c-jun/c-fos heterodimers regulate cellular genes via a
TRANSCRIPT

c-Junc-Fos heterodimers regulate cellular genesvia a newly identified class of methylated DNAsequence motifsMontse Gustems1 Anne Woellmer1 Ulrich Rothbauer2 Sebastian H Eck3
Thomas Wieland3 Dominik Lutter4 and Wolfgang Hammerschmidt1
1Research Unit Gene Vectors Helmholtz Zentrum Munchen German Research Center for Environmental Healthand German Centre for Infection Research (DZIF) Munich D-81377 Germany 2Biocenter at the Department ofBiology II Ludwig-Maximilians University Munich Martinsried D-82152 Germany 3Institute of Human GeneticsHelmholtz Zentrum Munchen German Research Center for Environmental Health Neuherberg D-85764Germany and 4Institute of Diabetes and Obesity Helmholtz Zentrum Munchen German Research Center forEnvironmental Health Garching D-85748 Germany
Received October 28 2013 Revised November 27 2013 Accepted November 28 2013
ABSTRACT
CpG methylation in mammalian DNA is known tointerfere with gene expression by inhibiting thebinding of transactivators to their cognatesequence motifs or recruiting proteins involved ingene repression An EpsteinndashBarr virus-encodedtranscription factor Zta was the first example of asequence-specific transcription factor that prefer-entially recognizes and selectively binds DNAsequence motifs with methylated CpG residuesreverses epigenetic silencing and activates genetranscription The DNA binding domain of Zta ishomologous to c-Fos a member of the cellularAP-1 (activator protein 1) transcription factorfamily which regulates cell proliferation andsurvival apoptosis transformation and oncogen-esis We have identified a novel AP-1 binding sitetermed meAP-1 which contains a CpG dinucleotideIf methylated meAP-1 sites are preferentially boundby the AP-1 heterodimer c-Junc-Fos in vitro and incellular chromatin in vivo In activated humanprimary B cells c-Junc-Fos locates to thesemethylated elements in promoter regions of tran-scriptionally activated genes Reminiscent of theviral Zta protein c-Junc-Fos is the first identifiedcellular member of the AP-1 family of transacti-vators that can induce expression of genes withmethylated hence repressed promoters reversingepigenetic silencing
INTRODUCTION
AP-1 (activator protein 1) is a family of transcriptionfactors involved in cell proliferation and survival apop-tosis transformation and oncogenesis AP-1 proteinscontain a basic leucine zipper (bZIP) domain and belongto three different subfamilies Jun (c-Jun JunB andJunD) Fos (c-Fos FosB Fra-1 and Fra-2) and ATF(activating transcription factor ATFa ATF2 ATF3ATF4 and B-ATF) To bind to DNA members of thedifferent subfamilies need to form dimers which showdifferent affinities for two classes of DNA sequencesThey are 12-O-Tetradecanoylphorbol-12-Acetate (TPA)response elements (TRE) and cyclic adenosinemonophosphate (cAMP) response elements (CRE)which encompass the consensus sequences 50-TGACGTCA-30 and 50-TGACGTCA-30 respectively Ingeneral JunJun and JunFos dimers show a higheraffinity for TRE sites whereas JunATF and FosATFdimers preferentially bind to CRE sites (1)Different cellular signaling pathways referred to as
mitogen-activated protein kinase (MAPK) cascadesregulate both the expression and post-translational modi-fications of AP-1 proteins The prototypic AP-1 member isthe c-Junc-Fos heterodimer and both proteins areregulated by MAPK pathways (2) Many different extra-cellular stimuli such as growth factors and cytokinestrigger the MAPK cascades consisting of a hierarchy ofsequential kinases that finally activate the three mainMAP kinases called JNK ERK and p38 These proteinscan phosphorylate different transcription factors which inturn activate the expression of c-jun and c-fos genes Onceexpressed the c-Jun and c-Fos proteins form dimers
To whom correspondence should be addressed Tel+49 8931 874 220 Fax+49 8931 874 225 Email montserratgustemshelmholtz-muenchendePresent addressesUlrich Rothbauer University of TubingenNatural and Medical Science Institute at the University of Tuebingen Reutlingen D-72770 GermanySebastian H Eck Centre for Human Genetics and Laboratory Medicine Dr Klein Dr Rost and Colleagues Martinsried D-82152 Germany
Published online 25 December 2013 Nucleic Acids Research 2014 Vol 42 No 5 3059ndash3072doi101093nargkt1323
The Author(s) 2013 Published by Oxford University PressThis is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (httpcreativecommonsorglicensesby-nc30) which permits non-commercial re-use distribution and reproduction in any medium provided the original work is properly cited For commercialre-use please contact journalspermissionsoupcom
and bind to their target DNA sequence motifs but theyalso need to be phosphorylated a modification that can beintroduced by MAPKs as well to activate transcription oftarget genes with AP-1 binding sites in their promoters (1)In eukaryotic cells transcriptional activation depends
on chromatin structure nucleosomal occupancy histonemodifications and DNA methylation which are diverseprocesses that play important roles in transcriptional regu-lation Transcriptionally repressed heterochromatin ischaracterized by the frequent occurrence of cytosine nu-cleotides with a methyl group at the 50 position of thecytosine pyrimidine ring followed by guanine nucleotidestermed CpG DNA methylation affects the transcriptionof genes in two ways First methylation of DNA itselfmay physically impede the binding of transcriptionalproteins to promoters and second methylated DNAmay be bound by proteins known as methyl-CpG-binding domain proteins (MBDs) MBD proteins thenrecruit additional proteins to the locus such as histonedeacetylases and other chromatin remodeling proteinsthat modify histones and promote the formation ofcompact inactive and silenced chromatin leading togene repression (3)CpG methylation is indicative of transcriptional repres-
sion but we and others have recently found that a viraltranscription (Zta) factor preferentially binds to CpG-methylated DNA sequence motifs activating epigeneticallyrepressed viral promoters (45) The Zta protein (alsotermed BZLF1 ZEBRA Z or EB1) is the major immedi-ate early transactivator of a human herpes virus EpsteinndashBarr virus (EBV) Ztarsquos bZIP domain is highly homolo-gous to cellular AP-1 members especially c-Fos (6) In asystematic approach we mapped all Zta responseelements (ZREs) within promoters of viral genes andidentified two classes of ZREs which are characterizedby two distinct binding motifs One class encompassesnearly all ZREs which bind to an AP-1-related DNAmotif The second class encompasses ZREs with a CpGmotif Zta binds to the majority of these ZREs only whenthe cytosine in the CG dinucleotide carries a 50-methylgroup Consequently these sites were termed meZREsZta binds to meZRE motifs and dramatically inducesviral gene expression but in a methylation-dependentfashion (5) This exceptional feature is essential for thisherpes virus to escape from its latent phase of infectionwhich is governed by transcriptional silencing of themajority of viral genes including extensive and widespreadCpG methylation of viral DNA (7)To our knowledge only two reports indicate that
cellular transcription factors also can target methylatedDNA sequence motifs supporting gene transcription inmetazoan cells in vivo Sp1Sp3 binding is not commonlyimpaired by CpG methylation (8) but rather enhanced inthe context of a tissue-specific promoter (9) Whenmethylated cAMP response elements (CRE) or compositeCRE and CEBP sequence elements generate novelbinding sites for CEBPa activating certain promoters(10) which is an unexpected finding (11)Herpes viruses acquire cellular genes through recombin-
ation with host cell DNA Cellular genes adopted andfurther modified by this class of viruses regulate apoptosis
control the cell cycle of infected cells and block hostimmune responses (12) Very likely the viral BZLF1 geneencoding Zta is derived from a cellular member of the AP-1family (6) has evolved to act as the critical immediate-earlygene of EBV to allow exit from latently infected cells (13)and activates resting primary human B cells the target ofEBV infection (14) As described earlier in the text thebZIP domain of Zta is highly conserved when comparedwith bZIP domains of AP-1members andmediates bindingto CpG-methylated DNA sequence motifs Zta might haveinherited this peculiar feature from cellular AP-1 membersthat could target and activate repressed cellular promotersencompassing CpG-methylated DNA as well This hypoth-esis would substantiate a previously unknown type of generegulation and constitute a new example of a cellular tran-scription factor capable of activating gene expression fromepigenetically repressed promoters through direct bindingto methylated DNA
MATERIALS AND METHODS
Cells
HEK293 Raji and K562 cells were maintained in RPMI1640 medium with 10 fetal calf serum (FCS) 1 peni-cillinndashstreptomycin and 1 sodium pyruvate at 37C and5 CO2 Wi-38 cells were maintained in DulbeccorsquosModified Eagle Medium (DMEM) medium with 10FCS 1 penicillinndashstreptomycin and 1 non-essentialamino acids at 37C and 5 CO2 Adenoid B cells weremaintained in RPMI 1640 medium supplemented with10 FCS 100 mgml streptomycin 100Uml penicillin100 nM sodium selenite and 1 mgml cyclosporin A at37C and 5 CO2 For cellular activation B cells werecultured on irradiated CD40L feeder cells in the presenceof 2 ngml IL-4 for 5 days as described (15)
Plasmids
The wild-type maxi-EBV plasmid (p2089) contains thecomplete genome of the EBV prototype B958 strain andhas been described (16) The DNA binding and dimeriza-tion domains of human c-Jun and c-Fos (amino acidresidues 123ndash331 and 1ndash206 respectively) separated bya flexible linker (SG4)5 (17) were cloned downstream ofthe coding region of enhanced green fluorescence protein(eGFP) in pEGFP-C1 (Clontech) to generate the expres-sion plasmid p4271 or downstream of the tandem StrepIIFLAG-tag (18) to yield the plasmid p4548 The plasmidexpressing VP16c-junc-fos (p4316) was generated byreplacing the gfp gene by the VP16 transactivationdomain (TAD) of the VP16 protein from HSV-1 (aminoacid residues 404ndash490) Luciferase plasmids were con-structed by inserting pentamers of single meAP-1 sitesinto a basic luciferase reporter plasmid with a minimalEF1a promoter The plasmid backbone of this reporterplasmid is free of CpGs (19)
DNA transfection
Transfection of DNA into HEK293 cells was performedusing polyethylenimine (Sigma-Aldrich) as previously
3060 Nucleic Acids Research 2014 Vol 42 No 5
described (14) For protein extracts 1 107 cells per 130-mm dish were seeded the day before transfection Eachplate was transfected with 30 mg of plasmid DNA Forreporter assays 5 105 HEK293 cells were seeded into6-well plates the day before transfection Each well wascotransfected with 05mg of reporter plasmid together with1 mg of transactivator and 001mg of DNA of a renilla-expressing plasmid as an internal control for datanormalization
In vitro DNA methylation
CpG methylation in vitro was performed with the de novomethyltransferase MSssI and S-adenosyl methionine asdescribed (520)
In vitro immunoprecipitation assays with GFPc-Junc-Fos
For the in vitro pull-down assays the plasmid p4271encoding gfpc-junc-fos was transiently transfected intoHEK293 cells Nuclear extracts containing the GFPc-Junc-Fos protein were used in in vitro immunopre-cipitation assays with Escherichia coli-derived genomicEBV DNA as described (5)
Recombinant adenovirus generation and cell infection
The adenoviral expression vector was created usingthe Gateway Recombination Cloning Technology(Invitrogen) Shortly the gfpc-junc-fos coding regionwas amplified by polymerase chain reaction (PCR)introduced in the PCR8GWTOPO vector and thentransferred to the pDEST122 vector The latter plasmidwas used to recombine the gfpc-junc-fos cassette into theadenoviral vector pCAGAdDu-GFP (provided by DrVigo Heissmeyer)
Stable transfection and establishment of cell lines
Five micrograms of p4271 DNA was transfected into5 106 Raji cells by electroporation in 250 ml medium at230V and 975 mF using a Biorad electroporation appar-atus in 4-mm cuvettes Immediately after electroporation400 ml of FCS was added to the cells which weretransferred to a flask containing 5ml of cell culturemedium Cells were incubated at 37C and 5 CO2 forone day The cells were plated into 96-well plates in 200 mlof cell culture mediumwell supplemented with puro-mycin (12 mgml) until resistant cells grew out whichwere further cultivated under selective pressure
Native chromatin immunoprecipitations
A total of 5 107 Raji cells expressing the GFPc-Junc-Fos protein were used for chromatin immunopre-cipitation (ChIP) experiments which were performed asdescribed previously (5)
Electromobility shift assays
Electromobility shift assays (EMSAs) were performed asdescribed previously [Bergbauer et al (5)]
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assays (ELISAs) wereperformed with purified protein from HEK293 cells
transiently transfected with StrepFLAGc-Junc-Fos(p4548) Two days post-transfection cells from three130-mm dishes were pooled and lysed in 2ml of RIPA-buffer (50mM Tris 150mM NaCl 1 NP40 05DOC 01 SDS pH 80) Cell lysates were sonicatedand StrepFLAGc-Junc-Fos was affinity purified withStrep-Tactin sepharose (iba-biotagnology) StrepFLAGc-Junc-Fos was eluted in 500 ml of Strep elutionbuffer Purified StrepFLAGc-Junc-Fos protein wascoated on a 96-well ELISA plate at 100 nM in phos-phate-buffered saline at 4C overnight Cy5-labeled oligo-nucleotides were provided by Metabion After washingand blocking with 5 bovine serum albumin 100 ml ofserial dilutions of annealed double-stranded Cy5-labeledoligos (in 20mM Hepes 75mM NaCl 1mM DTT 1glycerin 2mM MgCl2 001mgml bovine serum albuminand a 20molar excess of polydIdC) were then added andincubated for 2 h at room temperature Unbound oligowas removed with repeated washes in phosphate-buffered saline Finally the Cy5 levelswell weremeasured in the phosphoimager (FLA 5100 Fuji) Theobtained data were fit to the Hill equation with singlesite-specific binding using the Prism 5 software to calculatethe dissociation constant
Luciferase reporter assays
Forty-eight hours post-transfection the HEK293 cellswere analyzed with the Dual-Luciferase Reporter AssaySystem (Promega) Luciferase activity was measured in a96-well microplate luminometer (Victor2 Wallac)
Bisulfite sequencing
Bisulfite modification was done using the EpiTect BisulfiteKit (Qiagen) and the regions of interest were amplified bysuitably designed PCR primer pairs Primer sequences areavailable on request
Library construction
Sequence libraries were constructed with paired-end DNAsample preparation kits (Illumina) according to the manu-facturerrsquos recommendations with minor modifications andas described previously (5) In vitro immunoprecipitationswith nonndashCpG-methylated and fully CpG-methylatedEBV DNAs were converted into the librarieslsquo4271unmethylatedrsquo and lsquo4271methylatedrsquo respectivelyDNA obtained with ChIP of Raji cell chromatin was con-verted into the library lsquo4427Rajirsquo Wi-38 DNA used forChIP and ChIP DNA were converted into the librarieslsquo4673Wi38Inputrsquo and lsquo4673Wi38ChIPrsquo respectively
Read mapping
Sequencing reactions were performed on an IlluminaGenome Analyzer IIx or Illumina HiSeq 2000 machinesWe generated 32 million and 31 million 36-bp single-end reads for libraries lsquo4271unmethylatedrsquo andlsquo4271methylatedrsquo respectively 50 million 36-bp single-end reads for lsquo4427Rajirsquo and 50 and 47 million 100-bppair-end reads for the lsquo4673Wi38Inputrsquo andlsquo4673Wi38ChIPrsquo samples respectively Read mapping
Nucleic Acids Research 2014 Vol 42 No 5 3061
to the reference sequence and subsequent assembly wasperformed using the resequencing software Burrows-Wheeler Alignment (BWA) (v071 21) We used a com-bination of the recombinant EBV strain 2089 (16) and thebovine genome (build bosTau4) as reference sequences forthe two libraries lsquo4271unmethylatedrsquo and lsquo4271methylatedrsquo Calf thymus DNA was used as an unspecificcompetitor in the in vitro immunoprecipitation assays withGFPc-Junc-Fos The reference sequence for the in vivosamples lsquo4427Rajirsquo lsquo4673Wi38Inputrsquo and lsquo4673Wi38ChIPrsquo was the human reference sequence hg19
Transcription factor binding site detection
Genomic regions with a read depth above backgroundlevel are considered as regions containing transcriptionfactor binding sites We used two different programsSite Identification from Short Sequence Reads (SISSRs)(22) and QuEST (23) to identify these regionsTranscription factor binding sites are typically shorterthan the sequenced DNA fragments Binding sitesfor the sample libraries lsquo4271unmethylatedrsquo andlsquo4271methylatedrsquo were determined using SISSRsstandard parameters except for E (min number of direc-tional reads) which was set to 3200 for the librarylsquo4271unmethylatedrsquo and to 1600 for lsquo4271methylatedrsquoand F (average length of DNA fragments) which wasset to 200 in both cases Binding sites for the samplelsquo4427Rajirsquo were inferred using the default QuEST param-eters except for kernel density estimator and read countthreshold (th) which were set to 30 and 160 respectivelyIn the case of the sample lsquo4673Wi38ChIPrsquo it wasanalyzed by SISSRs with the results fromlsquo4673Wi38Inputrsquo as background file and setting th at 220 Toidentify consensus motifs within the discovered bindingsites the sequences containing possible binding siteswere extracted and used as input for the motif findingalgorithm Multiple EM for Motif Elicitation (MEME)(24)
ChIPs with an a-c-Jun antibody
ChIP experiments were conducted following standardprotocols Chromatin was cross-linked for 10minat room temperature and incubated with the antibodya-c-Jun (sc-44 Santa Cruz Biotechnology)
Quantitative real time PCR
Quantitative real time PCR (qRT-PCR) analysis was per-formed with a LightCycler 480 II instrument (Roche)with software version 150 SP4 according to the manu-facturerrsquos instructions Amplifications were monitoredwith the LightCycler FastStart reaction mix (SYBRgreen I Roche) The amplification of PCR products wasterminated after 45 cycles The following touchdown PCRprotocol was used (i) initial denaturation at 95 for 10minand (ii) 45 cycles of 95 (2 s) 65ndash62 (06 decrement inannealing temperaturecycle 10 s) 72 (10 s) For allprimer pairs a standard curve from serial dilutions ofthe PCR product with known concentration or the inputDNA was assessed Primer sequences are available onrequest
B cell isolation
Human primary B cells from adenoids were preparedand naıve B cells were sorted as described previously(1425) Anonymized adenoid samples from routineadenoidectomies were provided by the Department ofOtorhinolaryngology Klinikum Grosshadern Ludwig-Maximilians-University of Munich The institutionalreview board Ethikkommission of the KlinikumGrosshadern approved the study and did not requireprior informed patient consent
Bioinformatic identification of meAP-1 sites in primaryhuman B cells
In the reference human genome assembly GRCh37hg19we detectedAP-1 andmeAP-1 binding sites using the searchalgorithm Find Individual Motif Occurrences (FIMO) (26)based on our MEME output motifs Called sites weremapped to promoter sequences of RefSeq genes containedin theUCSCGenome Browser5 kb to+1kb of their tran-scription start sites (TSS) (27) Predicted meAP-1 bindingsite within promoter sequences were further evaluated forCpG methylation in naıve B cells (28) (C04M7ACXX_XFD088DACXX_XF D08E5ACXX_XF) For furtheranalysis we selected only meAP-1 binding sites that showat least 80 methylation in two of three replicates Toidentify genes upregulated in activated B cells we used ex-pression data obtained from unstimulated (GSM907730GSM907738 GSM907746) and stimulated (GSM907731GSM907739 GSM907747) B cell data sets available onGene Expression Omnibus (GSE36975) (29) We selectedtranscripts that were significantly expressed in 5 of 6samples Transcripts showing significant differential ex-pression between unstimulated and stimulated B cellswere identified performing a one way ANOVA test usingMATLAB
RESULTS
c-Junc-Fos binding motifs in genomic EBV DNA
We started our search with a synthetic model whichbuilds on the entire DNA genome of EBV cloned inE coli as a single plasmid (16) This DNA is large(180 kb) GC-rich (60) free of methylated cytosine nu-cleotides (which can be easily methylated in vitro)contains 85 genes and diverse promoter elements andhas been used previously for the analysis of Ztarsquos DNAbinding characteristics (5) We predicted that the DNArsquoscomplexity and size is suitable to identify putative DNAmotifs which are targeted by AP-1 if methylated Thesecond component in our model is a single-chain c-Junc-Fos protein designed to ensure the formation of a singledefined AP-1 heterodimer unaffected by other proteinsthat complex with either c-Jun or c-Fos Bakiri et al(17) showed that similar constructs with the murine se-quences of c-Jun and c-Fos specifically bind andtransactivate cellular promoters that contain the consen-sus AP-1 binding site
We used in vitro pull-down experiments as described (5)with randomly sheared nonndashCpG-methylated or fully
3062 Nucleic Acids Research 2014 Vol 42 No 5
CpG-methylated EBV DNA and a GFPc-Junc-Fosfusion protein purified from HEK293 cellsImmunoprecipitation of the fusion protein followed bydeep sequencing of the enriched DNA fragmentsprovided an extensive coverage of DNA sequences select-ively bound by the AP-1 heterodimer (SupplementaryFigure S1) A comparison of fully CpG-methylatedversus nonndashmethylated-DNA identified several regionspreferentially bound by c-Junc-Fos if CpG-methylated(Supplementary Figure S1 bottom) three of which werelocated at EBV promoters BFLF2 BBLF4 and BALF1(Supplementary Figure S2A) Algorithms suitable toidentify sequences of potential binding sites readilyfound the known palindromic AP-1 consensus bindingsite 50-TGACTCA-30 (Figure 1) in samples ofunmethylated as well as methylated DNA confirmingthe validity of our approach Putative AP-1 bindingmotifs identified in CpG-methylated DNA were groupedaccording to whether they encompassed CpG pairs or notThe reanalysis revealed a second novel motif 50-TGACGTCG-30 which is closely related to the consensus AP-1 butmay contain up to two CpG dinucleotides We termed thismotif lsquomeAP-1rsquo in analogy to meZRE motifs bound byZta (5) Lists of the identified AP-1 binding sites areprovided in Supplementary Tables S1ndashS3
To determine whether the biochemically identifiedmeAP-1 sites in EBV DNA are also bound by c-Junc-Fos in cells in vivo we engineered a derivative of thelatently EBV-infected cell line Raji with a constitutivelyexpressed GFPc-Junc-Fos chimera ChIP followed bydeep sequencing analysis (ChIP-seq) of DNA fragmentsspecifically bound to GFPc-Junc-Fos protein was per-formed mapped to the EBV genome and analyzedbioinformatically in conjunction with methylation data(7) Confirming our results in vitro the analyzed pro-moters in Raji cells were only enriched if the meAP-1sites were highly CpG-methylated (SupplementaryFigure S2)
c-Junc-Fos binds CpG-methylated meAP-1 sites withhigh affinity mediating transcriptional activation
The different meAP-1 motifs identified in the EBV genomemay contain up to two CpG pairs (Figure 1) but the threemore prominent meAP-1 sites (Supplementary Figure S1)located in three EBV promoters encompass the sequence50-TGACTCG-30 with one CpG pair only (Figure 2A)We addressed this uncertainty together with the questionof whether 50-methylcytosine residues present in the topbottom or both strands of the meAP-1 DNA sequenceaffect the affinity of c-Junc-Fos binding to DNA
Unmethylated or CpG-methylated synthetic oligo-nucleotides containing the meAP-1 binding sites in theBBLF4 BFLF2 and BALF1 promoters (Figure 2A) andthree promoters identified later in ChIP-seq experimentswith Raji cell chromatin (Supplementary Figure S3) wereanalyzed in EMSAs As shown in Figure 2B c-Junc-Fosbound almost exclusively to the CpG-methylated meAP-1motif 50-TGACTCG-30 that contains a single CpG pairbut methylation of a CpG pair in a more central positionof the motif decreased the binding of c-Junc-Fos
(Supplementary Figure S3B) Therefore we concentratedour analysis on meAP-1 binding sites with a CpG pair atthe distal position The consensus meAP-1 site consists ofthe sequence motif 50-TGACTCG-30 and only the methylgroup in 50 position of the cytosine in the bottom strand isnecessary for the methylation-specific binding of c-Junc-Fos to DNA (Figure 2C)To quantify DNA binding the apparent dissociation
constants (KD) of c-Junc-Fos were measured withunmethylated or methylated meAP-1 binding sites TheKD values of BBLF4 BALF1 and BFLF2 meAP-1 siteswere lower by a factor of 387 100 and 171 respectivelyif methylated indicating that the presence of50-methylcytosine increases the binding affinity of c-Junc-Fos to DNA (Figure 2D) The consensus AP-1 bindingsite from the human lsquocollagenasersquo promoter (TREcollag)with a published KD for c-Junc-Fos of 108 nM (30) wasused as a positive controlThe transactivation potential of the c-Junc-Fos
heterodimer was investigated in luciferase reporterassays Reporter plasmids containing pentamers of thethree selected meAP-1 sites were transiently transfectedinto HEK293 cells together with an expression plasmidencoding VP16c-Junc-Fos in which the VP16transactivator domain was fused to the single-chain AP-1 heterodimer VP16c-Junc-Fos induced the CpG-methylated reporter plasmids by a factor of 2ndash5 ascompared with their non-methylated controls (Figure 2E)
The human genome contains meAP-1 binding sites
Two different cell lines were used to identify AP-1 andmeAP-1 binding sites in vivo the Burkitt lymphoma B-cell line Raji and the human fibroblast cell line Wi-38As explained earlier in the text a Raji cell line expressingthe GFPc-Junc-Fos chimera was used in ChIP-seq
AP-1 (35 sites) meAP-1 (9 sites)
no CpG CpG
AP-1 (61 sites) (44 sites)
unmethylated methylated
1 2 3 4 5 6 7
2
1
0
bits
-1 1 2 3 4 5 6 7 8
2
1
0
bits
-1 1 2 3 4 5 6 7 8 9
2
1
0
bits
-1 1 2 3 4 5 6 7
2
1
0
bits
Figure 1 Bioinformatic identification of c-Junc-Fos binding motifs inthe EBV genome Deep sequencing data were analyzed with the SISSRsalgorithms for putative stretches of DNA which c-Junc-Fos bindsThe output files of SISSRs were used as training sets for MEMEwhich identifies gapless local and multiple sequence motifs A totalof 61 motifs were identified in the DNA free of CpG methylationand 44 motifs were identified in the fully CpG-methylated EBVDNA The identified motifs in methylated DNA were selected at thelevel of the SISSRs training set data and grouped into motifs with andwithout CpG dinucleotides followed by MEME analysis
Nucleic Acids Research 2014 Vol 42 No 5 3063
A B
C
E
agtcggctagcTGACTCAgatgtcc
gagcgaTGACTCGtgtgcg
ggtctgTGACTCGttaatta
tcagggTGACTCGctaccc
TREcollag
BBLF4
BFLF2
BALF1
α-GFP - - + - - +
prot - + + - + +
unme me
- - + - - +
- + + - + +
- - + - - +
- + + - + +
BFLF2 BBLF4 BALF1
unme me unme me
001 01 1 10 100 10000
200
400
600
800
1000 BBLF4meBBLF4
nM oligo
Cy5
inte
nsi
t y
001 01 1 10 100 10000
200
400
600BALF1meBALF1
nM oligo001 01 1 10 100 10000
200
400
600
800
1000 BFLF2meBFLF2
nM oligo001 01 1 10 100 10000
200
400
600
TREcollag
nM oligo
4382nM 1997nM 2174nM 126nM
113nM 199nM 127nM
unmethylated
methylated nax387 x100 x171
D
prot - + - + - + - + - +
meth -- ++ +- -+
meAP-1 AP-1
meAP-1 AP-1
TGACTCGACTGAGC
TGACTCGACTGAGC
TGACTCAACTGAGT
-+
-- TGACTCGACTGAGC++
TGACTCGACTGAGC
+-
5xBFLF2 5xBBLF4 5xBALF1
Figure 2 The c-Junc-Fos binds preferentially to and activates gene transcription through CpG-methylated meAP-1 motifs (A) Oligonucleotidescontaining meAP-1 binding sites identified in the viral BBLF4 BALF1 and BFLF2 promoters The TREcollag oligo containing a consensus AP-1binding site was used as a control for the KD measurements The meAP-1 sites and AP-1 site are indicated in bold capital letters and the CpG pairswithin the meAP-1 sites are underlined (B) Preferential binding of c-Junc-Fos to methylated meAP-1 sites from the EBV genome EMSAs wereperformed with affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Supershifts with a GFP antibodyconfirmed the identity of the proteinndashDNA complexes EMSAs of typical experiments are shown as examples (C) Only one methyl group is
3064 Nucleic Acids Research 2014 Vol 42 No 5
(continued)
experiments In a second experimental setting serum-starved Wi-38 cells were infected with a GFPc-Junc-Fos-expressing adenovirus Chromatin from Wi-38(two days post infection) and Raji cells was preparedimmunoprecipitated with an a-GFP antibody to enrichDNA fragments specifically bound by GFPc-Junc-Fosprotein and analyzed with next-generation sequencing(Figure 3)
In accordance with our first functional analysis shownin Supplementary Figure S3B and discussed earlier in thetext we adjusted the bioinformatic parameters and refinedthe characteristics of the two AP-1 motifs found previ-ously consensus AP-1 motifs lacking a CpG dinucleotideand putative meAP-1 motifs with a single distal CpG pairin positions six and seven (Figure 3A and B) The twopredominant motifs revealed in both cellular modelsconfirm the known AP-1 consensus (50-TGACTCA-30)and identified an infrequent second meAP-1 motif(50-TGACGTCG-30) (Figure 3A and B) SupplementaryTables S4 through S7 provide the location of the AP-1 andmeAP-1 sites in both cell types
The majority of all consensus AP-1 sites are located inintergenic regions (3132) with no assigned functions Toconcentrate our further analysis on meAP-1 sites in prox-imity of promoter elements meAP-1 sites located within5 kb upstream of annotated TSS were considered only ifno other conventional AP-1 was located within a windowof plusmn1kb Only a small number of cellular genes fulfilledour criteria From ChIP-seq experiments of Raji cells weincluded the promoters of three genes in our study PTK6P2RY5 and SUV39H1 In Wi-38 cells we concentrated ontwo genes CBR3 and LRRTM2 A positive control thecollagenase promoter MMP1 containing two AP-1binding sites (33) was also included
Visualization of the coverage of these promoters afterChIP-seq analysis indicated that they are differentiallybound by c-Junc-Fos in the two cell types (Figure 3C)suggesting that the extent of cytosine methylation mightalso differ Bisulfite sequencing of the five promotersrevealed only one promoter element P2RY5 that wasfully CpG-methylated in both cell lines but CpG pairsin other promoters with meAP-1 sites differed in a celltype-specific manner (Supplementary Figure S4)
50-methylcytosine-dependent binding of c-Junc-Fos tomeAP-1 sites in the human genome inducestranscriptional activation
Only 5 of meAP-1 sites identified in both cell types arelocated in putative promoter regions and therefore
presumably involved in directly regulating gene expres-sion Three candidate meAP-1 motifs with no AP-1 con-sensus binding sites in their close proximity were analyzedfurther the meAP-1 sites in the PTK6 P2RY5 andSUV39H1 promoters (Figure 4) As shown in Figure 4Bc-Junc-Fos bound almost exclusively to the CpG-methylated meAP-1 motif In ELISAs the apparent dis-sociation constants (KD) of PTK6 and P2RY5 meAP-1sites could only be determined with methylated oligo-nucleotides because KD values of the unmethylatedprobes were too high to be estimated (Figure 4C) TheKD for the SUV39H1 meAP-1 site could not becalculated but the results confirm that the bindingaffinity is higher when the meAP-1 site is methylated(Figure 4B and C) These results indicate that thepresence of 50-methylcytosine favors the binding ofc-Junc-Fos to the meAP-1 sequences identified in thehuman genomeThe ability of c-Junc-Fos to transactivate gene tran-
scription through the binding to the methylated meAP-1sites from the human genome was tested in reporterassays Reporter plasmids containing pentamers ofselected meAP-1 sites in the human promoters werecotransfected with an expression plasmid encoding thetransactivator VP16c-Junc-Fos or a negative controlDNA Luciferase activities with CpG-methylatedreporter plasmids were significantly higher as comparedwith unmethylated reporter plasmid DNAs (20ndash27-foldFigure 4D)
Promoters with methylated meAP-1 sites are bound byc-Junc-Fos in vivo
So far our studies were based on the ectopic expression ofartificial c-Junc-Fos dimers to identify potential meAP-1sites in cellular DNA In contrast Raha et al (31) mappedthe location of endogenous c-Jun and c-Fos protein in twosets of ChIP-seq experiments and identified gt40 000binding sites in the established human myeloblastic cellline K562 To correlate their findings and ours we per-formed ChIP assays in this cell line with a c-Jun specificantibody and investigated by quantitative PCR analysisthe enrichment of the five previously identified promoterelements PTK6 P2RY5 SUV39H1 CBR3 and LRRTM2along with negative and positive control regions (34) Asshown in Supplementary Figure S5 the previouslyidentified five promoter elements which contain onlymeAP-1 but no consensus AP-1 sites were enriched atlevels similar to or exceeding those of the positivecontrols in K562 chromatin indicating that endogenous
Figure 2 Continuedresponsible for the binding of c-Junc-Fos to methylated DNA The meAP-1 and AP-1 binding sites contained in the oligonucleotide used in EMSAsare shown the asterisks represent a 50-methylcytosine residue EMSAs using BBLF4 and TREcollag oligonucleotides with the sites depicted in theleft panel and GFPc-Junc-Fos-purified protein are shown One representative experiment out of three is provided (D) Determination of theapparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion proteintransiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values The obtained data werefitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data point indicates themean and standard deviation of three independent experiments (E) Functional identification of single meAP-1 binding sites Pentamers of singlemeAP-1 sites present in the promoters of BBLF4 BALF1 and BFLF2 EBV genes as in (A) were introduced into a basic luciferase reporter plasmidwith a minimal EF1a promoter and free of CpG dinucleotides Unmethylated and fully CpG-methylated reporter constructs were analyzed in thepresence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Each experiment was performed three times and the means andstandard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3065
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

and bind to their target DNA sequence motifs but theyalso need to be phosphorylated a modification that can beintroduced by MAPKs as well to activate transcription oftarget genes with AP-1 binding sites in their promoters (1)In eukaryotic cells transcriptional activation depends
on chromatin structure nucleosomal occupancy histonemodifications and DNA methylation which are diverseprocesses that play important roles in transcriptional regu-lation Transcriptionally repressed heterochromatin ischaracterized by the frequent occurrence of cytosine nu-cleotides with a methyl group at the 50 position of thecytosine pyrimidine ring followed by guanine nucleotidestermed CpG DNA methylation affects the transcriptionof genes in two ways First methylation of DNA itselfmay physically impede the binding of transcriptionalproteins to promoters and second methylated DNAmay be bound by proteins known as methyl-CpG-binding domain proteins (MBDs) MBD proteins thenrecruit additional proteins to the locus such as histonedeacetylases and other chromatin remodeling proteinsthat modify histones and promote the formation ofcompact inactive and silenced chromatin leading togene repression (3)CpG methylation is indicative of transcriptional repres-
sion but we and others have recently found that a viraltranscription (Zta) factor preferentially binds to CpG-methylated DNA sequence motifs activating epigeneticallyrepressed viral promoters (45) The Zta protein (alsotermed BZLF1 ZEBRA Z or EB1) is the major immedi-ate early transactivator of a human herpes virus EpsteinndashBarr virus (EBV) Ztarsquos bZIP domain is highly homolo-gous to cellular AP-1 members especially c-Fos (6) In asystematic approach we mapped all Zta responseelements (ZREs) within promoters of viral genes andidentified two classes of ZREs which are characterizedby two distinct binding motifs One class encompassesnearly all ZREs which bind to an AP-1-related DNAmotif The second class encompasses ZREs with a CpGmotif Zta binds to the majority of these ZREs only whenthe cytosine in the CG dinucleotide carries a 50-methylgroup Consequently these sites were termed meZREsZta binds to meZRE motifs and dramatically inducesviral gene expression but in a methylation-dependentfashion (5) This exceptional feature is essential for thisherpes virus to escape from its latent phase of infectionwhich is governed by transcriptional silencing of themajority of viral genes including extensive and widespreadCpG methylation of viral DNA (7)To our knowledge only two reports indicate that
cellular transcription factors also can target methylatedDNA sequence motifs supporting gene transcription inmetazoan cells in vivo Sp1Sp3 binding is not commonlyimpaired by CpG methylation (8) but rather enhanced inthe context of a tissue-specific promoter (9) Whenmethylated cAMP response elements (CRE) or compositeCRE and CEBP sequence elements generate novelbinding sites for CEBPa activating certain promoters(10) which is an unexpected finding (11)Herpes viruses acquire cellular genes through recombin-
ation with host cell DNA Cellular genes adopted andfurther modified by this class of viruses regulate apoptosis
control the cell cycle of infected cells and block hostimmune responses (12) Very likely the viral BZLF1 geneencoding Zta is derived from a cellular member of the AP-1family (6) has evolved to act as the critical immediate-earlygene of EBV to allow exit from latently infected cells (13)and activates resting primary human B cells the target ofEBV infection (14) As described earlier in the text thebZIP domain of Zta is highly conserved when comparedwith bZIP domains of AP-1members andmediates bindingto CpG-methylated DNA sequence motifs Zta might haveinherited this peculiar feature from cellular AP-1 membersthat could target and activate repressed cellular promotersencompassing CpG-methylated DNA as well This hypoth-esis would substantiate a previously unknown type of generegulation and constitute a new example of a cellular tran-scription factor capable of activating gene expression fromepigenetically repressed promoters through direct bindingto methylated DNA
MATERIALS AND METHODS
Cells
HEK293 Raji and K562 cells were maintained in RPMI1640 medium with 10 fetal calf serum (FCS) 1 peni-cillinndashstreptomycin and 1 sodium pyruvate at 37C and5 CO2 Wi-38 cells were maintained in DulbeccorsquosModified Eagle Medium (DMEM) medium with 10FCS 1 penicillinndashstreptomycin and 1 non-essentialamino acids at 37C and 5 CO2 Adenoid B cells weremaintained in RPMI 1640 medium supplemented with10 FCS 100 mgml streptomycin 100Uml penicillin100 nM sodium selenite and 1 mgml cyclosporin A at37C and 5 CO2 For cellular activation B cells werecultured on irradiated CD40L feeder cells in the presenceof 2 ngml IL-4 for 5 days as described (15)
Plasmids
The wild-type maxi-EBV plasmid (p2089) contains thecomplete genome of the EBV prototype B958 strain andhas been described (16) The DNA binding and dimeriza-tion domains of human c-Jun and c-Fos (amino acidresidues 123ndash331 and 1ndash206 respectively) separated bya flexible linker (SG4)5 (17) were cloned downstream ofthe coding region of enhanced green fluorescence protein(eGFP) in pEGFP-C1 (Clontech) to generate the expres-sion plasmid p4271 or downstream of the tandem StrepIIFLAG-tag (18) to yield the plasmid p4548 The plasmidexpressing VP16c-junc-fos (p4316) was generated byreplacing the gfp gene by the VP16 transactivationdomain (TAD) of the VP16 protein from HSV-1 (aminoacid residues 404ndash490) Luciferase plasmids were con-structed by inserting pentamers of single meAP-1 sitesinto a basic luciferase reporter plasmid with a minimalEF1a promoter The plasmid backbone of this reporterplasmid is free of CpGs (19)
DNA transfection
Transfection of DNA into HEK293 cells was performedusing polyethylenimine (Sigma-Aldrich) as previously
3060 Nucleic Acids Research 2014 Vol 42 No 5
described (14) For protein extracts 1 107 cells per 130-mm dish were seeded the day before transfection Eachplate was transfected with 30 mg of plasmid DNA Forreporter assays 5 105 HEK293 cells were seeded into6-well plates the day before transfection Each well wascotransfected with 05mg of reporter plasmid together with1 mg of transactivator and 001mg of DNA of a renilla-expressing plasmid as an internal control for datanormalization
In vitro DNA methylation
CpG methylation in vitro was performed with the de novomethyltransferase MSssI and S-adenosyl methionine asdescribed (520)
In vitro immunoprecipitation assays with GFPc-Junc-Fos
For the in vitro pull-down assays the plasmid p4271encoding gfpc-junc-fos was transiently transfected intoHEK293 cells Nuclear extracts containing the GFPc-Junc-Fos protein were used in in vitro immunopre-cipitation assays with Escherichia coli-derived genomicEBV DNA as described (5)
Recombinant adenovirus generation and cell infection
The adenoviral expression vector was created usingthe Gateway Recombination Cloning Technology(Invitrogen) Shortly the gfpc-junc-fos coding regionwas amplified by polymerase chain reaction (PCR)introduced in the PCR8GWTOPO vector and thentransferred to the pDEST122 vector The latter plasmidwas used to recombine the gfpc-junc-fos cassette into theadenoviral vector pCAGAdDu-GFP (provided by DrVigo Heissmeyer)
Stable transfection and establishment of cell lines
Five micrograms of p4271 DNA was transfected into5 106 Raji cells by electroporation in 250 ml medium at230V and 975 mF using a Biorad electroporation appar-atus in 4-mm cuvettes Immediately after electroporation400 ml of FCS was added to the cells which weretransferred to a flask containing 5ml of cell culturemedium Cells were incubated at 37C and 5 CO2 forone day The cells were plated into 96-well plates in 200 mlof cell culture mediumwell supplemented with puro-mycin (12 mgml) until resistant cells grew out whichwere further cultivated under selective pressure
Native chromatin immunoprecipitations
A total of 5 107 Raji cells expressing the GFPc-Junc-Fos protein were used for chromatin immunopre-cipitation (ChIP) experiments which were performed asdescribed previously (5)
Electromobility shift assays
Electromobility shift assays (EMSAs) were performed asdescribed previously [Bergbauer et al (5)]
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assays (ELISAs) wereperformed with purified protein from HEK293 cells
transiently transfected with StrepFLAGc-Junc-Fos(p4548) Two days post-transfection cells from three130-mm dishes were pooled and lysed in 2ml of RIPA-buffer (50mM Tris 150mM NaCl 1 NP40 05DOC 01 SDS pH 80) Cell lysates were sonicatedand StrepFLAGc-Junc-Fos was affinity purified withStrep-Tactin sepharose (iba-biotagnology) StrepFLAGc-Junc-Fos was eluted in 500 ml of Strep elutionbuffer Purified StrepFLAGc-Junc-Fos protein wascoated on a 96-well ELISA plate at 100 nM in phos-phate-buffered saline at 4C overnight Cy5-labeled oligo-nucleotides were provided by Metabion After washingand blocking with 5 bovine serum albumin 100 ml ofserial dilutions of annealed double-stranded Cy5-labeledoligos (in 20mM Hepes 75mM NaCl 1mM DTT 1glycerin 2mM MgCl2 001mgml bovine serum albuminand a 20molar excess of polydIdC) were then added andincubated for 2 h at room temperature Unbound oligowas removed with repeated washes in phosphate-buffered saline Finally the Cy5 levelswell weremeasured in the phosphoimager (FLA 5100 Fuji) Theobtained data were fit to the Hill equation with singlesite-specific binding using the Prism 5 software to calculatethe dissociation constant
Luciferase reporter assays
Forty-eight hours post-transfection the HEK293 cellswere analyzed with the Dual-Luciferase Reporter AssaySystem (Promega) Luciferase activity was measured in a96-well microplate luminometer (Victor2 Wallac)
Bisulfite sequencing
Bisulfite modification was done using the EpiTect BisulfiteKit (Qiagen) and the regions of interest were amplified bysuitably designed PCR primer pairs Primer sequences areavailable on request
Library construction
Sequence libraries were constructed with paired-end DNAsample preparation kits (Illumina) according to the manu-facturerrsquos recommendations with minor modifications andas described previously (5) In vitro immunoprecipitationswith nonndashCpG-methylated and fully CpG-methylatedEBV DNAs were converted into the librarieslsquo4271unmethylatedrsquo and lsquo4271methylatedrsquo respectivelyDNA obtained with ChIP of Raji cell chromatin was con-verted into the library lsquo4427Rajirsquo Wi-38 DNA used forChIP and ChIP DNA were converted into the librarieslsquo4673Wi38Inputrsquo and lsquo4673Wi38ChIPrsquo respectively
Read mapping
Sequencing reactions were performed on an IlluminaGenome Analyzer IIx or Illumina HiSeq 2000 machinesWe generated 32 million and 31 million 36-bp single-end reads for libraries lsquo4271unmethylatedrsquo andlsquo4271methylatedrsquo respectively 50 million 36-bp single-end reads for lsquo4427Rajirsquo and 50 and 47 million 100-bppair-end reads for the lsquo4673Wi38Inputrsquo andlsquo4673Wi38ChIPrsquo samples respectively Read mapping
Nucleic Acids Research 2014 Vol 42 No 5 3061
to the reference sequence and subsequent assembly wasperformed using the resequencing software Burrows-Wheeler Alignment (BWA) (v071 21) We used a com-bination of the recombinant EBV strain 2089 (16) and thebovine genome (build bosTau4) as reference sequences forthe two libraries lsquo4271unmethylatedrsquo and lsquo4271methylatedrsquo Calf thymus DNA was used as an unspecificcompetitor in the in vitro immunoprecipitation assays withGFPc-Junc-Fos The reference sequence for the in vivosamples lsquo4427Rajirsquo lsquo4673Wi38Inputrsquo and lsquo4673Wi38ChIPrsquo was the human reference sequence hg19
Transcription factor binding site detection
Genomic regions with a read depth above backgroundlevel are considered as regions containing transcriptionfactor binding sites We used two different programsSite Identification from Short Sequence Reads (SISSRs)(22) and QuEST (23) to identify these regionsTranscription factor binding sites are typically shorterthan the sequenced DNA fragments Binding sitesfor the sample libraries lsquo4271unmethylatedrsquo andlsquo4271methylatedrsquo were determined using SISSRsstandard parameters except for E (min number of direc-tional reads) which was set to 3200 for the librarylsquo4271unmethylatedrsquo and to 1600 for lsquo4271methylatedrsquoand F (average length of DNA fragments) which wasset to 200 in both cases Binding sites for the samplelsquo4427Rajirsquo were inferred using the default QuEST param-eters except for kernel density estimator and read countthreshold (th) which were set to 30 and 160 respectivelyIn the case of the sample lsquo4673Wi38ChIPrsquo it wasanalyzed by SISSRs with the results fromlsquo4673Wi38Inputrsquo as background file and setting th at 220 Toidentify consensus motifs within the discovered bindingsites the sequences containing possible binding siteswere extracted and used as input for the motif findingalgorithm Multiple EM for Motif Elicitation (MEME)(24)
ChIPs with an a-c-Jun antibody
ChIP experiments were conducted following standardprotocols Chromatin was cross-linked for 10minat room temperature and incubated with the antibodya-c-Jun (sc-44 Santa Cruz Biotechnology)
Quantitative real time PCR
Quantitative real time PCR (qRT-PCR) analysis was per-formed with a LightCycler 480 II instrument (Roche)with software version 150 SP4 according to the manu-facturerrsquos instructions Amplifications were monitoredwith the LightCycler FastStart reaction mix (SYBRgreen I Roche) The amplification of PCR products wasterminated after 45 cycles The following touchdown PCRprotocol was used (i) initial denaturation at 95 for 10minand (ii) 45 cycles of 95 (2 s) 65ndash62 (06 decrement inannealing temperaturecycle 10 s) 72 (10 s) For allprimer pairs a standard curve from serial dilutions ofthe PCR product with known concentration or the inputDNA was assessed Primer sequences are available onrequest
B cell isolation
Human primary B cells from adenoids were preparedand naıve B cells were sorted as described previously(1425) Anonymized adenoid samples from routineadenoidectomies were provided by the Department ofOtorhinolaryngology Klinikum Grosshadern Ludwig-Maximilians-University of Munich The institutionalreview board Ethikkommission of the KlinikumGrosshadern approved the study and did not requireprior informed patient consent
Bioinformatic identification of meAP-1 sites in primaryhuman B cells
In the reference human genome assembly GRCh37hg19we detectedAP-1 andmeAP-1 binding sites using the searchalgorithm Find Individual Motif Occurrences (FIMO) (26)based on our MEME output motifs Called sites weremapped to promoter sequences of RefSeq genes containedin theUCSCGenome Browser5 kb to+1kb of their tran-scription start sites (TSS) (27) Predicted meAP-1 bindingsite within promoter sequences were further evaluated forCpG methylation in naıve B cells (28) (C04M7ACXX_XFD088DACXX_XF D08E5ACXX_XF) For furtheranalysis we selected only meAP-1 binding sites that showat least 80 methylation in two of three replicates Toidentify genes upregulated in activated B cells we used ex-pression data obtained from unstimulated (GSM907730GSM907738 GSM907746) and stimulated (GSM907731GSM907739 GSM907747) B cell data sets available onGene Expression Omnibus (GSE36975) (29) We selectedtranscripts that were significantly expressed in 5 of 6samples Transcripts showing significant differential ex-pression between unstimulated and stimulated B cellswere identified performing a one way ANOVA test usingMATLAB
RESULTS
c-Junc-Fos binding motifs in genomic EBV DNA
We started our search with a synthetic model whichbuilds on the entire DNA genome of EBV cloned inE coli as a single plasmid (16) This DNA is large(180 kb) GC-rich (60) free of methylated cytosine nu-cleotides (which can be easily methylated in vitro)contains 85 genes and diverse promoter elements andhas been used previously for the analysis of Ztarsquos DNAbinding characteristics (5) We predicted that the DNArsquoscomplexity and size is suitable to identify putative DNAmotifs which are targeted by AP-1 if methylated Thesecond component in our model is a single-chain c-Junc-Fos protein designed to ensure the formation of a singledefined AP-1 heterodimer unaffected by other proteinsthat complex with either c-Jun or c-Fos Bakiri et al(17) showed that similar constructs with the murine se-quences of c-Jun and c-Fos specifically bind andtransactivate cellular promoters that contain the consen-sus AP-1 binding site
We used in vitro pull-down experiments as described (5)with randomly sheared nonndashCpG-methylated or fully
3062 Nucleic Acids Research 2014 Vol 42 No 5
CpG-methylated EBV DNA and a GFPc-Junc-Fosfusion protein purified from HEK293 cellsImmunoprecipitation of the fusion protein followed bydeep sequencing of the enriched DNA fragmentsprovided an extensive coverage of DNA sequences select-ively bound by the AP-1 heterodimer (SupplementaryFigure S1) A comparison of fully CpG-methylatedversus nonndashmethylated-DNA identified several regionspreferentially bound by c-Junc-Fos if CpG-methylated(Supplementary Figure S1 bottom) three of which werelocated at EBV promoters BFLF2 BBLF4 and BALF1(Supplementary Figure S2A) Algorithms suitable toidentify sequences of potential binding sites readilyfound the known palindromic AP-1 consensus bindingsite 50-TGACTCA-30 (Figure 1) in samples ofunmethylated as well as methylated DNA confirmingthe validity of our approach Putative AP-1 bindingmotifs identified in CpG-methylated DNA were groupedaccording to whether they encompassed CpG pairs or notThe reanalysis revealed a second novel motif 50-TGACGTCG-30 which is closely related to the consensus AP-1 butmay contain up to two CpG dinucleotides We termed thismotif lsquomeAP-1rsquo in analogy to meZRE motifs bound byZta (5) Lists of the identified AP-1 binding sites areprovided in Supplementary Tables S1ndashS3
To determine whether the biochemically identifiedmeAP-1 sites in EBV DNA are also bound by c-Junc-Fos in cells in vivo we engineered a derivative of thelatently EBV-infected cell line Raji with a constitutivelyexpressed GFPc-Junc-Fos chimera ChIP followed bydeep sequencing analysis (ChIP-seq) of DNA fragmentsspecifically bound to GFPc-Junc-Fos protein was per-formed mapped to the EBV genome and analyzedbioinformatically in conjunction with methylation data(7) Confirming our results in vitro the analyzed pro-moters in Raji cells were only enriched if the meAP-1sites were highly CpG-methylated (SupplementaryFigure S2)
c-Junc-Fos binds CpG-methylated meAP-1 sites withhigh affinity mediating transcriptional activation
The different meAP-1 motifs identified in the EBV genomemay contain up to two CpG pairs (Figure 1) but the threemore prominent meAP-1 sites (Supplementary Figure S1)located in three EBV promoters encompass the sequence50-TGACTCG-30 with one CpG pair only (Figure 2A)We addressed this uncertainty together with the questionof whether 50-methylcytosine residues present in the topbottom or both strands of the meAP-1 DNA sequenceaffect the affinity of c-Junc-Fos binding to DNA
Unmethylated or CpG-methylated synthetic oligo-nucleotides containing the meAP-1 binding sites in theBBLF4 BFLF2 and BALF1 promoters (Figure 2A) andthree promoters identified later in ChIP-seq experimentswith Raji cell chromatin (Supplementary Figure S3) wereanalyzed in EMSAs As shown in Figure 2B c-Junc-Fosbound almost exclusively to the CpG-methylated meAP-1motif 50-TGACTCG-30 that contains a single CpG pairbut methylation of a CpG pair in a more central positionof the motif decreased the binding of c-Junc-Fos
(Supplementary Figure S3B) Therefore we concentratedour analysis on meAP-1 binding sites with a CpG pair atthe distal position The consensus meAP-1 site consists ofthe sequence motif 50-TGACTCG-30 and only the methylgroup in 50 position of the cytosine in the bottom strand isnecessary for the methylation-specific binding of c-Junc-Fos to DNA (Figure 2C)To quantify DNA binding the apparent dissociation
constants (KD) of c-Junc-Fos were measured withunmethylated or methylated meAP-1 binding sites TheKD values of BBLF4 BALF1 and BFLF2 meAP-1 siteswere lower by a factor of 387 100 and 171 respectivelyif methylated indicating that the presence of50-methylcytosine increases the binding affinity of c-Junc-Fos to DNA (Figure 2D) The consensus AP-1 bindingsite from the human lsquocollagenasersquo promoter (TREcollag)with a published KD for c-Junc-Fos of 108 nM (30) wasused as a positive controlThe transactivation potential of the c-Junc-Fos
heterodimer was investigated in luciferase reporterassays Reporter plasmids containing pentamers of thethree selected meAP-1 sites were transiently transfectedinto HEK293 cells together with an expression plasmidencoding VP16c-Junc-Fos in which the VP16transactivator domain was fused to the single-chain AP-1 heterodimer VP16c-Junc-Fos induced the CpG-methylated reporter plasmids by a factor of 2ndash5 ascompared with their non-methylated controls (Figure 2E)
The human genome contains meAP-1 binding sites
Two different cell lines were used to identify AP-1 andmeAP-1 binding sites in vivo the Burkitt lymphoma B-cell line Raji and the human fibroblast cell line Wi-38As explained earlier in the text a Raji cell line expressingthe GFPc-Junc-Fos chimera was used in ChIP-seq
AP-1 (35 sites) meAP-1 (9 sites)
no CpG CpG
AP-1 (61 sites) (44 sites)
unmethylated methylated
1 2 3 4 5 6 7
2
1
0
bits
-1 1 2 3 4 5 6 7 8
2
1
0
bits
-1 1 2 3 4 5 6 7 8 9
2
1
0
bits
-1 1 2 3 4 5 6 7
2
1
0
bits
Figure 1 Bioinformatic identification of c-Junc-Fos binding motifs inthe EBV genome Deep sequencing data were analyzed with the SISSRsalgorithms for putative stretches of DNA which c-Junc-Fos bindsThe output files of SISSRs were used as training sets for MEMEwhich identifies gapless local and multiple sequence motifs A totalof 61 motifs were identified in the DNA free of CpG methylationand 44 motifs were identified in the fully CpG-methylated EBVDNA The identified motifs in methylated DNA were selected at thelevel of the SISSRs training set data and grouped into motifs with andwithout CpG dinucleotides followed by MEME analysis
Nucleic Acids Research 2014 Vol 42 No 5 3063
A B
C
E
agtcggctagcTGACTCAgatgtcc
gagcgaTGACTCGtgtgcg
ggtctgTGACTCGttaatta
tcagggTGACTCGctaccc
TREcollag
BBLF4
BFLF2
BALF1
α-GFP - - + - - +
prot - + + - + +
unme me
- - + - - +
- + + - + +
- - + - - +
- + + - + +
BFLF2 BBLF4 BALF1
unme me unme me
001 01 1 10 100 10000
200
400
600
800
1000 BBLF4meBBLF4
nM oligo
Cy5
inte
nsi
t y
001 01 1 10 100 10000
200
400
600BALF1meBALF1
nM oligo001 01 1 10 100 10000
200
400
600
800
1000 BFLF2meBFLF2
nM oligo001 01 1 10 100 10000
200
400
600
TREcollag
nM oligo
4382nM 1997nM 2174nM 126nM
113nM 199nM 127nM
unmethylated
methylated nax387 x100 x171
D
prot - + - + - + - + - +
meth -- ++ +- -+
meAP-1 AP-1
meAP-1 AP-1
TGACTCGACTGAGC
TGACTCGACTGAGC
TGACTCAACTGAGT
-+
-- TGACTCGACTGAGC++
TGACTCGACTGAGC
+-
5xBFLF2 5xBBLF4 5xBALF1
Figure 2 The c-Junc-Fos binds preferentially to and activates gene transcription through CpG-methylated meAP-1 motifs (A) Oligonucleotidescontaining meAP-1 binding sites identified in the viral BBLF4 BALF1 and BFLF2 promoters The TREcollag oligo containing a consensus AP-1binding site was used as a control for the KD measurements The meAP-1 sites and AP-1 site are indicated in bold capital letters and the CpG pairswithin the meAP-1 sites are underlined (B) Preferential binding of c-Junc-Fos to methylated meAP-1 sites from the EBV genome EMSAs wereperformed with affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Supershifts with a GFP antibodyconfirmed the identity of the proteinndashDNA complexes EMSAs of typical experiments are shown as examples (C) Only one methyl group is
3064 Nucleic Acids Research 2014 Vol 42 No 5
(continued)
experiments In a second experimental setting serum-starved Wi-38 cells were infected with a GFPc-Junc-Fos-expressing adenovirus Chromatin from Wi-38(two days post infection) and Raji cells was preparedimmunoprecipitated with an a-GFP antibody to enrichDNA fragments specifically bound by GFPc-Junc-Fosprotein and analyzed with next-generation sequencing(Figure 3)
In accordance with our first functional analysis shownin Supplementary Figure S3B and discussed earlier in thetext we adjusted the bioinformatic parameters and refinedthe characteristics of the two AP-1 motifs found previ-ously consensus AP-1 motifs lacking a CpG dinucleotideand putative meAP-1 motifs with a single distal CpG pairin positions six and seven (Figure 3A and B) The twopredominant motifs revealed in both cellular modelsconfirm the known AP-1 consensus (50-TGACTCA-30)and identified an infrequent second meAP-1 motif(50-TGACGTCG-30) (Figure 3A and B) SupplementaryTables S4 through S7 provide the location of the AP-1 andmeAP-1 sites in both cell types
The majority of all consensus AP-1 sites are located inintergenic regions (3132) with no assigned functions Toconcentrate our further analysis on meAP-1 sites in prox-imity of promoter elements meAP-1 sites located within5 kb upstream of annotated TSS were considered only ifno other conventional AP-1 was located within a windowof plusmn1kb Only a small number of cellular genes fulfilledour criteria From ChIP-seq experiments of Raji cells weincluded the promoters of three genes in our study PTK6P2RY5 and SUV39H1 In Wi-38 cells we concentrated ontwo genes CBR3 and LRRTM2 A positive control thecollagenase promoter MMP1 containing two AP-1binding sites (33) was also included
Visualization of the coverage of these promoters afterChIP-seq analysis indicated that they are differentiallybound by c-Junc-Fos in the two cell types (Figure 3C)suggesting that the extent of cytosine methylation mightalso differ Bisulfite sequencing of the five promotersrevealed only one promoter element P2RY5 that wasfully CpG-methylated in both cell lines but CpG pairsin other promoters with meAP-1 sites differed in a celltype-specific manner (Supplementary Figure S4)
50-methylcytosine-dependent binding of c-Junc-Fos tomeAP-1 sites in the human genome inducestranscriptional activation
Only 5 of meAP-1 sites identified in both cell types arelocated in putative promoter regions and therefore
presumably involved in directly regulating gene expres-sion Three candidate meAP-1 motifs with no AP-1 con-sensus binding sites in their close proximity were analyzedfurther the meAP-1 sites in the PTK6 P2RY5 andSUV39H1 promoters (Figure 4) As shown in Figure 4Bc-Junc-Fos bound almost exclusively to the CpG-methylated meAP-1 motif In ELISAs the apparent dis-sociation constants (KD) of PTK6 and P2RY5 meAP-1sites could only be determined with methylated oligo-nucleotides because KD values of the unmethylatedprobes were too high to be estimated (Figure 4C) TheKD for the SUV39H1 meAP-1 site could not becalculated but the results confirm that the bindingaffinity is higher when the meAP-1 site is methylated(Figure 4B and C) These results indicate that thepresence of 50-methylcytosine favors the binding ofc-Junc-Fos to the meAP-1 sequences identified in thehuman genomeThe ability of c-Junc-Fos to transactivate gene tran-
scription through the binding to the methylated meAP-1sites from the human genome was tested in reporterassays Reporter plasmids containing pentamers ofselected meAP-1 sites in the human promoters werecotransfected with an expression plasmid encoding thetransactivator VP16c-Junc-Fos or a negative controlDNA Luciferase activities with CpG-methylatedreporter plasmids were significantly higher as comparedwith unmethylated reporter plasmid DNAs (20ndash27-foldFigure 4D)
Promoters with methylated meAP-1 sites are bound byc-Junc-Fos in vivo
So far our studies were based on the ectopic expression ofartificial c-Junc-Fos dimers to identify potential meAP-1sites in cellular DNA In contrast Raha et al (31) mappedthe location of endogenous c-Jun and c-Fos protein in twosets of ChIP-seq experiments and identified gt40 000binding sites in the established human myeloblastic cellline K562 To correlate their findings and ours we per-formed ChIP assays in this cell line with a c-Jun specificantibody and investigated by quantitative PCR analysisthe enrichment of the five previously identified promoterelements PTK6 P2RY5 SUV39H1 CBR3 and LRRTM2along with negative and positive control regions (34) Asshown in Supplementary Figure S5 the previouslyidentified five promoter elements which contain onlymeAP-1 but no consensus AP-1 sites were enriched atlevels similar to or exceeding those of the positivecontrols in K562 chromatin indicating that endogenous
Figure 2 Continuedresponsible for the binding of c-Junc-Fos to methylated DNA The meAP-1 and AP-1 binding sites contained in the oligonucleotide used in EMSAsare shown the asterisks represent a 50-methylcytosine residue EMSAs using BBLF4 and TREcollag oligonucleotides with the sites depicted in theleft panel and GFPc-Junc-Fos-purified protein are shown One representative experiment out of three is provided (D) Determination of theapparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion proteintransiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values The obtained data werefitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data point indicates themean and standard deviation of three independent experiments (E) Functional identification of single meAP-1 binding sites Pentamers of singlemeAP-1 sites present in the promoters of BBLF4 BALF1 and BFLF2 EBV genes as in (A) were introduced into a basic luciferase reporter plasmidwith a minimal EF1a promoter and free of CpG dinucleotides Unmethylated and fully CpG-methylated reporter constructs were analyzed in thepresence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Each experiment was performed three times and the means andstandard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3065
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

described (14) For protein extracts 1 107 cells per 130-mm dish were seeded the day before transfection Eachplate was transfected with 30 mg of plasmid DNA Forreporter assays 5 105 HEK293 cells were seeded into6-well plates the day before transfection Each well wascotransfected with 05mg of reporter plasmid together with1 mg of transactivator and 001mg of DNA of a renilla-expressing plasmid as an internal control for datanormalization
In vitro DNA methylation
CpG methylation in vitro was performed with the de novomethyltransferase MSssI and S-adenosyl methionine asdescribed (520)
In vitro immunoprecipitation assays with GFPc-Junc-Fos
For the in vitro pull-down assays the plasmid p4271encoding gfpc-junc-fos was transiently transfected intoHEK293 cells Nuclear extracts containing the GFPc-Junc-Fos protein were used in in vitro immunopre-cipitation assays with Escherichia coli-derived genomicEBV DNA as described (5)
Recombinant adenovirus generation and cell infection
The adenoviral expression vector was created usingthe Gateway Recombination Cloning Technology(Invitrogen) Shortly the gfpc-junc-fos coding regionwas amplified by polymerase chain reaction (PCR)introduced in the PCR8GWTOPO vector and thentransferred to the pDEST122 vector The latter plasmidwas used to recombine the gfpc-junc-fos cassette into theadenoviral vector pCAGAdDu-GFP (provided by DrVigo Heissmeyer)
Stable transfection and establishment of cell lines
Five micrograms of p4271 DNA was transfected into5 106 Raji cells by electroporation in 250 ml medium at230V and 975 mF using a Biorad electroporation appar-atus in 4-mm cuvettes Immediately after electroporation400 ml of FCS was added to the cells which weretransferred to a flask containing 5ml of cell culturemedium Cells were incubated at 37C and 5 CO2 forone day The cells were plated into 96-well plates in 200 mlof cell culture mediumwell supplemented with puro-mycin (12 mgml) until resistant cells grew out whichwere further cultivated under selective pressure
Native chromatin immunoprecipitations
A total of 5 107 Raji cells expressing the GFPc-Junc-Fos protein were used for chromatin immunopre-cipitation (ChIP) experiments which were performed asdescribed previously (5)
Electromobility shift assays
Electromobility shift assays (EMSAs) were performed asdescribed previously [Bergbauer et al (5)]
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assays (ELISAs) wereperformed with purified protein from HEK293 cells
transiently transfected with StrepFLAGc-Junc-Fos(p4548) Two days post-transfection cells from three130-mm dishes were pooled and lysed in 2ml of RIPA-buffer (50mM Tris 150mM NaCl 1 NP40 05DOC 01 SDS pH 80) Cell lysates were sonicatedand StrepFLAGc-Junc-Fos was affinity purified withStrep-Tactin sepharose (iba-biotagnology) StrepFLAGc-Junc-Fos was eluted in 500 ml of Strep elutionbuffer Purified StrepFLAGc-Junc-Fos protein wascoated on a 96-well ELISA plate at 100 nM in phos-phate-buffered saline at 4C overnight Cy5-labeled oligo-nucleotides were provided by Metabion After washingand blocking with 5 bovine serum albumin 100 ml ofserial dilutions of annealed double-stranded Cy5-labeledoligos (in 20mM Hepes 75mM NaCl 1mM DTT 1glycerin 2mM MgCl2 001mgml bovine serum albuminand a 20molar excess of polydIdC) were then added andincubated for 2 h at room temperature Unbound oligowas removed with repeated washes in phosphate-buffered saline Finally the Cy5 levelswell weremeasured in the phosphoimager (FLA 5100 Fuji) Theobtained data were fit to the Hill equation with singlesite-specific binding using the Prism 5 software to calculatethe dissociation constant
Luciferase reporter assays
Forty-eight hours post-transfection the HEK293 cellswere analyzed with the Dual-Luciferase Reporter AssaySystem (Promega) Luciferase activity was measured in a96-well microplate luminometer (Victor2 Wallac)
Bisulfite sequencing
Bisulfite modification was done using the EpiTect BisulfiteKit (Qiagen) and the regions of interest were amplified bysuitably designed PCR primer pairs Primer sequences areavailable on request
Library construction
Sequence libraries were constructed with paired-end DNAsample preparation kits (Illumina) according to the manu-facturerrsquos recommendations with minor modifications andas described previously (5) In vitro immunoprecipitationswith nonndashCpG-methylated and fully CpG-methylatedEBV DNAs were converted into the librarieslsquo4271unmethylatedrsquo and lsquo4271methylatedrsquo respectivelyDNA obtained with ChIP of Raji cell chromatin was con-verted into the library lsquo4427Rajirsquo Wi-38 DNA used forChIP and ChIP DNA were converted into the librarieslsquo4673Wi38Inputrsquo and lsquo4673Wi38ChIPrsquo respectively
Read mapping
Sequencing reactions were performed on an IlluminaGenome Analyzer IIx or Illumina HiSeq 2000 machinesWe generated 32 million and 31 million 36-bp single-end reads for libraries lsquo4271unmethylatedrsquo andlsquo4271methylatedrsquo respectively 50 million 36-bp single-end reads for lsquo4427Rajirsquo and 50 and 47 million 100-bppair-end reads for the lsquo4673Wi38Inputrsquo andlsquo4673Wi38ChIPrsquo samples respectively Read mapping
Nucleic Acids Research 2014 Vol 42 No 5 3061
to the reference sequence and subsequent assembly wasperformed using the resequencing software Burrows-Wheeler Alignment (BWA) (v071 21) We used a com-bination of the recombinant EBV strain 2089 (16) and thebovine genome (build bosTau4) as reference sequences forthe two libraries lsquo4271unmethylatedrsquo and lsquo4271methylatedrsquo Calf thymus DNA was used as an unspecificcompetitor in the in vitro immunoprecipitation assays withGFPc-Junc-Fos The reference sequence for the in vivosamples lsquo4427Rajirsquo lsquo4673Wi38Inputrsquo and lsquo4673Wi38ChIPrsquo was the human reference sequence hg19
Transcription factor binding site detection
Genomic regions with a read depth above backgroundlevel are considered as regions containing transcriptionfactor binding sites We used two different programsSite Identification from Short Sequence Reads (SISSRs)(22) and QuEST (23) to identify these regionsTranscription factor binding sites are typically shorterthan the sequenced DNA fragments Binding sitesfor the sample libraries lsquo4271unmethylatedrsquo andlsquo4271methylatedrsquo were determined using SISSRsstandard parameters except for E (min number of direc-tional reads) which was set to 3200 for the librarylsquo4271unmethylatedrsquo and to 1600 for lsquo4271methylatedrsquoand F (average length of DNA fragments) which wasset to 200 in both cases Binding sites for the samplelsquo4427Rajirsquo were inferred using the default QuEST param-eters except for kernel density estimator and read countthreshold (th) which were set to 30 and 160 respectivelyIn the case of the sample lsquo4673Wi38ChIPrsquo it wasanalyzed by SISSRs with the results fromlsquo4673Wi38Inputrsquo as background file and setting th at 220 Toidentify consensus motifs within the discovered bindingsites the sequences containing possible binding siteswere extracted and used as input for the motif findingalgorithm Multiple EM for Motif Elicitation (MEME)(24)
ChIPs with an a-c-Jun antibody
ChIP experiments were conducted following standardprotocols Chromatin was cross-linked for 10minat room temperature and incubated with the antibodya-c-Jun (sc-44 Santa Cruz Biotechnology)
Quantitative real time PCR
Quantitative real time PCR (qRT-PCR) analysis was per-formed with a LightCycler 480 II instrument (Roche)with software version 150 SP4 according to the manu-facturerrsquos instructions Amplifications were monitoredwith the LightCycler FastStart reaction mix (SYBRgreen I Roche) The amplification of PCR products wasterminated after 45 cycles The following touchdown PCRprotocol was used (i) initial denaturation at 95 for 10minand (ii) 45 cycles of 95 (2 s) 65ndash62 (06 decrement inannealing temperaturecycle 10 s) 72 (10 s) For allprimer pairs a standard curve from serial dilutions ofthe PCR product with known concentration or the inputDNA was assessed Primer sequences are available onrequest
B cell isolation
Human primary B cells from adenoids were preparedand naıve B cells were sorted as described previously(1425) Anonymized adenoid samples from routineadenoidectomies were provided by the Department ofOtorhinolaryngology Klinikum Grosshadern Ludwig-Maximilians-University of Munich The institutionalreview board Ethikkommission of the KlinikumGrosshadern approved the study and did not requireprior informed patient consent
Bioinformatic identification of meAP-1 sites in primaryhuman B cells
In the reference human genome assembly GRCh37hg19we detectedAP-1 andmeAP-1 binding sites using the searchalgorithm Find Individual Motif Occurrences (FIMO) (26)based on our MEME output motifs Called sites weremapped to promoter sequences of RefSeq genes containedin theUCSCGenome Browser5 kb to+1kb of their tran-scription start sites (TSS) (27) Predicted meAP-1 bindingsite within promoter sequences were further evaluated forCpG methylation in naıve B cells (28) (C04M7ACXX_XFD088DACXX_XF D08E5ACXX_XF) For furtheranalysis we selected only meAP-1 binding sites that showat least 80 methylation in two of three replicates Toidentify genes upregulated in activated B cells we used ex-pression data obtained from unstimulated (GSM907730GSM907738 GSM907746) and stimulated (GSM907731GSM907739 GSM907747) B cell data sets available onGene Expression Omnibus (GSE36975) (29) We selectedtranscripts that were significantly expressed in 5 of 6samples Transcripts showing significant differential ex-pression between unstimulated and stimulated B cellswere identified performing a one way ANOVA test usingMATLAB
RESULTS
c-Junc-Fos binding motifs in genomic EBV DNA
We started our search with a synthetic model whichbuilds on the entire DNA genome of EBV cloned inE coli as a single plasmid (16) This DNA is large(180 kb) GC-rich (60) free of methylated cytosine nu-cleotides (which can be easily methylated in vitro)contains 85 genes and diverse promoter elements andhas been used previously for the analysis of Ztarsquos DNAbinding characteristics (5) We predicted that the DNArsquoscomplexity and size is suitable to identify putative DNAmotifs which are targeted by AP-1 if methylated Thesecond component in our model is a single-chain c-Junc-Fos protein designed to ensure the formation of a singledefined AP-1 heterodimer unaffected by other proteinsthat complex with either c-Jun or c-Fos Bakiri et al(17) showed that similar constructs with the murine se-quences of c-Jun and c-Fos specifically bind andtransactivate cellular promoters that contain the consen-sus AP-1 binding site
We used in vitro pull-down experiments as described (5)with randomly sheared nonndashCpG-methylated or fully
3062 Nucleic Acids Research 2014 Vol 42 No 5
CpG-methylated EBV DNA and a GFPc-Junc-Fosfusion protein purified from HEK293 cellsImmunoprecipitation of the fusion protein followed bydeep sequencing of the enriched DNA fragmentsprovided an extensive coverage of DNA sequences select-ively bound by the AP-1 heterodimer (SupplementaryFigure S1) A comparison of fully CpG-methylatedversus nonndashmethylated-DNA identified several regionspreferentially bound by c-Junc-Fos if CpG-methylated(Supplementary Figure S1 bottom) three of which werelocated at EBV promoters BFLF2 BBLF4 and BALF1(Supplementary Figure S2A) Algorithms suitable toidentify sequences of potential binding sites readilyfound the known palindromic AP-1 consensus bindingsite 50-TGACTCA-30 (Figure 1) in samples ofunmethylated as well as methylated DNA confirmingthe validity of our approach Putative AP-1 bindingmotifs identified in CpG-methylated DNA were groupedaccording to whether they encompassed CpG pairs or notThe reanalysis revealed a second novel motif 50-TGACGTCG-30 which is closely related to the consensus AP-1 butmay contain up to two CpG dinucleotides We termed thismotif lsquomeAP-1rsquo in analogy to meZRE motifs bound byZta (5) Lists of the identified AP-1 binding sites areprovided in Supplementary Tables S1ndashS3
To determine whether the biochemically identifiedmeAP-1 sites in EBV DNA are also bound by c-Junc-Fos in cells in vivo we engineered a derivative of thelatently EBV-infected cell line Raji with a constitutivelyexpressed GFPc-Junc-Fos chimera ChIP followed bydeep sequencing analysis (ChIP-seq) of DNA fragmentsspecifically bound to GFPc-Junc-Fos protein was per-formed mapped to the EBV genome and analyzedbioinformatically in conjunction with methylation data(7) Confirming our results in vitro the analyzed pro-moters in Raji cells were only enriched if the meAP-1sites were highly CpG-methylated (SupplementaryFigure S2)
c-Junc-Fos binds CpG-methylated meAP-1 sites withhigh affinity mediating transcriptional activation
The different meAP-1 motifs identified in the EBV genomemay contain up to two CpG pairs (Figure 1) but the threemore prominent meAP-1 sites (Supplementary Figure S1)located in three EBV promoters encompass the sequence50-TGACTCG-30 with one CpG pair only (Figure 2A)We addressed this uncertainty together with the questionof whether 50-methylcytosine residues present in the topbottom or both strands of the meAP-1 DNA sequenceaffect the affinity of c-Junc-Fos binding to DNA
Unmethylated or CpG-methylated synthetic oligo-nucleotides containing the meAP-1 binding sites in theBBLF4 BFLF2 and BALF1 promoters (Figure 2A) andthree promoters identified later in ChIP-seq experimentswith Raji cell chromatin (Supplementary Figure S3) wereanalyzed in EMSAs As shown in Figure 2B c-Junc-Fosbound almost exclusively to the CpG-methylated meAP-1motif 50-TGACTCG-30 that contains a single CpG pairbut methylation of a CpG pair in a more central positionof the motif decreased the binding of c-Junc-Fos
(Supplementary Figure S3B) Therefore we concentratedour analysis on meAP-1 binding sites with a CpG pair atthe distal position The consensus meAP-1 site consists ofthe sequence motif 50-TGACTCG-30 and only the methylgroup in 50 position of the cytosine in the bottom strand isnecessary for the methylation-specific binding of c-Junc-Fos to DNA (Figure 2C)To quantify DNA binding the apparent dissociation
constants (KD) of c-Junc-Fos were measured withunmethylated or methylated meAP-1 binding sites TheKD values of BBLF4 BALF1 and BFLF2 meAP-1 siteswere lower by a factor of 387 100 and 171 respectivelyif methylated indicating that the presence of50-methylcytosine increases the binding affinity of c-Junc-Fos to DNA (Figure 2D) The consensus AP-1 bindingsite from the human lsquocollagenasersquo promoter (TREcollag)with a published KD for c-Junc-Fos of 108 nM (30) wasused as a positive controlThe transactivation potential of the c-Junc-Fos
heterodimer was investigated in luciferase reporterassays Reporter plasmids containing pentamers of thethree selected meAP-1 sites were transiently transfectedinto HEK293 cells together with an expression plasmidencoding VP16c-Junc-Fos in which the VP16transactivator domain was fused to the single-chain AP-1 heterodimer VP16c-Junc-Fos induced the CpG-methylated reporter plasmids by a factor of 2ndash5 ascompared with their non-methylated controls (Figure 2E)
The human genome contains meAP-1 binding sites
Two different cell lines were used to identify AP-1 andmeAP-1 binding sites in vivo the Burkitt lymphoma B-cell line Raji and the human fibroblast cell line Wi-38As explained earlier in the text a Raji cell line expressingthe GFPc-Junc-Fos chimera was used in ChIP-seq
AP-1 (35 sites) meAP-1 (9 sites)
no CpG CpG
AP-1 (61 sites) (44 sites)
unmethylated methylated
1 2 3 4 5 6 7
2
1
0
bits
-1 1 2 3 4 5 6 7 8
2
1
0
bits
-1 1 2 3 4 5 6 7 8 9
2
1
0
bits
-1 1 2 3 4 5 6 7
2
1
0
bits
Figure 1 Bioinformatic identification of c-Junc-Fos binding motifs inthe EBV genome Deep sequencing data were analyzed with the SISSRsalgorithms for putative stretches of DNA which c-Junc-Fos bindsThe output files of SISSRs were used as training sets for MEMEwhich identifies gapless local and multiple sequence motifs A totalof 61 motifs were identified in the DNA free of CpG methylationand 44 motifs were identified in the fully CpG-methylated EBVDNA The identified motifs in methylated DNA were selected at thelevel of the SISSRs training set data and grouped into motifs with andwithout CpG dinucleotides followed by MEME analysis
Nucleic Acids Research 2014 Vol 42 No 5 3063
A B
C
E
agtcggctagcTGACTCAgatgtcc
gagcgaTGACTCGtgtgcg
ggtctgTGACTCGttaatta
tcagggTGACTCGctaccc
TREcollag
BBLF4
BFLF2
BALF1
α-GFP - - + - - +
prot - + + - + +
unme me
- - + - - +
- + + - + +
- - + - - +
- + + - + +
BFLF2 BBLF4 BALF1
unme me unme me
001 01 1 10 100 10000
200
400
600
800
1000 BBLF4meBBLF4
nM oligo
Cy5
inte
nsi
t y
001 01 1 10 100 10000
200
400
600BALF1meBALF1
nM oligo001 01 1 10 100 10000
200
400
600
800
1000 BFLF2meBFLF2
nM oligo001 01 1 10 100 10000
200
400
600
TREcollag
nM oligo
4382nM 1997nM 2174nM 126nM
113nM 199nM 127nM
unmethylated
methylated nax387 x100 x171
D
prot - + - + - + - + - +
meth -- ++ +- -+
meAP-1 AP-1
meAP-1 AP-1
TGACTCGACTGAGC
TGACTCGACTGAGC
TGACTCAACTGAGT
-+
-- TGACTCGACTGAGC++
TGACTCGACTGAGC
+-
5xBFLF2 5xBBLF4 5xBALF1
Figure 2 The c-Junc-Fos binds preferentially to and activates gene transcription through CpG-methylated meAP-1 motifs (A) Oligonucleotidescontaining meAP-1 binding sites identified in the viral BBLF4 BALF1 and BFLF2 promoters The TREcollag oligo containing a consensus AP-1binding site was used as a control for the KD measurements The meAP-1 sites and AP-1 site are indicated in bold capital letters and the CpG pairswithin the meAP-1 sites are underlined (B) Preferential binding of c-Junc-Fos to methylated meAP-1 sites from the EBV genome EMSAs wereperformed with affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Supershifts with a GFP antibodyconfirmed the identity of the proteinndashDNA complexes EMSAs of typical experiments are shown as examples (C) Only one methyl group is
3064 Nucleic Acids Research 2014 Vol 42 No 5
(continued)
experiments In a second experimental setting serum-starved Wi-38 cells were infected with a GFPc-Junc-Fos-expressing adenovirus Chromatin from Wi-38(two days post infection) and Raji cells was preparedimmunoprecipitated with an a-GFP antibody to enrichDNA fragments specifically bound by GFPc-Junc-Fosprotein and analyzed with next-generation sequencing(Figure 3)
In accordance with our first functional analysis shownin Supplementary Figure S3B and discussed earlier in thetext we adjusted the bioinformatic parameters and refinedthe characteristics of the two AP-1 motifs found previ-ously consensus AP-1 motifs lacking a CpG dinucleotideand putative meAP-1 motifs with a single distal CpG pairin positions six and seven (Figure 3A and B) The twopredominant motifs revealed in both cellular modelsconfirm the known AP-1 consensus (50-TGACTCA-30)and identified an infrequent second meAP-1 motif(50-TGACGTCG-30) (Figure 3A and B) SupplementaryTables S4 through S7 provide the location of the AP-1 andmeAP-1 sites in both cell types
The majority of all consensus AP-1 sites are located inintergenic regions (3132) with no assigned functions Toconcentrate our further analysis on meAP-1 sites in prox-imity of promoter elements meAP-1 sites located within5 kb upstream of annotated TSS were considered only ifno other conventional AP-1 was located within a windowof plusmn1kb Only a small number of cellular genes fulfilledour criteria From ChIP-seq experiments of Raji cells weincluded the promoters of three genes in our study PTK6P2RY5 and SUV39H1 In Wi-38 cells we concentrated ontwo genes CBR3 and LRRTM2 A positive control thecollagenase promoter MMP1 containing two AP-1binding sites (33) was also included
Visualization of the coverage of these promoters afterChIP-seq analysis indicated that they are differentiallybound by c-Junc-Fos in the two cell types (Figure 3C)suggesting that the extent of cytosine methylation mightalso differ Bisulfite sequencing of the five promotersrevealed only one promoter element P2RY5 that wasfully CpG-methylated in both cell lines but CpG pairsin other promoters with meAP-1 sites differed in a celltype-specific manner (Supplementary Figure S4)
50-methylcytosine-dependent binding of c-Junc-Fos tomeAP-1 sites in the human genome inducestranscriptional activation
Only 5 of meAP-1 sites identified in both cell types arelocated in putative promoter regions and therefore
presumably involved in directly regulating gene expres-sion Three candidate meAP-1 motifs with no AP-1 con-sensus binding sites in their close proximity were analyzedfurther the meAP-1 sites in the PTK6 P2RY5 andSUV39H1 promoters (Figure 4) As shown in Figure 4Bc-Junc-Fos bound almost exclusively to the CpG-methylated meAP-1 motif In ELISAs the apparent dis-sociation constants (KD) of PTK6 and P2RY5 meAP-1sites could only be determined with methylated oligo-nucleotides because KD values of the unmethylatedprobes were too high to be estimated (Figure 4C) TheKD for the SUV39H1 meAP-1 site could not becalculated but the results confirm that the bindingaffinity is higher when the meAP-1 site is methylated(Figure 4B and C) These results indicate that thepresence of 50-methylcytosine favors the binding ofc-Junc-Fos to the meAP-1 sequences identified in thehuman genomeThe ability of c-Junc-Fos to transactivate gene tran-
scription through the binding to the methylated meAP-1sites from the human genome was tested in reporterassays Reporter plasmids containing pentamers ofselected meAP-1 sites in the human promoters werecotransfected with an expression plasmid encoding thetransactivator VP16c-Junc-Fos or a negative controlDNA Luciferase activities with CpG-methylatedreporter plasmids were significantly higher as comparedwith unmethylated reporter plasmid DNAs (20ndash27-foldFigure 4D)
Promoters with methylated meAP-1 sites are bound byc-Junc-Fos in vivo
So far our studies were based on the ectopic expression ofartificial c-Junc-Fos dimers to identify potential meAP-1sites in cellular DNA In contrast Raha et al (31) mappedthe location of endogenous c-Jun and c-Fos protein in twosets of ChIP-seq experiments and identified gt40 000binding sites in the established human myeloblastic cellline K562 To correlate their findings and ours we per-formed ChIP assays in this cell line with a c-Jun specificantibody and investigated by quantitative PCR analysisthe enrichment of the five previously identified promoterelements PTK6 P2RY5 SUV39H1 CBR3 and LRRTM2along with negative and positive control regions (34) Asshown in Supplementary Figure S5 the previouslyidentified five promoter elements which contain onlymeAP-1 but no consensus AP-1 sites were enriched atlevels similar to or exceeding those of the positivecontrols in K562 chromatin indicating that endogenous
Figure 2 Continuedresponsible for the binding of c-Junc-Fos to methylated DNA The meAP-1 and AP-1 binding sites contained in the oligonucleotide used in EMSAsare shown the asterisks represent a 50-methylcytosine residue EMSAs using BBLF4 and TREcollag oligonucleotides with the sites depicted in theleft panel and GFPc-Junc-Fos-purified protein are shown One representative experiment out of three is provided (D) Determination of theapparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion proteintransiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values The obtained data werefitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data point indicates themean and standard deviation of three independent experiments (E) Functional identification of single meAP-1 binding sites Pentamers of singlemeAP-1 sites present in the promoters of BBLF4 BALF1 and BFLF2 EBV genes as in (A) were introduced into a basic luciferase reporter plasmidwith a minimal EF1a promoter and free of CpG dinucleotides Unmethylated and fully CpG-methylated reporter constructs were analyzed in thepresence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Each experiment was performed three times and the means andstandard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3065
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

to the reference sequence and subsequent assembly wasperformed using the resequencing software Burrows-Wheeler Alignment (BWA) (v071 21) We used a com-bination of the recombinant EBV strain 2089 (16) and thebovine genome (build bosTau4) as reference sequences forthe two libraries lsquo4271unmethylatedrsquo and lsquo4271methylatedrsquo Calf thymus DNA was used as an unspecificcompetitor in the in vitro immunoprecipitation assays withGFPc-Junc-Fos The reference sequence for the in vivosamples lsquo4427Rajirsquo lsquo4673Wi38Inputrsquo and lsquo4673Wi38ChIPrsquo was the human reference sequence hg19
Transcription factor binding site detection
Genomic regions with a read depth above backgroundlevel are considered as regions containing transcriptionfactor binding sites We used two different programsSite Identification from Short Sequence Reads (SISSRs)(22) and QuEST (23) to identify these regionsTranscription factor binding sites are typically shorterthan the sequenced DNA fragments Binding sitesfor the sample libraries lsquo4271unmethylatedrsquo andlsquo4271methylatedrsquo were determined using SISSRsstandard parameters except for E (min number of direc-tional reads) which was set to 3200 for the librarylsquo4271unmethylatedrsquo and to 1600 for lsquo4271methylatedrsquoand F (average length of DNA fragments) which wasset to 200 in both cases Binding sites for the samplelsquo4427Rajirsquo were inferred using the default QuEST param-eters except for kernel density estimator and read countthreshold (th) which were set to 30 and 160 respectivelyIn the case of the sample lsquo4673Wi38ChIPrsquo it wasanalyzed by SISSRs with the results fromlsquo4673Wi38Inputrsquo as background file and setting th at 220 Toidentify consensus motifs within the discovered bindingsites the sequences containing possible binding siteswere extracted and used as input for the motif findingalgorithm Multiple EM for Motif Elicitation (MEME)(24)
ChIPs with an a-c-Jun antibody
ChIP experiments were conducted following standardprotocols Chromatin was cross-linked for 10minat room temperature and incubated with the antibodya-c-Jun (sc-44 Santa Cruz Biotechnology)
Quantitative real time PCR
Quantitative real time PCR (qRT-PCR) analysis was per-formed with a LightCycler 480 II instrument (Roche)with software version 150 SP4 according to the manu-facturerrsquos instructions Amplifications were monitoredwith the LightCycler FastStart reaction mix (SYBRgreen I Roche) The amplification of PCR products wasterminated after 45 cycles The following touchdown PCRprotocol was used (i) initial denaturation at 95 for 10minand (ii) 45 cycles of 95 (2 s) 65ndash62 (06 decrement inannealing temperaturecycle 10 s) 72 (10 s) For allprimer pairs a standard curve from serial dilutions ofthe PCR product with known concentration or the inputDNA was assessed Primer sequences are available onrequest
B cell isolation
Human primary B cells from adenoids were preparedand naıve B cells were sorted as described previously(1425) Anonymized adenoid samples from routineadenoidectomies were provided by the Department ofOtorhinolaryngology Klinikum Grosshadern Ludwig-Maximilians-University of Munich The institutionalreview board Ethikkommission of the KlinikumGrosshadern approved the study and did not requireprior informed patient consent
Bioinformatic identification of meAP-1 sites in primaryhuman B cells
In the reference human genome assembly GRCh37hg19we detectedAP-1 andmeAP-1 binding sites using the searchalgorithm Find Individual Motif Occurrences (FIMO) (26)based on our MEME output motifs Called sites weremapped to promoter sequences of RefSeq genes containedin theUCSCGenome Browser5 kb to+1kb of their tran-scription start sites (TSS) (27) Predicted meAP-1 bindingsite within promoter sequences were further evaluated forCpG methylation in naıve B cells (28) (C04M7ACXX_XFD088DACXX_XF D08E5ACXX_XF) For furtheranalysis we selected only meAP-1 binding sites that showat least 80 methylation in two of three replicates Toidentify genes upregulated in activated B cells we used ex-pression data obtained from unstimulated (GSM907730GSM907738 GSM907746) and stimulated (GSM907731GSM907739 GSM907747) B cell data sets available onGene Expression Omnibus (GSE36975) (29) We selectedtranscripts that were significantly expressed in 5 of 6samples Transcripts showing significant differential ex-pression between unstimulated and stimulated B cellswere identified performing a one way ANOVA test usingMATLAB
RESULTS
c-Junc-Fos binding motifs in genomic EBV DNA
We started our search with a synthetic model whichbuilds on the entire DNA genome of EBV cloned inE coli as a single plasmid (16) This DNA is large(180 kb) GC-rich (60) free of methylated cytosine nu-cleotides (which can be easily methylated in vitro)contains 85 genes and diverse promoter elements andhas been used previously for the analysis of Ztarsquos DNAbinding characteristics (5) We predicted that the DNArsquoscomplexity and size is suitable to identify putative DNAmotifs which are targeted by AP-1 if methylated Thesecond component in our model is a single-chain c-Junc-Fos protein designed to ensure the formation of a singledefined AP-1 heterodimer unaffected by other proteinsthat complex with either c-Jun or c-Fos Bakiri et al(17) showed that similar constructs with the murine se-quences of c-Jun and c-Fos specifically bind andtransactivate cellular promoters that contain the consen-sus AP-1 binding site
We used in vitro pull-down experiments as described (5)with randomly sheared nonndashCpG-methylated or fully
3062 Nucleic Acids Research 2014 Vol 42 No 5
CpG-methylated EBV DNA and a GFPc-Junc-Fosfusion protein purified from HEK293 cellsImmunoprecipitation of the fusion protein followed bydeep sequencing of the enriched DNA fragmentsprovided an extensive coverage of DNA sequences select-ively bound by the AP-1 heterodimer (SupplementaryFigure S1) A comparison of fully CpG-methylatedversus nonndashmethylated-DNA identified several regionspreferentially bound by c-Junc-Fos if CpG-methylated(Supplementary Figure S1 bottom) three of which werelocated at EBV promoters BFLF2 BBLF4 and BALF1(Supplementary Figure S2A) Algorithms suitable toidentify sequences of potential binding sites readilyfound the known palindromic AP-1 consensus bindingsite 50-TGACTCA-30 (Figure 1) in samples ofunmethylated as well as methylated DNA confirmingthe validity of our approach Putative AP-1 bindingmotifs identified in CpG-methylated DNA were groupedaccording to whether they encompassed CpG pairs or notThe reanalysis revealed a second novel motif 50-TGACGTCG-30 which is closely related to the consensus AP-1 butmay contain up to two CpG dinucleotides We termed thismotif lsquomeAP-1rsquo in analogy to meZRE motifs bound byZta (5) Lists of the identified AP-1 binding sites areprovided in Supplementary Tables S1ndashS3
To determine whether the biochemically identifiedmeAP-1 sites in EBV DNA are also bound by c-Junc-Fos in cells in vivo we engineered a derivative of thelatently EBV-infected cell line Raji with a constitutivelyexpressed GFPc-Junc-Fos chimera ChIP followed bydeep sequencing analysis (ChIP-seq) of DNA fragmentsspecifically bound to GFPc-Junc-Fos protein was per-formed mapped to the EBV genome and analyzedbioinformatically in conjunction with methylation data(7) Confirming our results in vitro the analyzed pro-moters in Raji cells were only enriched if the meAP-1sites were highly CpG-methylated (SupplementaryFigure S2)
c-Junc-Fos binds CpG-methylated meAP-1 sites withhigh affinity mediating transcriptional activation
The different meAP-1 motifs identified in the EBV genomemay contain up to two CpG pairs (Figure 1) but the threemore prominent meAP-1 sites (Supplementary Figure S1)located in three EBV promoters encompass the sequence50-TGACTCG-30 with one CpG pair only (Figure 2A)We addressed this uncertainty together with the questionof whether 50-methylcytosine residues present in the topbottom or both strands of the meAP-1 DNA sequenceaffect the affinity of c-Junc-Fos binding to DNA
Unmethylated or CpG-methylated synthetic oligo-nucleotides containing the meAP-1 binding sites in theBBLF4 BFLF2 and BALF1 promoters (Figure 2A) andthree promoters identified later in ChIP-seq experimentswith Raji cell chromatin (Supplementary Figure S3) wereanalyzed in EMSAs As shown in Figure 2B c-Junc-Fosbound almost exclusively to the CpG-methylated meAP-1motif 50-TGACTCG-30 that contains a single CpG pairbut methylation of a CpG pair in a more central positionof the motif decreased the binding of c-Junc-Fos
(Supplementary Figure S3B) Therefore we concentratedour analysis on meAP-1 binding sites with a CpG pair atthe distal position The consensus meAP-1 site consists ofthe sequence motif 50-TGACTCG-30 and only the methylgroup in 50 position of the cytosine in the bottom strand isnecessary for the methylation-specific binding of c-Junc-Fos to DNA (Figure 2C)To quantify DNA binding the apparent dissociation
constants (KD) of c-Junc-Fos were measured withunmethylated or methylated meAP-1 binding sites TheKD values of BBLF4 BALF1 and BFLF2 meAP-1 siteswere lower by a factor of 387 100 and 171 respectivelyif methylated indicating that the presence of50-methylcytosine increases the binding affinity of c-Junc-Fos to DNA (Figure 2D) The consensus AP-1 bindingsite from the human lsquocollagenasersquo promoter (TREcollag)with a published KD for c-Junc-Fos of 108 nM (30) wasused as a positive controlThe transactivation potential of the c-Junc-Fos
heterodimer was investigated in luciferase reporterassays Reporter plasmids containing pentamers of thethree selected meAP-1 sites were transiently transfectedinto HEK293 cells together with an expression plasmidencoding VP16c-Junc-Fos in which the VP16transactivator domain was fused to the single-chain AP-1 heterodimer VP16c-Junc-Fos induced the CpG-methylated reporter plasmids by a factor of 2ndash5 ascompared with their non-methylated controls (Figure 2E)
The human genome contains meAP-1 binding sites
Two different cell lines were used to identify AP-1 andmeAP-1 binding sites in vivo the Burkitt lymphoma B-cell line Raji and the human fibroblast cell line Wi-38As explained earlier in the text a Raji cell line expressingthe GFPc-Junc-Fos chimera was used in ChIP-seq
AP-1 (35 sites) meAP-1 (9 sites)
no CpG CpG
AP-1 (61 sites) (44 sites)
unmethylated methylated
1 2 3 4 5 6 7
2
1
0
bits
-1 1 2 3 4 5 6 7 8
2
1
0
bits
-1 1 2 3 4 5 6 7 8 9
2
1
0
bits
-1 1 2 3 4 5 6 7
2
1
0
bits
Figure 1 Bioinformatic identification of c-Junc-Fos binding motifs inthe EBV genome Deep sequencing data were analyzed with the SISSRsalgorithms for putative stretches of DNA which c-Junc-Fos bindsThe output files of SISSRs were used as training sets for MEMEwhich identifies gapless local and multiple sequence motifs A totalof 61 motifs were identified in the DNA free of CpG methylationand 44 motifs were identified in the fully CpG-methylated EBVDNA The identified motifs in methylated DNA were selected at thelevel of the SISSRs training set data and grouped into motifs with andwithout CpG dinucleotides followed by MEME analysis
Nucleic Acids Research 2014 Vol 42 No 5 3063
A B
C
E
agtcggctagcTGACTCAgatgtcc
gagcgaTGACTCGtgtgcg
ggtctgTGACTCGttaatta
tcagggTGACTCGctaccc
TREcollag
BBLF4
BFLF2
BALF1
α-GFP - - + - - +
prot - + + - + +
unme me
- - + - - +
- + + - + +
- - + - - +
- + + - + +
BFLF2 BBLF4 BALF1
unme me unme me
001 01 1 10 100 10000
200
400
600
800
1000 BBLF4meBBLF4
nM oligo
Cy5
inte
nsi
t y
001 01 1 10 100 10000
200
400
600BALF1meBALF1
nM oligo001 01 1 10 100 10000
200
400
600
800
1000 BFLF2meBFLF2
nM oligo001 01 1 10 100 10000
200
400
600
TREcollag
nM oligo
4382nM 1997nM 2174nM 126nM
113nM 199nM 127nM
unmethylated
methylated nax387 x100 x171
D
prot - + - + - + - + - +
meth -- ++ +- -+
meAP-1 AP-1
meAP-1 AP-1
TGACTCGACTGAGC
TGACTCGACTGAGC
TGACTCAACTGAGT
-+
-- TGACTCGACTGAGC++
TGACTCGACTGAGC
+-
5xBFLF2 5xBBLF4 5xBALF1
Figure 2 The c-Junc-Fos binds preferentially to and activates gene transcription through CpG-methylated meAP-1 motifs (A) Oligonucleotidescontaining meAP-1 binding sites identified in the viral BBLF4 BALF1 and BFLF2 promoters The TREcollag oligo containing a consensus AP-1binding site was used as a control for the KD measurements The meAP-1 sites and AP-1 site are indicated in bold capital letters and the CpG pairswithin the meAP-1 sites are underlined (B) Preferential binding of c-Junc-Fos to methylated meAP-1 sites from the EBV genome EMSAs wereperformed with affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Supershifts with a GFP antibodyconfirmed the identity of the proteinndashDNA complexes EMSAs of typical experiments are shown as examples (C) Only one methyl group is
3064 Nucleic Acids Research 2014 Vol 42 No 5
(continued)
experiments In a second experimental setting serum-starved Wi-38 cells were infected with a GFPc-Junc-Fos-expressing adenovirus Chromatin from Wi-38(two days post infection) and Raji cells was preparedimmunoprecipitated with an a-GFP antibody to enrichDNA fragments specifically bound by GFPc-Junc-Fosprotein and analyzed with next-generation sequencing(Figure 3)
In accordance with our first functional analysis shownin Supplementary Figure S3B and discussed earlier in thetext we adjusted the bioinformatic parameters and refinedthe characteristics of the two AP-1 motifs found previ-ously consensus AP-1 motifs lacking a CpG dinucleotideand putative meAP-1 motifs with a single distal CpG pairin positions six and seven (Figure 3A and B) The twopredominant motifs revealed in both cellular modelsconfirm the known AP-1 consensus (50-TGACTCA-30)and identified an infrequent second meAP-1 motif(50-TGACGTCG-30) (Figure 3A and B) SupplementaryTables S4 through S7 provide the location of the AP-1 andmeAP-1 sites in both cell types
The majority of all consensus AP-1 sites are located inintergenic regions (3132) with no assigned functions Toconcentrate our further analysis on meAP-1 sites in prox-imity of promoter elements meAP-1 sites located within5 kb upstream of annotated TSS were considered only ifno other conventional AP-1 was located within a windowof plusmn1kb Only a small number of cellular genes fulfilledour criteria From ChIP-seq experiments of Raji cells weincluded the promoters of three genes in our study PTK6P2RY5 and SUV39H1 In Wi-38 cells we concentrated ontwo genes CBR3 and LRRTM2 A positive control thecollagenase promoter MMP1 containing two AP-1binding sites (33) was also included
Visualization of the coverage of these promoters afterChIP-seq analysis indicated that they are differentiallybound by c-Junc-Fos in the two cell types (Figure 3C)suggesting that the extent of cytosine methylation mightalso differ Bisulfite sequencing of the five promotersrevealed only one promoter element P2RY5 that wasfully CpG-methylated in both cell lines but CpG pairsin other promoters with meAP-1 sites differed in a celltype-specific manner (Supplementary Figure S4)
50-methylcytosine-dependent binding of c-Junc-Fos tomeAP-1 sites in the human genome inducestranscriptional activation
Only 5 of meAP-1 sites identified in both cell types arelocated in putative promoter regions and therefore
presumably involved in directly regulating gene expres-sion Three candidate meAP-1 motifs with no AP-1 con-sensus binding sites in their close proximity were analyzedfurther the meAP-1 sites in the PTK6 P2RY5 andSUV39H1 promoters (Figure 4) As shown in Figure 4Bc-Junc-Fos bound almost exclusively to the CpG-methylated meAP-1 motif In ELISAs the apparent dis-sociation constants (KD) of PTK6 and P2RY5 meAP-1sites could only be determined with methylated oligo-nucleotides because KD values of the unmethylatedprobes were too high to be estimated (Figure 4C) TheKD for the SUV39H1 meAP-1 site could not becalculated but the results confirm that the bindingaffinity is higher when the meAP-1 site is methylated(Figure 4B and C) These results indicate that thepresence of 50-methylcytosine favors the binding ofc-Junc-Fos to the meAP-1 sequences identified in thehuman genomeThe ability of c-Junc-Fos to transactivate gene tran-
scription through the binding to the methylated meAP-1sites from the human genome was tested in reporterassays Reporter plasmids containing pentamers ofselected meAP-1 sites in the human promoters werecotransfected with an expression plasmid encoding thetransactivator VP16c-Junc-Fos or a negative controlDNA Luciferase activities with CpG-methylatedreporter plasmids were significantly higher as comparedwith unmethylated reporter plasmid DNAs (20ndash27-foldFigure 4D)
Promoters with methylated meAP-1 sites are bound byc-Junc-Fos in vivo
So far our studies were based on the ectopic expression ofartificial c-Junc-Fos dimers to identify potential meAP-1sites in cellular DNA In contrast Raha et al (31) mappedthe location of endogenous c-Jun and c-Fos protein in twosets of ChIP-seq experiments and identified gt40 000binding sites in the established human myeloblastic cellline K562 To correlate their findings and ours we per-formed ChIP assays in this cell line with a c-Jun specificantibody and investigated by quantitative PCR analysisthe enrichment of the five previously identified promoterelements PTK6 P2RY5 SUV39H1 CBR3 and LRRTM2along with negative and positive control regions (34) Asshown in Supplementary Figure S5 the previouslyidentified five promoter elements which contain onlymeAP-1 but no consensus AP-1 sites were enriched atlevels similar to or exceeding those of the positivecontrols in K562 chromatin indicating that endogenous
Figure 2 Continuedresponsible for the binding of c-Junc-Fos to methylated DNA The meAP-1 and AP-1 binding sites contained in the oligonucleotide used in EMSAsare shown the asterisks represent a 50-methylcytosine residue EMSAs using BBLF4 and TREcollag oligonucleotides with the sites depicted in theleft panel and GFPc-Junc-Fos-purified protein are shown One representative experiment out of three is provided (D) Determination of theapparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion proteintransiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values The obtained data werefitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data point indicates themean and standard deviation of three independent experiments (E) Functional identification of single meAP-1 binding sites Pentamers of singlemeAP-1 sites present in the promoters of BBLF4 BALF1 and BFLF2 EBV genes as in (A) were introduced into a basic luciferase reporter plasmidwith a minimal EF1a promoter and free of CpG dinucleotides Unmethylated and fully CpG-methylated reporter constructs were analyzed in thepresence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Each experiment was performed three times and the means andstandard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3065
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

CpG-methylated EBV DNA and a GFPc-Junc-Fosfusion protein purified from HEK293 cellsImmunoprecipitation of the fusion protein followed bydeep sequencing of the enriched DNA fragmentsprovided an extensive coverage of DNA sequences select-ively bound by the AP-1 heterodimer (SupplementaryFigure S1) A comparison of fully CpG-methylatedversus nonndashmethylated-DNA identified several regionspreferentially bound by c-Junc-Fos if CpG-methylated(Supplementary Figure S1 bottom) three of which werelocated at EBV promoters BFLF2 BBLF4 and BALF1(Supplementary Figure S2A) Algorithms suitable toidentify sequences of potential binding sites readilyfound the known palindromic AP-1 consensus bindingsite 50-TGACTCA-30 (Figure 1) in samples ofunmethylated as well as methylated DNA confirmingthe validity of our approach Putative AP-1 bindingmotifs identified in CpG-methylated DNA were groupedaccording to whether they encompassed CpG pairs or notThe reanalysis revealed a second novel motif 50-TGACGTCG-30 which is closely related to the consensus AP-1 butmay contain up to two CpG dinucleotides We termed thismotif lsquomeAP-1rsquo in analogy to meZRE motifs bound byZta (5) Lists of the identified AP-1 binding sites areprovided in Supplementary Tables S1ndashS3
To determine whether the biochemically identifiedmeAP-1 sites in EBV DNA are also bound by c-Junc-Fos in cells in vivo we engineered a derivative of thelatently EBV-infected cell line Raji with a constitutivelyexpressed GFPc-Junc-Fos chimera ChIP followed bydeep sequencing analysis (ChIP-seq) of DNA fragmentsspecifically bound to GFPc-Junc-Fos protein was per-formed mapped to the EBV genome and analyzedbioinformatically in conjunction with methylation data(7) Confirming our results in vitro the analyzed pro-moters in Raji cells were only enriched if the meAP-1sites were highly CpG-methylated (SupplementaryFigure S2)
c-Junc-Fos binds CpG-methylated meAP-1 sites withhigh affinity mediating transcriptional activation
The different meAP-1 motifs identified in the EBV genomemay contain up to two CpG pairs (Figure 1) but the threemore prominent meAP-1 sites (Supplementary Figure S1)located in three EBV promoters encompass the sequence50-TGACTCG-30 with one CpG pair only (Figure 2A)We addressed this uncertainty together with the questionof whether 50-methylcytosine residues present in the topbottom or both strands of the meAP-1 DNA sequenceaffect the affinity of c-Junc-Fos binding to DNA
Unmethylated or CpG-methylated synthetic oligo-nucleotides containing the meAP-1 binding sites in theBBLF4 BFLF2 and BALF1 promoters (Figure 2A) andthree promoters identified later in ChIP-seq experimentswith Raji cell chromatin (Supplementary Figure S3) wereanalyzed in EMSAs As shown in Figure 2B c-Junc-Fosbound almost exclusively to the CpG-methylated meAP-1motif 50-TGACTCG-30 that contains a single CpG pairbut methylation of a CpG pair in a more central positionof the motif decreased the binding of c-Junc-Fos
(Supplementary Figure S3B) Therefore we concentratedour analysis on meAP-1 binding sites with a CpG pair atthe distal position The consensus meAP-1 site consists ofthe sequence motif 50-TGACTCG-30 and only the methylgroup in 50 position of the cytosine in the bottom strand isnecessary for the methylation-specific binding of c-Junc-Fos to DNA (Figure 2C)To quantify DNA binding the apparent dissociation
constants (KD) of c-Junc-Fos were measured withunmethylated or methylated meAP-1 binding sites TheKD values of BBLF4 BALF1 and BFLF2 meAP-1 siteswere lower by a factor of 387 100 and 171 respectivelyif methylated indicating that the presence of50-methylcytosine increases the binding affinity of c-Junc-Fos to DNA (Figure 2D) The consensus AP-1 bindingsite from the human lsquocollagenasersquo promoter (TREcollag)with a published KD for c-Junc-Fos of 108 nM (30) wasused as a positive controlThe transactivation potential of the c-Junc-Fos
heterodimer was investigated in luciferase reporterassays Reporter plasmids containing pentamers of thethree selected meAP-1 sites were transiently transfectedinto HEK293 cells together with an expression plasmidencoding VP16c-Junc-Fos in which the VP16transactivator domain was fused to the single-chain AP-1 heterodimer VP16c-Junc-Fos induced the CpG-methylated reporter plasmids by a factor of 2ndash5 ascompared with their non-methylated controls (Figure 2E)
The human genome contains meAP-1 binding sites
Two different cell lines were used to identify AP-1 andmeAP-1 binding sites in vivo the Burkitt lymphoma B-cell line Raji and the human fibroblast cell line Wi-38As explained earlier in the text a Raji cell line expressingthe GFPc-Junc-Fos chimera was used in ChIP-seq
AP-1 (35 sites) meAP-1 (9 sites)
no CpG CpG
AP-1 (61 sites) (44 sites)
unmethylated methylated
1 2 3 4 5 6 7
2
1
0
bits
-1 1 2 3 4 5 6 7 8
2
1
0
bits
-1 1 2 3 4 5 6 7 8 9
2
1
0
bits
-1 1 2 3 4 5 6 7
2
1
0
bits
Figure 1 Bioinformatic identification of c-Junc-Fos binding motifs inthe EBV genome Deep sequencing data were analyzed with the SISSRsalgorithms for putative stretches of DNA which c-Junc-Fos bindsThe output files of SISSRs were used as training sets for MEMEwhich identifies gapless local and multiple sequence motifs A totalof 61 motifs were identified in the DNA free of CpG methylationand 44 motifs were identified in the fully CpG-methylated EBVDNA The identified motifs in methylated DNA were selected at thelevel of the SISSRs training set data and grouped into motifs with andwithout CpG dinucleotides followed by MEME analysis
Nucleic Acids Research 2014 Vol 42 No 5 3063
A B
C
E
agtcggctagcTGACTCAgatgtcc
gagcgaTGACTCGtgtgcg
ggtctgTGACTCGttaatta
tcagggTGACTCGctaccc
TREcollag
BBLF4
BFLF2
BALF1
α-GFP - - + - - +
prot - + + - + +
unme me
- - + - - +
- + + - + +
- - + - - +
- + + - + +
BFLF2 BBLF4 BALF1
unme me unme me
001 01 1 10 100 10000
200
400
600
800
1000 BBLF4meBBLF4
nM oligo
Cy5
inte
nsi
t y
001 01 1 10 100 10000
200
400
600BALF1meBALF1
nM oligo001 01 1 10 100 10000
200
400
600
800
1000 BFLF2meBFLF2
nM oligo001 01 1 10 100 10000
200
400
600
TREcollag
nM oligo
4382nM 1997nM 2174nM 126nM
113nM 199nM 127nM
unmethylated
methylated nax387 x100 x171
D
prot - + - + - + - + - +
meth -- ++ +- -+
meAP-1 AP-1
meAP-1 AP-1
TGACTCGACTGAGC
TGACTCGACTGAGC
TGACTCAACTGAGT
-+
-- TGACTCGACTGAGC++
TGACTCGACTGAGC
+-
5xBFLF2 5xBBLF4 5xBALF1
Figure 2 The c-Junc-Fos binds preferentially to and activates gene transcription through CpG-methylated meAP-1 motifs (A) Oligonucleotidescontaining meAP-1 binding sites identified in the viral BBLF4 BALF1 and BFLF2 promoters The TREcollag oligo containing a consensus AP-1binding site was used as a control for the KD measurements The meAP-1 sites and AP-1 site are indicated in bold capital letters and the CpG pairswithin the meAP-1 sites are underlined (B) Preferential binding of c-Junc-Fos to methylated meAP-1 sites from the EBV genome EMSAs wereperformed with affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Supershifts with a GFP antibodyconfirmed the identity of the proteinndashDNA complexes EMSAs of typical experiments are shown as examples (C) Only one methyl group is
3064 Nucleic Acids Research 2014 Vol 42 No 5
(continued)
experiments In a second experimental setting serum-starved Wi-38 cells were infected with a GFPc-Junc-Fos-expressing adenovirus Chromatin from Wi-38(two days post infection) and Raji cells was preparedimmunoprecipitated with an a-GFP antibody to enrichDNA fragments specifically bound by GFPc-Junc-Fosprotein and analyzed with next-generation sequencing(Figure 3)
In accordance with our first functional analysis shownin Supplementary Figure S3B and discussed earlier in thetext we adjusted the bioinformatic parameters and refinedthe characteristics of the two AP-1 motifs found previ-ously consensus AP-1 motifs lacking a CpG dinucleotideand putative meAP-1 motifs with a single distal CpG pairin positions six and seven (Figure 3A and B) The twopredominant motifs revealed in both cellular modelsconfirm the known AP-1 consensus (50-TGACTCA-30)and identified an infrequent second meAP-1 motif(50-TGACGTCG-30) (Figure 3A and B) SupplementaryTables S4 through S7 provide the location of the AP-1 andmeAP-1 sites in both cell types
The majority of all consensus AP-1 sites are located inintergenic regions (3132) with no assigned functions Toconcentrate our further analysis on meAP-1 sites in prox-imity of promoter elements meAP-1 sites located within5 kb upstream of annotated TSS were considered only ifno other conventional AP-1 was located within a windowof plusmn1kb Only a small number of cellular genes fulfilledour criteria From ChIP-seq experiments of Raji cells weincluded the promoters of three genes in our study PTK6P2RY5 and SUV39H1 In Wi-38 cells we concentrated ontwo genes CBR3 and LRRTM2 A positive control thecollagenase promoter MMP1 containing two AP-1binding sites (33) was also included
Visualization of the coverage of these promoters afterChIP-seq analysis indicated that they are differentiallybound by c-Junc-Fos in the two cell types (Figure 3C)suggesting that the extent of cytosine methylation mightalso differ Bisulfite sequencing of the five promotersrevealed only one promoter element P2RY5 that wasfully CpG-methylated in both cell lines but CpG pairsin other promoters with meAP-1 sites differed in a celltype-specific manner (Supplementary Figure S4)
50-methylcytosine-dependent binding of c-Junc-Fos tomeAP-1 sites in the human genome inducestranscriptional activation
Only 5 of meAP-1 sites identified in both cell types arelocated in putative promoter regions and therefore
presumably involved in directly regulating gene expres-sion Three candidate meAP-1 motifs with no AP-1 con-sensus binding sites in their close proximity were analyzedfurther the meAP-1 sites in the PTK6 P2RY5 andSUV39H1 promoters (Figure 4) As shown in Figure 4Bc-Junc-Fos bound almost exclusively to the CpG-methylated meAP-1 motif In ELISAs the apparent dis-sociation constants (KD) of PTK6 and P2RY5 meAP-1sites could only be determined with methylated oligo-nucleotides because KD values of the unmethylatedprobes were too high to be estimated (Figure 4C) TheKD for the SUV39H1 meAP-1 site could not becalculated but the results confirm that the bindingaffinity is higher when the meAP-1 site is methylated(Figure 4B and C) These results indicate that thepresence of 50-methylcytosine favors the binding ofc-Junc-Fos to the meAP-1 sequences identified in thehuman genomeThe ability of c-Junc-Fos to transactivate gene tran-
scription through the binding to the methylated meAP-1sites from the human genome was tested in reporterassays Reporter plasmids containing pentamers ofselected meAP-1 sites in the human promoters werecotransfected with an expression plasmid encoding thetransactivator VP16c-Junc-Fos or a negative controlDNA Luciferase activities with CpG-methylatedreporter plasmids were significantly higher as comparedwith unmethylated reporter plasmid DNAs (20ndash27-foldFigure 4D)
Promoters with methylated meAP-1 sites are bound byc-Junc-Fos in vivo
So far our studies were based on the ectopic expression ofartificial c-Junc-Fos dimers to identify potential meAP-1sites in cellular DNA In contrast Raha et al (31) mappedthe location of endogenous c-Jun and c-Fos protein in twosets of ChIP-seq experiments and identified gt40 000binding sites in the established human myeloblastic cellline K562 To correlate their findings and ours we per-formed ChIP assays in this cell line with a c-Jun specificantibody and investigated by quantitative PCR analysisthe enrichment of the five previously identified promoterelements PTK6 P2RY5 SUV39H1 CBR3 and LRRTM2along with negative and positive control regions (34) Asshown in Supplementary Figure S5 the previouslyidentified five promoter elements which contain onlymeAP-1 but no consensus AP-1 sites were enriched atlevels similar to or exceeding those of the positivecontrols in K562 chromatin indicating that endogenous
Figure 2 Continuedresponsible for the binding of c-Junc-Fos to methylated DNA The meAP-1 and AP-1 binding sites contained in the oligonucleotide used in EMSAsare shown the asterisks represent a 50-methylcytosine residue EMSAs using BBLF4 and TREcollag oligonucleotides with the sites depicted in theleft panel and GFPc-Junc-Fos-purified protein are shown One representative experiment out of three is provided (D) Determination of theapparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion proteintransiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values The obtained data werefitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data point indicates themean and standard deviation of three independent experiments (E) Functional identification of single meAP-1 binding sites Pentamers of singlemeAP-1 sites present in the promoters of BBLF4 BALF1 and BFLF2 EBV genes as in (A) were introduced into a basic luciferase reporter plasmidwith a minimal EF1a promoter and free of CpG dinucleotides Unmethylated and fully CpG-methylated reporter constructs were analyzed in thepresence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Each experiment was performed three times and the means andstandard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3065
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

A B
C
E
agtcggctagcTGACTCAgatgtcc
gagcgaTGACTCGtgtgcg
ggtctgTGACTCGttaatta
tcagggTGACTCGctaccc
TREcollag
BBLF4
BFLF2
BALF1
α-GFP - - + - - +
prot - + + - + +
unme me
- - + - - +
- + + - + +
- - + - - +
- + + - + +
BFLF2 BBLF4 BALF1
unme me unme me
001 01 1 10 100 10000
200
400
600
800
1000 BBLF4meBBLF4
nM oligo
Cy5
inte
nsi
t y
001 01 1 10 100 10000
200
400
600BALF1meBALF1
nM oligo001 01 1 10 100 10000
200
400
600
800
1000 BFLF2meBFLF2
nM oligo001 01 1 10 100 10000
200
400
600
TREcollag
nM oligo
4382nM 1997nM 2174nM 126nM
113nM 199nM 127nM
unmethylated
methylated nax387 x100 x171
D
prot - + - + - + - + - +
meth -- ++ +- -+
meAP-1 AP-1
meAP-1 AP-1
TGACTCGACTGAGC
TGACTCGACTGAGC
TGACTCAACTGAGT
-+
-- TGACTCGACTGAGC++
TGACTCGACTGAGC
+-
5xBFLF2 5xBBLF4 5xBALF1
Figure 2 The c-Junc-Fos binds preferentially to and activates gene transcription through CpG-methylated meAP-1 motifs (A) Oligonucleotidescontaining meAP-1 binding sites identified in the viral BBLF4 BALF1 and BFLF2 promoters The TREcollag oligo containing a consensus AP-1binding site was used as a control for the KD measurements The meAP-1 sites and AP-1 site are indicated in bold capital letters and the CpG pairswithin the meAP-1 sites are underlined (B) Preferential binding of c-Junc-Fos to methylated meAP-1 sites from the EBV genome EMSAs wereperformed with affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Supershifts with a GFP antibodyconfirmed the identity of the proteinndashDNA complexes EMSAs of typical experiments are shown as examples (C) Only one methyl group is
3064 Nucleic Acids Research 2014 Vol 42 No 5
(continued)
experiments In a second experimental setting serum-starved Wi-38 cells were infected with a GFPc-Junc-Fos-expressing adenovirus Chromatin from Wi-38(two days post infection) and Raji cells was preparedimmunoprecipitated with an a-GFP antibody to enrichDNA fragments specifically bound by GFPc-Junc-Fosprotein and analyzed with next-generation sequencing(Figure 3)
In accordance with our first functional analysis shownin Supplementary Figure S3B and discussed earlier in thetext we adjusted the bioinformatic parameters and refinedthe characteristics of the two AP-1 motifs found previ-ously consensus AP-1 motifs lacking a CpG dinucleotideand putative meAP-1 motifs with a single distal CpG pairin positions six and seven (Figure 3A and B) The twopredominant motifs revealed in both cellular modelsconfirm the known AP-1 consensus (50-TGACTCA-30)and identified an infrequent second meAP-1 motif(50-TGACGTCG-30) (Figure 3A and B) SupplementaryTables S4 through S7 provide the location of the AP-1 andmeAP-1 sites in both cell types
The majority of all consensus AP-1 sites are located inintergenic regions (3132) with no assigned functions Toconcentrate our further analysis on meAP-1 sites in prox-imity of promoter elements meAP-1 sites located within5 kb upstream of annotated TSS were considered only ifno other conventional AP-1 was located within a windowof plusmn1kb Only a small number of cellular genes fulfilledour criteria From ChIP-seq experiments of Raji cells weincluded the promoters of three genes in our study PTK6P2RY5 and SUV39H1 In Wi-38 cells we concentrated ontwo genes CBR3 and LRRTM2 A positive control thecollagenase promoter MMP1 containing two AP-1binding sites (33) was also included
Visualization of the coverage of these promoters afterChIP-seq analysis indicated that they are differentiallybound by c-Junc-Fos in the two cell types (Figure 3C)suggesting that the extent of cytosine methylation mightalso differ Bisulfite sequencing of the five promotersrevealed only one promoter element P2RY5 that wasfully CpG-methylated in both cell lines but CpG pairsin other promoters with meAP-1 sites differed in a celltype-specific manner (Supplementary Figure S4)
50-methylcytosine-dependent binding of c-Junc-Fos tomeAP-1 sites in the human genome inducestranscriptional activation
Only 5 of meAP-1 sites identified in both cell types arelocated in putative promoter regions and therefore
presumably involved in directly regulating gene expres-sion Three candidate meAP-1 motifs with no AP-1 con-sensus binding sites in their close proximity were analyzedfurther the meAP-1 sites in the PTK6 P2RY5 andSUV39H1 promoters (Figure 4) As shown in Figure 4Bc-Junc-Fos bound almost exclusively to the CpG-methylated meAP-1 motif In ELISAs the apparent dis-sociation constants (KD) of PTK6 and P2RY5 meAP-1sites could only be determined with methylated oligo-nucleotides because KD values of the unmethylatedprobes were too high to be estimated (Figure 4C) TheKD for the SUV39H1 meAP-1 site could not becalculated but the results confirm that the bindingaffinity is higher when the meAP-1 site is methylated(Figure 4B and C) These results indicate that thepresence of 50-methylcytosine favors the binding ofc-Junc-Fos to the meAP-1 sequences identified in thehuman genomeThe ability of c-Junc-Fos to transactivate gene tran-
scription through the binding to the methylated meAP-1sites from the human genome was tested in reporterassays Reporter plasmids containing pentamers ofselected meAP-1 sites in the human promoters werecotransfected with an expression plasmid encoding thetransactivator VP16c-Junc-Fos or a negative controlDNA Luciferase activities with CpG-methylatedreporter plasmids were significantly higher as comparedwith unmethylated reporter plasmid DNAs (20ndash27-foldFigure 4D)
Promoters with methylated meAP-1 sites are bound byc-Junc-Fos in vivo
So far our studies were based on the ectopic expression ofartificial c-Junc-Fos dimers to identify potential meAP-1sites in cellular DNA In contrast Raha et al (31) mappedthe location of endogenous c-Jun and c-Fos protein in twosets of ChIP-seq experiments and identified gt40 000binding sites in the established human myeloblastic cellline K562 To correlate their findings and ours we per-formed ChIP assays in this cell line with a c-Jun specificantibody and investigated by quantitative PCR analysisthe enrichment of the five previously identified promoterelements PTK6 P2RY5 SUV39H1 CBR3 and LRRTM2along with negative and positive control regions (34) Asshown in Supplementary Figure S5 the previouslyidentified five promoter elements which contain onlymeAP-1 but no consensus AP-1 sites were enriched atlevels similar to or exceeding those of the positivecontrols in K562 chromatin indicating that endogenous
Figure 2 Continuedresponsible for the binding of c-Junc-Fos to methylated DNA The meAP-1 and AP-1 binding sites contained in the oligonucleotide used in EMSAsare shown the asterisks represent a 50-methylcytosine residue EMSAs using BBLF4 and TREcollag oligonucleotides with the sites depicted in theleft panel and GFPc-Junc-Fos-purified protein are shown One representative experiment out of three is provided (D) Determination of theapparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion proteintransiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values The obtained data werefitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data point indicates themean and standard deviation of three independent experiments (E) Functional identification of single meAP-1 binding sites Pentamers of singlemeAP-1 sites present in the promoters of BBLF4 BALF1 and BFLF2 EBV genes as in (A) were introduced into a basic luciferase reporter plasmidwith a minimal EF1a promoter and free of CpG dinucleotides Unmethylated and fully CpG-methylated reporter constructs were analyzed in thepresence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Each experiment was performed three times and the means andstandard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3065
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

experiments In a second experimental setting serum-starved Wi-38 cells were infected with a GFPc-Junc-Fos-expressing adenovirus Chromatin from Wi-38(two days post infection) and Raji cells was preparedimmunoprecipitated with an a-GFP antibody to enrichDNA fragments specifically bound by GFPc-Junc-Fosprotein and analyzed with next-generation sequencing(Figure 3)
In accordance with our first functional analysis shownin Supplementary Figure S3B and discussed earlier in thetext we adjusted the bioinformatic parameters and refinedthe characteristics of the two AP-1 motifs found previ-ously consensus AP-1 motifs lacking a CpG dinucleotideand putative meAP-1 motifs with a single distal CpG pairin positions six and seven (Figure 3A and B) The twopredominant motifs revealed in both cellular modelsconfirm the known AP-1 consensus (50-TGACTCA-30)and identified an infrequent second meAP-1 motif(50-TGACGTCG-30) (Figure 3A and B) SupplementaryTables S4 through S7 provide the location of the AP-1 andmeAP-1 sites in both cell types
The majority of all consensus AP-1 sites are located inintergenic regions (3132) with no assigned functions Toconcentrate our further analysis on meAP-1 sites in prox-imity of promoter elements meAP-1 sites located within5 kb upstream of annotated TSS were considered only ifno other conventional AP-1 was located within a windowof plusmn1kb Only a small number of cellular genes fulfilledour criteria From ChIP-seq experiments of Raji cells weincluded the promoters of three genes in our study PTK6P2RY5 and SUV39H1 In Wi-38 cells we concentrated ontwo genes CBR3 and LRRTM2 A positive control thecollagenase promoter MMP1 containing two AP-1binding sites (33) was also included
Visualization of the coverage of these promoters afterChIP-seq analysis indicated that they are differentiallybound by c-Junc-Fos in the two cell types (Figure 3C)suggesting that the extent of cytosine methylation mightalso differ Bisulfite sequencing of the five promotersrevealed only one promoter element P2RY5 that wasfully CpG-methylated in both cell lines but CpG pairsin other promoters with meAP-1 sites differed in a celltype-specific manner (Supplementary Figure S4)
50-methylcytosine-dependent binding of c-Junc-Fos tomeAP-1 sites in the human genome inducestranscriptional activation
Only 5 of meAP-1 sites identified in both cell types arelocated in putative promoter regions and therefore
presumably involved in directly regulating gene expres-sion Three candidate meAP-1 motifs with no AP-1 con-sensus binding sites in their close proximity were analyzedfurther the meAP-1 sites in the PTK6 P2RY5 andSUV39H1 promoters (Figure 4) As shown in Figure 4Bc-Junc-Fos bound almost exclusively to the CpG-methylated meAP-1 motif In ELISAs the apparent dis-sociation constants (KD) of PTK6 and P2RY5 meAP-1sites could only be determined with methylated oligo-nucleotides because KD values of the unmethylatedprobes were too high to be estimated (Figure 4C) TheKD for the SUV39H1 meAP-1 site could not becalculated but the results confirm that the bindingaffinity is higher when the meAP-1 site is methylated(Figure 4B and C) These results indicate that thepresence of 50-methylcytosine favors the binding ofc-Junc-Fos to the meAP-1 sequences identified in thehuman genomeThe ability of c-Junc-Fos to transactivate gene tran-
scription through the binding to the methylated meAP-1sites from the human genome was tested in reporterassays Reporter plasmids containing pentamers ofselected meAP-1 sites in the human promoters werecotransfected with an expression plasmid encoding thetransactivator VP16c-Junc-Fos or a negative controlDNA Luciferase activities with CpG-methylatedreporter plasmids were significantly higher as comparedwith unmethylated reporter plasmid DNAs (20ndash27-foldFigure 4D)
Promoters with methylated meAP-1 sites are bound byc-Junc-Fos in vivo
So far our studies were based on the ectopic expression ofartificial c-Junc-Fos dimers to identify potential meAP-1sites in cellular DNA In contrast Raha et al (31) mappedthe location of endogenous c-Jun and c-Fos protein in twosets of ChIP-seq experiments and identified gt40 000binding sites in the established human myeloblastic cellline K562 To correlate their findings and ours we per-formed ChIP assays in this cell line with a c-Jun specificantibody and investigated by quantitative PCR analysisthe enrichment of the five previously identified promoterelements PTK6 P2RY5 SUV39H1 CBR3 and LRRTM2along with negative and positive control regions (34) Asshown in Supplementary Figure S5 the previouslyidentified five promoter elements which contain onlymeAP-1 but no consensus AP-1 sites were enriched atlevels similar to or exceeding those of the positivecontrols in K562 chromatin indicating that endogenous
Figure 2 Continuedresponsible for the binding of c-Junc-Fos to methylated DNA The meAP-1 and AP-1 binding sites contained in the oligonucleotide used in EMSAsare shown the asterisks represent a 50-methylcytosine residue EMSAs using BBLF4 and TREcollag oligonucleotides with the sites depicted in theleft panel and GFPc-Junc-Fos-purified protein are shown One representative experiment out of three is provided (D) Determination of theapparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion proteintransiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values The obtained data werefitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data point indicates themean and standard deviation of three independent experiments (E) Functional identification of single meAP-1 binding sites Pentamers of singlemeAP-1 sites present in the promoters of BBLF4 BALF1 and BFLF2 EBV genes as in (A) were introduced into a basic luciferase reporter plasmidwith a minimal EF1a promoter and free of CpG dinucleotides Unmethylated and fully CpG-methylated reporter constructs were analyzed in thepresence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Each experiment was performed three times and the means andstandard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3065
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5
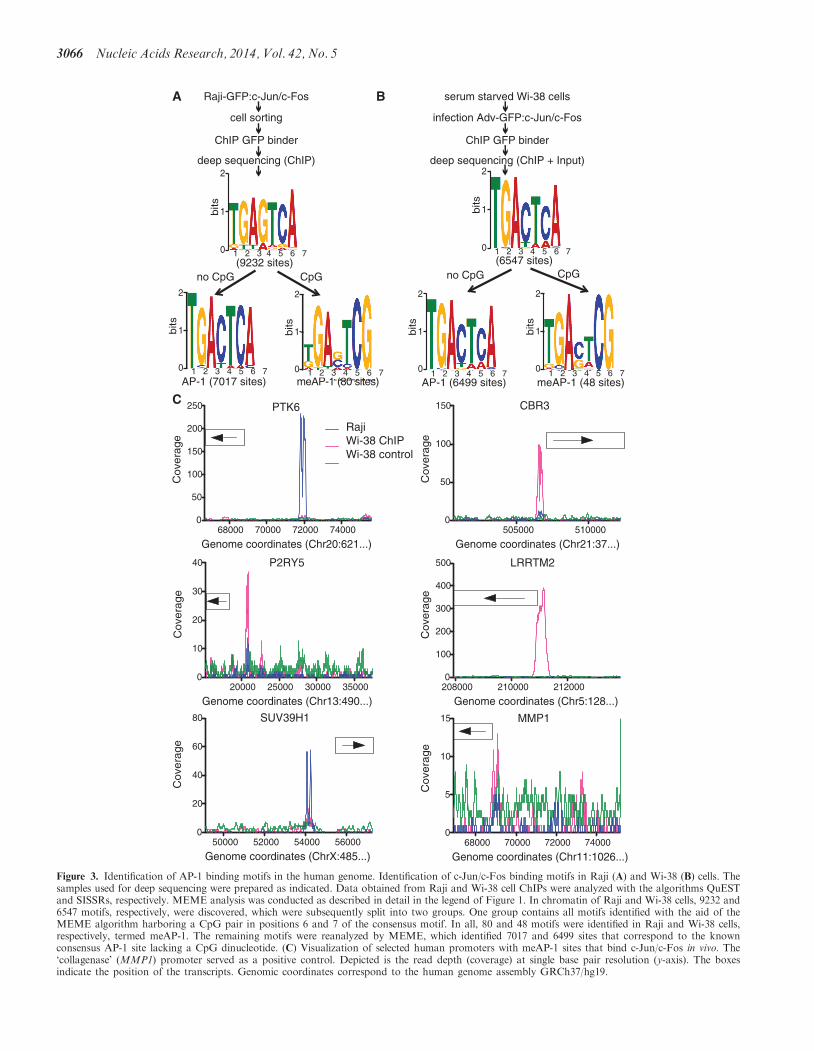
SUV39H1
50000 52000 54000 560000
20
40
60
80
Genome coordinates (ChrX485)
Cov
erag
e
MMP1
68000 70000 72000 740000
5
10
15
Genome coordinates (Chr111026)
Cov
erag
e
A B
C
serum starved Wi-38 cells
infection Adv-GFPc-Junc-Fos
ChIP GFP binder
deep sequencing (ChIP + Input)
AP-1 (6499 sites) meAP-1 (48 sites)
no CpG CpG
Raji-GFPc-Junc-Fos
cell sorting
ChIP GFP binder
deep sequencing (ChIP)
AP-1 (7017 sites) meAP-1 (80 sites)
no CpG CpG(9232 sites) (6547 sites)
PTK6
68000 70000 72000 740000
50
100
150
200
250
Genome coordinates (Chr20621)
Cov
erag
e
LRRTM2
208000 210000 2120000
100
200
300
400
500
Genome coordinates (Chr5128)
Cov
erag
e
P2RY5
20000 25000 30000 350000
10
20
30
40
Genome coordinates (Chr13490)
RajiWi-38 ChIPWi-38 control
Cov
erag
e
CBR3
505000 5100000
50
100
150
Genome coordinates (Chr2137)
Cov
erag
e
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
1 2 3 4 5 6 7
2
1
0
bits
Figure 3 Identification of AP-1 binding motifs in the human genome Identification of c-Junc-Fos binding motifs in Raji (A) and Wi-38 (B) cells Thesamples used for deep sequencing were prepared as indicated Data obtained from Raji and Wi-38 cell ChIPs were analyzed with the algorithms QuESTand SISSRs respectively MEME analysis was conducted as described in detail in the legend of Figure 1 In chromatin of Raji and Wi-38 cells 9232 and6547 motifs respectively were discovered which were subsequently split into two groups One group contains all motifs identified with the aid of theMEME algorithm harboring a CpG pair in positions 6 and 7 of the consensus motif In all 80 and 48 motifs were identified in Raji and Wi-38 cellsrespectively termed meAP-1 The remaining motifs were reanalyzed by MEME which identified 7017 and 6499 sites that correspond to the knownconsensus AP-1 site lacking a CpG dinucleotide (C) Visualization of selected human promoters with meAP-1 sites that bind c-Junc-Fos in vivo Thelsquocollagenasersquo (MMP1) promoter served as a positive control Depicted is the read depth (coverage) at single base pair resolution (y-axis) The boxesindicate the position of the transcripts Genomic coordinates correspond to the human genome assembly GRCh37hg19
3066 Nucleic Acids Research 2014 Vol 42 No 5
A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

A B
D
gcagtgTGACTCGtgcaga
gctgtcTGACTCGatggc
ggtagaTGACTCGtgttga
PTK6
SUV39H1
P2RY5
nd nd nd
171nM 193nM nd
unmethylated
methylated
1 10 100 10000
100
200
300
400
500SUB39H1meSUV39H1
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800
1000 P2RY5meP2RY5
nM oligo
Cy5
inte
nsity
1 10 100 10000
200
400
600
800 PTK6
mePTK6
nM oligo
Cy5
inte
nsity
C
prot - + - +
unme me
- + - +
SUV39H1 PTK6 P2RY5
unme me unme me
- + - +
5xSUV39H1 5xPTK6 5xP2RY5
Figure 4 Functional analysis of human meAP-1 sites (A) Oligonucleotides containing meAP-1 binding sites of the PTK6 P2RY5 and SUV39H1promoters The meAP-1 sites are indicated in bold capital letters and the CpG pairs within the meAP-1 sites are underlined (B) EMSAs showing thepreferential binding of c-Junc-Fos to the CpG-methylated meAP-1 sites of the PTK6 P2RY5 and SUV39H1 promoters EMSAs were performedwith affinity-purified GFP-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells EMSAs of typical experiments are shown(C) Determination of the apparent KD of c-Junc-Fos bound to meAP-1 ELISAs were performed with affinity-purified StrepFLAG-tagged c-Junc-Fos fusion protein transiently expressed in HEK293 cells Serial dilutions of Cy5 oligonucleotides were used to determine the KD values Thedata were fitted to the Hill equation with one site-specific binding to determine the dissociation constants which are indicated Each data pointindicates the mean and standard deviation of three independent experiments (D) The c-Junc-Fos activation of promoters with meAP-1 binding sitesis increased on CpG methylation Pentamers of short oligonucleotides shown in (A) which contain the meAP-1 sites present in the promoters of thehuman PTK6 P2RY5 and SUV39H1 genes were introduced into a basic luciferase reporter plasmid as described in Figure 2E Unmethylated andfully CpG-methylated reporter constructs were analyzed in the presence or absence of a cotransfected VP16c-Junc-Fos expression plasmid Eachexperiment was performed three times and means and standard deviations are depicted
Nucleic Acids Research 2014 Vol 42 No 5 3067
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5
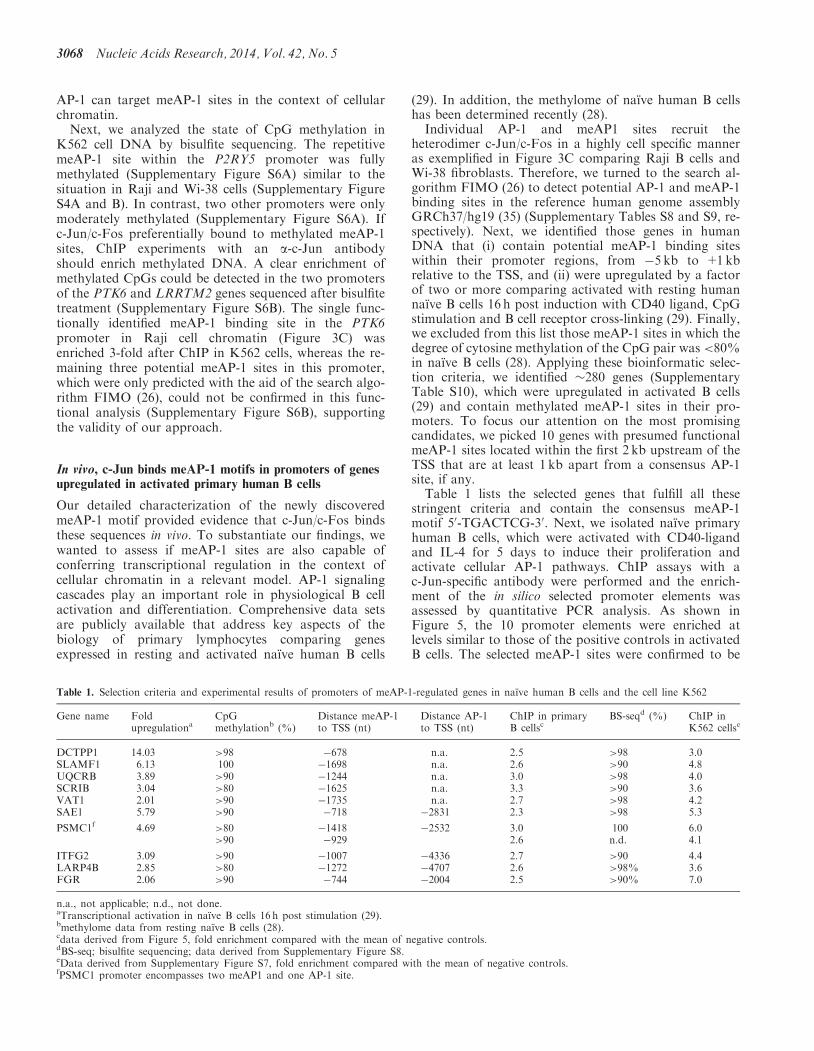
AP-1 can target meAP-1 sites in the context of cellularchromatinNext we analyzed the state of CpG methylation in
K562 cell DNA by bisulfite sequencing The repetitivemeAP-1 site within the P2RY5 promoter was fullymethylated (Supplementary Figure S6A) similar to thesituation in Raji and Wi-38 cells (Supplementary FigureS4A and B) In contrast two other promoters were onlymoderately methylated (Supplementary Figure S6A) Ifc-Junc-Fos preferentially bound to methylated meAP-1sites ChIP experiments with an a-c-Jun antibodyshould enrich methylated DNA A clear enrichment ofmethylated CpGs could be detected in the two promotersof the PTK6 and LRRTM2 genes sequenced after bisulfitetreatment (Supplementary Figure S6B) The single func-tionally identified meAP-1 binding site in the PTK6promoter in Raji cell chromatin (Figure 3C) wasenriched 3-fold after ChIP in K562 cells whereas the re-maining three potential meAP-1 sites in this promoterwhich were only predicted with the aid of the search algo-rithm FIMO (26) could not be confirmed in this func-tional analysis (Supplementary Figure S6B) supportingthe validity of our approach
In vivo c-Jun binds meAP-1 motifs in promoters of genesupregulated in activated primary human B cells
Our detailed characterization of the newly discoveredmeAP-1 motif provided evidence that c-Junc-Fos bindsthese sequences in vivo To substantiate our findings wewanted to assess if meAP-1 sites are also capable ofconferring transcriptional regulation in the context ofcellular chromatin in a relevant model AP-1 signalingcascades play an important role in physiological B cellactivation and differentiation Comprehensive data setsare publicly available that address key aspects of thebiology of primary lymphocytes comparing genesexpressed in resting and activated naıve human B cells
(29) In addition the methylome of naıve human B cellshas been determined recently (28)
Individual AP-1 and meAP1 sites recruit theheterodimer c-Junc-Fos in a highly cell specific manneras exemplified in Figure 3C comparing Raji B cells andWi-38 fibroblasts Therefore we turned to the search al-gorithm FIMO (26) to detect potential AP-1 and meAP-1binding sites in the reference human genome assemblyGRCh37hg19 (35) (Supplementary Tables S8 and S9 re-spectively) Next we identified those genes in humanDNA that (i) contain potential meAP-1 binding siteswithin their promoter regions from 5 kb to +1kbrelative to the TSS and (ii) were upregulated by a factorof two or more comparing activated with resting humannaıve B cells 16 h post induction with CD40 ligand CpGstimulation and B cell receptor cross-linking (29) Finallywe excluded from this list those meAP-1 sites in which thedegree of cytosine methylation of the CpG pair was lt80in naıve B cells (28) Applying these bioinformatic selec-tion criteria we identified 280 genes (SupplementaryTable S10) which were upregulated in activated B cells(29) and contain methylated meAP-1 sites in their pro-moters To focus our attention on the most promisingcandidates we picked 10 genes with presumed functionalmeAP-1 sites located within the first 2 kb upstream of theTSS that are at least 1 kb apart from a consensus AP-1site if any
Table 1 lists the selected genes that fulfill all thesestringent criteria and contain the consensus meAP-1motif 50-TGACTCG-30 Next we isolated naıve primaryhuman B cells which were activated with CD40-ligandand IL-4 for 5 days to induce their proliferation andactivate cellular AP-1 pathways ChIP assays with ac-Jun-specific antibody were performed and the enrich-ment of the in silico selected promoter elements wasassessed by quantitative PCR analysis As shown inFigure 5 the 10 promoter elements were enriched atlevels similar to those of the positive controls in activatedB cells The selected meAP-1 sites were confirmed to be
Table 1 Selection criteria and experimental results of promoters of meAP-1-regulated genes in naıve human B cells and the cell line K562
Gene name Foldupregulationa
CpGmethylationb ()
Distance meAP-1to TSS (nt)
Distance AP-1to TSS (nt)
ChIP in primaryB cellsc
BS-seqd () ChIP inK562 cellse
DCTPP1 1403 gt98 678 na 25 gt98 30SLAMF1 613 100 1698 na 26 gt90 48UQCRB 389 gt90 1244 na 30 gt98 40SCRIB 304 gt80 1625 na 33 gt90 36VAT1 201 gt90 1735 na 27 gt98 42SAE1 579 gt90 718 2831 23 gt98 53
PSMC1f 469 gt80 1418 2532 30 100 60gt90 929 26 nd 41
ITFG2 309 gt90 1007 4336 27 gt90 44LARP4B 285 gt80 1272 4707 26 gt98 36FGR 206 gt90 744 2004 25 gt90 70
na not applicable nd not doneaTranscriptional activation in naıve B cells 16 h post stimulation (29)bmethylome data from resting naıve B cells (28)cdata derived from Figure 5 fold enrichment compared with the mean of negative controlsdBS-seq bisulfite sequencing data derived from Supplementary Figure S8eData derived from Supplementary Figure S7 fold enrichment compared with the mean of negative controlsfPSMC1 promoter encompasses two meAP1 and one AP-1 site
3068 Nucleic Acids Research 2014 Vol 42 No 5
methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

methylated by analyzing the state of CpG methylationin genomic DNA of non-activated B cells by bisulfitesequencing (Table 1 and Supplementary Figure S8)Supporting our finding in primary B cells the 10promoter regions were also bound by c-Jun in the K562cell line (Supplementary Figure S7)
Table 1 summarizes the original data that led to the insilico identification of the 10 genes and the experimentalresults obtained in this work Our data indicate that thepredicted meAP-1 binding sites are CpG-methylated andtargeted by c-Jun in activated B cells in vivo suggestingthat individual meAP-1 sites contribute to transcriptionalactivation of highly CpG-methylated hence epigeneticallyrepressed genes in naıve B lymphocytes on their activa-tion One caveat remains that cannot be addressed experi-mentally mitogenic physiological signals in B cellproliferation and development-like B cell receptor cross-linking CD40 engagement and interleukin receptorsignals induce the MAPK cascades leading to AP-1 acti-vation but also engage additional signaling cascades thatmight also act on meAP-1-regulated genes assisting theirtranscriptional reactivation (36)
DISCUSSION
The aforementioned results confirm our starting hypoth-esis and indicate that AP-1 has the capacity to bind a classof previously unknown discrete DNA sequence motifs
with methylated cytosine residues in vivo It thus appearsthat Zta is not only a closely related member of the largeAP-1 family because it shares a highly conserved DNAbinding domain with c-Fos (6) but also may haveevolved from a predecessor of the AP-1 protein familywhich can specifically make contacts with the 50-methylgroup of cytosine nucleotides We propose that additionalmembers of the vast AP-1 family can also bind tomethylated DNA in a sequence-specific mannerA close inspection of ZREmeZRE and AP-1meAP-1
pairs (Figure 6) suggests that they are related but haveevolved differently (i) In general nucleotide transitions(but not transversions) cause the differences within theZREmeZRE and AP-1meAP-1 pairs (Figure 6A) reflect-ing only minor differences in the molecular structures ofthymine and 50-methylcytosine (Figure 6B) (ii) Theposition of the methylated cytosine in the consensus se-quences of meZRE and meAP-1 is offset by one nucleotide(Figure 6A) (iii) The consensus AP-1 motif is almost pal-indromic but the meAP-1 motif is not suggesting thateither c-Jun or c-Fos preferentially make contact withthe 50-methylcytosine group This view is supported bytwo detailed reports indicating that c-Fos and c-Jun areunequal siblings (3738) Interestingly Risse et al (38)delineated the importance of individual thymine residuesin the AP-1 consensus sequence and found that allthymines including the two distal positions crucially con-tribute to DNAndashprotein interactionsZta is the first example of a sequence-specific transcrip-
tion factor which preferentially recognizes and selectivelybinds DNA sequence motifs with methylated CpGresidues and activates gene transcription through thesedistinct motifs (45) Previously other mammalianfactors (RXF1 RBP-J ZBTB4) have been shown tobind DNA sequences encompassing 50-methylcytosine nu-cleotides in vitro only and no evidence for methylation-dependent binding of these factors exists in cells in vivo(39ndash42) Retrospectively this failure is not surprisingbecause consensus meAP-1 motifs are rarely targeted byc-Junc-Fos in cellular chromatin Accessible and there-fore functional meAP-1 sites account for lt2 of the45 000 c-Junc-Fos binding sites that are predicted inhuman genomic DNA (Figure 3A and B SupplementaryTable S9) This is in contrast to the high number of con-sensus AP-1 sites predicted in silico (32) which are readilybound by factors of the AP-1 family in human chromatin(3134)Interestingly a recent systematic in vitro analysis of a
large panel of transcription factors identified theirfrequent binding to sequence motifs with methylatedCpG dinucleotides (43) The search did not identify thenon-canonical meAP-1 motif reported here Rather itrevealed distinct and therefore unrelated motifs eitherfree of CpG dinucleotides or with methylated CpGstargeted by the same mammalian transcription factorThis observation is surprising and clearly in contrastto our findings which led to the discovery of themeAP-1 motif a close relative of the canonical AP-1 site(Figure 6)Our results indicate that the 50-methylcytosine residue
can functionally substitute for the crucial distal thymine in
PA
I 1
BC
L3
MM
P1
gusB
GA
PD
H
tubu
lin
DC
TP
P1
SLA
MF
1
UQ
CR
B
SC
RIB
VA
T1
SA
E1
PS
MC
1-1
PS
MC
1-2
ITF
G2
LAR
P4B
FG
R
00
05
10
15
20
25
positive negative
controlsmeAP-1
in
put
Figure 5 In vivo binding of c-Junc-Fos to cellular promoters withmeAP-1 sites in activated primary human B cells After immunopre-cipitation (ChIP) of chromatin from activated naıve B cells with an a-c-Jun antibody the enrichment of the indicated promoter regions wasassayed by quantitative PCR Shown are the results obtained fromthree promoter regions indicated in red that contain conventional pre-viously identified AP-1 sites (34) and three promoter regions indicatedin black which were identified in this study and are devoid of conven-tional AP-1 or meAP-1 sites Both sets of promoters served as positiveand negative controls respectively Ten promoters that were found tocontain meAP-1 sites (and lack conventional AP-1 sites in proximity)were analyzed by ChIP and the results are shown in green The PSMC1promoter contains two separate regions with one meAP-1 site eachdepicted as PSMC1-1 and -2 which were analyzed separately Detailsof all promoters investigated here are listed in Table 1 Means andstandard deviations of three independent experiments are shown
Nucleic Acids Research 2014 Vol 42 No 5 3069
one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

one strand of the AP-1 consensus motif providing similarmolecular structures for AP-1 binding (Figures 2C and 6)The structural resemblance of thymine to50-methylcytosine suggests that the c-Junc-Fosheterodimer binds similarly to AP-1 and meAP-1 sites(3944) As c-Junc-Fos heterodimers target meAP-1 sitesin vivo (Figures 3 and 5 Supplementary Figures S5 andS7) the status of DNA methylation determines AP-1binding and consequently responsiveness to AP-1-mediated transcriptional activation In this contextDNA methylation adds a new layer to gene regulationand presumably also cell type- and tissue-specific function-ality of signaling pathways that activate AP-1 familymembersThe majority of consensus AP-1 sites that were
identified in cellular chromatin are located in intergenicregions or are distant from transcriptional start sites andhave unknown functions (3134) We see a comparabledistribution of meAP-1 sites Of the 45 000 potentialmeAP-1 sites (Supplementary Table S9) only a smallfraction resides within 5 kb upstream of transcriptionalstart sites suggesting that these meAP-1 sites might servean important and direct role in regulating genes that aregoverned by epigenetic inhibition including DNA methy-lation The functions of the many consensus AP-1 andnon-canonical meAP-1 site localized in intergenic regionsremain an enigma that needs to be addressedOwing to the nature and frequency of potentially func-
tional meAP-1 binding sites in the cell lines and conditionsused in this study we had to restrict our search in estab-lished cell lines to promoters that are bound by c-Junc-Fos heterodimers and contain meAP-1 sites but lackconsensus AP-1 sites in proximity for obvious technicalreasons The strict filtering criteria resulted in a small setof promoter candidates that likely do not reflect profoundor even decisive functions of the identified genes in human
cells The situation is different in primary naıve B cells ofhuman origin in which we concentrated on meAP-1 in thepromoters of genes upregulated after induced B cell acti-vation The fact that AP-1 is binding these meAP-1binding sites suggests their having functional roles inmetazoan gene regulation in certain cell types andcertain situations eg during the transition from quies-cent resting B lymphocytes to proliferating B blasts Inthis context c-Junc-Fos differs from Zta which fre-quently binds to many viral meZRE-containing promotersand efficiently reverses their epigenetic repression(571445) These differences could be explained by thefact that the key functions of Zta in the EBV model areunderstood whereas the c-Junc-Fos transcription factormight have yet to be uncovered functional roles in differ-ent settings Although the functionality of the meAP-1binding sites cannot be fully uncovered with the experi-ments presented here the experiments document thein vivo existence of these elements which are bound pref-erentially if their DNA motif carries a 50-methylcytosineIt is likely that these elements play a critical role in thetranscriptional activation of their target genes Thesefindings now allow the investigation of other experimentalsettings such as different cell lines or primary cell types aswell as distinct stimuli stress signals or developmentalcues that activate cells induce their immediate differenti-ation or trigger cell fate decisions The goal is now todiscover more examples of meAP-1 sites that regulatethe expression of critical genes to unveil the full range ofmeAP-1 elements and their biological functions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Christine Gobel for technical advicewith the luciferase assays Dagmar Pich for her supportwith cell sorting and Vigo Heissmeyer for his adenoviralvector constructs advice and helpful discussions Theyalso thank Reiner Siebert (Institut fur HumangenetikUniversitatsklinikum Schleswig-Holstein Christian-Albrechts-Universitat zu Kiel) and Steve Hoffmann(Transcriptome Bioinformatics LIFE ResearchComplex University Leipzig) for the valuable access tothe methylome data of primary naıve B lymphocytes ofthree human donors and helpful suggestions anddiscussions
FUNDING
Alexander von Humboldt Foundation [Humboldt-Forschungsstipendium fur Postdoktoranden to MG]intramural grants the Deutsche Forschungsgemeinschaft[SPP1230 SFB 1054 SFB 1064 SFB-TR36] NationalInstitutes of Health [CA70723] Funding for open accesscharge Helmholtz Zentrum Munchen
Conflict of interest statement None declared
TGACTCAACTGAGT
TGAG AACTC T
TGACTCGACTGAGC
TGAGCGAACTCGCT
A
B
AP-1 meAP-1
ZRE meZRE
CATCGTAG
thymine 5rsquo-methylcytosine cytosine
12
34
5
61
2
34
5
61
2
34
5
6
CH3 CH3
Figure 6 Sequence motifs of AP-1 and ZRE binding sites and theirmolecular features (A) Comparison of AP-1 ZRE meAP-1 andmeZRE binding sites The CpG pairs in the meAP-1 and meZREmotifs are underlined (B) Structure of the DNA bases cytosine50-methylcytosine and thymine The dashed lines indicate thehydrogen bonds with the bases of the complementary DNA strand
3070 Nucleic Acids Research 2014 Vol 42 No 5
REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

REFERENCES
1 EferlR and WagnerEF (2003) AP-1 a double-edged sword intumorigenesis Nat Rev Cancer 3 859ndash868
2 HessJ AngelP and Schorpp-KistnerM (2004) AP-1 subunitsquarrel and harmony among siblings J Cell Sci 1175965ndash5973
3 ProkhortchoukE and DefossezPA (2008) The cell biology ofDNA methylation in mammals Biochim Biophys Acta 17832167ndash2173
4 BhendePM SeamanWT DelecluseHJ and KenneySC(2004) The EBV lytic switch protein Z preferentially binds toand activates the methylated viral genome Nat Genet 361099ndash1104
5 BergbauerM KallaM SchmeinckA GobelC RothbauerUEckS Benet-PagesA StromTM and HammerschmidtW(2010) CpG-methylation regulates a class of Epstein-Barr viruspromoters PLoS Pathog 6 e1001114
6 FarrellPJ RoweDT RooneyCM and KouzaridesT (1989)Epstein-Barr virus BZLF1 trans-activator specifically bindsto a consensus AP-1 site and is related to c-fos EMBO J 8127ndash132
7 WoellmerA Arteaga-SalasJM and HammerschmidtW (2012)BZLF1 governs CpG-methylated chromatin of Epstein-Barr virusreversing epigenetic repression PLoS Pathog 8 e1002902
8 HollerM WestinG JiricnyJ and SchaffnerW (1988) Sp1transcription factor binds DNA and activates transcription evenwhen the binding site is CpG methylated Genes Dev 21127ndash1135
9 HantuschB KaltR KriegerS PuriC and KerjaschkiD(2007) Sp1Sp3 and DNA-methylation contribute to basaltranscriptional activation of human podoplanin in MG63 versusSaos-2 osteoblastic cells BMC Mol Biol 8 20
10 RishiV BhattacharyaP ChatterjeeR RozenbergJ ZhaoJGlassK FitzgeraldP and VinsonC (2010) CpG methylation ofhalf-CRE sequences creates CEBPalpha binding sites thatactivate some tissue-specific genes Proc Natl Acad Sci USA107 20311ndash20316
11 Iguchi-ArigaSM and SchaffnerW (1989) CpG methylation ofthe cAMP-responsive enhancerpromoter sequence TGACGTCAabolishes specific factor binding as well as transcriptionalactivation Genes Dev 3 612ndash619
12 HolzerlandtR OrengoC KellamP and AlbaMM (2002)Identification of new herpesvirus gene homologs in the humangenome Genome Res 12 1739ndash1748
13 SinclairAJ (2003) bZIP proteins of human gammaherpesvirusesJ Gen Virol 84 1941ndash1949
14 KallaM SchmeinckA BergbauerM PichD andHammerschmidtW (2010) AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigeneticstate of the viral genome Proc Natl Acad Sci USA 107850ndash855
15 WiesnerM ZentzC MayrC WimmerR HammerschmidtWZeidlerR and MoosmannA (2008) Conditional immortalizationof human B cells by CD40 ligation PLoS One 3 e1464
16 DelecluseHJ HilsendegenT PichD ZeidlerR andHammerschmidtW (1998) Propagation and recovery of intactinfectious Epstein-Barr virus from prokaryotic to human cellsProc Natl Acad Sci USA 95 8245ndash8250
17 BakiriL MatsuoK WisniewskaM WagnerEF and YanivM(2002) Promoter specificity and biological activity of tethered AP-1 dimers Mol Cell Biol 22 4952ndash4964
18 GloecknerCJ BoldtK SchumacherA RoepmanR andUeffingM (2007) A novel tandem affinity purification strategyfor the efficient isolation and characterisation of native proteincomplexes Proteomics 7 4228ndash4234
19 KlugM and RehliM (2006) Functional analysis of promoterCpG methylation using a CpG-free luciferase reporter vectorEpigenetics 1 127ndash130
20 KallaM GobelC and HammerschmidtW (2012) Thelytic phase of Epstein-Barr virus requires a viral genomewith 5-methylcytosine residues in CpG sites J Virol 86447ndash458
21 LiH RuanJ and DurbinR (2008) Mapping short DNAsequencing reads and calling variants using mapping qualityscores Genome Res 18 1851ndash1858
22 JothiR CuddapahS BarskiA CuiK and ZhaoK (2008)Genome-wide identification of in vivo protein-DNA binding sitesfrom ChIP-Seq data Nucleic Acids Res 36 5221ndash5231
23 ValouevA JohnsonDS SundquistA MedinaC AntonEBatzoglouS MyersRM and SidowA (2008) Genome-wideanalysis of transcription factor binding sites based on ChIP-Seqdata Nat Methods 5 829ndash834
24 BaileyTL WilliamsN MislehC and LiWW (2006) MEMEdiscovering and analyzing DNA and protein sequence motifsNucleic Acids Res 34 W369ndashW373
25 SetoE MoosmannA GrommingerS WalzN GrundhoffAand HammerschmidtW (2010) Micro RNAs of Epstein-Barrvirus promote cell cycle progression and prevent apoptosis ofprimary human B cells PLoS Pathog 6 e1001063
26 GrantCE BaileyTL and NobleWS (2011) FIMOscanning for occurrences of a given motif Bioinformatics 271017ndash1018
27 MeyerLR ZweigAS HinrichsAS KarolchikD KuhnRMWongM SloanCA RosenbloomKR RoeG RheadB et al(2013) The UCSC Genome Browser database extensions andupdates 2013 Nucleic Acids Res 41 D64ndashD69
28 KulisM HeathS BibikovaM QueirosAC NavarroAClotG Martinez-TrillosA CastellanoG Brun-HeathIPinyolM et al (2012) Epigenomic analysis detects widespreadgene-body DNA hypomethylation in chronic lymphocyticleukemia Nat Genet 44 1236ndash1242
29 Le GallouS CaronG DelaloyC RossilleD TarteK andFestT (2012) IL-2 requirement for human plasma cellgeneration coupling differentiation and proliferation byenhancing MAPK-ERK signaling J Immunol 189 161ndash173
30 ChytilM PetersonBR ErlansonDA and VerdineGL (1998)The orientation of the AP-1 heterodimer on DNA strongly affectstranscriptional potency Proc Natl Acad Sci USA 9514076ndash14081
31 RahaD WangZ MoqtaderiZ WuL ZhongG GersteinMStruhlK and SnyderM (2010) Close association of RNApolymerase II and many transcription factors with Pol III genesProc Natl Acad Sci USA 107 3639ndash3644
32 ZhouH ZarubinT JiZ MinZ ZhuW DowneyJS LinSand HanJ (2005) Frequency and distribution of AP-1 sites in thehuman genome DNA Res 12 139ndash150
33 BenbowU and BrinckerhoffCE (1997) The AP-1 site andMMP gene regulation what is all the fuss about Matrix Biol15 519ndash526
34 LiM GeQ WangW WangJ and LuZ (2011) c-Junbinding site identification in K562 cells J Genet Genomics 38235ndash242
35 ChurchDM SchneiderVA GravesT AugerKCunninghamF BoukN ChenHC AgarwalaRMcLarenWM RitchieGR et al (2011) Modernizing referencegenome assemblies PLoS Biol 9 e1001091
36 RichardsS WatanabeC SantosL CraxtonA and ClarkEA(2008) Regulation of B-cell entry into the cell cycle ImmunolRev 224 183ndash200
37 LeonardDA and KerppolaTK (1998) DNA bendingdetermines Fos-Jun heterodimer orientation Nat Struct Biol 5877ndash881
38 RisseG JoossK NeubergM BrullerHJ and MullerR (1989)Asymmetrical recognition of the palindromic AP1 binding site(TRE) by Fos protein complexes EMBO J 8 3825ndash3832
39 BartelsSJ SpruijtCG BrinkmanAB JansenPWVermeulenM and StunnenbergHG (2011) A SILAC-basedscreen for methyl-CpG binding proteins identifies RBP-J as aDNA methylation and sequence-specific binding protein PLoSOne 6 e25884
40 NiesenMI OsborneAR YangH RastogiS ChellappanSChengJQ BossJM and BlanckG (2005) Activation of amethylated promoter mediated by a sequence-specific DNA-binding protein RFX J Biol Chem 280 38914ndash38922
41 ZhangXY Jabrane-FerratN AsieduCK SamacSPeterlinBM and EhrlichM (1993) The major histocompatibility
Nucleic Acids Research 2014 Vol 42 No 5 3071
complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5

complex class II promoter-binding protein RFX (NF-X)is a methylated DNA-binding protein Mol Cell Biol 136810ndash6818
42 SasaiN NakaoM and DefossezPA (2010) Sequence-specificrecognition of methylated DNA by human zinc-finger proteinsNucleic Acids Res 38 5015ndash5022
43 HuS WanJ SuY SongQ ZengY NguyenHN ShinJCoxE RhoHS WoodardC et al (2013) DNA methylationpresents distinct binding sites for human transcription factorsElife 2 e00726
44 GloverJN and HarrisonSC (1995) Crystal structure of theheterodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA Nature 373 257ndash261
45 KarlssonQH SchelcherC VerrallE PetosaC andSinclairAJ (2008) Methylated DNA recognition during thereversal of epigenetic silencing is regulated by cysteine and serineresidues in the Epstein-Barr virus lytic switch protein PLoSPathog 4 e1000005
3072 Nucleic Acids Research 2014 Vol 42 No 5