bvoc emission from populus canadensis saplings in response to acute uv-a radiation
TRANSCRIPT

Physiologia Plantarum 148: 51–61. 2013 Copyright © Physiologia Plantarum 2012, ISSN 0031-9317
BVOC emission from Populus × canadensis saplingsin response to acute UV-A radiationEmanuele Pallozzia,b, Alessio Fortunatia, Giovanni Marinoa, Francesco Loretoc, Giovanni Agatid andMauro Centrittoa,∗
aInstitute of AgroEnvironmental and Forest Biology, National Research Council, 00015, Monterotondo Scalo, RM, ItalybDepartment of Forest Environment and Resources, University of Tuscia, 01100, Viterbo, ItalycInstitute for Plant Protection, National Research Council, 50019, Sesto Fiorentino, FI, ItalydInstitute of Applied Physics ’N. Carrara’, National Research Council, 50019, Sesto Fiorentino, FI, Italy
Correspondence*Corresponding author,e-mail: [email protected]
Received 27 March 2012;revised 16 June 2012
doi:10.1111/j.1399-3054.2012.01687.x
Hybrid poplar (Populus × canadensis) saplings were subjected to acuteultraviolet-A (UV-A) irradiation (30 W m–2, ambient treatment, 60, 90 and120 W m–2 of UV-A irradiance) to determine the effects on photosynthesisand biogenic volatile organic compound (BVOC) emissions in two differentshort-term experiments (i.e. sequential increase in UV-A irradiance andUV-A intensity–response relationships). Both intensity–response experimentsshowed that the UV-A ambient treatment did not affect photosynthesis andBVOC emissions. Whereas exposition at 60, 90 and 120 W m–2 of UV-A (firstexperiment), increasingly inhibited photosynthesis. This increasing inhibitionwas also detected by decreasing trends of both photochemical reflectanceindex (PRI) and fluorescence yield. Isoprene emission resulted to be verysensitive to increasing UV-A irradiances. Methanol was also very sensitive tohigh UV-A radiation, suggesting the occurrence of strong damages of cellularstructures. The second experiment, which was performed both in the middleof July and repeated towards the end of the summer, showed a temporalvariations in the UV-A intensity–response relationships. In fact, there were nolonger significant differences in photosynthesis, PRI and isoprene emission inresponse to high UV-A radiation toward the end of the summer season. Theadaxial flavonoid level increased significantly over the period monitored,resulting 85% higher toward the end of the summer than during the middleof the summer. This dramatic increase in the adaxial flavonoids may haveplayed a protective role against UV-A radiation by shielding leaves. Ourfindings add to the understanding of physiological processes involved inplant response to UV radiation.
Introduction
The amount of ultraviolet (UV) radiation reaching theEarth’s surface is steadily increasing over the tropicsand southern high latitudes during spring and summer(Hegglin and Shepherd 2009). Increased UV is likely
Abbreviations – BVOC, biogenic volatile organic compound; Chl, chlorophyll; FLAV, flavonoid index; LEDs, light-emittingdiodes; PPFD, photosynthetic photon flux density; PRI, photochemical reflectance index; PTR-MS, proton transfer reactionmass spectrometer; RGBA, red, green, blue, amber; UV, ultraviolet.
to have both direct and indirect implications on photo-synthesis (Paul and Gwynn-Jones 2003) and, in turn, onbiogenic volatile organic compounds (BVOCs). Theseprimary and secondary metabolisms are now acceptedas important components of the biosphere’s response toclimate change (Centritto et al. 2011b) and consequently
Physiol. Plant. 148, 2013 51

there is great interest in determining their sensitivity todifferent radiation quality. This knowledge is particularlyrelevant for the biosynthesis of BVOCs, which, besideshaving an intriguing ecological role in emitting plants(Vickers et al. 2009), is very reactive and influences boththe atmospheric chemistry composition and physics.
Studies on the impact of either acute or chronicUV radiation on BVOC emission even on a single-species scale are few and show contrasting results.The first published study was performed by Harleyet al. (1996) on isoprene-emitting and non-emitting plantspecies subjected to both chronic and acute UV-B radi-ation (280–320 nm). They showed that chronic UV-Bexposure significantly increased isoprene emission inQuercus gambelii but not in Mucuna pruriens, bothstrong isoprene emitting species, and did not induceisoprene emission in Acer platanoides, a non-emittingspecies. Furthermore, when M. pruriens and A. pla-tanoides were exposed to high acute UV-B exposure,there were no apparent effects on either photosynthesisor isoprene emission. More recently, long-term effects ofUV enhancement on BVOC emission were analyzed ona pristine, mixed-species subarctic peatland ecosystem.Tiiva et al. (2007) found that enhanced UV-B signif-icantly increased the emission of isoprene during thewarm periods of the second growing season, and at theend of the fourth growing season. On the other hand,the authors did not detect significant UV-B effect on theemissions from subarctic fen ecosystem, during the warmperiod in the third growing season. Then, in a follow-up study on the same subarctic peatland ecosystem,Faubert et al. (2010) showed no overall UV-B effect onisoprene and monoterpene emissions, apart from tolueneand 1-octene. Furthermore, both Tiiva et al. (2007) andFaubert et al. (2010) found no effects of UV-A radiation(320–400 nm) on BVOC emissions from the subarcticfen ecosystem. In addition, long-term increase in UV-Bradiation did not affect terpene concentration in leaf andwood of Pinus sylvestris and Picea abies seedlings (Tur-tola et al. 2006), monoterpene and sesquiterpene emis-sions from P. abies (Blande et al. 2009). Similarly, Winterand Rostas (2008) found no differences in qualitative andquantitative composition of induced volatiles in soybeangrown under ambient or attenuated UV radiation.
Trees also emit significant amounts of methanol andacetaldehyde (Kreuzwieser et al. 2001, Huve et al.2007). These compounds have a major impact on OHradicals. Consequently, methanol and acetaldehydeemissions strongly influence the oxidative capacity ofthe lower atmosphere and may significantly contributeto the formation of photochemical ozone. Despite thesecompounds can be induced by environmental stresses,the literature does not contain any information about the
effects of UV radiation on methanol and acetaldehydeemissions.
The aim of this study was to assess the direct effects ofshort term, acute exposure to UV-A on BVOC emissionsfrom hybrid poplar saplings. Hybrid poplar leaves wereexposed to 30 W m–2 (a radiation intensity which usu-ally occurs on sunny summer days), 60 W m–2 (a highintensity that may occur in the Mediterranean Basin)(Abukassem and Bero 2012), 90 W m–2 (which representreal-world episodic UV-A radiation stress, i.e. a bad butrealistic scenario) (Gelsor et al. 2011) and 120 W m–2
(an unrealistic extreme intensity) of UV-A irradiance.We specifically examined the effects of acute UV-Aradiation on the emissions of isoprene, methanol andacetaldehyde, on carbon assimilation, photochemicalreflectance index (PRI) and chlorophyll fluorescenceat the same time. In particular we analyzed theseasonality-dependent plasticity of responses to acuteUV-A exposure, as seasonal changes in the amount offlavonoids accumulated in the epidermal tissue by deter-mining to what extent UV-A penetrates leaves couldcontribute to build up protection mechanisms againstharmful UV radiation. To our knowledge, there areno earlier data on seasonality and intensity-dependentUV-A effects on BVOC emissions from broadleaf trees.
Materials and methods
Plant material
One-year-old Populus × canadensis saplings, propa-gated from physiologically matured trees growing in aclonal provenance trial in Italy, were transplanted beforebudburst into 6 dm3 plastic pots containing standardpotting compost (sand:peat:loam mixture 1:5:3) andgrown outdoor near Monterotondo (RM), Italy (42.1◦Nlatitude; 123 m above sea level), under natural sunlightconditions. The plants were regularly watered to potwater capacity and fertilized with Hoagland solutiononce a week in order to supply mineral nutrients at freeaccess rates (Centritto et al. 1999, Centritto 2005).
Gas exchange and BVOC measurements
Photosynthesis and BVOC emission were simultane-ously measured between 10:00 and 14:00 h during twodifferent experiments carried out in the laboratory undercontrolled conditions. In both experiments, poplarleaves were exposed to 30 W m–2, i.e. a radiationintensity that often occurs on sunny summer days inthe area where the experiment was performed (Diemozet al. 2011) and for this reason it will be referred toas ambient treatment in the remainder of the text, and
52 Physiol. Plant. 148, 2013

A B
Fig. 1. Spectral power distribution of the radiation emitted per unit wavelength by the (A) UV-A LEDs and (B) RGBA (red, green, blue, amber) LEDs.
increasing UV-A radiation (60, 90 and 120 W m–2).A computer controlled illumination system (ENFIS Ltd,Swansea, UK), that combines four lamps with 4 cm2
arrays of RGBA (red, green, blue, amber) LEDs (light-emitting diodes; 140 W) generating white light with alamp with 4 cm2 arrays UV-A LEDs (100 W, wavelengthrange 335–395 nm, peak wavelength at 365 nm; Fig. 1)was used. To mimic the ambient UV-A radiation levelrecorded in full sunlight conditions during summertime,leaves were exposed to saturating photosynthetic photonflux density (PPFD) of 1000 μmol m–2 s–1 of white lightsupplemented with a fixed intensity of 30 W m–2 UV-A.The UV-A intensity at leaf level was measured with aquantum-photoradiometer and thermometer DO 9721(Delta Ohm S.r.l., Padua, Italy) equipped with a LP9021 UVA radiometric probe.
PPFD saturated-photosynthesis (A) was measured dur-ing the UV-A treatments using a portable gas-exchangesystem (LI-6400, LI-COR, Lincoln, NE, USA) by enclos-ing a portion of a single fully expanded leaf in a 6 cm2
gas-exchange cuvette. Leaf temperature was set at 30◦C,and relative humidity in the leaf cuvette ranged between45 and 55%. Leaves were exposed to a flux of syntheticair, free of contaminants and pollutants, comprising N2,O2 and CO2 in atmospheric concentrations (80%, 20%and 380 μmol mol–1, respectively). Leaf temperaturewas measured with an array of three thermocouplesnear to the center and at two margins of the abaxialleaf side. Measurements were recorded when A was insteady state.
BVOC emission was detected simultaneously with Aby connecting the exhaust line from the leaf cuvettes toa proton transfer reaction mass spectrometer (PTR-MS;Ionicon, Innsbruck, Austria). A 200 ml min–1 of the airflowing out of the cuvette was diverted to the PTR-MS.The PTR-MS was set in a single ion mode to record traceof protonated masses of methanol (m/z 33), acetalde-hyde (m/z 45) and isoprene (m/z 69). The instrument wascalibrated daily with gaseous standard for each com-pound (Rivoira, Milano, Italy) and measurements were
validated with gas-chromatography analysis (SyntechSpectras BTX Analyzer GC 855, Groningen, The Nether-lands). The instrument was sampled at 0.15 l min–1; theionization energy E/N (E = electric field; N = number ofdensity of the drift tube molecules) was set at approx-imately 130 Td. Drift tube pressure, temperature andvoltage were 2.00 mbar, 45◦C and 600 V, respectively.The count rate of NO+ was < 2% of the count rateof H3O+ (approximately 4.7 × 106 counts s–1). Detailson the theory and practice of PTR-MS technique arereported in Lindinger et al. (1998).
Sequential increase in UV-A irradiance experiments
The first experiment was performed between the end ofJune and the beginning of July 2010. Individual leaves offive plants were kept under ambient UV-A radiation forabout 2 h. Leaves were then exposed for a 20-min periodat the UV-A intensities of 60, 90 and 120 W m–2, respec-tively; each exposition period was followed by a recov-ery of 20 min at ambient UV-A conditions prior to furtherincrease the UV-A intensities (Fig. 2A). In parallel withall the aforementioned gas exchange measurements,leaf spectral reflectance and chlorophyll a fluorescencewere also measured. The fluorescence yield (i.e. thequantum yield of PSII in the light, �F/Fm
′) was measuredby using a saturating pulse (10 000 μmol m–2 s–1) ofwhite light with a pulse-amplitude-modulated fluorom-eter (MINI-PAM, Heinz Walz GmbH, Germany). Theelectron transport rate was measured by fluorescence asdescribed by Genty et al. (1989).
Leaf reflectance was measured by using a portablespectrometer (ASD FieldSpec 3, Analytical SpectralDevices, Inc., Boulder, CO, USA), operating in the spec-tral range between 350 and 1025 nm. The spectrometerwas equipped with an optic fiber probe with an angu-lar field of view of 25◦. The instrument automaticallycalculates the reflectance value as a ratio between theincident radiation reflected from the surface target, andthe incident radiation reflected by a reference panel(Spectralon, Labsphere, Inc., North Sutton, NH, USA).
Physiol. Plant. 148, 2013 53

A
B
C
D
E
F
Fig. 2. Time course of (A) acute UV-A treatments, (B) photosynthesis (A) and stomatal conductance (gs), (C) photochemical reflectance index (PRI)and fluorescence yield (�F/Fm
′), and emissions of (D) isoprene, (E) methanol and (F) acetaldehyde. Leaves of Populus × canadensis saplings, kept undersaturating PPFD of 1000 μmol m–2 s–1 of white light, were first exposed to ambient UV-A radiation (30 W m–2) for about 2 h and then subjected toincreasing acute UV-A treatments for 20 min, which was then followed by a 20-min recovery at ambient UV-A conditions prior to further increasethe UV-A intensities, as shown in the panel A. Results shown in the panels B and C are means of five replicates ± 1 SEM, whereas data shown in thepanels D–F, measured on line with the PTR-MS, are from a single leaf but are representative of experiments replicated five times on different plants.
Reflectance spectra were collected from a distance of5 cm from the adaxial leaf surface with an angle of 60◦,to avoid shading the analyzed leaf surface. Reflectancespectra were pre-processed by using the VIEWSPECPRO
(ASD, Analytical Spectral Devices, Inc., USA) software.PRI was calculated as the (R531 – R570)/(R531 + R570) ratio(Gamon et al. 1990, 1997); where R531 is the reflectanceat 531 nm, which is affected by the xanthophyll cyclestatus, and R570 indicates reflectance at 570 nm, i.e. a ref-erence waveband that normalizes the value of the index.
UV-A intensity–response relationship experiments
The second experiment was performed in the middle ofJuly 2010 and then repeated during the first half of themonth of September 2010. Leaves were first exposed toambient UV-A radiation for about 2 h and then treatedwith the UV-A intensities of 60, 90 and 120 W m–2 (fiveplants per treatment), respectively, for 2 h. Measurementsof leaf reflectance were performed on leaves before andimmediately after the exposition to UV-A radiation usingthe same method described above.
Epidermal flavonoids were optically estimated in vivousing the portable fluorimetric sensor Multiplex 3 (Mx)(FORCE-A, Orsay, France) (Ben Ghozlen et al. 2010,Agati et al. 2011). Mx measures the chlorophyll (Chl)fluorescence in both the red, at the 680–690 nm (RF),and far-red, at 730–780 nm (FRF), bands sequentiallyexcited at four different excitation wavelengths in theUV-A (385 nm), and in the blue (460 nm), green (525 nm)and red (625 nm) spectral regions. The fluorescencesignals were integrated over a 8-cm circular area. Forthe present experiment, the Chl fluorescence signalsFRFR, excited with red (R) light, and FRFUV, excited withUV radiation, were used to calculate the flavonoid index(FLAV) as: FLAV = FRFR/FRFUV. Chlorophyll (CHL) in thefirst layers of mesophyll was evaluated by the CHL index:CHL = FRFR/RFR, where RFR is the red emission excitedunder red light. The origin of these indices is detailedelsewhere (Bilger et al. 1997, Buschmann 2007, BenGhozlen et al. 2010). The adaxial and abaxial surfacesof three individual leaves per plant were measured oncea week from middle of July 2010 to middle of September2010 on three different plants.
54 Physiol. Plant. 148, 2013

Statistics
All measurements were performed on fully expandedmature leaves of the same age. Data were tested by asimple factorial ANOVA (two-way maximum interactions)to determine the main effects of UV-A and time on alldependent variables. Where appropriate, the treatmentmeans were compared using Tukey’s post hoc test.
Results and discussion
UV radiation reaching the Earth’ surface has increasedbetween 1979 and 2008 (Herman 2010). Despite thesuccessful implementation of the Montreal Protocol inlimiting stratospheric ozone depletion, the levels of UVirradiances that reach the Earth’ surface are expectedto be still elevated in the coming decades (McKenzieet al. 2011). Exposure to UV radiation is known tobe harmful to terrestrial plants (Li et al. 2010, Ballareet al. 2011). UV-B is typically the most destructiveform of UV radiation because it has enough energy tocause photochemical damage. Whereas UV-A, whichis the most commonly encountered type of UV lightbecause stratospheric ozone layer absorbs very littleof this part of the UV spectrum, is thought to have aminor biological effect. However, the UV-B illuminationsystems used in supplementation studies have a spectrumthat includes also a substantial fraction of UV-A radiationand, thus, it is impossible to dissect the effects of UV-Bexposure alone (Ren et al. 2010). Then, only a fewstudies have been conducted in which UV-A and UV-Btreatments were applied separately. These studies, whichwere mostly performed by manipulating UV radiationby using filters that either transmitted UV or excludedonly UV-B or both UV-B and UV-A, respectively, haveshown some degree of negative modifications in theplant physiological and growth responses to UV-A(Hader 1996, Day et al. 1999, Kirchgeßner et al. 2003,Kotilainen et al. 2008, Yang and Yao 2008, Morales et al.2011). Consequently, there are still major uncertainties
regarding the basic mechanisms of plant physiologicalresponses to high UVA levels.
Sequential increase in UV-A irradiance
We performed a first supplementation (addition of UV-Aradiation using a controlled illumination system) studyat the beginning of summer. Hybrid poplar saplingswere irradiated with increasing acute UV-A intensities,ranging from ambient to unrealistically high amounts,for about 20 min at each different intensity (Fig. 2A), inorder to elucidate the plasticity of innate physiologicaltraits to UV-A radiation alone, as UV-A intensity canvary during the day, depending not only solar anglebut also on cloudiness conditions and atmosphericclearness. These irradiances induced significant effectson the physiological status of poplar leaves. After 20 minof exposition at 60, 90 and 120 W m–2 of UV-A,A resulted significantly reduced by about 17%, 33and 42%, respectively, compared with the pre-stressvalue (Fig. 2B). It is noteworthy that A recoveredonly partially (i.e. approximately 80% of the pre-stressvalue) when plants were exposed back to the ambientUV-A radiation (30 W m–2) after each acute treatment.Stomatal conductance increased quickly, although notsignificantly, upon exposure at 60 W m–2, and thenrapidly declined by approximately 28% of the pre-stress value during the 20 min-exposition (Fig. 2B).Interestingly, gs showed a similar trend when exposedto the higher UV-A radiations. Differently from A, gs
did not recover significantly during the recovery phases(Table 1). The increasing inhibition of the photosyntheticefficiency was also detected by the decreasing trendof both PRI and fluorescence yield during the wholeduration of the UV-A treatments (Fig. 2C). PRI wasoriginally developed to estimate rapid changes in thexanthophyll cycle that occurs over a minute time scale(Gamon et al. 1990), but there are increasing evidencesthat PRI is related to changes in the photosyntheticlight use efficiency (Gamon et al. 1997, Sun et al.
Table 1. Average values of photosynthesis (A, μmol m−2 s−1), stomatal conductance (gs, mol m−2 s−1), photochemical reflectance index (PRI),fluorescence yield (�F/Fm
′), isoprene, methanol and acetaldehyde emissions (nmol m−2 s−1) during the UV-A treatments shown in Fig. 2. Results aremeans of five replicates ± 1 SEM. Letters (a–f) indicate significant differences at P < 0.05 in the same row.
Time (min) 0 20 40 60 80 100 130UV-A 30 W m−2 60 W m−2 30 W m−2 90 W m−2 30 W m−2 120 W m−2 30 W m−2
A 12.00 ± 0.45 a 9.91 ± 0.53 b 10.56 ± 0.49 ab 8.13 ± 0.45 c 9.77 ± 0.35 b 7.38 ± 0.46 c 9.66 ± 0.45 bg s 0.333 ± 0.024 a 0.324 ± 0.025 a 0.240 ± 0.011 b 0.255 ± 0.017 b 0.242 ± 0.016 b 0.261 ± 0.010 b 0.273 ± 0.012 b�F/Fm
′ 0.463 ± 0.016 a 0.375 ± 0.009 b 0.229 ± 0.009 c 0.226 ± 0.021 c 0.217 ± 0.004 c 0.247 ± 0.010 c 0.247 ± 0.009 cPRI 0.041 ± 0.004 a 0.029 ± 0.006 b 0.0243 ± 0.004 b 0.0218 ± 0.003 b 0.020 ± 0.003 b 0.017 ± 0.001 b 0.0180 ± 0.001 bIsoprene 3.00 ± 0.49 f 8.45 ± 0.73 cd 6.97 ± 0.57 e 11.21 ± 0.66 b 8.29 ± 0.65 cd 13.69 ± 0.46 a 7.79 ± 0.80 deMethanol 2.47 ± 0.11 e 5.16 ± 0.20 a 3.87 ± 0.18 c 4.68 ± 0.15 b 3.26 ± 0.14 d 3.84 ± 0.15 c 3.14 ± 0.09 dAcetaldehyde 2.68 ± 0.25 a 2.79 ± 0.18 a 2.36 ± 0.49 ab 2.46 ± 0.16 ab 2.22 ± 0.09 b 2.51 ± 0.14 a 2.19 ± 0.14 b
Physiol. Plant. 148, 2013 55

2008). Similarly, the fluorescence yield is a measureof the overall efficiency of PSII reaction centers of aleaf in bright light. The significant strong decline inboth PRI and �F/Fm
′ may indicate that damages at themetabolic level occurred as the intensity of the UV-Atreatments increased. However, the dynamics of thesetwo parameters differed from that of photosynthesis,because �F/Fm
′ and especially PRI showed a very littleincrease during the recovery phases which followedeach UV-A acute treatment. This different decliningtrend of A and of the two parameters describing theefficiency of the photochemical apparatus, may indicatethat the partial recovery of photosynthesis during theexposition at ambient UV-A which followed the acutetreatments, could have been mostly caused by reduceddiffusional limitations (Centritto et al. 2003, Loreto andCentritto 2008).
Differently from the pattern shown by the photosyn-thetic parameters, the increasing acute UV-A treatmentssignificantly triggered the emissions of isoprene andmethanol (Fig. 2D, E). Isoprene emission resulted to bevery sensitive to the UV-A irradiances, because it burstincreasingly and significantly as the UV-A intensityincreased (Fig. 2D), and quickly and significantlydeclined during the recovery phases (Table 1). At theend of the experiment, isoprene emission was still aboutthree times higher than the pre-stress level. This is afurther evidence that environmental stress uncouplesphotosynthesis from isoprene emission (Brilli et al.2007, Loreto and Schnitzler 2010, Centritto et al.2011a). Methanol emission was also very responsive toincreasing UV-A irradiances, but contrary to isoprene,methanol showed the highest emission peak afterthe first applied UV-A intensity (60 W m–2) and twoprogressively and significantly lower bursts at 90 and120 W m–2 (Fig. 2E). Because methanol is strongly watersoluble (Huve et al. 2007), reduced gs (Fig. 2B) mayhave affected its gas-phase diffusion when leaves wereexposed at increasing UV-A irradiances. However, theburst in methanol emission recorded at 120 W m–2
was significantly lower than that recorded at 90 W m–2
despite similar gs (Table 1). This indicates that theseprogressively reduced bursts in methanol emission can-not be fully attributed to gas-phase diffusion limitations.Methanol is a short-chained oxygenated compound inwhich emission is usually elicited early in stress devel-opment as a consequence of degradation of cell wallpectins (Loreto and Schnitzler 2010, Niinemets 2010).Thus, the coupled methanol and gs results suggest thatthe strongest damages of cellular structures occurredduring the first UV-A treatment, although an immediatestrong release from a large methanol pool already builtinside the leaf mesophyll cannot be ruled out (Huve
et al. 2007, Filella et al. 2009). As for isoprene, methanolemission quickly and significantly declined during therecovery phases (Table 1), and the emission was stillsignificantly higher at the end of the experiment than atthe pre-stress phase. Similar to methanol, acetaldehydeis also strongly water soluble (Huve et al. 2007) andits emission can be induced by environmental stresses(Kreuzwieser et al. 2001). However, the apparent effectsof increase in UV-A radiation on acetaldehyde emissionwere much less visible (Fig. 2F), and also in this casethe emission peaks declined although not significantlyas the applied UV-A intensities increased (Table 1).Furthermore, there was a significant decline in theemission during the recovery phases, which resultedin lower acetaldehyde emission at the end of theexperiment than at the pre-stress phase. However, thisemission pattern may have been probably affected bythe degradation of acetaldehyde in the gas phase by theUV-A treatments, which may have likely obscured theoverall acetaldehyde emission (Ohko et al. 1998).
UV-A intensity–response relationships
A second supplementation experiment was performed tostudy the UV-A intensity–response relationships. Poplarleaves were exposed for 2 h to 30 W m–2 of UV-A,a radiation intensity which usually occurs on sunnysummer days, 60 W m–2, a high intensity that may occurin the Mediterranean Basin (Abukassem and Bero 2012),90 W m–2, which represent real-world episodic UV-Aradiation stress (Gelsor et al. 2011), and 120 W m–2,which are unrealistically high amounts of UV-A. Thisexperiment was performed in the middle of summer andthen was repeated toward the end of the same summerseason. As expected, independently of the seasonality,the ambient treatment did not cause any significantchange in the time course of both A (Fig. 3A, C) andisoprene emission (Fig. 3B, D). Whereas, the response ofthese two parameters to high UV-A intensities showed asignificant temporal evolution. In fact, there was a clearresponse to the different irradiance levels immediatelyafter the UV-A stress application when the experimentwas made in the middle of summer. While 60 W m–2 ofUV-A radiation affected photosynthesis only transitorily(Table 2), A decreased rapidly in leaves exposed to higherUV-A intensities (Fig. 3A): A was already significantlyreduced after about 10 min in leaves exposed to 90 and120 W m–2, and after about 30 min from the onset of theUV-A treatments, A was further inhibited by about 17 and28% at 90 and 120 W m–2, respectively, and remainedsteadily inhibited to similar levels until the end of thetreatments (i.e. −20 and −34% at 90 and 120 W m–2,respectively; Table 2). The spectral reflectance analysis
56 Physiol. Plant. 148, 2013
A B
C D
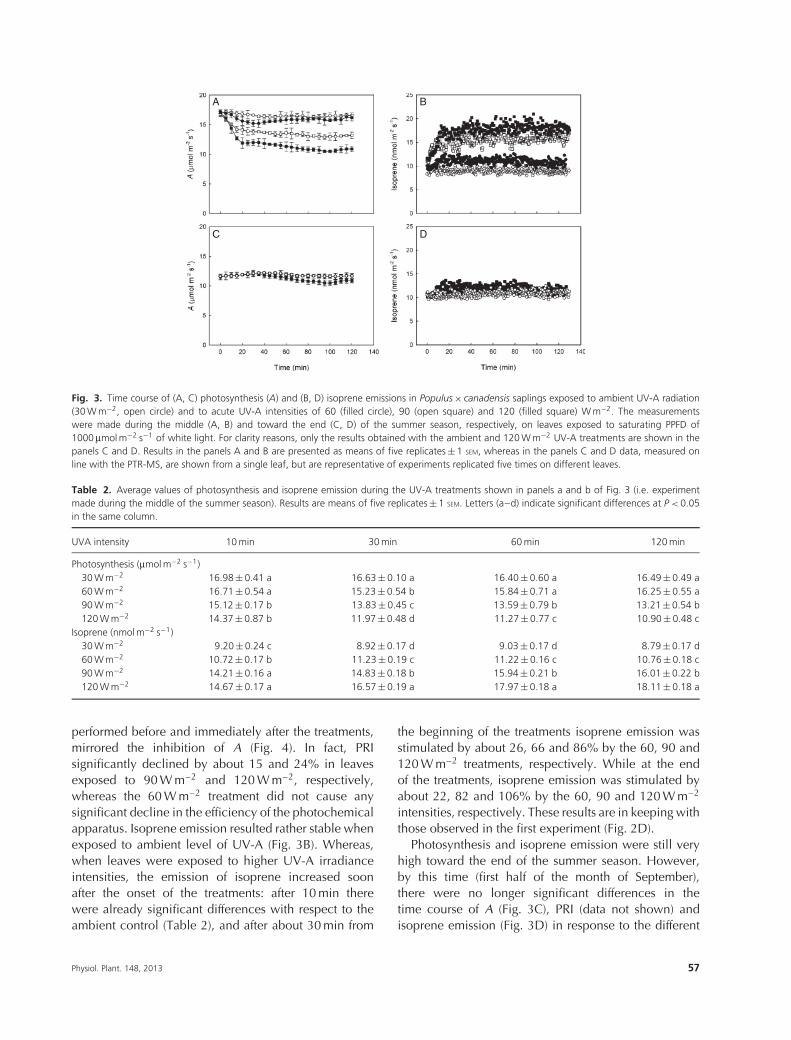
Fig. 3. Time course of (A, C) photosynthesis (A) and (B, D) isoprene emissions in Populus × canadensis saplings exposed to ambient UV-A radiation(30 W m−2, open circle) and to acute UV-A intensities of 60 (filled circle), 90 (open square) and 120 (filled square) W m−2. The measurementswere made during the middle (A, B) and toward the end (C, D) of the summer season, respectively, on leaves exposed to saturating PPFD of1000 μmol m−2 s−1 of white light. For clarity reasons, only the results obtained with the ambient and 120 W m−2 UV-A treatments are shown in thepanels C and D. Results in the panels A and B are presented as means of five replicates ± 1 SEM, whereas in the panels C and D data, measured online with the PTR-MS, are shown from a single leaf, but are representative of experiments replicated five times on different leaves.
Table 2. Average values of photosynthesis and isoprene emission during the UV-A treatments shown in panels a and b of Fig. 3 (i.e. experimentmade during the middle of the summer season). Results are means of five replicates ± 1 SEM. Letters (a–d) indicate significant differences at P < 0.05in the same column.
UVA intensity 10 min 30 min 60 min 120 min
Photosynthesis (μmol m−2 s−1)30 W m−2 16.98 ± 0.41 a 16.63 ± 0.10 a 16.40 ± 0.60 a 16.49 ± 0.49 a60 W m−2 16.71 ± 0.54 a 15.23 ± 0.54 b 15.84 ± 0.71 a 16.25 ± 0.55 a90 W m−2 15.12 ± 0.17 b 13.83 ± 0.45 c 13.59 ± 0.79 b 13.21 ± 0.54 b120 W m−2 14.37 ± 0.87 b 11.97 ± 0.48 d 11.27 ± 0.77 c 10.90 ± 0.48 c
Isoprene (nmol m−2 s−1)30 W m−2 9.20 ± 0.24 c 8.92 ± 0.17 d 9.03 ± 0.17 d 8.79 ± 0.17 d60 W m−2 10.72 ± 0.17 b 11.23 ± 0.19 c 11.22 ± 0.16 c 10.76 ± 0.18 c90 W m−2 14.21 ± 0.16 a 14.83 ± 0.18 b 15.94 ± 0.21 b 16.01 ± 0.22 b120 W m−2 14.67 ± 0.17 a 16.57 ± 0.19 a 17.97 ± 0.18 a 18.11 ± 0.18 a
performed before and immediately after the treatments,mirrored the inhibition of A (Fig. 4). In fact, PRIsignificantly declined by about 15 and 24% in leavesexposed to 90 W m–2 and 120 W m–2, respectively,whereas the 60 W m–2 treatment did not cause anysignificant decline in the efficiency of the photochemicalapparatus. Isoprene emission resulted rather stable whenexposed to ambient level of UV-A (Fig. 3B). Whereas,when leaves were exposed to higher UV-A irradianceintensities, the emission of isoprene increased soonafter the onset of the treatments: after 10 min therewere already significant differences with respect to theambient control (Table 2), and after about 30 min from
the beginning of the treatments isoprene emission wasstimulated by about 26, 66 and 86% by the 60, 90 and120 W m–2 treatments, respectively. While at the endof the treatments, isoprene emission was stimulated byabout 22, 82 and 106% by the 60, 90 and 120 W m–2
intensities, respectively. These results are in keeping withthose observed in the first experiment (Fig. 2D).
Photosynthesis and isoprene emission were still veryhigh toward the end of the summer season. However,by this time (first half of the month of September),there were no longer significant differences in thetime course of A (Fig. 3C), PRI (data not shown) andisoprene emission (Fig. 3D) in response to the different
Physiol. Plant. 148, 2013 57
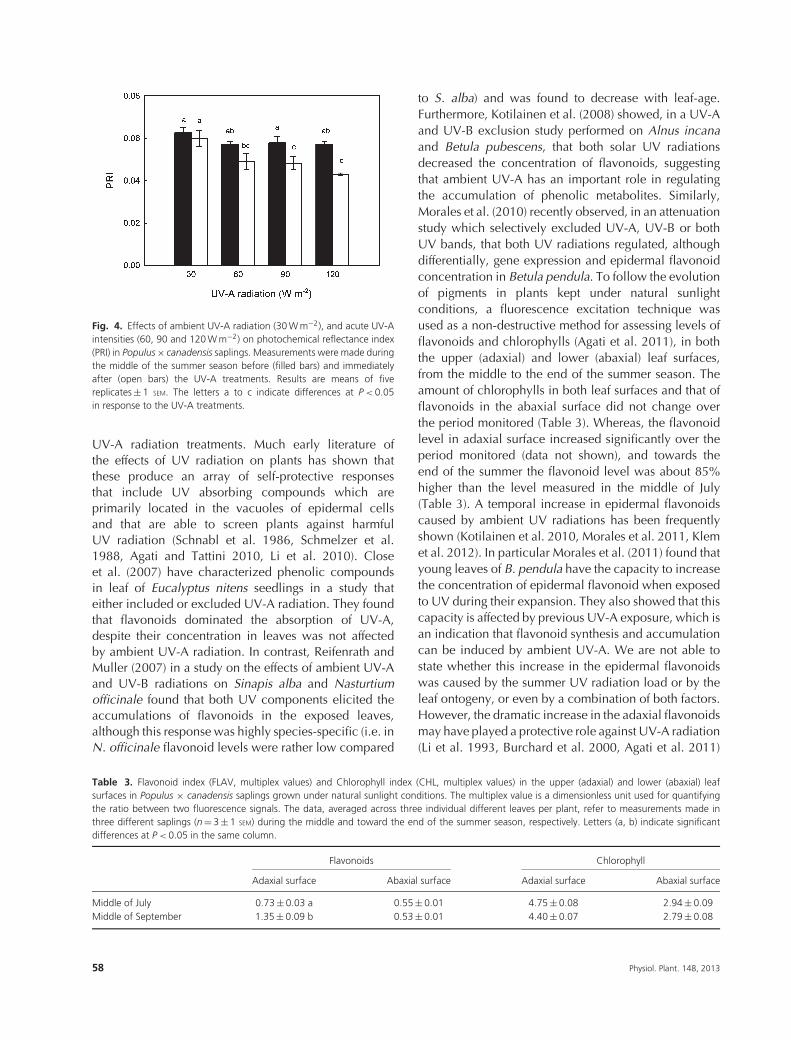
Fig. 4. Effects of ambient UV-A radiation (30 W m−2), and acute UV-Aintensities (60, 90 and 120 W m−2) on photochemical reflectance index(PRI) in Populus × canadensis saplings. Measurements were made duringthe middle of the summer season before (filled bars) and immediatelyafter (open bars) the UV-A treatments. Results are means of fivereplicates ± 1 SEM. The letters a to c indicate differences at P < 0.05in response to the UV-A treatments.
UV-A radiation treatments. Much early literature ofthe effects of UV radiation on plants has shown thatthese produce an array of self-protective responsesthat include UV absorbing compounds which areprimarily located in the vacuoles of epidermal cellsand that are able to screen plants against harmfulUV radiation (Schnabl et al. 1986, Schmelzer et al.1988, Agati and Tattini 2010, Li et al. 2010). Closeet al. (2007) have characterized phenolic compoundsin leaf of Eucalyptus nitens seedlings in a study thateither included or excluded UV-A radiation. They foundthat flavonoids dominated the absorption of UV-A,despite their concentration in leaves was not affectedby ambient UV-A radiation. In contrast, Reifenrath andMuller (2007) in a study on the effects of ambient UV-Aand UV-B radiations on Sinapis alba and Nasturtiumofficinale found that both UV components elicited theaccumulations of flavonoids in the exposed leaves,although this response was highly species-specific (i.e. inN. officinale flavonoid levels were rather low compared
to S. alba) and was found to decrease with leaf-age.Furthermore, Kotilainen et al. (2008) showed, in a UV-Aand UV-B exclusion study performed on Alnus incanaand Betula pubescens, that both solar UV radiationsdecreased the concentration of flavonoids, suggestingthat ambient UV-A has an important role in regulatingthe accumulation of phenolic metabolites. Similarly,Morales et al. (2010) recently observed, in an attenuationstudy which selectively excluded UV-A, UV-B or bothUV bands, that both UV radiations regulated, althoughdifferentially, gene expression and epidermal flavonoidconcentration in Betula pendula. To follow the evolutionof pigments in plants kept under natural sunlightconditions, a fluorescence excitation technique wasused as a non-destructive method for assessing levels offlavonoids and chlorophylls (Agati et al. 2011), in boththe upper (adaxial) and lower (abaxial) leaf surfaces,from the middle to the end of the summer season. Theamount of chlorophylls in both leaf surfaces and that offlavonoids in the abaxial surface did not change overthe period monitored (Table 3). Whereas, the flavonoidlevel in adaxial surface increased significantly over theperiod monitored (data not shown), and towards theend of the summer the flavonoid level was about 85%higher than the level measured in the middle of July(Table 3). A temporal increase in epidermal flavonoidscaused by ambient UV radiations has been frequentlyshown (Kotilainen et al. 2010, Morales et al. 2011, Klemet al. 2012). In particular Morales et al. (2011) found thatyoung leaves of B. pendula have the capacity to increasethe concentration of epidermal flavonoid when exposedto UV during their expansion. They also showed that thiscapacity is affected by previous UV-A exposure, which isan indication that flavonoid synthesis and accumulationcan be induced by ambient UV-A. We are not able tostate whether this increase in the epidermal flavonoidswas caused by the summer UV radiation load or by theleaf ontogeny, or even by a combination of both factors.However, the dramatic increase in the adaxial flavonoidsmay have played a protective role against UV-A radiation(Li et al. 1993, Burchard et al. 2000, Agati et al. 2011)
Table 3. Flavonoid index (FLAV, multiplex values) and Chlorophyll index (CHL, multiplex values) in the upper (adaxial) and lower (abaxial) leafsurfaces in Populus × canadensis saplings grown under natural sunlight conditions. The multiplex value is a dimensionless unit used for quantifyingthe ratio between two fluorescence signals. The data, averaged across three individual different leaves per plant, refer to measurements made inthree different saplings (n = 3 ± 1 SEM) during the middle and toward the end of the summer season, respectively. Letters (a, b) indicate significantdifferences at P < 0.05 in the same column.
Flavonoids Chlorophyll
Adaxial surface Abaxial surface Adaxial surface Abaxial surface
Middle of July 0.73 ± 0.03 a 0.55 ± 0.01 4.75 ± 0.08 2.94 ± 0.09Middle of September 1.35 ± 0.09 b 0.53 ± 0.01 4.40 ± 0.07 2.79 ± 0.08
58 Physiol. Plant. 148, 2013

by shielding leaves and then nullifying the impact onphotosynthesis (Fig. 3C) and isoprene emission (Fig. 3D).
In previous experiments performed on subarctic fenecosystem exposed either to ambient (Faubert et al.2010) or to enhanced UV-A radiation (Tiiva et al.2007) no effects were found on BVOC emissions. WhileFaubert et al.’s (2010) findings are in keeping with theresults obtained in our study on poplar saplings exposedto ambient UV-A intensities (i.e. 30 W m–2), Tiiva et al.’s(2007) results are in contrast with our findings. This isbecause Tiiva et al. (2007) did not found any stimulationin isoprene emitted by the subarctic fen ecosystemduring three different growing season under enhancedUV-A exposure, despite this ecosystem emitted highamount of isoprene, during the warm periods of thesecond growing season and at the end of the fourthgrowing season, when exposed to enhanced UV-Bradiation. Unfortunately, no information is provided byTiiva et al. (2007) about the UV-A intensities used intheir study. Consequently, it is impossible to understandwhether these contrasting findings resulted from differentintensity of UV-A radiations used or from species-specificresponses.
Conclusions
Growing evidence shows that human activities havedirect influences on the atmospheric physical and chem-ical properties. A major change is an increase in thelevels of UV irradiance reaching the earth’s surface. Theobjective of this supplementation study was to increaseknowledge of the mechanisms of primary and secondarymetabolism processes to UV-A irradiance. The UV-Acomputer-controlled illumination system employed inthis study allowed estimates of intensity–response func-tions. Our findings showed that both the mechanismsregulating photosynthesis and BVOC emissions are verysensitive to high UV-A. However, the responsiveness ofboth processes is reduced over the summer season likelybecause of the natural temporal increase in epidermalflavonoids in the adaxial surface which shield the leafinterior against UV. Taken collectively, our findingsshow that isoprene emissions are positively correlatedto UV-A radiation, but that there is an acclimation ofthe plants to UV radiation over the growing season. Thisis an important notion in the discussion of the effects ofUV radiations on the BVOC emissions which are majorcomponents in many global physicochemical processesas well as ecological process.
Acknowledgements – The authors would like to extend theirsincere thanks to Marco Giorgetti and Fabrizio Lelli fortheir technical assistance in the field. This work has been
funded by Italian National Research Council (project RSTL-DG.RSTL.010.003).
References
Abukassem I, Bero MA (2012) Radiometriccharacterization of radiochromic gel material appliedfor UVA radiation dosimetry. Radiat Meas 47: 34–39
Agati G, Tattini M (2010) Multiple functional roles offlavonoids in photoprotection. New Phytol 186:786–793
Agati G, Cerovic ZG, Pinelli P, Tattini M (2011)Light-induced accumulation of ortho-dihydroxylatedflavonoids as non-destructively monitored bychlorophyll fluorescence excitation techniques. EnvironExp Bot 73: 3–9
Ballare CL, Caldwell MM, Flint SD, Robinson A, BornmanJF (2011) Effects of solar ultraviolet radiation onterrestrial ecosystems. Patterns, mechanisms, andinteractions with climate change. Photochem PhotobiolSci 10: 226–241
Ben Ghozlen N, Cerovic ZG, Germain C, Toutain S,Latouche G (2010) Non-destructive optical monitoringof grape maturation by proximal sensing. Sensors 10:10040–10068
Bilger W, Veit M, Schreiber L, Schreiber U (1997)Measurement of leaf epidermal transmittance of UVradiation by chlorophyll fluorescence. Physiol Plant101: 754–763
Blande J, Turunen K, Holopainen JK (2009) Pine weevilfeeding on Norway spruce bark has a stronger impact onneedle VOC emissions than enhanced ultraviolet-Bradiation. Environ Pollut 157: 174–180
Brilli F, Barta C, Fortunati A, Lerdau M, Loreto F, CentrittoM (2007) Response of isoprene emission and carbonmetabolism to drought in white poplar (Populus alba)saplings. New Phytol 175: 244–254
Burchard P, Bilger W, Weissenbock G (2000) Contributionof hydroxycinnamates and flavonoids to epidermalshielding of UV-A and UV-B radiation in developing ryeprimary leaves as assessed by ultraviolet inducedchlorophyll fluorescence measurements. Plant CellEnviron 23: 1373–1380
Buschmann C (2007) Variability and application of thechlorophyll fluorescence emission ratio red/far-red ofleaves. Photosynth Res 92: 261–271
Centritto M (2005) Photosynthetic limitations and carbonpartitioning in cherry in response to water deficit andelevated [CO2]. Agric Ecosyst Environ 106: 233–242
Centritto M, Lee HSJ, Jarvis PG (1999) Long-term effects ofelevated carbon dioxide concentration and provenanceon four clones of Sitka spruce (Picea sitchensis) I. Plantgrowth, allocation and ontogeny. Tree Physiol 19:799–806
Physiol. Plant. 148, 2013 59

Centritto M, Loreto F, Chartzoulakis K (2003) The use oflow [CO2] to estimate diffusional and non-diffusionallimitations of photosynthetic capacity of salt-stressedolive saplings. Plant Cell Environ 26: 585–594
Centritto M, Brilli F, Fodale R, Loreto F (2011a) Differentsensitivity of isoprene emission, respiration, andphotosynthesis to high growth temperature coupled withdrought stress in black poplar (Populus nigra). TreePhysiol 31: 275–286
Centritto M, Tognetti R, Leitgeb E, Strelcova K, Cohen S(2011b) Above ground processes – Anticipating climatechange influences. In: Bredemeier M, Cohen S,Godbold DL, Lode E, Pichler V, Schleppi P (eds) ForestManagement and the Water Cycle: An Ecosystem-BasedApproach. Ecological Studies, Vol. 212. Springer,New York, pp 31–64
Close DC, Mcarthur C, Hagerman AE, Davies NW, BeadleCL (2007) Phenolic acclimation to ultraviolet-Airradiation in Eucalyptus nitens seedlings raised across anutrient environment gradient. Photosynthetica 45:36–42
Day TA, Ruhland CT, Grobe CW, Xiong F (1999) Growthand reproduction of Antarctic vascular plants inresponse to warming and UV radiation reductions in thefield. Oecologia 119: 24–35
Diemoz H, Siani AM, Casale GR, di Sarra A, Serpillo B,Petkov B, Scaglione S, Bonino A, Facta S, Fedele F,Grifoni D, Verdi L, Zipoli G (2011) First nationalintercomparison of solar ultraviolet radiometers in Italy.Atmos Meas Tech Discuss 4: 2789–2826
Faubert P, Tiiva P, Rinnan A, Rasanen J, Holopainen JK,Holopainen T, Kyro E, Rinnan R (2010) Non-methanebiogenic volatile organic compound emissions from asubarctic peatland under enhanced UV-B radiation.Ecosystems 13: 860–873
Filella I, Penuelas J, Seco R (2009) Short-chainedoxygenated VOC emissions in Pinus halepensis inresponse to changes in water availability. Acta PhysiolPlant 31: 311–318
Gamon JA, Field CB, Bilger W, Bjorkman O, Fredeen AL,Penuelas J (1990) Remote sensing of the xanthophyllcycle and chlorophyll fluorescence in sunflower leavesand canopies. Oecologia 85: 1–7
Gamon JA, Serrano L, Surfus JS (1997) The photochemicalreflectance index: an optical indicator of photosyntheticradiation use efficiency across species, functional types,and nutrient levels. Oecologia 112: 492–501
Gelsor N, Pingcuo N, Wangmu T, Kjeldstad B, Chen Y-C,Frette Ø, Stamnes JJ, Bhattarai BK, Dhalback A (2011)Solar UV irradiance measurements at four sites in Tibet.J Inst Eng 8: 75–86
Genty B, Briantais JM, Baker NR (1989) The relationshipbetween the quantum yield of photosynthetic electrontransport and quenching of chlorophyll fluorescence.Biochim Biophys Acta 990: 87–92
Hader DP (1996) Effects of solar radiation on local andGerman wheat seedlings in a Chilean high mountainstation. J Photochem Photobiol B: Biol 35: 181–187
Harley P, Deem G, Flint S, Caldwell M (1996) Effects ofgrowth under elevated UV-B on photosynthesis andisoprene emission in Quercus gambelii and Mucunapruriens. Glob Change Biol 2: 149–154
Hegglin MI, Shepherd TG (2009) Large climate-inducedchanges in ultraviolet index and stratosphere-to-troposphere ozone flux. Nat Geosci 2: 687–691
Herman JR (2010) Global increase in UV irradiance duringthe past 30 years (1979–2008) estimated from satellitedata. J Geophys Res 115: D04203
Huve K, Christ MM, Kleist E, Uerlings R, Niinemets U,Walter A, Wildt J (2007) Simultaneous growth andemission measurements demonstrate an interactivecontrol of methanol release by leaf expansion andstomata. J Exp Bot 58: 1783–1793
Kirchgeßner H-D, Reichert K, Hauff K, Steinbrecher R,Schnitzler J-P, Pfundel EE (2003) Light and temperature,but not UV radiation, affect chlorophylls andcarotenoids in Norway spruce needles (Picea abies (L.)Karst.). Plant, Cell Environ 26: 1169–1179
Klem K, Ac A, Holub P, Kovac D, Spunda V, Robson TM,Urban O (2012) Interactive effects of PAR and UVradiation on the physiology, morphology and leafoptical properties of two barley varieties. Environ ExpBot 75: 52–64
Kotilainen T, Tegelberg R, Julkunen-Tiitto R, Lindfors A,Aphalo PJ (2008) Metabolite specific effects of solarUV-A and UV-B on alder and birch leaf phenolics. GlobChange Biol 14: 1294–1304
Kotilainen T, Tegelberg R, Julkunen-Tiitto R, Lindfors A,O’Hara RB, Aphalo PJ (2010) Seasonal fluctuations inleaf phenolic composition under UV manipulationsreflect contrasting strategies of alder and birch trees.Physiol Plant 140: 297–309
Kreuzwieser J, Harren FJM, Laarhoven LJ, Boamfa I, teLintel-Hekkert S, Scheerer U, Huglin C, Rennenberg H(2001) Acetaldehyde emission by the leaves of trees -correlation with physiological and environmentalparameters. Physiol Plant 113: 41–49
Li J, Ou-Lee T, Raba R, Amundson RG, Last RL (1993)Arabidopsis flavonoid mutants are hypersensitive toUV-B Irradiation. Plant Cell 5: 171–179
Li F-R, Peng S-L, Chen B-M, Hou Y-P (2010) Ameta-analysis of the responses of woody andherbaceous plants to elevated ultraviolet-B radiation.Acta Oecologica 36: 1–9
Lindinger W, Hansel A, Jordan A (1998) On-linemonitoring of volatile organic compounds at pptvlevels by means of protontransfer reaction massspectrometry (PTR-MS): medical applications, foodcontrol and environmental research. Int J Mass Spectr173: 191–241
60 Physiol. Plant. 148, 2013

Loreto F, Centritto M (2008) Leaf carbon assimilation in awater-limited world. Plant Biosyst 142: 154–161
Loreto F, Schnitzler JP (2010) Abiotic stresses and inducedBVOCs. Trends Plant Sci 15: 154–166
McKenzie RL, Aucamp PJ, Bais AF, Bjorn LO, Ilyas M,Madronich S (2011) Ozone depletion and climatechange: impacts on UV radiation. Photochem PhotobiolSci 10: 182–198
Morales LO, Tegelberg R, Brosche M, Keinanen M,Lindfors A, Aphalo PJ (2010) Effects of solar UV-A andUV-B radiation on gene expression and phenolicaccumulation in Betula pendula leaves. Tree Physiol 30:923–934
Morales LO, Tegelberg R, Brosche M, Lindfors A, Siipola S,Aphalo PJ (2011) Temporal variation in epidermalflavonoids due to altered solar UV radiation ismoderated by the leaf position in Betula pendula.Physiol Plant 143: 261–270
Niinemets U (2010) Mild versus severe stress and BVOCs:thresholds, priming and consequences. Trends Plant Sci15: 145–153
Ohko Y, Tryk DA, Hashimoto K, Fujishima A (1998)Autoxidation of acetaldehyde initiated by TiO2
photocatalysis under weak UV illumination. J PhysChem B 102: 2699–2704
Paul ND, Gwynn-Jones D (2003) Ecological roles of solarUV radiation: towards an integrated approach. TrendsEcol Evol 18: 48–55
Reifenrath K, Muller C (2007) Species-specific and leaf-agedependent effects of ultraviolet radiation on twoBrassicaceae. Phytochemistry 68: 875–885
Ren W, Tan H, Wu J, Deng Y, Wu Y, Tang Y, Cui X (2010)UV light spectral response of photosyntheticphotochemical efficiency in alpine mosses. J Plant Ecol3: 17–24
Schmelzer E, Jahnen W, Hahlbrock K (1988) In situlocalization of light-induced chalcone-synthase mRNA,chalcone synthase, and flavonoid end products inepidermal cells of parsley leaves. Proc Natl Acad SciUSA 85: 2989–2993
Schnabl N, Weissenbock G, Scharf H (1986) In vivomicrospectrophotometric characterization of flavonolglycosides in Vicea faba guard and epidermal cells. JExp Bot 37: 61–72
Sun P, Grignetti A, Liu S, Casacchia R, Salvatori R,Pietrini F, Loreto F, Centritto M (2008) Associatedchanges in physiological parameters and spectralreflectance indices in olive (Olea europaea L.) leaves inresponse to different levels of water stress. Int RemoteSens 29: 1725–1743
Tiiva P, Rinnan R, Faubert P, Rasanen J, Holopainen T,Kyro E, Holopainen JK (2007) Isoprene emission from asubarctic peatland under enhanced UV-B radiation.New Phytol 176: 346–355
Turtola S, Sallas L, Holopainen JK, Julkunen-Tiitto R,Kainulainen P (2006) Long-term exposure to enhancedUV-B radiation has no significant effects on growth orsecondary compounds of outdoor-grown Scots pine andNorway spruce seedlings. Environ Pollut 144: 166–171
Vickers CE, Possell M, Cojocariu CI, Laothawornkitkul J,Ryan A, Mullineaux PM, Hewitt CN (2009) Isoprenesynthesis protects tobacco plants from oxidative stress.Plant Cell Environ 32: 520–531
Winter TR, Rostas M (2008) Ambient ultraviolet radiationinduces protective responses in soybean but does notattenuate indirect defense. Environ Pollut 155: 290–297
Yang YQ, Yao Y (2008) Photosynthetic responses to solarUV-A and UV-B radiation in low-and high-altitudepopulations of Hippophae rhamnoides. Photosynthetica46: 307–311
Edited by A. Krieger-Liszkay
Physiol. Plant. 148, 2013 61