castration modifies aortic vasoreactivity and serum fatty acids in a sucrose-fed rat model of...
TRANSCRIPT

ORIGINAL ARTICLE
Heart Vessels (2009) 24:147–155 © Springer 2009DOI 10.1007/s00380-008-1098-x
I. PerezDepartment of Pathology, Instituto Nacional de Cardiologia “Ignacio Chavez”, Mexico D.F., Mexico
M. El Hafi di · K. Carvajal · G. Baños (*)Department of Biochemistry, Instituto Nacional de Cardiologia “Ignacio Chavez”, Juan Badiano 1, Tlalpan. Col. Seccion XVI, 14080 Mexico D.F., MexicoTel. + 52-55-732911 ext. 1298; Fax + 52-55-730926e-mail: [email protected]
Israel Perez · Mohammed El Hafi di · Karla Carvajal Guadalupe Baños
Castration modifi es aortic vasoreactivity and serum fatty acids in a sucrose-fed rat model of metabolic syndrome
Abstract Levels of testosterone and estradiol infl uence the incidence of cardiovascular diseases: generally, estrogens in females are protective before menopause; coronaropathies, hypertension, and dyslipidemias in normal men are more frequent at comparable ages. We investigated the modula-tion by castration of in vitro vasoreactivity, serum lipid content, and systolic blood pressure (SBP) in rats with sucrose-induced metabolic syndrome. The main character-istics of the rat model are: hypertriglyceridemia, moderately high blood pressure, intra-abdominal accumulation of adipose tissue, hyperinsulinemia, nephropathy, increased oxidative stress, and altered vasoreactivity. Male weanling rats received 30% sucrose solution for 16 weeks (metabolic syndrome; MS), controls (C) had plain water; both had commercial rodent chow. They were subdivided into fi ve groups with two subgroups each: Group 1, intact C and MS rats, Groups 2–5, C and MS rats castrated for periods of 16, 12, 8, and 4 weeks. At the end of the study period, systolic blood pressure was measured, and blood and aortas were obtained for fatty acid determination and vasoreactivity assays, respectively. After 16 weeks’ sucrose treatment MS aortas showed hypercontractility and decreased vasodila-tion. Palmitic and palmitoleic acids were increased in MS versus C. Arachidonic acid levels in MS were lower than in intact or castrated C. Long-term castration of 16 weeks normalized the levels of palmitic and oleic acids. With the shorter periods of castration, contractility increased and relaxation decreased in C and MS, but it was more signifi -cant in C. Regarding fatty acid composition, long-term castration increased polyunsaturated (arachidonic and eicosapentaenoic) fatty acids. The shorter periods did not modify the fatty acid profi le in either C or MS. Metabolic
syndrome altered SBP, aortic reactivity, and levels of fatty acids; castration of long duration normalized them in some cases.
Key words Rat metabolic syndrome · Castration · Vascular reactivity · Free fatty acids
Introduction
The metabolic syndrome (MS) and particularly some of its main features such as dyslipidemia (hypertriglyceridemia with altered lipoprotein and fatty acid profi le), hyperten-sion, diabetes, and obesity, have become a worldwide epi-demic and, consequently, a subject of high-priority studies both for prevention and therapy. Although the pathology may only have a genetic origin in a portion of the popula-tion, in a large majority of cases it is the result of an inade-quate diet high in carbohydrates and/or fat, are associated with lack of physical exercise, which are environmental factors that contribute to a considerable increase in the prevalence of the syndrome.
Association is frequent between hypertension and meta-bolic disorders such as dyslipidemia and obesity. These abnormalities are risk factors in the development of cardio-vascular diseases. Peiris et al.1 have demonstrated that in obese subjects, visceral fat is a strong indicator of hyperten-sion. This fat is generated by an increase in the synthesis of free fatty acids (FFA) by the liver. Simon et al.2 found that a diet rich in saturated fatty acids increased systolic blood pressure (SBP), whereas a diet that included polyunsatu-rated and monounsaturated fatty acids decreased it, by reducing the levels of endothelial prostaglandins. Negative correlations between adiponectin serum levels and some variables associated with the metabolic syndrome, such as obesity and hypertension, have been described. In a study of healthy Japanese men it was found that there is an inde-pendent negative correlation between serum adiponectin levels and body mass index, but the correlation was positive when adiponectin and sleep duration were analyzed. These
Received: January 16, 2008 / Accepted: July 31, 2008

148
results evidence the infl uence of lifestyle factors upon a hormone whose functions include the regulation of the metabolism of lipids and glucose.3 On the other hand, for many years it has been known that gender may have an important role in the incidence of some of the pathological characteristics of the syndrome; females in their reproduc-tive years show some protection from disease development, such as cardiovascular complications, indicating that sex hormones participate in their modulation.4 Nevertheless, there are still controversial results concerning the role of hypertriglyceridemia as a cardiovascular risk factor and the possible protection by estrogens and by high levels of high density lipoproteins.
The effect of castration upon vasomotor responses and SBP in both male and female animals has been reported, as well as its reversal by hormone treatment.5,6 Sucrose-fed rat models of MS have been studied by several groups. Among their fi ndings, the participation of the sympathetic and renin–angiotensin systems has been described to affect renal hemodynamics and to elevate blood pressure.7
The role of androgens as modulators of blood pressure and vasoreactivity has been amply described. The observa-tions are somewhat controversial since the effects of the hormones vary: they have been found benefi cial in some cases of acute administration but detrimental in others, after chronic administration.8–11 The level of the hormones in in vivo or in vitro experiments may be a factor that deter-mines the elicited vascular response.12
We have developed a variant of the fructose- or sucrose-fed rat model fi rst described by Hwang et al.13 and Reaven and Ho,14 by administering 30% sucrose in the drinking water of weanling rats for periods of 20–24 weeks. The animals develop moderate hypertension, hypertriglyceride-mia, central accumulation of adipose tissue, altered serum free fatty acid levels, nephropathy, hyperinsulinemia, and in vitro vasoreactivity suggestive of endothelial damage.15,16 Blood glucose and cholesterol levels have been found to be normal.15 Most of these features characterize the metabolic syndrome.
In a study of in vivo plasma lipid oxidation in our model, we found that the degree of iron-induced lipoperoxidation was similar in sugar-fed males and females, whereas in con-trols the female rats had a lower degree than the males, suggesting that the apparent protection by estrogens might be lacking in the sugar-fed animals.17
Our own recent experience in studying the sucrose-fed model in which control and MS male rats were left intact or castrated with or without treatment with estradiol (E) or testosterone (T), has shown that SBP and levels of serum nitrates and nitrites (stable metabolites of nitric oxide) were modulated by castration and castration plus sex hormones. Castration alone and castration plus E in MS animals improved or normalized their altered vasoreactivity, reduced SBP, and increased serum nitrates and nitrites, whereas the administration of T reversed the improved responses to the castration. These observations show the infl uence of hormones in the vasoreactivity of the MS model as well as their relationship with endothelium-dependent function.18
As already mentioned, for many years it has been observed that testosterone has an important role in both normal and pathological cardiovascular physiology, although there are controversial reports.19,20 The purpose of this work was to investigate the effect of castration, at different stages of the treatment with sucrose, on the in vitro vascular reac-tivity of aortas as well as on body weight, SBP, and serum total fatty acids, in both male control and sucrose-fed rats (MS). The results of this study may give some indication as to the way sex hormones, particularly testosterone, and circulating fatty acids contribute to modulate vasoreactivity and SBP in MS animals.
Materials and methods
Animals
All studies were conducted in compliance with applicable laws and regulations as well as the principles expressed in the NIH, USPHS, and Guide for the Care and Use of Laboratory Animals, and the studies have been conducted on animals that were lawfully acquired. Use of animals has been approved by our Institution’s Animal Care and Use Committee.
Weanling male Wistar rats aged 4 weeks and weighing 62 ± 19 g were separated into two groups and each group into fi ve subgroups (n = 10 each): Group 1, control rats given tap water for drinking and Group 2, MS rats given 30% sucrose in their drinking water for 16 weeks (the actual experimental period). All animals were fed com-mercial rat chow ad libitum (Purina 5001. Richmond, IN, USA, containing 14.63 kJ/g, 23% protein, 12% fat, and 65% carbohydrate). The animals in the subgroups were either left intact or castrated at 4, 8, 12, and 16 weeks of age (Fig. 1).
The end of the sucrose treatment period was taken as the time when SBP was signifi cantly increased in at least 80% of the animals in the MS Group, i.e., 20 weeks of age. Monthly measurements of systolic arterial pressure were
Fig. 1. Time line diagram. The fi rst line indicates total experimental period in intact rats. Arrows indicate time of castration. Sucrose treat-ment was started at weaning (4 weeks of age)

149
taken by the tail-cuff method; the cuff was connected to a pneumatic pulse transducer (Narco Bio-systems, Health-dyne, Houston, TX, USA) and a programmed electrosphyg-momanometer (Narco Bio-systems). The recordings were taken in triplicate by means of a Grass polygraph (model 79, Grass Medical Instruments, Quincy, MA, USA). The last measurement was taken on the last week of the sucrose treatment.
Orchiectomy
The animals were anesthetized with pentobarbital sodium (63 mg/kg of body weight). The area of the scrotum was shaved, cleaned with soap, and disinfected with ethanol. A longitudinal incision of approximately 1 cm was made, the efferent duct of each testicle was ligated, and the testicles were removed immediately. After the extirpation the skin incision was sutured.
The procedure was performed both in C and MS groups at 4, 8, 12, or 16 weeks of age (Fig. 1). The groups were labeled according to the length of time the animals had been castrated, i.e., the 16-week group was castrated at 4 weeks of age simultaneously with the beginning of sucrose treatment, and a similar pattern was followed for the other groups. Intact C and MS groups were kept in parallel.
Blood samples
At the end of the treatment period, the animals were fasted overnight and then killed by decapitation. Blood samples were collected in tubes kept on ice, taking care to avoid hemolysis, and the serum was separated. Blood samples for plasma also were obtained in tubes containing 0.1% ethyl-enediamine tetraacetic acid; 0.005% butylated hydroxy-toluene (BHT) was added to the plasma as antioxidant; the samples were kept on ice. The blood was spun at 4°C and both serum and plasma were frozen at −70°C until required for assays. Triglycerides were measured by means of an enzymatic technique (Boehringer Mannheim, Mannheim, Germany) in an Abbott VP Series II Autoanalyzer (Irving, TX, USA), according to the method described by Nägele et al.21
Lipid extraction and analysis of fatty acid (FA) composition
Fatty acids were extracted from 100 μl of plasma in the presence of 10 μg of heptadecanoic acid (internal standard) using chloroform/methanol (2 : 1 V/V) containing 0.002% of BHT to prevent auto-oxidation, as described by Folch et al.22 The obtained lipid residue was dissolved in 2 ml methanol containing 2% of concentrated H2SO4, and FA were esterifi ed to their corresponding methyl esters by heating at 90°C during 2 h, as previously described.23 The concentration and composition of FA were evaluated with a Shimadzu gas chromatograph (GC 2010, Kyoto, Japan) equipped with autoinjector/autosampler (AOC-20i) and
Shimadzu GC solution software (version 2.3, Shimadzu, Kyoto Japan).
The areas of the peaks were calculated using the program supplied with the chromatograph; the identifi cation of each fatty acid methyl ester was made by comparing their reten-tion time with their corresponding standards.
Plasma testosterone
A Coat-A-Count (Siemens) kit was used. It is a 125I radioim-munoassay in solid phase designed for quantifi cation of serum or heparinized plasma testosterone, without extrac-tion. It uses a specifi c antibody against testosterone; the incubation time is of 3 h at 37°C. The interval for calibration was from 0.2 ng/ml to 16.0 ng/ml. The intra-assay variation coeffi cient was 6.52% and the inter-assay, 7.01%.
Vascular tissue preparation and tension recording
After decapitation, the upper thoracic aorta was immedi-ately excised and kept in an oxygenated standard Krebs solution of the following composition (mM): 118 NaCl, 4.7 KCl, 2.5 CaCl2, 1.2 MgSO4⋅7H2O, 24 NaHCO3, 1.2 KH2PO4, and 4.5 glucose. Under a dissecting microscope, the vessels were cleaned of surrounding tissue, arterial segments of 2–3 mm were cut from the aorta and two nichrome S-shaped hooks were inserted into the lumen to measure the tension developed transversally. One of the hooks was fi xed to the bottom of a double-walled organ chamber, and the other was attached to an isometric force-displacement transducer (FT03, Grass Medical Instruments). The transducers were connected to a Grass polygraph model 79 D. A basal passive tension of 2 g was applied to the aortic rings. The Krebs fl uid was continuously bubbled with a gas mixture of 95% O2 − 5% CO2, keeping it warm (37°C), and was changed every 20 min. The rings were allowed to equilibrate for 1 h.
Contractions were induced by increasing concentrations, 2 × 10−9 to 2 × 10−5 M norepinephrine (NE). After the devel-oped tension was recorded, the rings were washed with normal Krebs solution and were allowed to return to their basal tension level. The procedure was repeated and the mean value of the force developed was taken as a 100% response.
Relaxation curves were obtained by precontracting the aortic rings with 2 × 10−7 M NE and then adding acetyl-choline (ACh) at increasing concentrations (2 × 10−9 to 2 × 10−5 M).
Statistical analyses
Concentration–response curves for vascular reactivity were obtained using the program GraphPad Prism 3.1, which compares whole curves (nonlinear regression, sigmoid dose–response curves), to calculate statistical signifi cance. Two-way analysis of variance (ANOVA), considering the length of the castration period and the MS condition as the
150
two independent variables, and Bonferroni’s post-test, were applied to determine differences between groups. The levels of fatty acids are expressed as percentages. Numerical values are means and standard error of the means, (n = 7–10).
Results
Body weight
The average body weight in intact MS rats was signifi cantly lower than in intact C animals at the end of the experimen-tal period of 20 weeks: 412 ± 11 vs 465 ± 12 g, respectively (P = 0.001). In castrated rats, the 16- and 12-week groups showed a signifi cantly decreased average body weight when compared with that of the intact groups (C = 438.5 ± 15, MS = 369 ± 12, P < 0.05 vs intact). The other groups, i.e., 8- and 4-week-castration, did not show signifi cant differences when compared with their intact controls (Table 1).
Systolic blood pressure
After 16 weeks of sucrose treatment, the increase in SBP in intact MS rats was signifi cant when compared with that of intact control animals: 139 ± 1 vs 121 ± 0.8 mmHg, respec-tively (P = 0.001). In castrated rats, the 16-, 12-, and 4-week castrated MS groups showed a signifi cantly reduced SBP (129.8 ± 1, 128 ± 5, and 127 ± 3 mmHg, respectively) when compared with that of the intact MS (139 ± 1, P = 0.001). The SBP was not signifi cantly modifi ed by castration in any of the C groups or in the 8-week castrated MS; in the latter it remained signifi cantly higher than in C. Table 1 shows SBP values for all groups at the end of the experimental period.
Serum triglycerides
These were only measured in intact C and MS groups and castrated C and castrated MS of the 16-week castration groups. Their concentrations were (mg/dl): intact C 71 ± 3, intact MS 136 ± 5 (P < 0.0001), castrated C 45 ± 14, and castrated MS 86 ± 4 (P = 0.02).
Plasma testosterone
The levels of testosterone were 7.9 ± 1.6 and 1.14 ± 0.03 ng/ml (P = 0.001 C vs MS) in intact C and intact MS, respec-tively. After castration they decreased to undetectable in both.
Vascular reactivity
Figure 2 shows the contraction and relaxation curves of concentration response to NE and ACh in the groups in which the orchiectomy lasted 16 and 12 weeks and in which the rats were treated with sucrose for a total of 16 weeks. The resulting graphs indicate that the contractility in the MS aortas was signifi cantly larger than in C rings and that the relaxation was also signifi cantly reduced in MS aortic rings when compared with that of C preparations. Castra-tion only modifi ed the responses of MS rings, decreasing contraction and increasing relaxation, both to normal levels.
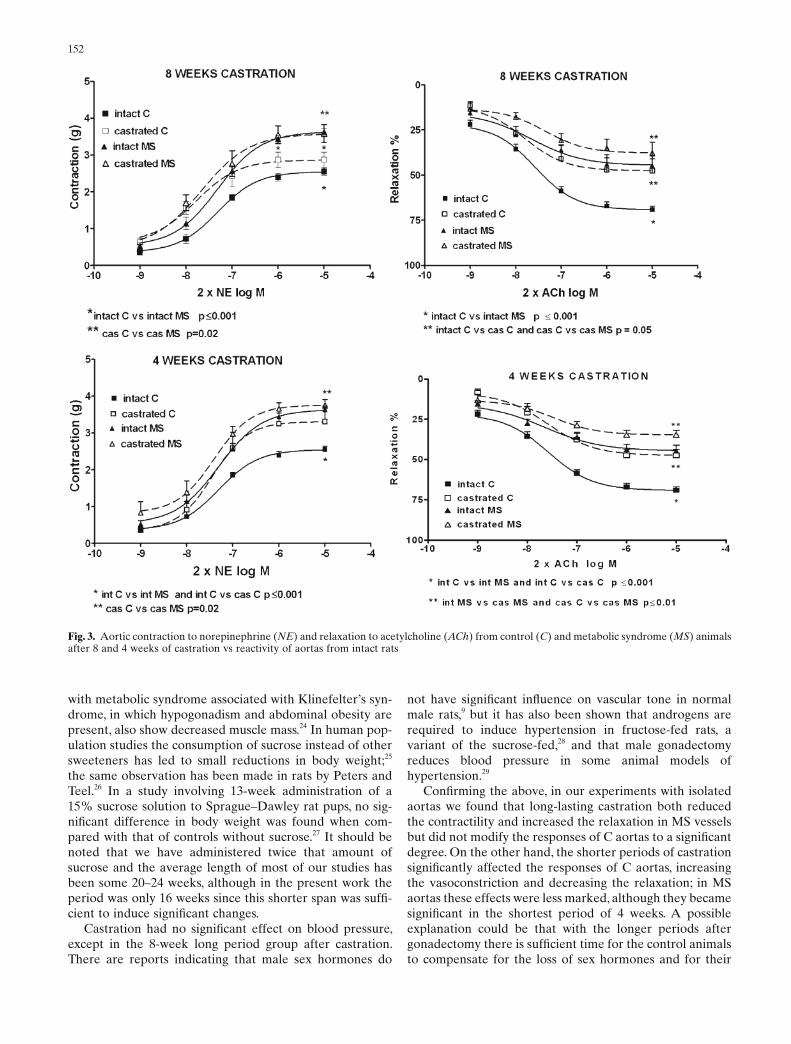
When castration lasted 8 weeks (Fig. 3), we observed that the contractility in castrated C and castrated MS rings increased when compared with the longer periods of castra-tion, although the difference in response between the two was signifi cant, the MS being larger and not now signifi -cantly different from that of intact MS. The same relation-ships were found when comparing the relaxation curves: the responses decreased in both castrated C and castrated MS, but the difference with intact rings was only signifi cant in castrated C aortic rings. The reactivity in the group with 4 weeks’ castration followed the same pattern as that of 8 weeks’ castration, except that the decrease in relaxation became signifi cant in both the castrated C and castrated MS aortas when compared with the intact C and intact MS, respectively.
Serum fatty acid composition
The areas of the peaks were calculated using the program supplied with the chromatograph; the identifi cation of each fatty acid methyl-ester was made by comparing their reten-tion time with their corresponding standards (Table 2). The proportions of palmitic, palmitoleic, and oleic acids were signifi cantly higher in intact MS than in intact C, and this
Table 1. Body weight and systolic pressure
Intact 16 weeks castration 12 weeks castration 8 weeks castration 4 weeks castration
C MS C MS C MS C MS C MS
Body weight at 20 weeks of age (g)465 ± 12 412 ± 11* 448 ± 18 367 ± 12* 428 ± 21 370 ± 12*** 469 ± 19 405 ± 18** 420 ± 18 382 ± 27
Systolic pressure at 20 weeks of age (mmHg)121 ± 0.8 139 ± 1* 125 ± 6 129.8 ± 1 123 ± 4 128 ± 5 117 ± 2 132 ± 4* 121 ± 2 127 ± 3
C, control; MS, metabolic syndrome* P < 0.01, ** P = 0.03, *** P = 0.04. The P values were obtained by comparing intact or castrated MS with their corresponding intact or castrated C.

151
difference was not modifi ed by castration, except in the 16-week group in the case of palmitic and oleic acids.
On the other hand, the proportions of linoleic, arachi-donic, and docosahexaenoic acids were lower in the intact MS than in the intact C groups; castration only increased arachidonic acid in the 12- and 16-week groups. Eicosapen-taenoic acid proportion was also higher in the 16-week castrated MS group than in castrated C.
Discussion
The most relevant fi ndings in this work were that the vas-cular reactivity in this model of metabolic syndrome was
altered, suggesting that endothelial function was impaired, among other possible abnormalities, such as those that can be induced by fl uctuations in levels of sex hormones or fatty acids. The castration in the long term modifi ed those altera-tions and the reactivity was normalized. The composition of serum fatty acids was also affected and with long-term castration in some cases they tended to normality.
Regarding body weight, the lower gain in MS rats can be explained by their associated lower intake of solid food, as previously reported, although they do accumulate a large amount of intraperitoneal adipose tissue on account of a higher intake of calories from the sucrose solution they drink. This increase is not, however, refl ected in the total body weight.15 It is possible that the muscle mass decreases, but we have yet to evaluate this. In this respect, patients
Fig. 2. Aortic contraction to norepinephrine (NE) and relaxation to acetylcholine (ACh) from control (C) and metabolic syndrome (MS) animals after 16 and 12 weeks of castration vs reactivity of aortas from intact rats
152
with metabolic syndrome associated with Klinefelter’s syn-drome, in which hypogonadism and abdominal obesity are present, also show decreased muscle mass.24 In human pop-ulation studies the consumption of sucrose instead of other sweeteners has led to small reductions in body weight;25 the same observation has been made in rats by Peters and Teel.26 In a study involving 13-week administration of a 15% sucrose solution to Sprague–Dawley rat pups, no sig-nifi cant difference in body weight was found when com-pared with that of controls without sucrose.27 It should be noted that we have administered twice that amount of sucrose and the average length of most of our studies has been some 20–24 weeks, although in the present work the period was only 16 weeks since this shorter span was suffi -cient to induce signifi cant changes.
Castration had no signifi cant effect on blood pressure, except in the 8-week long period group after castration. There are reports indicating that male sex hormones do
Fig. 3. Aortic contraction to norepinephrine (NE) and relaxation to acetylcholine (ACh) from control (C) and metabolic syndrome (MS) animals after 8 and 4 weeks of castration vs reactivity of aortas from intact rats
not have signifi cant infl uence on vascular tone in normal male rats,9 but it has also been shown that androgens are required to induce hypertension in fructose-fed rats, a variant of the sucrose-fed,28 and that male gonadectomy reduces blood pressure in some animal models of hypertension.29
Confi rming the above, in our experiments with isolated aortas we found that long-lasting castration both reduced the contractility and increased the relaxation in MS vessels but did not modify the responses of C aortas to a signifi cant degree. On the other hand, the shorter periods of castration signifi cantly affected the responses of C aortas, increasing the vasoconstriction and decreasing the relaxation; in MS aortas these effects were less marked, although they became signifi cant in the shortest period of 4 weeks. A possible explanation could be that with the longer periods after gonadectomy there is suffi cient time for the control animals to compensate for the loss of sex hormones and for their

153
arteries to recover normal vasoreactivity, due to mecha-nisms such as increased expression of nitric oxide synthase induced by a release of estradiol through possible aromati-zation, as indicated by a previous study.18 In MS rats, in which the endothelium-dependent responses suggest impairment, this recovery may not take place in the short-term castration and the vasoreactivity remains altered. It should be mentioned that aortic tissue preparations have been examined to determine morphological or mass changes but none have been detected.
Regarding the results of the ANOVA test on the reactiv-ity data (Table 3), there was a signifi cant effect in the maximal response, either of contraction or of relaxation. The test suggests that the effect was given both by the pathological condition and by the length of the castration, the latter contributing more to the results (38.4% for NE and 45% for ACh). There was a synergism of the two effects that participated with 13% of the variation in the case of the ACh. No variations in EC were found, which suggests that changes are unlikely in the responsiveness of the organ to the agonist. Thus it is possible to think that such a response may be due to perturbations in other mechanisms contributing to the contraction and relaxation processes, such as synthesis and release of NO. Indeed, NOS activity has been suggested to be diminished in the sucrose-fed hypertriglyceridemic rat.15
The participation of alpha-adrenergic receptors and their corresponding signal mechanisms have been investigated in patients with the metabolic syndrome. With the use of blockers, it has been demonstrated that vasoconstriction of coronary microcirculation is indeed mediated by adrenergic receptors.30 A similar association has been found in obese Zucker rats where skeletal muscle perfusion is defective as a result of alpha-adrenergic-induced vasoconstriction.31 A recent study in a novel model of MS in rats has coincided with that observation.32 On the other hand, there was a sig-nifi cant difference in the maximal contraction and relax-ation responses between intact MS and castrated MS of long duration, indicative of the reversal, by castration, of the damage induced by the MS condition.
It is worth mentioning that androgens have dual effects on vessels: low levels found in conditions such as the metabolic syndrome may induce high blood pressure and increased vascular contractility, whereas high levels of testosterone generally normalize high blood pressure and decrease hyper-reactivity of isolated arteries;33 some of its metabolites such as 5-beta-dihydrotestosterone are potent in vitro vasodilators and their absence might favor increased contractility and reduced relaxation in the short term.34 Some of the mechanisms involved after castration may include genomic effects on sex hormone receptors, such as production of estradiol from testosterone, through aroma-tase activity.35
Another pathological variable present in the MS is insulin resistance, which has recently been shown to be involved in endothelium-dependent vasodilatory effects in peripheral circulation. In nondiabetic patients insulin resis-tance does reduce coronary dilatation, which is associated with the degree of insulin resistance in those patients, Ta
ble
2. P
erce
ntag
e of
fat
ty a
cids
in t
he r
at s
erum
(m
ean
± SE
M)
Fatt
y ac
idIn
tact
CIn
tact
MS
16 w
eeks
cas
trat
ion
12 w
eeks
cas
trat
ion
8 w
eeks
cas
trat
ion
4 w
eeks
cas
trat
ion
CM
SC
MS
CM
SC
MS
Pal
mit
ic24
.2 ±
1.4
30.1
± 1
.5*#
23 ±
229
.8 ±
523
.4 ±
131
.5 ±
2.4
*#26
.4 ±
1.8
31.9
± 4
.9*#
25.8
± 1
.633
.5 ±
2.6
*#
Pal
mit
olei
c3.
0 ±
0.3
9.4
± 0.
7*#
3.3
± 0.
18.
6 ±
1.4*
#2.
6 ±
0.4
7.2
± 1.
0*#
2.65
± 1
.27.
8 ±
1.3*
#3
± 0.
59.
4 ±
0.9*
#
Ole
ic20
.15
± 2
29.7
± 1
.8*#
20.6
± 2
28.3
± 4
19.5
5 ±
0.3
28.1
± 2
.5*#
17.5
± 3
32.4
± 5
*#18
.0 ±
1.3
29.3
± 1
.6*#
Lin
olei
c23
.5 ±
0.8
513
.3 ±
0.7
*#20
.6 ±
4.0
12 ±
2.2
*#22
.26
± 0.
211
.66
± 0.
6*#
21.5
± 2
.69.
9 ±
1.5*
#24
.15
± 1.
49.
4 ±
1*#
Gam
ma
Lin
olen
ic0.
7 ±
0.1
0.6
± 0.
060.
8 ±
0.8
0.70
± 0
.20.
6 ±
0.6
0.65
± 0
.65
0.7
± 0.
10.
65 ±
0.1
0.7
± 0.
080.
6 ±
0.06
Ara
chid
onic
10.2
± 1
.44.
3 ±
0.2*
#17
.2 ±
5.6
7.5
± 1.
4*#
11.7
± 0
.86.
4 ±
1.6*
#9.
1 ±
1.36
4.1
± 0.
7*#
11.1
± 1
.74.
7 ±
0.8*
#
Eic
osap
enta
enoi
c1.
7 ±
0.6
1.1
± 0.
21.
3 ±
0.2
2.1
± 0.
7#0.
7 ±
0.1
0.7
± 0.
071.
6 ±
0.4
1.4
± 0.
31.
7 ±
0.2
1.1
± 0.
25D
ocos
ahex
aeno
ic2.
8 ±
0.6
2.0
± 0.
21.
8 ±
0.6
1.1
± 0.
22.
9 ±
0.8
1.2
± 0.
12.
4 ±
.0.5
1.15
± 0
.21.
8 ±
0.4
1.7
± 0.
3
C, c
ontr
ol; M
S, m
etab
olic
syn
drom
e*
Den
otes
dif
fere
nce
vs c
ontr
ol u
nder
the
sam
e co
ndit
ion
(tim
e of
cas
trat
ion)
aft
er B
onfe
rron
i’s p
ost-
test
# In t
hese
gro
ups
the
MS
cond
itio
n ac
coun
ts f
or 3
0%–6
5% o
f th
e ob
serv
ed d
iffe
renc
e, r
egar
dles
s of
the
len
gth
of c
astr
atio
n. R
egar
ding
the
lev
els
of f
atty
aci
ds, t
he i
nter
acti
on b
etw
een
the
MS
cond
itio
n an
d ca
stra
tion
was
not
sig
nifi c
ant.
Two-
way
ana
lysi
s of
var
ianc
e w
as p
erfo
rmed
con
side
ring
the
MS
cond
itio
n an
d th
e le
ngth
of
cast
rati
on a
s th
e tw
o fa
ctor
s af
fect
ing
the
resu
lts
for
each
fat
ty a
cid

154
besides generally being a risk factor for the development of coronary atherosclerosis.36
With regard to the fatty acids, both saturated fatty acids and triglycerides impair the endothelium-dependent relaxation and this, in turn, alters the vasoreactivity.37–39 Therefore, in our experiments with intact and 4- and 8 week-castration MS aortas, we could perhaps speculate that the increases in fatty acids affected the reactivity, increasing contraction and decreasing relaxation. The results of the ANOVA test indicate that contrary to what happens in the reactivity, it is the pathological condition, particularly the hypertriglyceridemia, which is responsible for the alter-ations in the fatty acid composition, and which in turn may be modifi ed by the length of castration.
The analysis of rat serum fatty acids in the intact MS group indicated a larger proportion of saturated fatty acids and a smaller of polyunsaturated, when compared with those of intact C, as previously observed.16,23,40 The increased proportion of membrane saturated fatty acids and decreased polyunsaturated fatty acids have been associated with insulin sensitivity in intraplatelet arginine–vasopressin calcium and sodium stimulation in patients with hyperten-sion and hyperinsulinemia.41
N-3 fatty acids and eicosapentaenoic acid oral supple-mentation increase membrane eicosapentaenoic acid content and reduce SBP in patients with essential hyperten-sion. Based on the association between the increase in membrane eicosapentaenoic acid content and the decrease in intracellular sodium concentration, eicosapentaenoic acid may lower blood pressure by altering the activities of the membrane sodium transport systems.42
In our study, the level of eicosapentaenoic acid tended to be lower than that of its corresponding control, except after the longest period of castration, when it became signifi cantly higher than its castrated control. This result suggests that long-term castration may improve SBP by participation of increases in the proportions of polyunsatu-rated fatty acids.
Table 3. Analysis of variance test on the reactivity data
Intact 16 weeks castration 12 weeks castration 8 weeks castration 4 weeks castration
Log EC50 NE Control −7.56 ± 0.42 −7.56 ± 0.42 −7.84 ± 0.22 −7.93 ± 0.42 −7.62 ± 0.23
n = 7 n = 7 n = 9 n = 7 n = 10 MS −8.14 ± 0.76 −7.65 ± 0.41 −8.11 ± 0.31 −7.43 ± 0.44 −7.48 ± 0.35
n = 7 n = 9 n = 10 n = 9 n = 10% Maximal relaxation ACh Control 68.4 ± 6.16 68.4 ± 6.2 71.7 ± 3.3 46.9 ± 4.2† 47.4 ± 2.5†
n = 7 n = 7 n = 9 n = 7 n = 7 MS 30.8 ± 3.9* 68.2 ± 4.4# 72.0 ± 4.1# 37.0 ± 3.2 35.0 ± 2.3
n = 7 n = 9 n = 10 n = 9 n = 10
Two-way analysis of variance test was performed considering time of castration and MS condition as the two factors contributing to the variance in the results. A Bonferroni post-test was performed to fi nd difference between MS and control groups and between the intact condition and different times of castration within the same groupNE, norepinephrine; ACh, acetylcholine* P < 0.05 vs Control intact. # P < 0.05 vs MS intact. † P < 0.05 vs Control intact. In this case, interaction between factors (MS condition and time of castration) was signifi cant (P < 0.0001) and it accounts for 12% of the results observed; MS condition accounts for only 9.6% (P < 0.0001) and time of castration for 45% (P < 0.0001)
The decrease in triglycerides in castrated rats probably contributed to the signifi cant differences found in some of the fatty acids, for instance in oleic acid, which has a pressor effect in humans and rats,43 although it did not decrease after castration. Arachidonic acid is a precursor of several metabolites with vasopressor effects, such as thromboxanes and most prostaglandins. The fact that its proportion was found to be reduced in MS serum suggested that its consumption was increased, with enhanced release of its metabolites, thus contributing to alterations in blood pres-sure and vasoreactivity. Measurements of the levels of pre-cursor and metabolites are required to confi rm the validity of this hypothesis.
The changes in vasoreactivity with the shorter periods of evolution of castration (8 and 4 weeks) in both C and MS aortas, are diffi cult to explain, particularly in the controls, since they were unexpected; in MS the effects were only signifi cant with the shortest period. Calcium handling may also be altered on account of hormone imbalance. There are reports which describe the participation of endogenous testosterone in the regulation of calcium fl uxes and the effect of short term castration in vascular smooth muscle.23,44 Another possibility may be that the differences in testoster-one levels before and after castration are larger in controls and this would be refl ected in the more signifi cant changes in responses in this group whereas, in the MS group, the basal levels of testosterone are low18 and castration would make a smaller difference than in the control. In conclusion, in a male rat model of metabolic syndrome in which the pathological characteristics include hypertension, altered lipid profi le, and apparent endothelium malfunction, castra-tion of long duration contributes to normalization of the vasoreactivity in isolated aortas and some of the levels of serum fatty acids.
Acknowledgments This work was partially supported by Grant Nos. 3210-PM and 43229-M from the National Council for Science and Technology (CONACyT) to G.B. and were the only sources of funding.

155
References
1. Peiris AN, Sothmann MS, Hoffmann RG (1989) Adiposity, fat distribution, and cardiovascular risk. Ann Int Med 110:867–872
2. Simon JA, Fong J, Bernert J (1996) Fatty acids and blood pressure. Hypertension 27:303–307
3. Kotani K, Sakane N, Saiga K, Kato M, Ishida K, Kato Y, Kurozawa Y (2007) Serum adiponectin levels and lifestyle factors in Japanese men. Heart Vessels 22(5):291–296
4. Galipeau D, Verma S, McNeill JH (2002) Female rats are pro-tected against fructose-induced changes in metabolism and blood pressure. Am J Physiol Heart Circ Physiol 283(6):H2478–H2484
5. Fischer GM, Swain ML (1977) Effect of sex hormones on blood pressure and vascular connective tissue in castrated and non-castrated male rats. Am J Physiol 232:H617–H621
6. Paredes-Carbajal MC, Juarez-Oropeza MA, Ortiz-Mendoza CM, Mascher D (1995) Effects of acute and chronic estrogenic treat-ment on vasomotor responses of aortic rings from ovariectomized rats. Life Sci 57:443–486
7. Freitas RR, Lopes KL, Carillo BA, Bergamaschi CT, Carmona AK, Casarini DE, Furukawa L, Heimann JC, Campos RR, Dolnikoff MS (2007) Sympathetic and renin-angiotensin systems contribute to increased blood pressure in sucrose-fed rats. Am J Hypertens 20(6):692–698
8. Reckelhoff JF (2005) Sex steroids, cardiovascular disease, and hypertension: unanswered questions and some speculations. Hypertension 45(2):170–174
9. Orshal JM, Khalil RA (2004) Gender, sex hormones and vascular tone. Am J Physiol Regul Integr Comp Physiol 286:R233–R249
10. Malkin CJ, Jones RD, Jones TH, Channer KS (2006) Effect of testosterone on ex vivo vascular reactivity in man. Clin Sci (Lond) 111(4):265–274
11. Dubey RK, Oparil S, Imthurn B, Jackson EK (2002) Sex hormones and hypertension. Cardiovasc Res 53(3):688–708
12. Iliescu R, Reckelhoff JF (2006) Testosterone and vascular reactiv-ity Clin Sci (Lond) 111(4):251–252
13. Hwang IS, Ho H, Hoffman BB, Reaven GM (1987) Fructose-induced insulin resistance and hypertension in rats. Hypertension 10(5):512–516
14. Reaven GM, Ho H (1991) Sugar-induced hypertension in Sprague-Dawley rats. Am J Hypertens 4(7):610–614
15. Baños G, Carvajal K, Cardoso G, Zamora J, Franco M (1997) Vascular reactivity and effects of serum in a rat model of hyperten-sion and hypertriglyceridemia. Am J Hypertens 10:379–388
16. El Hafi di M, Valdez R, Baños G (2000) Possible relationship between altered fatty acid composition of serum, platelets, and aorta and hypertension induced by sugar feeding in rats. Clin Exper Hypertens 22 (1):99–108
17. El Hafi di M, Baños G (1997) In vivo plasma lipid oxidation in sugar-induced rat hypertriglyceridemia and hypertension. Hyper-tension 30:624–628
18. Torres IP, El Hafi di M, Zamora-G J, Infante O, Chavira R, Baños G (2007) Modulation of aortic vascular reactivity by sex hormones in a male rat model of metabolic syndrome. Life Sci 80:2170–2180
19. Reckelhoff J F, Granger J P (1999) Role of androgens in mediating hypertension and renal injury. Clin Exp Pharm Physiol 26(2):127–131
20. Jones RD, Hugh Jones T, Channer KS (2004) The infl uence of testosterone upon vascular reactivity. Eur J Endocrinol 151(1):29–37
21. Nägele U, Hagele EO, Sauer G, Wiedemann E, Lehmann P, Wahlefeld AW, Gruber W (1984) Reagent for the enzymatic deter-mination of serum total triglycerides with improved lipolytic effi -ciency. J Clin Chem Clin Biochem 22(2):165–174
22. Folch L, Lees M, Sloane-Stanley CH (1957) A simple method for the isolation and purifi cation of total lipid from animal tissues. J Biol Chem 226:497–509
23. El Hafi di M, Cuellar A, Ramirez J, Banos G (2001) Effect of sucrose addition to drinking water, that induces hypertension in the rats, on liver microsomal Delta 9- and Delta 5-desaturase activities. J Nutr Biochem 12(7):396–403
24. Bojesen A, Kristensen K, Birkebaek NH, Fedder J, Mosekilde L, Bennett P, Laurberg P, Frystyk J, Flyvbjerg A, Christiansen JS, Gravholt CH (2006) The metabolic syndrome is frequent in
Klinefelter’s syndrome and is associated with abdominal obesity and hypogonadism. Diabetes Care 29(7):1591–1598
25. Vermunt SH, Pasman WJ, Schaafsma G, Kardinaal AF (2003) Effects of sugar intake on body weight: a review. Obes Rev 4(2):91–99
26. Peters LP, Teel RW (2003) Effects of high sucrose diet on body and liver weight and hepatic enzyme content and activity in the rat. In vivo 17(1):61–65
27. Hulman S, Falkner B (1994) The effect of excess dietary sucrose on growth, blood pressure, and metabolism in developing Sprague-Dawley rats. Pediatr Res 36(1 Pt 1):95–101
28. Song D, Arikawa E, Galipeau D, Battell M, McNeill JH (2004) Androgens are necessary for the development of fructose-induced hypertension. Hypertension 43(3):667–672
29. Martin DS, Biltoft S, Redetzke R, Vogel E (2005) Castration reduces blood pressure and autonomic venous tone in male spon-taneously hypertensive rats. J Hypertens 23(12):2229–2236
30. Camici PG, Marraccini P, Gistri R, Salvadori PA, Sorace O, L’Abbate A (1994) Adrenergically mediated coronary vasocon-striction in patients with syndrome X. Cardiovasc Drugs Ther 8(2):221–226
31. Frisbee JC (2004) Enhanced arteriolar alpha-adrenergic constric-tion impairs dilator responses and skeletal muscle perfusion in obese Zucker rats J Appl Physiol 97(2):764–772
32. Grisk O, Frauendorf T, Schlüter T, Klöting I, Kuttler B, Krebs A, Lüdemann J, Rettig R (2007) Impaired coronary function in Wistar Ottawa Karlsburg W rats – a new model of the metabolic syn-drome. Pfl ugers Arch 454(6):1011–1021
33. El Hafi di M, Pérez I, Carrillo S, Cardoso G, Zamora J, Chavira R, Baños G (2006) Effect of sex hormones on non-esterifi ed fatty acids, intra-abdominal fat accumulation, and hypertension induced by sucrose diet in male rats. Clin Exp Hypertens 28(8):669–681
34. Perusquia M, Villalon CM (1999) Possible role of Ca2+ channels in the vasodilating effect of 5beta-dihydrotestosterone in rat aorta. Eur J Pharm 371(2–3):169–178
35. Drago F, Lo Presti L, Nardo F, Panella I, Matera M, Scapagnini U (1982) Aromatization of testosterone by adipose tissue and sexual behavior of castrated male rats. Biol Reprod 27(4):765–770
36. Fujii N, Tsuchihashi K, Sasao H, Eguchi M, Miurakami H, Hase M, Higashiura K, Yuda S, Hashimoto A, Miura T, Ura N, Shimamoto K (2008) Insulin resistance functionally limits endothelium-dependent coronary vasodilation in nondiabetic patients Heart Vessels 23(1):9–15
37. Steinberg HO, Tarshoby M, Monestel R, Hook G, Cronin J, Johnson A, Bayazeed B, Baron AD (1997) Elevated circulating free fatty acid levels impair endothelium-dependent vasodilation. J Clin Invest 100:1230–1239
38. Steinberg HO, Paradisi G, Hook G, Crowder K, Cronin J, Baron AD (2000) Free fatty acid elevation impairs insulin-mediated vaso-dilation and nitric oxide production. Diabetes 49:1231–1238
39. Vigili de Kreutzenberg S, Crepaldi C, Marchetto S, Calo L, Tiengo A, delPrato S, Avogaro A (2000) Plasma free fatty acids and endothelium-dependent vasodilation: effect of chain length and cyclooxygenase inhibition. J Clin Endocrinol Metab 85:793–798
40. Perez I, El Hafi di M, Sanchez C, Baños G (1999) Effect of sugar-induced hypertension in rats on the pattern of serum arachidonic, dihomo-gamma-linoleic and linoleic acids. Med Sci Res 27:847–849
41. Maekawa K, Tsujino T, Saito K, Kim JI, Ikeda Y, Emoto N, Yokoyama M (2006) Inhibitory effect of insulin on vasopressin-induced intracellular calcium response is blunted in hyper-insulinemic hypertensive patients: role of membrane fatty acid composition. Heart Vessels 21(4):205–212
42. Miyajima T, Tsujino T, Saito K, Yokoyama M (2001) Effects of eicosapentaenoic acid on blood pressure, cell membrane fatty acids, and intracellular sodium concentration in essential hyperten-sion. Hypertens Res 24:537–542
43. Grekin R, Vollmer A, Sider R (1995) Pressor effects of portal venous oleate infusion, a proposed mechanism for obesity hyper-tension. Hypertension 26:193–198
44. Bowles DK, Maddali KK, Ganjam VK, Rubin LJ, Tharp DL, Turk JR, Heaps CL (2004) Endogenous testosterone increases L-type Ca2+ channel expression in porcine coronary smooth muscle. Am J Physiol Heart Circ Physiol 287:H2091–H2098