chameleon species composition and density estimates of three
TRANSCRIPT

Introduction.
Madagascar is extraordinary, not just in its species diversity, but also in its staggering levels of endemism (Goodman and Benstead, 2005; Randrianandianina et al., 2003). This is particularly notable in its reptilian fauna, with 92% of the 363 species being endemic (Glaw and Vences 2007).
Chameleons are of particular conservation interest. As with much of the Malagasy flora and fauna, they are becoming increasingly threatened because of anthropogenic pressures. These pressures are chiefly due to habitat loss and the pet trade, with chameleons being perhaps the most targeted of Madagascar’s herpetofauna among animal collectors (Jenkins et al. 1999; Glaw and Vences 2007; Raselimanana and Rakotomalala 2003).
Among the most threatened chameleon habitats are the dry deciduous forests of northern Madagascar (Smith 1997). This loss of habitat may have future implications on the many known species occurring in this region (Glaw and Vences 2007).
This study examines three unprotected dry deciduous forest fragments, located between Montagne d’Ambre Parc National and Ankarana Réserve Spéciale. The different environmental conditions present in Montagne d’Ambre and Ankarana have given rise to some locally endemic reptile, amphibian, and bird species (Morris and Hawkins 1998; Glaw and Vences 2007). However, it is not known whether populations of other species are
Herpetology Notes, volume 5: 107-113 (2012) (published online on 25 April 2012)
Chameleon species composition and density estimates of three unprotected dry deciduous forests between Montagne d’Ambre Parc National
and Ankarana Réserve Spéciale in northern Madagascar
Andrew J. Lowin
Society for Environmental Exploration / Frontier, 50-52 Rivington Street, London EC2A 3QP, United Kingdom, e-mail: [email protected]
Abstract. Due to anthropogenic pressures, the chameleons of the dry deciduous forests in Northern Madagascar are rapidly losing their habitat. This study uses an already tried and tested methodology, based upon distance sampling, to monitor chameleon populations in three forest fragments between Montagne d’Ambre Parc National and Ankarana Réserve Spéciale. In total, 190 chameleons comprised of four species: Brookesia stumpffi, Furcifer oustaleti, Furcifer pardalis, Furcifer petteri were recorded during the study. Recorded at the highest pooled density was B. stumpffi (149.82 ha-1), followed by F.pardalis (70.48 ha-1), F. petteri (65.56 ha-1), and F.oustaleti (54.12 ha-1). Both F. pardalis and F. oustaleti were absent from one of the forest fragments. All densities were estimated using the computer program DISTANCE. Disturbance surveys in each forest revealed a variety of levels of human disturbances which may well be impacting the chameleon populations. The forest fragments between Montagne d’Ambre Parc National and Ankarana Réserve Spéciale are currently unprotected and may well be home to some of the last vestiges of now isolated chameleon populations. Without further in depth information, such as those provided by this study, successful conservation management strategies for the future will be difficult.
Keywords. Chameleons, Population densities, Forest fragments, Dry deciduous forests, Corridor, Anthropogenic pressures.
Figure 1. Map of study sites, (a) Ankaz, (b) Antsolipa, and(c) Bat Roost.

Andrew J. Lowin108
continuous between the two protected areas and at what densities they occur. The forest fragments surveyed in this study presumably represent some of the last vestiges of forest habitat that once linked the two protected areas of Montagne d’Ambre and Ankarana. With intense anthropogenic pressures recorded in the area (Lowin 2011, Durkin et al. 2011), the threats to these fragments are serious.
Jenkins et al. (2003) in particular, calls for species inventories and density data in order to monitor and compare future programmes of conservation. This study compares the three forest fragments for their chameleon species composition, population density estimates, and levels of disturbance.
Materials and Methods. Study sites
(a) The three study sites: Ankaz (12º 45’03.7’S, 049º 08’57.1’’E), (b) Antsolipa (12º 46’07.3’S, 049º 09’23.6’ E), and (c) Bat roost (12º 45’16.1’S, 049º 10’01.3’E) are situated ap-proximately 90km south of Antsiranana (Diego Suarez), between Montagne d’Ambre Parc National to the north and Ankarana Ré-serve Spéciale to the south, near the small commercial settlement of Anivorano (Figure 1.). Each Forest fragment varies in size: (a) 363ha, (b) 138ha, and (c) 66ha.
The geographical location of the study area is in a ‘transition zone’ between two already protected areas. The region is characte-rized by heavy degradation with several pockets of secondary dry deciduous forest in the surrounding valley.
Transect SurveysSurveys were carried out from November 2009 to May 2010
following the methods of Brady and Griffiths (2003) in order to form comparisons. Within each forest, line transects were establis-hed randomly from forest paths. Transects consisted of three 30m straight lines arranged in parallel and separated by a distance of 20m. Lines all started from a path within the forest and followed a compass bearing to ensure that they remained parallel. Measuring tapes were used to mark out each 30m line and then flagging tape was attached to trees at breast height marking every metre. Every other piece of flagging tape was marked with the distance from the path (i.e., every two metres).
To limit the effects of disturbance, transects were left for a mi-nimum of 24 hours before being surveyed. Roosting chameleons become very pale at night (Parcher 1974) and are relatively easy to detect with a flash light (Raxworthy 1988). Thus, each 3m x 30m transect was surveyed at night by a team of two people. The-se methods assume that all chameleons were seen within 1.5m of the transect line and with a vertical detection rate of up to 6m. Search speed was dependant on the observer and also the terrain being surveyed but was estimated at a mean search speed of 1.80m/min. Each transect was surveyed three times, again leaving 24 hours between surveying to minimize disturbance.
Plate 1. Adult male Furcifer petteri Plate 2. Brookesia stumpffi
Plate 3. Adult male Furcifer pardalis Plate 4. Adult female Furcifer oustaleti

Chameleon densities in northern Madagascar 109
Upon finding a chameleon, its species, sex, and perpendicular distance from the transect were recorded. The computer program DISTANCE 5.0 (Laake et al., 1993) was used to estimate the den-sities of all species.
DisturbancePaths within each of the three forests were chosen at random for
the vegetation surveys, with survey points at every twenty steps. Ideally twenty points would be made during each survey but this was not possible in some cases due to the size of the forest and the location of the paths (i.e. some paths left the forest during a survey). At each point, evidences of the following disturbances were recorded: Cut trees, burning, animal traps, cattle grazing, cultivation, introduced species, and a path wider than one metre.
Results
Species composition and density estimates Chameleons were surveyed along 21 transects in each
of the three forests (Survey effort = 1890m per forest). A total of 190 chameleons comprised of four species: Brookesia stumpffi, Furcifer oustaleti, Furcifer pardalis, Furcifer petteri were recorded during the study. The most frequently recorded species was B. stumpffi (n=126), followed by F. petteri (n=39), F. pardalis (n=13), and F. oustaleti (n=6).
The highest estimated mean densities of chameleons
(117.61 ha-1) were recorded at Ankaz, followed by Antsoli-pa (79.44 ha-1), and Bat Roost (41.28 ha-1). The two species F. petteri and F. oustaleti were not record-ed in Bat Roost and notably high densities of B. stumpffi were recorded in Ankaz forest (Table 1).
Population StructureA higher proportion of male F. oustaleti and F.
pardalis were recorded in all forests except Bat Roost where neither species was recorded (Table 2). A notably high proportion of B. stumpffi hatchlings were recorded in Ankaz forest.
Distribution and Protective statusAll four species recorded are listed in appendix II of
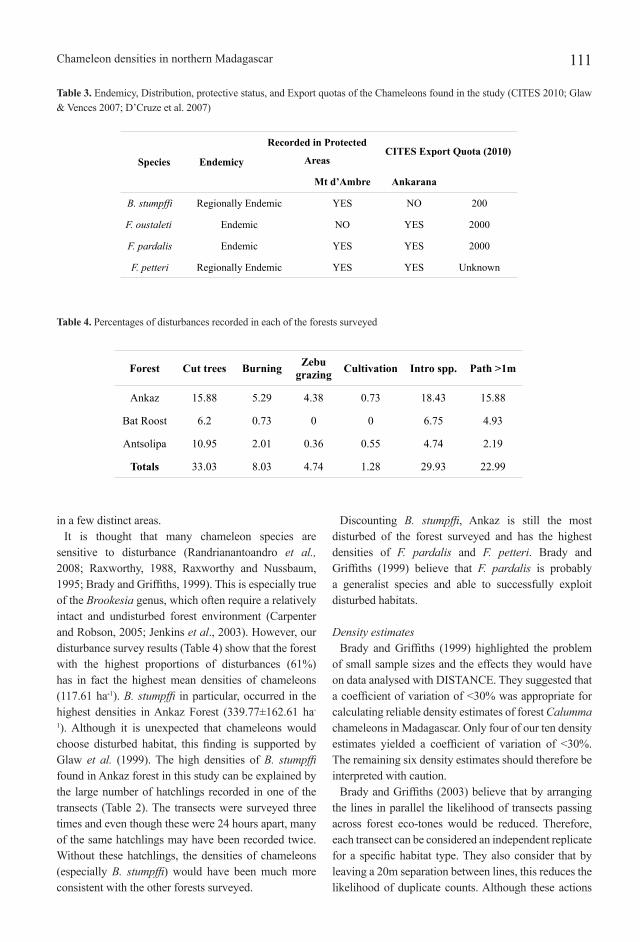
CITES 2010, limiting their export quotas (Table 3). Both B. stumpffi and F. petteri are regionally endemic to the area.
Disturbance
A total of 100 points were surveyed in each of the three forests for evidence of disturbances. The forest with the highest proportion of disturbances recorded was Ankaz with 61%, followed by Antsolipa with 21%, and Bat Roost with 18%.
Study Area Species n Density (ha-1) ± SE % CV 95% CI
B. stumpffi 81 339.77±162.61 47.86 123.71 – 933.15
F. oustaleti 2 37.35±18.66 49.99 23.22 – 90.69
F. pardalis 7 81.14±24.89 30.68 41.50 – 158.63 Ankaz
F. petteri 10 74.58±28.73 38.52 32.86 – 169.13
B. stumpffi 31 59.81±14.13 23.62 37.43 – 95.52
F. oustaleti 4 70.89±22.98 32.43 25.91 – 193.91
F. pardalis 6 59.81±29.69 49.87 19.96 – 177.24 Antsolipa
F. petteri 12 48.11±14.42 29.98 25.50 – 90.74
B. stumpffi 14 49.87±13.94 27.95 27.90 - 89.09
Bat Roost F. petteri 17 73.99±16.72 22.6 46.64 – 117.37
n, Number of observations; % CV, per cent coefficient of variation; 95% CI, 95 per cent confidence intervals.
Table 1. DISTANCE calculated estimated densities for chameleon populations in the three forests surveyed

Discussion
DistributionTable 3 summarizes the data relevant to our species
of interest found by D’Cruze et al. 2007, Jenkins et al. 1999, and Glaw and Vences 2007. They show that only F. pardalis and F. petteri are found in both Ankarana and Montagne d’Ambre. B. stumpffi is only recorded in Montagne d’Ambre and F. oustaleti is only recorded in Ankarana.
Interestingly in this study, F. oustaleti was recorded in all forests except Bat Roost forest, which is approximately 2km from the southern park limit of Montagne d’Ambre and is the closest of the three forests surveyed to the National Park. The next closest forest to Bat Roost is Ankaz (approx 5km from Montagne d’Ambre), where F. oustaleti was recorded. This suggests that Ankaz forest may be the limit of F. oustaleti’s distribution from Ankarana. Furcifer spp. are regarded to only inhabit dry forests (Andreone et al. 2005) but the two forests of Bat Roost and Ankaz are very similar in structure and species composition as the outer limits of Montagne d’Ambre (a montane rain-forest). Outside of the survey work, F. oustaleti was often seen in a variety of highly disturbed habitats between forest fragments, so in the author’s opinion, this species does in fact not only occur in all of the fragments between the two protected areas but also potentially in the lower foot hills of Montagne d’Ambre itself. Further survey work in this area would be needed to confirm this.
Because B. stumpffi was recorded at all three sites, it is impossible to know the full extent of its range without
surveying the remaining forests between our survey sites and Ankarana. As B. stumpffi has been found in both rainforest and dry forest habitat (Carpenter and Robson, 2005; Glaw and Vences 2007), it would be interesting to study where the species does disappear from the section of forest fragments between our study sites and Ankarana, and why it does not occur in the dry forests of Ankarana.
Threats and legislation The major threats to all chameleon species are habitat
destruction and the trade industry (Carpenter and Robson, 2005; Randrianantoandro et al., 2008; Brady and Griffiths, 1999). All four of the species recorded are listed on appendix II of CITES 2010, under which their trade is controlled to avoid utilization compromising their future survival. B. stumpffi is the least widespread species that we recorded and is regionally endemic to distinct parts of northern Madagascar (Glaw and Vences 2007; Carpenter and Robson, 2005). This species has the lowest CITES export quota (200 in 2010) of the four species we recorded, reflecting its vulnerability. F. pardalis and F. oustaleti both have export quotas of 2000 and it is suggested by Andreone et al. (2005) that this collection rate has little impact on the population of F. pardalis. So if we were to make recommendations based on these findings, it would be that the export quota for B. stumpffi could in fact be raised to a similar level as the Furcifer species we recorded. However, we know that B. stumpffi is a regionally endemic species (Table 3) and therefore may only be found in these high densities
Andrew J. Lowin110
Table 2. Population structure of chameleons recorded in the three study sites
Study Area Species Male Female Unknown Hatchling/Adult
Ankaz
B. stumpffi - - 1.00 0.70/0.30
F. oustaleti 0.62 0.38 - 0.1/0.99
F. pardalis 0.55 0.45 - 0.00/1.00
F. petteri 0.41 0.59 - 0.00/1.00
Antsolipa
B. stumpffi - - 1.00 0.38/0.62
F. oustaleti 0.59 0.41 - 0.00/1.00
F. pardalis 0.67 0.33 - 0.00/1.00
F. petteri 0.46 0.54 - 0.00/1.00
Bat RoostB. stumpffi - - 1.00 0.17/0.83
F. petteri 0.39 0.61 - 1.00
Data represents proportions of total observations of n (Table 2). B. stumpffi could not
be sexed. Every chameleon that was recorded was classed as either a hatchling or an
adult as juveniles were unable to be identified.
in a few distinct areas. It is thought that many chameleon species are
sensitive to disturbance (Randrianantoandro et al., 2008; Raxworthy, 1988, Raxworthy and Nussbaum, 1995; Brady and Griffiths, 1999). This is especially true of the Brookesia genus, which often require a relatively intact and undisturbed forest environment (Carpenter and Robson, 2005; Jenkins et al., 2003). However, our disturbance survey results (Table 4) show that the forest with the highest proportions of disturbances (61%) has in fact the highest mean densities of chameleons (117.61 ha-1). B. stumpffi in particular, occurred in the highest densities in Ankaz Forest (339.77±162.61 ha-
1). Although it is unexpected that chameleons would choose disturbed habitat, this finding is supported by Glaw et al. (1999). The high densities of B. stumpffi found in Ankaz forest in this study can be explained by the large number of hatchlings recorded in one of the transects (Table 2). The transects were surveyed three times and even though these were 24 hours apart, many of the same hatchlings may have been recorded twice. Without these hatchlings, the densities of chameleons (especially B. stumpffi) would have been much more consistent with the other forests surveyed.
Discounting B. stumpffi, Ankaz is still the most disturbed of the forest surveyed and has the highest densities of F. pardalis and F. petteri. Brady and Griffiths (1999) believe that F. pardalis is probably a generalist species and able to successfully exploit disturbed habitats.
Density estimates Brady and Griffiths (1999) highlighted the problem
of small sample sizes and the effects they would have on data analysed with DISTANCE. They suggested that a coefficient of variation of <30% was appropriate for calculating reliable density estimates of forest Calumma chameleons in Madagascar. Only four of our ten density estimates yielded a coefficient of variation of <30%. The remaining six density estimates should therefore be interpreted with caution.
Brady and Griffiths (2003) believe that by arranging the lines in parallel the likelihood of transects passing across forest eco-tones would be reduced. Therefore, each transect can be considered an independent replicate for a specific habitat type. They also consider that by leaving a 20m separation between lines, this reduces the likelihood of duplicate counts. Although these actions
Chameleon densities in northern Madagascar 111
Table 3. Endemicy, Distribution, protective status, and Export quotas of the Chameleons found in the study (CITES 2010; Glaw & Vences 2007; D’Cruze et al. 2007)
Species Endemicy
Recorded in Protected
AreasCITES Export Quota (2010)
Mt d’Ambre Ankarana
B. stumpffi Regionally Endemic YES NO 200
F. oustaleti Endemic NO YES 2000
F. pardalis Endemic YES YES 2000
F. petteri Regionally Endemic YES YES Unknown
Table 4. Percentages of disturbances recorded in each of the forests surveyed
Forest Cut trees BurningZebu
grazingCultivation Intro spp. Path >1m
Ankaz 15.88 5.29 4.38 0.73 18.43 15.88
Bat Roost 6.2 0.73 0 0 6.75 4.93
Antsolipa 10.95 2.01 0.36 0.55 4.74 2.19
Totals 33.03 8.03 4.74 1.28 29.93 22.99

may help to reduce these potential biases we must consider their possible implications to the estimates given.
In Randrianantoandro et al. (2008), differences in the density of chameleons between sites are thought to be related to vegetation structure/habitat quality, illicit collection (past or present), seasonality or altitude. Without vegetation surveys it is difficult to assess whether vegetation structure affected our density estimates. It appears that in this study disturbance increases chameleon densities. It is not known whether illicit collection occurs in the area but in the author’s opinion it would not be by local people as they are still very superstitious about chameleons. As the three survey sites were all at approximately the same altitude it is also difficult to say whether altitude effected density estimates.
Seasonality does definitely affect densities. For example, as Brady and Griffiths (2003) also mention, during the dry season (May to August) Brookesia are undetectable as they are thought to enter a state of aestivation. If we had conducted the survey work during this time we may have concluded that B. stumpffi was not present in the area as opposed to being the chameleon occurring in the highest densities. We must also assume that not all the chameleons present on our surveys were in fact recorded. This maybe particularly relevant to B. stumpffi which are often very cryptic. Seasonality may also have affected the other species recorded during the study. Base camp was situated in Antsolipa forest and from July to September (outside the survey work for this study) F. oustaleti appeared to be much more abundant. The reason for this is uncertain but it is possible that the true density of F. oustaleti was underestimated due to the time of year when the survey work was carried out.
ConclusionThe survey work in this paper provides abundance
and species diversity information about chameleons in forests that are under varying levels of anthropogenic disturbance. This study continues the work of Jenkins et al. (2003), Randrianantoandro et al. (2008), and Brady and Griffiths (1999) who all provided information on abundance and species diversity in order to target future conservation efforts. Our results show that the study area may well be part of a broken corridor between two protected areas. The area is also shown to be under pressure from human disturbance which is affecting the chameleon population to varying degrees.
Many studies of chameleons have mostly focused on
site-specific inventories (Raxworthy, 1988; Raxworthy and Nussbaum, 1995), with little known about the abundance of individual species. For example, Carpenter and Robson, 2005 stated that at the time of their research, no studies were currently investigating population sizes or trends in any species of Brookesia.
Unless species composition and density data are available, how are we able to make comparisons and monitor programmes in the future. Comparisons of the effects of disturbance and the pet trade will be impossible without these data.
Acknowledgements. I would like to thank all of the students and staff from Frontier who helped with the field work, especially Andy Allen. Finally, I would like to thank Andrea Baden for all of her editing support.
References
Andreone, F., Guarino, F.M., and Randrianirina, J.E. (2005): Life history traits, age profile, and conservation of the panther chameleon, Furcifer pardalis (Cuvier 1829), at Nosy Be, NW Madagascar. Tropical Zoology 18: 209-225.
Brady, L.D. & Griffiths, R.A. (1999): Status Assessment of Cha-meleons in Madagascar. IUCN Species Survival Commission, Gland, Switzerland.
Brady, L.D. & R.A. Griffiths (2003): Chameleon population es-timates, pp. 970-972, The Natural History of Madagascar. The University of Chicago Press and London.
Carpenter, A.I & Robson, O. (2005): A review of the endemic chameleon genus Brookesia from Madagascar, and the ratio-nale for its listing on CITES Appendix II) Oryx Vol 39 No 4.
CITES (2010): Convention on international trade in endangered species of wild fauna and flora. Http://www.cites.org [accessed 1 June 2011].
D’cruze, N., Sabel, J., Dawson, J., Gardner, C, Robinson, J., Starkie, G., Vences, M., Glaw, F. (2007): The First compre-hensive survey of amphibians and reptiles at Montagne des Francais, Madagascar. Herpetological Conservation and Bio-logy 2(2):87-99.
Durkin, L., Steer, M.D., and Belle, E. (2011): Herpetological Sur-veys of forest fragments between Montagne D’Ambre Natio-nal Park and Ankarana Special Reserve, Northern Madagascar. Herpetological Conservation and Biology 6(1):114-126.
Goodman, S., and Benstead, J. (2005): Updated estimates of bio-tic diversity and endemism for Madagascar. Oryx, 39,73-77.
Glaw, F. & Vences, M. (2007): A field guide to the amphibians and reptiles of Madagascar, 3rd edition. Vences and Glaw Ver-lag, Koln.
Glaw, F., Vences, M., Ziegler, T., Bohme, W. & Kohler, J. (1999): Specific distinctness and biogeography of the dwarf chame-leons Brookesia minima, B. peyrierasi and B. Tuberculatab (Reptilia: Chamaeleonidae): evidence from hemipenial and external morphology. Journal of Zoology, London, 247,225–238.
Andrew J. Lowin112

Jenkins, R.K.B., Brady, L.D., Huston, K., Kauffmann, J.L.D., Re-bearivony, J., Raveloson, G., and Rowcliffe, M. (1999): The population status of chameleons within Ranomafana National Park, Madagascar, and recommendations for future monito-ring. Oryx 33:38-46.
Jenkins, R.K.B., Brady, L.D., Bisoa, M., Rabearivony, J. & Grif-fiths, R. (2003): Forest disturbance and river proximity influ-ence chameleon abundance in Madagascar. Biological
Conservation, 109, 407–415.Laake, J.L., Buckland, S.T., Anderson, D.R. & Burnham, K.P.
(1993): DISTANCE User’s Guide. Version 2.0. Colorado Coo-perative Fish and Wildlife Research Unit, Colorado State Uni-versity, Fort Collins.
Lowin, A. (2011): Lepilemur feeding observations from Northern Madagascar. Lemur News Vol.15.
Mittermeier, R.A., Konstant, W.R., and Rylands, A.B. (2003): Lemur Conservation. pp.1538-1545, The Natural History of Madagascar. The University of Chicago Press and London.
Morris, P., and F. Hawkins. (1998): Birds of Madagascar: A Pho-tographic Guide. Yale University Press, New Haven, Connec-ticut, USA.
Parcher, S.R. (1974): Observations on the natural histories of six Malagasy chamaeleontidae. Z.Tierpsychol. 34:500-523.
Raselimanana A.P. and Rakotomalala D. (2003): Chamaeleoni-dae, Chameleons. pp. 960-970, The Natural History of Mada-gascar. The University of Chicago Press and London.
Raxworthy, C.J. (1998): Reptiles, rainforest and conservation in Madagascar. Biological Conservation 43: 181-211.
Raxworthy, C.J. (2003): Introduction to reptiles, pp. 934-949, The Natural History of Madagascar. The University of Chicago Press and London.
Raxworthy, C.J., and Nussbaum, R.A. (1995): Systematics, spe-ciation and biogeography of the dwarf chameleons (Brookesia Reptilia, Squamata, Chamaeleontidae) of northern Madagas-car. Journal of Zoology 235, 525–558.
Randrianandianina, B.N., L.R. Andriamahaly, F.M. Harisoa and M.E. Nicoll. (2003). The Role of the Protected Areas in the Management of the Island’s Biodiversity. In: The Natural His-tory of Madagascar (eds. S.M. Goodman and J.P. Benstead), pp. 1423-1432. University of Chicago Press, Chicago.
Randrianantoandro, C.J, Randrianavelona, R., Andriantsimana-rilafy, R., Hantalalaina, E.F., Rakotondravony, D., Randria-nasolo, M., Ravelomanantsoa, H.L., and Jenkins R.B. (2008): Identifying important areas for the conservation of dwarf chameleons (Brookesia spp.) in Tsingy de Bemaraha National Park, western Madagascar. Oryx 42(4) 578-583
Smith, A.P. (1997): Deforestation, fragmentation, and reserve design in western Madagascar. pp.415-441 in Tropical Forest Remnants, Ecology, Management and Conservation of frag-mented Communities (eds. Lawrence, W. & O.W. Bierreg-aard). – University Press, Chicago.
Chameleon densities in northern Madagascar 113
Accepted by Zoltan T. Nagy