cathepsin g increases mmp expression in normal human fibroblasts through fibronectin fragmentation,...
TRANSCRIPT

Letters to the Editor / Journal of Dermatological Science 53 (2009) 146–165150
Zhi-Rong Lu1
Department of Environmental Health,
School of Public Health and Tropical Medicine,
The Southern Medical University,
Guangzhou 510515, PR China
Tae-Ho Park1
Department of Dermatology, Kangbuk Samsung Hospital,
Seoul, Korea
Eil-Soo Lee
Department of Dermatology,
Sungkyunkwan University School of Medicine,
Samsung Medical Center, Seoul, Korea
Kea-Jeung Kim
Department of Dermatology, Kangbuk Samsung Hospital,
Seoul, Korea
Daeui Park
Byoung-Chul Kim
Seong-Woong Cho
Jong Bhak
Korean BioInformation Center (KOBIC), KRIBB,
Daejeon, Republic of Korea
A R T I C L E I N F O
Keywords:
Fibronectin
Fibronectin fragments
Extracellular matrix
Skin aging
Cathepsin G
Yong-Doo Park
Fei Zou*
Department of Environmental Health,
School of Public Health and Tropical Medicine,
Southern Medical University, Guangzhou 510515, PR China
Jun-Mo Yang**
Department of Dermatology,
Sungkyunkwan University School of Medicine,
Samsung Medical Center, Seoul, Republic of Korea
*Corresponding author. Tel.: +86 2061648301; fax: +86 2061648324
E-mail address: [email protected]
(F. Zou)
**Corresponding author. Tel.: +822 3410 3541; fax: +822 3410 3869
E-mail address: [email protected]
(J.-M. Yang)
1These authors contributed equally to this study.
doi:10.1016/j.jdermsci.2008.08.004
Letter to the Editor
Cathepsin G increases MMP expression in normal humanfibroblasts through fibronectin fragmentation, and induces theconversion of proMMP-1 to active MMP-1
Fibronectin (Fn) is a glycoprotein that is found in the plasmaand is a component of the extracellular matrix (ECM). It binds tocollagen and fibulin, as well as a class of the cell surfaceadhesion receptors termed integrins, and has been shown to beinvolved in various biological activities, including cell attach-ment, cell migration, and wound healing [1]. Fn is readilydegraded into fibronectin fragment (Fn-frs) through the actionof many different types of proteases [2] containing cathepsin G[3]. The expression of MMP (matrix metalloproteinase) by Fn-frs(Fn-f29; N-terminal heparin binding fragment, 45 kDa; Fn-f45,collagen binding fragment, and 110 kDa; Fn-f110 cell bindingfragment) has been studied in chondrocytes [4–6], however,little is known about the roles of Fn-frs, or cathepsin G-mediatedfragmentation of Fn, in aged human skin and normal humanfibroblasts (NHFs).
The purpose of this study was to investigate the relationshipbetween cathepsin G-mediated fragmentation of Fn and theinduction of MMP expression in NHFs and aged human skin.
We examined the expression of Fn in facial and sun-protectedbuttock skin biopsies from four young (men, mean age 33 years, agerange 31–35 years) and six aged (four men, mean age 68 years, agerange 60–75 years; two women, mean age 66 years, age range 64–68years) Koreans by fluorescence immunohistochemistry. Fn localizedto the ECM, in particular, to the dermo–epidermal junctions of theskin samples. The levels of Fn were higher in young skin than in agedskin (Fig. 1a). This pattern of localization of Fn is consistent with theFyrand [7], who reported that Fn is located in the basementmembrane zone in the papillary and the reticular dermis. When weexamined the sun-protected buttock biopsies by Western blot for Fnfragmentation, we found that the 200 and 190 kDa forms of Fn werepresent in young (n = 4), while there were at least 4–5 smallermolecular weight fragments of Fn in aged skin (n = 4) (Fig. 1b).
We also examined the effect of purified Fn-frs (Sigma, St.Louis, MO) on MMP-1 gene and protein expression in NHFs. NHFswere cultured in the presence of 0.5 mM Fn-f45 or -f70 (N-terminal heparin/gelatin binding fragment) for 48 h, and thentotal RNA from treated and untreated cell was subjected to RT-PCR analysis. Fn-f45 and -f70 increased the levels of MMP-1 genetranscription by 130 � 23% (p < 0.01) and 140 � 9.4% (p < 0.01),respectively (Fig. 1c). When we examined cell supernatants byWestern blot, we found that Fn-f45 and -f70 increased MMP-1secretion by 158 � 44% and 206 � 60% (Fig. 1d). To determinewhether Fn-frs regulated MMP-2 and MMP-9 expression, weanalyzed the activity of MMP-2 and MMP-9 by zymography. Whileboth of the Fn-frs increased proMMP-2 activity in NHFs, only Fn-f70increased the level of the active MMP-2 (Fig. 1e). Fn-frs increasedMMP-9 activity of chondrocytes in OA [8] but not NHFs (Fig. 1e).These results suggest that Fn-f45 and -f70 may promote the releaseof MMPs in NHFs.
To determine whether the inhibition of cathepsin G inhibitedFn fragmentation, cathepsin G (1 mU/ml) was added at the same
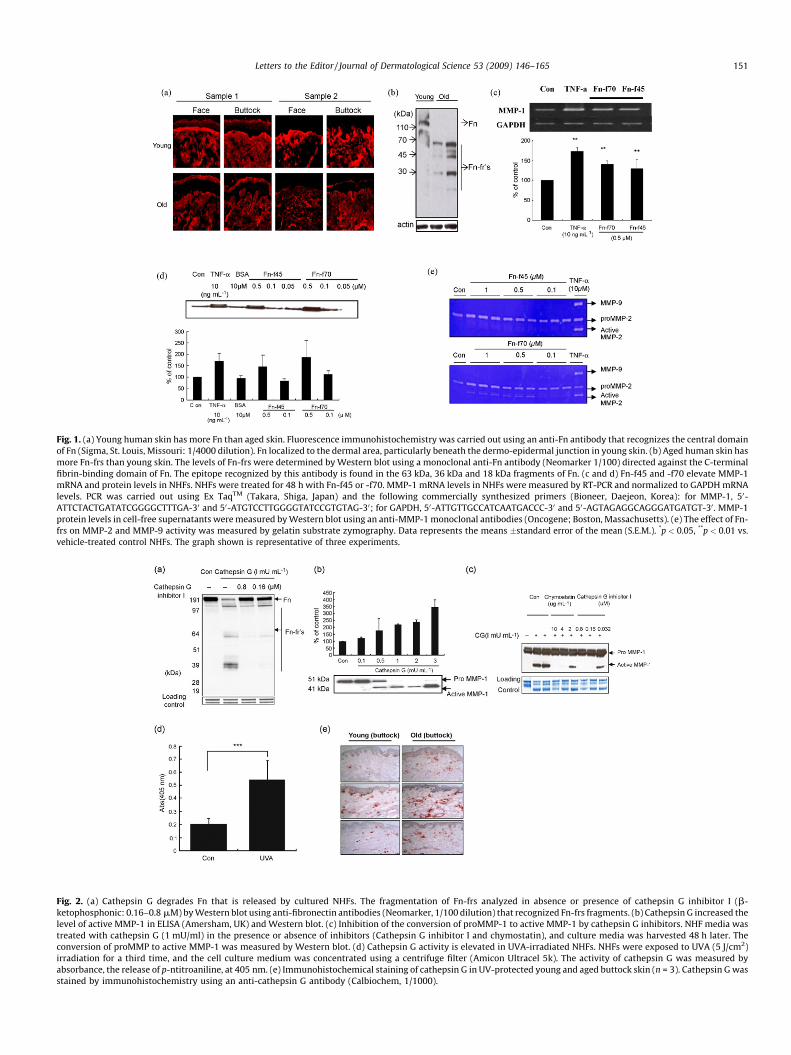
Fig. 1. (a) Young human skin has more Fn than aged skin. Fluorescence immunohistochemistry was carried out using an anti-Fn antibody that recognizes the central domain
of Fn (Sigma, St. Louis, Missouri: 1/4000 dilution). Fn localized to the dermal area, particularly beneath the dermo-epidermal junction in young skin. (b) Aged human skin has
more Fn-frs than young skin. The levels of Fn-frs were determined by Western blot using a monoclonal anti-Fn antibody (Neomarker 1/100) directed against the C-terminal
fibrin-binding domain of Fn. The epitope recognized by this antibody is found in the 63 kDa, 36 kDa and 18 kDa fragments of Fn. (c and d) Fn-f45 and -f70 elevate MMP-1
mRNA and protein levels in NHFs. NHFs were treated for 48 h with Fn-f45 or -f70. MMP-1 mRNA levels in NHFs were measured by RT-PCR and normalized to GAPDH mRNA
levels. PCR was carried out using Ex TaqTM (Takara, Shiga, Japan) and the following commercially synthesized primers (Bioneer, Daejeon, Korea): for MMP-1, 50-
ATTCTACTGATATCGGGGCTTTGA-30 and 50-ATGTCCTTGGGGTATCCGTGTAG-30; for GAPDH, 50-ATTGTTGCCATCAATGACCC-30 and 50-AGTAGAGGCAGGGATGATGT-30 . MMP-1
protein levels in cell-free supernatants were measured by Western blot using an anti-MMP-1 monoclonal antibodies (Oncogene; Boston, Massachusetts). (e) The effect of Fn-
frs on MMP-2 and MMP-9 activity was measured by gelatin substrate zymography. Data represents the means �standard error of the mean (S.E.M.). *p < 0.05, **p < 0.01 vs.
vehicle-treated control NHFs. The graph shown is representative of three experiments.
Fig. 2. (a) Cathepsin G degrades Fn that is released by cultured NHFs. The fragmentation of Fn-frs analyzed in absence or presence of cathepsin G inhibitor I (b-
ketophosphonic: 0.16–0.8 mM) by Western blot using anti-fibronectin antibodies (Neomarker, 1/100 dilution) that recognized Fn-frs fragments. (b) Cathepsin G increased the
level of active MMP-1 in ELISA (Amersham, UK) and Western blot. (c) Inhibition of the conversion of proMMP-1 to active MMP-1 by cathepsin G inhibitors. NHF media was
treated with cathepsin G (1 mU/ml) in the presence or absence of inhibitors (Cathepsin G inhibitor I and chymostatin), and culture media was harvested 48 h later. The
conversion of proMMP to active MMP-1 was measured by Western blot. (d) Cathepsin G activity is elevated in UVA-irradiated NHFs. NHFs were exposed to UVA (5 J/cm2)
irradiation for a third time, and the cell culture medium was concentrated using a centrifuge filter (Amicon Ultracel 5k). The activity of cathepsin G was measured by
absorbance, the release of p-ntitroaniline, at 405 nm. (e) Immunohistochemical staining of cathepsin G in UV-protected young and aged buttock skin (n = 3). Cathepsin G was
stained by immunohistochemistry using an anti-cathepsin G antibody (Calbiochem, 1/1000).
Letters to the Editor / Journal of Dermatological Science 53 (2009) 146–165 151

Letters to the Editor / Journal of Dermatological Science 53 (2009) 146–165152
time to NHFs in the presences or absence of cathepsin G inhibitor I(b-ketophosphonic acid). We also examined the effect ofcathepsin G on the levels of active MMP-1 in cell-free super-natants, and whether the inhibition of cathepsin G effected theconversion of proMMP-1 to active MMP-1. Fn fragmentation wasreduced in media treated with cathepsin G plus inhibitor ascompared to cathepsin G treatment alone (Fig. 2a). Fragmentationpattern of Fn from cell supernatants was very similar to thefragmentation pattern of purified Fn by 10 mU cathepsin G (datanot shown).
Cathepsin G increased the level of active MMP-1 in a dose-dependent manner (Fig. 2b), as promoted the conversion ofproMMP-1 (52/54 kDa) into active MMP-1 (42/44 kDa). Theaddition of cathepsin G inhibitors (Cathepsin G Inhibitor I andchymostatin) prevented the conversion of proMMP-1 to activeMMP-1 (Fig. 2c).
Also, while MMP-1 protein and active MMP-2 were induced inNHFs following exposure to UVB (50 mJ/cm2) irradiation, we didnot detect Fn-frs in the medium (data not shown). Thus, it islikely that MMPs, including MMP-1 and MMP-2, are not themajor enzymes that digest Fn. Our results indicate that cathepsinG plays an important role in digestion of Fn and the activation ofMMP-1.
After NHFs exposed to Bio-Sun UVA (5 J/cm2) irradiation(Vilbert-Lourmat, France), the activity of cathepsin G in cell-freesupernatants was increased 2.6 folds (Fig. 2d). We also detectedthe levels of cathepsin G in young and aged human skin (n = 3) byimmunohistochemistry. Cathepsin G was strongly expressed in theECM and was present at higher levels in aged human skin than inyoung human skin (Fig. 2e). These results suggested that cathepsinG activity is upregulated in aged skin, that in vitro, the activity isupregulated to UV irradiation.
In the current study, we demonstrated that the extent of Fnfragmentation is greater in aged skin than young skin in vivo, andthat cathepsin G is expressed at higher levels in aged human skinthan in young skin. We also showed that UVA irradiation of NHFs inculture increased cathepsin G activity. Fn was readily degraded bycathepsin G into four or five fragments, and Fn-f45 and Fn-f70increased MMP-1 mRNA and protein levels and MMP-2 activity inNHFs. However, only Fn-f70 increased the levels of active MMP-2in NHFs. These observations suggest that cathepsin G and Fnfragments contribute to matrix damage in aged skin.
Acknowledgment
This study was supported in part by a grant of the Korea Health21 R&D Project, Ministry of Health & Welfare, Republic of Korea(A050432).
References
[1] Hynes Ro. Fibronectins. New York: Springer-Verlag; 1990.[2] Labat-Robert J. Fibronectin in malignancy. Semin Cancer Biol 2002;12:187–95.
[3] Vartio T. Characterization of the binding domains in the fragments cleaved bycathepsin G from human plasma fibronectin. Eur J Biochem 1982;123:223–33.
[4] Stanton H, Ung L, Fosang AJ. The 45 kDa collagen-binding fragment of fibro-nectin induces MMP-13 synthesis by chrondrocytes and aggrecan degradationby aggrecanases. Biochem J 2002;364:181–90.
[5] Xie DL, Hui F, Meyers R, Homandberg GA. Cartilage chondrolysis by fibronectinfragments is associated with release of several proteinases; stromelysin plays amajor role in chondrolysis. Arch Biochem Biophys 1994;311:205–12.
[6] Loeser RF, Forsyth CB, Samarel AM, Im HJ. Fibronectin fragment activation ofproline-rich tyrosine kinase PYK2 mediates integrin signals regulating collage-nase-3 expression by human chondrocytes through a protein kinase C-depen-dent pathway. J Biol Chem 2003;278:24577–85.
[7] Fyrand O. Studies on fibronectin in the skin. V. Indirect immunofluorescencestudies in dermatitis herpetiformis. Dermatologica 1981;162:220–9.
[8] Homandberg GA, Wen C, Hui F. Cartilage damaging activities of fibronectinfragments derived from cartilage and synovial fluid. Osteoarthritis Cartilage1998;6:231–44.
Eui Dong Son
Hyaekyoung Kim
Hyunjung Choi
So Hee Lee
Jin Young Lee
Sujung Kim
AmorePacific Corp/R&D Center, 314-1 Bora-dong,
Giheung-gu, Yongin-si, Kyeonggi-do,
Republic of Korea
B. Closs
Silab, 19108 Brive Cedex, France
Sera Lee
Jin Ho Chung
Department of Dermatology,
Seoul National University College of Medicine and
Institute of Dermatological Science,
Seoul National University,
Seoul, Republic of Korea
Jae Sung Hwang*
Skin research Institute AmorePacific R&D Center,
314-1 Bora-dong, Giheung-gu,
Yongin-si, Kyeonggi-do 446-729,
Republic of Korea
*Corresponding author. Tel.: +82 31 280 5811;
fax: +82 31 282 6063
E-mail address: [email protected] (J.S. Hwang)
22 April 2008
doi:10.1016/j.jdermsci.2008.08.006