cellular damage to phytophthora infestans in tomato leaves treated with oxadixyl: an ultrastructural...
TRANSCRIPT

J. Phytopathology 132, 116—128 (1991)© 1991 Paul Parey Scientific Publishers, Berlin and HamburgISSN 0931-1785
Universitat Hohenheim, Institut fur PhytomedizinPostfach 700562, 7000 Stuttgart 70, Federal Republic of Germany
Cellular Damage to Phytophthora infestans in Tomato LeavesTreated with Oxadixyl: an Ultrastructural Investigation
YoNG JIANG and F. GROSSMANN
Authors' addresses: Y. JIANG, Department of Crop Protection, Waite Agricultural Research Institute,Glen Osmond, SA 5064 (Australia). F. GROSSMANN, Universitat Hohenheim, Institut fiir Phytomedi-
zin (360), Postfach 700562, D-7000 Stuttgart 70 (Fed. Rep. of Germany).
With 12 figures
Received March 14, 1990; accepted January 9, 1991
Abstract
Ukrastructural observations showed massive accumulation of electron dense deposits adjacentto plasmalemma and mitochondria and mitochondrial degeneration, which probably leads to celldeath, in some hyphae and haustoria of Phytophthora infestans in infected tomato leaves treated withthe systemic fungicide oxadixyl (8 ;Ug/ml). However, no changes in the endoplasmic reticulum andnumber of ribosomes were observed. The cell walls of some hyphae were markedly thickened afteroxadixyl treatment. Degenerated haustoria were partly present in intact host cells. Evidently, fungalcell damage by treatment with oxadixyl is not correlated with host cell damage.
Zusammenfassung
Zellschaden bei Phytophthora infestans in Tomatenblatternnach Behandlung mit Oxadixyl: Eine ultrastrukturelle Untersuchung
Ultrastrukturelle Untersuchungen zeigten eine massive Akkumulation von elektronendichtenAblagerungen in unmittelbarer Nachbarschaft des Plasmalemmas und der Mitochondrien sowieDegeneration der Mitochondrien, die wahrscheinlich zum Zelltod fiihrt, in einigen Hyphen undHaustorien von Phytophthora infestans in infizierten Tomatenblattern, die mit dem systemischenFungizid Oxadixyl (8 Mg/ml) behandelt worden waren. Veranderungen des endoplasmatischen Reticu-lums und der Zahl der Ribosomen wurden jedoch nicht beobachtet. Ferner waren die Zellwandeeiniger Hyphen nach der Behandlung betrachtlich verdickt. Degenerierte Haustorien wurden teil-weise in intakten Wirtszellen gefunden. Offensichtlich korreliert die Schadigung der pilzlichen Zellennach Behandlung mit Oxydixyl nicht mit der Schadigung der Wirtszellen.
U.S. Copyright Clearance Center Code Statement: 0931-1785/91/3202-0116$02.50/0

Cellular Damage to Phytophthora infestans Treated with Oxadixyl 117
Oxadixyl is a systemic fungicide acting mainly on the Peronosporales ofoomycetous fungi (FULLER and Gisi 1985). Based on the similarity of the chemicalstructure and the cross-resistance to metalaxyl, a systemic fungicide in use sincemany years, it is expected that oxadixyl may have the same inhibitory effect onrRNA synthesis of Oomycetes as metalaxyl (COHEN and COFFEY 1986, DAVIDSE
et al. 1988). Ukrastructural studies revealed that ribosomes of Phytophthorainfestans were indeed reduced after treatment with the fungicide (unpublished).Moreover, the haustorium development of the fungus was also inhibited (unpub-lished) and interpreted as the consequence of the fungistatic effect. However,application of oxadixyl in combination with the protective fungicide, mancozeb,successfully controls plant diseases caused by pathogens which are even resistantto metalaxyl (Gisi et al. 1985) and oxadixyl displays less activity in vitro than, butthe same activity in vivo as, metalaxyl (COHEN and COFFEY 1986). Therefore,differences between oxadixyl and metalaxyl in their mode of action seem to exist.
In an ukrastructural study with Phytophthora infestans in infected tomatoleaves treated with oxadixyl, it was observed that oxadixyl exhibited not onlyfungistatic, but also fungitoxic effects on the fungus, which have not yet beenreported on metalaxyl-treated fungi in vitro and in vivo (HiCKY and COFFEY
1980, GROHMANN and HOFFMANN 1982, COHEN and COFFEY 1986). Hence, thepossibility of another action site of oxadixyl is implied to be present.
Materials and Methods
Fungus material
Phytophthora infestans Mont (de Bary), race 1.4, was cultured on potato discs (cv. 'Bintje') at20 °C. Four days after inoculation, sporangia were collected in distilled water and incubated at 12 °Cfor 3—4 h until zoospores were released. The resulting zoospore suspension was used for inoculationof leaves.
Plant material
Seedlings of Lycopersicum esculentum (cv. 'Bonny Best'), susceptible to the above mentionedrace, were planted in 9-cm-diameter plastic pots containing a mixture of Humosoil, perlite and sand(1 : 1 : 1, v/v) and cultivated in the greenhouse at 18—26 °C. Five-week-old plants were used for theexperiments.
Fungicide and application
Oxadixyl [2-methoxy-N-(2-oxo-l,3-oxazolidin-3-yl)acet-2',6'xylidide], technical grade, 96 %pure, was obtained from Sandoz Ltd. It was dissolved in tap water (8 jug/ml, equal to the EC50 value)and sprayed onto the upper surfaces of tomato leaves 24 h before inoculation. As a control, only waterwas sprayed.
Inoculation
Small filter paper discs (5 mm in diameter) were dipped in a freshly prepared zoosporesuspension (10* ml"') and placed on the lower surfaces of the third and fourth leaves, in order to avoiddirect contact between the fungicide and the fungus before infection. After inoculation, all plants werekept in a moist chamber at 20 °C.
Electron microscopy
15 and 18 h after inoculation, the inoculated areas of the leaves were excised with a razor bladeand placed in buffered glutaraldehyde (3 % in 0.6 mol/1 cacodyiate, pH 7.0, containing 3 mmol/1
l ) , fixed at room temperature for 2 h. After washing (3 x 20 tnin) in the same buffer, the
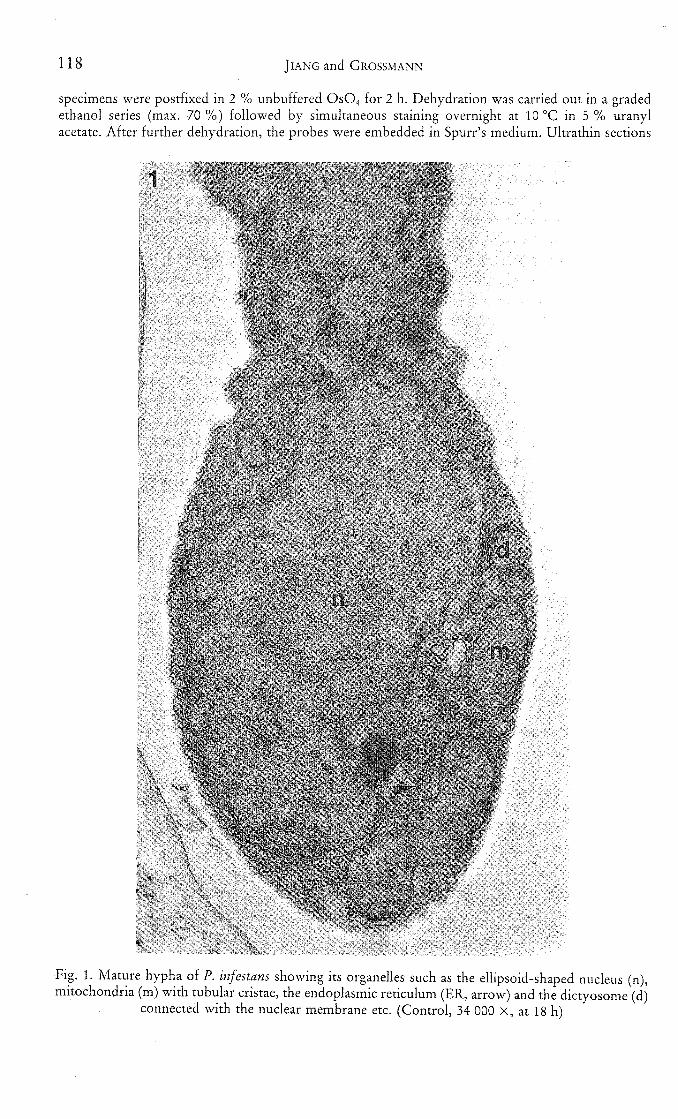
118 JIANG and GROSSMANN
specimens were postfixed m 2 % unbuffered OSO4 for 2 h. Dehydration was carried out in a gradedethanol series (max. 70 %) followed by simultaneous staining overnight at 10 °C in 5 % uranylacetate. After further dehydration, the probes were embedded in Spurr's medium. Ultrathin sections
1
Fig. 1. Mature hypha of P. infestans showing its organelles such as the ellipsoid-shaped nucleus (n),mitochondria (m) with tubular cristae, the endoplasmic reticulum (ER, arrow) and the dictyosome (d)
connected with the nuclear membrane etc. (Control, 34 000 X, at 18 h)
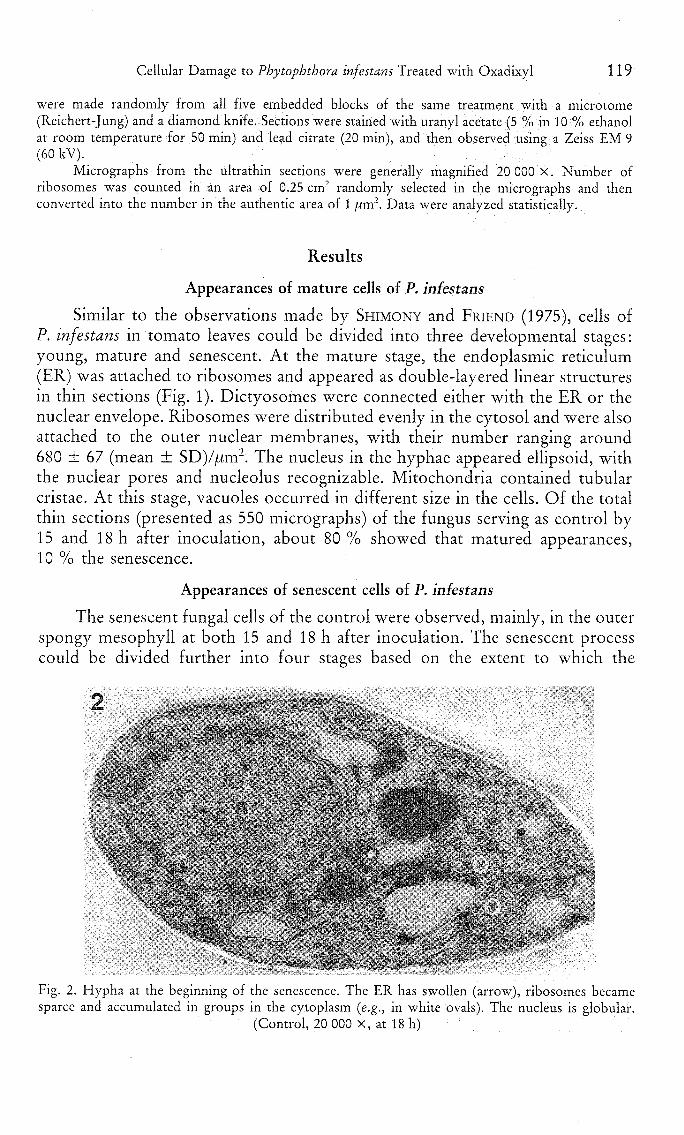
Cellular Damage to Phytophthora infestans Treated with Oxadixyl 119
were made randomly from all five embedded blocks of the same treatment with a microtome(Reichert-Jung) and a diamond knife. Sections were stained with uranyl acetate (5 % in 10 % ethanolat room temperature for 50 min) and lead citrate (20 min), and then observed using a Zeiss EM 9(60 kV).
Micrographs from the ultrathin sections were generally magnified 20 000 X. Number ofribosomes was counted in an area of 0.25 cm^ randomly selected in the micrographs and thenconverted into the number in the authentic area of 1 jum .̂ Data were analyzed statistically.
Results
Appearances of mature cells of P. infestans
Similar to the observations made by SHIMONY and FRIEND (1975), cells ofP. infestans in tomato leaves could be divided into three developmental stages:young, mature and senescent. At the mature stage, the endoplasmic reticulum(ER) was attached to ribosomes and appeared as double-layered linear structuresin thin sections (Fig. 1). Dictyosomes were connected either with the ER or thenuclear envelope. Ribosomes were distributed evenly in the cytosol and were alsoattached to the outer nuclear membranes, with their number ranging around680 ± 67 (mean ± SD)//im^. The nucleus in the hyphae appeared ellipsoid, withthe nuclear pores and nucleolus recognizable. Mitochondria contained tubularcristae. At this stage, vacuoles occurred in different size in the cells. Of the totalthin sections (presented as 550 micrographs) of the fungus serving as control by15 and 18 h after inoculation, about 80 % showed that matured appearances,10 % the senescence.
Appearances of senescent cells of P. infestans
The senescent fungal cells of the control were observed, mainly, in the outerspongy mesophyll at both 15 and 18 h after inoculation. The senescent processcould be divided further into four stages based on the extent to which the
Fig. 2. Hypha at the beginning of the senescence. The ER has swollen (arrow), ribosomes becamesparce and accumulated in groups in the cytoplasm {e.g., in white ovals). The nucleus is globular.
(Control, 20 000 X, at 18 h)

120 JIANG and GROSSMANN
austorium in the middle stage of the senescence. Electron dense deposits appear at iFig. 3. A haustorium in the middle stage of the senescence. Electron dense deposits appear at nuclear(n) and mitochondrial (m) membranes and at tonoplasts of some vacuoles. The host cell has
degenerated already, showing no extrahaustorial membrane. (Control, 20 000 X, at 15 h)
ukrastructural changes in organelles, especially mitochondria, occurred in thisstudy: 1. Primary stage of the senescence; appearance of many organelles exceptmitochondria were changed. The ER was fragmented and swelled, but remainedattached to ribosomes, which, however, were not in contact with the nuclearenvelope. The nucleus did not show its fine structure and became globular. In thecytosol, the number of ribosomes decreased by 25 ± 14 % (P < 0.05 afterStudent's test) compared to the mature cells. Most of the ribosomes accumulatedtogether as groups, probably as polyribosomes (Fig. 2). Vacuoles appeared asmany small ones or as an enlarged central one. However, in some sections,vacuoles were not evident due to presence of other organelles. 2. Middle stage ofthe senescence; amorphous electron dense material was deposited at mitochon-drial membrane, nuclear envelope and tonoplasts (Fig. 3). The ribosomes were nolonger evident. The tonoplasts disappeared partly. 3. Late stage of the senescence;mitochondrial cristae disappeared to a great extent or completely, whereas the ERwas more dilated but maintained the membrane structure (Fig. 4). Meanwhile,nuclear membranes, especially the inner ones, partly disappeared. The tonoplastsdisappeared completely. However, the plasma membrane remained intact. 4.Final stage of the senescence; all organelles became unrecognizable (Fig. 5),

Cellular Damage to Phytophthora infestans Treated with Oxadixyl 121
• ••••:'.• • •• ':
' ' l .
•• A
Fig. 4. A hypha in the late stage of senescence. Mitochondrial (m) cristae have disappeared. The ER ismore dilated. Tonoplasts of most vacuoles (v) become degenerated. Only plasmalemma remains
intact. (Control, 20 000 X, at 15 h)
Fig. 5. Collapsed cytoplasm of a haustorium without recognizable organelles. The host cell possessesno visible structure except the remaining rest of the extrahaustorial membranes (arrows). (Control,
20 000 X, at 18 h)

122 JIANG and GROSSMANN
indicating the thorough loss of the compartmentation. The collapsed cytoplasmappeared electron dense and then very electron lucent. It is noteworthy ofmentioning that the host cells containing the senescent haustoria had alreadycollapsed (Fig. 5).
: ; • ; - : : . • ^ ; ; - ; . . ^ • . = • - / • - • • • > ; ; • ^ • ^ • • : . ' i ' ^ ^ . .
' ! " - • ' - " . - " • ' . - ' - . • ! • . " > , / i ' ^ / - " ' , ^ ; ? . • - • • • • . ' ' " • ' " . • . ' - . . ' " • . ; • V > - . • ' : . • • , ' ' • . • - . • • '
\ , - ' , ' : v • • • • . ; ' " • ; • " ; • -• ' • • ' . • ' • ' ' \ - / ' • ' ' l ^ " , . : O W - - ^ ' ' '
: " ' • V / , • . • . - , " r i ; ; ; . ' . • , - • . ; • • . . . • • ' . . . • . . : • . : i . ; • - . > v / . • . .
Fig. 6. Hypha after treatment with oxadixyl. Some electron dense deposits are visible at plasmalemma(arrows), whereas the ER, dictyosome and mitochondria are not changed. (Oxadixyl treatment,
20 000 X, at 15 h)
^" « i " . * " ' . " - i " ' . ' ; • - ' • " " - . ' • ' " ' ' ' ' - • : • * . - • ' ' • ' ' . ' " ' * ' * * ' ' ^ ' * ' * , - * ' ' : • : ' * * . • , . • . * - '
Fig. 7. The electron dense deposits develop extensively from the plasmalemma to some region of thecytoplasm. The plasmalemma (arrows) is located outside the electron dense deposits. Mitochondria(m) have degenerated and are coated by the deposits as well. The ER remains as double-layered linear
structure despite its attachment to some deposits. (Oxadixyl treatment, 20 000 X, at 15 h)
Cellular Damage to Phytophthora infestans Treated with Oxadixyl 123
Appearances of damaged cells of P. infestans after treatment with oxadixyl
By 15 and 18 h after inoculation, the treatment with oxadixyl caused strikingdifferences in the appearance of the fungal cells. In addition to other observationsdemonstrated as fungistatic effects (unpublished), also about 10 % of the thinsections (presented as 570 micrographs) made from the same plant tissue in the
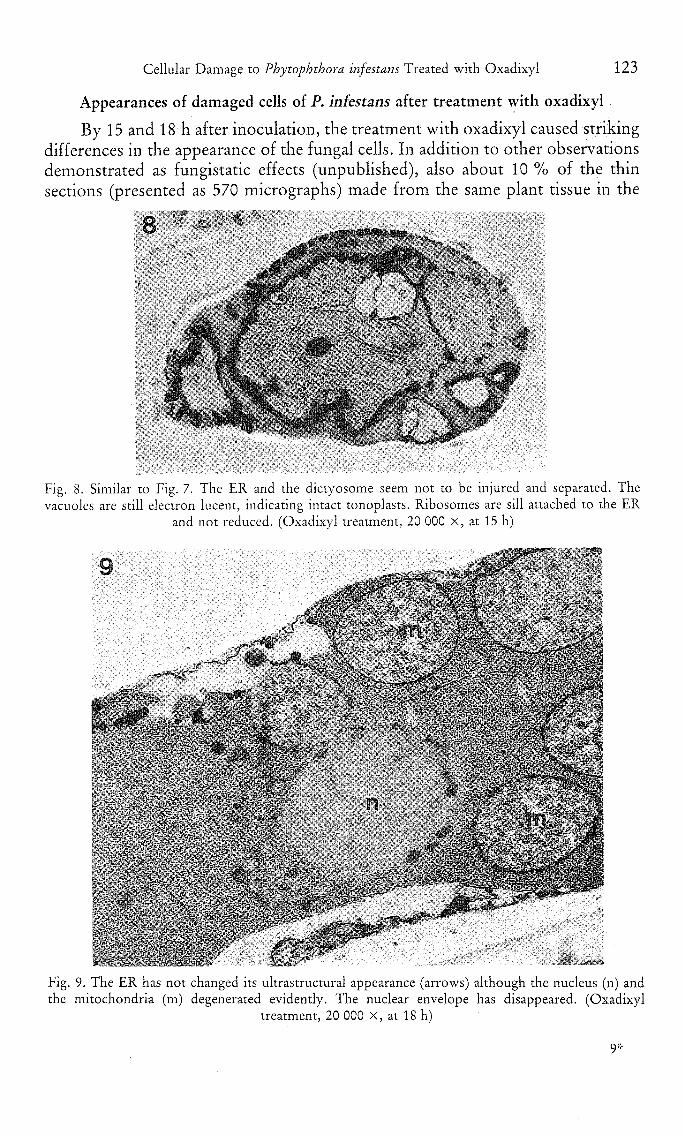
Fig. 8. Similar to Fig. 7. The ER and the dictyosome seem not to be injured and separated. Thevacuoles are still electron lucent, indicating intact tonoplasts. Ribosomes are sill attached to the ER
and not reduced. (Oxadixyl treatment, 20 000 X, at 15 h)
G -k-̂ "̂-'
:C:'^y.:^\.^:>-Z'-':o::
• . \ • •
\^., t. 'f.'-' ',.-VS5»=~
Fig. 9. The ER has not changed its ultrastructural appearance (arrows) although the nucleus (n) andthe mitochondria (m) degenerated evidently. The nuclear envelope has disappeared. (Oxadixyl
treatment, 20 000 X, at 18 h)

124 JIANG and CROSSMANN
oxadixyl treatment, exhibited the following changes which were characterized bythe accumulation of electron dense deposits in the fungal cells. The electron densedeposits occurred near or at the plasma membranes and/or at mitochondrialmembranes (Fig. 6—7). At the same time, however, the ER did not change andthe distribution pattern and number (about 640 ± 81/jdxn^) of ribosomes showedno significant difference from those of the mature cells of the control. Mitochon-dria became degenerated during the time of accumulation of the electron densedeposits (Fig. 7 and 9), with the fine structure of the cristae lost. With the time,the electron dense deposits accumulated massively in the cytosol near the plasmamembrane and extended towards the central area of the cell (Fig. 7—8). Theplasma membrane itself did not become electron dense (Fig. 7), but could becovered by the massive deposits. During the process, changes in the fine structureof the nuclei could not be observed. However, the nuclear membranes dis-appeared and were also in contact with the electron dense deposits (Fig. 9). Thewhole nucleus degenerated. Interestingly, the ER remained still unchanged or lessmodified, even in spite of attachment to some electron dense deposits. Theribosomes were recognizable. Finally, the collapsed cytoplasm was electron denseand/or electron lucent, with the plasmalemma and the ER lost. The phenomenondescribed here occurred occasionally also in small haustoria, together with thedegenerated mother hyphae. In addition, degenerated haustoria were evidentlypresent in intact host cells (Fig. 10).
Fig. 10. A degenerated haustorium with the unchanged ER and collapsed mitochondria (m) present inthe host cell which shows the intact mitochondrion, tonoplasts and chloroplasts. (Oxadixyl treatment,
20 000 X, at 18 h)

Cellular Damage to Phytophthora infestans Treated with Oxadixyl 125
Fig. 11. Thickened cell walls of a hypha. The cytoplasm is filled with some vacuoles. (Oxadixyltreatment, 7000 X, at 18 h)
In some intercellular hyphae, cell walls were thickened considerably(Fig. 11). These cells generally showed the seemingly fragmented but not dilatedER with ribosomes. In addition, vacuolation of the cells was also very conspicu-ous. However, electron dense deposits were seldom observed. It seemed that the
\
• - ' . . • , ' :
Fig. 12. Some cytoplasmic vesicles are fusing with the plasmalemma of the hypha (arrows), probablyleading to the thickening of the cell walls. Vacuoles are evident in the cytoplasm. (Oxadixyl treatment,
20 000 X, at 18 h)

126 JIANG and GROSSMANN
thickening of the cell walls was due to the accumulation of the cytoplasmic,probably dictyosome, vesicles near the cell walls and their fusion with theplasmalemma (Fig. 12).
Discussion
The results showed that in both oxadixyl treatment and the untreated controlthere were appearances of damaged cell organelles of P. infestans due to thefungicide treatment and the physiological senescence, respectively. Hence, it isnecessary to clarify the differences between the mature and senescent cells in thecontrol and the cells affected by the fungicide to draw conclusions here.
The senescent process occurring in P. infestans is mainly timed as reductionin ribosomes, dilation of the ER, degeneration of mitochondria and plas-maiemma. Changes in ribosomes and ER may reflected increased activity ofribonucleases and proteolysis (MATILE 1971, GAHAN 1981). It seems likely thatchanges in the ER and appearance of the electron dense deposits are ultrastruc-tural markers of the senescence, whereas the degeneration of mitochondria is themarker in the senescent process leading to death, as described in irreversibleevents occurring in necrosis of animal cells (WYLLIE 1981). Degeneration ofmitochondria and instability of the mitochondrial genome are closely correlatedwith senescence of fungi (MUNKRES 1985). In many cases, the dead cells of higherplants are finally characterized by irreversible plasmolysis, disruption of cellularmembranes with a concomitant loss of compartmentation, loss of respiratoryactivity etc. (GAHAN 1981). Occurrence of one of these features indicates that thecell is dead. Evidently, the degenerated mitochondria may have no respiratoryactivity and present the irreversible "switch on" of the step in cell senescenceleading to cell death. Thus, any change in the cells after mitochondrial degenera-tion could only be a post-mortem event. It was considered that loss of compart-mentation of cells can be an indicator of the moment of cell death (MATILE 1975).But it is more likely that cell death has occurred before the loss of compartmenta-tion (GAHAN 1981). This seems to be the case here, i.e., the cell death might occurafter degeneration of mitochondria but before disappearance of the plasmalemma,especially in treatment with oxadixyl.
The genetically programmed cell senescence (MUNKRES 1985, 1987) and celldeath of P. infestans seems to be strikingly different from the cellular damagecaused by treatment with oxadixyl. Although the ultrastructural appearance ofthe fungus in oxadixyl treatment is similar to that of the untreated control at thelate stage, i.e., mitochondrial degeneration and degeneration of plasmalemma, theER, the possible marker of the senescence, surprisingly remains unchanged evenafter mitochondrial degeneration. The most notable difference lies in the exten-sive occurrence of the electron dense deposits and their location at plasmalemma.The deposits here are similar to the cytoplasmic densities observed in this andother fungi after treatment with other fungicides (MULLER and BuRTH 1983,HOCH and SYKOLNIK 1979). Presumably, the deposits adjacent to the plas-malemma and in mitochondria could be denatured proteins (WYLLIE 1981) whichwere formed m involvement of the fungal plasmalemma and mitochondrialmembranes.

Cellular Damage to Phytophthora infestans Treated with Oxadixyl 127
It is not known whether the occurrence of the cell damage of the type asdescribed above only in a minority of the fungal cells (10 % in all thin sections) isdue to a relatively high concentration of the fungicide encountered by the fungusin vivo or to the high sensitivity of these fungal cells. The latter seems to be morepossible in that different appearances of the fungus such as cell wall thickeningand fungitoxic phenomena occur in different hyphae located in the same planttissue. In vivo and in vitro, many oomycetous fungi show different sensitivity tometalaxyl in the same species (COFFEY and BOWER 1984, COFFEY et al. 1984, COOK
and ZHANG 1985, HERZOG and SCHOEPP 1985, HUNGER et al. 1982, SHEW 1984),dependent on the growth environment (FULLER and Gisi 1985). Additionally,with P. infestans, isolates are frequently found that are resistant to metalaxyl invitro and yet highly sensitive to it in vivo (COHEN and COFFEY 1986). This couldbe due to the induction of plant resistance by metalaxyl (BORNER et al. 1981,CAHILL and WARD 1989). However, with oxadixyl, another possibility seems tobe the fungitoxic action on some hyphae which become sensitive to oxadixyl afterinfection of the host plants. Further investigations are needed to support thesuggestion.
The work was supported by the Deutsche Gesellschaft fiir Technische Zusammenarbeit. Mrs.G. MOLL developed part of the photographs.
Literature
BORNER, H., G. SCHATZ, and H. GRISEBACH, 1983: Influence of the systemic fungicide metalaxyl onglyceollin accumulation in soybean infected with Phytophthora megasperma f. sp. glycinea.Physiol. Plant Pathol. 23, 145—152.
CAHILL, D . M. , and E. W. B. WARD, 1989: Effects of metalaxyl on elicitor activity, stimulation ofglyceollin production and growth of sensitive and tolerant isolates of Phytophthora mega-sperma f. sp. glycinea. Physiol. Mol. Plant Pathol. 17, 259—267.
COEFEY, M. D., and L. A. BowER, 1984: In vitro variability among isolates of six Phytophthoraspecies in response to metalaxyl. Phytopathology 74, 502—506.
, L. J. KLURE, and L. A. BOWER, 1984: Variability in sensitivity to metalaxyl of isolates ofPhytophthora cinnamomi and P. citricola. Phytopathology 74, 417—422.
COHEN, Y., and M. D. COFFEY, 1986: Systemic fungicides and the control of Oomycetes. Ann. Rev.Phytopath. 24, 311—338.
COOK, R. J., and B. X. ZHANG, 1985: Degrees of sensitivity within the Pythium spp. pathogenic towheat in the Pacific Northwest. Plant Dis. 69, 686—688.
DAVIDSE, L. C , O . C . M. GERRITSMA, J. IDELER, K. PIE, and G. G. M. VELTHUIS, 1988: Antifungalmodes of action of metalaxyl, cyprofuram, benalaxyl and oxadixyl in phenylamide-sensitiveand phenylamide-resistant strains of Phytophthora megasperma f. sp. medicaginis and Phy-tophthora infestans. Crop Protection 7, 347—355.
FULLER, M. S., and U. GiSI, 1985: Comparative study of the in vitro activity of the fungicidesoxadixyl and metalaxyl. Mycologia 77, 424—432.
GAHAN, P. B., 1981: Cell senescence and death in plants. In: BOWEN, I. D., and R. A. LOGKSHIN(eds). Cell death in biology and pathology, pp. 145—169. Chapman and Hall, London.
GiSI, U., H. BINDER, and E. RIMBAGH, 1985: Synergistic interactions of fungicides with differentmodes of action. Trans. Br. Mycol. Soc. 85, 299—306.
GROHMANN, U. , und G. M. HOFFMANN, 1982: Licht und elektronenoptische Untersuchungen zurWirkung von Metalaxyl bei Pythium- und Phytophthora-Anen. Z. PflKrankh. PflSchutz 89,435—446.

128 JIANG and GROSSMANN, Cellular Damage to Phytophthora infestans Treated with Oxadixyl
HERZOG, J., and H. SGHUEPP, 1985: Three types of sensitivity to metalaxyl in Plasmopara viticola.Phytopathol. Z. 114, 90—93.
HOCH, H . C , and M. SYKOLNIK, 1979: Viability of Venturia inaequalis in chlorotic flecks resultingfrom fungicide application to infected Malus leaves. Phytopathology 69, 456—462.
HUNGER, R. M., P. B. HAMM, C. E. HORNER, and E. M. HANSEN, 1982: Tolerance oi Phytophthoramegasperma isolates to metalaxyl. Plant Dis. 66, 645—649.
MATILE, P., 1975: The lyxic compartment of plant cells. Springer Verlag, Wien, New York., and F. WiNKENBACH, 1971: Function of lysosomes and lysosomal enzymes in the senescing
corolla of the morning glory (Ipomea purpurea). J. Exp. Bot. 22, 759—771.MULLER, H . M., und U. BURTH, 1983: Veranderungen der Ultrastruktur in Hyphen von Phyto-
phthora infestans (Mont.) de Bary durch 2-Chlor-6-methyl-4-benzylphenol. Arch. Phy-topath. PflSchutz 19, 357—359.
MUNKRES, K. D. , 1985: Aging of fungi. Rev. Biol. Res. Aging 2, 29—43., 1987: Aging of fungi. Rev. Biol. Res. Aging 3, 41—50.
SHEW, H . D. , 1984: In vitro growth response oi Phytophthora parasitica var. nicotianae to metalaxylexposure. Plant Dis. 68, 764—766.
SHIMONY, C , and J. FRIEND, 1975: Ultrastructure of the interaction between Phytophthora infestansand leaves of two cultivars of potato {Solanum tuberusum L.) Orion and Majestic. NewPhytol. 74, 59—65.
WYLLIE, A. H., 1981: Cell death: a new classification separating apoptosis from necrosis. In: BowEN,I. D., and R. A. LOCKSHIN (eds). Cell death in biology and pathology, pp. 1—33. Chapmanand Hall, London.
