bmb 170 lecture 10 nucleic acids, october 26thsaf.bio.caltech.edu/bi170/bmb170_2017_lecture10.pdfbmb...
TRANSCRIPT

EM of T2 coliphage - Kleinschmidt et al (1962) BBA 61:857-64
BMB 170 Lecture 10Nucleic Acids, October 26th
Today-Basicsandstructure
DoodlePoll!!!

The basic bases in DNA and RNA
RNAonly
DNAonly
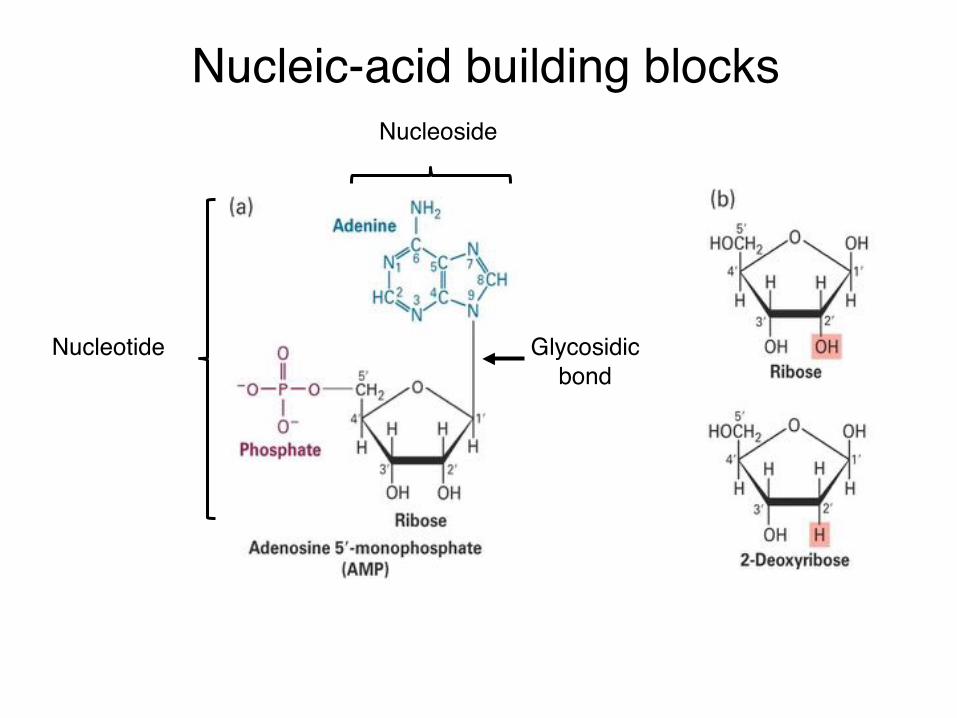
Nucleic-acid building blocksNucleoside
Nucleotide Glycosidicbond

Single strand of RNA
Chain is directional with the convention being 5’ → 3’
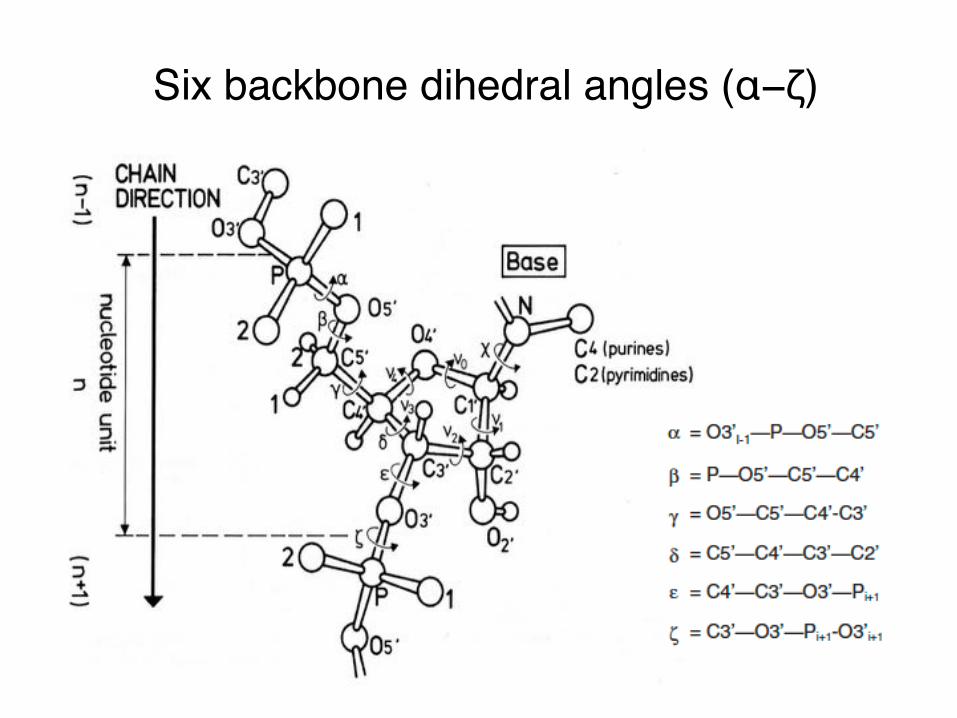
Six backbone dihedral angles (α−ζ)
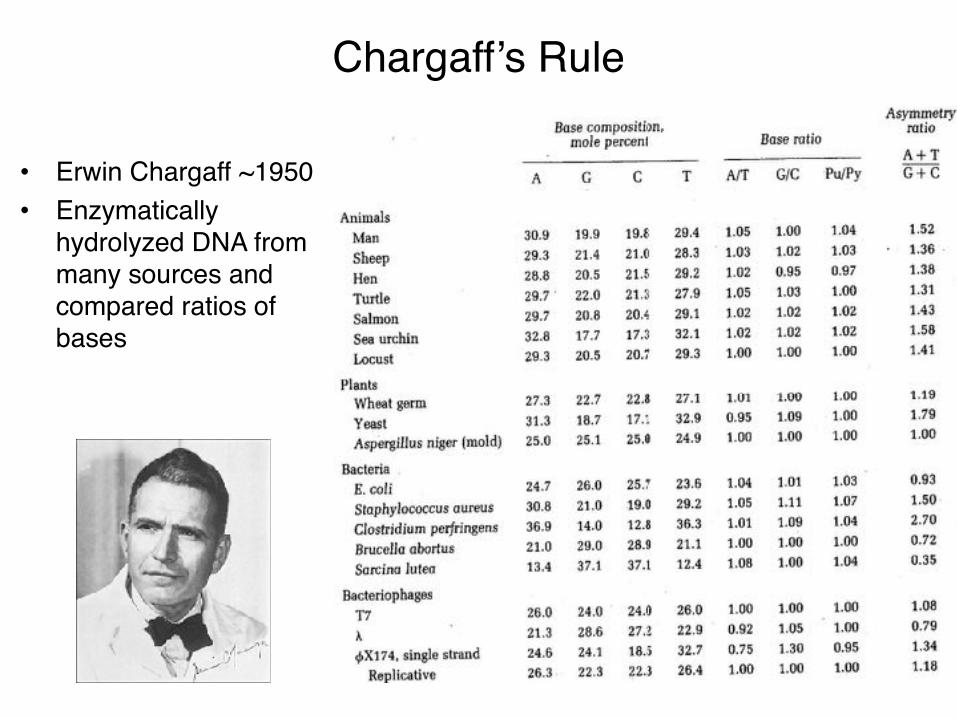
Chargaff’s Rule
• Erwin Chargaff ~1950• Enzymatically
hydrolyzed DNA from many sources and compared ratios of bases

“Tautomers”: Keto and amino forms occur >99.99% of the time under physiological conditions.
D = H-bond donor; A = H-bond acceptor
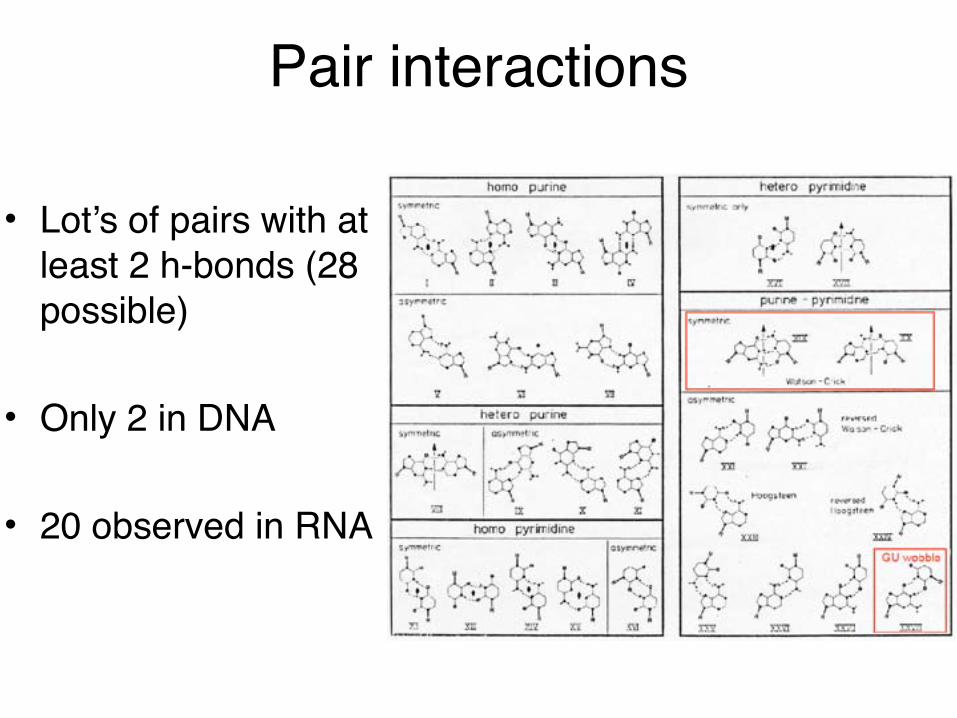
Pair interactions
• Lot’s of pairs with at least 2 h-bonds (28 possible)
• Only 2 in DNA
• 20 observed in RNA

Common base pairs
Fig. 3.25-3.27AdenineCrystals

Sugar pucker
RNA DNA

Rosalind Franklin
Conclusion: Helix with 10 bp/repeat and 3.4 Å between bps Died at 37 from cancer
A physical chemist who refined methods for DNA fiber diffraction first identifying A and B forms of DNA.
Photograph 51

DNA structure determinationRosalind Franklin Watson & Crick
A B
Franklin & Gosling Acta Cryst (1953) 6:673Watson & Crick Nature (1953) 171:737
Nobel in Chemistry 1962 (w/ M. Wilkins)

ScienceMuseum-LondonMorganBeeby,ImperialCollege
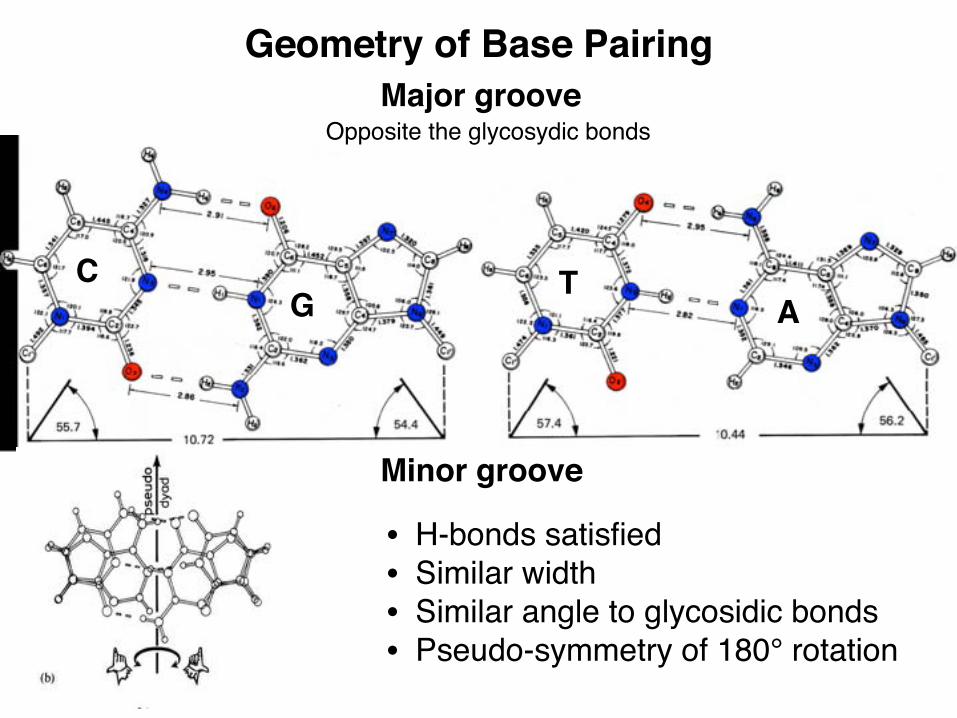
Geometry of Base Pairing
C
• H-bonds satisfied• Similar width• Similar angle to glycosidic bonds• Pseudo-symmetry of 180° rotation
G TA
Major groove
Minor groove
Opposite the glycosydic bonds

Comparison of B-DNA and A-DNABP on axis BP off axis
A-DNA10.92.9Å
31˚20˚
Interior5.9Å
~26Ånarrow/deepwide/shallow
B-DNABP/Turn 10Rise/base3.4ÅHelical twist 36˚Base tilt 6˚Pi Backbone OutPi-Pi 6.9ÅDiameter ~20ÅMajor groove wideMinor groove narrow
Maj
Maj
Min
Min

vdw
Sequence-specific recognition of double helical nucleic acids by proteins
Seeman et al (1976) PNAS 73:804-8
Major Minor
• Major groove: all 4• Minor groove: GC/CG vs AT/TA.• Seeman et al proposed that you
need two H-bonds for discrimination (bidentate interactions).

Dickerson B-DNA (Caltech!)• First structure of DNA double
helix• 19° bend/12 bp• Core GAATTC: B-like with 9.8
bp/turn• Flanking CGCG more
complex, but P-P distance = 6.7 Å (like B)
• BPs not flat– Propeller twist
• 11° for GC • 17° for AT
• Very hydrated
Wing et al Nature (1980) 28:755 (1bna)

DNA parameter descriptors
• Relative to helix• Propeller twist: dihedral
angle of base planes• Displacement: distance
from helix axis to bp center
• Twist: relative rotation around helix axis
• Roll: rotation angle of mean bp plane around C8-C6 line
• Tilt: rotation of bp plane around pseudo-dyad perpendicular to twist and roll axes
Fig. 3.17

Helical parameters of the dodecamer
C1/G24
G12/C13
Range 4.9-18.6° 32.2-41.4° 8.1-11.2 3.14-3.54 Å

Effects of parameters
Fig. 3.18

Calculated base stacking energies
• Can vary quite a lot• Accommodating base geometries
affects stacking energy• ~3-10 kcal/mole (slightly stronger than
an H-bond)
Florián, Sponer and Warshel (1999) J. Phys Chem B 103:884

Tm depends on G+C content

Tm depends on ionic strength
High KCl stabilizes duplex DNA

Predicting secondary structure• Calculate all of the
energies involved• Penalties for loops and
mismatches• Dependent on solvent
considerations
Tinoco et al Nature (1971) 230:362

• 1542 bases in E. coli– Often several copies in a
genome• Highly conserved• Used to classify genus• First model from 100
genomes (Noller lab)
16S Ribosomal RNA
Central
3’ Major
3’ Minor
5’
Woese et al NAR (1980) 8:2275

Current models much more refined• Better free energy
minimization and phylogenetic comparisons
• DNA parameters relatively defined/RNA pretty good
• A number of algorithms• Not good at
pseudoknots
• NUPACK (Pierce lab)– http://www.nupack.org
• mFold (Zuker lab) – now UNAFold– http://
mfold.rna.albany.edu/?q=unafold-man-pages

Using nucleic acids as design tools
• Pierce lab• Winfree lab• Rothemund lab

Self-assembly of polyhedra
He et al (2008) Nature 452:198-201

Reconfigurable topologies
• Uses DNA origami to generate möbius strips
• Strand displacement can yield novel structures
Han et al (2010) Nature Nano 5:712-7


G-tetrads
Fig. 3.29, 3.30 & 3.31 RNA quadruplex UGGGGU (1j8g)

RNA structure (A-form)• RNA
– Steric clashes force A-form to dominate– Can form complicated tertiary structure
• Large complexes– Spliceosome– Ribosome
• Lot’s of structures– Small RNA pieces – tRNA– Ribozymes
• Self splicing/cleaving• Introns (261), hammerhead, HDV, hairpin
– Ribosomes (catalytic RNA?)• 30S (1500), 23S (3400), 5S (120)
– Signal Recognition Particle– Spliceosome components
Reviewed in Chen & Varani FEBS Journal (2005) 272:2088-97
MajMin

tRNA • Links genetic code to amino acid code• Predicted by Frances Crick
– The Sequence Hypothesis “assumes that the specificity of a piece of nucleic acid is expressed solely by the sequence of its bases, and that this sequence is a (simple) code for the amino acid sequence of a particular protein”
– The Central Dogma “the transfer of information from nucleic acid to nucleic acid, or from nucleic acid to protein may be possible, but transfer from protein to protein, or from protein to nucleic acid is impossible”
– The Adaptor Hypothesis “One would expect, therefore, that whatever went on to the template in a specific way did so by forming hydrogen bonds. It is therefore a natural hypothesis that the amino acid is carried to the template by an ‘adaptor’ molecule, and that the adaptor is the part which actually fits on to the RNA. In its simplest form one would require twenty adaptors, one for each amino acid”
Crick"OnProteinSynthesis”SympSocExpBiol(1958)12:138

tRNA secondary structure• All tRNAs are 73 to 93
nucleotides• 7-15% of the bases are
modified• The cloverleaf• Many tRNAs, all have
same overall structure
RajBhandary & Chang JBC (1962) 598-608
Receptor Stem
Anti-codonStem
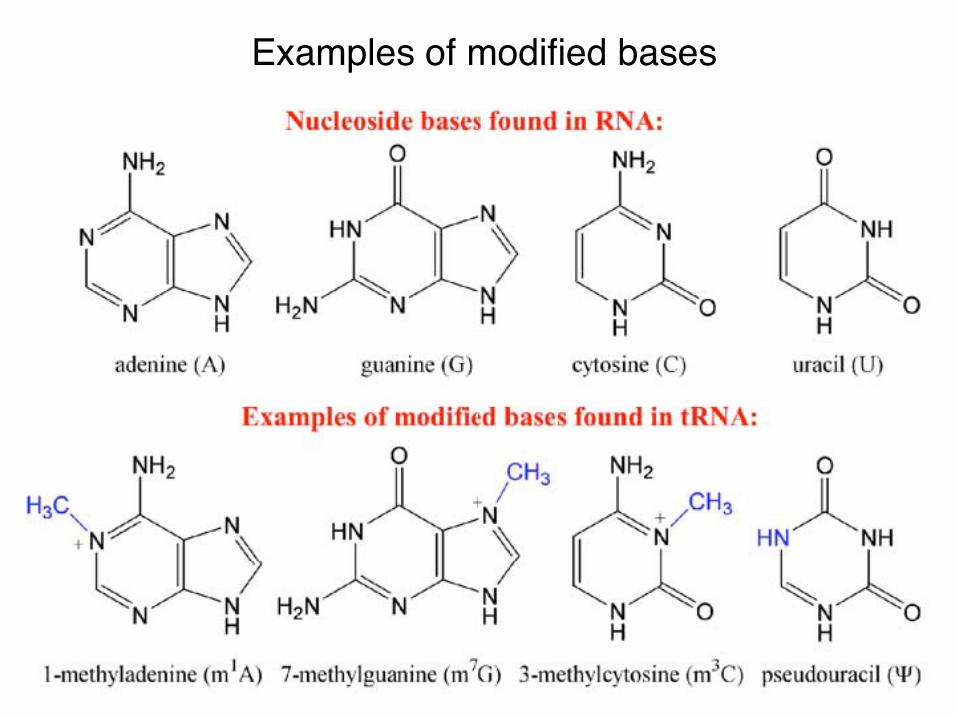
Examples of modified bases

AnQcodonofIle-tRNA2CATforcodonAUA
More modifications
Fig.3.32

Race to the tRNA structure (1974)
3Å resolutionRobertus, Ladner, Finch, Rhodes, Brown, Clark & Klug Nature (Aug 1974) 250:546
3Å resolutionSuddath, Quigley, McPherson, Sneden, Kim, Kim & Rich Nature (Mar 1974) 248:20

31st July, 1974
Dr. Alex“Rich, Department of Biology Massachusetts Institute of Technology Cambridge Mass. 02139
/ U.S.A.
Dear Alex,
Does your name stink. Aaron was convinced that once you had weedled out the details of his structure you would attempt to publish it as your own, This is exactly what has happened, I real& that you had already gone some distance along the same lines, but the fact remains that you said nothing aIjout this at all in public at the Madison meeting and that Kim obtained the details of the Cambridge structure from Robertus at the Gordon Conference. There is absolutely nothing to suggest that you would have actually published a revised structure at this time except for the knowledge you obtained of the Can&rids structure, Moreover you did not even have the elementary courtesy to ack- knowledge the Cambridge work. In addition to use your special influence with Science to rush into publication is quite inexcusable.
Unless you are prepared to make a suitable apology in public I must tell you that your visits to Cambridge in future will not he welcomed,
F, H. C: Crick
copy R. Sinsheimer .

tRNA Structure
tRNAPhe
Rhodes lab at the LMB15 year old xtals (1evv)
Jovine et al JMB (2000) 301:401

tRNA folding
Fig. 3.52 & 3.53

Detailed interactions
Interaction between D- and T- loops
Sharp turns in the tRNA structure (Anticodon, T-loop, D-loop, 9-11 U-turn)
Fig. 3.54 & 3.55

Modified bases• Conserved• Not all the roles are
clear yet• Often aids in stabilizing
long range interactions
Ψ
m5U
m1A

Mg2+ stabilizes tertiary structure• Tertiary structure
brings lots of negative charge together
• Divalent metal ions do the trick
• Note the hexavalent coordination
Jovine et al JMB (2000) 301:401

Base triples
Fig. 3.56

JuncQons
Fig.3.42&3.43

“Hammerhead” ribozyme
Fig. 3.58 & 3.59

Reaction
Hypothetical transition state

Doudna & Cech Labs: Cate et al Science (1996) 273:1678-85
• First ribozyme described (Tom Cech - Nobel) Cech et al Cell (1981) 27:487-96
• Self catalyzes removal of intron• Stable tertiary domain of P4-P6
Tetrahymena group I intron
P4-P6 domain (1gid)

Structure (from the text)
Fig. 3.62 & 3.63
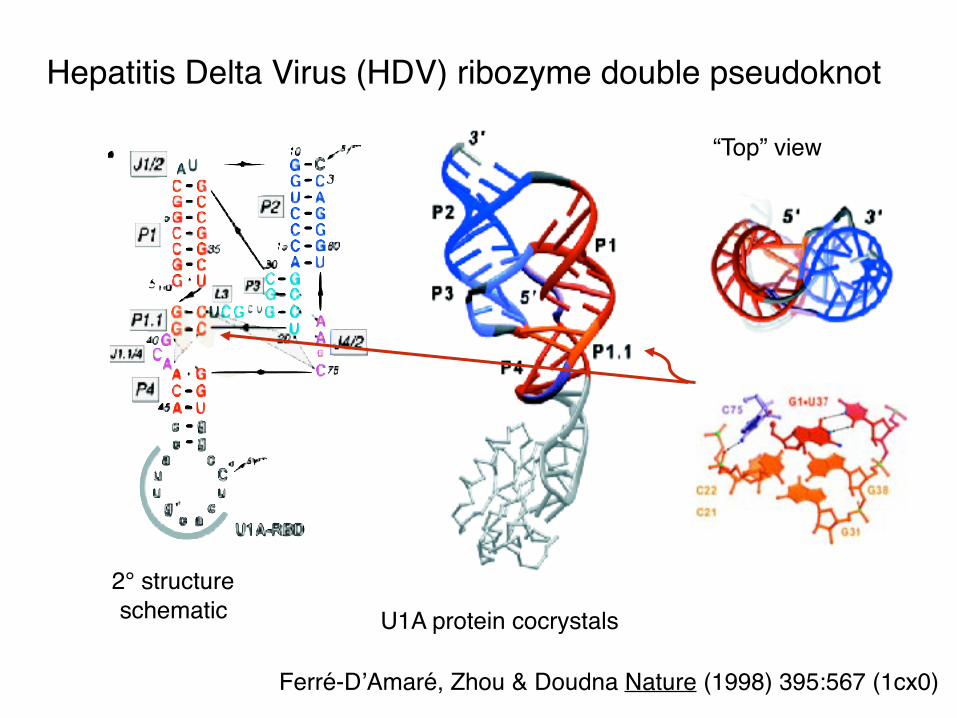
Hepatitis Delta Virus (HDV) ribozyme double pseudoknot
“Top” view
U1A protein cocrystals2° structureschematic
Ferré-D’Amaré, Zhou & Doudna Nature (1998) 395:567 (1cx0)

Pseudoknot structure
Fig. 3.46 & 3.47 Biotin bound pseudoknot aptamer (1f27)
Apseudoknotstructurecontainsmorethanonestem-loopswhereatleastoneisintercalatedintoanother.
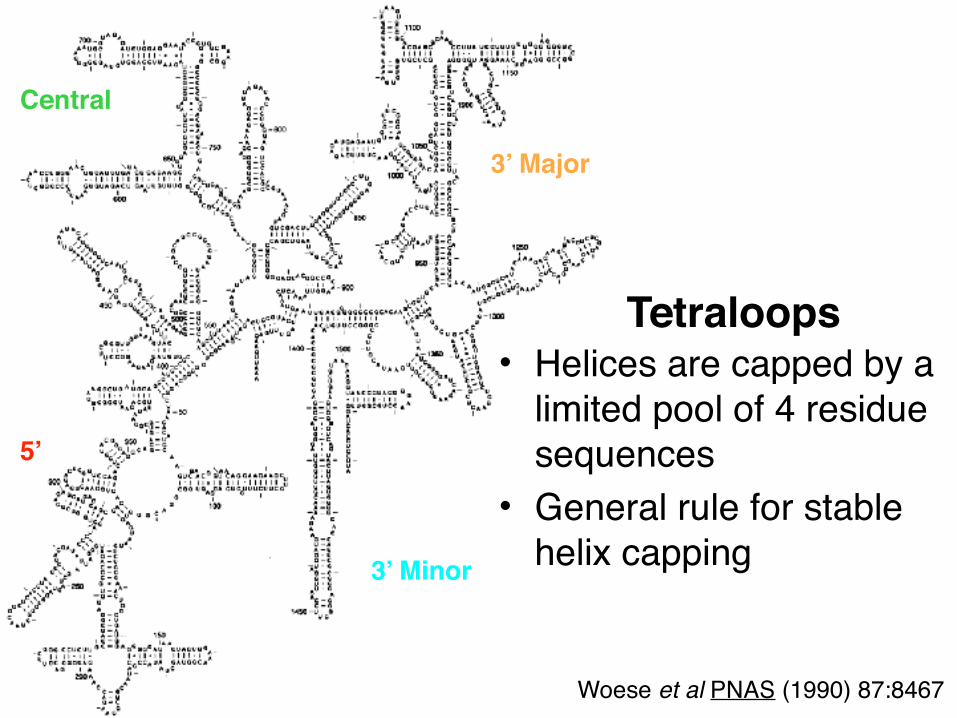
• Helices are capped by a limited pool of 4 residue sequences
• General rule for stable helix capping
Tetraloops
Central
3’ Major
3’ Minor
5’
Woese et al PNAS (1990) 87:8467
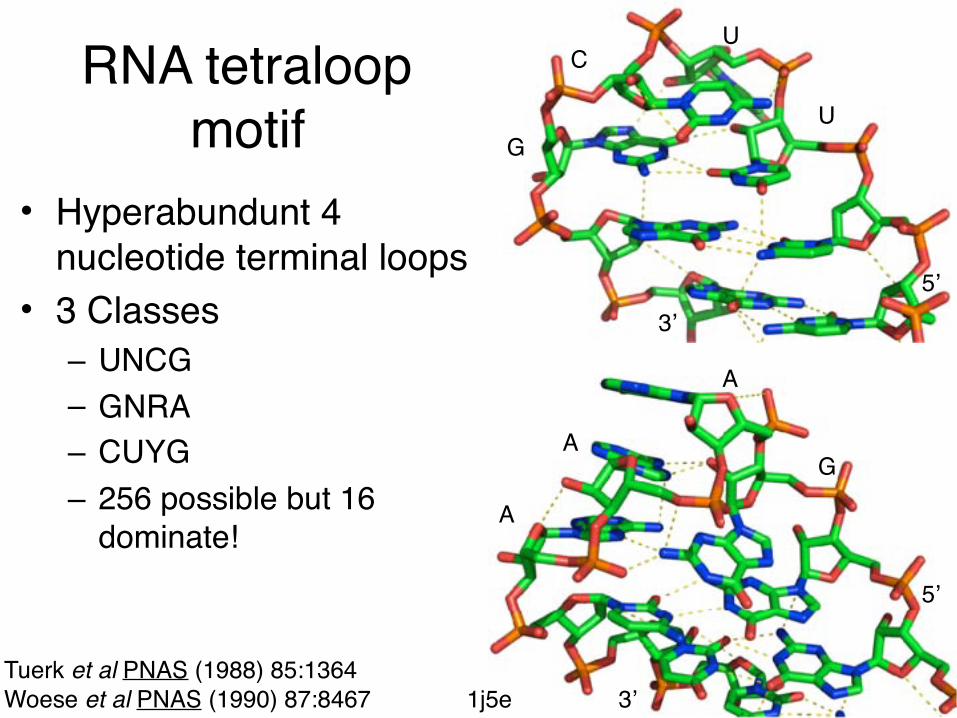
RNA tetraloop motif
• Hyperabundunt 4 nucleotide terminal loops
• 3 Classes– UNCG– GNRA– CUYG– 256 possible but 16
dominate!
Tuerk et al PNAS (1988) 85:1364Woese et al PNAS (1990) 87:8467
5’3’
U
UC
G
5’
3’
G
A
A
A
1j5e

RNA tertiary interactions• Several large RNA structures– Ribozymes• Self splicing/cleaving• Introns (261), hammerhead, HDV, hairpin
– Ribosome (catalytic RNA?)• 30S (1500), 23S (3400), 5S (120)
– Structural motifs• Surprisingly few long range motifs
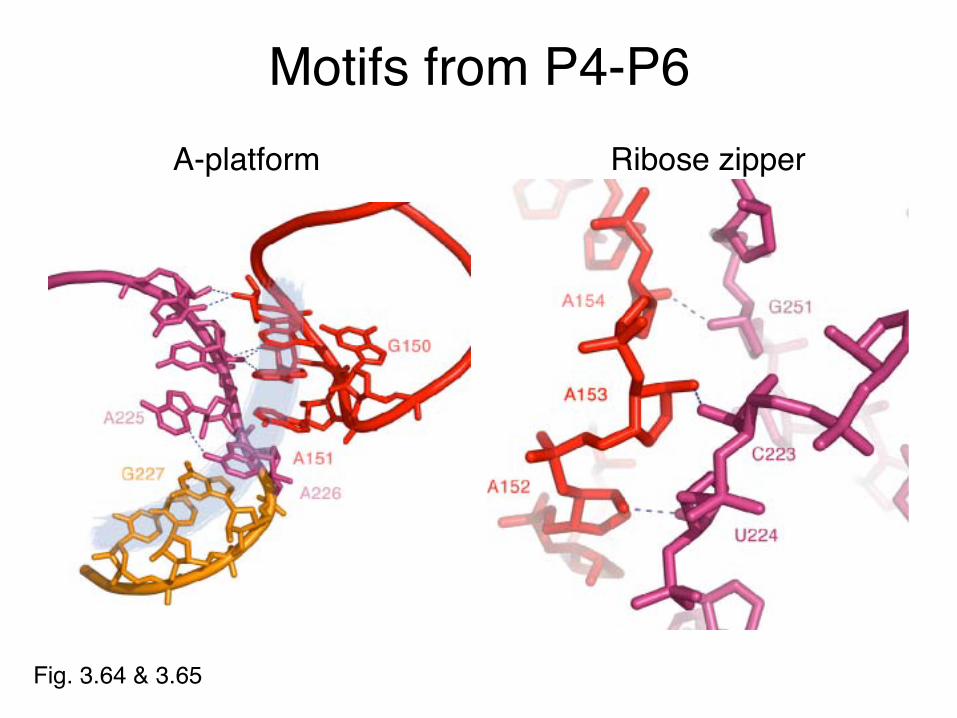
Motifs from P4-P6
Fig. 3.64 & 3.65
A-platform Ribose zipper

A-minor motif• First noted in P4-P6
domain, also found in ribosome
• Long range stabilizing interaction
• Minor groove bulge – Exposed A-platform– Triplet
• GAAA tetraloop docks in minor groove
• Stabilized by π-stacking• Adenines can measure
minor groove1gid
Steitz & Moore Labs: Nissen et al PNAS (2001) 98:4899-903

A-minorinteracQonsfrom23SrRNA
Fig.3.49

Minor groove packing
• Adenine stabilized (most typical)
• Phosphate ridge to minor groove (usually stabilized by guanine N2s)
• End on mode, unpaired purine mediates helices at right angles
Examples taken from the 30S ribosome

hZps://www.youtube.com/watch?v=u9dhO0iCLww
Directedin1971byRobertAlanWeissfortheDepartmentofChemistryofStanfordUniversityandimprintedwiththe"freelove"auraoftheperiod,thisshortfilmconQnuestobeshowninbiologyclasstoday.IthassincespawnaseriesofsimilarfunnyaZemptsatvulgarizingproteinsynthesis.NarratedbyPaulBerg,1980NobelprizeforChemistry.