bones, teeth and genes
TRANSCRIPT

BONES, TEETH AND GENES: A Genomic Homage to HarrySicher’s “Axial Movement of Teeth”
Sean Holliday1,2, Bernard Schneider1, Maria Therese S. Galang1,2, Tadayoshi Fukui3, AkiraYamane4, Xianghong Luan1,2, and Thomas G.H. Diekwisch2,1,*1 Department of Orthodontics, UIC College of Dentistry, Chicago, IL, U.S.A2 Department of Oral Biology, UIC College of Dentistry, Chicago, IL, U.S.A3 Department of Orthodontics, Tsurumi University School of Dental Medicine, Yokohama, Japan4 Department of Pharmacology, Tsurumi University School of Dental Medicine, Yokohama, Japan
AbstractAims—We have used the model of the un-opposed rodent molar to study morphologic and geneticmechanisms of tooth eruption.
Methods—Left maxillary molar teeth of 12-day old Swiss-Webster mice were extracted underanesthesia and mandibular molars were allowed to super-erupt. To trace areas of tissue remodelingand to determine areas of new tissue formation, mice were injected with fluorescent dyes,tetracycline, alizarin red, and calcein blue. Subsequent to sacrifice, mandible tissue blocks wereprepared for ultrathin ground sections, fluorescent microscopy, and von Kossa’s mineral detectionprocedure. A second set of specimen was prepared for RNA extraction and microarray analysis.
Results—Our data established significant eruption of first and second mandibular mouse molarsfollowing complete extraction of antagonists by 0.13 mm after 12 days. Labeled tissue sectionsrevealed significant amounts of new bone and cementum apposition on the un-opposed sidecompared to the control side as revealed by fluorescent markers and ultrathin ground sections.Microarray transcript level comparisons between the experimental and the control groupdemonstrated significant (more than 2-fold) increase in gene expression of elastin and tenascin Cextracellular matrix proteins; brevican, lumican, and biglycan proteoglycans; as well as fibroblastgrowth factor 9.
Conclusion—In this study we have established the un-opposed mouse molar as a model to studytissue dynamics during the axial movement of teeth. Our data indicated significant new formationof bone and cementum in tandem with increased expression of extracellular matrix-related genes.
IntroductionIn a series of landmark papers between 1941 and 1944 Harry Sicher and Joseph-PeterWeinmann established the biological foundation for tooth movement as a consequence of bothroot cementum and bone growth 1,2,3,4. Prior to Sicher and Weinmann’s pioneering work, theorthodontic theory of tooth movement suggested that bone behaves as a passive tissue duringtooth movement 5, 6. It was on the basis of Sicher and Weinmann’s radiographic evidence andalizarin-labeling studies that dynamic changes in bone morphology in the periphery of teeth
*Author for Correspondence: Thomas G.H. Diekwisch, D.D.S., Ph.D., Professor and Head, Department of Oral Biology, Allan G. BrodieSr. Endowed Chair for Orthodontic Research, Director, Allan G. Brodie Laboratory for Craniofacial Genetics, UIC College of Dentistry,801 South Paulina, Chicago, IL 60612.
NIH Public AccessAuthor ManuscriptWorld J Orthod. Author manuscript; available in PMC 2009 July 23.
Published in final edited form as:World J Orthod. 2005 ; 6(1): 61–70.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

during physiological movement were documented 4. From their studies they concluded:“Developmental movements of tooth germs, eruptive movements of teeth, and physiologicmovements of teeth during their functional period are the result of differential growth of thetooth and the surrounding bone” 4. In their classic textbook “Bone and Bones”, the authorswent even one step further and argued for a superior role of bone growth over tooth growth:“While the dependence of bone growth upon the development of teeth is slight, the reverserelation, that is, the dependence of tooth development and, especially, tooth eruption upongrowth of bone and bones, is considerable” 7. The establishment of a dynamic relationshipbetween bone growth and tooth movement became the dawn of a new era of orthodontics: theera of biological research.
Sicher and Weinmann retained their interest in understanding the true mechanisms of toothmovement throughout their long and distinguished careers. One of the issues of debate at thattime was the mechanism behind the drift of the dentition since it had been documented that inhumans the drift was directed mesially while in rodents (e.g. rats and mice) the molars hadbeen shown to drift distally. Sicher and Weinmann were interested in finding out whetherexternal forces such as occlusion and bite force or intrinsic genetic mechanisms wereresponsible for the physiologic drift of the dentition. In order to address this question, theyproposed the model of the un-opposed molar as a means to determine the forces behind thedrift of the rodent dentition that was then tested by their colleague Julia Meyer and a graduatestudent 8. Briefly, Sicher and Weinmann suggested removing the upper molars unilaterally,and thus allowing the lower molars to erupt without an antagonist. Their model was based onprevious studies, in which it had been demonstrated that rodent molars erupt continuouslythroughout life 4. Sicher and Weinmann believed that if molars continued to drift in the absenceof antagonists, the forces behind the drift of the dentition were intrinsic and genetic 4. Insubsequent studies their model was validated and rat molars were documented to drift distallyin the absence of an opposing occlusion8, 9, 10. Moreover, Sicher and Weinmann’s modelproved to be a useful tool not only to study the drift of the dentition, but also to study otheraspects of tissue remodeling during axial tooth movement 8, 9, 10.
In the current study we have returned to Sicher and Weinmann’s historic model of the un-opposed rodent molar. Moreover, we have decided to re-visit their original question about themechanisms behind physiological tooth movement. However, we have employed modernpreparation and imaging techniques as well as state-of-the-art microarray genomics to oncemore ask the question as to which factors and forces might be involved in physiological toothmovement. Our data not only confirm Sicher and Weinmann’s original findings, but also offera genomic explanation for the processes that might lead to orthodontic tooth movement.
Materials and MethodsExperimental model and tissue preparation
A total of forty 35-day old Swiss-Webster mice were acquired and separated into variousexperimental groups. Prior to unilateral extraction of maxillary molars, mice were anesthetizedusing Ketamine (100mg/kg) and Xylazine (5mg/kg). Following anesthesia, all three left,maxillary molars were extracted (Figure 2). Then, pressure was applied to the area to achievehemostasis. Buprenorphine was administered as an analgesic subcutaneously at a dose of 0.05mg/kg before ending the procedure. Doses of analgesic were administered every 12 hours forup to two days post-procedure. The right side served as the control side, while the left side wasthe experimental side of each mouse. The mice were weighed daily to ensure they were thriving(Figure 3).
Holliday et al. Page 2
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Tissue processingA group of 15 mice received an intraperitoneal injection of tetracycline hydrochloride (50 mg/kg) as a vital stain at the time of extraction. They also received a vital stain of alizarin red (25mg/kg) at 4 days, and calcein blue (30 mg/kg) at 8 days. They were sacrificed at day 12 byusing CO2 gas, and stored in 70% ethanol for fixation overnight. The tissues were placed inincreasing concentrations of ethanol, followed by acetone, and then infiltrated with resin(Technovit 2000®, EXAKT Technologies Inc.). This sample was embedded and prepared intoundecalcified ground sections for fluorescent microscopy.
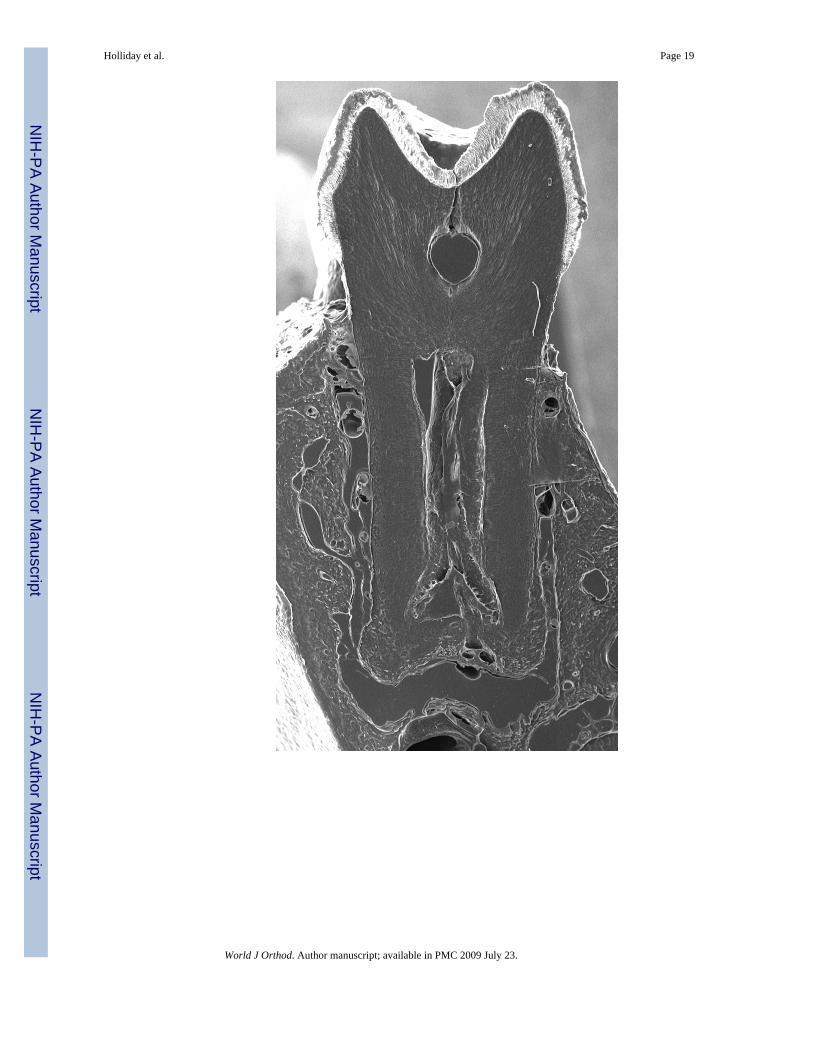
Scanning Electron microscopyFour mouse skulls were skeletonized by removing all soft tissue. Two skulls were from micethat had no teeth extracted, serving as controls. Another two mice had their maxillary first,second, and third molars extracted. Both groups were sacrificed after 12 days of theexperimental mandibular molars being in an un-opposed situation. Their mandibles weremounted into Technovit 2000® (EXAKT Technologies, Inc.) blocks and sectioned in atransverse plane across their first molars (Figure 4). These sections were subsequently preparedfor viewing via scanning electron microscopy (SEM). A layer of 3 nanometers of platinum-palladium coating was applied to each sample using a Cressington sputter coater 208HR® anda Cressington Thickness controller mtm20® to prepare for scanning electron microscopy. Thesamples were viewed under variable pressure with the Hitachi S-3000N VP® scanning electronmicroscope.
Von Kossa’s staining for mineralized calciumOriginally designed as a technique to detect inorganic phosphates via silver nitrate, von Kossa’stechnique has found wide acceptance as a mineralized tissue marker. For this technique,mandibles from 12day-unopposed molars and controls were dissected, fixed in 10% bufferedformalin, de-hydrated, and processed for paraffin sectioning. Tissue sections were treated witha 5% silver nitrate solution as previously described11. During subsequent four hours of sunlightexpose, calcium was reduced and replaced by metallic silver deposits, which appear blackunder the microscope.
Microarray technologyFor microarray analysis, five mice were sacrificed after molars were subjected to an un-opposedsituation for 6 days. Root apices from un-opposed molars were removed and stored in liquidnitrogen. Another five mice were euthanized at day zero. Control root apices were collectedand immediately placed in liquid nitrogen. Subsequently, mRNA was extracted from root apextissues using TRIZOL® LS Reagent (Invitrogen ™ life technologies) according to themanufacturer’s instructions. For transcript screening, we used the AtlasTM Mouse 1.2 Array(Clontech, Palo Alto, CA). The PolyA+ RNA was enriched using magnetic beads (Clontech,Palo Alto, CA) and used to generate cDNA probes via MMLV reverse transcriptase with 32P-dATP and random Atlas array specific primers. cDNA from mouse molar apex regions wasgenerated using the SMART™ cDNA technology according to the manufacturer’s instruction(Clontech, Palo Alto, CA). DNA targets on the arrays were located using grid overlays andspot intensities were measured using AtlasImage 1.5 software. Relative levels of geneexpression between arrays were calculated by dividing the normalized intensities of spots onone array by normalized spot intensities on a second array. Spots were normalized to all ninehouse keeping genes provided on the Atlas 1.2 array.
Holliday et al. Page 3
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ResultsOn the un-opposed side, molars erupted 0.35mm above the occlusal plane
The left mandibular mouse molars were kept in an unopposed situation for 12 days. Following12 days, mandibles were skeletonized and the difference between molar heights in the un-opposed and opposed situation was determined (Figure 1). Molar height in the un-opposedmodel was compared with molar height in a control group in which opposing molars maintainedocclusion. The molars on the experimental side (i.e. the un-opposed side) exceeded the controlside (i.e. the opposed side) by 0.13 mm (Figure 1).
Change in mouse weight after extraction of maxillary molarsMonitoring the weight of the mice was done daily to evaluate the effects of tooth extractionson eating habits. The experimental mice did thrive, despite the intra-oral surgery, and the lackof occlusion on one side. They showed a small drop in weight of 4 grams +/− 2.5 grams (lessthan 15% of their total body weight), during the initial period of wound healing. They startedto gain weight again after 2 days, reaching their starting weights within the experimental period(Figure 3).
37% Increase in un-opposed molar cellular cementum area compared to control molarsScanning electron microscopy (SEM) was used to view transverse sections of skeletonizedmandibles at the first molar. SEM revealed that the area of cellular cementum was increasedat the root apex of the un-opposed molar by 37% (Figure 5).
Significant increase of cellular cementum and alveolar bone thickness at the un-opposedmolar tooth apex as revealed by vital stain labeling
During the 12 days in which mouse molars were kept in an un-opposed situation, fluorescentdyes were injected at selected time intervals to mark significant events of tissue remodeling atpre-selected time points.
Following each of 4, 8, and 12 day intervals, significant amounts of new cementum and alveolarbone apposition were detected at the apical tip of the root and its corresponding alveolar bonesurface. The experimental apposition rates of bone and cementum were approximately threetimes that of the control side. In all stages of mineralized tissue remodeling, the mean differencebetween the experimental and control sides was shown to be significant. The analysis of thedata documented that differences between mineral apposition rates were significant betweengroups while there was no significant change in the periodontal ligament space dimensions.
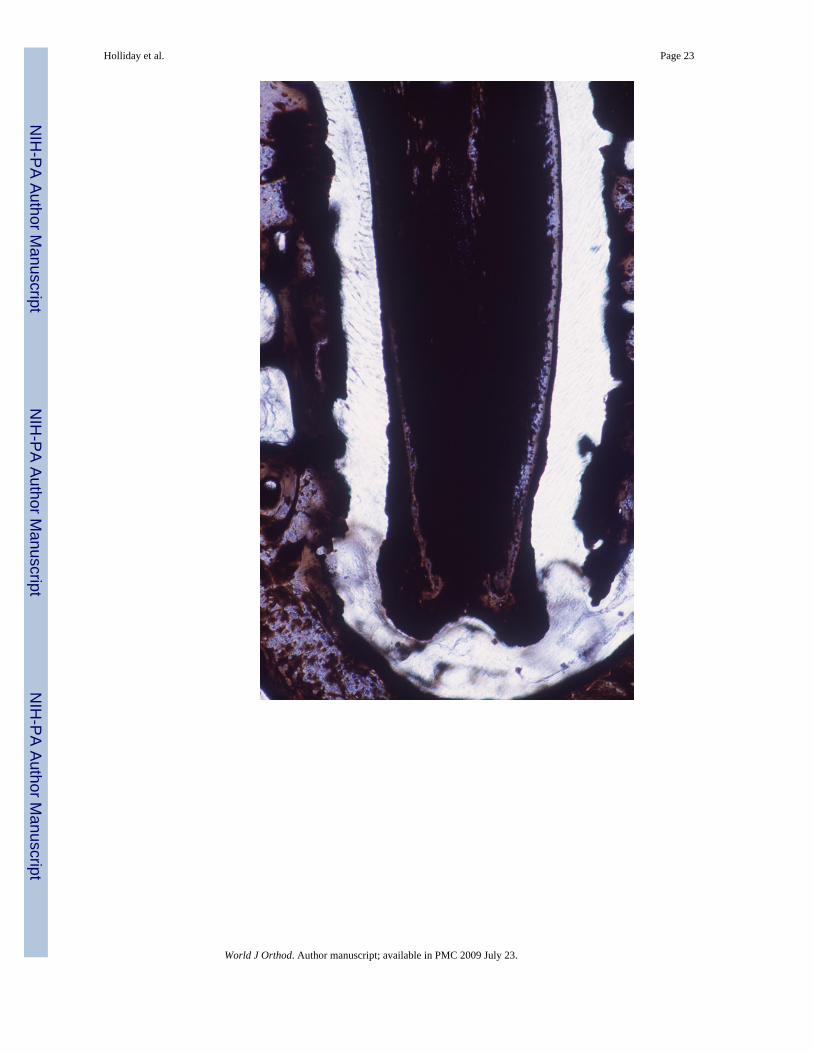
New mineralized tissue formation at the apex of un-opposed molars demonstrated by vonKossa’s method
Inorganic phosphates as common constituents of mineralized tissues were labeled using vonKossa’s procedure and apical cementum layers were compared between the control side andthe un-opposed molar (Figs. 7A and B). The cementum layer of the unopposed molar root wassignificantly greater in diameter than the control molar cementum layer and completelyblackened using von Kossa’s method (Fig. 7B). At high magnification, stages of intracellularcementoblast mineralization at the cementoblast/periodontal ligament interface were revealed,while the ligament itself was devoid of phosphate precipitates (Fig. 7C).
Genomic analysis revealed significant changes in extracellular matrix gene expressionduring tooth eruption
Extracellular matrix microarray analysis demonstrated significantly (> 2-fold) increased levelsof elastin and tenascin C extracellular matrix proteins; brevican, lumican, and biglycan
Holliday et al. Page 4
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

proteoglycans; as well as fibroblast growth factor 9 in the experimental group versus thecontrol. In contrast, the laminin expression was greatly reduced, as were the proteases cathepsinD and MMP11 (Table 1).
DiscussionThe model system of the un-opposed rodent molar has been suggested by Harry Sicher andJoseph-Peter Weinmann to document directions of tooth drift during craniofacial growth anddevelopment. We have used this model to determine tissue dynamics and genomic eventsduring vertical tooth movements in the adult. Our results have confirmed previous findings ontooth eruption and drift of the dentition, but added cellular detail and genomic analysis.Together, our findings have validated the model of the un-opposed mouse molar as a meansto study tooth movement. The model of the unopposed mouse molar has allowed us to re-visitquestions of axial movement of teeth that have been originally asked by Sicher and Weinmannmore than 50 years ago 2, 3, 4.
In our un-opposed molars we reported a 0.13mm over-eruption above the occlusal plane 12days past extraction of antagonists. Our results were based on skeletal preparations of jaws thathad been either subjected to extraction of antagonizing molars or remained in occlusion forcontrol study purposes. Our findings of continued eruption after the tooth had reached the planeof occlusion are supported by previous studies using the un-opposed molar model in rats 8, 9,10. The issue of super-eruption after the tooth had reached the plane of occlusion had been thetopic of a historic debate between Bernhard Gottlieb and Oskar Weski, the later of the twoclaiming that super-eruption did not occur once the tooth had reached the plane of occlusion12. Our findings confirm Gottlieb’s original view of continuous eruption throughout the life ofthe tooth, a view that was shared in Sicher and Weinmann’s model of the un-opposed rodentmolar. We have assessed and verified our findings of significant over-eruption of un-opposedmolars using a variety of techniques, including morphometry of skeletonized mandibles,morphometry of scanning electron micrographs, and use of fluorescent markers following vitalstain injections and ultrathin ground sections. Together, these three very different analysisstrategies have validated and confirmed each other, yielding almost identical overall values forsuper-eruption (0.13mm – 0.17mm after 12 days).
Our studies have revealed that new cementum and alveolar bone were formed at the apex ofthe un-opposed molar after 12 days. This deposition of new mineralized tissue formation seenin this study supports previous findings in the rat model 8. Other studies either reported noincrease in cementum formation 10 or a lesser degree of cementum formation 8, 9. Thesediscrepancies might be explained either by species differences between the mouse and the ratmodel or by less sophisticated imaging and labeling techniques used in those studies (e.g. leadacetate or alizarin red in combination with thick ground sections). Our analysis of scanningelectron micrographs revealing a 37% enlarged cementum area per mid-root cross section ascompared to cementum area in control sections adds additional support for our report ofincreased cementum thickness in the un-opposed experimental molars in comparison to thecontrols. Together, both techniques have clearly established the formation of new and cellularcementum at the root apex of un-opposed tooth molars.
Our data indicated that newly formed apical cellular cementum was highly mineralized.Patterns of initial cementum deposition were similar to those observed during physiologicaltooth development. We therefore argue that axial tooth movement as it is triggered in an un-opposed situation re-initiates the process of apical tissue deposition that occurs during tootheruption and root development. We further speculate that this process may have been broughtto a halt as long as occlusion with the opposing molar prevails and tissue growth is inhibited.It is only in the un-opposed situation that processes of mineralized tissue deposition are re-
Holliday et al. Page 5
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

initiated late in development. Tissue remodeling events as observed in the model of the un-opposed molar may therefore resemble physiological events of tooth eruption or toothmovement without the inhibitory event of root resorption or occluding forces. We thereforebelieve that the model of the un-opposed mouse molar is a powerful model to study geneticevents as they occur during tooth eruption and tooth movement.
In order to determine possible mechanisms by which the tissue remodeling during toothmovement might occur, we have determined changes in extracellular matrix (ECM) geneexpression profiles using microarray technology. We have focused on extracellular matrix genearrays because the extracellular matrix is at once the premier signaling environment thatcontrols and regulates changes in cell shape, adhesion, fate, and proliferation as well as thescaffold in which actual changes in tissue morphology are manifested. Following tooth-displacement in the un-opposed mouse molar, we determined significantly increased levels inthe expression of specific matrix-related genes, including elastin, tenascin C, brevican,lumican, and biglycan as well as FGF9. Other matrix-related genes and proteases such aslaminin, cathepsin D, and MMP11 were downregulated.
Together, our results indicate significant changes in the presence and transcriptional activityof extracellular matrix-related genes during the apical remodeling that occurs as a result of aloss of antagonistic tooth forces. Our results related to extracellular matrix glycoproteins arein congruence with several other studies that have been either based on gingival crevicularfluid analysis during orthodontic tooth movement13 or on simulated orthodontic forces in cellculture14,15. Interestingly, even though all three model systems are based on completelydifferent approaches toward a molecular analysis of factors involved in tooth movement,findings related to extracellular matrix gene expression yielded similar trends and conclusions.Notably, our studies confirm previous studies on the significant role of proteoglycans, e.g. thesmall proteoglycans biglycan and lumican, during orthodontic tooth movement13. Findings onbiglycan upregulation appear especially meaningful in light of biglycan’s putative role inelastic fiber formation16 and previous findings on tropoelastin upregulation after simulationof orthodontic force14. We thus speculate that synergistic actions of both elastin and biglycanlead to the formation of new elastic ligament fibers, which are comprised to 90% of elastin.However, among the extracellular matrix proteoglycans we studied, the large proteoglycanbrevican demonstrated the greatest degree of upregulation, and thus might play a significantrole in our future understanding of orthodontic tooth movement. Another extracellularglycoprotein, tenascin, that was also upregulated in our model, had been previously associatedwith responses to mechanical forces in the periodontal ligament15.
Many of the glycoproteins listed above are tightly controlled by growth factors and/or mediatethe presentation of growth factors within the extracellular matrix. One of the growth factorsthat might be associated with bone formation at this level is fibroblast growth factor 917, whichwas significantly upregulated in our study. Within the extracellular matrix, numerousmetalloproteinases are involved in the proteolytic cleavage of these growth factors and theirsubsequent access to individual cells18. Matrix metalloproteinase 11, which wasdownregulated in our study, might only be one of many proteases involved in the equilibriumof factors, MMPs, and TIMPs, which contribute to extracellular matrix mediated cell growthand migration during tooth movement. Other proteases, such as cathepsin D, might be moretightly associated with the function of specific cells such as odontoclasts19. Cathepsin Ddownregulation, as observed in our model, may not only indicate an absence of bone orcementum resorption, but possibly even reflect a general downregulation of osteoclast/odontoclast activity in the absence of antagonistic molar contact. Intestingly, several majorsubunits of one of the major matrix proteins, laminin, were also downregulated, which mayindicate that laminin disintegration might play an important role to facilitate tooth movement.
Holliday et al. Page 6
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ConclusionIn their classic textbook “Bone and Bones”, Sicher and Weinmann wrote a monograph aboutthe all-encompassing role of bone and bones 7. While they understood tooth movement as anexpression of the differential growth of tooth and bone 4, they nevertheless emphasized thesuperior role of bone and bones as it pertains to tooth movement 7. We have re-visited Sicherand Weinmann’s classic model of the unopposed rodent molar to determine tissue dynamicsand possible genetic mechanisms involved in the axial movement of teeth. Our resultsdocumented significant levels of super-eruption in un-opposed mouse molars, continuouslevels of cellular cementum and alveolar bone apposition during the period of super-eruption,and increased levels of elastin, tenascin, brevican, lumican, and biglycan extracellular matrix-related gene expression during axial movement of teeth. Together, our studies document thatboth alveolar bone and tooth cementum re-modeling equally contribute to tooth movement bymodulating gene expression in the extracellular matrix and cell surface. We are thereforepresenting a case for “Bones, Teeth, and Genes” as the important contributors to the movementof teeth.
AcknowledgmentsThe authors of this paper would like to dedicate this article to the memory of the late Donald and Barbara Brodie,whose generosity facilitated the creation of the Brodie Laboratory for Craniofacial Genetics in the honor of DonaldBrodie’s father, the former Dean and Orthodontic Department Head of the University of Illinois College of Dentistry,Dr. Allan G. Brodie. It were Allan G. Brodie’s tireless efforts to promote research on the “Biologic Aspects ofOrthodontia” 20 that built the foundation for a biologically oriented school of orthodontic therapy at the Universityof Illinois. Together with the contributions of his colleagues Isaac Schour, Joseph-Peter Weinmann, and Harry Sicher,Allan G. Brodie’s program in orthodontics had a profound impact on generations of students all over the world.
Funding for this study was provided by the NIH grant R01 DE15425 to TGHD, the Brodie endowment to theDepartment of Orthodontics, the research award from the Odontographic Society of Chicago, and funds from theRehak endowment to the Department of Orthodontics. Skulls were prepared by Shawn Heflick at artbyevolution.com.Additional help by Dr. Therese M. Galang and other members of the Brodie Laboratory for Craniofacial Genetics isgratefully acknowledged.
References1. Weinmann JP. Bone changes related to eruption of the teeth. Angle Orthod 1941;11:83.2. Sicher H. Tooth eruption: The axial movement of continuously growing teeth. J Dent Res 1942a;
21:201–210.3. Sicher H. Tooth eruption: Axial movement of teeth with limited growth. J Dent Res 1942b;21:395–
402.4. Sicher H, Weinmann JP. Bone growth and physiologic tooth movement. Am J Orthod 1944;30:109–
132.5. Kingsley, N. A treatise on oral deformities as a branch of mechanical surgery. D. Appleton; New York:
1880. p. 576. MacDowell, JN. Orthodontia. E. G. Colgrove; Chicago: 1901. p. 1027. Weinmann, JP.; Sicher, H. Fundamentals of Bone Biology. Mosby: St. Louis; 1944. Bone and Bones;
p. 978. Schneider B, Meyer J. Experimental studies on the interrelations of condylar growth and alveolar bone
formation. Angle Orthod 1965;35:187–199. [PubMed: 14331019]9. Cohn S. Disuse atrophy of the periodontium in mice following partial loss of function. Arch Oral Biol
1966;11:95–105. [PubMed: 5226737]10. Levy G, Maillard M. Histologic study of the effects of occlusal hypofunction following antagonist
tooth extraction in the rat. J Periodontol 1980;51:393–399. [PubMed: 6930479]11. Diekwisch TGH, Berman BJ, Gentner S, Slavkin HC. Initial enamel crystals are not spatially
associated with mineralized dentine. Cell Tiss Res 1995;279:149–167.
Holliday et al. Page 7
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

12. Carranza, F.; Shklar, G. History of Periodontology. Quintessence Publishing Co; Chicago: 2003. p.122
13. Waddington RJ, Embery G. Proteoglycans and Orthodontic Tooth Movement. J Orthod 2001;28:281–290. [PubMed: 11709593]
14. Redlich M, Roos HA, Reichenberg E, Zaks B, Mussig D, Baumert U, et al. Expression of tropoelastinin human periodontal ligament fibroblasts after simulation of orthodontic force. Archs Oral Biol2004;49:119–124.
15. Theilig C, Bernd A, Leyhausen G, Kaufmann R, Geurtsen W. Effects of mechanical force on primaryhuman fibroblasts derived from the gingiva and the periodontal ligament. J Dent Res 2001;80:1777–1780. [PubMed: 11669493]
16. Kielty CM, Sherratt MJ, Shuttleworth CA. Elastic fibres. J Cell Science 2002;115:2817–2828.[PubMed: 12082143]
17. Ornitz DM, Marie PJ. FGF signaling pathways in endochondral and intramembranous bonedevelopment and human genetic disease. Genes and Development 2002;16:1446–1465. [PubMed:12080084]
18. Chang C, Werb Z. The many faces of metalloproteases: cell growth, invasion, angiogenesis andmetastasis. Trends Cell Biol 2001;11:S37–S43. [PubMed: 11684441]
19. Gotz W, Quondamatteo F, Ragotzki A, Affeldt J, Jager A. Localization of cathepsin D in humanodontoclasts. A light and electron micrsocopical immunocytochemical study. Conn Tiss Res2000;41:185–194.
20. Brodie, AG. Biologic Aspects of Orthodontia. In: Gordon, SM., editor. Dental Science and DentalArts. Lea & Febiger; Philadelphia: 1938.
Holliday et al. Page 8
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1.
Holliday et al. Page 9
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Holliday et al. Page 10
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Holliday et al. Page 11
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Holliday et al. Page 12
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Figure 2.
Holliday et al. Page 13
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3.
Holliday et al. Page 14
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Holliday et al. Page 15
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Holliday et al. Page 16
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4.
Holliday et al. Page 17
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Holliday et al. Page 18
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Holliday et al. Page 19
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
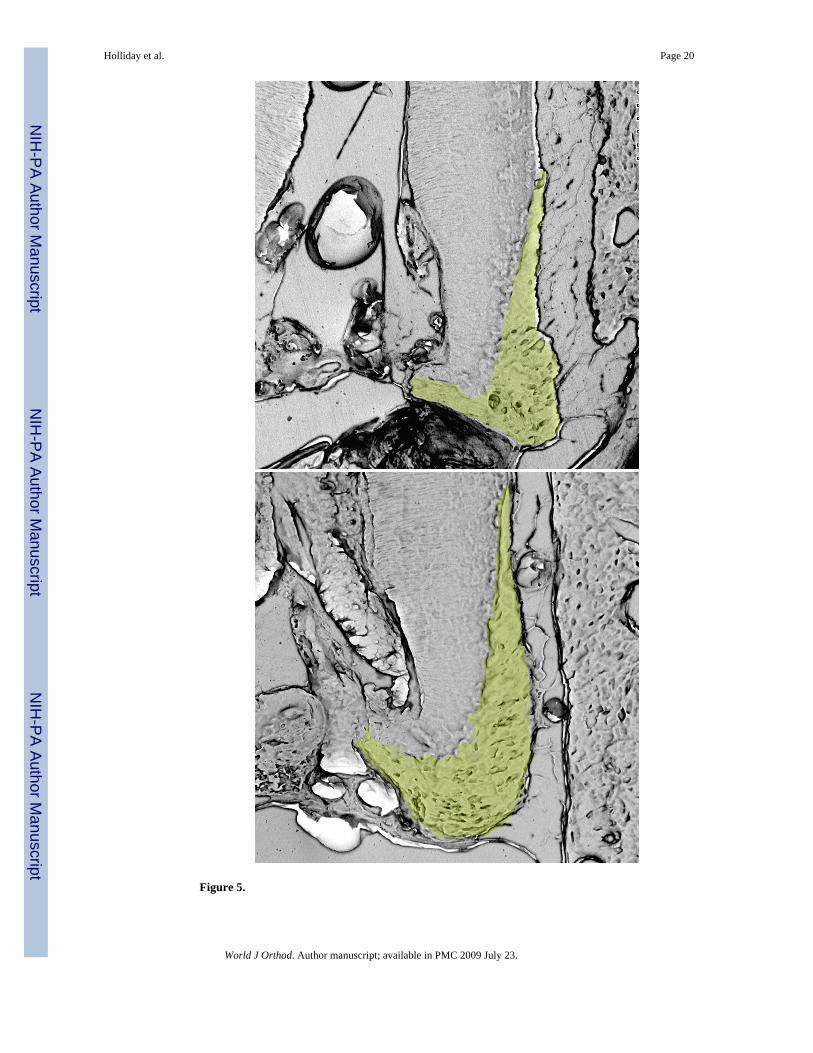
Figure 5.
Holliday et al. Page 20
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Holliday et al. Page 21
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6.
Holliday et al. Page 22
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Holliday et al. Page 23
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Holliday et al. Page 24
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
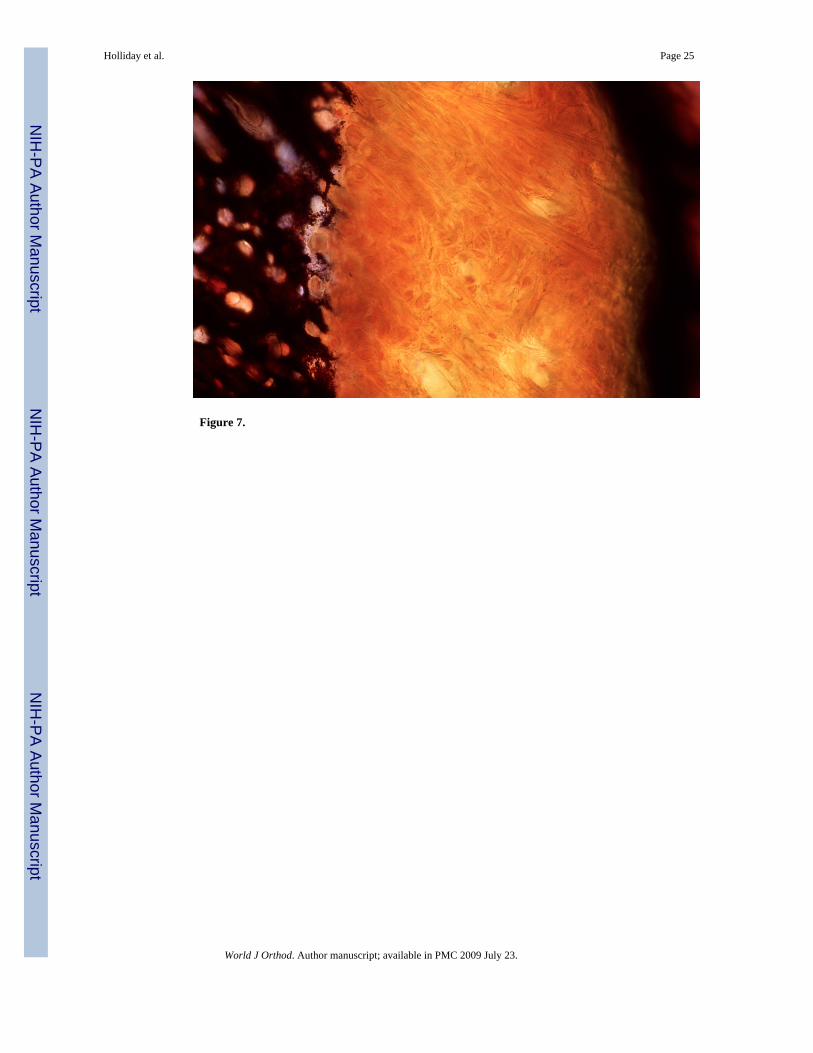
Figure 7.
Holliday et al. Page 25
World J Orthod. Author manuscript; available in PMC 2009 July 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Holliday et al. Page 26
Table 1Differentially expressed Extracellular Matrix Genes, Proteases, and Growth Factors.Gene expression microarray analysis of the un-opposed side root apex compared to control side root apex. Notesignificantly increased levels in the expression of specific matrix-related genes, including elastin, tenascin C, brevican,lumican, and biglycan as well as FGF9. Other matrix-related genes and proteases such as laminin, cathepsin D, andMMP11 were downregulated.
Gene Genbank # Ratio Day 10 vs. Control
Elastin NM_007925 11.83
Laminin beta 2 X75928 −11.81
Tenascin C NM_011607 2.57
Brevican NM_007529 46.77
Lumican NM_008524 3.79
Biglycan L20276 2.37
Cathepsin D X52886 −2.55
MMP11 Z12604 −20.21
FGF9 U33535 2.84
World J Orthod. Author manuscript; available in PMC 2009 July 23.