breaking onion bulb endodormancy with hydrogen peroxide
TRANSCRIPT

HORTSCIENCE 54(10):1694–1702. 2019. https://doi.org/10.21273/HORTSCI14118-19
Breaking Onion Bulb Endodormancywith Hydrogen PeroxideChristopher J. D’AngeloIllinois Foundation Seeds, Inc., Plover, WI
Irwin L. GoldmanDepartment of Horticulture, University of Wisconsin-Madison, 1575 LindenDrive, Madison, WI 53706
Additional index words. Allium cepa, breeding, dormancy, vernalization
Abstract. Long-day storage onion bulbs (Allium cepa) undergo a period of endodormancythat begins before harvest and lasts for several weeks, depending on the genotype. Onionseed production relies on cold treatment to break endodormancy and vernalize the bulbs.When bulbs are planted shortly after harvest, endodormancy results in delayed growthand, in turn, slower flowering and seed production. Through this work we sought toexplore the use of hydrogen peroxide as a treatment to break dormancy in onion bulbs.Endodormant bulbs of two long-day cultivars, Cortland and Sherman, were treated withhydrogen peroxide solutions at various concentrations in a series of experiments over a 3-year period and were monitored for root and leaf (sprout) development. We found a 2- to4-hour exogenous treatment of 20% (weight by volume) hydrogen peroxide to be highlyeffective at initiating uniform root growth in endodormant bulbs.When compared with apurified water control, the 20% treatment resulted in a 61.3% average reduction in thetime to rooting in 2016. We also observed improved uniformity in rooting time between‘Cortland’ and ‘Sherman’ in all 3 years of this work. We propose this novel method as atool for breeders, researchers, and seed producers seeking rapid, uniform endodormancyrelease in onion bulbs to hasten seed production.
Onion (Allium cepa) is a globally impor-tant vegetable crop. It is grown in more than160 countries and on more than 4.9 millionha. Global onion production has been in-creasing steadily since 1990 (FAO, 2016).The most widely produced marketable prod-uct of common onion is a dormant bulb. It isimportant for the bulbs to remain unsproutedwhile in storage because premature sproutingresults in poor-quality bulbs with low con-sumer acceptance. Dormancy plays an in-tegral role in onion bulb storage longevityand sprout suppression, but can also delaygrowth when it is desired. Dormancy release,along with vernalization, are key processesfor seed production during which rapid,uniform growth and development are valu-able. There is great economic interest incontrolling dormancy release for both breed-ing and seed production.
Dormancy is a temporary suspension ofvisible growth of any plant structure contain-ing a meristem (Lang et al., 1987). There arethree classifications of dormancy, which are
defined by the origin of initial growth sup-pression cues: endodormancy, paradormancy,and ecodormancy. Endodormancy occurswhenthe signal to suspend growth originates withinthe affected structure. Paradormancy is a re-sponse to a signal that originates in an organother than the affected structure; ecodormancyis a response to unfavorable and extremeenvironmental conditions, such as very highor low temperatures or water availability (Langet al., 1987). All three types of dormancy mayoccur simultaneously within a plant, but thetype present in each structure depends on thelife stage, physiology, and environmental con-ditions. For onion bulbs that have been har-vested and put into cold storage, endodormancyand ecodormancy are the primary processesthat regulate growth (Chope et al., 2012a).
Bulb endodormancy is prevalent in long-day storage onion germplasm and begins totake effect in the weeks before harvest. Asonions growing in the field complete theirbulbing phase, they cease production of newleaves and the remaining leaves topple asthey senesce. This senescence is often usedby growers as an indication that the bulbs areready to be harvested. When removed fromthe field, bulbs are taken into storage wherethey remain dormant. After �3 weeks, thebulbs resume preharvest levels of cellulardivision and transcription (Chope et al.,2012b; Pak et al., 1995). Pak et al. (1995)found that as cellular division resumes, bulbsare capable of rooting when exposed to highmoisture. This marks the transition fromendodormancy to ecodormancy.
In onion, endodormancy is typically bro-ken through exposure to cold temperatures.These cold treatments also serve to vernal-ize the bulbs. Little is known about themechanism of endodormancy release inmost plant species, but in seeds, which havebeen investigated most thoroughly, there isgenerally a strong association with abscisicacid degradation and gibberellic acid syn-thesis as dormancy is broken (N�ee et al.,2017). Successful use of chemical treat-ments to break endodormancy in seeds andthe buds of woody plants using compoundssuch as hydrogen cyanamide have beenreported (Horvath et al., 2003; Mohamedet al., 2012; Vergara and P�erez, 2010).Hydrogen cyanamide acts as a catalaseinhibitor, which is a key enzyme that removesreactive oxygen species from plant cells.When administered to endodormant grapevinebuds, hydrogen cyanamide was shown to in-crease levels of hydrogen peroxide in cellsbefore breaking dormancy (Mazzitelli et al.,2007; Mohamed et al., 2012; P�erez and Lira,2005). P�erez and Lira (2005) hypothesizedthat hydrogen peroxide acts as a secondarymessenger to signal the release of endodor-mancy in grapevines (Vitis vinifera L.). Thishypothesis was later supported by Mohamedet al. (2012) upon finding that, before dor-mancy release, an accumulation of hydrogenperoxide causes a temporary oxidative stressin grape buds following an exogenous appli-cation of hydrogen cyanamide. Similar hy-potheses have been posited regardinghydrogen peroxide’s role as a signal in seeddormancy release (Oracz et al., 2007). Chem-ical signaling under stress is a common role forreactive oxygen species, including hydrogenperoxide. Specifically, hydrogen peroxide hasbeen shown to play a role in a range ofcellular processes including programmedcell death, response to wounding, and absci-sic acid-mediated stomatal closure (El-Maarouf-Bouteau and Bailly, 2008). Anexogenous application of hydrogen perox-ide has also been used successfully to breakdormancy in seeds (Liu et al., 2011). Muchlike the findings in endodormant buds andseeds of other plant species, transcriptionalanalysis of onion bulbs during dormancyrelease shows that transcripts associatedwith genes related to defense and stressresponse are highly upregulated during thetransition from endodormancy to ecodor-mancy (Chope et al., 2012b).
The hypothesis by P�erez and Lira (2005),in combination with the demonstrated suc-cess in using hydrogen peroxide to break seeddormancy, made it an interesting candidatefor use on onion bulbs. Through this work, wesought to test the effects of an exogenousapplication of hydrogen peroxide on endo-dormant onion bulbs and to determinewhether these treatments could serve as aneffective way to break endodormancy.
Materials and Methods
Field design. A series of experiments totest the effects of exogenous hydrogen peroxide
Received for publication 9 Apr. 2019. Accepted forpublication 29 July 2019.We thank Ned, John, and Joan Crescio of Jack’sPride Farms in Randolph, WI, for their assistanceand for donating the bulbs used in this study.We claim no affiliation with Acros Organics, BDH,or their respective parent companies, ThermoFisher Scientific and VWR International/Avantor.This study does not constitute or imply an endorse-ment of either company or their products.I.L.G. is the corresponding author. E-mail:[email protected].
1694 HORTSCIENCE VOL. 54(10) OCTOBER 2019

treatments on onion bulb endodormancywas conducted from 2014 to 2016. The2014 experiment was conducted as a pilotstudy. Bulbs of the F1 hybrid cultivarsCortland and Sherman (Bejo Seeds, Oceano,CA) were used in all years of this experi-ment. The bulbs were grown under com-mercial growing conditions on muck soil atJack’s Pride Farm in Randolph, WI, andwere replicated at a second location inMarkesan, WI, during 2015 and 2016. Seedwas planted between late April and earlyMay on raised beds at a density of 36 plants/m using a modified Planet Junior (ColePlanter Company, Albany, GA) planterequipped with a cone seeder attachment in3.66-m rows with a 1.22-m alley and 30-cmrow spacing. No maleic hydrazide or othersprout inhibitors were applied to the plantsin the field. All bulbs were topped in thefield, harvested in early September, andcured for 1 week in a dark, well-ventilatedstorage room at ambient temperature. Bulbsfrom the 2014 experiment were only har-vested from the Randolph, WI, location. Sixbulbs per treatment, cultivar, and locationwere used in all years of this study; however,in 2016, there were two replications perlocation from which bulbs were harvested.The 2014 and 2015 experiments used bulbsthat were harvested from only one replica-tion per location. The diameter of each bulbwas measured transversely at the widestpoint using Vernier calipers.
Chemical source. Two concentrated sour-ces of hydrogen peroxide were used in theseexperiments. The Acros Organics hydrogenperoxide 50% by weight solution in water(AC302865000; Acros Organics, Geel, Bel-gium) was first used in 2014. In 2015, theBDH1 hydrogen peroxide 30% stabilizedACS (BDH7690-1; VWR, Radnor, PA) so-lution was used. Both products use additivesto stabilize the hydrogen peroxide and toslow degradation of the product during stor-age. However, stabilizer use can differ bymanufacturer, and each product used differ-ent stabilizers. The Acros Organics product isstabilized with up to 200 ppm phosphate froman undisclosed source, whereas the BDHproduct uses sodium stannate. Followingthe 2015 experiment, the experiment in2016 was conducted using both products toassess whether these differently stabilizedsolutions influenced dormancy release. Forboth products and in all years of this study,the hydrogen peroxide stock solution wasdiluted to the appropriate final concentrationusing purified water (E-pure Water Purifica-tion model 7117; Thermo Fisher Scientific,Waltham, MA) immediately before treatingthe bulbs.
Bulb treatment. The top third of eachbulb was cut so that the basal plate and twothirds of the bulb remained. Bulbs wereplaced into a plastic container with the cutside facing down while the appropriatehydrogen peroxide solution was added tothe container. Initially, in 2014, 10%, 20%,and 30% hydrogen peroxide solutions weretested and compared with a control group
that used only purified water (0% hydrogenperoxide). After finding all treatment groupsto be effective in 2014, a lower concentra-tion was included in the subsequent exper-iments. A 4% hydrogen peroxide solutionwas chosen to be included in the 2015 and2016 experiments because this is typical ofthe concentration found in the consumer-grade product and is widely available instores across the United States. Enoughsolution was added to each container so thatthe cut side of the bulb was submerged in�1.5 cm of liquid (Fig. 1A). The bulbs wereleft to soak uncovered in the solution for 4 hat ambient temperature. An alternate 2-hsoak was also evaluated in 2015 alongsidethe 4-h treatment. Upon completion of the
treatment, the bulbs were removed from thesolution and rinsed under cool tap water toremove any excess solution. The bulbs werethen planted in a greenhouse in a completelyrandomized design (Fig. 1B). Bulbs wereinspected every 2 to 3 d for rooting andsprouting. The date was recorded for theinitial root and sprout observations for eachplant.
Greenhouse culture. All bulbs were plantedin black plastic pots. The potting media was a2:1mix of silty loamcompost soil collected fromtheWestMadison Agricultural Research Stationand soilless medium (MetroMix 366; Sun GroHorticulture, Agawam, MA). The greenhousetemperature in 2014 was kept at 16 �C usingsteam baseboard radiators and a combination of
Fig. 1. (A) Cut onion bulbs soaking in 30%hydrogen peroxide solutions from the 2016 experiment. The topbucket is filled with sodium stannate-stabilized hydrogen peroxide; the bottom bucket containsphosphate-stabilized hydrogen peroxide. Note the difference in foaming between the two solutions. (B)Onions planted in the greenhouse following treatment with hydrogen peroxide. (C) Rooting onion inthe greenhouse. Bulbs were lifted from the soil to observe and record initial root growth. (D) Sprouting‘Sherman’ onion from the 20% treatment group.
HORTSCIENCE VOL. 54(10) OCTOBER 2019 1695

manual and automated vents. In 2015–16,air-conditioned greenhouses with forced-airheating were used to maintain a temperatureof 20 �C. In all years, high-pressure sodiumsupplemental lighting was used to maintaina 16-h daylength with a photon flux densityof 103 mmol·m–2·s–1 in 2014, and 343mmol·m–2·s–1 in 2015 and 2016 (ModelQMSS; Apogee Instruments, Logan, UT).Plants were watered as needed daily andwere fertilized as needed with a 400-mg·L–1
mixture of 20N–4.365P–16.602K fertilizerwith micronutrients (Peters ProfessionalPeat-Lite Special; ICL Specialty Fertilizers,Dublin, OH).
Data collection. Following treatment andsubsequent planting in the greenhouse, bulbswere monitored every 2 to 3 d for signs ofroot development and sprouting (Fig. 1C andD). The date that each event was first ob-served was recorded.
Statistics. Natural logarithmic transfor-mations were performed on all days-to-rooting and days-to-sprouting data. Individualbulbs were treated as experimental unitswith replication over time. Measuring in-dividual bulbs allowed us to assess theresponse of bulb-to-bulb variation treat-ment with hydrogen peroxide. Analysis ofvariance (ANOVA) was performed using the‘car’ package in R with type three sums ofsquares and effect sizes were measured usingan h2 calculation from the ‘sjstats’ package.Tukey’s highly significant difference (HSD) testwas also performed in R using the ‘stats’package (R Development Core Team, 2018).Nonsignificant interaction terms, such as con-centration:location and concentration:varietywere not included in the linear models forthis analysis.
Results
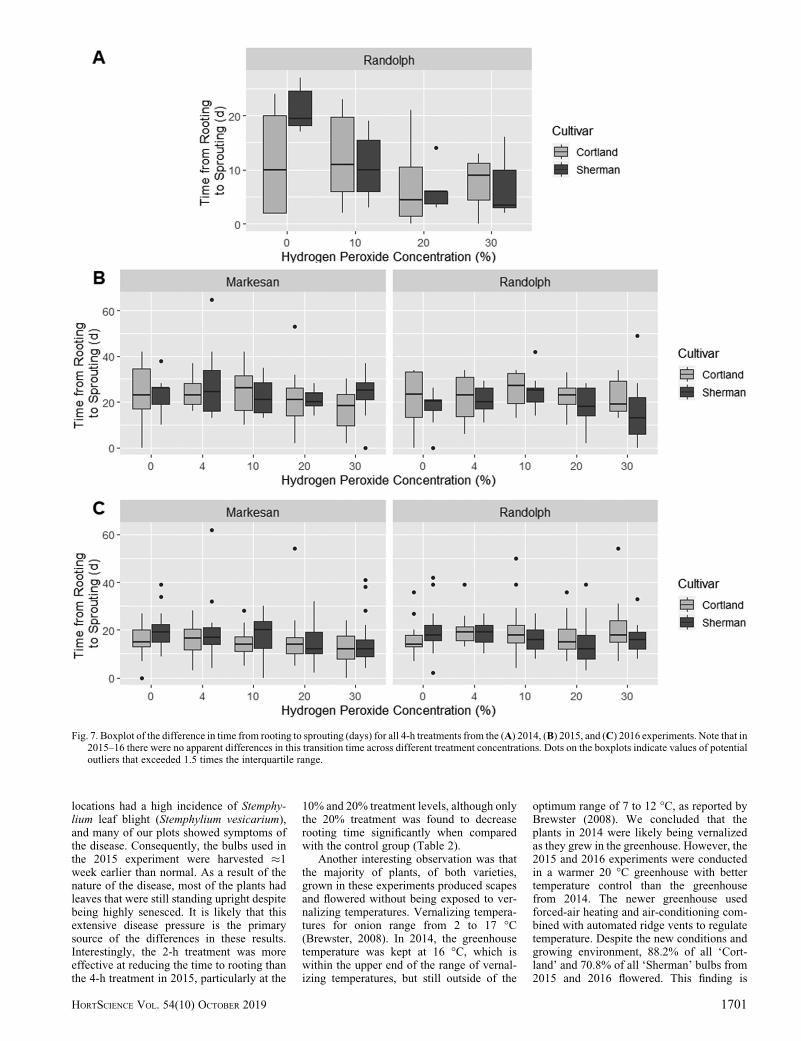
The 2014 and 2016 experiments foundthat ‘Cortland’ and ‘Sherman’ bulbs treatedwith 10%, 20%, and 30% solutions of hydro-gen peroxide exhibited expedited rooting andsprouting when compared with the 0% con-trol group (Figs. 2–7). For 2016, the 20%treatment resulted in a 61.3% average re-duction in the time to rooting. These datashow that the 20% hydrogen peroxide treat-ment was associated with the lowest meantime to rooting and sprouting, with littlespread (Figs. 2, 5, and 6). Except for ‘Cort-land’ bulbs harvested from Markesan in2015, all varieties and treatment durationsshowed fast, uniform rooting after the 20%treatment. The sprouting data show a sim-ilar trend, although the spread of the datafor each treatment group is larger than forroot development. There was no apparentdifference in time from rooting to sproutingacross treatment groups in 2015 and 2016,although a weak negative trend was ob-served in 2014 as the treatment concentra-tion increased (Fig. 7).
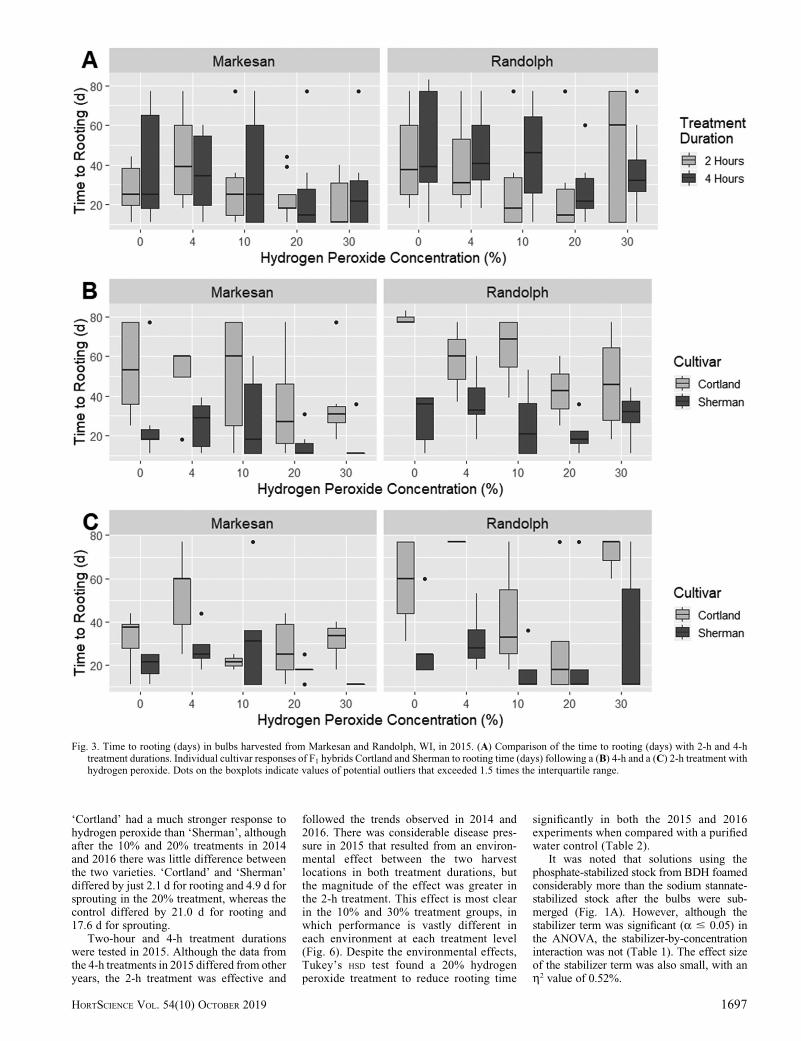
Although data from 2015 showed im-provements in the time to rooting for sometreatment groups, the trends are less clearlydefined than in 2014 and 2016 (Figs. 3 and
4). These data showed greater variances at the10%, 20%, and 30% concentration treatmentgroups for time to rooting than in 2014 and2016 (Fig. 3). In addition, there is a reducedeffect on the mean time to sprouting across alltreatment groups when compared with the 0%control (Fig. 4). In both ‘Cortland’ and ‘Sher-man’, a 4% treatment, regardless of duration,increased the time to sprouting compared withthe control. Similarly, the 30% treatment in2015 resulted in very poor uniformity in the
time to rooting, particularly in bulbs harvestedfrom Randolph.
‘Cortland’ and ‘Sherman’ bulbs averagedrooting in 11.1 d and sprouting in 28.6 dacross two environments after a 4-h treatmentwith 20% hydrogen peroxide in 2016. Thereis a strong varietal main effect in the time torooting and, to a lesser degree, in the time tosprouting (Table 1). The greatest differencebetween the two varieties can be observed inthe control and 4% treatment groups.
Fig. 2. Effects of hydrogen peroxide on onion bulb dormancy from the initial 2014 experiment. (A)Comparison of the time to rooting (days) after treatment with hydrogen peroxide in the F1 hybridcultivars Cortland and Sherman. (B) Comparison of the time to sprouting (days) after being treatedwith hydrogen peroxide in the F1 hybrid cultivars Cortland and Sherman. Dots on the boxplots indicatevalues of potential outliers that exceeded 1.5 times the interquartile range.
Table 1. Degrees of freedom (df) and F values from an analysis of variance (ANOVA) for time to rooting(measured in days) in ‘Cortland’ and ‘Sherman’ after treatment with hydrogen peroxide solutions atfive concentrations in the 2015 and 2016 experiments.
ANOVA 2015 2016
Source df F df F
Concentration 4 5.35 *** 4 36.05 ***Cultivar 1 26.96 *** 1 32.78 ***Location 1 4.69 * 1 12.22 ***Bulb diameter 1 2.44 1 30.16 ***Stabilizer — — 1 4.16 *Replication — — 1 0.04Hours 1 2.63 — —Cultivar:location 1 0.02 1 10.76 **Residuals 171 453
*, **, ***Significant at P # 0.05, 0.01, or 0.001, respectively.— = Terms not included in the linear model for the given analysis.
1696 HORTSCIENCE VOL. 54(10) OCTOBER 2019
‘Cortland’ had a much stronger response tohydrogen peroxide than ‘Sherman’, althoughafter the 10% and 20% treatments in 2014and 2016 there was little difference betweenthe two varieties. ‘Cortland’ and ‘Sherman’differed by just 2.1 d for rooting and 4.9 d forsprouting in the 20% treatment, whereas thecontrol differed by 21.0 d for rooting and17.6 d for sprouting.
Two-hour and 4-h treatment durationswere tested in 2015. Although the data fromthe 4-h treatments in 2015 differed from otheryears, the 2-h treatment was effective and
followed the trends observed in 2014 and2016. There was considerable disease pres-sure in 2015 that resulted from an environ-mental effect between the two harvestlocations in both treatment durations, butthe magnitude of the effect was greater inthe 2-h treatment. This effect is most clearin the 10% and 30% treatment groups, inwhich performance is vastly different ineach environment at each treatment level(Fig. 6). Despite the environmental effects,Tukey’s HSD test found a 20% hydrogenperoxide treatment to reduce rooting time
significantly in both the 2015 and 2016experiments when compared with a purifiedwater control (Table 2).
It was noted that solutions using thephosphate-stabilized stock from BDH foamedconsiderably more than the sodium stannate-stabilized stock after the bulbs were sub-merged (Fig. 1A). However, although thestabilizer term was significant (a # 0.05) inthe ANOVA, the stabilizer-by-concentrationinteraction was not (Table 1). The effect sizeof the stabilizer term was also small, with anh2 value of 0.52%.
Fig. 3. Time to rooting (days) in bulbs harvested from Markesan and Randolph, WI, in 2015. (A) Comparison of the time to rooting (days) with 2-h and 4-htreatment durations. Individual cultivar responses of F1 hybrids Cortland and Sherman to rooting time (days) following a (B) 4-h and a (C) 2-h treatment withhydrogen peroxide. Dots on the boxplots indicate values of potential outliers that exceeded 1.5 times the interquartile range.
HORTSCIENCE VOL. 54(10) OCTOBER 2019 1697

Bulb diameter exhibited a significant effecton time to rooting in 2016, but not in 2015. Theh2 value indicates that bulb diameter onlyaccounted for 3.4% of the variance in the timeto rooting, which is likely a result, in part, ofthe uniformity of the bulbs in the experiment.The mean bulb diameter was 68.1 ± 7.5 mm in2016 and 72.6 ± 8.1 mm in 2015. The 2016data revealed that each additionalmillimeter inbulb diameter resulted in a 2.8% averagedecrease in time to rooting.
Several of the bulbs in all treatment groupsas well as the control sprouted before de-veloping roots. This was not observed in
2014, but was found in 3.9% of the bulbs thatsprouted in 2015 and in 1.1% of the bulbs thatsprouted in 2016. In some cases, root initiationbegan within several days of sprouting, butothers produced roots after several weeks, if atall. There was no pattern to suggest thattreatment concentration increases the likeli-hood of this phenomenon, but there was a clearvarietal and stabilizer effect. A total of 78.6%of the bulbs in this group were from thecultivar Sherman and 93.3% were treated withsodium stannate-stabilized hydrogen peroxide.
Many of the plants grown in this studyflowered without vernalization, which was an
unexpected finding. A total of 86.1% of all‘Cortland’ and 70.2% of all ‘Sherman’ bulbsgrown in this study that sprouted also producedscapes, without being grown at vernalizingtemperatures. This occurred in all 3 experimen-tal years at high percentages. Excluding thedata from 2014, which used slightlycooler greenhouse temperatures, the in-cidence of scape emergence increases to88.2% in all ‘Cortland’ and 70.8% in all‘Sherman’ bulbs. In 2016, on average,scapes emerged from ‘Cortland’ bulbs in121.0 ± 19.4 d and from ‘Sherman’ bulbsin 128.9 ± 18.0 d.
Fig. 4. Time to sprouting (days) in bulbs harvested from Markesan and Randolph, WI, in 2015. (A) Comparison of the time to sprouting (days) with 2-h and 4-htreatment durations. Individual cultivar responses of F1 hybrids Cortland and Sherman to sprouting time (days) following a (B) 4-h and a (C) 2-h treatmentwith hydrogen peroxide. Dots on the boxplots indicate values of potential outliers that exceeded 1.5 times the interquartile range.
1698 HORTSCIENCE VOL. 54(10) OCTOBER 2019

Discussion
The presence of endodormancy in onionbulbs delays visible growth until dormancy isbroken. Endodormancy is usually broken inonion bulbs after chilling for several weeks,
but in the absence of chilling, dormancybreak and initiation of visible growth is lessuniform. As observed in this study, there areboth varietal and environmental effects thataffect the time to rooting and sprouting(Figs. 5–7). For growth in controlled envi-
ronments where uniformity is desirable, dif-ferences in growth from year to year andacross genotypes presents a challenge. Aprimary goal of this study was to determinewhether an exogenous application of hydro-gen peroxide to endodormant onion bulbs
Table 2. Tukey’s honestly significant difference test results for time to rooting in the 2015 and 2016 experiments. Each treatment concentration is compared todetermine whether the difference in rooting time is significantly different from zero. Natural logarithmic transformations of the rooting time data were used inthese tests.
Comparison ofconcn treatments
2015 2016
Difference P value Significance Difference P value Significance
4% to 0% 0.0838 0.9740 –0.1655 0.565210% to 0% –0.1811 0.6729 –0.8260 0.0000 ***20% to 0% –0.4712 0.0054 ** –1.0165 0.0000 ***30% to 0% –0.2860 0.1896 –0.8208 0.0000 ***10% to 4% –0.2649 0.3475 –0.6605 0.0000 ***20% to 4% –0.5550 0.0012 ** –0.8509 0.0000 ***30% to 4% –0.3698 0.0611 –0.6553 0.0000 ***20% to 10% –0.2901 0.2302 –0.1904 0.430330% to 10% –0.1049 0.9374 0.0053 1.000030% to 20% 0.1852 0.6357 0.1957 0.4253
**, ***Significant at P # 0.01 or 0.001, respectively.
Fig. 5. Time to rooting (days) in bulbs harvested from Markesan and Randolph, WI, in 2016. (A) Comparison of the time to rooting (days) with phosphate andsodium stannate-stabilized hydrogen peroxide stock solutions across both cultivars. (B) Individual cultivar responses of F1 hybrids Cortland and Sherman torooting time following a 4-h treatment with hydrogen peroxide. Note the difference in rooting time response across the two locations at lower treatmentconcentrations. Dots on the boxplots indicate values of potential outliers that exceeded 1.5 times the interquartile range.
HORTSCIENCE VOL. 54(10) OCTOBER 2019 1699

would break dormancy and elicit uniformgrowth when planted in a greenhouse.
Throughout the 3 years during which thisstudy was conducted, we found soakingdormant bulbs in a 20% hydrogen peroxidesolution for 2 to 4 h to be an effective methodfor breaking dormancy (Table 2). Treatingwith a 20% solution reduced the time torooting and improved the uniformity of initialroot development, which signified that dor-mancy was broken (Lang et al., 1987). Wealso found that the time to sprouting isdecreased with a 20% hydrogen peroxidetreatment; however, this finding was attrib-uted primarily to the treatment’s effect onroot development because there was noapparent difference in the time from rootingto sprouting between the control and any ofthe treatment groups in 2015 and 2016(Fig. 7). By breaking dormancy and initiatinggrowth earlier, these plants appear to besprouting faster, but are progressing fromrooting to sprouting at the same rate as thecontrol.
The results from 2014 and 2016 demon-strate the effectiveness of the 20% treatmentonminimizing varietal differences in the timeto root development (Figs. 2 and 5). Thedifference in mean time to rooting between‘Cortland’ and ‘Sherman’ in 2016 was 21.0 din the control, but was reduced to 2.1 d in the20% treatment group. ‘Sherman’ is generallyconsidered to be earlier maturing than ‘Cort-land’, so the difference in rooting timebetween the two varieties in the control groupwas consistent with our expectations. How-ever, these results suggest that this treatmentmay break dormancy synchronously acrossa range of long-day genotypes.
The 2015 results were a departure fromthe very clear effects of the 10% to 30%treatment groups observed in 2014 and 2016.Although many of the trends that wereidentified in the other years are still presentin the 2015 data, they were less clearlydefined in the 4-h treatment and were mutedin the 2-h treatment. The experimental designand methods were the same between 2014
and 2015, only expanded to include a secondlocation and two treatment durations. How-ever, there were two factors that could havecontributed to these differences in results.The first was that a different brand of hydro-gen peroxide was used in 2015 than in 2014.At the time, we were unaware that hydrogenperoxide solutions were stabilized and thatthe stabilizing compound differed by manu-facturer. After discovering that each stocksolution used a different stabilizer, bothproducts were included in the 2016 experi-ment. Despite the significance of the stabi-lizer term in the ANOVA (Table 1), a paired ttest found that the difference between stabi-lizers for rooting time in the 10%, 20%, and30% treatment groups was not significant atthe a = 0.05 level. This suggests that the useof differentially stabilized hydrogen peroxidesolutions between 2014 and 2015 was likelynot a major contributor to the difference inresults. The second factor worth consideringis that there was high disease pressure at bothRandolph and Markesan in 2015. Both
Fig. 6. Time to sprouting (days) in bulbs harvested fromMarkesan and Randolph,WI, in 2016. (A) Comparison of the time to sprouting (days) with phosphate andsodium stannate-stabilized hydrogen peroxide stock solutions across both cultivars. (B) Individual cultivar responses of F1 hybrids Cortland and Sherman tosprouting time following a 4-h treatment with hydrogen peroxide. Dots on the boxplots indicate values of potential outliers that exceeded 1.5 times theinterquartile range.
1700 HORTSCIENCE VOL. 54(10) OCTOBER 2019
locations had a high incidence of Stemphy-lium leaf blight (Stemphylium vesicarium),and many of our plots showed symptoms ofthe disease. Consequently, the bulbs used inthe 2015 experiment were harvested �1week earlier than normal. As a result of thenature of the disease, most of the plants hadleaves that were still standing upright despitebeing highly senesced. It is likely that thisextensive disease pressure is the primarysource of the differences in these results.Interestingly, the 2-h treatment was moreeffective at reducing the time to rooting thanthe 4-h treatment in 2015, particularly at the
10% and 20% treatment levels, although onlythe 20% treatment was found to decreaserooting time significantly when comparedwith the control group (Table 2).
Another interesting observation was thatthe majority of plants, of both varieties,grown in these experiments produced scapesand flowered without being exposed to ver-nalizing temperatures. Vernalizing tempera-tures for onion range from 2 to 17 �C(Brewster, 2008). In 2014, the greenhousetemperature was kept at 16 �C, which iswithin the upper end of the range of vernal-izing temperatures, but still outside of the
optimum range of 7 to 12 �C, as reported byBrewster (2008). We concluded that theplants in 2014 were likely being vernalizedas they grew in the greenhouse. However, the2015 and 2016 experiments were conductedin a warmer 20 �C greenhouse with bettertemperature control than the greenhousefrom 2014. The newer greenhouse usedforced-air heating and air-conditioning com-bined with automated ridge vents to regulatetemperature. Despite the new conditions andgrowing environment, 88.2% of all ‘Cort-land’ and 70.8% of all ‘Sherman’ bulbs from2015 and 2016 flowered. This finding is
Fig. 7. Boxplot of the difference in time from rooting to sprouting (days) for all 4-h treatments from the (A) 2014, (B) 2015, and (C) 2016 experiments. Note that in2015–16 there were no apparent differences in this transition time across different treatment concentrations. Dots on the boxplots indicate values of potentialoutliers that exceeded 1.5 times the interquartile range.
HORTSCIENCE VOL. 54(10) OCTOBER 2019 1701

deserving of further investigation. If vernal-ization in long-day onion is facultative, ratherthan obligate, procedures might be designedto reduce the time necessary to complete theonion plant’s life cycle (D’Angelo and Gold-man, 2018). This, in turn, would hasten seedproduction and many breeding methods.
The 30% treatment effectively reducedthe rooting time to levels that were compa-rable to the 10% and 20% treatments in 2014and 2016. However, across all years, �20%of the bulbs in the 30% treatment became softand rotten. The rot often also included moldthat grew on the cut surface of the bulb andusually resulted in plant death or stuntedgrowth. The 10% treatments were effectiveat reducing the rooting time in 2014, 2016,and in the 2015 2-h treatment group, but weresensitive to environmental effects and possi-bly genotype-by-environment interactions(Figs. 3C and 5B). In the 2-h 10% treatmentgroup from 2015, there was a change in rankfor rooting time between ‘Cortland’ and‘Sherman’ over the two harvest locations:Markesan and Randolph. The 20% treatmentconsistently had the best performance acrossyears, harvest locations, treatment durations,and in each of the varieties. In addition, the20% treatment had the effect of breakingdormancy in ‘Cortland’ and ‘Sherman’ within2.1 d of each other, on average. We havefound the 20% treatment to be an effective,versatile tool for breaking dormancy inonion bulbs.
The ability to break dormancy in bulbsuniformly is an important advance for onionresearch. The novel method we are pro-posing offers researchers greater controlover the timing and duration of treatmentsto onions grown from bulbs and may aid inoffering insight to the molecular control ofdormancy release. In addition, it has poten-tial for synchronizing growth in diversegermplasm, which can be beneficial in thecontext of research, breeding, and seedproduction. There is an economic interestin synchronous growth in the seed industrybecause asynchronous growth affects seed
yield negatively and limits cross-pollinationsto plants at a similar reproductive maturity.This method may also prove to be a usefultool in the development of an annual-cyclebreeding system that does not rely onseedling vernalization. Depending on therelationship between vernalization anddormancy, if nondormant bulbs can vernal-ize at a faster rate than dormant bulbs, asreported by Bertaud (1988) and Shishidoand Saito (1977), then rapid, uniform dor-mancy release may be key in solving theannualization puzzle. Further research intothe effects of treatment duration and theimpact of high disease pressure on theefficacy of these treatments is necessary.
Literature Cited
Bertaud, D.S. 1988. Effects of chilling duration,photoperiod and temperature on floral initiationand development in sprouted and unsproutedonion bulbs, p. 254–261. In: Symposium onAllium: EUCARPIA - Section Vegetables, 4thCoventry 6–9 Sept. 1988 United Kingdom GB.
Brewster, J.L. 2008. Onions and other vegetablealliums. 2nd ed. Crop production science inhorticulture 15. CAB International, Walling-ford, UK.
Chope, G.A., K. Cools, J.P. Hammond, A.J.Thompson, and L.A. Terry. 2012a. Physiolog-ical, biochemical and transcriptional analysisof onion bulbs during storage. Ann. Bot.109:819–831.
Chope, G.A., K. Cools, L.A. Terry, J.P. Hammond,and A.J. Thompson. 2012b. Association of geneexpression data with dormancy and sprout sup-pression in onion bulbs using a newly developedonion microarray. Acta Hort. 969:169–174.
D’Angelo, C.J. and I.L. Goldman. 2018. Temporalaspects of vernalization and flowering in longday storage onion. J. Amer. Soc. Hort. Sci.43:446–453.
El-Maarouf-Bouteau, H. and C. Bailly. 2008. Oxi-dative signaling in seed germination and dor-mancy. Plant Signal. Behav. 3:175–182.
FAO. 2016. FAOSTAT crops data. 4 Feb. 2018.<http://www.fao.org/faostat/en/#data/QC>.
Horvath, D.P., J.V. Anderson, W.S. Chao, andM.E. Foley. 2003. Knowing when to grow:Signals regulating bud dormancy. Trends PlantSci. 8:534–540.
Lang, G.A., J.D. Early, G.C.Martin, andR.L. Darnell.1987. Endo-, para-, and eco-dormancy: Physio-logical terminology and classification for dor-mancy research. HortScience 3:371–377.
Liu, X., Z. Deng, H. Cheng, X. He, and S. Song.2011. Nitrite, sodium nitroprusside, potassiumferricyanide and hydrogen peroxide releasedormancy of Amaranthus retroflexus seeds ina nitric oxide-dependent manner. Plant GrowthRegulat. 64:155–161.
Mazzitelli, L., R.D. Hancock, S. Haupt, P.G.Walker, S.D.A. Pont, J. McNicol, L. Cardle,J. Morris, R. Viola, R. Brennan, P.E. Hedley,and M.A. Taylor. 2007. Co-ordinated geneexpression during phases of dormancy releasein raspberry (Rubus idaeus L.) buds. J. Expt.Bot. 58:1035–1045.
Mohamed, H.B., A.M. Vadel, J.M.C. Geuns, andH. Khemira. 2012. Effects of hydrogen cyana-mide on antioxidant enzymes’ activity, prolineand polyamine contents during bud dormancyrelease in Superior Seedless grapevine buds.Acta Physiol. Plant. 34:429–437.
N�ee, G., Y. Xiang, and W.J. Soppe. 2017. Therelease of dormancy, a wake-up call for seeds togerminate. Curr. Opin. Plant Biol. 35:8–14.
Oracz, K., H.E. Bouteau, J.M. Farrant, K. Cooper,M. Belghazi, C. Job, D. Job, F. Corbineau, andC. Bailly. 2007. ROS production and proteinoxidation as a novel mechanism for seeddormancy alleviation. Plant J. 50:452–465.
Pak, C., L.H.W. van der Plas, and A.D. de Boer.1995. Importance of dormancy and sink strengthin sprouting of onions (Allium cepa) duringstorage. Physiol. Plant. 94:277–283.
P�erez, F.J. and W. Lira. 2005. Possible role ofcatalase in post-dormancy bud break in grape-vines. J. Plant Physiol. 162:301–308.
R Development Core Team. 2018. R: A language andenvironment for statistical computing.RFoundationfor Statistical Computing. Vienna, Austria. 13 July2019. <https://www.R-project.org>.
Shishido, Y. and T. Saito. 1977. Studies on theflower bud formation in onion plants III.Effects of physiological conditions on the lowtemperature induction of flower buds in bulbs.J. Jpn. Soc. Hort. Sci. 46:310–316.
Vergara, R. and F.J. P�erez. 2010. Similaritiesbetween natural and chemically induced bud-endodormancy release in grapevine Vitis vinif-era L. Scientia Hort. 125:648–653.
1702 HORTSCIENCE VOL. 54(10) OCTOBER 2019