which an amide bond links the amino group of one -amino acid and the carboxyl group of another.
An amide bond of this type is often referred to as a peptide bond.
The amino acids in a peptide are called residues. Since a water was removed to make the peptide bond each residue does not represent a complete amino acid.
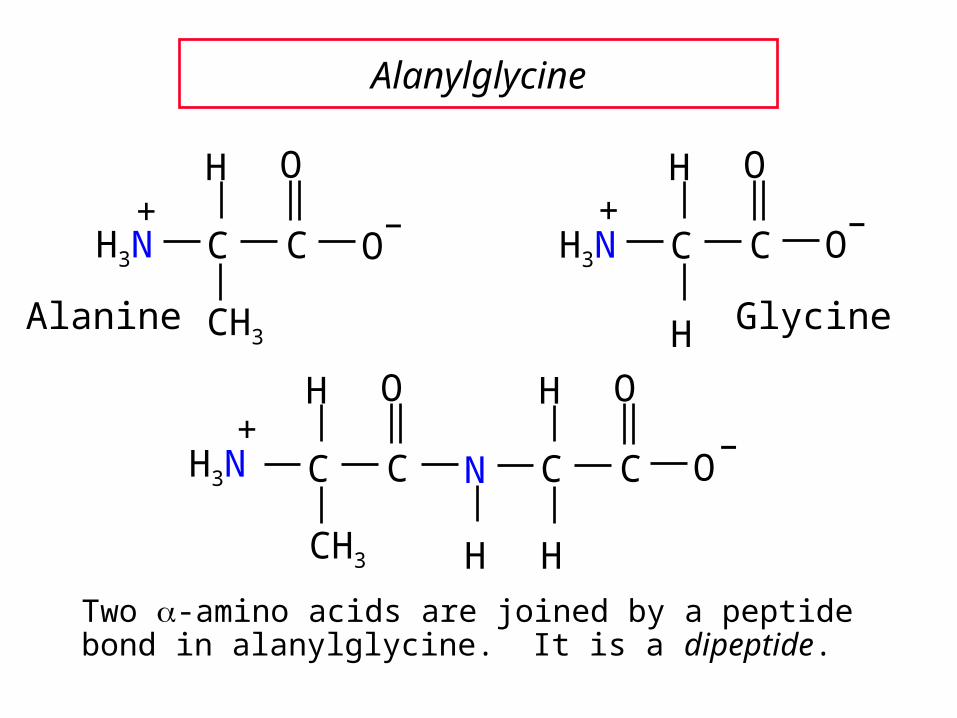
Alanylglycine
CH3
O
CH3N+
H
C
O
CN
H
H
C O–
H
Two -amino acids are joined by a peptide bond in alanylglycine. It is a dipeptide.
CH3
O
C+
H
C O–
H3N
O
C
H
H
CH3N+
O–
GlycineAlanine
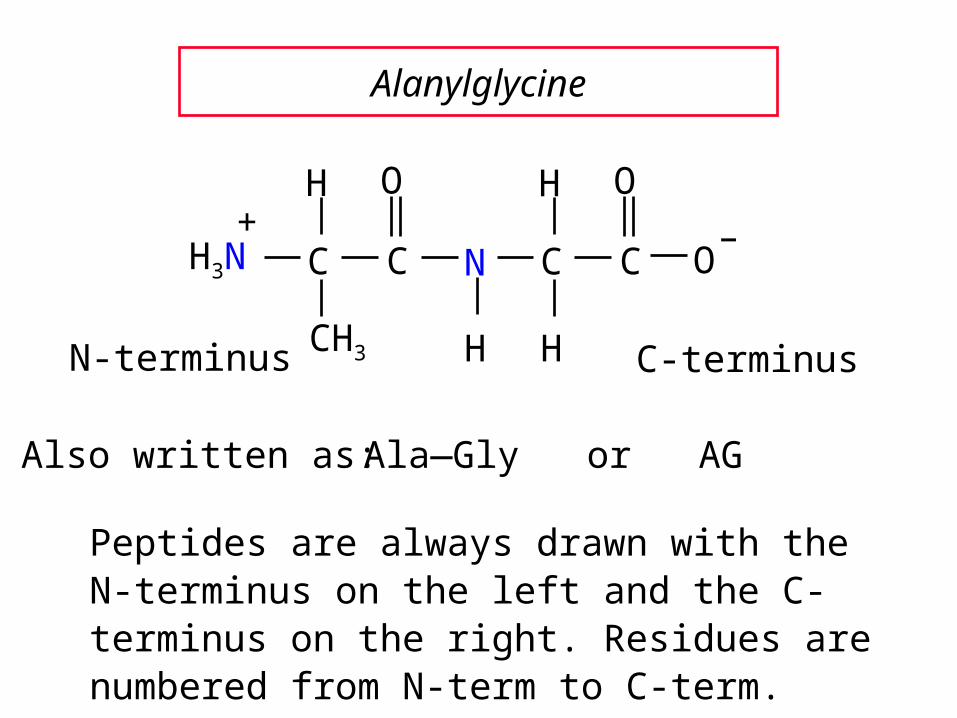
Alanylglycine
Ala—Gly or AG
N-terminus C-terminusCH3
O
CH3N+
H
C
O
CN
H
H
C O–
H
Also written as:
Peptides are always drawn with the N-terminus on the left and the C-terminus on the right. Residues are numbered from N-term to C-term.
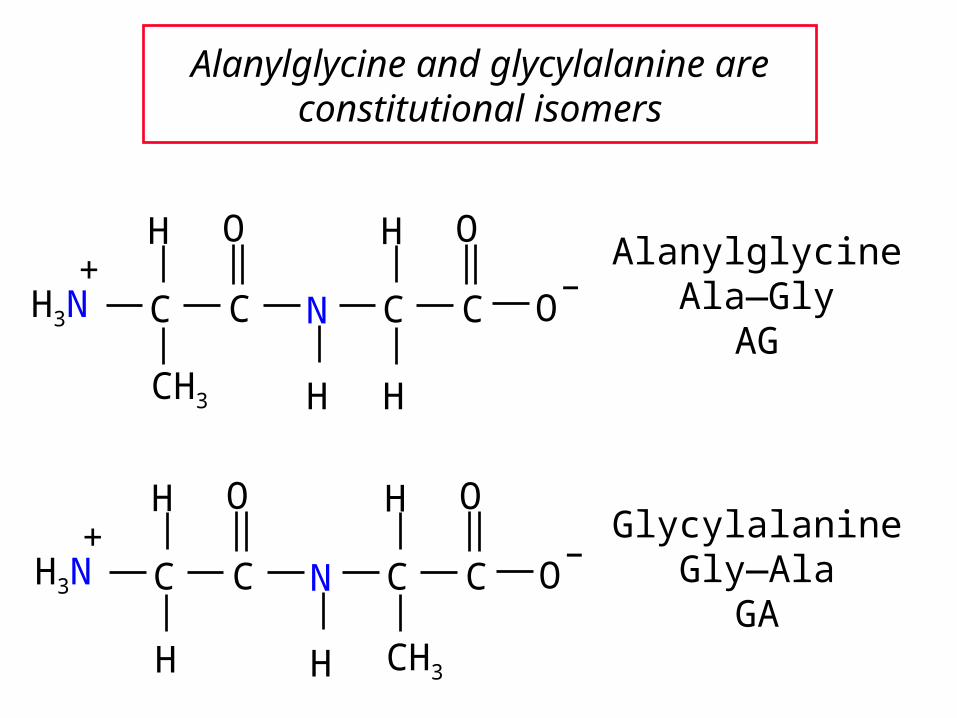
Alanylglycine and glycylalanine are constitutional isomers
H
O
CH3N+
H
C
O
CN
H
CH3
C O–
H
AlanylglycineAla—Gly
AG
GlycylalanineGly—Ala
GA
CH3
O
CH3N+
H
C
O
CN
H
H
C O–
H

Alanylglycine
The peptide bond is characterized by a planar geometry.
The carbonyl O and the amide H are anti to each other.
The six atoms in the box are coplanar.
CH3
O
CH3N+
H
C
O
CN
H
H
C O–
H

Higher Peptides
Peptides are classified according to the number of amino acids linked together, similar to the saccharides, e.g.:
dipeptides, tripeptides, tetrapeptides, etc.
Leucine enkephalin (next slide) is an example of a pentapeptide.
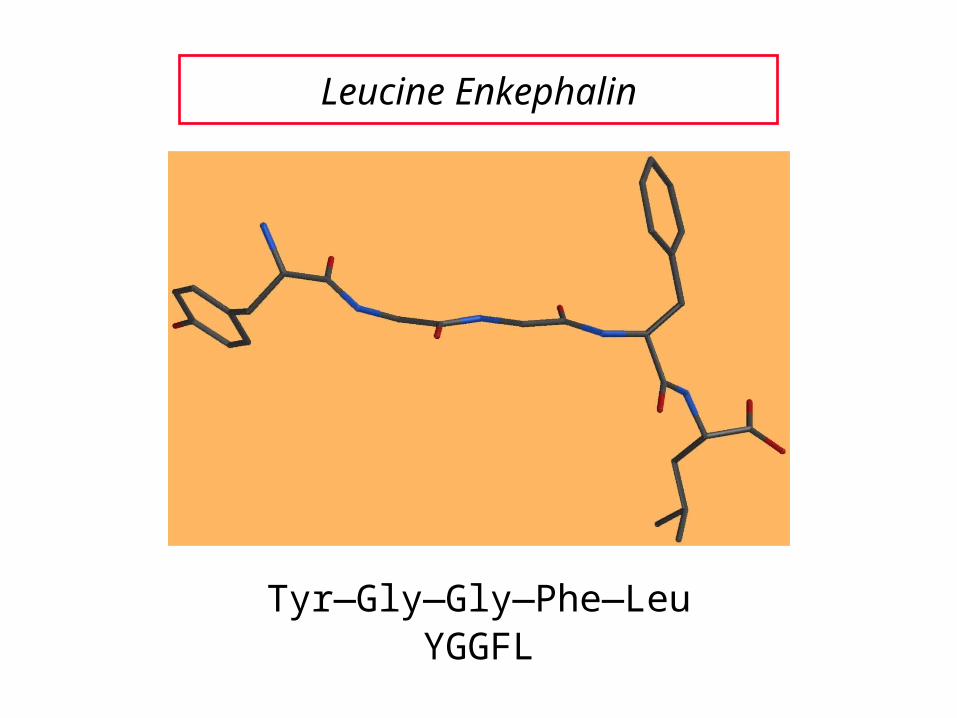
Leucine Enkephalin
Tyr—Gly—Gly—Phe—LeuYGGFL

Oxytocin
Oxytocin is a cyclic nonapeptide.
Instead of having its amino acids linked in an extended chain, two cysteine residues are joined by an S—S bond.
N-terminus
C-terminusIle—Gln—Asn
Tyr
Cys S S
Cys—Pro—Leu—GlyNH2
1
2
34 5
6 7 8 9

Oxytocin
S—S bond
An S—S bond between two cysteines isoften referred to as a disulfide bridge.

25.825.8Introduction to PeptideIntroduction to PeptideStructure DeterminationStructure Determination

Primary Structure
The primary structure of a peptide or protein is its specific sequence of amino acids including any disulfide linkages.
A general strategy for determining primary structure is given on the next slide.

Classical Strategy (Sanger)
1. Determine what amino acids are present and their molar ratios (amino acid composition).
2. Cleave the peptide into smaller fragments, and determine the amino acid composition of each smaller fragment.
3. Identify the N-terminus and C-terminus in the parent peptide and in each fragment.
4. Organize the information so that the sequences of small fragments can be overlapped to reveal the full sequence.

25.925.9Amino Acid AnalysisAmino Acid Analysis

Amino Acid Analysis
Acid-hydrolysis of a peptide gives a mixture of amino acids (heat at 100o in 6 M HCl for 24 hr).
The mixture is separated by ion-exchange chromatography. This method depends on the differences in isoelectric point (pI) among the various amino acids.
Amino acids are detected using ninhydrin.
An automated analyzer requires only 10-1 to 101
micrograms (μg) of peptide.

25.1025.10Partial Hydrolysis of Partial Hydrolysis of
PeptidesPeptides

Partial Hydrolysis of Peptides and Proteins
Acid-hydrolysis of the peptide as mentioned previously cleaves all of the peptide bonds.
Partial hydrolysis cleaves some, but not all, of the peptide bonds and yields smaller peptides.
These small fragments are then separated and the amino acids present in each fragment determined.
Enzyme-catalyzed cleavage is one method used to produce cleavages.

Partial Hydrolysis of Peptides and Proteins
Partial acid hydrolysis is performed using 6 N HCl at 100o for 30 minutes or at 37o for 2-3 days or at lower acid concentrations, etc.
Enzymes that catalyze the hydrolysis of peptide bonds are generally called peptidases, proteases, or proteolytic enzymes, e.g. trypsin, -chymotrypsin, sa protease, etc.

Trypsin
Trypsin is selective for cleaving the peptide bond to the carboxyl group of lysine or arginine.
NHCHC
O
R'
NHCHC
O
R"
NHCHC
O
R
lysine or arginine

Chymotrypsin
Chymotrypsin is selective for cleaving the peptidebond to the carboxyl group of amino acids withan aromatic side chain.
NHCHC
O
R'
NHCHC
O
R"
NHCHC
O
R
phenylalanine, tyrosine or tryptophan

SA Protease
SA Protease is selective for cleaving the peptide bond to the carboxyl group of Asp or Glu.
NHCHC
O
R'
NHCHC
O
R"
NHCHC
O
R
aspartic or glutamic

Carboxypeptidase
proteinH3NCHC
O
R
+NHCHCO
O
R
–C
O
Carboxypeptidase is selective for cleavingthe peptide bond to the C-terminal amino acid.

25.1125.11End Group AnalysisEnd Group Analysis

End Group Analysis
Amino sequence is ambiguous unless we know whether to read it left-to-right or right-to-left.
One needs to know the identity of the N-terminal and C-terminal amino acids.
The C-terminal amino acid can be determined by carboxypeptidase-catalyzed hydrolysis.
Several chemical methods have been developed for identifying the N-terminus. They depend on the fact that the amino N at the terminus is more nucleophilic than any of the amide nitrogens.

Sanger's Method
The key reagent in Sanger's method for identifying the N-terminus is 1-fluoro-2,4-dinitrobenzene (FDNB).
1-Fluoro-2,4-dinitrobenzene is very reactive toward nucleophilic aromatic substitution (Chapter 12).
FO2N
NO2

Sanger's Method
1-Fluoro-2,4-dinitrobenzene reacts with the amino nitrogen of the N-terminal amino acid.
FO2N
NO2
NHCH2C NHCHCO
CH3
NHCHC
CH2C6H5
H2NCHC
O OOO
CH(CH3)2
–+
O2N
NO2
NHCH2C NHCHCO
CH3
NHCHC
CH2C6H5
NHCHC
O OOO
CH(CH3)2
–
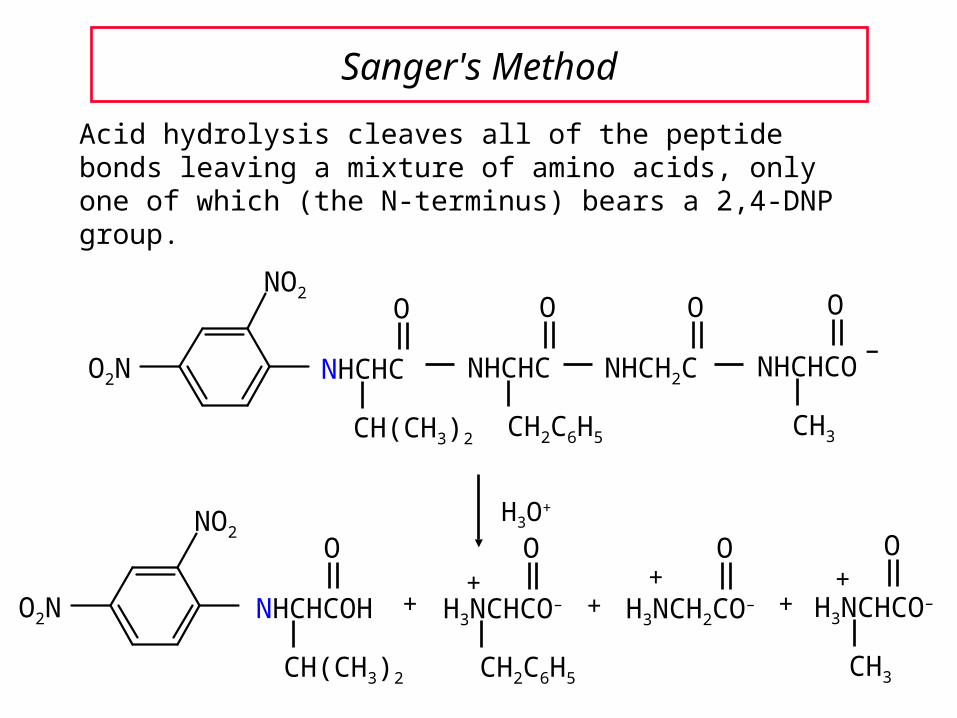
Sanger's Method
Acid hydrolysis cleaves all of the peptide bonds leaving a mixture of amino acids, only one of which (the N-terminus) bears a 2,4-DNP group.
H3O+
O
O2N
NO2
NHCHCOH
CH(CH3)2
H3NCHCO–
CH3
+H3NCH2CO–
O O
+
O
H3NCHCO–
CH2C6H5
++ +
+
O2N
NO2
NHCH2C NHCHCO
CH3
NHCHC
CH2C6H5
NHCHC
O OOO
CH(CH3)2
–

25.1325.13The Edman Degradation andThe Edman Degradation and
Automated Sequencing of PeptidesAutomated Sequencing of Peptides

Edman Degradation
1. Another method for determining N-terminal amino acid.
2. This method sequentially cleaves one residue at a time from the N-terminus. Using the automated method one can usually determine the first 20 or so amino acids from the N-terminus.
3. Only about 10-10 g of sample is needed.

Edman Degradation
The key reagent in the Edman degradation is phenyl isothiocyanate.
The carbon atom between the N and S is very electrophilic and is attacked by the N-term –NH2 of the peptide.
N C S
phenyl isothiocyanate

Edman Degradation
peptideC6H5NHCNHCHC
O
R
NH
S
peptideH3NCHC
O
R
+NHC6H5N C S +

Edman Degradation
The initial product is a phenylthiocarbamoyl (PTC)derivative.
The PTC derivative is then treated with HCl in an anhydrous solvent. The N-terminal amino acid is cleaved from the remainder of the peptide.
peptideC6H5NHCNHCHC
O
R
NH
S

Edman Degradation
HCl
peptideH3N+
+C6H5NH C
SC
N CH
R
O
peptideC6H5NHCNHCHC
O
R
NH
S
The product is a thiazolone which rearranges to a phenylthiohydantoin (PTH) derivative.
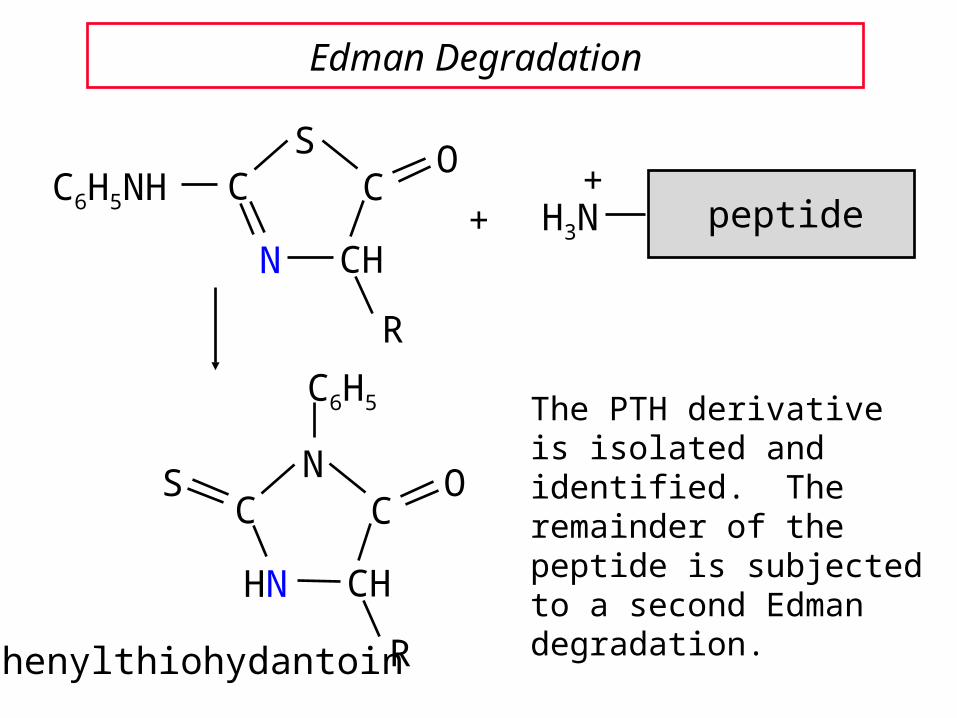
Edman Degradation
The PTH derivative is isolated and identified. The remainder of the peptide is subjected to a second Edman degradation.
peptideH3N+
+C6H5NH C
SC
N CH
R
O
CCN
HN CH
R
OS
C6H5
phenylthiohydantoin

25.1425.14The Strategy of PeptideThe Strategy of Peptide
SynthesisSynthesis

General Considerations
Making peptide bonds between amino acids is not difficult.
The challenge is connecting amino acids in the correct sequence.
Random peptide bond formation in a mixture of phenylalanine and glycine, for example, four dipeptides could be formed.
Phe—Phe Gly—Gly Phe—Gly Gly—Phe

General Strategy
1. The number of possibilities is limited by "protecting" the nitrogen of one amino acid and the carboxyl group of the other.
N-Protectedphenylalanine
C-Protectedglycine
NHCHCOH
CH2C6H5
O
X H2NCH2C
O
Y

General Strategy
2. Couple the two protected amino acids.
NHCH2C
O
YNHCHC
CH2C6H5
O
X
NHCHCOH
CH2C6H5
O
X H2NCH2C
O
Y
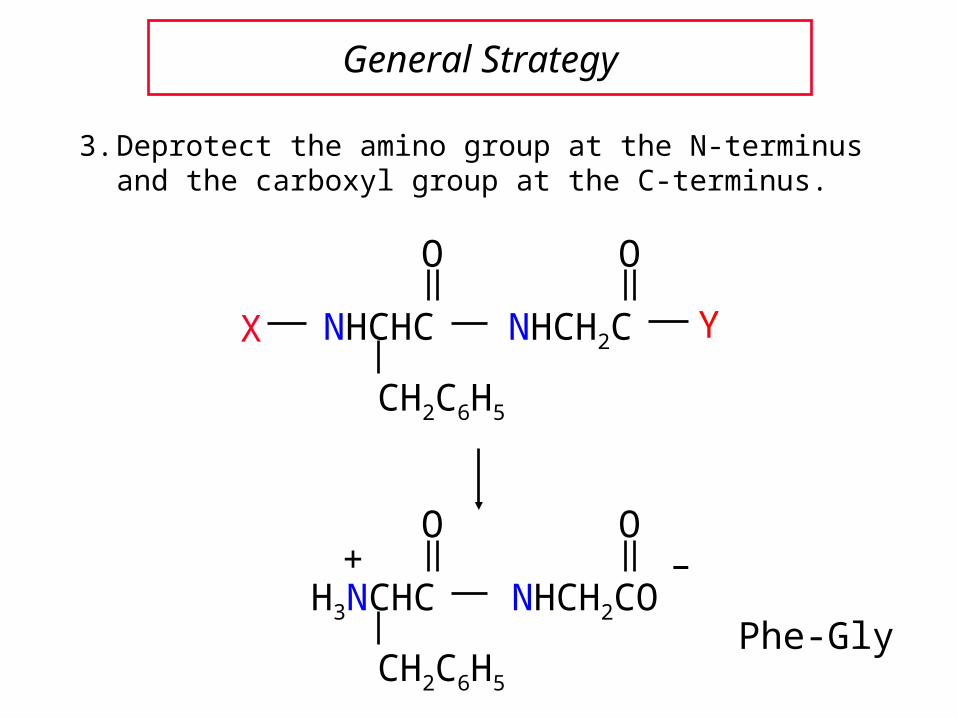
General Strategy
3. Deprotect the amino group at the N-terminus and the carboxyl group at the C-terminus.
NHCH2CO
O
H3NCHC
CH2C6H5
O+ –
Phe-Gly
NHCH2C
O
YNHCHC
CH2C6H5
O
X

25.1725.17Peptide Bond FormationPeptide Bond Formation

The two major methods are:
1. coupling of suitably protected amino acids using N,N'-dicyclohexylcarbodiimide (DCCI)
2. via an active ester of the N-terminal amino acid.
Forming Peptide Bonds
C6H11N C NC6H11
DicyclohexylcarbodiimideThe central C is very electrophilic.

DCCI-Promoted Coupling
ZNHCHCOH
CH2C6H5
O
+ H2NCH2COCH2CH3
O
DCCI, chloroform
ZNHCHC
CH2C6H5
O
NHCH2COCH2CH3
O
(83%)
Overall reaction:

Mechanism of DCCI-Promoted Coupling
ZNHCHCOH
CH2C6H5
O
+ C6H11N C NC6H11
CH2C6H5
O
C6H11N C
C6H11N
H
OCCHNHZ
Oxygen of the acid group attacks DCCI which activates the C=O.

Mechanism of DCCI-Promoted Coupling
H2NCH2COCH2CH3
O
C6H11N C
C6H11NH
H
O + ZNHCHC
CH2C6H5
O
NHCH2COCH2CH3
O
CH2C6H5
O
C6H11N C
C6H11N
H
OCCHNHZ +
Dicyclohexylurea New peptide
Free –NH2 of another amino acid attacks the activated C=O.

25.1925.19Primary and Secondary Structures of Primary and Secondary Structures of
Peptides and Proteins Peptides and Proteins

Levels of Protein Structure
Primary structure = the amino acid sequence plus disulfide links.
Secondary structure = conformational relationship between nearest neighbor amino acids.
helix pleated sheet

Secondary Protein Structure
Planar geometry of peptide bondAnti conformation of main chainHydrogen bonds between N—H and O=C
The -helix and pleated sheet are both characterized by:
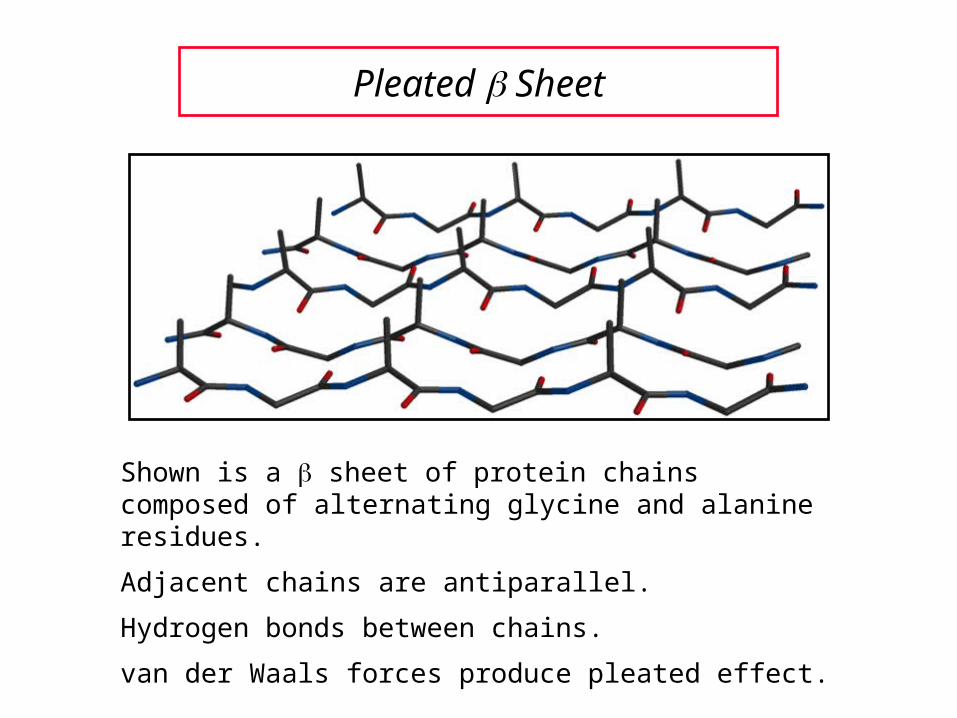
Pleated Sheet
Shown is a sheet of protein chains composed of alternating glycine and alanine residues.
Adjacent chains are antiparallel.
Hydrogen bonds between chains.
van der Waals forces produce pleated effect.

Pleated Sheet
Sheet is most commonly seen with amino acids having small side chains (glycine, alanine, serine).
80% of fibroin (main protein in silk) is repeating sequence of —Gly—Ser—Gly—Ala—Gly—Ala—.
Sheet is flexible, but resists stretching.
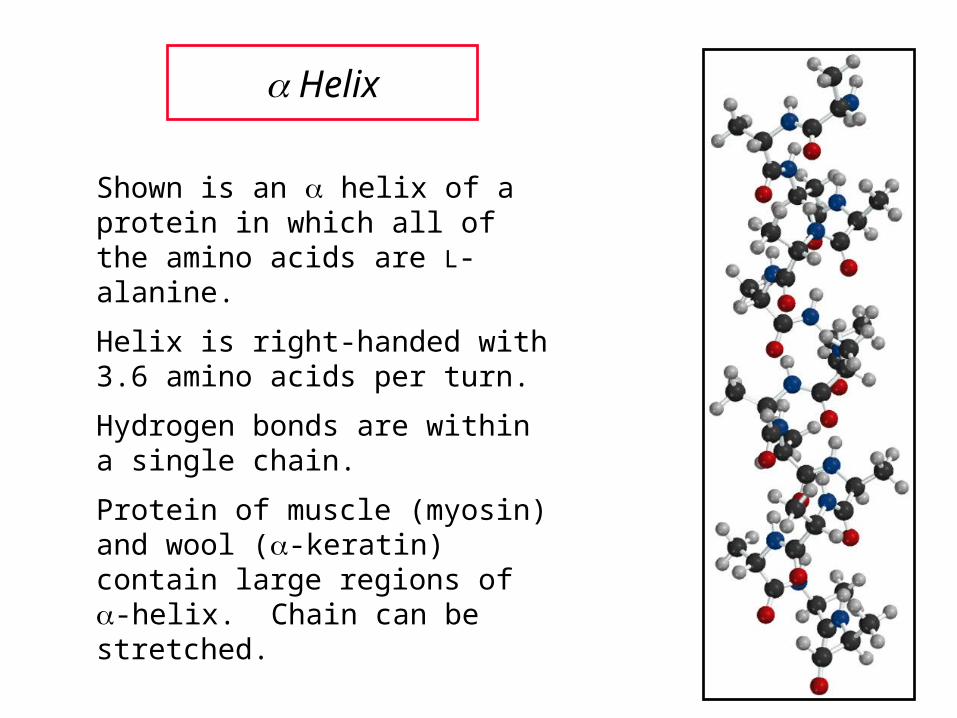
Helix
Shown is an helix of a protein in which all of the amino acids are L-alanine.
Helix is right-handed with 3.6 amino acids per turn.
Hydrogen bonds are within a single chain.
Protein of muscle (myosin) and wool (-keratin) contain large regions of -helix. Chain can be stretched.

25.2025.20Tertiary and Quaternary Structures Tertiary and Quaternary Structures
of of Polypeptides and Proteins Polypeptides and Proteins

Tertiary Structure
Refers to overall shape (how the chain is folded).
Fibrous proteins (hair, tendons, wool) have elongated shapes.
Globular proteins are approximately spherical.
Most enzymes are globular proteins.
An example is carboxypeptidase.

Protein Quaternary Structure
Some proteins are assemblies of two or more chains (3o subunits). The association of these subunits is called the quaternary structure.
Hemoglobin, for example, isa tetramer consisting of 4 subunits.
It has 2 chains (identical) and 2 chains (also identical).
Each subunit contains one heme and each is about the size of myoglobin (monomeric).

End of Chapter 25End of Chapter 25Amino Acids, Peptides, Amino Acids, Peptides,
and Proteinsand Proteins