blazevich 2006
TRANSCRIPT

Sports Med 2006; 36 (12): 1003-1017LEADING ARTICLE 0112-1642/06/0012-1003/$39.95/0
2006 Adis Data Information BV. All rights reserved.
Effects of Physical Training andDetraining, Immobilisation, Growthand Aging on HumanFascicle GeometryAnthony J. Blazevich
Centre for Sports Medicine and Human Performance, Brunel University, Uxbridge,Middlesex, UK
In addition to its size and the extent of its neural activation, a muscle’sAbstractgeometry (the angles and lengths of its fibres or fascicles) strongly influences itsforce production characteristics. As with many other tissues within the body,muscle displays significant plasticity in its geometry. This review summarisesgeometric differences between various athlete populations and describes researchexamining the plasticity of muscle geometry with physical training, immobilisa-tion/detraining, growth and aging. Typically, heavy resistance training in youngadults has been shown to cause significant increases in fascicle angle of vastuslateralis and triceps brachii as measured by ultrasonography, while high-speed/plyometrics training in the absence of weight training has been associated withincreases in fascicle length and a reduction in angles of vastus lateralis fascicles.These changes indicate that differences in geometry between various athleticpopulations might be at least partly attributable to their differing training regimes.Despite some inter-muscular differences, detraining/unloading is associated withdecreases in fascicle angle, although little change was shown in muscles such asvastus lateralis and triceps brachii in studies examining the effects of prolongedbed rest. No research has examined the effects of other interventions such asendurance or chronic stretching training. Few data exist describing geometricadaptation during growth and maturation, although increases in gastrocnemiusfascicle angle and length seem to occur until maturation in late adolescence.Although some evidence suggests that a decrease in both fascicle angle and lengthaccompanies the normal aging process, there is a paucity of data examining theissue; heavy weight training might attenuate the decline, at least in fascicle length.A significant research effort is required to more fully understand geometricadaptation in response to physical training, immobilisation/detraining, growth andaging.

1004 Blazevich
Historically, changes in human fascicle geome- changes after training and detraining interventionsand/or aging and development.try1 have been impossible to examine since the
excision and subsequent study of muscle could onlybe performed in cadavers. More recently, imaging
1. Geometry of Human Musclestechniques such as magnetic resonance (MRI)[1] andultrasound[2,3] imaging (figure 1) have allowed the invivo examination of geometric, i.e. fascicle length Many human muscles contain fascicles that doand angle, changes.[2,4-8] Results of such research not run directly from origin to insertion, but arehave predominately shown geometry to be highly angled and attach to the muscles’ aponeuroses (fig-changeable. An understanding of how different ure 2). These ‘pennate’ muscles can have complexforms of muscle loading, or exercise training, affects fascicular arrangements that differ markedly be-geometry would allow us to bring about deliberate tween muscles. Their fascicle arrangement is proba-and specific changes that would influence the mus- bly a greater determinant of general muscle functioncles’ force-generating properties. There are, howev- than other aspects such as fibre type.[9] As a generaler, a limited number of studies that have examined rule, muscles with large fascicle attachment anglesthe plasticity of fascicle geometry, or that have and correspondingly short fascicles are suited tocompared populations of individuals who have per- contractions involving high forces[10] and/or pro-formed different exercise training. The present re- duce forces over a relatively short range of mo-view summarises research that quantifies changes in tion.[11] Examples include the large proximal mus-fascicle geometry with exercise and detraining, and cles of the legs (e.g. vastus lateralis; see table I),development and aging. A greater understanding of smaller muscles such as those involved in mastica-
tion,[10] and those involved in postural control suchfascicle geometric adaptation could inform bothas soleus and tibialis posterior. Large fascicle anglesphysical performance and rehabilitative trainingare also seen in muscles that attach to long tendons,programmes, as well as allowing the development ofsuch as gastrocnemius lateralis, which produce suf-muscle models that predict physical performanceficient force to remain relatively isometric duringthe propulsive phase of stretch-shorten contractionsin walking,[12-14] running[12,14] and jumping with[15,16]
and without[12,14,17] countermovement, and allow op-timum elongation and recoil of the tendon.[12] Mus-cles that commonly participate in movements re-quiring significant length change or a high shorten-ing velocity tend to be characterised by longfascicles attaching at relatively small angles to thetendon.[9,18] Examples include adductor magnus andlongus, whose longer fascicles attaching at smallerangles allows force generation over a large range ofmotion at the hip during flexion/adduction tasks,and the topologically similar long head of bicepsfemoris, which is highly active during sprint run-ning.[19-21]
b
ACSA
a
α
θβ
Muscle length
lf = ∆a − b
PCSA
Fig. 1. Muscle architectural parameters include: fibre length = dis-tance between ends of a fibre (a to b); pennation angle (θ) =fascicle angle (relative to the aponeurosis [α]) minus the aponeuro-sis angle (relative to the tendon [β]); muscle length; and anatomical(ACSA) or physiological (PCSA) cross-sectional area. The PCSAcan be calculated as (V/t) • sinθ for a simple, uni-pennate muscle,where V is the muscle volume, t is the muscle thickness from oneaponeurosis to the other, and θ is the pennation angle. In morecomplex muscles, PCSA is calculated as V/ lf • cosθ where lf is themean fibre/fascicle length.
1 The term ‘fascicle geometry’ as used here describes the angulation and length of muscle fascicles. The broader term‘muscle architecture’ will be reserved for the description of the whole muscle structure including fascicle geometry,muscle length and muscle volume (or physiological cross-sectional area).
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

Plasticity of Fascicle Geometry 1005
2. Effects of Geometry onForce Production
2.1 Fibre Length
Muscle-specific geometric differences can be ra-tionalised by considering the effects of differentfascicle arrangements on force production. Musclescontaining long fascicles would produce forces overlarge length ranges and at high shortening speedsbecause they have a large number of simultaneouslycontracting, serially arranged sarcomeres. Moreo-ver, since the shortening speed of each sarcomere ina fibre or fascicle would be slower for a given speedof whole-fibre shortening when there are moresarcomeres in series, sarcomere force would notdecrease as rapidly as fibre-shortening speeds in-crease, according to the force-velocity relationship.Therefore, at high shortening velocities, longer fas-cicles are capable of generating greater force. None-theless, the increase in sarcomere number wouldincrease the energy cost of force production, sinceforce output is not improved with the increase inenergy-consuming sarcomeres. Indeed, it is theoreti-cally possible that energy consumption per sarcom-ere is increased due to the absorption of energy byneighbouring sarcomeres in protein structures suchas titin,[23-26] cross-bridges,[27,28] and actin and myo-sin filaments,[29-31] as well as by the re-arrangementof the z-band lattice,[32] which has been shown to
100
m
Tendon/aponeurosis
Fascicleforce
θ = 5°
θ
Tend
on fo
rce
(% m
uscl
e fo
rce)
80
θ = 5° θ = 25°
θ = 45°
60
40
20
05 15
Fascicle angle (°)25 35 45 55
m
θ = 25°
θ
m
θ = 45°
θ
Fig. 2. The effect of fascicle angle on the quantity of force directedalong the tendon axis. As fascicle angle (θ) increases, the propor-tion of fibre force directed along the tendon decreases (tendonforce = sum of fibre forces × cos[fibre angle]), where the fibre forceis represented by the arrow attached to the tendon/aponeurosis.The tendon is attached to a mass (m) representing the inertia of thesystem on which the muscle-tendon complex does work. The effectof fascicle angulation on the proportion of force directed along thetendon is minimal when fascicle angle is moderate (e.g. <25°), butincreases non-linearly as fascicle angle increases, as shown in thegraph.
extend by approximately 20% in tetanised fibres.[33]
Thus, muscle containing longer fascicles produce angulation will have a greater physiological cross-force over long length ranges and at high shortening section and thus a greater force-generating capaci-speeds, but the relative energy cost of force produc- ty.[34] Essentially, this fascicular geometry allows ation is high. greater amount of contractile tissue to attach to a
given area of tendon or aponeurosis.[7] Second, fas-2.2 Fibre Angle cicle angulation probably increases force by al-
lowing fibres to operate closer to their optimumWhile increases in fascicle angulation, or penna-length. Fibres in pennate muscles rotate as theytion, reduces the proportion of fibre force directedshorten[35] so tendon excursion is greater than thealong the tendon (tendon force = sum of fibre forcesshortening distance of the individual fibres. Accord-× cos[fibre angle]), this effect is minimal whening to the length-tension relationship, there will befascicle angle is moderate (e.g. <25°), as shown inan optimum sarcomere length at which fibres pro-figure 2. However, angulation probably improvesduce their greatest active force. Since optimumforce generation in three main ways. First, for the
same muscle volume, a muscle with larger fascicle sarcomere length seems to occur at lengths where
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)
1006 Blazevich
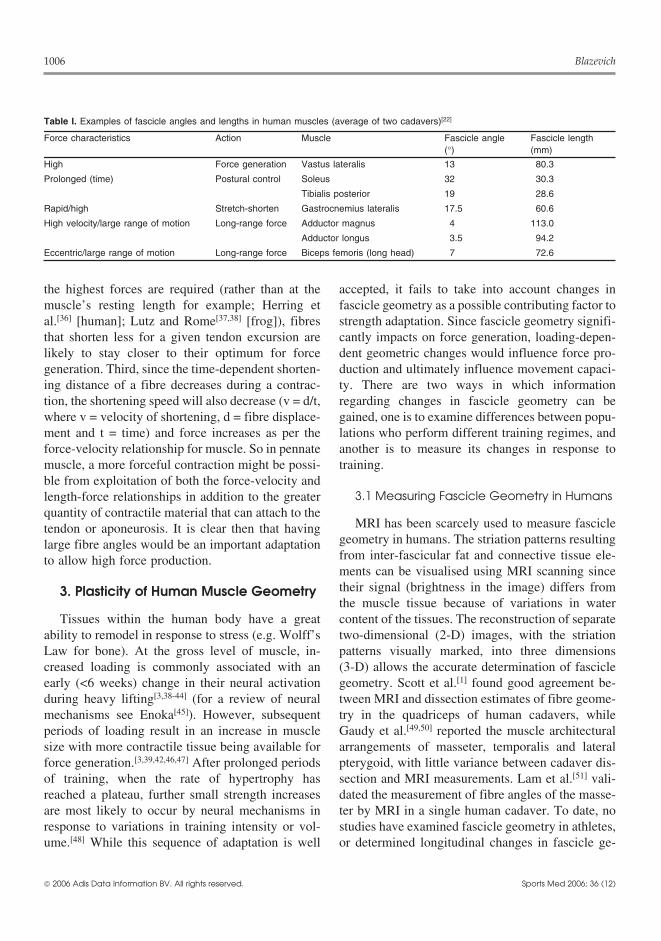
Table I. Examples of fascicle angles and lengths in human muscles (average of two cadavers)[22]
Force characteristics Action Muscle Fascicle angle Fascicle length(°) (mm)
High Force generation Vastus lateralis 13 80.3
Prolonged (time) Postural control Soleus 32 30.3
Tibialis posterior 19 28.6
Rapid/high Stretch-shorten Gastrocnemius lateralis 17.5 60.6
High velocity/large range of motion Long-range force Adductor magnus 4 113.0
Adductor longus 3.5 94.2
Eccentric/large range of motion Long-range force Biceps femoris (long head) 7 72.6
the highest forces are required (rather than at the accepted, it fails to take into account changes inmuscle’s resting length for example; Herring et fascicle geometry as a possible contributing factor toal.[36] [human]; Lutz and Rome[37,38] [frog]), fibres strength adaptation. Since fascicle geometry signifi-that shorten less for a given tendon excursion are cantly impacts on force generation, loading-depen-likely to stay closer to their optimum for force dent geometric changes would influence force pro-generation. Third, since the time-dependent shorten- duction and ultimately influence movement capaci-ing distance of a fibre decreases during a contrac- ty. There are two ways in which informationtion, the shortening speed will also decrease (v = d/t, regarding changes in fascicle geometry can bewhere v = velocity of shortening, d = fibre displace- gained, one is to examine differences between popu-ment and t = time) and force increases as per the lations who perform different training regimes, andforce-velocity relationship for muscle. So in pennate another is to measure its changes in response tomuscle, a more forceful contraction might be possi- training.ble from exploitation of both the force-velocity and
3.1 Measuring Fascicle Geometry in Humanslength-force relationships in addition to the greaterquantity of contractile material that can attach to the
MRI has been scarcely used to measure fascicletendon or aponeurosis. It is clear then that havinggeometry in humans. The striation patterns resultinglarge fibre angles would be an important adaptationfrom inter-fascicular fat and connective tissue ele-to allow high force production.ments can be visualised using MRI scanning sincetheir signal (brightness in the image) differs from3. Plasticity of Human Muscle Geometrythe muscle tissue because of variations in water
Tissues within the human body have a great content of the tissues. The reconstruction of separateability to remodel in response to stress (e.g. Wolff’s two-dimensional (2-D) images, with the striationLaw for bone). At the gross level of muscle, in- patterns visually marked, into three dimensionscreased loading is commonly associated with an (3-D) allows the accurate determination of fascicleearly (<6 weeks) change in their neural activation geometry. Scott et al.[1] found good agreement be-during heavy lifting[3,38-44] (for a review of neural tween MRI and dissection estimates of fibre geome-mechanisms see Enoka[45]). However, subsequent try in the quadriceps of human cadavers, whileperiods of loading result in an increase in muscle Gaudy et al.[49,50] reported the muscle architecturalsize with more contractile tissue being available for arrangements of masseter, temporalis and lateralforce generation.[3,39,42,46,47] After prolonged periods pterygoid, with little variance between cadaver dis-of training, when the rate of hypertrophy has section and MRI measurements. Lam et al.[51] vali-reached a plateau, further small strength increases dated the measurement of fibre angles of the masse-are most likely to occur by neural mechanisms in ter by MRI in a single human cadaver. To date, noresponse to variations in training intensity or vol- studies have examined fascicle geometry in athletes,ume.[48] While this sequence of adaptation is well or determined longitudinal changes in fascicle ge-
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

Plasticity of Fascicle Geometry 1007
ometry using MRI, perhaps because of the difficulty cles, larger errors in fascicle length estimates havebeen observed when compared with 3-D ultrasoundof manually tracing fascicles and then reconstruct-images (2.4–14.0%).[65] Fascicle length is also diffi-ing 3-D images.cult to accurately measure when the width of theMore recently, other MRI-based techniques haveprobe (often 3.8 or 6cm) is insufficient to capture anbeen developed. Diffusion tensor imaging relies onentire fascicle. In these instances, linear approxima-the diffusion of water in the muscle being con-tions are calculated from muscle thickness and fasci-strained by membranes and other cellular constitu-cle angle measures (length = muscle thickness/sin ×
ents, so in muscles the diffusion occurs largely alongfascicle angle). These do not account for fibre curva-
the plane of the fibres. The diffusion tensor can beture that is present in some hypertrophied or con-
calculated from the measurement of the direction oftracting muscles and errors range from approximate-
the diffusion.[52,53] This technique is more efficiently 0% (non-contracted gastrocnemius) to 6.6% (con-
than traditional methods of MRI determination be-tracted gastrocnemius) according to Muramatsu et
cause the fibres can be tracked rapidly by a fibre-al.,[66] or 2.4% in the non-contracted tibialis anterior
tracking algorithm[54,55] and the fibre orientationsmuscle.[67] Fascicle length can also be estimated by
can be quickly reconstructed. While the techniquemeasuring the visible portion directly and then esti-
has been used to determine fascicle geometry inmating the non-visible portion. Errors of 2–7% have
animal muscles,[53,56-59] it has yet to be used to been reported for this method.[68,69] Regardless, ul-quantify human fascicle geometry adaptations. In trasound imaging techniques allow valid and relia-addition to the diffusion tensor technique, magnetic ble measures of fascicle geometry in vivo, and haveresonance elastography has recently been shown to been commonly used to examine population differ-provide information with regard to fascicle geome- ences and longitudinal adaptations.try in humans.[60] After low frequency (<1000Hz)oscillations are induced in the muscle by a pneumat-
3.2 Population Differences inic or mechanical driver, MRI is used to measure theFascicle Geometryspread of shear waves travelling through the muscle.
The spread of waves is constrained by the stiffnessA muscle’s phenotype is at least partly reflectiveof muscle-based tissues, where lateral (with respect
of its long-term activity patterns. Although a per-to the muscle fibres) transmission is affected byson’s genetic predisposition might play a role, it isparallel layers of connective tissue but longitudinal
transmission is relatively unimpeded along the fib-res. Again, this technique has not yet been used toquantify muscle fascicle geometric changes longitu-dinally; however, both diffusion tensor imaging andelastography show great promise as fascicle imag-ing techniques.
The most common method of measuring fasciclegeometry is via the acquisition of 2-D longitudinalimages of the muscle using ultrasound[2,4-7,61,62] (fig-ure 3). When the ultrasound transducer (probe) isoriented in the plane of the fascicles, the fascicleangles and lengths can be accurately measured.Measures of fascicle angle (≤1.5°)[2,63,64] and fasci-cle length (≤1.5mm)[2] have been shown to be simi-lar to those measured directly in cadavers. When thetransducer is not oriented in the plane of the fasci-
Skin/fat layer
Vastuslateralis
Vastusintermedius
Bone
ba
Fig. 3. Images of human muscles can be obtained using ultrasoundimaging. In this longitudinal section of the thigh, the skin/fat layer,femur (bone), and vastus lateralis and vastus intermedius musclesare clearly visible. The aponeurosis (apo) and line of fascicles (fas)are also visible, from which fascicle angle can be determined, andfascicle length can be estimated using standard trigonometric pro-cedures.[5-7,68,69] Physical training can alter a muscle’s fascicle ge-ometry. Here, a period of heavy strength training has resulted in anincrease in fascicle angle concomitant with the increase in musclethickness (a vs b).
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

1008 Blazevich
expected that the physical work capacity of athletes, relatively regardless of cost. These data would sug-gest that there are significant differences in thewho have consistently trained with specific move-fascicle geometry of well trained sprinters comparedment patterns for a period of many years, wouldwith lesser-trained sprinters, endurance runners orvary according to the adaptations elicited by theircontrol subjects.training. Indeed, significant, training-specific differ-
Unique geometry is also seen in weight-trainedences in work capacity have been shown amongindividuals. As expected, muscle thickness mea-groups of athletes with varied training histo-sured at 13 sites on muscles including tricepsries.[70-76] Thus, one method of considering training-brachii, vastus lateralis and gastrocnemius lateralisdependent fascicle geometric adaptations is to com-of powerlifters was significantly correlated withpare populations with different physical activity his-squat, bench-press and deadlift performance (r =tories.0.63–0.91[80]). However, fascicle length of the vas-A significant quantity of research has examinedtus lateralis was also positively correlated with squatdifferences in fascicle geometry between differentlift (r = 0.50) and deadlift (r = 0.54) strength, andathlete populations. For example, both vastus later-triceps fascicle length was correlated with bench-
alis and gastrocnemius in highly trained male sprint-press strength (r = 0.52). While muscle thickness
ers (100m in <11.0 seconds) were found to containand fascicle angle were positively correlated (triceps
longer fascicles attached at smaller angles than inbrachii r = 0.64, p < 0.01; gastrocnemius r = 0.48, p
lesser-trained sprinters (100m in >11.0 seconds[77]).< 0.05), triceps brachii fascicle angle was negatively
In females, sprinters have been shown to have small-correlated with bench-press performance (relative to
er fascicle angles in vastus lateralis than non-trained fat-free mass, r = –0.45) and gastrocnemius fasciclecontrols (with no differences in gastrocnemius) and angle was negatively correlated with deadlift per-longer fascicles in vastus lateralis and gastrocnemi- formance (relative to fat-free mass, r = –0.56). Inus lateralis, which were significantly correlated with another study, bodybuilders were shown to have100m best time (r = –0.51 and r = –0.44, respective- greater muscle thickness and fascicle angles thanly[78]). A comparison of male and female sprinters non-weight-trained controls.[2] There is also somerevealed no differences in absolute or relative (to leg evidence that muscle size, fascicle angle and fasci-length) fascicle length.[78] When the fascicle geome- cle length are simultaneously greater in athletes whotry of sprinters was compared with well trained require a large body mass and physical strength asendurance runners, differences were also seen.[79]
well as fast movement velocities (e.g. Sumo wres-While endurance runners (13.5–14.5 minutes for tlers[81]), than they are in the normal population.5000m) had a lesser muscle thickness in the vastuslateralis and gastrocnemius muscles than sprinters 3.3 Training Effects on Fascicle Geometry(10.0–10.9 seconds for 100m), their fascicles were
While the assessment of population differencesalso shorter and their fascicle angles greater. Com-provides an insight into possible activity-dependentpared with non-active subjects, young enduranceadaptations in fascicle geometry, a more satisfactoryrunners had larger fascicle angles (13.7%) but amethod of examining the plasticity of fascicle geom-similar fascicle length in gastrocnemius medialis;etry is to directly measure the longitudinal adapta-there were no differences in fascicle geometry in thetions.vastus lateralis.[61] At least for the gastrocnemius,
muscles in endurance runners appear to be adapted 3.3.1 Resistance Trainingto produce forces with minimal metabolic cost (i.e. Early research was suggestive of there being littleshort fibres or fascicles require less energy for a change in geometry in response to resistance train-given level of force production because they have ing. Rutherford and Jones[8] showed no change infewer sarcomeres in series), whereas in sprinters vastus lateralis or intermedius fascicle angles after 3they are adapted for high-speed force development months of quadriceps strength training in seven men
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

Plasticity of Fascicle Geometry 1009
Table II. Muscle architecture changes after strength training in humansa
Study Method Muscles Change
Rutherford and Jones[8] 3mo weight training (previously Vastus lateralis, vastus No change in fascicle angle or lengthuntrained) intermedius
Kawakami et al.[7] 16wk elbow extensor training Triceps brachii (lateral head) 29.1% increase in fascicle angle, no(previously untrained) changes in fascicle length (0.9%)
Aagaard et al.[4] 14wk leg extensor training Vastus lateralis 33.8% (2.7°) increase in fascicle angle(previously untrained)
Blazevich and Giorgi[5] 12wk bench-press and triceps Triceps brachii (lateral head) No change in fascicle angleextension training (well trained)
12wk bench-press and triceps Triceps brachii (lateral head) 39.5% (3.2°) increase in fascicle angleextension training + testosteroneinjections (well trained)
Blazevich et al.[84] 5wk isokinetic leg extension Vastus lateralis, vastus No change in fascicle angle or length(previously untrained) intermedius, vastus medialis
a Significant geometric change seems possible in previously untrained subjects, but these seem more limited in highly trainedathletes.
and five women. There was also no relationship range of motion most influenced the changes (sec-tion 5). Thus, recent research has shown a signifi-between fascicle angle and force-generating capaci-cant adaptability of fascicle geometry in response toty, although the authors reported a moderately highlonger-term resistance training.coefficient of variation (13.5%), making small
changes (<2°) in fascicle angle difficult to detect. Other research has examined changes in wellFurther research showed that significant changes in trained, athletic populations. A small change in tri-fascicle geometry occurred in response to training ceps brachii muscle thickness (+13.6%) after a(see example in figure 3 and a summary in table II). 12-week period of heavy resistance training in pre-Kawakami et al.[7] reported significant increases in viously weight-trained men (n = 5) was shown totriceps brachii physiological cross-sectional area occur without fascicle geometry or strength(PCSA) [+33.3%] and fascicle angle (+29.1%, 4.8°) changes.[5] However, in the same study, anotherwith no changes in fascicle length (–0.9%) after 16 group (n = 4) showed a significant strength increaseweeks of elbow extensor training in five men. Aag- (15.8%, bench-press) accompanied by increases inaard et al.[4] found that fascicle angle (+33.8%, 2.7°; fascicle angle (39.5%, 3.2°) after receiving once-vastus lateralis) increased significantly after 14 weekly intramuscular testosterone enanthate injec-weeks of leg extensor training in 11 men simultane- tions (3.5 mg/kg). Little change in geometry orously with anatomical cross-sectional area (+9.7%; strength appears to occur, therefore, in well trainedquadriceps total area) and muscle fibre cross-sec- lifters who continue to perform similar training,tional area (+12.9%; vastus lateralis). More recently, although the administration of anabolic steroidsGondin et al.[82] reported fascicle angle increases in might allow greater strength increases accompaniedvastus lateralis (14%) after 8 weeks of static electro- by significant geometric adaptation. In anothermyostimulation training of the knee extensors that study,[6] however, two resistance training groupsresulted in an increase of 27% in maximum contrac- comprising competitive athletes (n = 7 and n = 8)tion force and a 6% increase in quadriceps cross with at least 1 year of resistance training experience,section. Also, Alegre et al.[83] reported increases in showed increases in vastus lateralis fascicle anglefascicle length in vastus lateralis after 13 weeks of after 5 weeks of heavy (first session in the week) andstrength training when the concentric phase was explosive (second session) squat lift training thatperformed with maximum speed. These data indi- were significantly greater than a group that omittedcate that fascicle length is also adaptable, although it its strength training, but performed only sprint/jump
training (n = 8; no change). In the same study, vastusis presently unclear whether the training velocity or
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

1010 Blazevich
lateralis fascicle length increased only for the sprint/ no changes in muscle size and geometry of elbowextensors of an untrained limb in five men after 16jump-trained group. Muscle thickness of both vastusweeks of training, although no strength changeslateralis and rectus femoris increased in all trainingwere found either. Blazevich et al.[84] have reportedgroups also, although there were no differences inthat significant increases in contralateral knee exten-performance (sprint, jump and strength tests) be-sor strength after 5 weeks of isokinetic trainingtween the groups after training. Similar to previousoccurred without a change in geometry. Thus, al-studies on untrained subjects,[7,83] these results showthough few data are available, there is no currentthat changes in fascicle geometry occur in responseevidence to suggest that fascicle geometric changesto resistance training in well trained subjects. Theare a factor affecting short-term strength increases inresults may have differed from previous studiesa contralateral limb.where little change was noticed in trained subjects
because the training programme differed sufficient-3.3.2 Endurance Trainingly from that which the athletes had previously per-No research has examined changes in geometryformed. These data were the first to show short-term
after long-term endurance training. It is thereforechanges in geometry in response to training, andnot known whether endurance training can elicitalso show that increases in fascicle length and de-geometric adaptations, or how different modes ofcreases in fascicle angle can occur after high-veloc-training (e.g. cycling, running, lifting) might affectity movement training in humans; it is not known,them. It makes intuitive sense that if a muscle couldhowever, whether other factors such as the traininggenerate force more efficiently, then muscle endur-range of motion influence the architectural adapta-ance would improve. Given that muscles with largetion.pennation and shorter fascicles would generate force
Given that rapid (5-week) changes in geometrywith less metabolic cost, such an adaptation might
have been shown to occur in athletes, one mightbe expected with endurance training. Certainly, effi-
speculate that rapid adaptation is a mechanism byciency during stretch-shortening contractions could
which early strength increases occur in previously be expected to increase following such a geometricuntrained subjects. However, recent data[84] showed adaptation. This reasoning is consistent with theno changes in quadriceps muscle size or geometry in finding that vastus lateralis and gastrocnemius fasci-previously untrained men (n = 7) and women (n = 8) cles were shorter and fascicle angles greater in adespite significant increases in concentric and ec- group of endurance runners compared with sprint-centric isokinetic strength of the quadriceps muscle ers.[79] However, it could also be argued that longergroup after 5 weeks of isokinetic knee extensor fibres would benefit muscles that are commonlytraining. Also, Gondin et al.[82] did not find signifi- recruited during endurance tasks where forces arecant increases in vastus lateralis fascicle angle after produced over large ranges of motion, since such4 weeks of electromyostimulation training, despite fibre geometry is uniquely adapted to perform suchthere being significant increases after 8 weeks. It is work. Also, intramuscular pressure (IMP) is greatesttherefore likely that changes in fascicle geometry in muscles with large fibre angles;[88,89] therefore,are not a significant factor influencing early strength occlusion of blood flow during muscle contractionincreases in previously untrained subjects, although would be greater. This hypothesis is consistent withwhether rapid adaptations occur after training using the decreases in blood volume and oxygen satura-other modes (e.g. isotonic/isoinertial) is not tion seen during exercise (1Hz plantar flexion) in theknown.[7]
distal region of gastrocnemius medialis, where fas-Geometric changes are probably not a factor in- cicle angle and length are greatest.[90] During loco-
volved in strength increases that have been reported motion, IMP increases with movement speed[91] inin the untrained contralateral limb in response to line with the known positive relationship betweenunilateral training.[85-87] Kawakami et al.[7] reported IMP and muscle force,[92-95] so the limitation of
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

Plasticity of Fascicle Geometry 1011
Table III. Muscle architecture changes after detraining/unloading in humansa
Study Method Muscles Change
Abe et al.[103] 20d bed rest Gastrocnemius medialis, No change in vastus lateralis or triceps brachii,vastus lateralis, triceps brachii significant decrease in gastrocnemius (5.5%)
measured standing, but not lying (4.7% decrease)
Kawakami et al.[98] 20d bed rest Vastus lateralis, plantarflexors, No change in fascicle angle or lengthtriceps brachii
Narici and Cerretelli[62] Disuse/injury Gastrocnemius lateralis 16.4% decrease in fascicle angle, 12.7% decrease infascicle length
Bleakney and Maffulli[102] Disuse/injury Vastus lateralis 4.8° decrease in fascicle angle, 7.5mm decrease infascicle length
a Some data are suggestive of decreases in fascicle angle and length in line with muscle atrophy. Differences between studies mightbe related to study duration or the length at which muscles were held during unloading.
blood flow at increased speeds of locomotion might li[62] found that disuse atrophy of gastrocnemiusbe expected to impact on muscle endurance. Of lateralis (23.1% reduction in anatomical cross-sec-course, during most forms of locomotion, muscle tional area) in an injured leg was associated with acontractions are phasic and blood flow will not be significant decrease in fascicle angle and lengthsignificantly affected when the duty cycle is below a (compared with uninjured leg; time period of injurythreshold limit (e.g. <40%),[93] so the effect of ge- not reported). Also, although measurements wereometry on endurance performance is probably com- taken at varying times after injury, Bleakney andplex. It would seem likely that fibre type is a more Maffulli[102] found significant decreases in fibre an-important muscle-based determinant of muscle en- gle and length of vastus lateralis in legs of subjectsdurance while fascicle geometry affects more the recovering from tibia/femur fracture, compared withlength-tension and force-velocity characteristics of a the non-injured limb (time after fracture, mean = 7.6muscle. months, range = 14 days to 2 years, 7 months). Abe
et al.[103] reported mixed results in subjects who3.3.3 Detraining/Unloading underwent 20 days of bed rest where there was aNumerous studies have investigated the effect of decrease in gastrocnemius medialis fascicle angle
detraining/unloading on muscle size and strength. measured during standing (5.5%) and a non-signifi-For example, significant reductions in strength and cant reduction during lying (4.7%), but no changesmuscle size follow prolonged bed rest[96-98] along
in triceps brachii or vastus lateralis muscles. There-with decreases in fibre size[99] and changes in fibre
fore, while there is some discrepancy in the litera-type (slow- to fast-twitch transformation[100]). How-
ture, there is good evidence that fascicle geometryever, few studies have measured fascicle geometry
changes can occur in response to unloading/de-after a period of detraining/unloading, or have not
training in humans.reported changes in geometry when it has been
The reasons for the disparate results in de-measured for the determination of PCSA. In onetraining/unloading studies are not clear; however,study, Kawakami et al.[98] found no changes in vas-the interaction effects of pre-unloading fascicle ge-tus lateralis, plantarflexors or triceps brachii fascicleometry and the muscle length during immobilisationlengths or angles in subjects (n = 5) after 20 days ofmight be two factors affecting the magnitude ofbed rest despite significant reductions in PCSA (ta-geometric change during detraining. Studies on cat,ble III); a similar finding was reported in four sub-mouse, rat and rabbit muscles have shown signifi-jects in a later publication.[97] These results are simi-cant decreases in fibre length after periods of un-lar to those reported in rat studies where immobilisa-loading when muscles were immobilised at lengthstion of gastrocnemius and soleus muscles atshorter than normal rest length.[94,104-108] These ef-shortened lengths was not associated with changes
in fibre angles.[101] However, Narici and Cerretel- fects have been reduced or reversed when muscles
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

1012 Blazevich
were immobilised in anatomical[109] or a lengthened 4. Fascicle Geometry through Growthand Agingposition.[93,106,108,109] Immobilisation in a lengthened
position has also been shown to counteract the slow-Fascicle geometric changes have yet to be de-to fast-twitch transformation that often accompanies
scribed in most human muscles through develop-immobilisation[110] and reduces the loss of wholement from infant to adult. Binzoni et al.[112] mea-
muscle mass.[111] These data on animal muscles sured medial gastrocnemius fascicle angle in 134show that the length of immobilisation has an im- subjects (87 men and 47 women) in the age range ofpact on muscle and fibre characteristics, although 0–70 years. Very small fascicle angles at birth in-this has not been fully examined in humans. In creased through childhood and adolescence to reachparticular, given that the fibre angle of a shortened their maxima in early adulthood. A comparison of
bodyweight and height, and tibia length indicated apennate muscle is greater than in a lengthened mus-strong relationship between fascicle angle and phys-cle, one might speculate that immobilisation of mus-ical size, possibly indicating that increases in fasci-cles in a shortened position might maintain the ge-cle angle of the gastrocnemius were related to theometry of highly pennate, short-fibred muscles. Fi-loads imposed on it. No research has examined othernally, given that immobilisation in lengthenedmuscles, making it difficult to predict fascicle geo-
positions,[106] or the application of moderate stretch- metric change with development, particularly ining (30 min/day),[107] has been shown to prevent non-weight-bearing muscles.fibre shortening during disuse, it is also pertinent to More data are available with regard to fascicledetermine whether such interventions might prevent geometry with aging from early adult to old age.the fibre shortening that has sometimes been shown Aging is associated with a loss of muscle size, orto accompany disuse in humans.[62] Certainly the sarcopenia,[113-117] resulting from a loss of fibres/
motor units[118] and a significant reduction in fibreaddition of small amounts of stretching/mobilisa-area, particularly of the fast-twitch fibres.[116,119-121]tion[99] during detraining has been shown to arrestThese morphological changes in muscle have beenmuscle atrophy in the human soleus.implicated, along with possible reductions in muscleactivation,[122,123] in the significant loss of muscle3.3.4 Chronic Stretchingforce[114,115,119,124-126] and power[114,115,126,127] seen in
No research has examined changes in fascicle aged individuals. Changes in fascicle geometry aregeometry after acute or chronic stretching of mus- therefore likely given the positive relationship be-cles in humans. Evidence from animal studies indi- tween fascicle angle and muscle size. Data collectedcates that chronic stretching might have a significant on 229 women from the ages of 20–79 years indicat-effect on fibre length. Immobilisation of muscle or ed that pennation of vastus lateralis (r = –0.50, p <
0.001), but not medial gastrocnemius or tricepsmuscle fibres in a lengthened position for periods ofbrachii, might be reduced with age, although theredays to weeks has resulted in increases in the num-was no evidence for a change in fascicle lengthber of serially arranged sarcomeres and overall fibre(relative to limb length).[128] The lesser pennation oflength in animal muscles.[106,108,110] Interestingly,vastus lateralis in elderly compared with young, butmoderate stretching performed for only 30 min/daylack of differences in gastrocnemius or triceps
was enough to reduce or reverse the fibre shorteningbrachii, or in the length of fascicles in any of these
that accompanied short-length immobilisation in rat muscles, has also been reported for men.[129] In amuscle.[107] It is not known whether flexibility train- smaller (n21–32y = 19, n60–69y = 30), but welling in humans results in an increase in fibre/fascicle matched (anthropometrically) sample, Karamanidislength. It is also not clear what effect, if any, chronic and Arampatzis[61] found no differences in gas-stretching would have on fascicle angle. trocnemius medialis or vastus lateralis fascicle
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

Plasticity of Fascicle Geometry 1013
lengths or angles, regardless of whether the subjects geometry shown with aging are influenced by exer-cise interventions.were habitual endurance runners or not. Nonethe-
less, other data have shown significant differencesin gastrocnemius pennation (young [27–42 years] > 5. Conclusions and Future Directions13.2% [elderly = 70–81 years]) and fascicle length
Human fascicle geometry is highly adaptable.(young >10.2%) in subjects matched for height,Heavy weight training in physically active individu-mass and physical activity.[117] These data suggestals is often associated with increases in fasciclethat the aging process is accompanied by a signifi-angle (with some evidence for fascicle shortening)cant change in fascicle geometry that cannot neces-with these changes amplified by anabolic steroidsarily be explained by physical activity levels. Fur-use. In the elderly, the prominent adaptation seemsthermore, Morse et al.[130] showed that decreases into be an increase in fascicle length, although it is notmuscle volume of soleus and medial and lateralclear how geometry changes in response to longer-gastrocnemius were associated with decreases interm training. Both detraining and aging are associ-both fascicle angle and length. These decreases wereated with decreases in fascicle angle and/or de-similar amongst the three muscles indicating thatcreases in fascicle length, although there appears tothese muscles, which fulfil different functionalbe some inter-muscular differences. These changesroles, are affected similarly during aging. Thus,have important consequences for force generationwhile it seems that fascicle geometry changes fromsince increases in fascicle angle allow higher muscleadult to old age, more research is required to sub-forces, while increases in fascicle length allowstantiate these early findings and further examinehigher shortening speeds and for forces to be gener-inter-muscular differences.ated over larger length ranges.
The significant improvements in both muscle While there are significant data showing the plas-strength[131-133] and power[127] after extended periods ticity of fascicle geometry, more research is requiredof resistance exercise in older individuals are associ- to fully understand the effects of physical training.ated with increases in cross-sectional area or volume Not least, no research has examined the effects ofof muscles.[127-130] This increase in cross-sectional endurance or chronic stretching training. Further-area is often largely explained by increases in areas more, in order to more fully understand how fascicleof both type I and II muscle fibres.[131,134] Few geometry responds to heavy loading (e.g. resistanceresearchers have examined the effects of changing training), a number of research lines need to begeometry on muscle size, or indeed the effects of explored. For example, despite a well establishedgeometry on force development. One study[133] has link between fascicle geometry and function, andshown that an increase in muscle volume was attrib- significant evidence of geometric changes in re-uted partly to an increase in fascicle length of the sponse to resistance training, relatively little re-vastus lateralis. This change largely accounted for search has examined geometric changes in responsethe increase in muscle PCSA, and suggests that to athletic training interventions. Only one researchsignificant changes in geometry might occur with study[6] has examined changes in geometry in re-training in aged subjects. In this case, the change sponse to training in athletic subjects who performtoward longer fascicle lengths is at odds with the mixed training regimens, and only one study[128] hasincreases in fascicle angle (with no change or a examined changes in older individuals, so the ef-slight reduction in fascicle length) seen after training fects of training programme manipulations in ath-in younger subjects. This might reflect an adaptation letes and the aged remain relatively unstudied. Also,in less mobile older subjects to a greater movement no research has specifically investigated the effectsrange of motion used in exercise training compared of training mode, contraction type or range of mo-with their normal daily activities. More research is tion. It is unclear whether, for example, adaptationsrequired to determine how the changes in fascicle to isokinetic training are different to isotonic/
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

1014 Blazevich
9. Burkholder TJ, Fingado B, Baron S, et al. Relationship betweenisoinertial training. Despite changes in sarcomeremuscle fiber types and sizes and muscle architectural proper-
number being reported after periods of eccentric ties in the mouse hindlimb. J Morphol 1994; 221: 177-9010. Van Eijden TMGJ, Korfage JAM, Brugman P, et al. Architec-training in rats (downhill running),[135,136] which
ture of the human jaw-closing and jaw-opening muscles. Anatmight suggest that overall fibre/fascicle length also Rec 1997; 248: 464-74increases with such training, no research has ex- 11. Lieber RL, Friden J. Functional and clinical significance of
skeletal muscle architecture. Muscle Nerve 2000; 23: 1647-66amined differences between the effects of concen-12. Hof AL. Muscle mechanics and neuromuscular control. J Bi-tric, eccentric or isometric training programmes on omech 2003; 36: 1031-8
geometric adaptation. Also, while range of motion- 13. Hof AL, Geelen BA, Van den Berg J. Calf muscle moment,work and efficiency in level walking: role of series elasticity. Jspecific strength adaptations cannot be fully ex-Biomech 1983, 37
plained by neural mechanisms,[87] no research has 14. Hof AL, Van Zandwijk JP, Bobbert MF. Mechanics of humantriceps surae muscle in walking, running and jumping. Actaexamined the effects of training range of motion. APhysiol Scand 2002; 174: 17-30greater understanding of these parameters could
15. Kawakami Y, Muraoka T, Ito S, et al. In-vivo muscle-fibrehave significant benefits for injury/illness rehabilita- behaviour during counter-movement exercise in humans
reveals significant role of tendon elasticity. J Physiol 2002;tion in that maintenance of muscle strength during540: 635-46relative immobilisation might accelerate physical 16. Kubo K, Kanehisa H, Takeshita D, et al. In vivo dynamics of
recovery. No less, a more complete understanding human medial gastrocnemius muscle-tendon complex duringstrength-shortening cycle exercise. Acta Physiol Scand 2000;might also inform athletic training practices.170: 127-35
17. Kurokawa S, Fukunaga T, Fukashiro S. Behavior of fasciclesand tendinous structures of human gastrocnemius during verti-Acknowledgements cal jumping. J Appl Physiol 2001; 90: 1349-58
18. Lieber RL, Jacobson MD, Fazelle BM, et al. Architecture ofselected muscles of the arm and forearm: anatomy and impli-Special thanks to Prof. Craig Sharp for his insightfulcations for tendon transfer. J Hand Surg 1992; 17A: 787-98comments and discussion during the preparation of this arti-
19. Elliott BC, Blanksby BA. The synchronization of muscle activi-cle. This research was partly funded by an American Societyty and body segment movements during a running cycle. Medof Biomechanics research grant (ASB-406015). The author Sci Sports 1979; 11: 322-7
has no conflicts of interest that are directly related to the 20. Wiemann K, Tidow G. Relative activity of hip and knee exten-content in this article. sors in sprinting: implications for training. N Studies Athl
1995; 10: 29-4921. Wood GA. Optimal performance criteria and limiting factors in
sprint running. N Studies Athl 1986; 2: 55-63References22. Friederich JA, Brand RA. Muscle fiber architecture in the1. Scott SJ, Engstrom CM, Loeb GE. Morphometry of human
human lower limb. J Biomech 1990; 23: 91-5thigh muscles: determination of fascicle architecture by mag-23. Keller TCS. Molecular bungees. Nature 1997; 387: 233-5netic resonance imaging. J Anat 1993; 182: 249-5724. Politou AS, Thomas DJ, Pastore A. The folding and stability of2. Kawakami Y, Abe T, Fukunaga T. Muscle-fiber pennation
titin immunoglobulin-like modules, with implications for theangles are greater in hypertrophied than in normal muscles. Jmechanism of elasticity. Biophys J 1995; 96: 2601-10Appl Physiol 1993; 74: 2740-4
25. Tskhovrebova L, Trinick J, Sleep JA, et al. Elasticity and3. Narici MV, Hoppeler H, Kayser B, et al. Human quadricepsunfolding of single molecules of the giant muscle protein titin.cross-sectional area, torque and neural activation during 6 moNature 1997; 387: 308-12strength training. Acta Physiol Scand 1996; 157: 175-86
26. Tskhovrebova L, Trinick J. Extensibility in the titin molecule4. Aagaard P, Andersen JL, Dyhre-Poulsen P, et al. A mechanismand its relation to muscle elasticity. Adv Exp Med Biol 2000;for increased contractile strength of human pennate muscle in481: 163-78response to strength training: changes in muscle architecture. J
27. Linari M, Dobbie I, Reconditi M, et al. The stiffness of skeletalPhysiol 2001; 534: 613-23muscle in isometric contraction and rigor: the fraction of5. Blazevich AJ, Giorgi A. Effect of testosterone administrationmyosin heads bound to actin. Biophys J 1998; 74: 2459-73and weight training on muscle architecture. Med Sci Sports
Exerc 2001; 33: 1688-93 28. Tawada K, Kimura M. Stiffness of glycerinated rabbit psoasfibers in the rigor state: filament-overlap relation. Biophys J6. Blazevich AJ, Gill ND, Bronks R, et al. Training-specific mus-1984; 45: 593-602cle architecture adaptation after 5-wk concurrent training in
athletes. Med Sci Sports Exerc 2003; 35: 2013-22 29. Huxley HE, Stewart A, Sosa H, et al. X-ray diffraction measure-ments of the extensibility of actin and myosin filaments in7. Kawakami Y, Abe T, Kuno S-Y, et al. Training-inducedcontracting muscle. Biophys J 1994; 67: 2411-21changes in muscle architecture and specific tension. Eur J Appl
Physiol 1995; 72: 37-43 30. Martyn DA, Chase PB, Regnier M, et al. A simple model with8. Rutherford OM, Jones DA. Measurement of fibre pennation myofilament compliance predicts activation-dependent cross-
using ultrasound in the human quadriceps in vivo. Eur J Appl bridge kinetics in skinned skeletal fibers. Biophys J 2002; 83:Physiol 1992; 65: 433-7 3425-34
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

Plasticity of Fascicle Geometry 1015
31. Wakabayashi K, Sugimoto Y, Tanaka H, et al. X-ray diffraction 52. Basser PJ, Mattiello J, LeBihan D. MR diffusion tensor spec-evidence for the extensibility of actin and myosin filaments troscopy and imaging. Biophys J 1994; 66: 259-67during contraction. Biophys J 1997; 67: 2422-35 53. van Donkelaar CC, Kretzers LJ, Bovendeerd PH, et al. Diffu-
32. Schroeter P. P, Bretaudiere JP, Sass RL, et al. Three-dimension- sion tensor imaging in biomechanical studies of skeletal mus-al structure of the Z band in a normal mammalian skeletal cle function. J Anat 1999; 194: 79-88muscle. J Cell Biol 1996; 133: 571-83 54. Mori S, Crain BJ, Chacko VP, et al. Three-dimensional tracking
33. Goldstein MA, Michael LH, Schroeter JP, et al. The Z-band of axonal projections in the brain by magnetic resonancelattice in skeletal muscle before, during and after tetanic con- imaging. Ann Neurol 1999; 45: 265-9traction. J Muscle Res Cell Motil 1986; 7: 527-36 55. Mori S, van Zijl PC. Fiber tracking: principles and strategies – a
34. Fukunaga T, Miyatani M, Tachi M, et al. Muscle volume is a technical review. NMR Biomed 2002; 15: 468-80major determinant of joint torque in humans. Acta Physiol 56. Heemskerk AM, Strijkers GJ, Vilanova A, et al. DeterminationScand 2001; 172: 249-55 of mouse skeletal muscle architecture using three-dimensional
35. Muhl ZF. Active length-tension relation and the effect of muscle diffusion tensor imaging. Mag Res Med 2005; 53: 1333-40pinnation of fiber lengthening. J Morphol 1982; 173: 285-92 57. van Doorn A, Bovendeerd PH, Nicolay K, et al. Determination
36. Herring SW, Grimm AF, Grimm BR. Regulation of sarcomere of muscle fibre orientation using diffusion-weighted MRI. Eurnumber in skeletal muscle: a comparison of hypotheses. Mus- J Morphol 1996; 34: 5-10cle Nerve 1984; 7: 161-73 58. Damon BM, Ding Z, Anderson AW, et al. Validation of diffu-
37. Lutz GJ, Rome LC. Muscle function during jumping in frogs I: sion tensor MRI-based muscle fiber tracking. Mag Res Medsarcomere length change, EMG pattern, and jumping perform- 2002; 48: 97-104ance. Am J Physiol 1996; 271: C563-70 59. Cleveland GG, Chang DC, Hazlewood CF, et al. Nuclear mag-
38. Lutz GJ, Rome LC. Muscle function during jumping in frogs II: netic resonance measurement of the intracellular water. Bi-mechanical properties of muscle – implications for system ophys J 1976; 16: 1043-53design. Am J Physiol 1996; 271: C571-8 60. Bensamoun SF, Ringleb SI, Littrell L, et al. Determination of
39. Hakkinen K, Komi PV. Effect of explosive type strength train- thigh muscle stiffness using magnetic resonance elastography.ing on electromyographic and force production characteristics J Mag Res Imag 2006; 23: 242-7of leg extensor muscles during concentric and various stretch- 61. Karamanidis K, Arampatzis A. Mechanical and morphologicalshortening cycle exercises. Scand J Sports Sci 1985; 7: 65-76 properties of human quadriceps femoris and triceps surae
40. Hakkinen K, Komi PV. Electromyographic changes during muscle-tendon unit in relation to aging and running. J Biomechstrength training and detraining. Med Sci Sports Exerc 1983; 2006; 39: 406-1715: 455-60 62. Narici M, Cerretelli P. Changes in human muscle architecture in
41. Hortobagyi T, Barrier J, Beard D, et al. Greater initial adapta- disuse-atrophy evaluated by ultrasound imaging. J Gravittions to submaximal muscle lengthening than maximal short- Physiol 1998; 5: 73-4ening. J Appl Physiol 1996; 81: 1677-82 63. Chleboun GS, France AR, Crill MT, et al. In vivo measurement
42. Moritani T, DeVries HA. Neural factors versus hypertrophy in of fascicle length and pennation angle of the human bicepsthe time course of muscle strength gain. Am J Phys Med 1979; femoris muscle. Cells Tiss Org 2001; 169: 401-958: 115-30 64. Narici MV, Binzoni T, Hiltbrand E, et al. Changes in human
43. Patten C, Kamen G, Rowland D, et al. Rapid adaptations of gastrocnemius architecture with joint angle, at rest and withmotor unit firing rate during the initial phase of strength isometric contraction, evaluated in vivo. J Physiol 1996; 496:development. Med Sci Sports Exerc 1995; 27: S6 287-97
44. Ploutz LL, Tesch PA, Biro RL, et al. Effect of resistance training 65. Kurihara T, Oda T, Chino K, et al. Use of three-dimensionalon muscle use during exercise. J Appl Physiol 1994; 76: ultrasonography for the analysis of the fascicle length of1675-81 human gastrocnemius muscle during contractions. Int J Sport
45. Enoka RM. Neural adaptations with chronic physical activity. J Health Sci 2005; 3: 226-34Biomech 1997; 30: 447-55 66. Muramatsu T, Muraoka T, Kawakami Y, et al. In vivo determi-
46. Chilibeck PD, Calder AW, Sale DG, et al. A comparison of nation of fascicle curvature in contracting human skeletalstrength and muscle mass increases during resistance training muscles. J Appl Physiol 2002; 92: 129-34in young women. Eur J Appl Physiol 1998; 77: 170-5 67. Reeves ND, Narici MV. Behavior of human muscle fascicles
47. Hickson RC, Hidaka K, Foster C, et al. Successive time courses during shortening and lengthening contractions in vivo. J Applof strength development and steroid hormone responses to Physiol 2003; 95: 1090-6heavy-resistance training. J Appl Physiol 1994; 76: 663-70 68. Finni T, Ikegawa S, Komi PV. Concentric force enhancement
48. Hakkinen K, Komi PV, Alen M, et al. EMG, muscle fibre and during human movement. Acta Physiol Scand 2001; 173:force production characteristics during a 1 year training period 369-77in elite weight-lifters. Eur J Appl Physiol 1987; 56: 419-27 69. Finni T, Ikegawa S, Lepola V, et al. Comparison of force-
49. Gaudy JF, Zouaoui A, Bravetti P, et al. Functional anatomy of velocity relationships of vastus lateralis muscle in isokineticthe human temporal muscle. Surg Radiol Anat 2001; 23: and in stretch-shortening cycle exercises. Acta Physiol Scand389-98 2003; 177: 483-91
50. Gaudy JF, Zouaoui A, Bravetti P, et al. Functional organization 70. Izquerdo M, Ibanez J, Hakkinen K, et al. Maximal strength andof the human masseter muscle. Surg Radiol Anat 2000; 22: power, muscle mass, endurance and serum hormones in181-90 weightlifters and road cyclists. J Sport Sci 2004; 22: 465-78
51. Lam EW, Hannam AG, Christiansen EL. Estimation of tendon- 71. Izquierdo M, Hakkinen K, Gonzalez-Badillo JJ, et al. Effects ofplane orientation within human masseter muscle from recon- long-term training specificity on maximal strength and powerstructed magnetic resonance images. Arch Oral Biol 1991; 36: of the upper and lower extremities in athletes from different845-53 sports. Eur J Appl Physiol 2002; 87: 264-71
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

1016 Blazevich
72. Coyle EF. Improved muscular efficiency displayed as Tour de 92. Aratow M, Ballard RE, Crenshaw AG, et al. IntramuscularFrance champion matures. J Appl Physiol 2005; 98: 2191-6 pressure and electromyography as indexes of force during
isokinetic exercise. J Appl Physiol 1993; 74: 2634-4073. Alexander MJL. The relationship between muscle strength andsprint kinematics in elite sprinters. Can J Sport Sci 1989; 14: 93. Degens H, Salmons S, Jarvis JC. Intramuscular pressure, force148-57 and blood flow in rabbit tibialis anterior muscles during single
and repetitive contractions. Eur J Appl Physiol Occup Physiol74. Taylor NAS, Cotter JD, Stanley SN, et al. Functional torque-1998; 78: 13-9velocity and power-velocity characteristics of elite athletes.
Eur J Appl Physiol 1991; 62: 116-21 94. Jarvinen MJ, Einola SA, Virtanen EO. Effect of the position ofimmobilization upon the tensile properties of the rat gas-75. Blazevich AJ, Jenkins DG. Physical performance differencestrocnemius muscle. Arch Phys Med Rehab 1992; 73: 253-7between weight-trained sprinters and weight trainers. J Sci
Med Sport 1998; 1: 12-21 95. Sejersted OM, Hagens AR, Kardel KR, et al. Intramuscular fluid76. Sleivert GG, Backus RD, Wenger HA. Neuromuscular differ- pressure during isometric contraction of human skeletal mus-
ences between volleyball players, middle distance runners and cle. J Appl Physiol 1984; 56: 287-95untrained controls. Int J Sport Med 1995; 16: 390-8 96. Akima H, Kuno S, Suzuki Y, et al. Effects of 20 days of bed rest
77. Kumagai K, Abe T, Brechue WF, et al. Sprint performance is on physiological cross-sectional area of human thigh and legrelated to muscle fascicle length in male 100-m sprinters. J muscles evaluated by magnetic resonance imaging. J GravitAppl Physiol 2000; 88: 811-6 Physiol 1997; 4: S15-21
78. Abe T, Fukashiro S, Harada Y, et al. Relationship between 97. Kawakami Y, Akima H, Kubo K, et al. Changes in muscle size,sprint performance and muscle fascicle length in female sprint- architecture, and neural activation after 20 days of bed resters. J Physiol Anthropol Appl Human Sci 2001; 20: 141-7 with and without resistance exercise. Eur J Appl Physiol 2001;
79. Abe T, Kumagai K, Brechue WF. Fascicle length of leg muscles 84: 7-12is greater in sprinters than distance runners. Med Sci Sports 98. Kawakami Y, Muraoka Y, Kubo K, et al. Changes in muscleExerc 2000; 32: 1125-9 size and architecture following 20 days of bed rest. J Gravit
80. Brechue WF, Abe T. The role of FFM accumulation and skeletal Physiol 2000; 7: 53-60muscle architecture in powerlifting performance. Eur J Appl 99. Yamashita-Goto K, Okuyama R, Honda M, et al. Maximal andPhysiol 2002; 86: 327-36 submaximal forces of slow fibers in human soleus after bed
81. Kearns CF, Abe T, Brechue WF. Muscle enlargement in sumo rest. J Appl Physiol 2001; 91: 417-24wrestlers includes increased muscle fascicle length. Eur J Appl 100. Andersen JL, Gruschy-Knudsen T, Sandri C, et al. Bed restPhysiol 2000; 83: 289-96 increases the amount of mismatched fibers in human skeletal
82. Gondin J, Guette M, Ballay Y, et al. Electromyostimulation muscle. J Appl Physiol 1999; 86: 455-60training effects on neural drive and muscle architecture. Med 101. Heslinga JW, Huijing PA. Effects of short length immobiliza-Sci Sports Exerc 2005; 37: 1291-9 tion of medial gastrocnemius muscle of growing young adult
83. Alegre LM, Jimenez F, Gonzalo-Orden JM, et al. Effects of rats. Eur J Morphol 1992; 30: 257-73dynamic resistance training on fascicle length and isometric
102. Bleakney R, Maffulli N. Ultrasound changes to intramuscularstrength. J Sports Sci 2006; 24: 501-8architecture of the quadriceps following intramedullary nail-
84. Blazevich AJ, Gill ND, Deans N, et al. Lack of human muscle ing. J Sports Med Phys Fit 2002; 42: 120-5architectural adaptation after short-term strength training.
103. Abe T, Kawakami Y, Suzuki Y, et al. Effects of 20 days bed restMuscle Nerve. Epub 2006 Oct 12on muscle morphology. J Gravit Physiol 1997; 4: S10-4
85. Hakkinen K, Kallinen M, Linnamo V, et al. Neuromuscular104. Baker JH, Matsumoto DE. Adaptation of skeletal muscle toadaptations during bilateral versus unilateral strength training
immobilization in a shortened position. Muscle Nerve 1988;in middle-aged and elderly men and women. Acta Physiol11: 231-44Scand 1996; 158: 77-88
105. Hayat A, Tardieu C, Tabary JC, et al. Effects of denervation on86. Tanaguchi Y. Laterality specificity in resistance training: thethe reduction of sarcomere number in cat soleus muscle immo-effect of bilateral and unilateral training. Eur J Appl Physiolbilized in shortened position during seven days. J Physiol1997; 75: 144-50(Paris) 1978; 74: 563-787. Weir JP, Housh DJ, Housh TJ, et al. The effect of unilateral
106. Tabary JC, Tabary C, Tardieu C, et al. Physiological and struc-concentric weight training and detraining on joint angle speci-tural changes in the cat’s soleus muscle due to immobilizationficity, cross-training, and the bilateral deficit. J Orthop Sportsat different lengths by plaster casts. J Physiol 1972; 224:Phys Ther 1997; 25: 264-70231-4488. Jarvholm U, Palmerud G, Karlsson D, et al. Intramuscular
107. Williams PE. Use of intermittent stretch in the prevention ofpressure and electromyography in four shoulder muscles. Jserial sarcomere loss in immobilised muscle. Ann Rheum DisOrthop Res 1991; 9: 609-191990; 49: 316-789. Sejersted OM, Hagens AR. Intramuscular pressures for moni-
108. Williams PE, Goldspink G. Changes in sarcomere length andtoring different tasks and muscle conditions. Adv Expl Medphysiological properties in immobilized muscle. J Anat 1978;Biol 1995; 384: 339-50127: 459-6890. Miura H, McCully K, Nioka S, et al. Relationship between
109. Nicks DK, Beneke WM, Key RM, et al. Muscle fibre size andmuscle architectural features and oxygenation status deter-number following immobilisation atrophy. J Anat 1989; 163:mined by near infrared device. Eur J Appl Physiol 2004; 91:1-5273-8
91. Ballard RE, Watenpaugh DE, Breit GA, et al. Leg intramuscular 110. Leterme D, Cordonnier C, Mounier Y, et al. Influence of chron-pressures during locomotion in humans. J Appl Physiol 1998; ic stretching upon rat soleus muscle during non-weight-bear-84: 1976-81 ing conditions. Eur J Physiol 1994; 429: 274-9
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)

Plasticity of Fascicle Geometry 1017
111. Herbert RD, Balnave RJ. The effect of position of immobilisa- 125. Roos MR, Rice CL, Connelly DM, et al. Quadriceps muscletion on resting length, resting stiffness, and weight of the strength, contractile properties, and motor unit firing rates insoleus muscle of the rabbit. J Orthop Res 1993; 11: 358-6 young and old men. Muscle Nerve 1999; 22: 1094-103
112. Binzoni T, Bianchi S, Hanquinet S, et al. Human gastrocnemius 126. Valour D, Ochala J, Ballay Y, et al. The influence of ageing onmedialis pennation angle as a function of age: from newborn to the force-velocity-power characteristics of human elbow flex-the elderly. J Physiol Anthropol Appl Hum Sci 2001; 20: 293-8
or muscles. Exp Gerontol 2003; 38: 387-95113. Hakkinen K, Pakarinen A. Muscle strength and serum hormones
127. Newton RU, Hakkinen K, Hakkinen A, et al. Mixed-methodsin middle-aged and elderly men and women. Acta Physiol
resistance training increases power and strength of young andScand 1993; 148: 199-207older men. Med Sci Sports Exerc 1999; 34: 1367-75
114. Izquierdo M, Ibanez J, Gorostiaga E, et al. Maximal strength128. Kubo K, Kanehisa H, Azuma K, et al. Muscle architecturaland power characteristics in isometric and dynamic actions of
characteristics in women aged 20-79 years. Med Sci Sportsthe upper and lower extremities in middle-aged and older men.Acta Physiol Scand 1999; 167: 57-68 Exerc 2003; 35: 39-44
115. Izquerdo M, Hakkinen K, Anton A, et al. Maximal strength and 129. Kubo K, Kanehisa H, Azuma K, et al. Muscle architecturalpower, endurance performance, and serum hormones in mid- characteristics in young and elderly men and women. Int Jdle-aged and elderly men. Med Sci Sports Exerc 2001; 33: Sports Med 2003; 24: 125-301577-87
130. Morse CI, Thom JM, Birch KM, et al. Changes in triceps surae116. Klitgaard H, Mantoni M, Schiaffino S, et al. Function, morphol- muscle architecture with sarcopenia. Acta Physiol Scand 2005;
ogy and protein expression of ageing skeletal muscle: a cross-183: 291-8sectional study of elderly men with different training back-
grounds. Acta Physiol Scand 1990; 140: 41-54 131. Hakkinen K, Kraemer WJ, Pakarinen A, et al. Effects of heavy
resistance/power training on maximal strength, muscle mor-117. Narici MV, Maganaris CN, Reeves ND, et al. Effect of aging onphology, and hormonal response patterns in 60-75-year-oldhuman muscle architecture. J Appl Physiol 2003; 95: 2229-34men and women. Can J Appl Physiol 2002; 27: 213-31118. Lexell J, Taylor CC, Sjostrom M. What is the cause of the aging
132. Izquerdo M, Hakkinen K, Ibanez J, et al. Effects of strengthatrophy? Total number, size and proportion of different fibertypes studies in whole vastus lateralis muscle from 15- to training on submaximal and maximal endurance performance83-year-old men. J Neurol Sci 1988; 84: 275-94 capacity in middle-aged and older men. J Strength Cond Res
119. Hortobagyi T, Zheng D, Weidner M, et al. The influence of 2003; 17: 129-39aging on muscle strength and muscle fiber characteristics with 133. Reeves ND, Narici MV, Maganaris CN. Effect of resistancespecial reference to eccentric strength. J Gerontol A: Biol Sci
training on skeletal muscle-specific force in elderly humans. JMed Sci 1995; 50: B399-406Appl Physiol 2004; 96: 885-92
120. Larsson L, Grimby G, Karlsson J. Muscle strength and speed of134. Trappe S, Williamson D, Godard M, et al. Effect of resistancemovement in relation to age and muscle morphology. J Appl
training on single muscle fiber contractile function in olderPhysiol 1979; 46: 451-6men. J Appl Physiol 2000; 89: 143-52121. Poggi P, Marchettic C, Scelsi R. Automatic morphometric anal-
135. Lynn R, Morgan DL. Decline running produced more sarcomer-ysis of skeletal muscle fibers in the aging man. Anat Rec 1987;217: 30-4 es in rat vastus intermedius muscle fibers than does incline
running. J Appl Physiol 1994; 77: 1439-44122. Hakkinen K, Alen M, Kallinen M, et al. Neuromuscular adapta-tion during prolonged strength training, detraining and re- 136. Butterfield TA, Leonard TR, Herzog W. Differential serialstrength-training in middle-aged and elderly people. Eur J sarcomere number adaptations in knee extensor muscles of ratsAppl Physiol 2000; 83: 51-62
is contraction type dependent. J Appl Physiol 2005; 99: 1352-8123. Hakkinen K, Kallinen M, Izquierdo M, et al. Changes in ago-
nist-antagonist EMG, muscle CSA, and force during strengthtraining in middle-aged and older people. J Appl Physiol 1998; Correspondence and offprints: Dr Anthony J. Blazevich,84: 1341-9
Centre for Sports Medicine and Human Performance,124. Knight CA, Kamen G. Adaptations in muscular activation of the
Brunel University, Uxbridge, UB8 3PH, UK.knee extensor muscles with strength training in young andE-mail: [email protected] adults. J Electromyogr Kinesiol 2001; 11: 405-12
2006 Adis Data Information BV. All rights reserved. Sports Med 2006; 36 (12)