ccr5 blockade modulates inflammation and alloimmunity in
TRANSCRIPT

of November 27, 2018.This information is current as
Alloimmunity in PrimatesCCR5 Blockade Modulates Inflammation and
DeMartinoMiller, Irwin I. Singer, Agnes M. Azimzadeh and Julie A. Mudgett, Kathy Lyons, Sander G. Mills, Geraldine G.Siciliano, Susan Iliff, Julia M. Ayala, Min Lu, John S. Guosheng Wu, Tianshu Zhang, Martin S. Springer, Sal J.Nguyen, Douglas W. Kawka, Laurence B. Peterson, Carsten Schröder, Richard N. Pierson III, Bao-Ngoc H.
http://www.jimmunol.org/content/179/4/2289doi: 10.4049/jimmunol.179.4.2289
2007; 179:2289-2299; ;J Immunol
Referenceshttp://www.jimmunol.org/content/179/4/2289.full#ref-list-1
, 17 of which you can access for free at: cites 51 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

CCR5 Blockade Modulates Inflammation and Alloimmunityin Primates1
Carsten Schroder,2*† Richard N. Pierson III,2,3*† Bao-Ngoc H. Nguyen,2*† Douglas W. Kawka,‡
Laurence B. Peterson,‡ Guosheng Wu,*† Tianshu Zhang,*† Martin S. Springer,‡
Sal J. Siciliano,‡ Susan Iliff,‡ Julia M. Ayala,‡ Min Lu,‡ John S. Mudgett,‡ Kathy Lyons,‡
Sander G. Mills,‡ Geraldine G. Miller,§ Irwin I. Singer,2‡ Agnes M. Azimzadeh,*†
and Julie A. DeMartino2‡
Pharmacologic antagonism of CCR5, a chemokine receptor expressed on macrophages and activated T cells, is an effectiveantiviral therapy in patients with macrophage-tropic HIV infection, but its efficacy in modulating inflammation and immunity isonly just beginning to be investigated. In this regard, the recruitment of CCR5-bearing cells into clinical allografts is a hallmarkof acute rejection and may anticipate chronic rejection, whereas conventionally immunosuppressed renal transplant patientshomozygous for a nonfunctional �32 CCR5 receptor rarely exhibit late graft loss. Therefore, we explored the effects of a potent,highly selective CCR5 antagonist, Merck’s compound 167 (CMPD 167), in an established cynomolgus monkey cardiac allograftmodel. Although perioperative stress responses (fever, diminished activity) and the recruitment of CCR5-bearing leukocytes intothe graft were markedly attenuated, anti-CCR5 monotherapy only marginally prolonged allograft survival. In contrast, relativeto cyclosporine A monotherapy, CMPD 167 with cyclosporine A delayed alloantibody production, suppressed cardiac allograftvasculopathy, and tended to further prolong graft survival. CCR5 therefore represents an attractive therapeutic target forattenuating postsurgical stress responses and favorably modulating pathogenic alloimmunity in primates, including man. TheJournal of Immunology, 2007, 179: 2289–2299.
U ntil we learn how to achieve tolerance, progress in hearttransplantation will depend on better control of patho-genic influences on the donor heart. Chemotactic cyto-
kines or “chemokines” play a critical role in activation of innateimmunity (1), ischemia/reperfusion injury (2), and induction ofadaptive immune responses (3). Because each of these pathways isassociated with important injury in transplanted organs, chemo-kines and their receptors are attractive candidates for new inter-ventions to protect allografts from immunologic injury and for
induction of tolerance (4). The current inflammation paradigmholds that chemokine receptors afford homing and activation sig-nals critical to the migration and function of lymphohemopoieticcells in a variety of circumstances, including embryonic develop-ment, growth, and inflammation (1, 5, 6). Despite promiscuityamong the many possible chemokine ligand-receptor pairs and“cross-talk” between different receptors on individual cells, a fewspecific chemokine receptors, particularly CCR1, CCR2, CCR5,and CXCR3, have emerged in murine models as promising targetsto prevent allograft injury and to attenuate innate immune vascularinjury (7–11).
CCR5 is best known as a major coreceptor for macrophage-tropic human immunodeficiency viruses. Its natural ligands in-clude RANTES (CCL5), MIP-1� and MIP-1� (CCL3 and 4), andMCP-2 (CCL8). CCR5 is expressed on the surfaces of residenttissue monocytes and dendritic cells and on activated T cells, mac-rophages, and NK cells in both lymphoid and nonlymphoid tissues(5, 6). Human peripheral blood monocytes and T cells exhibit lowlevels of CCR5, but expression is increased several weeks after invitro stimulation or in vivo in the setting of established Th1-typeinflammation (8), and particularly on monocytes and activated/memory Th1 lymphocytes. Because these cells are capable of en-tering sites of inflammation and executing pathogenic effectorfunctions, blockade of CCR5 might be expected to prevent injuryto the graft by CCR5� helper or effector T cell subsets or by otherimportant immunocyte populations (dendritic cells, macrophages)that express CCR5.
The absence of functional CCR5 is well tolerated in humanshomozygotic for the CCR5�32 mutation (12), suggesting that“normal CCR5 function is well compensated or redundant” (5).Similarly, CCR5 knockouts and mice treated with CCR5 inhibitorsexhibit only subtly diminished stress response to endotoxin and
*Division of Cardiac Surgery, Department of Surgery, University of Maryland andBaltimore Veterans Administration Medical Center, Baltimore, MD 21201; †Depart-ment of Cardiothoracic Surgery, Vanderbilt University and Nashville Veterans Ad-ministration Medical Center, Nashville, TN 37232; ‡Merck Research Laboratories,Rahway, NJ 07065; and §Division of Infectious Disease, Department of Medicine,Vanderbilt University and Nashville Veterans Administration Medical Center, Nash-ville, TN 37232
Received for publication June 15, 2006. Accepted for publication June 1, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was presented in part orally and in abstract form at the American Trans-plant Congress and the International Society for Heart and Lung Transplantationannual meetings 2002–2006. This work was supported by a Merck Research Labo-ratory sponsored research agreement and gift, National Institutes of Health Grant U01AI 066719, and an American Heart Association Award (0455722U) (to R.N.P.); Na-tional Institutes of Health Grant F32 HL079818 and Thoracic Surgery Foundation forResearch and Education resident research awards (to B.-N.H.N.); Deutsche For-schungsgemeinschaft research award (to C.S.); and an Other Tobacco Related Dis-eases research grant from the Maryland Cigarette Restitution Fund Program.2 C.S. and R.N.P. contributed equally to the first authorship of this study. C.S. andB.-N.H.N. were primarily responsible for the conduct of the in vivo studies, and C.S.made the cardinal observation regarding the effect of CCR5 inhibition on the febrileresponse. In conjunction with J.A.D. and I.I.S., R.N.P. was primarily responsible forstudy design and manuscript authorship.3 Address correspondence and reprint requests to Dr. Richard N. Pierson III, Divisionof Cardiac Surgery, University of Maryland, 22 South Greene Street, Room N4W94,Baltimore, MD 21201. E-mail address: [email protected]
The Journal of Immunology
www.jimmunol.org
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
preserved or enhanced Th1 and Th2 responses (13). Together,these data suggest that CCR5 inhibition is not likely to be associ-ated with other clinically important immune deficits (in contrast toCCR2; for example, see Ref. 14). Based on promising rodent trans-plant data (10, 15–18) and these clinical considerations, this study wasundertaken to investigate the role of CCR5 in primate cardiac allograftrejection using compound 167 (CMPD 167 (N-[(1R,3S,4S)-3-((4-(3-benzyl-1-ethylpyrazol-5-yl)piperidin-1-yl)methyl)-4-(3-fluorophenyl)cyclopentan-1-yl]-N-methyl-D-valine).4 This compound is anallosteric inhibitor of CCR5 that has similar affinity for the G pro-tein-coupled and uncoupled states of the receptor (M. S. Springerand P. E. Finke, manuscript in preparation). Consistent with thisprofile, CMPD 167 blocks the binding of all known ligands ofCCR5, including chemokines, R-5 tropic gp120 envelope proteins(including YU2), and nonpeptidyl antagonist ligands with similaraffinities ranging from 0.05 to 2.5 nM. In addition, it binds withhigh affinity to the CCR5 receptor expressed on human PBMCs.The compound has similar affinities for the human, chimpanzee,rhesus monkey, and cynomolgus monkey, suppresses chemokineligand-induced CCR5–mediated functions (G protein-dependentcalcium signaling, chemotaxis, and MAPK (ERK1/2) activation)
over a broad range of physiologically achievable serum concen-trations, and prevents infection by SIV in macaques (19, 20). Ourinitial observations in this regard have also led to additional pre-liminary efforts to elucidate a potential role for CCR5 in the no-ciceptive and febrile response to surgical stress in the cynomolgusmonkey heart transplant model.
Materials and MethodsCMPD 167
Merck’s CMPD 167 is a lead compound selected for exquisitely specifichigh-affinity binding to human and cynomolgus monkey CCR5 (21, 22).The structure, as well as the pharmacokinetic properties of this proprietaryacidic cyclopentyl derivative in preclinical species, has been published(World Intellectual Property Organization patent WO 00/76972, Decem-ber, 2000; A1; 69). CMPD 167 was administered in a twice daily (bid; bisin die) dosing paradigm via a tether jacket system implanted on the day oftransplant to maintain trough levels of �100 nM, its IC90 value in wholemonkey blood.
Cynomolgus monkeys
Wild-caught cynomolgus monkeys (Macaca fascicularis) were obtainedfrom Coulston Foundation (Alamogordo, NM) and Charles River Labora-tories. All procedures were approved by the institutional animal care anduse committees of Vanderbilt University (Nashville, TN), University ofMaryland (Baltimore, MD), and Merck and were conducted in compliancewith National Institutes of Health guidelines for the care and use of labo-ratory animals (23). Males weighing 3.8 to 4.3 kg, were selected as organrecipients of AB blood type-compatible, MHC class II-mismatched (stim-ulation index: mean 7.7, median 6.2, range 3.1–26) donor hearts (24, 25).Donor-recipient pairs were determined so as to maximize MLR responsewithin groups of blood type-compatible animals.
4 Abbreviations used in this paper: CMPD 167, compound 167 (N-[(1R,3S,4S)-3-((4-(3-benzyl-1-ethylpyrazol-5-yl)piperidin-1-yl)methyl)-4-(3-fluorophenyl)cyclopentan-1-yl]-N-methyl-D-valine; bid, twice daily (bis in die); CAV, cardio allograft vascu-lopathy; CsA, cyclosporine; ISHLT, International Society of Heart and LungTransplantation.
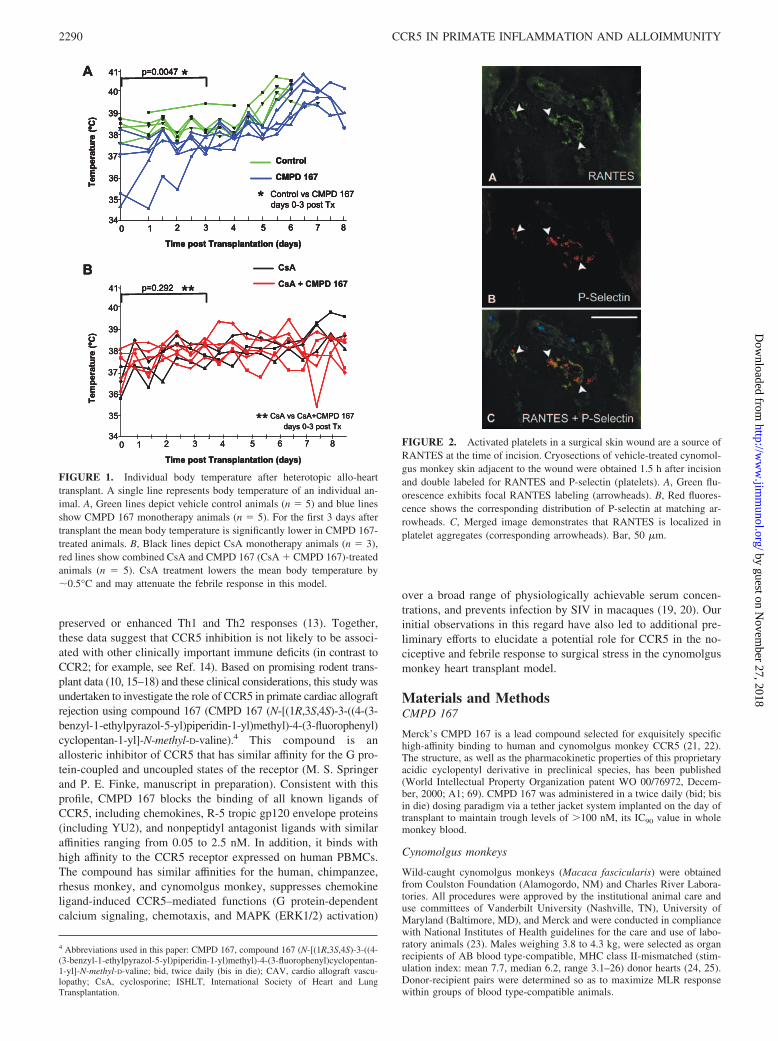
FIGURE 1. Individual body temperature after heterotopic allo-hearttransplant. A single line represents body temperature of an individual an-imal. A, Green lines depict vehicle control animals (n � 5) and blue linesshow CMPD 167 monotherapy animals (n � 5). For the first 3 days aftertransplant the mean body temperature is significantly lower in CMPD 167-treated animals. B, Black lines depict CsA monotherapy animals (n � 3),red lines show combined CsA and CMPD 167 (CsA � CMPD 167)-treatedanimals (n � 5). CsA treatment lowers the mean body temperature by�0.5°C and may attenuate the febrile response in this model.
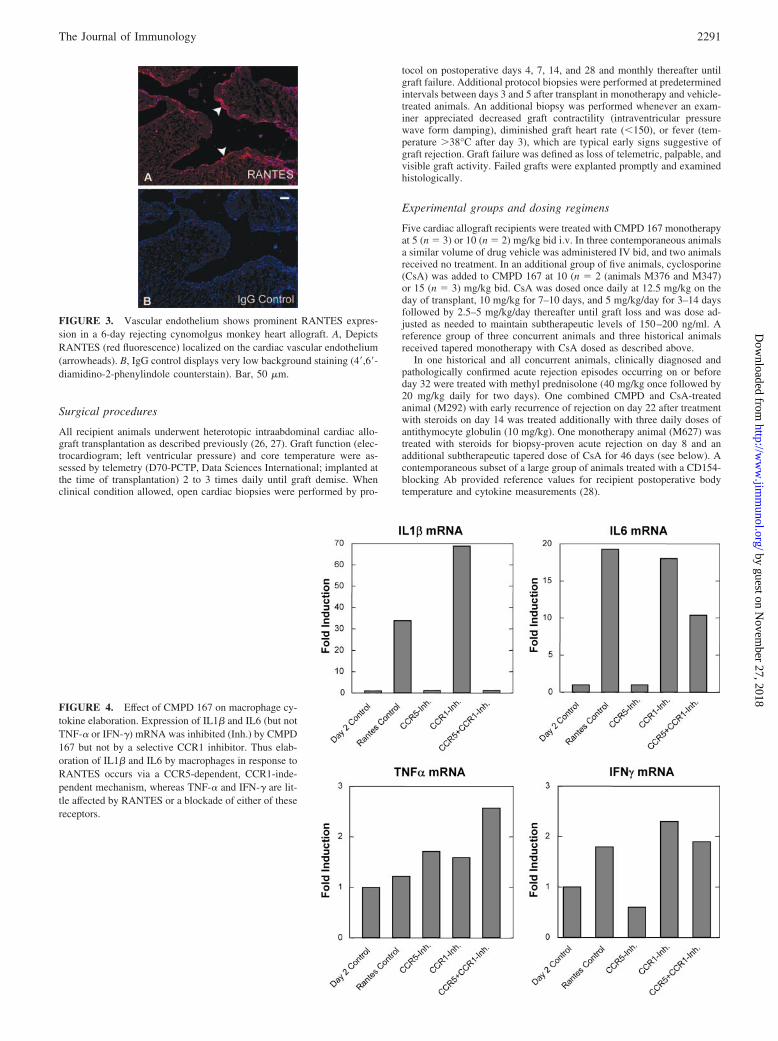
FIGURE 2. Activated platelets in a surgical skin wound are a source ofRANTES at the time of incision. Cryosections of vehicle-treated cynomol-gus monkey skin adjacent to the wound were obtained 1.5 h after incisionand double labeled for RANTES and P-selectin (platelets). A, Green flu-orescence exhibits focal RANTES labeling (arrowheads). B, Red fluores-cence shows the corresponding distribution of P-selectin at matching ar-rowheads. C, Merged image demonstrates that RANTES is localized inplatelet aggregates (corresponding arrowheads). Bar, 50 �m.
2290 CCR5 IN PRIMATE INFLAMMATION AND ALLOIMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Surgical procedures
All recipient animals underwent heterotopic intraabdominal cardiac allo-graft transplantation as described previously (26, 27). Graft function (elec-trocardiogram; left ventricular pressure) and core temperature were as-sessed by telemetry (D70-PCTP, Data Sciences International; implanted atthe time of transplantation) 2 to 3 times daily until graft demise. Whenclinical condition allowed, open cardiac biopsies were performed by pro-
tocol on postoperative days 4, 7, 14, and 28 and monthly thereafter untilgraft failure. Additional protocol biopsies were performed at predeterminedintervals between days 3 and 5 after transplant in monotherapy and vehicle-treated animals. An additional biopsy was performed whenever an exam-iner appreciated decreased graft contractility (intraventricular pressurewave form damping), diminished graft heart rate (�150), or fever (tem-perature �38°C after day 3), which are typical early signs suggestive ofgraft rejection. Graft failure was defined as loss of telemetric, palpable, andvisible graft activity. Failed grafts were explanted promptly and examinedhistologically.
Experimental groups and dosing regimens
Five cardiac allograft recipients were treated with CMPD 167 monotherapyat 5 (n � 3) or 10 (n � 2) mg/kg bid i.v. In three contemporaneous animalsa similar volume of drug vehicle was administered IV bid, and two animalsreceived no treatment. In an additional group of five animals, cyclosporine(CsA) was added to CMPD 167 at 10 (n � 2 (animals M376 and M347)or 15 (n � 3) mg/kg bid. CsA was dosed once daily at 12.5 mg/kg on theday of transplant, 10 mg/kg for 7–10 days, and 5 mg/kg/day for 3–14 daysfollowed by 2.5–5 mg/kg/day thereafter until graft loss and was dose ad-justed as needed to maintain subtherapeutic levels of 150–200 ng/ml. Areference group of three concurrent animals and three historical animalsreceived tapered monotherapy with CsA dosed as described above.
In one historical and all concurrent animals, clinically diagnosed andpathologically confirmed acute rejection episodes occurring on or beforeday 32 were treated with methyl prednisolone (40 mg/kg once followed by20 mg/kg daily for two days). One combined CMPD and CsA-treatedanimal (M292) with early recurrence of rejection on day 22 after treatmentwith steroids on day 14 was treated additionally with three daily doses ofantithymocyte globulin (10 mg/kg). One monotherapy animal (M627) wastreated with steroids for biopsy-proven acute rejection on day 8 and anadditional subtherapeutic tapered dose of CsA for 46 days (see below). Acontemporaneous subset of a large group of animals treated with a CD154-blocking Ab provided reference values for recipient postoperative bodytemperature and cytokine measurements (28).
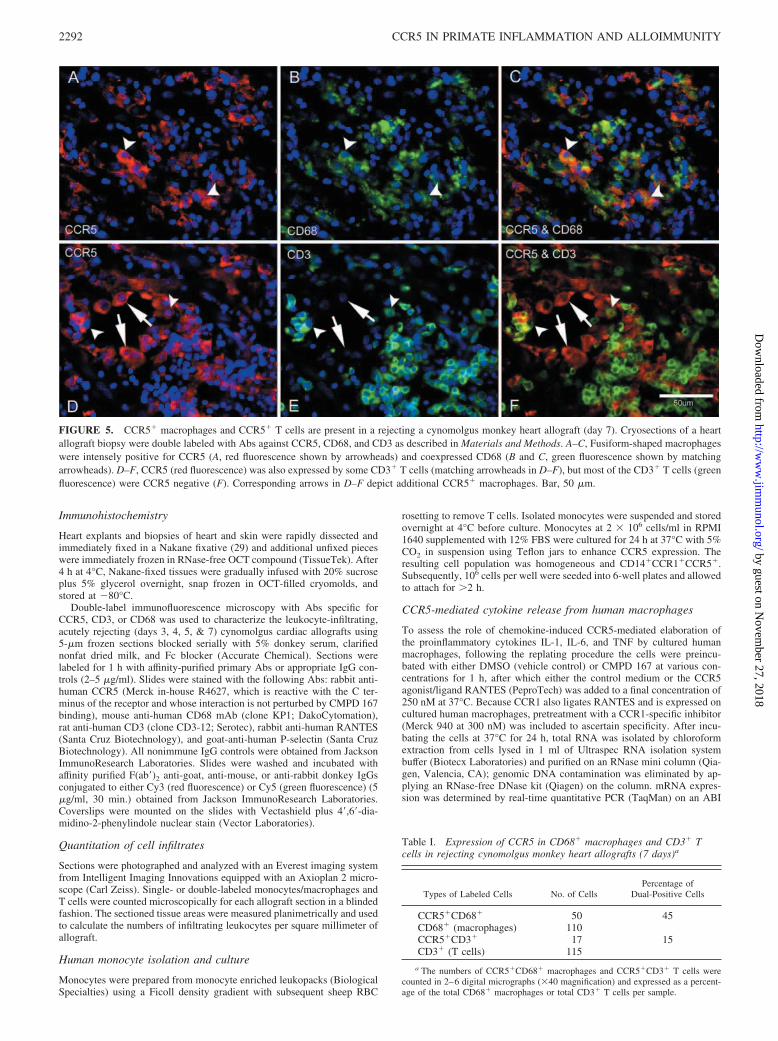
FIGURE 3. Vascular endothelium shows prominent RANTES expres-sion in a 6-day rejecting cynomolgus monkey heart allograft. A, DepictsRANTES (red fluorescence) localized on the cardiac vascular endothelium(arrowheads). B, IgG control displays very low background staining (4�,6�-diamidino-2-phenylindole counterstain). Bar, 50 �m.
FIGURE 4. Effect of CMPD 167 on macrophage cy-tokine elaboration. Expression of IL1� and IL6 (but notTNF-� or IFN-�) mRNA was inhibited (Inh.) by CMPD167 but not by a selective CCR1 inhibitor. Thus elab-oration of IL1� and IL6 by macrophages in response toRANTES occurs via a CCR5-dependent, CCR1-inde-pendent mechanism, whereas TNF-� and IFN-� are lit-tle affected by RANTES or a blockade of either of thesereceptors.
2291The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Immunohistochemistry
Heart explants and biopsies of heart and skin were rapidly dissected andimmediately fixed in a Nakane fixative (29) and additional unfixed pieceswere immediately frozen in RNase-free OCT compound (TissueTek). After4 h at 4°C, Nakane-fixed tissues were gradually infused with 20% sucroseplus 5% glycerol overnight, snap frozen in OCT-filled cryomolds, andstored at �80°C.
Double-label immunofluorescence microscopy with Abs specific forCCR5, CD3, or CD68 was used to characterize the leukocyte-infiltrating,acutely rejecting (days 3, 4, 5, & 7) cynomolgus cardiac allografts using5-�m frozen sections blocked serially with 5% donkey serum, clarifiednonfat dried milk, and Fc blocker (Accurate Chemical). Sections werelabeled for 1 h with affinity-purified primary Abs or appropriate IgG con-trols (2–5 �g/ml). Slides were stained with the following Abs: rabbit anti-human CCR5 (Merck in-house R4627, which is reactive with the C ter-minus of the receptor and whose interaction is not perturbed by CMPD 167binding), mouse anti-human CD68 mAb (clone KP1; DakoCytomation),rat anti-human CD3 (clone CD3-12; Serotec), rabbit anti-human RANTES(Santa Cruz Biotechnology), and goat-anti-human P-selectin (Santa CruzBiotechnology). All nonimmune IgG controls were obtained from JacksonImmunoResearch Laboratories. Slides were washed and incubated withaffinity purified F(ab�)2 anti-goat, anti-mouse, or anti-rabbit donkey IgGsconjugated to either Cy3 (red fluorescence) or Cy5 (green fluorescence) (5�g/ml, 30 min.) obtained from Jackson ImmunoResearch Laboratories.Coverslips were mounted on the slides with Vectashield plus 4�,6�-dia-midino-2-phenylindole nuclear stain (Vector Laboratories).
Quantitation of cell infiltrates
Sections were photographed and analyzed with an Everest imaging systemfrom Intelligent Imaging Innovations equipped with an Axioplan 2 micro-scope (Carl Zeiss). Single- or double-labeled monocytes/macrophages andT cells were counted microscopically for each allograft section in a blindedfashion. The sectioned tissue areas were measured planimetrically and usedto calculate the numbers of infiltrating leukocytes per square millimeter ofallograft.
Human monocyte isolation and culture
Monocytes were prepared from monocyte enriched leukopacks (BiologicalSpecialties) using a Ficoll density gradient with subsequent sheep RBC
rosetting to remove T cells. Isolated monocytes were suspended and storedovernight at 4°C before culture. Monocytes at 2 106 cells/ml in RPMI1640 supplemented with 12% FBS were cultured for 24 h at 37°C with 5%CO2 in suspension using Teflon jars to enhance CCR5 expression. Theresulting cell population was homogeneous and CD14�CCR1�CCR5�.Subsequently, 106 cells per well were seeded into 6-well plates and allowedto attach for �2 h.
CCR5-mediated cytokine release from human macrophages
To assess the role of chemokine-induced CCR5-mediated elaboration ofthe proinflammatory cytokines IL-1, IL-6, and TNF by cultured humanmacrophages, following the replating procedure the cells were preincu-bated with either DMSO (vehicle control) or CMPD 167 at various con-centrations for 1 h, after which either the control medium or the CCR5agonist/ligand RANTES (PeproTech) was added to a final concentration of250 nM at 37°C. Because CCR1 also ligates RANTES and is expressed oncultured human macrophages, pretreatment with a CCR1-specific inhibitor(Merck 940 at 300 nM) was included to ascertain specificity. After incu-bating the cells at 37°C for 24 h, total RNA was isolated by chloroformextraction from cells lysed in 1 ml of Ultraspec RNA isolation systembuffer (Biotecx Laboratories) and purified on an RNase mini column (Qia-gen, Valencia, CA); genomic DNA contamination was eliminated by ap-plying an RNase-free DNase kit (Qiagen) on the column. mRNA expres-sion was determined by real-time quantitative PCR (TaqMan) on an ABI
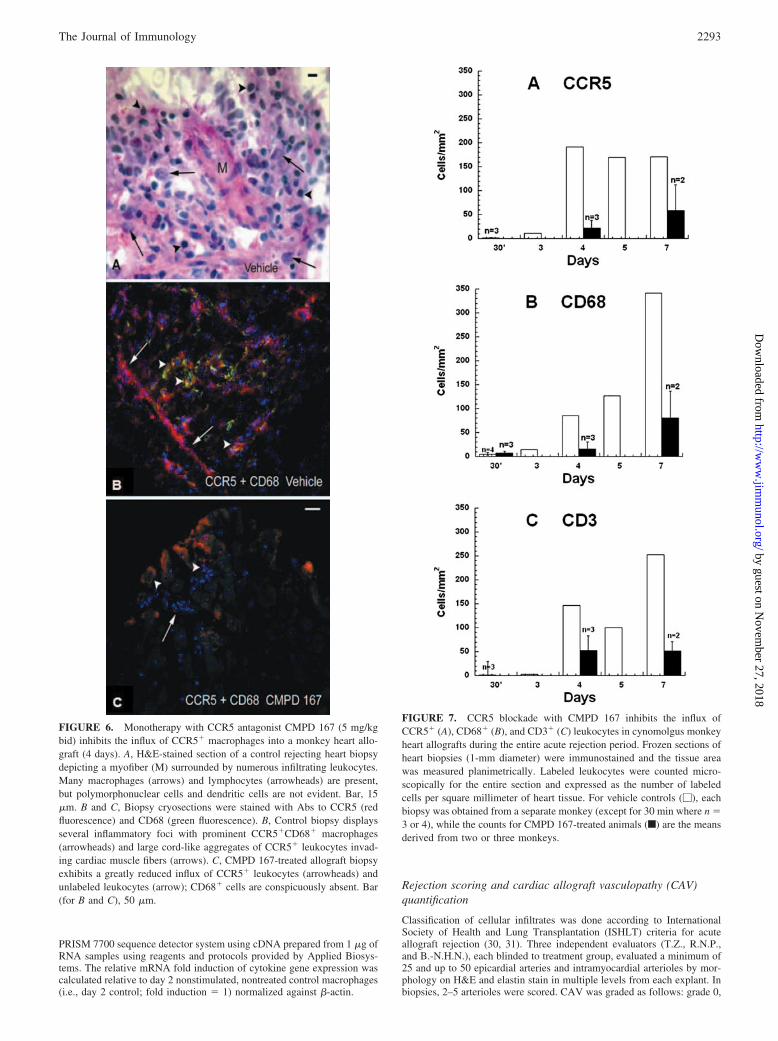
FIGURE 5. CCR5� macrophages and CCR5� T cells are present in a rejecting a cynomolgus monkey heart allograft (day 7). Cryosections of a heartallograft biopsy were double labeled with Abs against CCR5, CD68, and CD3 as described in Materials and Methods. A–C, Fusiform-shaped macrophageswere intensely positive for CCR5 (A, red fluorescence shown by arrowheads) and coexpressed CD68 (B and C, green fluorescence shown by matchingarrowheads). D–F, CCR5 (red fluorescence) was also expressed by some CD3� T cells (matching arrowheads in D–F), but most of the CD3� T cells (greenfluorescence) were CCR5 negative (F). Corresponding arrows in D–F depict additional CCR5� macrophages. Bar, 50 �m.
Table I. Expression of CCR5 in CD68� macrophages and CD3� Tcells in rejecting cynomolgus monkey heart allografts (7 days)a
Types of Labeled Cells No. of CellsPercentage of
Dual-Positive Cells
CCR5�CD68� 50 45CD68� (macrophages) 110CCR5�CD3� 17 15CD3� (T cells) 115
a The numbers of CCR5�CD68� macrophages and CCR5�CD3� T cells werecounted in 2–6 digital micrographs (40 magnification) and expressed as a percent-age of the total CD68� macrophages or total CD3� T cells per sample.
2292 CCR5 IN PRIMATE INFLAMMATION AND ALLOIMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
PRISM 7700 sequence detector system using cDNA prepared from 1 �g ofRNA samples using reagents and protocols provided by Applied Biosys-tems. The relative mRNA fold induction of cytokine gene expression wascalculated relative to day 2 nonstimulated, nontreated control macrophages(i.e., day 2 control; fold induction � 1) normalized against �-actin.
Rejection scoring and cardiac allograft vasculopathy (CAV)quantification
Classification of cellular infiltrates was done according to InternationalSociety of Health and Lung Transplantation (ISHLT) criteria for acuteallograft rejection (30, 31). Three independent evaluators (T.Z., R.N.P.,and B.-N.H.N.), each blinded to treatment group, evaluated a minimum of25 and up to 50 epicardial arteries and intramyocardial arterioles by mor-phology on H&E and elastin stain in multiple levels from each explant. Inbiopsies, 2–5 arterioles were scored. CAV was graded as follows: grade 0,
FIGURE 6. Monotherapy with CCR5 antagonist CMPD 167 (5 mg/kgbid) inhibits the influx of CCR5� macrophages into a monkey heart allo-graft (4 days). A, H&E-stained section of a control rejecting heart biopsydepicting a myofiber (M) surrounded by numerous infiltrating leukocytes.Many macrophages (arrows) and lymphocytes (arrowheads) are present,but polymorphonuclear cells and dendritic cells are not evident. Bar, 15�m. B and C, Biopsy cryosections were stained with Abs to CCR5 (redfluorescence) and CD68 (green fluorescence). B, Control biopsy displaysseveral inflammatory foci with prominent CCR5�CD68� macrophages(arrowheads) and large cord-like aggregates of CCR5� leukocytes invad-ing cardiac muscle fibers (arrows). C, CMPD 167-treated allograft biopsyexhibits a greatly reduced influx of CCR5� leukocytes (arrowheads) andunlabeled leukocytes (arrow); CD68� cells are conspicuously absent. Bar(for B and C), 50 �m.
FIGURE 7. CCR5 blockade with CMPD 167 inhibits the influx ofCCR5� (A), CD68� (B), and CD3� (C) leukocytes in cynomolgus monkeyheart allografts during the entire acute rejection period. Frozen sections ofheart biopsies (1-mm diameter) were immunostained and the tissue areawas measured planimetrically. Labeled leukocytes were counted micro-scopically for the entire section and expressed as the number of labeledcells per square millimeter of heart tissue. For vehicle controls (�), eachbiopsy was obtained from a separate monkey (except for 30 min where n �3 or 4), while the counts for CMPD 167-treated animals (f) are the meansderived from two or three monkeys.
2293The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
normal arterial morphology; grade 1, activated endothelial cells with en-larged nuclei and/or adherent leukocytes and without luminal narrowing(�10%); grade 2, distinct neointimal thickening and luminal narrowing�50%; grade 3, extensive neointimal proliferation with �50% luminalocclusion (for illustrative histology see Fig. 9a) Individual CAV scoreswere tabulated for each explant, and mean and median scores were calcu-lated. After unblinding, the median of the three final CAV measurementswas accepted as the score for that sample. In four instances where discrep-ancies occurred (scoring range between observers of �0.5), the score wasadjudicated by pathologic review (B.-N.H.N. and R.N.P.).
Alloantibody assay
Recipient serum specimens were collected weekly during the first month,biweekly for 2 months, and monthly thereafter until graft rejection andstored at �80°C until use. Alloantibodies were measured retrospectivelyby flow cytometry using frozen donor splenocytes as described previously(32). Results were expressed as the percentage of positive T cells (definedas CD3�DR�) as compared with recipient pretransplant serum. Donor se-rum and archived historical samples were used as internal negative andpositive controls, respectively. Two animals treated with CsA were notevaluated because viable donor cells were not available.
Statistical analysis
Unless otherwise mentioned, all data are presented as means and SD for allvariables. Continuous variables were checked for normality by plottinghistograms. Variables that were not normally distributed were analyzedusing the Kruskal-Wallis test and the Mann-Whitney U test. Those thatwere normally distributed were assessed with a one-way ANOVA andStudent’s t test. A �2 test for trend was used for scoring variables and theirchanges over time. Survival statistics were calculated by the Kaplan-Meiermethod using the log-rank test for significance analysis. Correlations be-tween parameters were studied using the Pearson or Spearman correlationtest. p values of �0.05 were considered statistically significant. All testswere two tailed. All statistical analyses were performed on a personal com-puter with the statistical package SPSS for Windows (version 13.0) exceptfor the power analyses, which were performed using the shareware PS:
Power and Sample Size Calculations, version 2.1 (W. D. Dupont and W. D.Plummer, Vanderbilt University).
ResultsCCR5 inhibition blunts the febrile response
The average temperature over the first 3 days after surgery in an-imals treated with CMPD 167 (37.3 0.5°C, n � 5) was unex-pectedly �1°C lower than in vehicle-treated or untreated animals(38.5 0.4°C, n � 5, p � 0.0047; Fig. 1) or animals treated witha mAb directed at CD154 (IDEC-131, 38.4 0.7, n � 4, p � 0.05;data not shown), and 0.3°C below CsA-treated animals (n � 5, p �NS). As independently documented by the veterinary animal caretechnicians and veterinarians, animals treated with CMPD 167were more active and resilient following multiple surgical proce-dures within the first 2 wk (transplant, protocol biopsies on days 4and 7, and graft explant) than were animals in any other treatmentgroup.
Inhibition of CCR5 could potentially blunt fever by interferingwith the proximal triggers of innate immune activation in thewound or the revascularized graft by inhibiting the release of as-sociated proinflammatory cytokines or by modulating the responseof constitutively CCR5� cells to these cytokines. To explorewhether CCR5 ligands are expressed locally in the wound and thusmight initiate the fever consistently seen in these animals, we eval-uated RANTES expression at the surgical site. As shown in Fig. 2,RANTES expression is prominent in the skin incision 90 min afterwounding. Similar to inflamed and atherosclerotic endothelia (33),RANTES is associated with aggregated CD62P� platelets that ap-pear sequestered at the edge of the surgical wound as they are in
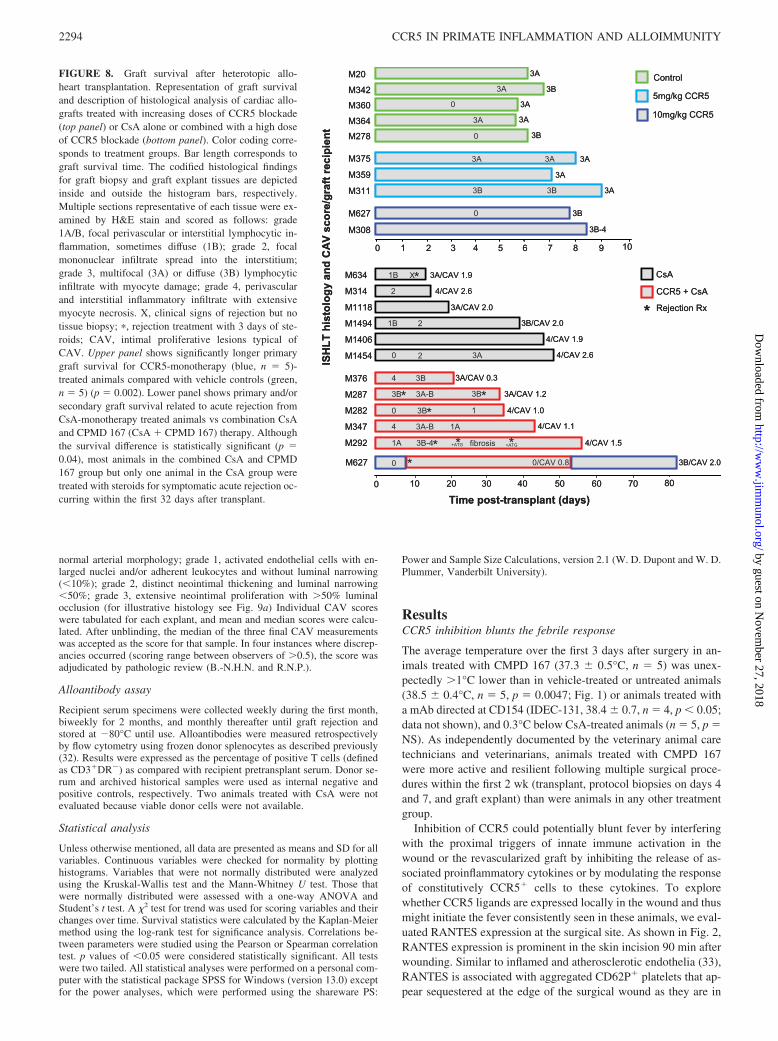
FIGURE 8. Graft survival after heterotopic allo-heart transplantation. Representation of graft survivaland description of histological analysis of cardiac allo-grafts treated with increasing doses of CCR5 blockade(top panel) or CsA alone or combined with a high doseof CCR5 blockade (bottom panel). Color coding corre-sponds to treatment groups. Bar length corresponds tograft survival time. The codified histological findingsfor graft biopsy and graft explant tissues are depictedinside and outside the histogram bars, respectively.Multiple sections representative of each tissue were ex-amined by H&E stain and scored as follows: grade1A/B, focal perivascular or interstitial lymphocytic in-flammation, sometimes diffuse (1B); grade 2, focalmononuclear infiltrate spread into the interstitium;grade 3, multifocal (3A) or diffuse (3B) lymphocyticinfiltrate with myocyte damage; grade 4, perivascularand interstitial inflammatory infiltrate with extensivemyocyte necrosis. X, clinical signs of rejection but notissue biopsy; �, rejection treatment with 3 days of ste-roids; CAV, intimal proliferative lesions typical ofCAV. Upper panel shows significantly longer primarygraft survival for CCR5-monotherapy (blue, n � 5)-treated animals compared with vehicle controls (green,n � 5) (p � 0.002). Lower panel shows primary and/orsecondary graft survival related to acute rejection fromCsA-monotherapy treated animals vs combination CsAand CPMD 167 (CsA � CPMD 167) therapy. Althoughthe survival difference is statistically significant (p �0.04), most animals in the combined CsA and CPMD167 group but only one animal in the CsA group weretreated with steroids for symptomatic acute rejection oc-curring within the first 32 days after transplant.
2294 CCR5 IN PRIMATE INFLAMMATION AND ALLOIMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
the rejecting graft. Intense RANTES expression, presumably aris-ing from platelets surrounding skin incisions shortly after wound-ing and on the vascular endothelia of rejecting cardiac allografts(Fig. 3), strongly implies that RANTES plays a role in leukocytechemotaxis in these settings.
We next assessed the effect of CMPD 167 on macrophage cy-tokine elaboration during in vitro stimulation of CCR5�CCR1�
cultured human monocytes with RANTES and found that expres-sion of IL1� and IL6 (but not TNF-� or IFN-�) mRNA was in-hibited by a CCR5-dependent, CCR1-independent mechanism(Fig. 4). Because suppression of fever was an unanticipated findingin our preliminary studies, serum samples for reference levels ofinflammatory markers were not systematically obtained at clini-cally relevant intervals (2–6 and 12–36 h after surgery) in mostCMPD 167 monotherapy-, vehicle-, or CsA monotherapy-treatedanimals. However, initial findings from the blood of a small subsetof the experimental subjects reported here indicate that IL1�, IL6,IL8, C-reactive protein, and soluble CD154 elaborations on day 1after transplant or at the time of graft demise are highly variablebetween individual animals (data not shown) and suggest that avery large number of observations (�10–25 per group) would berequired to determine whether serum cytokine levels are signifi-cantly different between groups of CMPD-167-treated animals and
relevant reference populations. Sample sizes of this magnitude areimpractical in this model.
CCR5 modulates graft leukocyte infiltration
We defined the kinetics of leukocyte infiltration into cardiac allo-grafts and studied the efficacy of CMPD 167 to block this immi-gration. Intense multifocal perivascular and interstitial infiltrationof leukocytes (lymphocytes, macrophages, and monocytes) intothe graft reliably occurs by days 4–7 in untreated animals (Fig. 5).The greatest CCR5 expression was observed on CD68� macro-phages (45%) with a minority of graft-infiltrating CCR5� T cells(15%) (Fig. 5 and Table I). These data suggest a prominent role forCCR5�CD68� leukocytes in cardiac allograft rejection. His-topathological analysis of rejecting control biopsies shows a prom-inent leukocytic infiltrate consisting of macrophages and lympho-cytes without polymorphonuclear cells or dendritic cells (Fig. 6A).In animals treated with CMPD 167, infiltration of CCR5�,CD68�, and CD3� leukocytes was inhibited in association withCCR5 blockade on days 4 and 7 and at rejection (Figs. 6 and 7)relative to vehicle-treated control animals at similar intervals. Im-portantly, infiltration by CD3� cells was also inhibited relative to
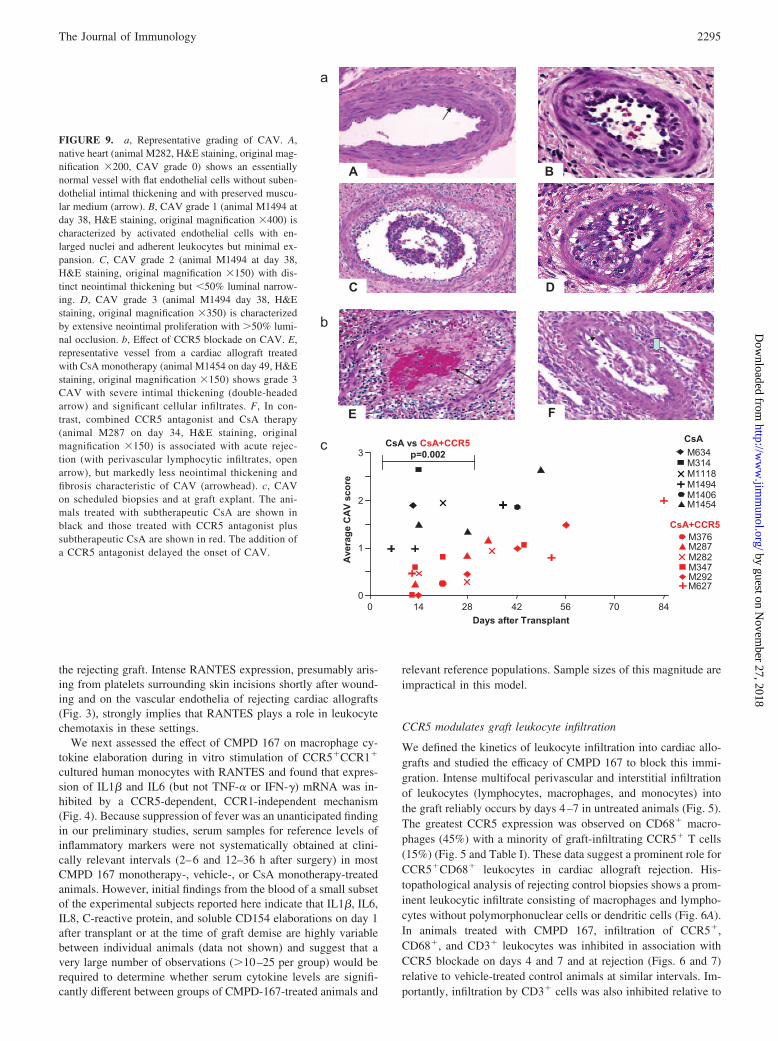
FIGURE 9. a, Representative grading of CAV. A,native heart (animal M282, H&E staining, original mag-nification 200, CAV grade 0) shows an essentiallynormal vessel with flat endothelial cells without suben-dothelial intimal thickening and with preserved muscu-lar medium (arrow). B, CAV grade 1 (animal M1494 atday 38, H&E staining, original magnification 400) ischaracterized by activated endothelial cells with en-larged nuclei and adherent leukocytes but minimal ex-pansion. C, CAV grade 2 (animal M1494 at day 38,H&E staining, original magnification 150) with dis-tinct neointimal thickening but �50% luminal narrow-ing. D, CAV grade 3 (animal M1494 day 38, H&Estaining, original magnification 350) is characterizedby extensive neointimal proliferation with �50% lumi-nal occlusion. b, Effect of CCR5 blockade on CAV. E,representative vessel from a cardiac allograft treatedwith CsA monotherapy (animal M1454 on day 49, H&Estaining, original magnification 150) shows grade 3CAV with severe intimal thickening (double-headedarrow) and significant cellular infiltrates. F, In con-trast, combined CCR5 antagonist and CsA therapy(animal M287 on day 34, H&E staining, originalmagnification 150) is associated with acute rejec-tion (with perivascular lymphocytic infiltrates, openarrow), but markedly less neointimal thickening andfibrosis characteristic of CAV (arrowhead). c, CAVon scheduled biopsies and at graft explant. The ani-mals treated with subtherapeutic CsA are shown inblack and those treated with CCR5 antagonist plussubtherapeutic CsA are shown in red. The addition ofa CCR5 antagonist delayed the onset of CAV.
2295The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
vehicle-treated controls on days 4 and 7 (Fig. 7C) despite the ab-sence of detectable CCR5 expression on 85% of T cells found inuntreated grafts.
Anti-CCR5 monotherapy prolongs allograft survival
Five cynomolgus monkeys treated with CMPD 167 monotherapyat 5 (n � 3) or 10 mg/kg/bid (n � 2) exhibited significantly pro-longed mean graft survival with monotherapy (8.1 days 0.4SEM, chemotaxis index 7.4–8.9), relative to 6.4 0.2 (confidenceinterval, 6.1–6.6, p � 0.002) for five vehicle-treated or untreatedanimals (Fig. 8, top panel). Rejected grafts in all groups exhibiteda typical intense myocardial infiltrate rich in lymphocytes, mono-cytes, and macrophages, including CCR5� cells, although (asnoted above) the proportion of CCR5� cells was lower in CMPD167-treated animals at rejection than in vehicle-treated referenceanimals. Thus, CCR5� cells are capable of graft infiltration duringCCR5 inhibition as used here, and acute allograft rejection canoccur despite apparently efficient CCR5 antagonism.
In one case (animal M627), the rejecting CMPD 167 mono-therapy graft was successfully treated with “rescue” steroids ini-tiated on day 8. The addition of cyclosporine from day 8 to 54 was
associated with survival for 85 days, 31 days after cyclosporinewithdrawal.
Synergy studies: acute cellular rejection
Twelve animals were treated with CsA dosed to achieve subthera-peutic CsA levels (100–300 ng/ml in our cynomolgus monkeyheart model) 10–14 days after transplant, 11 as primary therapyand one after acute rejection treatment on day 8. Mean time to thefirst symptomatic acute rejection episode was 30 7 days (days11, 15, 21, 39, 46, and 49) with subtherapeutic CsA compared with�20 6 days (days 7, 14, 16, �21, and 44) with CsA plus CMPD167 ( p � 0.32) (Fig. 8, bottom panel). Two early acute rejectionepisodes in the CsA group (days 15 and 21) were not treated withsteroids and were associated with graft failure as expected. Acuterejection episodes occurring in two CsA plus CMPD 167 animalswithin the first month were successfully treated with steroids. OneCsA monotherapy was rejected on day 13 despite steroid treatmenton day 11, and one CsA plus CMPD 167 animal exhibited recur-rent rejection on day 25 (M292), 1 wk after steroid therapy, thatresponded to antithymocyte globulin. Two animals treated usingcombined CMPD 167 and CsA were sacrificed on days 21 (as per
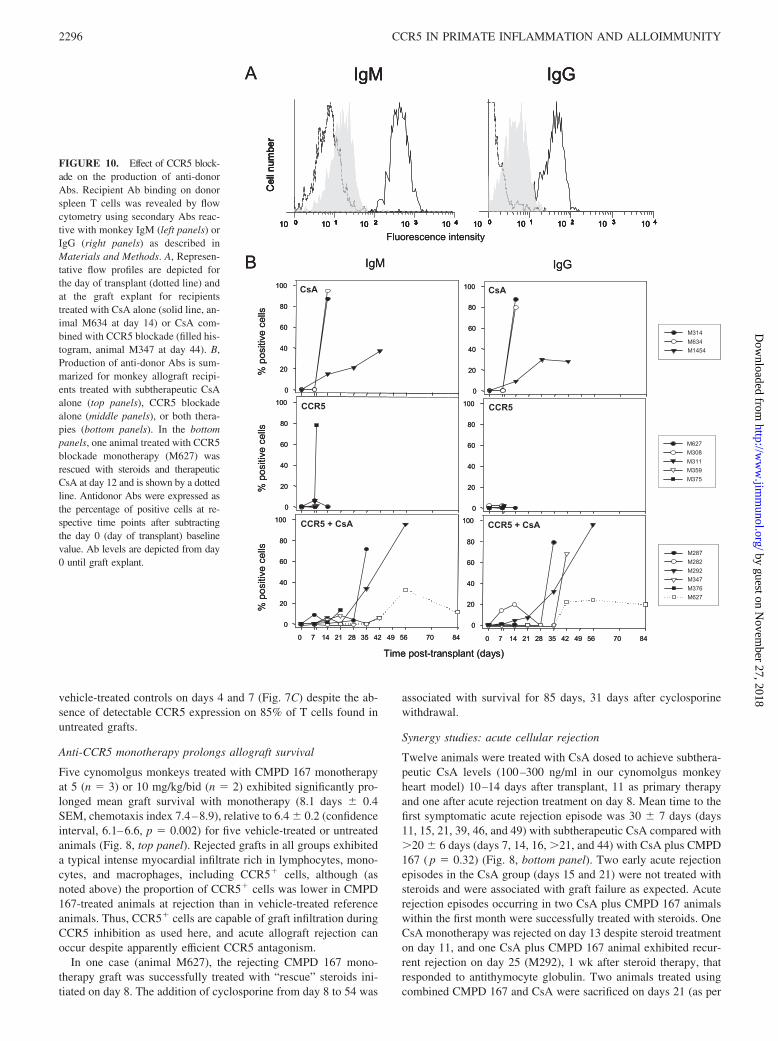
FIGURE 10. Effect of CCR5 block-ade on the production of anti-donorAbs. Recipient Ab binding on donorspleen T cells was revealed by flowcytometry using secondary Abs reac-tive with monkey IgM (left panels) orIgG (right panels) as described inMaterials and Methods. A, Represen-tative flow profiles are depicted forthe day of transplant (dotted line) andat the graft explant for recipientstreated with CsA alone (solid line, an-imal M634 at day 14) or CsA com-bined with CCR5 blockade (filled his-togram, animal M347 at day 44). B,Production of anti-donor Abs is sum-marized for monkey allograft recipi-ents treated with subtherapeutic CsAalone (top panels), CCR5 blockadealone (middle panels), or both thera-pies (bottom panels). In the bottompanels, one animal treated with CCR5blockade monotherapy (M627) wasrescued with steroids and therapeuticCsA at day 12 and is shown by a dottedline. Antidonor Abs were expressed asthe percentage of positive cells at re-spective time points after subtractingthe day 0 (day of transplant) baselinevalue. Ab levels are depicted from day0 until graft explant.
2296 CCR5 IN PRIMATE INFLAMMATION AND ALLOIMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the IACUC protocol for �10% weight loss despite normal activityand food intake) and 35 (aspiration) with normal graft functionwithout exhibiting acute rejection. Overall, spontaneously resolv-ing asymptomatic (biopsies with ISHLT score �1) or symptomatic(fever, bradycardia, or diminished graft contractility) acute rejec-tion episodes occurred at similar intervals and with similar fre-quencies in both groups.
Effect of CCR5 antagonism on CAV
CAV scores from day 0–28 in the CsA group (1.6 0.6, n � 7)were significantly higher than with combined CMPD and CsA(0.4 0.3, n � 11, p � 0.002) (Fig. 9C). Upon histology atexplant, although all rejected grafts had moderate to severe cellularinfiltrates (ISHLT grades 3A to 4), only mild to moderate or mildCAV was observed with combined CMPD and CsA, even in thethree longest-surviving grafts (animal M347, CAV score 1.1;M292, CAV score 1.5; 627, CAV score 0.8 on day 54) as illus-trated in Fig. 9C. This result stands in distinct contrast to graftssurviving for similar intervals in animals treated with CsA, with orwithout steroid rescue therapy, in which CAV is relatively severe(�2) in most animals.
Effect of CCR5 antagonism on anti-donor Ab
We reasoned that CCR5 might attenuate CAV by inhibiting adap-tive immunity to alloantigens and by delaying elaboration of thealloantibody. Thus, anti-donor Ab levels were measured serially inthree vehicle-treated animals, three animals treated with CsAmonotherapy, five animals treated with CMPD 167 monotherapy,five animals with CsA plus CMPD 167, and animal M627 (delayedaddition of CsA after steroid rescue). Animals treated with CsAalone or with vehicle exhibited high levels of antidonor IgM andIgG Ab within 3 wk (Fig. 10). In contrast, anti-donor Ab wasabsent or present at low titers at 3 wk in animals treated withCMPD 167 alone. Alloantibody elaboration and class switching(from IgM to IgG) was significantly delayed with combined CsAand CMPD 167 relative to CsA alone.
DiscussionCCR5 inhibition using CMPD 167 monotherapy modestly but sig-nificantly delayed the onset of acute cellular rejection and pro-longed graft survival, confirming in primates observations initiallymade in rodents (10, 17, 18, 34, 35). However, the effect of de-laying infiltration of CCR5� cells and attenuating acute allograftinjury, while qualitatively similar to what is seen rodents (10, 18),is quantitatively much less efficacious in protecting the allograftfrom lethal injury. Although incomplete receptor blockade byCMPD 167 cannot be excluded based on our study, in our esti-mation it is unlikely. Trough drug levels in treated animals wereconsistently at or above the in vitro whole blood IC90, and infil-tration of the allograft by CCR5� cells was significantly inhibitedeven at the time of graft failure. We conclude that, as expected,other chemokines and receptors can mediate graft infiltration inprimates despite CCR5 blockade to a degree that is clinically moreimportant than in rodents. We infer that CCR5 blockade as mono-therapy or with subtherapeutic conventional immunosuppression isunlikely to attenuate acute rejection in humans.
Although CCR5 was expressed in 45% of graft-infiltratingmonocytes and macrophages but was only detected in 15% ofgraft-associated T cells, the influx of T cells was significantly in-hibited in CMPD-treated animals. Whether CCR5 blockade atten-uates acute cellular rejection kinetics by a direct effect on CCR5�
monocytes or by indirect effects on T cells that enter the graftremains to be determined. Despite its salutary effect on earlymonocyte infiltration, the early febrile response, and late CAV,
CCR5 inhibition in conjunction with CsA is associated with anincidence of steroid-responsive acute rejection at least as high asthat in the CsA-treated reference group, although allograft survivalis prolonged. Alloantibody elaboration is delayed, an observationanticipated by some rodent studies (36), but not prevented. Ourdata leave open the possibility that a CCR5 blockade exerts com-peting effects on alloimmunity, both delaying infiltration of patho-genic CCR5� (and CCR5�) immunocytes into the graft and in-terfering with the emergence or function of regulatory pathways,because CCR5 is expressed on regulatory T cells and plasmacytoiddendritic cells (37–40). If so, clinical application could be targetedto the first days of the transplant episode to inhibit surgical stressresponses and, after stable graft function is established, to sup-press CAV after donor-specific T regulatory cells have becomeestablished.
That CCR5 modulates inflammation (fever) and malaise occur-ring in the context of surgical stress is a novel observation withpotentially important implications for the clinical care of patientsundergoing cardiac surgery or with infection or trauma. The mech-anisms causing fever remain incompletely understood (41, 42),complicating our efforts to explain our observations. The currentparadigm holds that fever is mediated centrally by hypothalamicPGE1 elaboration. Because CMPD 167 poorly penetrates the CNS,we presume that this agent does not directly modulate prostanoid-driven signaling in the brain. Increased activity and appetite on thefirst days after surgery in CMPD 167-treated monkeys are consis-tent with an anti-nociceptive effect on CCR5� postganglionic neu-rons, but this mechanism, if operative, would not likely directlyaccount for the antipyrexial effect associated with CCR5 antago-nism. CsA treatment partially attenuated the febrile response, aneffect perhaps ascribable to the blunting of cyclophilin-mediated,NFAT- and/or NF-�B-dependent cytokine pathways. In contrast,CsA monotherapy had no effect on appetite or malaise, suggestingthat these features of the CCR5 blockade phenotype are distinctfrom cyclophilin-driven innate immune responses and perhapsspecific to this pathway.
To explore how CCR5 might modulate inflammation, we rea-soned that peripheral events elicited by wounding might trigger theelaboration of CCR5-dependent humoral mediators of inflamma-tion pivotal to fever. In support of this notion, we find platelet-associated RANTES in surgical wounds and show that RANTESamplifies IL-1 and IL-6 mRNA in cultured monocytes by a CCR5-dependent mechanism. Based on these observations, we posit thatCCR5 blockade attenuates fever by inhibiting the amplification ofplatelet-derived RANTES-driven inflammation in the wound,which in turn modulates the elaboration of cytokines by residentCCR5� tissue monocytes and dendritic cells or blood monocytoidpopulations activated by coagulation and the associated inflamma-tion in the wound.
Our observations and derivative hypotheses build upon theevolving paradigm linking innate immunity to the classical alloan-tigen-driven pathogenic adaptive immune response to an allograft.In this study we demonstrate that CCR5 plays an important role ininnate immune responses to surgical stress following transplanta-tion and in CAV. Innate immunity is increasingly recognized as animportant modulator of adaptive responses to donor Ags through“danger” signals (43–48) and in vascular disease (35, 49–52).Whether this effect on innate immunity accounts in part or whollyfor the delay in Ab elaboration and, conversely, how Ag presen-tation and lymphocyte responses are affected by CCR5 blockadeare important remaining questions and the focus of ongoing work.As the broad array of reagents needed to perform informative stud-ies becomes available for the cynomolgus monkey, comprehensiveand pathway-specific gene expression profiling in tissues archived
2297The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

from these studies should shed additional light on the mechanismsinvolved.
We conclude that CCR5 antagonism modulates perioperativeinflammation, delays graft infiltration by monocytes/macrophagesand T cells, and slightly prolongs acute cardiac allograft survivalin cynomolgus monkeys. Combined with CsA, CMPD 167 delays(but does not prevent) alloantibody elaboration and confers signif-icant protection from CAV relative to CsA monotherapy. Attenu-ation of CAV despite failure to prevent intense cellular infiltrates,alloantibody elaboration, and myocyte injury suggests that CCR5plays an important role in vascular remodeling downstream fromalloimmune injury. The number of observations to date is rela-tively small, and ongoing work with this and other CCR5 antag-onists will be required to confirm the specific role of this promisingtherapeutic target in allograft injury and to determine how CCR5affects innate and adaptive immunity (6, 38–40). If efficacy insafely inhibiting chronic rejection is confirmed, CCR5 inhibitioncould substantially improve long-term outcomes after transplanta-tion of the heart and other organs and address a pressing and cur-rently unmet clinical need.
AcknowledgmentsWe acknowledge the important contributions of James Atkinson for grad-ing cardiac rejection scores and Qi Feng, Amal Laaris, Athba Hammed,Hui Zhou, Chris Avon, and Nitin Sangrampurkar for technical assistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Springer, T. A. 1995. Traffic signals on endothelium for lymphocyte recirculation
and leukocyte emigration. Annu. Rev. Physiol. 57: 827–872.2. Flach, R., N. Spieled, S. Flohe, J. Borgermann, I. G. Dresen, J. Erhard, and
F. U. Schade. 1998. Analysis of intragraft cytokine expression during early reper-fusion after liver transplantation using semi-quantitative RT-PCR. Cytokine 10:445–451.
3. Gu, L., S. Tseng, R. M. Horner, C. Tam, M. Loda, and B. J. Rollins. 2000.Control of TH2 polarization by the chemokine monocyte chemoattractant pro-tein-1. Nature 404: 407–411.
4. Hancock, W. W., W. Gao, K. L. Faia, and V. Csizmadia. 2000. Chemokines andtheir receptors in allograft rejection. Curr. Opin. Immunol. 12: 511–516.
5. Murphy, P. M., M. Baggiolini, I. F. Charo, C. A. Hebert, R. Horuk,K. Matsushima, L. H. Miller, J. J. Oppenheim, and C. A. Power. 2000. Interna-tional Union of Pharmacology, XXII: nomenclature for chemokine receptors.Pharmacol. Rev. 52: 145–176.
6. Luster, A. D. 1998. Chemokines: chemotactic cytokines that mediate inflamma-tion. N. Engl. J. Med. 338: 436–445.
7. Foxman, E. F., J. J. Campbell, and E. C. Butcher. 1997. Multistep navigation andthe combinatorial control of leukocyte chemotaxis. J. Cell Biol. 139: 1349–1360.
8. Qin, S., J. B. Rottman, P. Myers, N. Kassam, M. Weinblatt, M. Loetscher,A. E. Koch, B. Moser, and C. R. Mackay. 1998. The chemokine receptorsCXCR3 and CCR5 mark subsets of T cells associated with certain inflammatoryreactions. J. Clin. Invest. 101: 746–754.
9. Miura, M., K. Morita, H. Kobayashi, T. A. Hamilton, M. D. Burdick,R. M. Strieter, and R. L. Fairchild. 2001. Monokine induced by IFN-� is a dom-inant factor directing T cells into murine cardiac allografts during acute rejection.J. Immunol. 167: 3494–3504.
10. Gao, W., P. S. Topham, J. A. King, S. T. Smiley, V. Csizmadia, B. Lu,C. J. Gerard, and W. W. Hancock. 2000. Targeting of the chemokine receptorCCR1 suppresses development of acute and chronic cardiac allograft rejection.J. Clin. Invest. 105: 35–44.
11. Melter, M., A. Exeni, M. I. J. Reinders, J. C. Fang, G. McMahon, P. Ganz,W. W. Hancock, and D. M. Briscoe. 2001. Expression of the chemokine receptorCXCR3 and its ligand IP-10 during human cardiac allograft rejection. Circulation104: 2558–2564.
12. Fischereder, M., B. Luckow, B. Hocher, R. P. Wuthrich, U. Rothenpieler,H. Schneeberger, U. Panzer, R. A. Stahl, I. A. Hauser, K. Budde, et al. 2001. CCchemokine receptor 5 and renal-transplant survival. Lancet 357: 1758–1761.
13. Zhou, Y., T. Kurihara, R. P. Ryseck, Y. Yang, C. Ryan, J. Loy, G. Warr, andR. Bravo. 1998. Impaired macrophage function and enhanced T cell-dependentimmune response in mice lacking CCR5, the mouse homologue of the majorHIV-1 coreceptor. J. Immunol. 160: 4018–4025.
14. Kuziel, W. A., S. J. Morgan, T. C. Dawson, S. Griffin, O. Smithies, K. Ley, andN. Maeda. 1997. Severe reduction in leukocyte adhesion and monocyte extrav-asation in mice deficient in CC chemokine receptor 2. Proc. Natl. Acad. Sci. USA94: 12053–12058.
15. Hancock, W. W., W. Gao, V. Csizmadia, K. L. Faia, N. Shemmeri, andA. D. Luster. 2001. Donor-derived IP-10 initiates development of acute allograftrejection. J. Exp. Med. 193: 975–980.
16. Koga, S., M. B. Aurerbach, T. M. Engeman, A. C. Novick, H. Toma, andR. L. Fairchild. 1999. T cell infiltration into class II MHC-disparate allografts andacute rejection is dependent on the IFN-�-induced chemokine Mig. J. Immunol.163: 4878–4885.
17. Hancock, W. W., B. Lu, B. Gao, V. Csizmadia, K. Faia, J. A. King, S. T. Smiley,M. Ling, N. P. Gerard, and C. Gerard. 2000. Requirement of the chemokinereceptor CXCR3 for acute allograft rejection. J. Exp. Med. 192: 1515–1519.
18. Gao, W., K. L. Zaia, V. Csizmadia, S. T. Smiley, D. Soler, J. King, T. M. Danoff,and W. W. Hancock. 2001. Beneficial effects of targeting CCR5 in allograftrecipients. Transplantation 72: 1199–1205.
19. Veazey, R. S., P. J. Klasse, T. J. Ketas, J. D. Reeves, M. Piatak, Jr., K. Kunstman,S. E. Kuhmann, P. A. Marx, J. D. Lifson, J. Dufour, et al. 2003. Use of a smallmolecule CCR5 inhibitor in macaques to treat simian immunodeficiency virusinfection or prevent simian-human immunodeficiency virus infection. J. Exp.Med. 198: 1551–1562.
20. Wolinsky, S. M., R. S. Veazey, K. J. Kunstman, P. J. Klasse, J. Dufour,A. J. Marozsan, M. S. Springer, and J. P. Moore. 2004. Effect of a CCR5 inhibitoron viral loads in macaques dual-infected with R5 and X4 primate immunodefi-ciency viruses. Virology 328: 19–29.
21. Hale, J. J., R. J. Budhu, S. G. Mills, M. MacCoss, L. Malkowitz, S. Siciliano,S. L. Gould, J. A. DeMartino, and M. S. Springer. 2001. 1,3,4-Trisubstitutedpyrrolidine CCR5 receptor antagonists, part 1: discovery of the pyrrolidine scaf-fold and determination of its stereochemical requirements. Bioorg. Med. Chem.Lett. 11: 1437–1440.
22. Hale, J. J., R. J. Budhu, E. B. Holson, P. E. Finke, B. Oates, S. G. Mills,M. MacCoss, S. L. Gould, J. A. DeMartino, M. S. Springer, et al. 2001. 1,3,4-Trisubstituted pyrrolidine CCR5 receptor antagonists, part 2: lead optimizationaffording selective, orally bioavailable compounds with potent anti-HIV activity.Bioorg. Med. Chem. Lett. 11: 2741–2745.
23. National Institutes of Health, Public Health Service. 1985. National Institutes ofHealth Publication 86-23: Guide for the Care and Use of Laboratory Animals.National Institutes of Health, Bethesda, MD.
24. Socha, W. W., and S. G. Lasano. 1997. Monoclonal antibodies against humanblood group related antigens in tests with non-human primate red cells. Transfus.Clin. Biol. 4: 111–114.
25. Blancher, A., M. E. Reid, and W. W. Socha. 2000. Cross-reactivity of antibodiesto human and primate red cell antigens. Transfus. Med. Rev. 14: 161–179.
26. Pierson, R. N. III, A. C. Chang, M. G. Blum, K. S. Blair, M. A. Scott,J. B. Atkinson, B. J. Collins, J. P. Zhang, D. W. Thomas, L. C. Burkly, andG. G. Miller. 1999. Prolongation of primate cardiac allograft survival by treat-ment with ANTI-CD40 ligand (CD154) antibody. Transplantation 68:1800–1805.
27. Pierson, R. N. III, J. E. Crowe, Jr., S. Pfeiffer, J. Atkinson, A. Azimzadeh, andG. G. Miller. 2001. CD40-ligand in primate cardiac allograft and viral immunity.Immunol. Res. 23: 253–262.
28. Azimzadeh A. M., S. Pfeiffer, G. Wu, C. Schroder, G. L. Zorn, III,S. S. Kelishadi, E. Ozkaynak, M. Kehry, J. B. Atkinson, G. G. Miller, andR. N. Pierson, III. 2006. Alloimmunity in primate heart recipients with CD154blockade: evidence for alternative costimulation mechanisms. Transplantation81: 255–264.
29. McLean, I. W., and P. K. Nakane. 1974. Periodate-lysine-paraformaldehyde fix-ative: a new fixation for immunoelectron microscopy. J. Histochem. Cytochem.22: 1077–1083.
30. Billingham, M. E., N. R. Cary, M. E. Hammond, J. Kemnitz, C. Marboe,H. A. McCallister, D. C. Snovar, G. L. Winters, and A. Zerbe. 1990. A workingformulation for the standardization of nomenclature in the diagnosis of heart andlung rejection: Heart Rejection Study Group. J. Heart Transplant. 6: 587–593.
31. Billingham, M. E. 1992. Histopathology of graft coronary disease. J. Heart LungTransplant. 11: 38–44.
32. Azimzadeh, A. M., S. Pfeiffer, G. S. Wu, C. Schroder, H. Zhou, G. L. Zorn, III,M. Kehry, G. G. Miller, M. L. Rose, and R. N. Pierson, III. 2005. Humoralimmunity to vimentin is associated with cardiac allograft injury in nonhumanprimates. Am. J. Transplant. 5: 2349–2359.
33. von Hundelshausen, P., K. S. Weber, Y. Huo, A. E. Proudfoot, P. J. Nelson,K. Ley, and C. Weber. 2002. RANTES desposition by platelets triggers monocytearrest on inflamed and atherosclerotic endothelium. Circulation 103: 1772–1777.
34. Yun, J. J., M. P. Fischbein, H. Laks, M. C. Fishbein, M. L. Espejo, K. Ebrahimi,Y. Irie, J. Berliner, and A. Ardehali. 2000. Early and late chemokine productioncorrelates with cellular recruitment in cardiac allograft vasculopathy. Transplan-tation 69: 2515–2524.
35. Yun, J. J., M. P. Fischbein, H. Laks, Y. Irie, M. L. Espejo, M. C. Fishbein,J. A. Berliner, and A. Ardehali. 2001. RANTES production during developmentof cardiac allograft vasculopathy. Transplantation 71: 1649–1656.
36. Amano, H., A. Bickerstaff, C. G. Orosz, A. C. Novick, H. Toma, andR. L. Fairchild. 2005. Absence of recipient CCR5 promotes early and increasedallospecific antibody responses to cardiac allografts. J. Immunol. 174:6499–6508.
37. Loetscher, M., B. Gerber, P. Loetscher, S. A. Jones, L. Piali, I. Clark-Lewis,M. Baggiolini, and B. Moser. 1996. Chemokine receptor specific for IP10 andmig: structure, function, and expression in activated T-lymphocytes. J. Exp. Med.184: 963–969.
2298 CCR5 IN PRIMATE INFLAMMATION AND ALLOIMMUNITY
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

38. Sozzni, S., P. Allavena, G. D’Amico, W. Loini, G. Bianchi, M. Kataura, T. Imai,O. Yoshie, R. Bonecchi, and A. Mantovani. 1998. Cutting edge: differential reg-ulation of chemokine receptors during dendritic cell maturation: a model for theirtrafficking properties. J. Immunol. 161: 1083–1086.
39. Sallusto, F., P. Schaerli, P. Loetscher, C. Schaniel, D. Lenig, C. R. Mackay,S. Qin, and A. Lanzavecchia. 1998. Rapid and coordinated switch in chemokinereceptor expression during dendritic cell maturation. Eur. J. Immunol. 28:2760–2769.
40. Wysocki, C. A., Q. Jiang, A. Panoskaltsis-Mortari, P. A. Taylor, K. P. McKinnon,L. Su, B. R. Blazar, and J. S. Serody. 2005. Critical role for CCR5 in the functionof donor CD4�CD25� regulatory T cells during acute graft-versus-host disease.Blood 106: 3300–3307.
41. Roth, J., and G. E. De Souza. 2001. Fever induction pathways: evidence fromresponses to systemic or local cytokine formation. Braz. J. Med. Biol. Res. 34:301–314.
42. Dalai, S., and D. S. Zhukovsky. 2006. Pathophysiology and management of fever.J. Support. Oncol. 4: 9–16.
43. Matzinger, P. 1994. Tolerance, danger, and the extended family. Annu. Rev.Immunol. 12: 991–1045.
44. Gallucci, S., M. Lolkema, and P. Matzinger. 1999. Natural adjuvants: endoge-nous activators of dendritic cells. Nat. Med. 5: 1249–1255.
45. Gallucci, S., and P. Matzinger. 2001. Danger signals: SOS to the immune system.Curr. Opin. Immunol. 13: 114–119.
46. Soares, M. P., Y. Lin, K. Sato, K. M. Stuhlmeier, and F. H. Back. 1999. Ac-commodation. Immunol. Today 20: 434–437.
47. Anderson, C. C., J. M. Carroll, S. Gallucci, J. P. Ridge, A. W. Cheever, andP. Matzinger. 2001. Testing time-, ignorance-, and danger-based models of tol-erance. J. Immunol. 166: 3663–3671.
48. Matzinger, P. 2002. The danger model: a renewed sense of self. Science 296:301–305.
49. Pethig, K., B. Heublein, I. Kutschka, and A. Haverich. 2000. Systemic inflam-matory response in cardiac allograft vasculopathy: high-sensitive C-reactive pro-tein is associated with progressive luminal obstruction. Circulation 102:233–236.
50. Bholasingh, R., J. H. Cornel, O. Kamp, J. P. van Straalen, G. T. Sanders,L. Dijksman, J. G. Tijssen, and R. J. de Winter. 2003. The prognostic value ofmarkers of inflammation in patients with troponin T-negative chest pain beforedischarge from the emergency department. Am. J. Med. 115: 521–528.
51. Kawai, T., M. Seki, K. Hiromatsu, J. W. Eastcott, G. F. Watts, M. Sugai,D. J. Smith, S. A. Porcelli, and M. A. Taubman. 1999. Selective diapedesis ofTh1 cells induced by endothelial cell RANTES. J. Immunol. 163: 3269–3278.
52. Horiguchi, K., S. Kitagawa-Sakakida, Y. Sawa, Z. Z. Li, N. Fukushima,R. Shirakura, and H. Matsuda. 2002. Selective chemokine and receptor geneexpressions in allografts that develop transplant vasculopathy. J. Heart LungTransplant. 21: 1090–1100.
2299The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from