carbon-13 nuclear resonance analysis of metabolism soybeans … · a single soybean plant (glycine...
TRANSCRIPT

Plant Physiol. (1975) 55, 1048-1053
Carbon-13 Nuclear Magnetic Resonance Analysis of
Metabolism in Soybeans Labeled by 13CO2Received for publication August 19, 1974 and in revised form February 18, 1975
JACOB SCHAEFER, E. 0. STEJSKAL, AND C. F. BEARD'Corporate Research Department and Monsanto Agricultural Products Company, Monsanto Company, St. Louis,
Missouri 63166
ABSTRACT
Fourier transform 13C nuclear magnetic resonance spectrahave been obtained of intact, fresh soybean ovules (Glycine maxL. cv. Dare) harvested from pods subtended by a trifoliolateexposed to "3CO2 1 to 3 days earlier. The high resolution spectraare interpreted in terms of the labeled sugars and lipids in theovule. Comparison of the spectra taken over the 3-day periodpermits qualitative estimates of sugar metabolism and rates oflipid synthesis. The spectra also contain information about thedistribution of labels within the lipid chains. This informationleads to a method of estimating the extent to which glucosedegradation in the synthesizing soybean ovule is involved in thereactions of the phosphogluconate pathway.
production, but also, in part, the distribution of the '2C labelswithin the lipids.
This distribution in the lipids is complicated by the effect ofrecycling of hexose phosphates in the pentose cycle duringsynthesis in the ovule. That is, the label, when in the precursorsugar, is subject to a redistribution among unlabeled sugars dueto reactions of the complete pentose cycle, as well as by thereversible transaldolase-transketolase reactions which form thenondehydrogenase part of the pentose cycle (13). The oxida-tion of sugar by the reactions of the pentose phosphate cycleis a source of NADPH for the synthesizing ovule (3). Theinformation concerning pairs of labels available from a '2Clabeling and NMR experiment leads to a fast and simple wayof estimating the amount of glucose in an intact synthesizingsoybean ovule which is involved in the reactions of the pentosecycle, relative to that utilized by other metabolic pathways.
Carbon-1 3 labels can be introduced into soybean ovules byfeeding `CO,2 to a single soybean trifoliolate in a closed systemphotosynthesis cell. About 200 mg of '2CO2 are fixed in a 4-hourexperiment. The fixed 'C is transported to the ovules in theform of massively labeled sucrose, which is then used in thesynthesis of fatty acids, amino acids, and various insolublematerials (3). Analysis of the '3C NMR2 spectra of intactovules clearly identifies the labeled lipids present, as well asthose labeled sugars in solution. The spectra are well resolved,displaying widely spaced, relatively narrow lines, featurescommon to the 22C NMR spectra of many nonrigid systems (7,8).
Although the use of 1'C labels, followed by an NMR de-tection of the label, has been reported before, previous experi-ments have been, for the most part, restricted to the intro-duction of labeled intermediates into bacterial or mammaliansystems (9). There are at least two important advantages inusing '2CO2, rather than labeled intermediates, in the study ofcertain types of metabolism in soybeans. First, the label canbe introduced under completely normal conditions. Second, byadministering the "CO-2 for relatively short periods, it becomespossible to detect (from characteristic spin-multiplet patternsin the '2C NMR spectra) the concentration of isolated pairs of3C-'3C label present in fatty acid chains synthesized in theovules from the labeled sugar, relative to the concentration ofisolated single labels in the same chains. Thus, the `2C NMRanalysis can be used to establish not only the rate of lipid
I Present address: 113 Blue Rock Road, Wilmington, Del. 19809.- Abbreviation: NMR: nuclear niagnetic resonance.
MATERIALS AND METHODS
A single soybean plant (Glycine max, L. cv. Dare) waslabeled with `2CO, between 8 AM and noon on a day in the be-ginning of its log growth ovule-filling period (6). The labelingwas performed using a photosynthesis system in which a singlesoybean trifoliolate could be enclosed and sealed in a box witha transparent top, while still remaining attached to the wholeplant. The seal was achieved with a gummy rubber acting as agasket between the petiole and the box. Circulating air withadjusted temperature, humidity, and `2CO2 level entered the boxand passed over the trifoliolate. Progress of photosynthesiswas monitored by the rate of fixation of "2CO2 as determined bya differential infrared gas analyzer, by changes in the leaf tem-perature, and by changes in the relative humidity in the cell.The photosynthetic rate was determined to be in excess of 20mg of CO2 fixed/ dm2 leaf area hr and was reasonably constantthroughout the labeling period. Air temperature was main-tained at 31 + 2 C, leaf temperature at 29 ± 1 C, relativehumidity at 10 + 1%, and '2CO2 level at 300 ± 75 ppm. Leaftemperature was always less than air temperature. Relativehumidity was kept around 10% to help maintain a low leaftemperature during active photosynthesis. The relatively largerange on the CO2 level was the result of adding 1CO2 to thesystem in measured bursts rather than continuously. The timebetween bursts of "3CO2 was about 6 min. Isotopic purity of the'CO. was 90% as determined experimentally by mass spec-trometry. The flow rate of air through the system was 10 1/min.A complete purging of the system of 12CO2 and replacementwith "'CO2 required 5 min. The system operated at close toatmospheric pressure and was judged to be free of leaks asdetermined by the absence of a CO2 signal from the gasanalyzer (using N2 as the reference gas) when no plant was in
1048 www.plantphysiol.orgon March 15, 2020 - Published by Downloaded from
Copyright © 1975 American Society of Plant Biologists. All rights reserved.

C-13 NMR OF SOYBEANS
the system. The leak-free condition was confirmed at the endof the labeling period by the observation of a typical compensa-tion point for soybeans of about 40 p,l/l CO2 (1). The lightsource used in the experiment produced 950 jLeinsteins/m -secas determined by a photosynthetically active radiation quantumsensor (Lambda Instruments Co., Lincoln, Neb.). This corre-sponds to roughly 4500 ft-c and is a saturating light level.A trifoliolate was chosen for labeling which was in the
middle to upper middle region of the plant and which sub-tended a node having two pods, each of approximately thesame size and each containing two ovules of about equal size.One pod was harvested 24 hr after the completion of the label-ing and the second after 72 hr. One ovule was taken from eachof the two pods for examination by 'C NMR. In addition, asimilar pod from a comparable, unlabeled soybean plant wasalso harvested for purposes of obtaining a natural abundance'C NMR spectrum of a fresh ovule at the same stage ofdevelopment. Finally, in separate labeling experiments, sugarswere extracted from the leaves and from the subtending petiole,after exposure to 3CO2. This was done in order to obtain ahigh resolution 13C NMR spectrum of the labeled sugar beingsupplied to the ovule.
Carbon-I 3 NMR spectra were obtained at 22.6 MHz and 25C by standard pulsed techniques (2) using a Bruker HFX spec-trometer, some details of which have been described earlier (10).Repetitive, intense radiofrequency pulses, spaced 3 sec apart,excited the "C spin system, and the resulting NMR transientresponses, or free induction decays, were digitized and ac-cumulated in a Nicolet 1085 laboratory computer. After 1 hra suitably strong signal had been accumulated, and a Fouriertransform was performed. This calculation produced an ab-sorption NMR spectrum (2) whose line positions and inten-sities could be interpreted in the usual way, assuming thespacing of the repetitive pulses was sufficiently long (2). Spectrawere obtained using a spinning sample and a 10-mm diameterreceiver coil. All 'C NMR spectra were obtained under con-ditions of proton decoupling so that each line in a naturalabundance "3C spectrum represented a magnetically uniquechemical environment (11).
Experiments were performed on intact ovules and weremade with the ovule suspended in a dry, inert powder of poly-vinyl chloride. This technique ensured high resolution NMRmeasurements by minimizing magnetic susceptibility varia-tions within the sample coil, while at the same time not affect-ing the integrity of the intact ovule.
RESULTS AND DISCUSSION
Assignment of Spectra. The '1C NMR spectra of somelabeled and natural abundance fresh, intact soybean ovules areshown in Figure 1. At the time of harvest, an individual ovulehad a fresh weight of about 200 mg and a dry weight of about50 mg. The 1 % natural abundance "1C NMR spectrum of thiskind of ovule, which contains only about one-sixth as muchoil as a fully mature soybean (6), is therefore relatively weak.The '1C NMR spectra of single ovules harvested from thenode whose trifoliolate had been treated with l3CO2 1 or 3 daysearlier are, on the other hand, about 10 times as strong as thatof the unlabeled ovule. This is a result of the fact that for asoybean ovule at the beginning of its log-growth phase, thedaily production of oil is relatively large, even though the totaloil reserves are still small (6).The NMR lines appearing around 175 ppm from tetramethyl-
silane (as an external reference) are due to carboxyl carbons ofvarious types mostly in fatty acids in the form of lipids (tri-acylglycerols). The lines from the olefinic carbons of fatty
3 days after trcatmcnt.
1 day aftcr trcatmnit
naturaL abundance
I180 160 140 120 100 80 60 40 20 0
dc- (ppnt fron TMS)FIG. 1. Carbon-13 NMR spectra of intact fresh soybean ovules.
The upper two spectra are of ovules whose subtending trifoliolatehad been treated with '3CO2. Tetramethylsilane (TMS) is the externalreference. The magnetic field increases from left to right.
acids appear at 130 ppm, the lines between 60 and 100 ppmarise, for the most part, from the different kinds of carbons inthe soluble sugars in the ovule, while the group of linescentered about 30 ppm can be attributed to the methylene andmethyl carbons of the fatty acid chains (8). Immobilizedmaterials such as rigid proteins and carbohydrates not in solu-tion are subject to extensive carbon-proton dipolar broadening,and hence do not give rise to an observable "'C NMR signalunder the conditions used in these experiments.Comparison of the spectra of the ovules harvested at dif-
ferent times shows that 3 days after the 1"CO2 treatment muchof the sugar signal has disappeared, while the labeled lipidlines are considerably stronger, that is, have an increased totalintegrated intensity. Ignoring differences between ovules aboutequal in size, this comparison constitutes a qualitative measureof fatty acid synthesis as a function of the dynamics of turn-over in the sugar pool of the intact ovule. Clearly, any con-clusion based on data of this quality should not be consideredas an attempt at some kind of definitive statement about carbonmetabolism in soybeans. Rather, these experiments should beconsidered illustrative of the ease with which the I3C labelingtechnique, when combined with an NMR analysis of the intactplant material, can provide chemically useful information, andstill avoid the tedious and time-consuming chemical and phys-ical manipulations common in metabolism studies. While theestimates of rates of fatty acid synthesis obtained from theseparticular experiments require comparisons of different labeledplants, it is physically possible to design an NMR probe whichcan accept a selected soybean pod still attached to the plant.Because of the nondestructiveness of the '3C NMR analysis,
Plant Physiol. Vol. 55, 1975 1049
www.plantphysiol.orgon March 15, 2020 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.
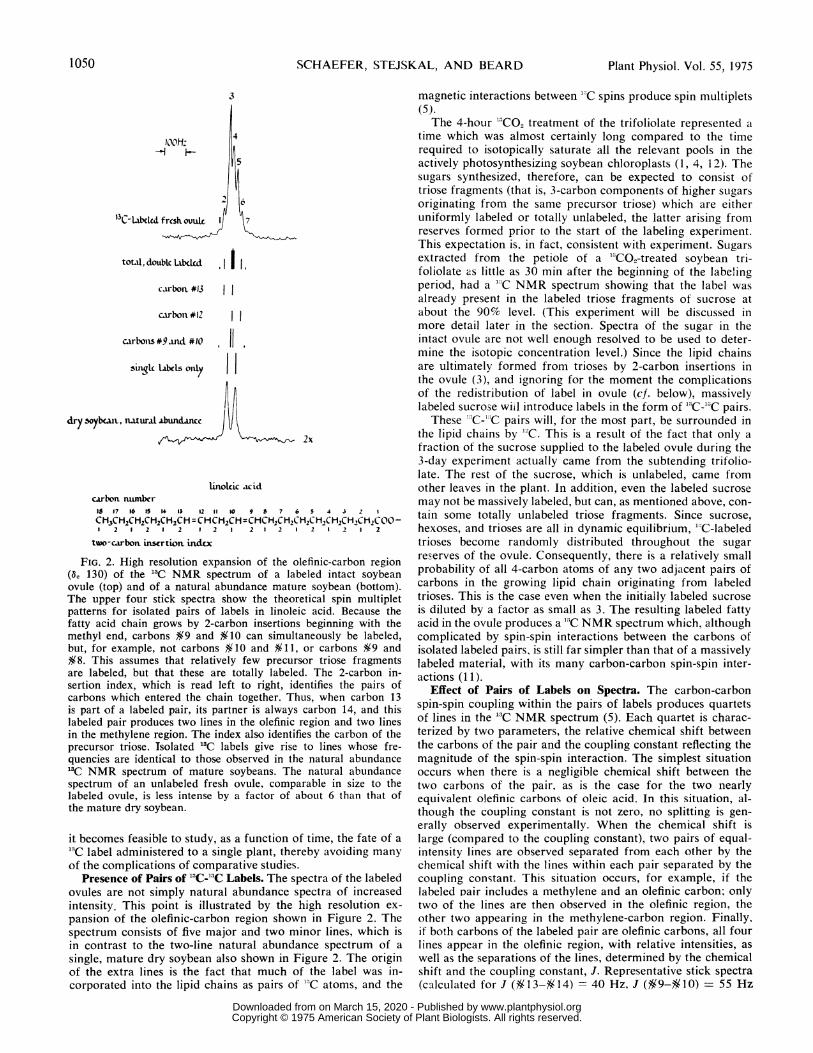
SCHAEFER, STEJSKAL, AND BEARD
3
IOOH:
13 Chc-tck frcsh ovukt
toal, doublcLitbcd. I1.
carbon.#13
carbon #12
carbonis #9and #10
sidrSnat.bl s onty
dry soybean.. naturat .a.bundancc
I I
11.
2x
tinokic acid
carbonnumbnr1 1716 IS IS 10 7 5
CH.3CH2CH2CH1=2CH1-2CH =CHCH2CHCHcH2c2CH2CH2CHC2CH2CI-H2C0-1 2 2 2 2 2
two-carbon. insertion.indix
FIG. 2. High resolution expansion of the olefinic-carbon(6c 130) of the '3C NMR spectrum of a
ovule (top) and of a natural abundance mature
The upper four stick spectra show the theoretical
patterns for isolated pairs of labels in linoleic
fatty acid chain grows by 2-carbon insertions
methyl end, carbons $9 and%£10 can simultaneously
but, for example, not carbons $10 and Y11, §O8. This assumes that relatively few precursor
are labeled, but that these are totally labeled. 2-carbon
sertion index, which is read left to right,
carbons which entered the chain together. Thus,
is part of a labeled pair, its partner is always
labeled pair produces two lines in the olefinic
in the methylene region. The index also identifies
precursor triose. Isolated13C labels give rise
quencies are identical to those observed in
'IC NMR spectrum of mature soybeans. The
spectrum of an unlabeled fresh ovule, comparable
labeled ovule, is less intense by a factor of
the mature dry soybean.
it becomes feasible to study, as a function
'3C label administered to a single plant, thereby
of the complications of comparative studies.
Presence of Pairs of13C-'3C Labels. The spectra
ovules are not simply natural abundance
intensity. This point is ilILstrated by the
pansion of the olefinic-carbon region shown
spectrum consists of five major and two
in contrast to the two-line natural abundance
single, mature dry soybean also shown in
of the extra lines is the fact that much
corporated into the lipid chains as pairs
magnetic interactions between l C spins produce spin multiplets(5).The 4-hour `2CO2 treatment of the trifoliolate represented a
time which was almost certainly long compared to the timerequired to isotopically saturate all the relevant pools in theactively photosynthesizing soybean chloroplasts (1, 4, 12). Thesugars synthesized, therefore, can be expected to consist oftriose fragments (that is, 3-carbon components of higher sugars
originating from the same precursor triose) which are eitheruniformly labeled or totally unlabeled, the latter arising fromreserves formed prior to the start of the labeling experiment.
This expectation is, in fact, consistent with experiment. Sugarsextracted from the petiole of a 'CO2-treated soybean tri-foliolate as little as 30 min after the beginning of the labelingperiod, had a'C NMR spectrum showing that the label was
already present in the labeled triose fragments of sucrose at
about the 90% level. (This experiment will be discussed in
more detail later in the section. Spectra of the sugar in theintact ovule are not well enough resolved to be used to deter-mine the isotopic concentration level.) Since the lipid chainsare ultimately formed from trioses by 2-carbon insertions inthe ovule (3), and ignoring for the moment the complicationsof the redistribution of label in ovule (cf. below), massivelylabeled sucrose wiil introduce labels in the form of'"C-';C pairs.
These C-' C pairs will, for the most part, be surrounded inthe lipid chains by'lC. This is a result of the fact that only a
fraction of the sucrose supplied to the labeled ovule during the3-day experiment actually came from the subtending trifolio-late. The rest of the sucrose, which is unlabeled, came fromother leaves in the plant. In addition, even the labeled sucrose
may not be massively labeled, but can, as mentioned above, con-
tain some totally unlabeled triose fragments. Since sucrose,
hexoses, and trioses are all in dynamic equilibrium, ':C-labeledtrioses become randomly distributed throughout the sugar
reserves of the ovule. Consequently, there is a relatively smallprobability of all 4-carbon atoms of any two adjacent pairs ofcarbons in the growing lipid chain originating from labeledtrioses. This is the case even when the initially labeled sucrose
is diluted by a factor as small as 3. The resulting labeled fattyacid in the ovule produces al;C NMR spectrum which, althoughcomplicated by spin-spin interactions between the carbons ofisolated labeled pairs, is still far simpler than that of a massivelylabeled material, with its many carbon-carbon spin-spin inter-actions(11).
Effect of Pairs of Labels on Spectra. The carbon-carbonspin-spin coupling within the pairs of labels produces quartetsof lines in the'3C NMR spectrum (5). Each quartet is charac-terized by two parameters, the relative chemical shift betweenthe carbons of the pair and the coupling constant reflecting themagnitude of the spin-spin interaction. The simplest situationoccurs when there is a negligible chemical shift between thetwo carbons of the pair, as is the case for the two nearlyequivalent olefinic carbons of oleic acid. In this situation, al-though the coupling constant is not zero, no splitting is gen-
erally observed experimentally. When the chemical shift islarge (compared to the coupling constant), two pairs of equal-intensity lines are observed separated from each other by thechemical shift with the lines within each pair separated by thecoupling constant. This situation occurs, for example, if thelabeled pair includes a methylene and an olefinic carbon; onlytwo of the lines are then observed in the olefinic region, theother two appearing in the methylene-carbon region. Finally.if both carbons of the labeled pair are olefinic carbons, all fourlines appear in the olefinic region, with relative intensities, as
well as the separations of the lines, determined by the chemicalshift and the coupling constant, J. Representative stick spectra(calculated for J (§ 13-§ 14) =40 Hz, J (§9-§ 10) --55 Hz
1050 Plant Physiol. Vol. 55, 1975
www.plantphysiol.orgon March 15, 2020 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

C-13 NMR OF SOYBEANS
(11) and relative chemical shifts as shown) illustrate what canbe expected from pair-wise labeling of linoleic acid, the majorcomponent of soybean lipid, and are shown in Figure 2. Alsoshown in that figure is a 2-carbon insertion index illustratingexactly which pairs of carbons can be labeled together for thatfatty acid.The sum total of the calculated double labeled spectra can
account for the first, second, fourth, sixth, and seventh lines ofthe observed, labeled fresh ovule spectrum. The third line, the
Table I. Calculated Contributionts to Linies 5 and 6 ofOlefinzic-Carbon Spectrum of 'IC-labeled
Soybean Ovules
13CConcnt
90
60
CarbonPair
#12-#1 1
#10X9
#12-#11
#l0-#9
IsotopicPairing
13C-13C13C-12C12C.13C12C-12C13C-13C13C-12C12C-13C
12C-12C
3C-13C13C _12C12C-13C12C-12C13C-13C13C-12C12C13,312_12C,2
RandomPairProb-ability
9 X 99X 11 X91 X 19 x 99X 11 X91 X I
664466414
X 6X 4X 6X 4X 6X 4X 6X 4
CarbonsContri-buting
toOlefinicLine 5
No.
01000100
01000100
CarbonsContri-buting
toOlefinicLine 6
No.
10
00000
100000'00
Spin-SpinMulti-plet
Factor
1
I,
00
1I
00
'2100
12
I001~I
I
0
TotalRela-tive
Contri-bution
toOlefinicLine 5
09000900
18
0240002400
48
P-D-GbkosX
strongest in the olefinic-carbon spectrum, can be assigned, inpart, to the labeled, nearly equivalent carbons of oleic acid,which is the other major lipid component present in soybeanovules in significant amounts (6). (For the purposes of thisdiscussion, the presence of relatively small amounts of linolenicacid changes none of the arguments and therefore will beignored.)The fifth line in the spectrum cannot, however, be accounted
for by the presence of pairs of labels. In the natural abundancespectrum, this line arises, for the most part, from carbons 10and 12 of linoleic acid (8). However, the natural abundancespectrum of an immature ovule having a dry weight of about50 mg can only account for about 10% of the intensity of line5. The presence of a strong line at exactly this frequency inthe labeled fresh ovule spectrum is unambiguous proof of thepresence of substantial concentrations of single '3C labels,resulting from a `C-`C 2-carbon insertion. Naturally, this kindof insertion must occur to some extent since the isotopic purityof the "CO2 used in these experiments was only 90%. Thus,some '2C02 is present to join with 'CO2 in the formation ofnewly synthesized labeled trioses. This process will result insingle labels associated with lines whose frequencies are thesame as those in the natural abundance spectrum. However,a 10% concentration of 'lCO2 leads to a prediction of 18:40for the intensity ratio of the fifth line, relative to the sixth line,of the labeled, fresh ovule olefinic-carbon spectrum. This pre-diction, obtained from Table I, is too small by a factor of about6 to account for the experimentally observed ratio.The high concentration of single '3C labels in the fatty acids
can be shown not to arise from any dilution of the label duringsugar formation in the leaf, or, as mentioned earlier, duringsugar transport into the ovule. The "C NMR spectrum ofmixed sugars extracted from leaves exposed to "CO2 forseveral hours is, as shown in Figure 3, extremely complicated.The complications are due to multiplets arising from the manypossible spin-spin interactions present in massively labeledmaterial. Nevertheless, one region of the sugar spectrum isrelatively simple. This is the part due to the anomeric carbonsof either glucose or sucrose. As illustrated in Figure 3, ananomeric carbon has only one carbon nearest neighbor, andso will produce what amounts to a doublet if that neighbor is"C (because of spin-spin coupling), or a singlet if the neighbor
1af cxtractc
InaraL. abundanceFIG. 3. Carbon-13 NMR spectrum of sugars extracted from an actively photosynthesizing soybean trifoliolate exposed to 13CO2 for several
hours (top) and of a solution of natural abundance glucose (bottom). The spread of chemical shifts in the natural abundance spectrum is 35ppm.
Plant Physiol. Vol. 55, 1975 1051
www.plantphysiol.orgon March 15, 2020 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

SCHAEFER, STEJSKAL, AND BEARD
carbon. nwolulc10C
CC-P
(,x,y .:)(X.y.z)(X,y,:)(xyz)(x,y.:)(x,y,z)
2CW(x 2C(y)3Cx.x) KOY2C('0) 3c(-)C(x) Cly)C(x) CO(y)C- P(x)J - P(y)
,.
_C( J O
P(z) CC,-PO:
3 glucosc-6-P 2 frucwosc-6-P +glyccraldchyc-3-P + 3 C02
carbon. mokecule1C203C
(II) C
C0p
(x,yJ lC(x)(x,y) 2C(x)x,y) ,:, lC(y)
3C(x)3Cly)
+ 20y)C
C CC-P C-P
2 fructosc-6-P 2 fructosc-6-PFIG. 4. Redistribution of label by the complete pentose cycle
(I) and the nondehydrogenase part of the pentose cycle (II).
is '2C. The position of the singlet will be identical to that ob-served in a natural abundance spectrum of the same sugar.Since the ratio of integrated intensity of anomeric-carbonsinglet to doublet (summed) is about 1:8, the isotopic con-centration of the labeled triose fragments of the sugar is closeto the 90% level of the labeling 3CO2. This is true even thoughthe total isotopic enrichment of the sugar as a whole may beless than 90%, due to the presence of totally unlabeled triosefragments.
Redistribution of Label. The high concentration of single'3C labels in the fatty acid is the result of a substantial, but farfrom complete, redistribution of the label in the precursorsugars due to recycling of hexose phosphates in the ovule priorto synthesis. (Turnover in synthesized storage fats and proteinsin the ovule is assumed to be negligible.) This recycling canbe due either to the complete pentose cycle (reaction I), or thenondehydrogenase part of the pentose cycle (reaction 11) (13),both of which mix labeled and unlabeled sugars together.
Reactions I and II are illustrated in Figure 4. In reaction I,if either the x-th or y-th molecules are massively labeled, butthe z-th molecule is not, then one pass will have no effect onthe concentration of pairs of "3C labels in the fatty acid chains,since only carbons 1 and 2 of a triose are ultimately involvedin the 2-carbon insertion. For the same reason, if the z-thmolecule is labeled but the other two are not, one pass willhave no effect on the fatty acid spectrum. Recycling by re-action I (that is, more than one pass) can, however, have aneffect. For example, if the x-th molecule is labeled and all theothers are not, two passes will place a 13C label in the 1-carbonposition with '2C in the 2- and 3-carbon positions. The 2-carboninsertion into a fatty acid chain will therefore result in a singleisolated "3C label. Since the total amount of label in the sugarpools of the ovule is small, we can neglect the possibility oftwo labeled molecules becoming involved within the samethree-pass sequence.
If either the x- or y-th molecule is the initially massivelylabeled species, an isolated 13C label will never be found at the2-carbon position resulting from just one or two passes throughreaction I. However, two passes will put an isolated label at the2 position, if the z-th molecule is initially labeled, with threepasses putting the label at the 1-carbon position. It is clear thatafter just a few passes the label has been redistributed between
carbon positions 1 and 2 by reaction I in a distinctly non-random fashion (13). This is in contrast to the redistributionafter many passes through reaction I, and in contrast to theredistribution resulting from reaction II, in both of whichsituations the isolated label occurs at the 1- and 2-carbonpositions with equal probability.Whether or not the isolated single label distribution is indeed
random can be determined by NMR analysis. One particularlystraightforward way in which to evaluate the randomness is tocompare the concentration of isolated single labels as evaluatedfrom the methyl-carbon region of the fatty acid spectrum(which, as shown in Figure 5, depends only upon single labelfrom the 1-carbon position) with the concentration of isolatedsingle labels as evaluated from the ratio of olefinic-carbonlines 5 and 6 (which depends upon single labels from both the1- and 2-carbon positions). The extent of the disagreementbetween the two evaluations is a measure of the nonrandomnessof the single label distribution. As long as the total amount oflabel present is small, this determination is independent of therelative proportion of '3C-labeled sugar to 12C sugar present inthe ovule pools. From Figure 5, we see that the central line ofthe methyl-carbon region is about 40% of the total intensity,indicating an effective 13C isotopic labeling concentration(after label redistribution) of 60%, rather than the 90%actually used. Thus, the calculated random value for the ratioof olefinic-carbon lines 5 and 6 is, using Table I, 48/18, or 2.7.This is within experimental error of the observed value of 3 ±0.3. Therefore, in these experiments, in order to account forthe concentration as well as the distribution of single labels,about 30% of the sugar in the ovules is totally randomized bycontact with either the nondehydrogenase part of the pentosecycle, or repeated passes through the complete cycle. In orderto account for the concentrations of pairs of labels, 70% ofall hexose phosphate either makes only one pass through the
|3c- LibcEd jO; iiV,
frcslh oviulc "i
:~I. ;
3C H3 12CH2-\ '3CHPCH --
/r4x
dry soybc.xn. C'H:n.atural abundance K | H-
2x
inuthyknc mcthyl
FIG. 5. High resolution expansion of the methylene- and methyl-carbon regions (a& 20-30) of the "3C NMR spectrum of a labeledintact soybean ovule (top) and of a natural abundance maturesoybean (bottom). Both regions of the ovule spectrum show theeffects of spin-spin coupling resulting from pairs of labels in thelipid chains. The spectrum of an unlabeled fresh ovule, com-
parable in size to the labeled ovule, is less intense by a factor ofabout 6 than that of the mature dry soybean.
1052 Plant Physiol. Vol. 55, 1975
I
I"
www.plantphysiol.orgon March 15, 2020 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

C-13 NMR OF SOYBEANS
complete cycle, or else simply avoids both the complete cycleand the nondehydrogenase part of the complete cycle.The practical value of these results is not in establishing
the fact that in synthesizing soybean ovules some of the sugaris oxidized via the pentose phosphate cycle, since this is al-ready well known. Rather, we feel the value lies in the demon-stration of the ease and speed with which semiquantitativeestimates of the extent of the oxidation could be made in anintact plant system using 'C labeling and NMR analysis. Hope-fully, more complicated, as yet unsolved, problems in carbonmetabolism can also be approached by the techniques illus-trated here.
Acknowledgment-The authors thank Prof. Richard Jensen for his commentsand criticisms.
LITERATURE CITED
1. BLACK, C. C. 1973. Photosynthetic carbon fixation in relation to net C02 UP-take. Annu. Rev. Plant Physiol. 24: 253-286.
2. FARRAR, T. C. AND E. D. BECKER. 1971. Pulse and Fourier Transform NMR.Academic Press, New York.
1053
3. GREENBERG, D. M. (ed.). 1968. Metabolic Pathways, Vol. II. Academic Press,New York.
4. JACKSON, W. A. AND R. J. VOLK. 1970. Photorespiration. Annu. Rev. PlantPhysiol. 24: 385-432.
5. POPLE, J. A., W. G. SCHNEIDER, AND H. J. BERNSTEIN. 1959. High-ResolutionNuclear Magnetic Resonance. McGraw-Hill, New York, p. 122.
6. RUBEL, A., R. W. RINNE, AND D. T. CANVINx. 1972. Protein, oil, and fattyacid in developing soybean seeds. Crop Sci. 12: 739-741.
7. SCHAEFER, J. 1972. Comparisons of the Carbon-13 nuclear magnetic resonancespectra of some solid cis- and trans-polyisoprenes. Macromolecules 5: 427-440.
8. SCHAEFER, J. AND E. 0. STEJSKAL. 1974. Carbon-13 nuclear magnetic resonancemeasurement of oil composition in single viable soybeans. J. Am. Oil Chem.Soc. 51: 210-213.
9. SEQUIN, U. AND A. IAN ScoTT. 1974. Carbon-13 as a label in biosyntheticstudies. Science 186: 101-107.
10. STEJSKAL, E. 0. AND J. SCHAEFER. 1974. Comparisons of quadrature and single-phase Fourier transform NMR. J. Magn. Res. 14: 160-170.
11. STOTHERS, J. B. 1972. Carbon-13 NMR Spectroscopy. Academic Press, NewYork.
12. VOLK, R. J. AND W. A. JACKSON. 1972. Photorespiratory phenomena in maize.Plant Physiol. 49: 218-223.
13. WOOD, H. G. AND J. KATZ. 1958. The distribution of 14C in the hexose phos-phates and the effect of recycling in the pentose cycle. J. Biol. Chem. 233:1279-1282.
Plant Physiol. Vol. 55, 1975
www.plantphysiol.orgon March 15, 2020 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.