calretinin-like immunoreactivity in mormyrid and
TRANSCRIPT

Calretinin-Like Immunoreactivityin Mormyrid and Gymnarchid
Electrosensory and ElectromotorSystems
MATTHEW A. FRIEDMAN1* AND MASASHI KAWASAKI2
1Section of Neurobiology and Behavior, Cornell University, Ithaca, New York 148532Department of Biology, University of Virginia, Charlottesville, Virginia 22903
ABSTRACTCalretinin-like immunoreactivity was examined in the electrosensory and electromotor
systems of the two families of mormyriform electric fish. Mormyrid fish showed the strongestimmunoreactivity in the knollenorgan electroreceptor pathway; in the nucleus of theelectrosensory lateral line lobe (ELL) and the big cells of the nucleus exterolateralis parsanterior. Mormyromast and ampullary zones of the ELL showed calretinin-like immunoreac-tivity in the ganglion, granule, and intermediate cell and fiber layers. Mormyromast zonesadditionally showed labeling of apical dendrites and commissural cells, but the ampullaryzone did not. In the electromotor system, two nuclei in the corollary discharge pathwayshowed labeling: in the paratrigeminal command-associated nucleus and the juxtalobarnucleus.
Gymnarchus niloticus (Gymnarchidae) showed strongest calretinin-like immunoreactiv-ity in part of the phase-coding pathway; in S-type electroreceptor afferents. Zones of the ELLnot receiving phase-coder input had weak labeling. The electromotor system showed labelingin the lateral relay nucleus and less strongly in the medullary relay nucleus, but none in thepacemaker.
The concentration of calcium-binding proteins in mormyrid and gymnarchid time-codingelectrosensory pathways is consistent with the hypothesis that they play a role in preservingtemporal information across synapses. Cell types that encode temporal characteristics ofstimuli in precise spike times have high levels of calcium-binding proteins, but cells thatre-code temporal information into presence or magnitude of activity have low levels.
Some cell types in the electromotor pathways and early in the time-coding electrosensorypathways do not follow this hypothesis, and therefore preserve temporal information using amechanism independent of calcium-binding proteins. In particular, electromotor systems mayuse extensive electrotonic coupling within nuclei to ensure precise timing. J. Comp. Neurol.387:341–357, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: temporal coding; electric fish; recoding; calcium-binding protein
In the course of sensory processing, parallel pathwaysmay become specialized to analyze different features ofsignals (Deyoe and van Essen, 1988; Konishi, 1991; Carr,1993). Some of the most striking specializations are foundin systems that analyze temporal features of signals, i.e.,so-called time-coding pathways. Time-coding pathwaysencode temporal features of stimuli by the timing ofspikes, and therefore to allow fine discrimination, spikelatencies after a stimulus vary only slightly, on the order oftens of microseconds; that is, they show low temporal‘‘jitter’’ (Carr, 1986). To reduce the introduction of jitter dueto propagation through axons and synapses, time-coding
pathways show anatomical specializations (such as heavymyelination, large axons, calyceal synapses, and aden-dritic somata) and physiological specializations (Carr,1993). One remarkable physiological specialization is thatsome time-coding pathways have higher concentrations of
Grant sponsor: NIMH; Grant numbers: MH15793, MH37972.*Correspondence to: Matthew Friedman, currently at: Department of
Neurobiology, Harvard Medical School, 220 Longwood Avenue, Boston, MA02115. E-mail: [email protected]
Received 19 April 1996; Revised 10 April 1997; Accepted 12 May 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 387:341–357 (1997)
r 1997 WILEY-LISS, INC.

calcium-binding proteins than pathways in the same sen-sory modality that analyze other stimulus features, suchas amplitude (Carr, 1986). Calcium-binding proteins havebeen found in time-coding auditory pathways of birds andmammals (Rogers, 1987; Takahashi et al., 1987; Zettel etal., 1991; Resibois and Rogers, 1992; Vater and Braun,1994) and electrosensory pathways of South Americangymnotiform fish (Maler et al., 1984; Losier and Matsub-ara, 1990).African mormyriform electric fish have indepen-dently evolved electrosensory systems that specialize inthe analysis of temporal information, and therefore wedescribe their immunoreactivity for the calcium-bindingprotein, calretinin, to obtain further support for the involve-ment of calretinin in time-coding pathways.
Calretinin is a 28 kDa protein belonging to the 6-EFhand subfamily of calcium-binding proteins (Baimbridgeet al., 1992; Andressen et al., 1993; Seto-Ohshima, 1994).Several calcium-binding proteins, including calretinin,calbindin, parvalbumin, neurocalcin, and hippocalcin, areexpressed mainly in the central nervous system (CNS) anddiffuse throughout entire neurons, making them usefulhistological markers (Seto-Ohshima, 1994). The specificimportance to time-coding pathways of intracellular cal-cium binding is not clearly understood. Faithful transmis-sion of spike times through chemical synapses may requireshort integration times and therefore short synaptic poten-tials, so that presynaptic calcium must be brought backdown to baseline levels quickly. Likewise, calcium buildupcould activate calcium-dependent channels that coulddistort repetitive signals by adaptation. Preservation of
fine temporal information may require large synapses ontolarge somata, with large pre- and post-synaptic currentsthat could cause high calcium loads that are potentiallytoxic. For example, chick magnocellularis neurons haveAMPA (a-amino-3-hydroxy-5-methyl-4-isoxazoleproprion-ate) glutamate receptors with unusually high calciumpermeability (Otis et al., 1995). The high activity of thesereceptors probably induces a high calcium load. However,the question of whether calcium-binding proteins conferresistance to excitotoxicity is unresolved (Baimbridge etal., 1992; Andressen et al., 1993).
The anatomical and physiological specializations to pre-serve temporal information in precise spike times are nolonger necessary once fine temporal discriminations havebeen made and the information is re-coded into anotherformat, such as presence or magnitude of responses, or acomputational map. In barn owls and Eigenmannia, cal-cium-binding proteins are strongly present in low-ordernuclei of time-coding pathways, and are absent in high-order nuclei (Maler et al., 1984; Carr, 1986; Takahashi etal., 1987; Losier and Matsubara, 1990).
Motor systems may also need to preserve fine temporalinformation to co-ordinate muscle fibers or muscle groups.For electric organs to generate a strong current during thedischarge, the electrocytes have to be activated simulta-neously (Bennett, 1968). If calcium-binding proteins helpin temporal fidelity, then they might also be concentratedin the electromotor system. High immunoreactivity tocalbindin was found in part of the electromotor system inEigenmannia in the medullary relay nucleus (Maler et al.,
Abbreviations
bc bulbocerebellar tractBCA bulbar command-associated nucleus (M)c commissure of ELL (G, M)C1–C4 cerebellum, lobes C1–C4 (G, M)COM command nucleus (M)cs superior cerebellar commissureDF deep fiber layer of MZ (G)dll decussation of the lateral lemniscusDLZ dorsolateral zone of ELL (M)DZ dorsal zone of ELL (G)EG eminentia granularis (G, M)EGp eminentia granularis posterior (G, M)ELa anterior exterolateral toral nucleus (M)ELL electrosensory lateral line lobe (G, M)ELp posterior exterolateral toral nucleus (M)EMN electromotorneuron (G, M)EOD electric organ dischargeGA ganglion cell layer of ELL (M)gc giant cells (G)GR granule cell layer of ELL (M)IC inner cell layer of MZ (G)ICF intermediate cell and fiber layer of ELL (M)IG isthmic granule nucleusIL inferior lobe of diencephalonJL juxtalobar nucleus (M)jlem juxtalemniscal nucleus (M)L lateral toral nucleus (G, M)LC caudal cerebellar lobe (G, M)LCmp posterior molecular zone of LC (G, M)ll lateral lemniscus (G, M)LRN lateral relay nucleus (G)MCA mesencephalic command-associated nucleus (M)MD mediodorsal toral nucleus (G, M)MF deep fiber layer of medial zone of ELL (M)MGM mesencephalic granule mass
Nomenclature for mormyrids (M) from Bell and Szabo (1986) or, where lacking, from an unpublished atlas of Gnathonemus petersii by T. Szabo.Nomenclature for Gymnarchus (G) from Bass and Hopkins (1982). All brain areas without M or G indicators have been found in mormyrids butnot indicated in Bass and Hopkins (1982) for Gymnarchus.
ML molecular layer of ELL (G, M)mlf medial longitudinal fasciculus (G, M)MLMO molecular layer of medial octavolateral nucleusMO medial octavolateral nucleusMR medullary relay nucleus (G, M)MV medioventral toral nucleus (G, M)MZ medial zone of ELL (G, M)nALL anterior lateral line nerve (G, M)NELL nucleus of the ELL (M)nPLL posterior lateral line nerve (G, M)OB olfactory bulb (G, M)OC outer cell layer of MZ (G)OT optic tectum (G, M)P plexiform layer of ELL (G, M)PCA paratrigeminal command-associated nucleus (M)PE preeminential nucleus (G, M)PN pacemaker nucleus (G)scc superior commissure in cerebellum, lobe C2spc spinocerebellar tractsublem sublemniscal nucleusTel telencephalon (G, M)TL torus longitudinalis (G, M)TM tectum mark (G, M)tp toro-preeminential tractTS torus semicircularis (G, M)V trigeminal nerve (G, M)VA valvula of cerebellum (G, M)VIII statoacoustic nerve root (G, M)VLZ ventrolateral zone of ELL (M)Vm trigeminal motor nucleus (G, M)vmc ventral mesencephalic commissureVP ventroposterior toral nucleus (G, M)vped valvular peduncle (G, M)VZ ventral zone of ELL (G)
342 M.A. FRIEDMAN AND M. KAWASAKI

1984). Furthermore, mormyrids have a corollary dischargepathway, whereby the motor command to generate adischarge travels not just to the electric organ, but also toprimary electrosensory areas to prepare them for reaffer-ent sensory information (see Fig. 2C; Bell et al., 1983; Bell,1989). If the precise temporal relationship between theactual and the expected discharge must be maintained,then calcium-binding proteins might be concentrated inthe corollary discharge pathway.
MATERIALS AND METHODS
For this study, we used nine mormyrid fish (four Brieno-myrus brachyistius, three B. niger, and two Gnathonemuspetersii, all ,10–12 cm, non-breeding individuals) andthree juveniles (11–13 cm) of Gymnarchus niloticus. Ourprotocol was approved by the Institutional Animal Careand Use Committee at Cornell University (number 85-1-95). Fish were anesthetized by immersion in MS-222(500–1000 mg/l) and then perfused through the heart withsaline (0.9% NaCl) followed by 4% buffered paraformalde-hyde. We removed the brains and post-fixed them for 1–2days in 4% buffered paraformaldehyde.
Brains to be cut on the Vibratome (Pelco 101, Redding,CA) were embedded in 4% agar just prior to sectioning andwere cut at 50 µm. Agar-embedded sections were moredifficult to handle throughout the histological procedure,so in later experiments we cut brains as gelatin-embeddedfrozen sections. Brains were cleaned of meninges, post-fixed for a second time in 4% buffered paraformaldehyde-10% sucrose for 1 day, embedded in 10% 100 bloomgelatin-10% sucrose, and post-fixed for another 1–3 days.We cut the brains on a sliding microtome (AmericanOptical, Buffalo, NY) at 50 µm.
Sections were washed three times in phosphate-bufferedsaline (PBS; 0.02 M phosphate buffer, pH 7.6, 0.9% NaCl),blocked in 5% normal goat serum (Vector Laboratories,Burlingame, CA) in PBS with 0.3–1% Triton X-100 (PBST;Aldrich Chemical Co., Milwaukee, WI), and incubated inanti-calretinin primary antibody (Swiss Antibodies, Bell-inzona, Switzerland, lot 25392) with 1% normal goatserum in PBST for 2 days. Dilutions ranged from 1:3,000 to1:14,000, all of which produced quite satisfactory results. ATriton X-100 concentration of 1% and a 30 minute wash inmethanol before blocking were necessary to facilitateantibody penetration into highly myelinated regions, suchas fiber tracts.
Sections were then washed in PBS three times andincubated for 2 hours in secondary antibody (biotinylatedgoat anti-rabbit; Vector Laboratories, Burlingame, CA) ata dilution of 1:300 with 2% normal goat serum in PBST.Sections were washed three times in PBS, incubated inavidin-biotin complex (ABC) solution (Vector Laborato-ries) for 2 hours, washed two times in PBS, and two timesin 0.1 M, pH 7.2, tris buffer (TB). Sections were soaked for15 minutes in a solution of 10 mg DAB (Sigma ChemicalCo., St. Louis, MO) in 50 ml TB. We added 30 µl 3% H2O2and let the reaction continue for 15 minutes. Finally, wewashed sections four times in TB, and then mounted,dehydrated, and coverslipped them under Permount orDepex.
To control for non-specific 2° antibody binding, represen-tative sections from one Gymnarchus and one B. brachyis-tius were treated as above, except that they were notincubated in primary antibody. None of these showed any
immunoreactivity whatsoever. From most series, we alsotook every fourth or fifth section to generate a Nissl seriesusing neutral red or cresyl violet.
The primary antibody used in this study is a polyclonalantibody raised in rabbits against recombinant humancalretinin produced in E. coli (Schwaller et al., 1993). Toverify antibody specificity in the species used here, wedeeply anesthetized one animal of each species in 0.1%MS-222 and removed their brains. We homogenized eachbrain in lysis buffer (20 mM Tris, pH 7.2, 2 mM EDTA, 25µg/ml leupeptin, 2.5 µg/ml aprotinin, 10 µg/ml pepstatin,and 1 mM pefabloc), assessed the total protein (proteinassay kit, Bio-Rad, Hercules, CA), and ran 30 µg of proteinon a 12% SDS gel. We transferred the gel to a polyscreenPVDF membrane (Dupont-NEN, Boston, MA), blocked themembrane in 4.5% nonfat dry milk, and incubated it withthe calretinin antibody at a dilution of 1:10,000 overnight.We visualized the 1° antibody by incubating the membranewith horseradish peroxidase (HRP)-conjugated mouse anti-rabbit for 1 hour, soaking it for 1 minute in DupontChemiluminescence kit, and exposing it to X-ray film for10–30 seconds. When three species showed two bands (Fig.1A), we performed another Western blot to compare onespecies (Gymnarchus) against 250 ng pure calretinin andcalbindin protein (SWant), using the calretinin antibodyand a monoclonal calbindin antibody (Sigma Chemical Co.,St. Louis, MO; 1:1,000).
Anatomical figures were transferred to a Kodak photoCD, and gels were scanned on an Epson ES-1200C scan-ner. We made minor brightness and contrast adjustmentsand applied the unsharp mask filter to the digitizedimages using Adobe Photoshop 3.0. Images were as-sembled in Macromedia Freehand 5.0.
RESULTS
Antibody specificity and staining
We verified the specificity of the calretinin antibody usedin this study by Western blot of all the species described in
Fig. 1. Calretinin antibody specificity. A: Western blot of Gymnar-chus niloticus (Gy), Gnathonemus petersii (Gp), Brienomyrus niger(Bn), and B. brachyistius (Bb), immunolabeled with anti-calretininantibody. All samples show a dark band at ,30 kDa, and Gymnarchus,G. petersii, and B. niger show an additional faint band at ,28 kDa. B:Two Western blots of Gymnarchus, compared with pure calretininprotein (cr) and pure calbindin protein (cb), immunolabeled withanti-calretinin and anti-calbindin antibodies. Both bands in Gymnar-chus (lane 1) match bands in pure calretinin (lane 2), suggesting thatthey are calretinin. The calretinin antibody also recognizes calbindin(lane 3). Gymnarchus contains a third protein recognized by theanti-calbindin antibody (lane 4), which matches the position of calbin-din (lanes 3, 5).
CALRETININ IN MORMYRIFORMS 343

anatomical detail below (Fig. 1A). In all species, theantibody recognized a band at 30 kDa (close to the reportedmolecular weight of 29 kDa) (Rogers, 1987; Schwaller etal., 1993), but a faint band at 28 kDa was visible forGymnarchus niloticus, Brienomyrus niger, and Gnathone-mus petersii.
Because we were concerned about cross-reactivity withcalbindin, we performed a Western blot of Gymnarchuswith pure calretinin protein and calbindin protein, stainedwith the calretinin antibody, and we stained a duplicatemembrane with a monoclonal calbindin antibody (Fig. 1B).We found that both bands on the fish matched bands in thepure calretinin protein lane (compare Fig. 1B, lanes 1 and2), suggesting that both bands are calretinin. However, incontrast to the results of Schwaller et al. (1993), thecalretinin antibody did recognize pure calbindin protein(Fig. 1B, lane 3), and this band was at a different molecularweight (,29 kDa) from either of the calretinin bandsobserved in Gymnarchus (Fig. 1B, lane 1). Also, we foundthat the monoclonal calbindin antibody recognized a faint29 kDa band in Gymnarchus (Fig. 1B, lane 4), whichaligned with the pure calbindin protein (Fig. 1B, lane 5),but which was distinct from either of the calretinin bands(Fig. 1B, lane 2). This result is unexpected because somestudies have reported that calbindin is not present inteleosts (Parmentier et al., 1987; Schwaller et al., 1993Arevalo et al., 1995). We could not see any bands at 29 kDalabeled by the calretinin antibody in the fish samples (Fig.1A, all lanes, and Fig. 1B, lane 1), so we conclude thatcross-reactivity is low and most of the staining reportedbelow is due to calretinin.
Histological sections generally showed well-differenti-ated staining. No within-species variation was seen in anyof the cell types described here, although there weredifferences between species as described below.
Mormyrid electrosensory areas
The four types of electroreceptors in mormyrid fish havedifferent temporal sensitivities. Since they follow distinctpathways in the CNS (Figs. 2A, 3; Szabo and Fessard,1974; Bell, 1989), they can be analyzed individually fortheir calretinin-like immunoreactivity.
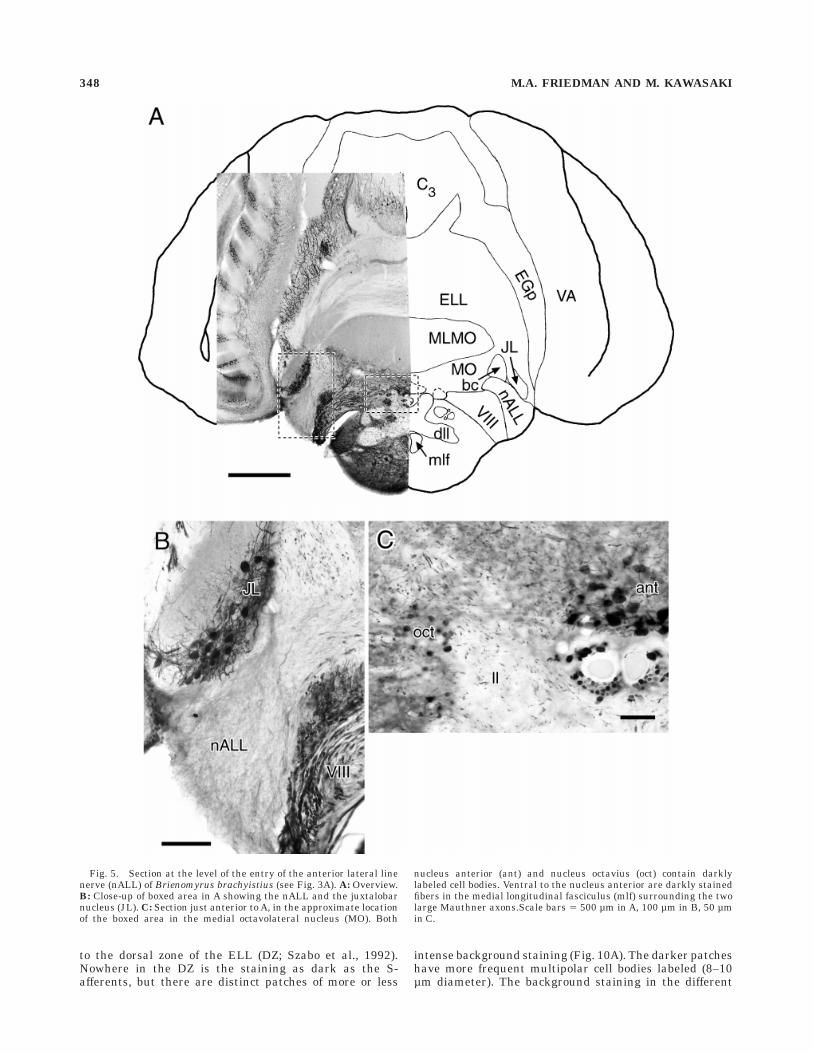
The knollenorgan electroreceptor encodes the temporalstructure of brief stimuli ( , 1 ms) with precise spikelatencies; that is, it is a time coder (Szabo et al., 1979;Amagai et al., 1993). The knollenorgan primary afferentsrun in the posterior and anterior lateral line nerves (nPLL,nALL), which show weak calretinin-like immunoreactivity(Fig. 5). This weak labeling is not a general feature ofcranial nerves, as for example the statoacoustic (VIII)nerve was darkly labeled (Fig. 5). Knollenorgan primaryafferents project to the nucleus of the ELL (NELL; Fig. 2A;Zipser and Bennett, 1976; Szabo et al., 1983; Bell et al.,1989). The NELL shows heavy calretinin-like immunoreac-tivity in Brienomyrus brachyistius and B. niger (Fig. 4).Adendritic round somata (10–15 µm diameter) are dark asare the initial segments of axons. The axons project intothe lateral lemniscus (ll), which has scattered, labeledfibers in these two species (Figs. 6–8). In Gnathonemuspetersii by contrast, although labeling in the rest of theELL is identical, the NELL and lateral lemniscus areunlabeled. Therefore, the labeled axons in the laterallemniscus of B. niger and B. brachyistius probably arisesolely from NELL somata.
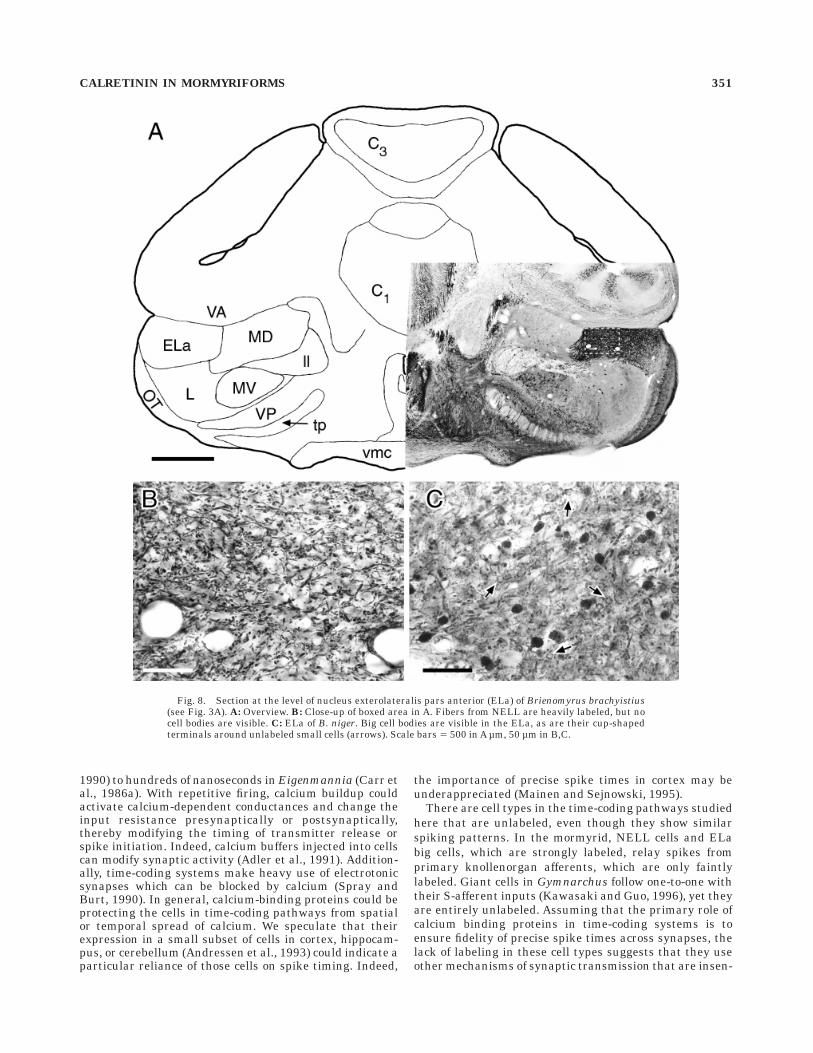
The NELL projects bilaterally to the midbrain nucleusexterolateralis pars anterior (ELa), and ends on big andsmall cell types (Fig. 2A; Mugnaini and Maler, 1987a;Amagai, 1993; Amagai et al., 1993). The ELa receives itsonly input from the NELL. The ELa is very darkly labeled(Fig. 8). In Brienomyrus brachyistius, the labeling is dueonly to fibers from the NELL (Fig. 8B). However, in B.niger, round somata of diameter 10–15 µm are also stronglylabeled. The ELa contains only two cell types (Mugnainiand Maler, 1987a), and these labeled cells are judged bytheir size to be the ELa big cells (also called ‘‘interstitialcells’’). Their characteristic cup-shaped terminals aroundthe unlabeled small cells are also labeled (diameter ,5µm; Fig. 8C). In Gnathonemus petersii, the ELa showsstrong labeling in ELa big cell somata and a dense networkof fibers, which probably arise from the big cells becausethe NELL in G. petersii was unlabeled.
ELa small cells project to the ELp, and there immunore-activity also varies across species. In B. brachyistius,labeling in the ELp is weak (Fig. 7), although thereappears the suggestion of a gradient of labeling of a fewirregularly shaped cells (6–8 µm diameter), from darkestat the lateral edge to lightest at the medial edge (Fig. 7B).In the ELp of G. petersii, a large number of round (6–10 µmdiameter) and elongate (, 5 3 10 µm) cells are lightly todarkly labeled, with one or two processes visible abovebackground. In B. niger, no ELp cells are labeled.
Mormyromast electroreceptors are used in active electro-location and detect the fish’s own electric organ discharge(EOD) as it is affected by objects near the fish (Szabo andHagiwara, 1967). The mormyromast type A, which projectsto the medial zone (MZ) of the ELL cortex, responds toimpedance-induced EOD amplitude changes, whereas themormyromast type B, which projects to the dorsolateralzone (DLZ), is more sensitive to capacitance-induced phase-shifts in the waveform (Fig. 2A; von der Emde andBleckmann, 1992). Both mormyromast types encode stimuliby using spike latency and number of spikes (Bell, 1990),and therefore spike latencies and timing differences be-tween types A and B encode electrical characteristics ofobjects near the fish.
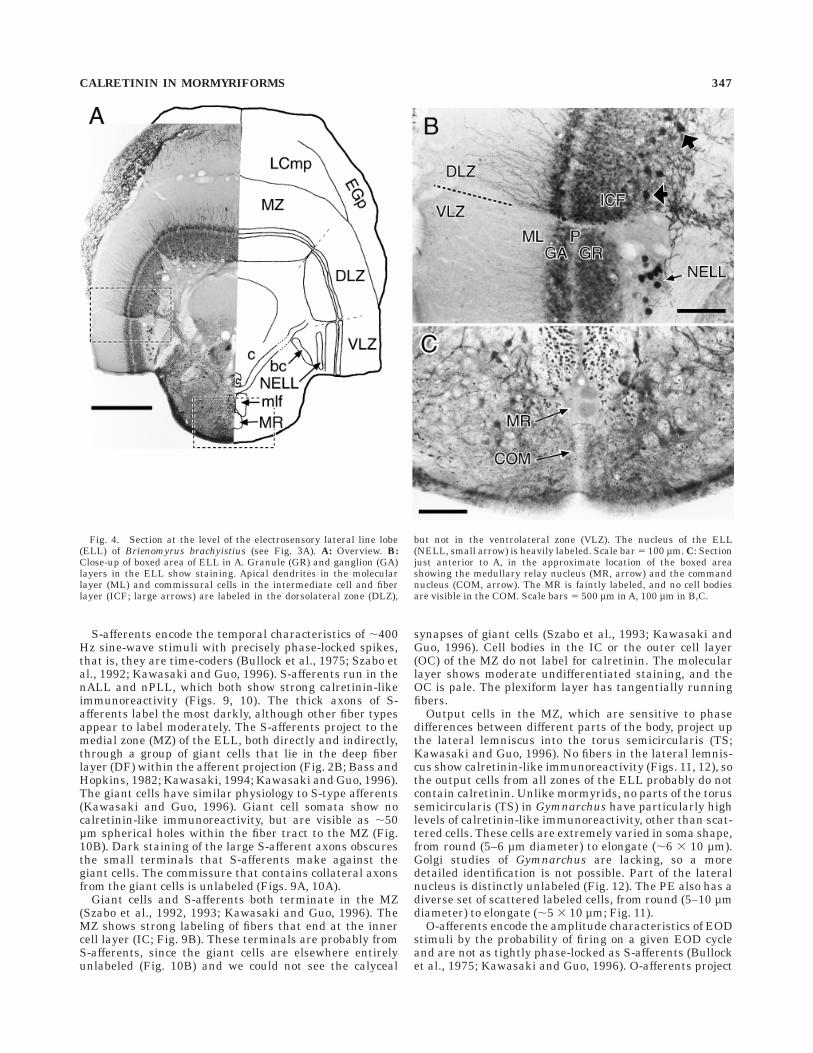
Ganglion (GA), granule (GR), and intermediate cell andfiber (ICF) layers in mormyromast zones show strongcalretinin-like immunoreactivity in all three mormyridspecies (Fig. 4). Most of the labeling is small round andoval cells (diameter 5–7 µm). The plexiform (P) layershows fibers running radially through it. We also labeledlarge round cells (diameter 10–15 µm) in the ICF, whichare probably the commissural cells that connect left andright halves of the ELL (Fig. 4; Bell et al., 1981). Addition-ally, apical dendrites in the molecular layer (ML) arelabeled (Fig. 4), which are difficult to ascribe to a particu-lar cell type. We saw no output cell types labeled.
Ampullary receptors encode the amplitudes of low-frequency electrical stimuli by spike rate and have highspontaneous activity (Bell and Russell, 1978). Therefore,the importance of precise spike times is expected to becomparatively low. Ampullary primary afferents terminatein the ventrolateral zone (VLZ). Like the mormyromastzones, many small round cells are labeled in the GA, GR,and ICF layers (Fig. 4). However, unlike the mormyromastzones, we see no label either in apical dendrites in the MLor in commissural cells in the ICF layer.
All three zones of the ELL cortex project to the preemi-nential nucleus (PE) and the lateral toral nucleus (L; Fig.
344 M.A. FRIEDMAN AND M. KAWASAKI

Fig. 2. Electromotor and electrosensory pathways in mormyrid and gymnarchid. The mormyridelectromotor pathway includes a corollary discharge pathway that affects electrosensory areas. Items inboldface were found, in this study, to show strong calretinin-like immunoreactivity. Dotted lines indicatetime-coding pathways.
CALRETININ IN MORMYRIFORMS 345

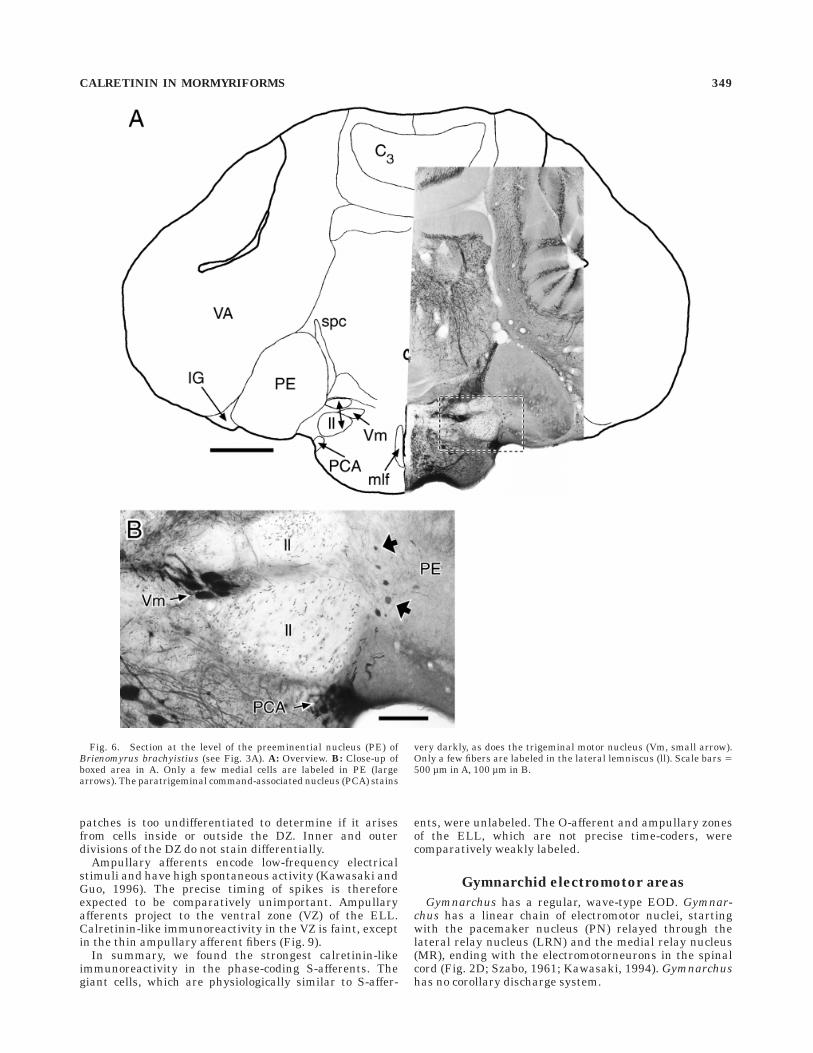
2A). In the L of all three mormyrid species, there is avaried population of round (5–12 µm diameter) to elongate(5–7 3 15–18 µm) cells, scattered at low density through-out the nucleus. They appear to be mainly multipolar, butbackground labeling precludes detailed description of dif-ferent cell types. The PE shows labeling only in about 30oval multipolar cells (,11 3 18 µm) along the medialborder for B. brachyistius and G. petersii (Fig. 6). Theyhave thin, moderately branched processes that extend outmore than 100 µm along the medial border of the PE andlaterally into the PE. Large cells in this position have beendescribed as projecting bilaterally to the L (Finger et al.,1981) and to the ELL (Bell et al., 1981), but double labelingis necessary to confirm this identification. These cells werenot labeled in B. niger.
To summarize, we found the strongest calretinin-likeimmunoreactivity in the time-coding knollenorgan path-way, in NELL cells and ELa big cells. All three zones of the
ELL cortex showed strong labeling in certain layers, butonly the mormyromast zones, which may have greaterdependence on temporal information than the ampullaryzone, also show labeling in apical dendrites in the molecu-lar layer and in the commissural cells.
Mormyrid electromotor areas
Mormyrids have irregular, pulse-type EODs. The com-mand nucleus (COM) initiates the discharge (Grant et al.,1986). It projects to the medullary relay nucleus (MR) bothdirectly and indirectly through the bulbar command-associated nucleus (BCA; Fig. 2C; Bell et al., 1983). Theelectromotor pathway does not show labeling. The COMhas densely labeled fibers running through it, but novisible cell bodies (Fig. 4C). The BCA is unlabeled, and themedullary relay nucleus (MR) is only lightly labeled(Fig. 4C).
Five nuclei have been described so far in the corollarydischarge pathway from the BCA to the electrosensoryareas (Fig. 2C; Bell et al., 1983, 1995; Bell, 1989; Bell andvon der Emde, 1995). The paratrigeminal command-associated nucleus (PCA) is very darkly labeled withsomata from round (17–20 µm diameter) to oval (,12 3 20µm), embedded in very dense fibers (Fig. 6B). The juxtalo-bar nucleus (JL) shows ,13 3 20 µm oval somata anddense fibers (Fig. 5B). No label above background is foundin the medullary command-associated nucleus, the sublem-niscal nucleus, or the juxta-lemniscal nucleus. Just thesetwo nuclei in the corollary discharge pathway, and none inthe direct electromotor pathway, show strong calretinin-like immunoreactivity.
Mormyrid auditory areas
The statoacoustic (VIII) cranial nerve showed strongcalretinin-like immunoreactivity (Fig. 5B), but because itcontains afferents from several end organs (the sacculus,the utriculus, the lagena, and the semi-circular canals;Bell, 1981a), it is not possible to tell from which end organthey originated. Two nuclei that receive input from thestatoacoustic nerve were conspicuously labeled (Fig. 5C).The anterior nucleus, which receives input from saccularand lagenar afferents (Bell, 1981a), showed strong labelingin round somata of 13–16 µm diameter (this nucleus is alsocalled the dorsal zone of the descending octaval nucleus, ordzD; McCormick, 1992; Crawford, 1997a,b). The somataappear adendritic. The nucleus octavius, which receivesinput from all octavolateral organs except electrorecep-tors, showed strong labeling in 6–9 µm diameter roundsomata. Background labeling in octavius obscured anyprocesses.
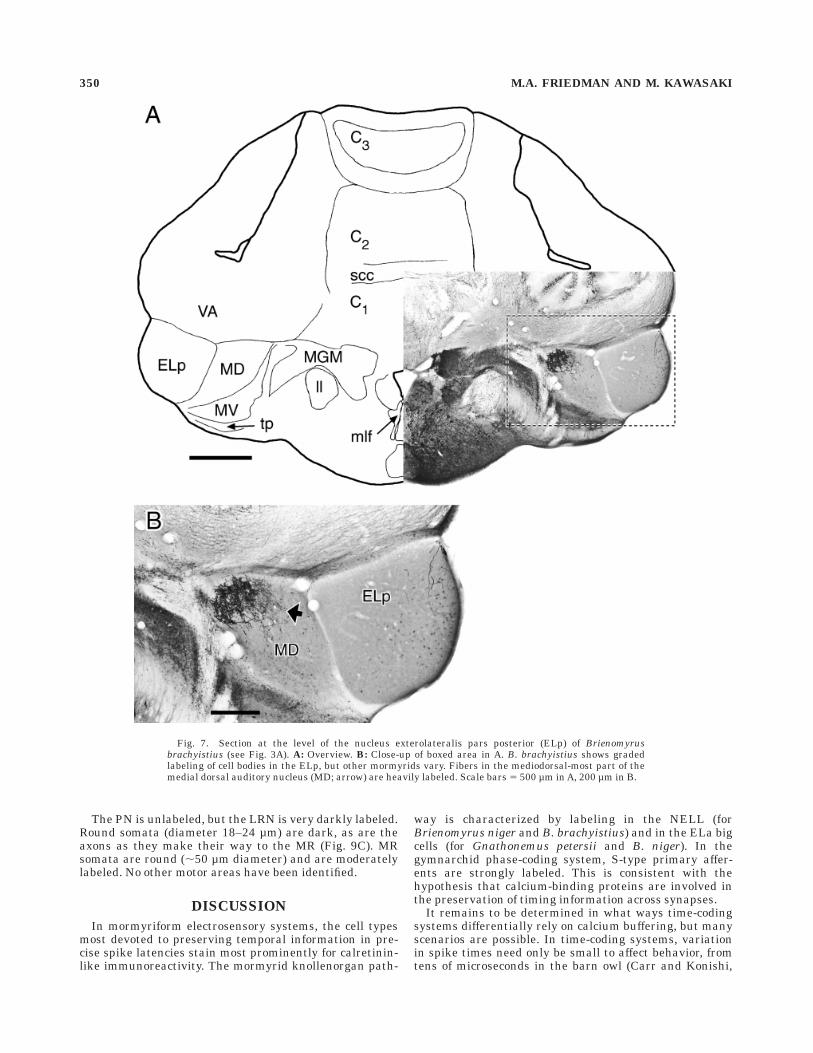
The medial octavolateral nucleus projects to the toralnucleus medialis dorsalis (MD; Bell, 1981b; Haugede-Carre, 1983), and responses to auditory stimuli have beenrecorded there (Crawford, 1993; Crawford, 1997a). In themediodorsal part of the MD, a dense network of thickfibers was labeled (Fig. 7), but no cell bodies were visible.
Gymnarchid electrosensory areas
Similarly to the mormyrid, the three types of electrore-ceptors in Gymnarchus have different temporal sensitivi-ties and follow distinct pathways in the CNS (Szabo andFessard, 1974; Bullock et al., 1975; Kawasaki and Guo,1996), so we could analyze their calretinin-like immunore-activity individually (Fig. 2B).
Fig. 3. Dorsal views of mormyrid and gymnarchid brains, with andwithout valvula, showing approximate positions of transverse slices inFigures 4–12. A: Mormyrid brain (Brienomyrus brachyistius). B:Gymnarchid brain (Gymnarchus niloticus). Drawing modified fromBass and Hopkins (1982). Scale bars 5 1 mm in A, 2 mm in B.
346 M.A. FRIEDMAN AND M. KAWASAKI
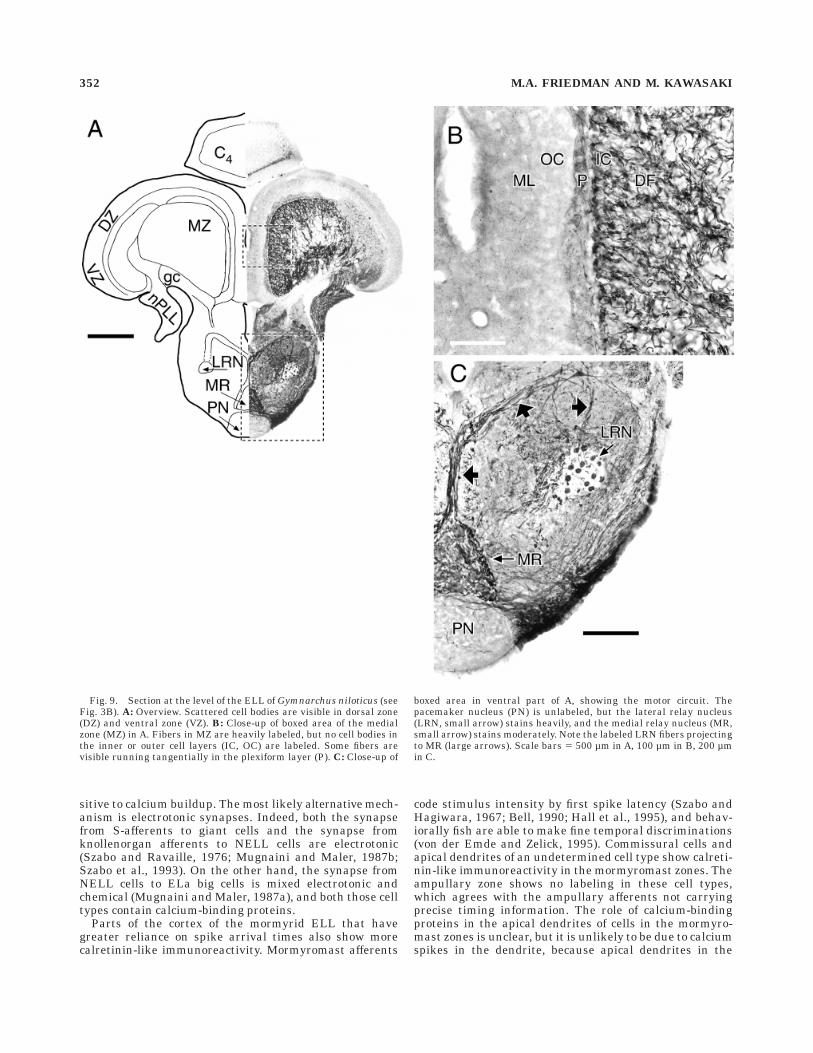
S-afferents encode the temporal characteristics of ,400Hz sine-wave stimuli with precisely phase-locked spikes,that is, they are time-coders (Bullock et al., 1975; Szabo etal., 1992; Kawasaki and Guo, 1996). S-afferents run in thenALL and nPLL, which both show strong calretinin-likeimmunoreactivity (Figs. 9, 10). The thick axons of S-afferents label the most darkly, although other fiber typesappear to label moderately. The S-afferents project to themedial zone (MZ) of the ELL, both directly and indirectly,through a group of giant cells that lie in the deep fiberlayer (DF) within the afferent projection (Fig. 2B; Bass andHopkins, 1982; Kawasaki, 1994; Kawasaki and Guo, 1996).The giant cells have similar physiology to S-type afferents(Kawasaki and Guo, 1996). Giant cell somata show nocalretinin-like immunoreactivity, but are visible as ,50µm spherical holes within the fiber tract to the MZ (Fig.10B). Dark staining of the large S-afferent axons obscuresthe small terminals that S-afferents make against thegiant cells. The commissure that contains collateral axonsfrom the giant cells is unlabeled (Figs. 9A, 10A).
Giant cells and S-afferents both terminate in the MZ(Szabo et al., 1992, 1993; Kawasaki and Guo, 1996). TheMZ shows strong labeling of fibers that end at the innercell layer (IC; Fig. 9B). These terminals are probably fromS-afferents, since the giant cells are elsewhere entirelyunlabeled (Fig. 10B) and we could not see the calyceal
synapses of giant cells (Szabo et al., 1993; Kawasaki andGuo, 1996). Cell bodies in the IC or the outer cell layer(OC) of the MZ do not label for calretinin. The molecularlayer shows moderate undifferentiated staining, and theOC is pale. The plexiform layer has tangentially runningfibers.
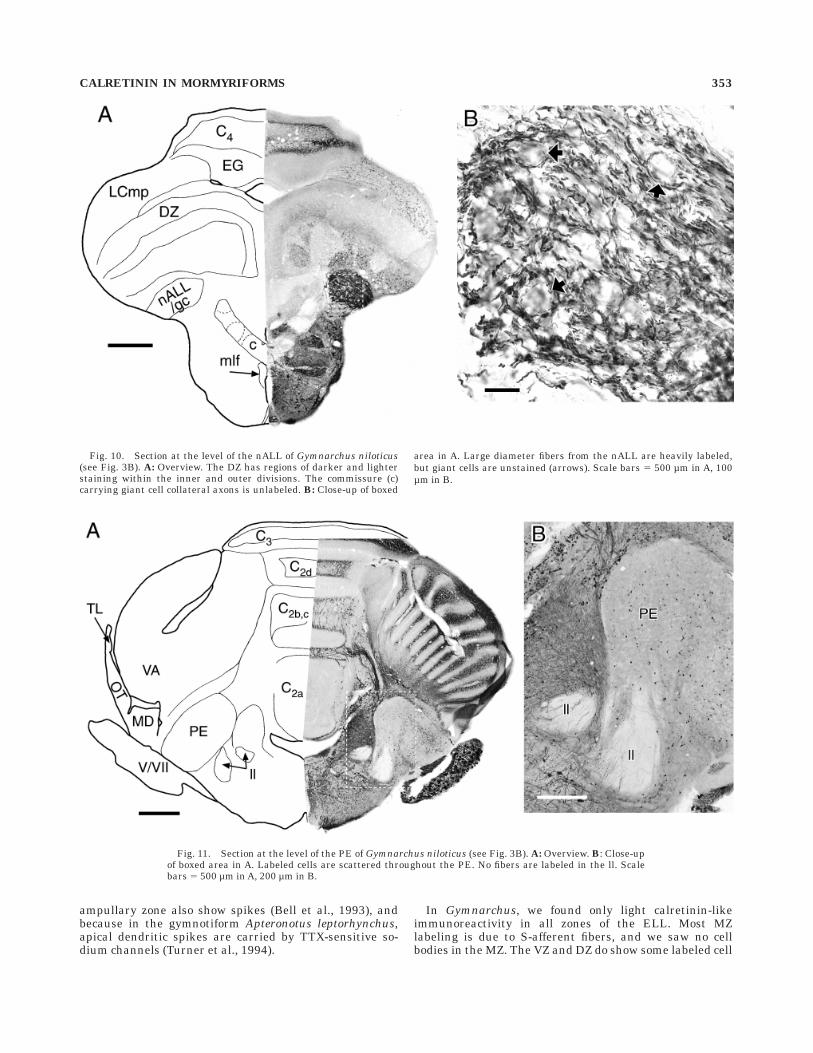
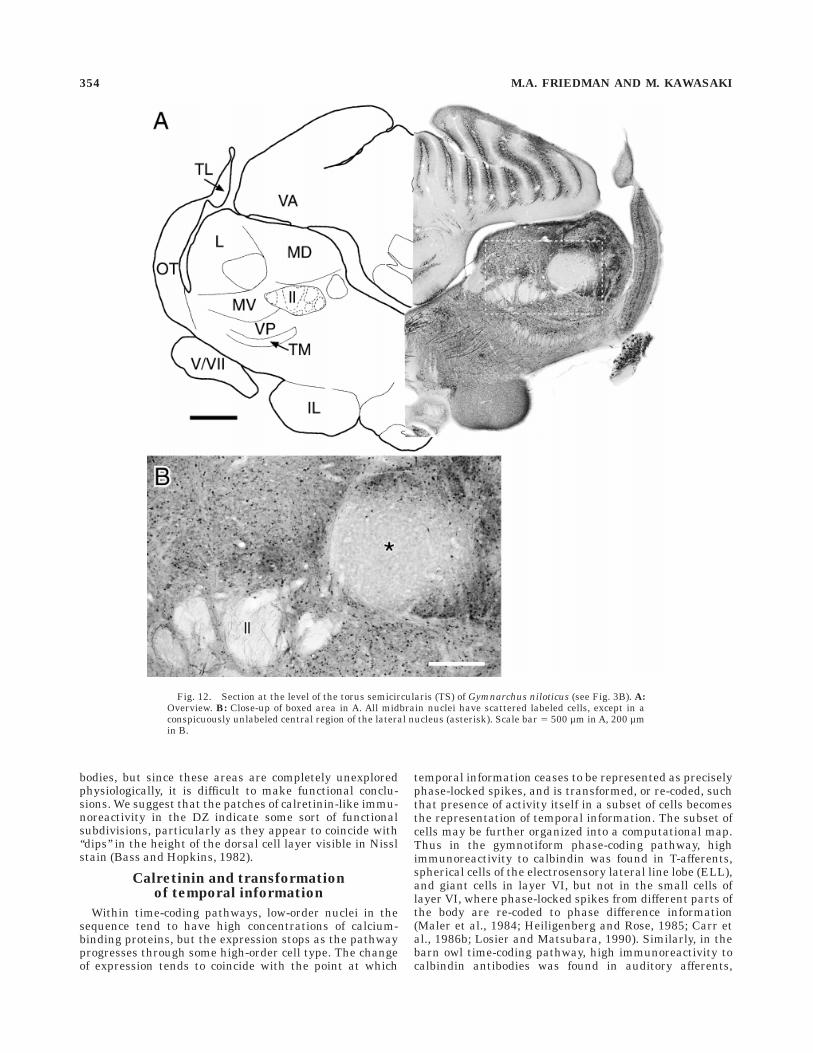
Output cells in the MZ, which are sensitive to phasedifferences between different parts of the body, project upthe lateral lemniscus into the torus semicircularis (TS;Kawasaki and Guo, 1996). No fibers in the lateral lemnis-cus show calretinin-like immunoreactivity (Figs. 11, 12), sothe output cells from all zones of the ELL probably do notcontain calretinin. Unlike mormyrids, no parts of the torussemicircularis (TS) in Gymnarchus have particularly highlevels of calretinin-like immunoreactivity, other than scat-tered cells. These cells are extremely varied in soma shape,from round (5–6 µm diameter) to elongate (,6 3 10 µm).Golgi studies of Gymnarchus are lacking, so a moredetailed identification is not possible. Part of the lateralnucleus is distinctly unlabeled (Fig. 12). The PE also has adiverse set of scattered labeled cells, from round (5–10 µmdiameter) to elongate (,5 3 10 µm; Fig. 11).
O-afferents encode the amplitude characteristics of EODstimuli by the probability of firing on a given EOD cycleand are not as tightly phase-locked as S-afferents (Bullocket al., 1975; Kawasaki and Guo, 1996). O-afferents project
Fig. 4. Section at the level of the electrosensory lateral line lobe(ELL) of Brienomyrus brachyistius (see Fig. 3A). A: Overview. B:Close-up of boxed area of ELL in A. Granule (GR) and ganglion (GA)layers in the ELL show staining. Apical dendrites in the molecularlayer (ML) and commissural cells in the intermediate cell and fiberlayer (ICF; large arrows) are labeled in the dorsolateral zone (DLZ),
but not in the ventrolateral zone (VLZ). The nucleus of the ELL(NELL, small arrow) is heavily labeled. Scale bar 5 100 µm. C: Sectionjust anterior to A, in the approximate location of the boxed areashowing the medullary relay nucleus (MR, arrow) and the commandnucleus (COM, arrow). The MR is faintly labeled, and no cell bodiesare visible in the COM. Scale bars 5 500 µm in A, 100 µm in B,C.
CALRETININ IN MORMYRIFORMS 347
to the dorsal zone of the ELL (DZ; Szabo et al., 1992).Nowhere in the DZ is the staining as dark as the S-afferents, but there are distinct patches of more or less
intense background staining (Fig. 10A). The darker patcheshave more frequent multipolar cell bodies labeled (8–10µm diameter). The background staining in the different
Fig. 5. Section at the level of the entry of the anterior lateral linenerve (nALL) of Brienomyrus brachyistius (see Fig. 3A). A: Overview.B: Close-up of boxed area in A showing the nALL and the juxtalobarnucleus (JL). C: Section just anterior to A, in the approximate locationof the boxed area in the medial octavolateral nucleus (MO). Both
nucleus anterior (ant) and nucleus octavius (oct) contain darklylabeled cell bodies. Ventral to the nucleus anterior are darkly stainedfibers in the medial longitudinal fasciculus (mlf) surrounding the twolarge Mauthner axons.Scale bars 5 500 µm in A, 100 µm in B, 50 µmin C.
348 M.A. FRIEDMAN AND M. KAWASAKI
patches is too undifferentiated to determine if it arisesfrom cells inside or outside the DZ. Inner and outerdivisions of the DZ do not stain differentially.
Ampullary afferents encode low-frequency electricalstimuli and have high spontaneous activity (Kawasaki andGuo, 1996). The precise timing of spikes is thereforeexpected to be comparatively unimportant. Ampullaryafferents project to the ventral zone (VZ) of the ELL.Calretinin-like immunoreactivity in the VZ is faint, exceptin the thin ampullary afferent fibers (Fig. 9).
In summary, we found the strongest calretinin-likeimmunoreactivity in the phase-coding S-afferents. Thegiant cells, which are physiologically similar to S-affer-
ents, were unlabeled. The O-afferent and ampullary zonesof the ELL, which are not precise time-coders, werecomparatively weakly labeled.
Gymnarchid electromotor areas
Gymnarchus has a regular, wave-type EOD. Gymnar-chus has a linear chain of electromotor nuclei, startingwith the pacemaker nucleus (PN) relayed through thelateral relay nucleus (LRN) and the medial relay nucleus(MR), ending with the electromotorneurons in the spinalcord (Fig. 2D; Szabo, 1961; Kawasaki, 1994). Gymnarchushas no corollary discharge system.
Fig. 6. Section at the level of the preeminential nucleus (PE) ofBrienomyrus brachyistius (see Fig. 3A). A: Overview. B: Close-up ofboxed area in A. Only a few medial cells are labeled in PE (largearrows). The paratrigeminal command-associated nucleus (PCA) stains
very darkly, as does the trigeminal motor nucleus (Vm, small arrow).Only a few fibers are labeled in the lateral lemniscus (ll). Scale bars 5500 µm in A, 100 µm in B.
CALRETININ IN MORMYRIFORMS 349
The PN is unlabeled, but the LRN is very darkly labeled.Round somata (diameter 18–24 µm) are dark, as are theaxons as they make their way to the MR (Fig. 9C). MRsomata are round (,50 µm diameter) and are moderatelylabeled. No other motor areas have been identified.
DISCUSSION
In mormyriform electrosensory systems, the cell typesmost devoted to preserving temporal information in pre-cise spike latencies stain most prominently for calretinin-like immunoreactivity. The mormyrid knollenorgan path-
way is characterized by labeling in the NELL (forBrienomyrus niger and B. brachyistius) and in the ELa bigcells (for Gnathonemus petersii and B. niger). In thegymnarchid phase-coding system, S-type primary affer-ents are strongly labeled. This is consistent with thehypothesis that calcium-binding proteins are involved inthe preservation of timing information across synapses.
It remains to be determined in what ways time-codingsystems differentially rely on calcium buffering, but manyscenarios are possible. In time-coding systems, variationin spike times need only be small to affect behavior, fromtens of microseconds in the barn owl (Carr and Konishi,
Fig. 7. Section at the level of the nucleus exterolateralis pars posterior (ELp) of Brienomyrusbrachyistius (see Fig. 3A). A: Overview. B: Close-up of boxed area in A. B. brachyistius shows gradedlabeling of cell bodies in the ELp, but other mormyrids vary. Fibers in the mediodorsal-most part of themedial dorsal auditory nucleus (MD; arrow) are heavily labeled. Scale bars 5 500 µm in A, 200 µm in B.
350 M.A. FRIEDMAN AND M. KAWASAKI
1990) to hundreds of nanoseconds in Eigenmannia (Carr etal., 1986a). With repetitive firing, calcium buildup couldactivate calcium-dependent conductances and change theinput resistance presynaptically or postsynaptically,thereby modifying the timing of transmitter release orspike initiation. Indeed, calcium buffers injected into cellscan modify synaptic activity (Adler et al., 1991). Addition-ally, time-coding systems make heavy use of electrotonicsynapses which can be blocked by calcium (Spray andBurt, 1990). In general, calcium-binding proteins could beprotecting the cells in time-coding pathways from spatialor temporal spread of calcium. We speculate that theirexpression in a small subset of cells in cortex, hippocam-pus, or cerebellum (Andressen et al., 1993) could indicate aparticular reliance of those cells on spike timing. Indeed,
the importance of precise spike times in cortex may beunderappreciated (Mainen and Sejnowski, 1995).
There are cell types in the time-coding pathways studiedhere that are unlabeled, even though they show similarspiking patterns. In the mormyrid, NELL cells and ELabig cells, which are strongly labeled, relay spikes fromprimary knollenorgan afferents, which are only faintlylabeled. Giant cells in Gymnarchus follow one-to-one withtheir S-afferent inputs (Kawasaki and Guo, 1996), yet theyare entirely unlabeled. Assuming that the primary role ofcalcium binding proteins in time-coding systems is toensure fidelity of precise spike times across synapses, thelack of labeling in these cell types suggests that they useother mechanisms of synaptic transmission that are insen-
Fig. 8. Section at the level of nucleus exterolateralis pars anterior (ELa) of Brienomyrus brachyistius(see Fig. 3A). A: Overview. B: Close-up of boxed area in A. Fibers from NELL are heavily labeled, but nocell bodies are visible. C: ELa of B. niger. Big cell bodies are visible in the ELa, as are their cup-shapedterminals around unlabeled small cells (arrows). Scale bars 5 500 in A µm, 50 µm in B,C.
CALRETININ IN MORMYRIFORMS 351
sitive to calcium buildup. The most likely alternative mech-anism is electrotonic synapses. Indeed, both the synapsefrom S-afferents to giant cells and the synapse fromknollenorgan afferents to NELL cells are electrotonic(Szabo and Ravaille, 1976; Mugnaini and Maler, 1987b;Szabo et al., 1993). On the other hand, the synapse fromNELL cells to ELa big cells is mixed electrotonic andchemical (Mugnaini and Maler, 1987a), and both those celltypes contain calcium-binding proteins.
Parts of the cortex of the mormyrid ELL that havegreater reliance on spike arrival times also show morecalretinin-like immunoreactivity. Mormyromast afferents
code stimulus intensity by first spike latency (Szabo andHagiwara, 1967; Bell, 1990; Hall et al., 1995), and behav-iorally fish are able to make fine temporal discriminations(von der Emde and Zelick, 1995). Commissural cells andapical dendrites of an undetermined cell type show calreti-nin-like immunoreactivity in the mormyromast zones. Theampullary zone shows no labeling in these cell types,which agrees with the ampullary afferents not carryingprecise timing information. The role of calcium-bindingproteins in the apical dendrites of cells in the mormyro-mast zones is unclear, but it is unlikely to be due to calciumspikes in the dendrite, because apical dendrites in the
Fig. 9. Section at the level of the ELL of Gymnarchus niloticus (seeFig. 3B). A: Overview. Scattered cell bodies are visible in dorsal zone(DZ) and ventral zone (VZ). B: Close-up of boxed area of the medialzone (MZ) in A. Fibers in MZ are heavily labeled, but no cell bodies inthe inner or outer cell layers (IC, OC) are labeled. Some fibers arevisible running tangentially in the plexiform layer (P). C: Close-up of
boxed area in ventral part of A, showing the motor circuit. Thepacemaker nucleus (PN) is unlabeled, but the lateral relay nucleus(LRN, small arrow) stains heavily, and the medial relay nucleus (MR,small arrow) stains moderately. Note the labeled LRN fibers projectingto MR (large arrows). Scale bars 5 500 µm in A, 100 µm in B, 200 µmin C.
352 M.A. FRIEDMAN AND M. KAWASAKI
ampullary zone also show spikes (Bell et al., 1993), andbecause in the gymnotiform Apteronotus leptorhynchus,apical dendritic spikes are carried by TTX-sensitive so-dium channels (Turner et al., 1994).
In Gymnarchus, we found only light calretinin-likeimmunoreactivity in all zones of the ELL. Most MZlabeling is due to S-afferent fibers, and we saw no cellbodies in the MZ. The VZ and DZ do show some labeled cell
Fig. 10. Section at the level of the nALL of Gymnarchus niloticus(see Fig. 3B). A: Overview. The DZ has regions of darker and lighterstaining within the inner and outer divisions. The commissure (c)carrying giant cell collateral axons is unlabeled. B: Close-up of boxed
area in A. Large diameter fibers from the nALL are heavily labeled,but giant cells are unstained (arrows). Scale bars 5 500 µm in A, 100µm in B.
Fig. 11. Section at the level of the PE of Gymnarchus niloticus (see Fig. 3B). A: Overview. B: Close-upof boxed area in A. Labeled cells are scattered throughout the PE. No fibers are labeled in the ll. Scalebars 5 500 µm in A, 200 µm in B.
CALRETININ IN MORMYRIFORMS 353
bodies, but since these areas are completely unexploredphysiologically, it is difficult to make functional conclu-sions. We suggest that the patches of calretinin-like immu-noreactivity in the DZ indicate some sort of functionalsubdivisions, particularly as they appear to coincide with‘‘dips’’ in the height of the dorsal cell layer visible in Nisslstain (Bass and Hopkins, 1982).
Calretinin and transformationof temporal information
Within time-coding pathways, low-order nuclei in thesequence tend to have high concentrations of calcium-binding proteins, but the expression stops as the pathwayprogresses through some high-order cell type. The changeof expression tends to coincide with the point at which
temporal information ceases to be represented as preciselyphase-locked spikes, and is transformed, or re-coded, suchthat presence of activity itself in a subset of cells becomesthe representation of temporal information. The subset ofcells may be further organized into a computational map.Thus in the gymnotiform phase-coding pathway, highimmunoreactivity to calbindin was found in T-afferents,spherical cells of the electrosensory lateral line lobe (ELL),and giant cells in layer VI, but not in the small cells oflayer VI, where phase-locked spikes from different parts ofthe body are re-coded to phase difference information(Maler et al., 1984; Heiligenberg and Rose, 1985; Carr etal., 1986b; Losier and Matsubara, 1990). Similarly, in thebarn owl time-coding pathway, high immunoreactivity tocalbindin antibodies was found in auditory afferents,
Fig. 12. Section at the level of the torus semicircularis (TS) of Gymnarchus niloticus (see Fig. 3B). A:Overview. B: Close-up of boxed area in A. All midbrain nuclei have scattered labeled cells, except in aconspicuously unlabeled central region of the lateral nucleus (asterisk). Scale bar 5 500 µm in A, 200 µmin B.
354 M.A. FRIEDMAN AND M. KAWASAKI

nucleus magnocellularis neurons, and nucleus laminarisneurons, where time-of-arrival of sounds at the two ears isre-coded into a place map of interaural delay, but not in thepostsynaptic targets of nucleus laminaris (Carr, 1986;Takahashi et al., 1987; Carr and Konishi, 1990). Theimmunohistochemical border between presence and ab-sence of calcium-binding proteins correlates with thefunctional border between representing fine temporal infor-mation in spike arrival times and re-coding it into acomputational map.
As this study shows, in some mormyrids, NELL cells(Fig. 4) and ELa big cells (Fig. 8) had calretinin-likeimmunoreactivity, but the ELa small cells did not. Thesmall cells have been hypothesized to re-code temporalinformation because they are targets of an excitatorydirect input from the NELL and a presumably inhibitoryindirect input through the ELa big cells (Mugnaini andMaler, 1987a; Amagai, 1993). Small cells also lack many ofthe anatomical specializations that correlate with preser-vation of fine temporal information: they have smallsomata, thin axons, less heavy myelination, and smallsynapses (Mugnaini and Maler, 1987a; Friedman, 1995).Lack of calretinin-like immunoreactivity reinforces thehypothesis that ELa small cells re-code temporal informa-tion.
A parallel case appears in the mormyrid auditory sys-tem. Statoacoustic nerve afferents (Fig. 5B), cells in thehindbrain medial octavolateral nucleus (Fig. 5C), andfibers in the mediodorsal-most part of the midbrain MD(Fig. 7) showed calretinin-like immunoreactivity, but so-mata in the MD did not. On the basis of calretinin-likeimmunoreactivity, therefore, this pathway may processtemporal characteristics of auditory signals, in whichre-coding occurs in MD. In fact, in a related species ofmormyrid, Pollimyrus, this part of MD has been found tohave complex response properties to auditory stimuli, suchas responding to particular temporal characteristics (inter-click interval; Crawford, 1993, 1997a).
The relationship between calretinin-like immunoreactiv-ity and re-coding is less clear in Gymnarchus; however.Kawasaki and Guo (1996) showed that giant cells have thesame phase-locked spike pattern as S-type afferents, butthe giant cells do not label for calretinin (Fig. 10). In thissystem, re-coding is performed by cells in the inner celllayer in the medial zone, which responds to phase differ-ences between distant parts of the body, by using acomputational algorithm similar to that of the small cellsof Eigenmannia (Kawasaki, 1993). Giant cells may nothave high levels of calretinin because they contain adifferent calcium binding protein, because they accumu-late less calcium, or because it affects them less, forunknown reasons.
Electromotor systems
Electromotor systems also require preservation of tim-ing information to ensure that the electrocytes are acti-vated simultaneously. In Gymnarchus, the LRN has highcalretinin-like immunoreactivity (Fig. 9), and the MR isalso somewhat labeled, which is consistent with the hypoth-esis that calretinin helps in the preservation of temporalinformation from the PN through the LRN to the MR.However, the electromotor system of mormyrids does notshow labeling in the COM, BCA, or MR. The electromotorsystem of mormyrids may use mechanisms to preservetemporal information that do not require calcium-binding
proteins. In particular, cells within the COM and the MRin mormyrids are strongly coupled electrotonically, and thesynapse between the COM and the MR is mixed chemicaland electrotonic (Bennett et al., 1967; Elekes and Szabo,1985, 1986; Grant et al., 1986). Since all the neurons in anelectromotor nucleus should be active simultaneously,electrotonic coupling can increase fidelity without degrad-ing utility. Interestingly, in the gymnotiform Eigenman-nia, MR cells are not electrotonically coupled with eachother (Elekes and Szabo, 1981), and they are heavilycalbindin immunoreactive (Maler et al., 1984).
Unlike motor systems, sensory systems cannot increasetemporal fidelity by electrotonically coupling units withinone level of a sensory pathway, because this mechanismwould degrade spatial resolution. Sensory systems aretherefore constrained to optimize synapses to preservetemporal information across them. Sensory systems do useconvergence, but this is primarily effective for reducingtemporal variation among peripheral receptors, whosesensitivity makes them prone to noise (Carr et al., 1986a;Carr, 1993). Convergence sacrifices spatial resolution forsensitivity. Indeed, reducing jitter through convergencerelies on temporal fidelity of synapses so that no additionaltemporal noise is introduced at the synapse (Amagai,1993; Carr and Amagai, 1996).
Mormyrids show significant labeling in only two nucleiof the corollary discharge pathway. The PCA (Fig. 6B) andthe JL (Fig. 5B) both show staining, but no interveningnuclei show staining. The unlabeled nuclei may preservetemporal information with a mechanism that does notrequire calretinin, such as using electrotonic coupling, orthey may not have the same requirement for temporalfidelity. The ultrastructure of the corollary discharge path-way has not been described. It is interesting to note thatthe two nuclei that do show calretinin-like immunoreactiv-ity project out of the corollary discharge pathway. Onemight speculate that the synaptic specializations the restof the pathway uses to insure temporal fidelity may not beavailable at this juncture.
Species differences in calretinin expression
We found species differences in calretinin-like immuno-reactivity in NELL cells, ELa big cells (Fig. 8) and cells inELp. Species differences in calcium-binding proteins arecommon (Baimbridge et al., 1992). Losier and Matsubara(1990) found that calbindin immunoreactivity in the gym-notiform electrosensory time-coding pathway correlatedwith EOD discharge rate. Apteronotus (500–1,000 Hz) andEigenmannia (250–500 Hz) both had strong reactivity, butthe low-frequency Sternopygus (50–150 Hz) had none. Thisis consistent with the hypothesis that calcium-bindingproteins keep intracellular calcium low, since time-codingunits in Apteronotus and Eigenmannia fire more fre-quently than in Sternopygus, and therefore could beaccumulating higher calcium loads. In the mormyridsstudied here, however, EOD characteristics are very simi-lar. All three species tested have similar EOD duration(,0.5 ms), peak power content (3.5–5 kHz), and interpulseintervals.
Part of the variation in expression could be that othercalcium-binding proteins are expressed in place of calreti-nin in calretinin-negative areas. Assessing the expressionof different calcium-binding proteins has historically hadtrouble due to antibody specificity problems. Antibodies tocalbindin have recognized more than one protein, such as
CALRETININ IN MORMYRIFORMS 355

calretinin (Rogers, 1987) or perhaps a second ‘‘form’’ ofcalbindin (Parmentier et al, 1987). So, for example, thecalbindin reactivity in the barn owl described by Takaha-shi et al. (1987) was probably due to cross-reactivity withcalretinin (Rogers, 1987). Likewise, calbindin reactivity inEigenmannia reported by Maler et al. (1984) and Losierand Matsubara (1990) overlaps with reactivity to thecalretinin antibody used in this study (Calvin Wong,personal communication). As we show here (Fig. 1), thisantibody has similar problems, although the cross-reactivity is low. Despite reports that antibodies againstcalretinin or calbindin recognize the same single band inWestern blots of teleosts (Parmentier et al., 1987; Schwalleret al., 1993), we found evidence of both calretinin andcalbindin in Gymnarchus niloticus. The expression offurther calcium-binding proteins in mormyrid and gymnar-chid fish will require more study.
ACKNOWLEDGMENTS
We thank Carl Hopkins for support and help throughoutthis project, Glenn Northcutt for introducing us to thisantibody, Calvin Wong and Grace Kennedy for help withthe antibody staining protocol, Margie Nelson for themormyrid brain drawing, Andy Bass for information aboutGymnarchus, Rufeng Xu for help with the Western blots,and Garry Harned for technical help. We also thank twoanonymous reviewers for their comments. This work wassupported by NIMH MH15793 to M.A.F. and MH37972 toCarl Hopkins.
LITERATURE CITED
Adler, E.M., G.J. Augustine, S.N. Duffy, and M.P. Charlton (1991) Alienintracellular calcium chelators attenuate neurotransmitter release atthe squid giant synapse. J. Neurosci. 11:1496–1507.
Amagai, S. (1993) Neural representation of time-domain information in theelectrosensory system of a mormyrid fish. PhD Thesis, Cornell Univer-sity.
Amagai, S., M.A. Friedman, and C.D. Hopkins (1993) Anatomy andphysiology of cells in the time-coding pathway of mormyrid electric fishI: Nucleus exterolateralis pars anterior. J. Comp. Physiol. A 173:747.
Andressen, C., I. Blumcke, and M.R. Celio (1993) Calcium-binding proteins:Selective markers of nerve cells. Cell Tissue Res. 271:181–208.
Arevalo, R., J.R. Alonso, A. Porteros, J.G. Brinon, C. Crespo, J. Lara, and J.Aijon (1995) Calretinin-like immunoreactivity in the optic tectum of thetench (Tinca tinca L.). Brain Res. 671:112–118.
Baimbridge, K.G., M.R. Celio, and J.H. Rogers (1992) Calcium-bindingproteins in the nervous system. TINS 15:303–308.
Bass, A.H., and C.D. Hopkins (1982) Comparative aspects of brain organiza-tion of an African ‘‘wave’’ electric fish, Gymnarchus niloticus. J. Mor-phol. 174:313–334.
Bell, C.C. (1981a) Central distribution of octavolateral afferents andefferents in a teleost (Mormyridae). J. Comp. Neurol. 195:391–414.
Bell, C.C. (1981b) Some central connections of medullary octavolateralcenters in a mormyrid fish. In W.N. Tavolga, A.N. Popper, and R.R. Fay(eds): Hearing and Sound Communication in Fishes. New York: Springer-Verlag, pp. 383–391.
Bell, C.C. (1989) Sensory coding and corollary discharge effects in mor-myrid electric fish. J. Exp. Biol. 146:229–253.
Bell, C.C. (1990) Mormyromast electroreceptor organs and their afferentfibers in mormyrid fish. III. Physiological differences between twomorphological types of fibers. J. Neurophysiol. 63:319–332.
Bell, C.C., A. Caputi, K. Grant, and J. Serrier (1993) Storage of a sensorypattern by anti-Hebbian synaptic plasticity in an electric fish. Proc.Natl. Acad. Sci. U.S.A. 90:4650–4654.
Bell, C.C., K. Dunn, C. Hall, and A. Caputi (1995) Electric organ corollarydischarge pathways in mormyrid fish. I. The mesencephalic commandassociated nucleus. J. Comp. Physiol. A 177:449–462.
Bell, C.C., T.E. Finger, and C.J. Russell (1981) Central connections of theposterior lateral line lobe in mormyrid fish. Exp. Brain Res. 42:9–22.
Bell, C.C., S. Libouban, and T. Szabo (1983) Pathways of the electric organdischarge command and its corollary discharges in mormyrid fish. J.Comp. Neurol. 216:327–338.
Bell, C.C., and C.J. Russell (1978) Effect of electric organ discharge onampullary receptors in a mormyrid. Brain Res. 145:85–96.
Bell, C.C., and T. Szabo (1986) Electroreception in mormyrid fish: Centralanatomy. In T.H. Bullock and W. Heiligenberg (eds): Electroreception.New York: John Wiley and Sons, pp. 375–421.
Bell, C.C., and G. von der Emde (1995) Electric organ corollary dischargepathways in mormyrid fish. II. The medial juxtalobar nucleus. J. Comp.Physiol. A 177:463–479.
Bell, C.C., H. Zakon, and T.E. Finger (1989) Mormyromast electroreceptororgans and their afferent fibers in mormyrid fish. I. Morphology. J.Comp. Neurol. 286:391–407.
Bennett, M.V.L. (1968) Neural Control of Electric Organs. In D. Ingle (ed):The Central Nervous System and Fish Behavior. Chicago: University ofChicago Press, pp. 147–169.
Bennett, M.V.L., G.D. Pappas, E. Aljure, and Y. Nakajima (1967) Physiologyand ultrastructure of electrotonic junctions. II. Spinal and medullaryelectromotor nuclei in mormyrid fish. J. Neurophysiol. 30:180–208.
Bullock, T.H., K. Behrend, and W. Heiligenberg (1975) Comparison of thejamming avoidance responses in gymnotoid and gymnarchid electricfish: A case of convergent evolution of behavior and its sensory basis. J.Comp. Physiol. A 103:97–121.
Carr, C.E. (1986) Time coding in electric fish and barn owls. Brain Behav.Evol. 28:122–133.
Carr, C.E. (1993) Processing of temporal information in the brain. Annu.Rev. Neurosci. 16:223–243.
Carr, C.E., and S. Amagai (1996) Processing of temporal information in thebrain. In M.A. Pastor and J. Artieda (eds): Time, Internal Clocks andMovement. New York: Elsevier, pp. 27–52.
Carr, C.E., W. Heiligenberg, and G.J. Rose (1986a) A time-comparisoncircuit in the electric fish midbrain. I. Behavior and Physiology. J.Neurosci. 6:107–119.
Carr, C.E., and M. Konishi (1990) A circuit for detection of interaural timedifferences in the brain stem of the barn owl. J. Neurosci. 10:3227–3246.
Carr, C.E., L. Maler, and B. Taylor (1986b) A time-comparison circuit in theelectric fish midbrain. II. Functional morphology. J. Neurosci. 6:1372–1383.
Crawford, J.D. (1993) Central auditory neurophysiology of a sound-producing fish: The mesencephalon of Pollimyrus isidori (Mormyridae).J. Comp. Physiol. A 172:139–152.
Crawford, J.D. (1997a) Feature-detecting auditory neurons in the brain of asound-producing fish. J. Comp. Physiol. A 180:439–450.
Crawford, J.D. (1997b) Hearing and acoustic communication in mormyridelectric fishes. Mar. Fresh. Behav. Physiol. 29: (in press).
Deyoe, E.A., and D.C. van Essen (1988) Concurrent processing streams inmonkey visual cortex. TINS 11:219–226.
Elekes, K., M. Ravaille, C.C. Bell, S. Libouban, and T. Szabo (1985) Themormyrid brainstem II: The medullary electromotor relay nucleus: Anultrastructural horseradish peroxidase study. Neuroscience 15:417–429.
Elekes, K., and T. Szabo (1981) Comparative synaptology of the pacemakernucleus in the brain of weakly electric fish (Gymnotidae). In T. Szaboand G. Czeh (eds): Sensory Physiology of Aquatic Lower Vertebrates.Oxford: Pergamon, pp. 107–127.
Elekes, K., and T. Szabo (1985) The mormyrid brainstem III. Ultrastruc-tural and synaptic organization of the medullary ‘‘pacemaker’’ nucleus.Neuroscience 15:431–443.
Finger, T.E., C.C. Bell, and C.J. Russell (1981) Electrosensory pathways tothe valvula cerebelli in mormyrid fish. Exp. Brain Res. 42:23–33.
Friedman, M.A. (1995) The projection within the nucleus exterolateralis inthe knollenorgan pathway of mormyrid fish. Soc. Neurosci. Abstr.21:187.
Grant, K., C.C. Bell, S. Clausse, and M. Ravaille (1986) Morphology andphysiology of the brainstem nuclei controlling the electric organ dis-charge in mormyrid fish. J. Comp. Neurol. 245:514–530.
Hall, C., C. Bell, and R. Zelick (1995) Behavioral evidence of a latency codefor stimulus intensity in mormyrid electric fish. J. Comp. Physiol. A177:29–39.
Haugede-Carre, F. (1983) The mormyrid mesencephalon. II. The medio-dorsal nucleus of the torus semicircularis: Afferent and efferent connec-tions studied with the HRP method. Brain Res. 268:1–14.
356 M.A. FRIEDMAN AND M. KAWASAKI

Heiligenberg, W., and G. Rose (1985) Phase and amplitude computations inthe midbrain of an electric fish: Intracellular studies of neuronsparticipating in the jamming avoidance response of Eigenmannia. J.Neurosci. 5:515–531.
Kawasaki, M. (1993) Independently evolved jamming avoidance responsesemploy identical computational algorithms: A behavioral study of theAfrican electric fish, Gymnarchus niloticus. J. Comp. Physiol. A 173:9–22.
Kawasaki, M. (1994) The African wave-type electric fish, Gymnarchusniloticus, lacks corollary discharge mechanisms for electrosensorygating. J. Comp. Physiol. A 174:133–144.
Kawasaki, M., and Y.-X. Guo (1996) Neuronal circuitry for comparison oftiming in the electrosensory lateral line lobe of an African wave-typeelectric fish, Gymnarchus niloticus. J. Neurosci. 16:380–391.
Konishi, M. (1991) Deciphering the brain’s codes. Neural Computat. 3:1–18.Losier, B.J., and J.A. Matsubara (1990) Comparison of calbindin D 28K and
cytochrome c oxidase in electrosensory nuclei of high- and low-frequency weakly electric fish (Gymnotiformes). Cell Tissue Res. 260:29–39.
Mainen, Z.F., and T.J. Sejnowski (1995) Reliability of spike timing inneocortical neurons. Science 268:1503–1506.
Maler, L., S. Jande, and E.M. Lawson (1984) Localization of vitaminD-dependent calcium binding protein in the electrosensory and electro-motor system of high frequency gymnotid fish. Brain Res. 301:166–170.
McCormick, C.A. (1992) Evolution of central auditory pathways in anamni-otes. In D.B. Webster, R.R. Fay, and A.N. Popper (eds): The Evolution-ary Biology of Hearing. New York: Springer-Verlag, pp. 323–350.
Mugnaini, E., and L. Maler (1987a) Cytology and immunocytochemistry ofthe nucleus exterolateralis anterior of the mormyrid brain: Possible roleof GABAergic synapses in temporal analysis. Anat. Embryol. 176:313–336.
Mugnaini, E., and L. Maler (1987b) Cytology and immunocytochemistry ofthe nucleus of the lateral line lobe in the electric fish Gnathonemuspetersii (Mormyridae): Evidence suggesting that GABAergic synapsesmediate an inhibitory corollary discharge. Synapse 1:32–56.
Otis, T.S., I.M. Raman, and L.O. Trussell (1995) AMPA receptors with highCa21 permeability mediate synaptic transmission in the avian auditorypathway. J. Physiol. 482:309–315.
Parmentier, M., M. Ghysens, F. Rypens, D.E.M. Lawson, J.L. Pasteels, andR. Pochet (1987) Calbindin in vertebrate classes: Immunohistochemicallocalization of Western blot analysis. Gen. Comp. Endocrinol. 65:399–407.
Resibois, A., and J.H. Rogers (1992) Calretinin in rat brain: An immunohis-tochemical study. Neuroscience 46:101–134.
Rogers, J.H. (1987) Calretinin: A gene for a novel calcium-binding proteinexpressed principally in neurons. J. Cell Biol. 105:1343–1353.
Schwaller, B., P. Buchwald, I. Blumcke, M.R. Celio, and W. Hunziker (1993)Characterization of a polyclonal antiserum against the purified humanrecombinant calcium-binding protein calretinin. Cell Calcium 14:639–648.
Seto-Ohshima, A. (1994) Calcium-binding proteins in the central nervoussystem. Acta Histochem. Cytochem. 27:93–106.
Spray, D.C., and J.M. Burt (1990) Structure-activity relations of the cardiacgap junction channel. Am. J. Physiol. 258:C195–C205.
Szabo, T. (1961) Anatomo-physiologie des centres nerveux specifiques dequelques organes electriques. In C. Chagas (ed): Bioelectrogenesis. NewYork: Elsevier, pp. 185–201.
Szabo, T., A.H. Bass, R.G. Northcutt, and M. Ravaille-Veron (1992) Centralprojections of the lateral line nerves in Gymnarchus niloticus identifiedby HRP axonal transport. Soc. Neurosci. Abstr. 18:348.
Szabo, T., A.H. Bass, R.G. Northcutt, and M. Ravaille-Veron (1993) Theelectrosensory ‘‘time coding’’ pathway in Gymnarchus niloticus. Soc.Neurosci. Abstr. 19:376.
Szabo, T., P.S. Enger, and S. Libouban (1979) Electrosensory systems in themormyrid fish, Gnathonemus petersii: Special emphasis on the fastconducting pathway. J. Physiol. (Paris) 75:409–420.
Szabo, T., and A. Fessard (1974) Physiology of Electroreceptors. In A.Fessard (ed): Handbook of Sensory Physiology III/3. New York: Springer-Verlag, pp. 59–124.
Szabo, T., and S. Hagiwara (1967) A latency-change mechanism involved insensory coding of electric fish (mormyrids). Physiol. Behav. 2:331–335.
Szabo, T., and M. Ravaille (1976) Synaptic structure of the lateral line lobenucleus in mormyrid fish. Neurosci. Lett. 2:127–131.
Szabo, T., M. Ravaille, S. Libouban, and P.S. Enger (1983) The mormyridrhombencephalon. I. Light and EM investigations on the structure andconnections of the lateral line lobe nucleus with HRP labeling. BrainRes. 266:1–19.
Takahashi, T.T., C.E. Carr, N. Brecha, and M. Konishi (1987) Calciumbinding protein-like immunoreactivity labels the terminal field ofnucleus laminaris of the barn owl. J. Neurosci. 7:1843–1856.
Turner, R.W., L. Maler, T. Deerinck, S.R. Levinson, and M.H. Ellisman(1994) TTX-sensitive dendritic sodium channels underlie oscillatorydischarge in a vertebrate sensory neuron. J. Neurosci. 14:6453–6471.
Vater, M., and K. Braun (1994) Parvalbumin, calbindin D-28k, andcalretinin immunoreactivity in the ascending auditory pathway ofhorseshoe bats. J. Comp. Neurol. 341:534–558.
von der Emde, G., and H. Bleckmann (1992) Differential responses of twotypes of electroreceptive afferents to signal distortions may permitcapacitance measurement in a weakly electric fish, Gnathonemuspetersii. J. Comp. Physiol. A 171:683–694.
von der Emde, G., and R. Zelick (1995) Behavioral detection of electricsignal waveform distortion in the weakly electric fish, Gnathonemuspetersii. J. Comp. Physiol. A 177:493–501.
Zettel, M.L., C.E. Carr, and W.E. O’Neill (1991) Calbindin-like immunoreac-tivity in the central auditory system of the mustached bat, Pteronotusparnelli. J. Comp. Neurol. 313:1–16.
Zipser, B., and M.V.L. Bennett (1976) Responses of cells of posterior lateralline lobe to activation of electroreceptors in a mormyrid fish. J.Neurophysiol. 39:693–712.
CALRETININ IN MORMYRIFORMS 357