biological pretreatment of cellulose: enhancing enzymatic hydrolysis rate using cellulose-binding...
TRANSCRIPT

Bioresource Technology 102 (2011) 2910–2915
Contents lists available at ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Biological pretreatment of cellulose: Enhancing enzymatic hydrolysisrate using cellulose-binding domains from cellulases
Mélanie Hall a, Prabuddha Bansal b, Jay H. Lee c, Matthew J. Realff b, Andreas S. Bommarius b,⇑a Department of Chemistry, Organic and Bioorganic Chemistry, Heinrichstraße 28, 8010 Graz, Austriab School of Chemical and Biomolecular Engineering, Georgia Institute of Technology, 311 Ferst Drive, N.W., Atlanta, GA 30332-0100, USAc Department of Chemical and Biomolecular Engineering, Korea Advanced Institute of Science and Technology (KAIST), 291 Daehak-ro, Yuseong-gu, Daejeon 305-701, Korea
a r t i c l e i n f o a b s t r a c t
Article history:Received 13 August 2010Received in revised form 27 October 2010Accepted 1 November 2010Available online 9 November 2010
Keywords:Cellulose-binding domainsBiofuelCellulose hydrolysisPretreatmentCrystallinity
0960-8524/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.biortech.2010.11.010
⇑ Corresponding author. Tel.: +1 404 385 1334; faxE-mail address: [email protected].
In this study, cellulose-binding domains (CBDs) of cellulases from Trichoderma reesei were used in a pre-treatment step and were found to effectively reduce the crystallinity of cellulose (both Avicel and fibrouscellulose). This, in turn, led to higher glucose concentrations (up to 25% increase) in subsequent hydro-lysis of cellulose using a mixture of cellulases and without the need for any intermediate purificationstep. CBDs were shown to be active in a range of temperatures (up to 50 �C), while cellulase hydrolyticactivity was greatly reduced after incubation at 50 �C. This was explained by retention of full bindingcapacity after incubation at 50 �C for 15 h. Our findings suggest that CBDs may be a valuable tool in pre-treating cellulose and eventually afford faster enzymatic conversion of cellulose to glucose, thus contrib-uting to more affordable processes in the production of biofuels.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
The current and urgent need for alternative renewable energiesto supplement petroleum-based fuels has led to the developmentof various technologies based on wind, solar, tidal energy or bio-mass. The use of biomass as a source for biofuels relies on the greatabundance of cellulose, as it is one of the most abundant sources oforganic material on our planet. Currently, widespread productionof biofuels from lignocellulosic materials is prevented by highprocessing costs. To render lignocellulosic biofuels more competi-tive, improvements in the enzymatic degradation of cellulose arecrucial (Himmel et al., 2007; Lynd et al., 2008). The enzymatichydrolysis of cellulose, a polysaccharide consisting of repeatedcellobiose units (glucose dimers), requires the synergistic actionof various cellulases (endoglucanases (EC 3.2.1.4), exoglucanases(EC 3.2.1.91) and b-glucosidase (EC 3.2.1.21)) to eventually pro-duce glucose, a valuable key intermediate that can be fermentedto bioalcohols or transformed into various other chemicals (Daviesand Henrissat, 1995; Lynd et al., 2002). Various process limitationsaccount for overall slow reaction and decreasing rates as the con-version proceeds, and can be directly related to cellulose properties(insoluble nature, crystallinity, surface area, porosity, degree ofpolymerization) and enzymatic features (deactivation, inhibition,
ll rights reserved.
: +1 404 894 2295.edu (A.S. Bommarius).
jamming, clogging, adsorption, processivity) (Bansal et al., 2009;Hall et al., 2010; Zhang and Lynd, 2004).
Cellulases are glycosyl hydrolases of greatly varying structure,though they all catalyze the hydrolysis of 1,4-b-D-glycosidic bonds(Davies and Henrissat, 1995). Most cellulases consist of three do-mains: a catalytic domain (the locus of hydrolysis), a cellulose-binding domain (that anchors the whole enzyme onto the cellulosesurface and orients the cellulose fiber towards the tunnel contain-ing the active site) and a glycosylated flexible linker connecting thetwo other domains (and providing sufficient spatial separation re-quired between them; it allows processive motion and likely alsoplays a role in energy storage) (Gilkes et al., 1991; Zhao et al.,2008). Cellulose-binding domains (CBDs) of cellulases from Tricho-derma reesei (belonging to family I CBD) form a wedge-like fold,where a flat face provides key (aromatic) residues strongly inter-acting with crystalline cellulose (Linder et al., 1995; Takashimaet al., 2007). Recent investigation of the effect of CBDs (withoutthe catalytic unit) on cellulose provides evidence that family ICBD may play a role in the hydrolysis of crystalline cellulose byweakening and splitting off the hydrogen bond network in the cel-lulose crystal (Gao et al., 2001; Wang et al., 2008; Xiao et al., 2001).Given our latest study on the role of cellulose crystallinity in deter-mining hydrolysis rate (Hall et al., 2010), and given the processivenature of cellulases (Beckham et al., 2010), we hypothesized thatCBDs could be used in a separate step prior to the hydrolysis,and without the requirement for change of solvent, such as inthe case of pretreatment with ionic liquids (Brennan et al., 2010;

M. Hall et al. / Bioresource Technology 102 (2011) 2910–2915 2911
Zhao et al., 2009) or organic solvents such as N-methyl-morpholine-N-oxide (NMMO) (Jeihanipour et al., 2010). Our goalwas to render cellulose less recalcitrant to hydrolysis by havingCBDs interact with the cellulose surface. Here, we report the effectof CBD on the structure of crystalline cellulose and its direct impli-cation in the subsequent hydrolysis step with full-lengthcellulases.
2. Methods
2.1. General
Avicel PH-101, fibrous cellulose from cotton linters (SigmaC6288, medium), cellulases from T. reesei (159 FPU mL�1)), andb-glucosidase (from almonds, 2.32 U mg�1)) were obtained fromSigma (St. Louis, MO, USA). The (micro) BCA protein assay kit andthe Coomassie (Bradford) protein assay kit were obtained fromThermo Fischer Scientific (Rockford, IL, USA). Papain was obtainedfrom Fluka as lyophilized powder from Carica papaya(9.84 U mg�1). Centrifugal devices with polyethersulfone mem-branes were from Pall Life Sciences (Ann Arbor, MI, USA). All exper-iments and assays were run at least in duplicate.
2.2. Cel7A purification
Cel7A was purified from T. reesei cellulase cocktail by means ofanion-exchange chromatography as previously published (Hallet al., 2010). Purity was confirmed by SDS–PAGE. After purification,Cel7A buffer was exchanged to sodium acetate buffer (50 mM, pH5) using a polyethersulfone membrane (molecular weight cut-offof 10 kDa) in a Macrosep device.
2.3. Cellulase proteolysis and CBD isolation
Cellulase cleavage using papain was adapted from publishedprocedures (Lemos et al., 2000). Cellulase preparation (mixturefrom T. reesei) was diluted (5�) in sodium acetate buffer (50 mM,pH 5) and concentrated in a Jumbosep centrifugal device (polye-thersulfone membrane, molecular weight cut-off of 30 kDa) to re-move smaller molecules. Papain was activated in ammoniumacetate buffer (50 mM, pH 6) at 30 �C and 170 rpm for 30 minand added to the cellulase solution (w/w 1:5, v/v 1:5). The diges-tion was performed at 30 �C for 2 h (completion checked viaSDS–PAGE). The mixture was then filtered through 30 kDa mem-brane and both filtrate and concentrate were analyzed for proteincontent using the micro BCA assay.
2.4. X ray diffraction and crystallinity measurement
X ray diffraction patterns of cellulose samples (buffer and CBDpretreated) obtained after freeze-drying (at �55 �C and 0.03 mbarfor 16 h) were recorded with an X’Pert PRO X ray diffractometer(PANanalytical BV, Almelo, the Netherlands) at room temperaturefrom 10 to 35 �C, using Cu/Ka1 irradiation (1.54 Å) at 45 kV and40 mA. The scan speed was 0.021425� s�1 with a step size of0.0167�. The crystallinity index of cellulose was measured from Xray diffraction data using an analytical method developed previ-ously (Bansal et al., 2010).
2.5. Cellulose pretreatment and hydrolysis
In a typical experiment, CBDs were added in various amounts(up to 12 lg mg�1) to cellulose (Avicel or fibrous cellulose, total56 mg, 40 mg mL�1) in sodium acetate buffer (50 mM, pH 5) in aglass vial (containing a stir bar). The mixture was incubated for15 h at various temperatures in a shaker incubator at 170 rpm
and was then diluted with buffer to lower cellulose concentrationto 20 mg mL�1 for subsequent hydrolysis. Hydrolysis was initiatedafter the pretreatment step by adding cellulases (24 mL L�1,3.4 g L�1 total protein) and b-glucosidase (15 kU L�1) to the reac-tion mixture, and was carried out at 50 �C and 170 rpm. The courseof the reaction was monitored by performing DNS assay (Hall et al.,2010) on small aliquots taken after 30 min, 1 and 2.5 h.
For the thermostability experiment, cellulases and catalytic do-mains were incubated at 50 �C for 15 h and then added(300 lg mL�1 and 125 lg mL�1, respectively), to a cellulose mix-ture (Avicel, 20 mg mL�1 in sodium acetate buffer, 50 mM, pH 5)that had been incubated at 50 �C for 1 h at 900 rpm. The hydrolysiswas run at 50 �C and 900 rpm and the conversion was monitoredafter 6 and 22 h by performing DNS assay on the supernatant.
2.6. Enzyme adsorption experiments
Avicel samples (20 mg mL�1) in sodium acetate buffer (50 mM,pH 5) were incubated at 50 �C for 1 h at 900 rpm, and then cooleddown to 4 �C. Cellulases (which had been incubated in sodium ace-tate buffer at 50 �C for 15 h or stored at 4 �C since purchase) wereadded in various amounts and the mixture was further agitated for30 min at 4 �C. After centrifugation, the supernatant was collectedand protein content analysis was performed using the BCA proteinassay kit.
2.7. MALDI-TOF mass spectrometry
Purified CBD was obtained as described above, except that thebuffer was ammonium acetate 50 mM, pH 6.0 (used in place of so-dium acetate in order to improve the signal-to-noise ratio of thepeaks). MALDI-TOF MS was performed on an Applied Biosystems4700 Proteomics Analyzer. The spectra were acquired in linearmode, and the matrix used was alpha-cyano-4-hydroxycinnamicacid (CHCA). A 10 mg mL solution of the matrix was made in50:50 (v:v) water:acetonitrile containing 0.1% formic acid by vol-ume. Equal volumes of the matrix solution and the CBD solutionwere mixed, and the resulting solution was spotted onto theMALDI plate. A total of 2000 laser shots were combined to producethe final spectrum.
3. Results and discussion
3.1. Cellulase proteolysis and CBD isolation
Both cellulase mixture (endoglucanases and exoglucanases) andpurified cellobiohydrolase Cel7A from T. reesei were used in theproteolysis experiment using papain as cleaving agent. Cleavagewas complete in both cases (as observed by the disappearance offull-length enzymes) and isolation of CBD through filtrationyielded a mixture of CBDs or pure CBDCel7A, and correspondingcatalytic domains (CDs) from the retentate could be observed bySDS–PAGE (Fig. 1). Silver-staining (known to be a highly sensitivedetection method) was also employed with the SDS–PAGE andshowed that the CBD samples were not contaminated by other lar-ger species (that could have leaked through the filter (data notshown)). Additionally, the CBD solution was evaluated by MALDI-TOF mass spectrometry to ascertain the purity of the sample aswell as the size of the molecule (CBD with linker). A mass distribu-tion between 8.4 and 9.4 kDa was obtained (overall mass corre-sponding to the difference between Cel7A and CD masses onFig. 1), with peaks separated by �162 Da (respective major peaks:8426.3, 8589.0, 8750.6, 8913.4, 9074.4, 9235.0, 9399.0 Da;spectrum not shown). This mass corresponds to the molecularweight of hexose sugars that are O-linked to the linker residues

Fig. 1. SDS–PAGE before and after papain cleavage of cellulases. Lane 1, ladder; lane2, cellulase cocktail from Trichoderma reesei after filtration through 30 kDamembrane; lane 3, catalytic domains (CDs) of cellulases; lane 4, purified Cel7Aafter anion-exchange chromatography; lane 5, CD of purified Cel7A (to rule out thepresence of impurities and other species, samples from lanes 2 and 3 wereoverloaded on SDS–PAGE and appeared unchanged, data not shown).
Table 1Effect of 15 h incubation with buffer and cellulose-binding domains on cellulosecrystallinity (CrI) as determined via X ray diffraction (Bansal et al., 2010).
Cellulose substrate Avicel (%) Fibrous cellulose
CrI on dry sample (untreated) 60 72%CrI after incubation in buffera 65 74%CrI after pretreatment with CBDs 58 68%CrI after pretreatment with CBDCel7A
b 53–56 n.d.CrI after incubation with CDCel7A
b 64 n.d.
n.d. Not determined.a Sodium acetate (pH 5.0, 50 mM).b Longer incubation times (>48 h).
2912 M. Hall et al. / Bioresource Technology 102 (2011) 2910–2915
(distribution pattern observed previously (Jeoh et al., 2008)). Nohigher mass could be detected so that the CBD solution was con-sidered pure and not contaminated by CD or Cel7A.
3.2. Cellulose incubation with CBDs and crystallinity
There have been a few reports on the effect of CBD binding oncellulose (mostly ramie fibers, Whatman cellulose fibers, cotton fi-bers, Avicel or filter paper). The binding of CBD on cellulose wasshown to have an influence on the hydrogen bond network ofthe cellulose crystal, by weakening and splitting of bonds (asshown via FT-IR) (Ciolacu et al., 2010; Gao et al., 2001; Xiaoet al., 2001). Given the nature of crystalline cellulose, intermolecu-lar hydrogen bonds are the key to a tight crystal and one of the rea-sons for its recalcitrance to enzymatic attack. Also, it has beenproposed that CBDs help the overall hydrolysis process by pullingcellulose chains from the crystal and feeding them into the cata-lytic domain, where they are hydrolyzed (Arantes and Saddler,2010) (which might explain why the overall crystallinity ofcellulose remains unchanged during the conversion (Hall et al.,2010)). The source of energy required for the processive motionis still unclear but it makes sense to assume that it partly stemsfrom the catalytic event itself (Sinnott, 1998). Employing CBDs inan incubation step would therefore require some form of energyso that they could possibly move along (or through) the cellulosesurface and affect the overall structure of the crystal.
In the experiments reported here, vigorous stirring was pro-vided using a shaker incubator and the addition of a stir bar tothe system. Avicel (CrI 60%) and fibrous cellulose (CrI 72%) wereemployed, as their high crystalline content allows for easier mon-itoring of any possible change occurring during pretreatment. Afterpretreatment of cellulose with CBDs in buffer at 42 �C for 15 h un-der vigorous agitation, the samples were collected and their crys-tallinity content was assessed by X ray diffraction (XRD), whilethe supernatant was analyzed for the presence of soluble reducingends. No reducing ends were released from the buffer control sam-ple. While the amount of soluble reducing ends obtained with
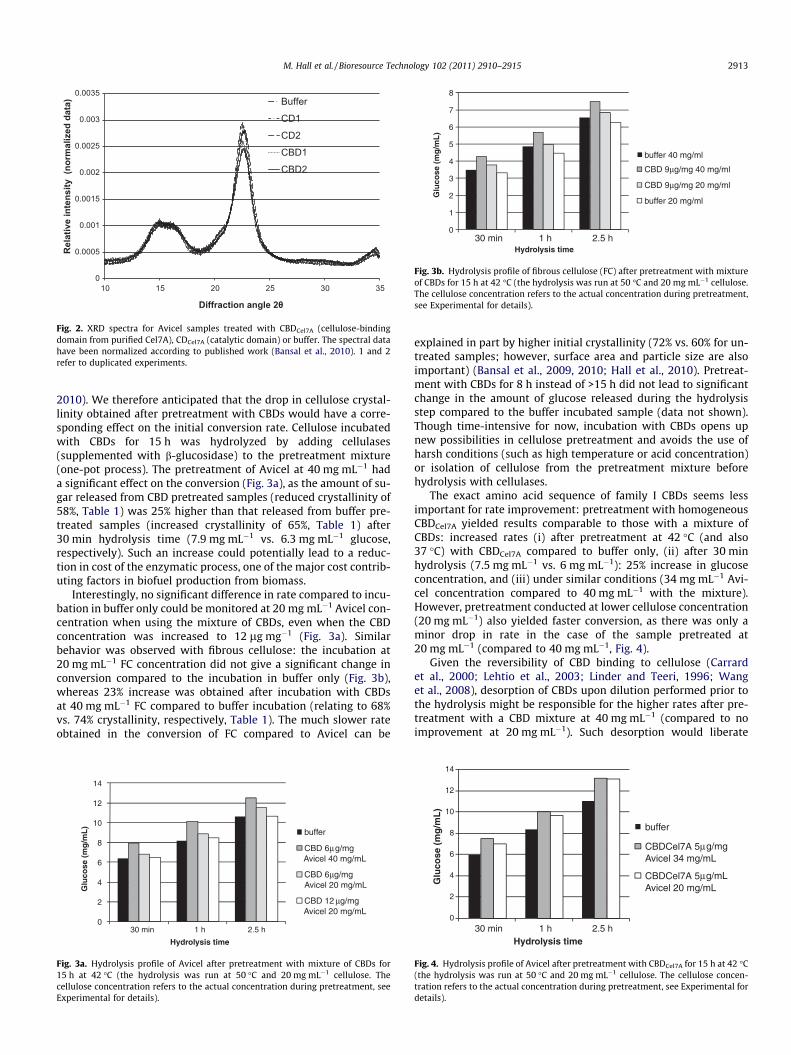
CBDs was very low (<120 lg mL�1, starting from cellulose concen-tration at 20 mg mL�1), it was consistent and above buffer back-ground. Family I CBDs most likely do not have any hydrolyticactivity; rather, small soluble oligomers were released from thecellulose surface upon binding/processing of CBDs. No disruptingactivity could be observed (i.e. no agglomerate formation nor fasterprecipitation without CBDs), as opposed to what has been reportedfor CBDs from other families (release of insoluble small particles/short fibers) (Din et al., 1991; Gao et al., 2001). After incubationin buffer only (control), the crystallinity of both types of cellulose(Avicel and fibrous) was shown to have slightly increased (Table 1),a phenomenon referred to as ‘‘recrystallization’’ (Ouajai andShanks, 2006; Wormald et al., 1996). The gain was noticeable(from 60% up to 65–66% on average for Avicel, and 72% up to74% for fibrous cellulose) and cannot be attributed to the removalof non-crystalline material from cellulose, as no soluble reducingends were released in the buffer control sample. Also, the freeze-drying has been shown to have no effect on cellulose crystallinity(Bansal et al., 2010; Hall et al., 2010) so that only extended incuba-tion in buffer resulted in such a dramatic increase in crystallinityindex (additionally, both buffer and CBD pretreated samples werefreeze-dried before XRD analysis). The use of CBDs (up to10 lg mg�1 at 40 mg mL�1 cellulose in a typical pretreatment step)resulted in a different trend, as it not only prevented the substratefrom recrystallizing, but also reduced the initial crystallinity of cel-lulose. Avicel crystallinity reached 58% after incubation with a mix-ture of CBDs, while fibrous cellulose crystallinity reached 68%, asignificant drop from the crystallinity indices obtained after incu-bation in buffer only. When cellulose was incubated with pureCBDs from cellobiohydrolase I Cel7A (CBDCel7A) over extensiveperiods of incubation (>48 h), an even higher reduction in crystal-linity was obtained with Avicel (down to 53%, Fig. 2). Though thedifference between the spectra appears limited to one portion ofthe XRD patterns, our analytical method recently developed forinterpretation of XRD data of cellulose allowed accurate determi-nation of the crystallinity index. The drop in intensity observedat a Bragg angle of 22.5� ((2 0 0) plane) accounts for the overallreduction in crystallinity. Thus, CBDs from T. reesei (belonging tofamily I CBD, along with CBDs of cellulases from Phanerochaetechrysosporium or Penicillium janthinellum), though not catalyticallyactive with cellulose, had a strong effect on the cellulose structure,as overall crystallinity was significantly reduced. Interestingly, thecrystal plane (2 0 0) seems to be the only one affected by thesechanges (Fig. 2). Overall, CBDs are responsible for splitting of inter-molecular hydrogen bonds to such an extent that the cellulosecrystallinity is noticeably reduced. This most likely proceeds via adynamic process (processive motion on cellulose surface required),which is still poorly understood.
3.3. Hydrolysis of CBD pretreated cellulose
The degree of crystallinity of cellulose has been shown to corre-late inversely with initial enzymatic hydrolysis rates (Hall et al.,
0
0.0005
0.001
0.0015
0.002
0.0025
0.003
0.0035
10 15 20 25 30 35
Rel
ativ
e in
tens
ity (
norm
aliz
ed d
ata)
Diffraction angle 2θ
Buffer
CD1
CD2
CBD1
CBD2
Fig. 2. XRD spectra for Avicel samples treated with CBDCel7A (cellulose-bindingdomain from purified Cel7A), CDCel7A (catalytic domain) or buffer. The spectral datahave been normalized according to published work (Bansal et al., 2010). 1 and 2refer to duplicated experiments.
0
1
2
3
4
5
6
7
8
30 min 1 h 2.5 h
Glu
cose
(m
g/m
L)
Hydrolysis time
buffer 40 mg/ml
CBD 9 µg/mg 40 mg/ml
CBD 9 µg/mg 20 mg/ml
buffer 20 mg/ml
Fig. 3b. Hydrolysis profile of fibrous cellulose (FC) after pretreatment with mixtureof CBDs for 15 h at 42 �C (the hydrolysis was run at 50 �C and 20 mg mL�1 cellulose.The cellulose concentration refers to the actual concentration during pretreatment,see Experimental for details).
M. Hall et al. / Bioresource Technology 102 (2011) 2910–2915 2913
2010). We therefore anticipated that the drop in cellulose crystal-linity obtained after pretreatment with CBDs would have a corre-sponding effect on the initial conversion rate. Cellulose incubatedwith CBDs for 15 h was hydrolyzed by adding cellulases(supplemented with b-glucosidase) to the pretreatment mixture(one-pot process). The pretreatment of Avicel at 40 mg mL�1 hada significant effect on the conversion (Fig. 3a), as the amount of su-gar released from CBD pretreated samples (reduced crystallinity of58%, Table 1) was 25% higher than that released from buffer pre-treated samples (increased crystallinity of 65%, Table 1) after30 min hydrolysis time (7.9 mg mL�1 vs. 6.3 mg mL�1 glucose,respectively). Such an increase could potentially lead to a reduc-tion in cost of the enzymatic process, one of the major cost contrib-uting factors in biofuel production from biomass.
Interestingly, no significant difference in rate compared to incu-bation in buffer only could be monitored at 20 mg mL�1 Avicel con-centration when using the mixture of CBDs, even when the CBDconcentration was increased to 12 lg mg�1 (Fig. 3a). Similarbehavior was observed with fibrous cellulose: the incubation at20 mg mL�1 FC concentration did not give a significant change inconversion compared to the incubation in buffer only (Fig. 3b),whereas 23% increase was obtained after incubation with CBDsat 40 mg mL�1 FC compared to buffer incubation (relating to 68%vs. 74% crystallinity, respectively, Table 1). The much slower rateobtained in the conversion of FC compared to Avicel can be
0
2
4
6
8
10
12
14
Glu
cose
(m
g/m
L)
Hydrolysis time
buffer
CBD 6 µg/mg Avicel 40 mg/mL
CBD 6 µg/mg Avicel 20 mg/mL
CBD 12 µg/mg Avicel 20 mg/mL
30 min 1 h 2.5 h
Fig. 3a. Hydrolysis profile of Avicel after pretreatment with mixture of CBDs for15 h at 42 �C (the hydrolysis was run at 50 �C and 20 mg mL�1 cellulose. Thecellulose concentration refers to the actual concentration during pretreatment, seeExperimental for details).
explained in part by higher initial crystallinity (72% vs. 60% for un-treated samples; however, surface area and particle size are alsoimportant) (Bansal et al., 2009, 2010; Hall et al., 2010). Pretreat-ment with CBDs for 8 h instead of >15 h did not lead to significantchange in the amount of glucose released during the hydrolysisstep compared to the buffer incubated sample (data not shown).Though time-intensive for now, incubation with CBDs opens upnew possibilities in cellulose pretreatment and avoids the use ofharsh conditions (such as high temperature or acid concentration)or isolation of cellulose from the pretreatment mixture beforehydrolysis with cellulases.
The exact amino acid sequence of family I CBDs seems lessimportant for rate improvement: pretreatment with homogeneousCBDCel7A yielded results comparable to those with a mixture ofCBDs: increased rates (i) after pretreatment at 42 �C (and also37 �C) with CBDCel7A compared to buffer only, (ii) after 30 minhydrolysis (7.5 mg mL�1 vs. 6 mg mL�1): 25% increase in glucoseconcentration, and (iii) under similar conditions (34 mg mL�1 Avi-cel concentration compared to 40 mg mL�1 with the mixture).However, pretreatment conducted at lower cellulose concentration(20 mg mL�1) also yielded faster conversion, as there was only aminor drop in rate in the case of the sample pretreated at20 mg mL�1 (compared to 40 mg mL�1, Fig. 4).
Given the reversibility of CBD binding to cellulose (Carrardet al., 2000; Lehtio et al., 2003; Linder and Teeri, 1996; Wanget al., 2008), desorption of CBDs upon dilution performed prior tothe hydrolysis might be responsible for the higher rates after pre-treatment with a CBD mixture at 40 mg mL�1 (compared to noimprovement at 20 mg mL�1). Such desorption would liberate
0
2
4
6
8
10
12
14
30 min 1 h 2.5 h
Glu
cose
(m
g/m
L)
Hydrolysis time
buffer
CBDCel7A 5 µg/mg Avicel 34 mg/mL
CBDCel7A 5 µg/mL Avicel 20 mg/mL
Fig. 4. Hydrolysis profile of Avicel after pretreatment with CBDCel7A for 15 h at 42 �C(the hydrolysis was run at 50 �C and 20 mg mL�1 cellulose. The cellulose concen-tration refers to the actual concentration during pretreatment, see Experimental fordetails).

2914 M. Hall et al. / Bioresource Technology 102 (2011) 2910–2915
binding spots for cellulases on the cellulose surface and thus in-crease overall hydrolysis rate. However, the experimental limita-tion in differentiating between CBD and cellulase binding and thefairly low amount of CBDs provided during pretreatment renderthis suggestion experimentally difficult to verify. Progress in quan-tification techniques of bound CBDs (Jervis et al., 1997; Pinto et al.,2008, 2006) may shed light on this intriguing behavior.
Though incubation in buffer led to an increase rather than de-crease in overall cellulose crystallinity (Table 1), it provided hydra-tion of the sample and led to nearly 2-fold increase in hydrolysisrate in comparison to employing dry cellulose without pretreat-ment (data not shown). Thus, it seems that the increase in rate ob-tained after pretreatment with CBDs is a combination of hydrationeffect from the buffer and reduction in crystallinity from CBDs.
Increased CBD concentration during pretreatment led to en-hanced production of glucose in the hydrolysis step (Fig. 5). Giventhat 150 lg mg�1 cellulase at total loading of 1230 lg mg�1 bind tothe cellulose surface (Hall et al., 2010), the present loading of
0
2
4
6
8
10
12
14
16
30 min 1 h 2.5 h
Glu
cose
(m
g/m
L)
Hydrolysis time
buffer 40 mg/ml
CBD 3 µg/mg 40 mg/ml
CBD 6 µg/mg 40 mg/ml
CBD 9 µg/mg 40 mg/ml
Fig. 5. Effect of various CBD concentrations in the pretreatment step (15 h at 42 �C)on hydrolysis rate (the hydrolysis was run at 50 �C and 20 mg mL�1 cellulose. Thecellulose concentration refers to the actual concentration during pretreatment, seeExperimental for details).
0
2
4
6
8
10
12
14
16
18
20
30 min 1 h 30 min 1 h 2.5 h
Glu
cose
(m
g/m
L)
No stirbar Large stirbars
buffer
CBD Avicel 40 mg/mL
CBD Avicel 20 mg/mL
Fig. 6. Effect of stir bar during pretreatment of cellulose with CBDs on hydrolysisrates. The pretreatment was run at 42 �C for 15 h without stir bar (10 lg mg�1), orwith large stir bars (7.5 lg mg�1). The hydrolysis was run at 50 �C and 20 mg mL�1
cellulose concentration.
Table 2Effect of 15 h incubation at 50 �C on catalytic activity of cellulases and catalytic domains
Incubation temperature Activity (glucose concentration in mg mL�1)
Cellulase cocktail CDs
4 �C After 6 h: 5.6 After 22 h: 8.2 After 6
50 �C After 6 h: 3.2 After 22 h: 3.6 After 6
9 lg mg�1 is far from saturating the surface. Also, Ståhlberg et al.could not demonstrate saturation of Avicel employing up to470 lg mg�1 CBDCel7A (corresponding to 49 lg mg�1 bound CBD)(Stahlberg et al., 1991).
In the absence of stirring during pretreatment (but when shak-ing was still provided), overall reaction rates were 40% lower andincubation with CBDs surprisingly did not lead to an increase inglucose concentration (Fig. 6). The use of stir bars 72% as large asthe inner diameter of the vials resulted in up to 1.5-fold higherconversion levels in buffer, a positive effect of CBD incubation,but reduced reproducibility (most likely due to inhomogeneousstirring owing to geometric constraints in the reaction vial). Hence-forth, stir bars of 55% of inner vial diameter were used. Likely, thelack of continual convection in the absence of stirring prevents lat-eral displacement of CBD molecules and thus limits their effective-ness, given the absence of catalytic events as a potential energysource for CBD movement along the cellulose surface.
Contrary to previous publications (Esteghlalian et al., 2001;Stahlberg et al., 1991), we showed that preincubation of cellulosewith CBDs lowered its index of crystallinity and resulted in higherglucose concentration in the subsequent hydrolysis reaction. Giventhat experimental conditions are key to achieving improved hydro-lytic rates (vigorous stirring, proper cellulose concentration duringpretreatment, sufficient incubation time), it can be expected thatCBDs from other families may also prove useful in pretreating cel-lulose. Also, simultaneous addition of CBDs to non-pretreated cel-lulose samples did not show any improvement in conversion levels(data not shown), in contrast to the work of Lemos et al. performed,however, at much higher CBD concentration, ca. 4 mg on filter pa-per compared to microcrystalline cellulose (Lemos et al., 2003).This justifies the use of CBDs in a pretreatment step rather thana one-step process combining addition of CBDs and full-length en-zymes. Simultaneous addition of CBDs to the hydrolysis mixturemost likely prevents reduction of crystallinity by CBDs due to com-petitive binding with cellulases and hydrolysis of cellulose bycellulases.
It should be noted that various pretreatment methods are avail-able and have been deeply investigated, but the vast majority ofthem focuses on delignification and removal of hemicellulose, hav-ing as beneficial consequence a decrease in cellulose crystallinity(Cardona et al., 2010). For instance, biological pretreatment ofhardwoods and softwoods using the white-rot fungus Echinodon-tium taxodii resulted in the degradation and alteration of ligninand thus enhanced the overall enzymatic hydrolysis rate by severalfolds, though over non-practical pretreatment times (i.e. severalweeks) (Yu et al., 2009).
3.4. Thermostability of CBDs
Similarly to the conditions used in the enzymatic hydrolysis ofpretreated cellulose, pretreatment with CBDs was also performedat 50 �C. After 15 h pretreatment at 50 �C and hydrolysis for30 min, conversion was only slightly lower than after pretreatmentat 42 �C (data not shown), so that CBDs can be considered as rela-tively thermostable. This observation was confirmed with bindingexperiments, where full-length cellulases were shown to bind
(CDs) and binding of cellulases on Avicel (compared to standard storage at 4 �C).
Binding capacity (bound cellulases lg mg�1)
h: 1.2 After 22 h: 2.2 37.3 (total added 90 lg mg�1)38.4 (total added 150 lg mg�1)
h: 0.5 After 22 h: 0.7 41.4 (total added 90 lg mg�1)44.0 (total added 150 lg mg�1)

M. Hall et al. / Bioresource Technology 102 (2011) 2910–2915 2915
similarly to Avicel after 15 h of incubation at 50 �C, compared tostorage at 4 �C (Table 2). The activity of cellulases, however, wasgreatly affected by the same incubation period at 50 �C (1.8-folddecrease after 6 h hydrolysis, up to 2.3 after 22 h). The catalyticactivity of CDs was even more reduced after incubation at 50 �C(2.4- and 3.1-fold decrease in glucose concentration after 6 and22 h, respectively). It can thus be concluded that the observed ther-molability of cellulases is mostly due to loss of catalytic activityrather than binding capacity, as pretreatment of cellulose withCBDs can be performed at 50 �C. CBDCel7A is a small peptide (37residues) and made up of an irregular triple-stranded antiparallelb-sheet (Kraulis et al., 1989). The whole structure is stabilized bytwo disulfide bridges, and this compact structure may be indeedmore stable at higher temperatures, compared to the more com-plex catalytic domain structure.
4. Conclusions
CBDs of cellulases from T. reesei had a noticeable impact on cel-lulose structure upon incubation. The crystallinity of differenttypes of cellulose (microcrystalline and fibrous) was reduced afterpretreatment with CBDs, rendering the substrates less recalcitrantto enzymatic hydrolysis, and an increase up to 25% in glucose con-centration during hydrolysis was obtained. This provides an alter-native pretreatment method for cellulosic material that reliesexclusively on biological molecules and mild conditions and doesnot require change of solvent. Lastly, this study demonstrates thatbinding capacity of CBDs is retained after extensive incubation at50 �C while there is significant loss of catalytic activity.
Acknowledgements
Chevron Corporation is gratefully acknowledged for financialsupport. We would like to thank Nicholas Johnson and Sang BeomKim for their help in the acquisition of data and the production ofCBDs. We thank David Bostwick from the Bioanalytical Mass Spec-trometry Facility at the School of Chemistry and Biochemistry atGeorgia Tech for the MALDI-TOF MS analysis and Jonathan Rubinfor his great efforts and contribution to help finalizing this study.
References
Arantes, V., Saddler, J.N., 2010. Access to cellulose limits the efficiency of enzymatichydrolysis: the role of amorphogenesis. Biotechnol. Biofuels 3.
Bansal, P., Hall, M., Realff, M.J., Lee, J.H., Bommarius, A.S., 2009. Modeling cellulasekinetics on lignocellulosic substrates. Biotechnol. Adv. 27, 833–848.
Bansal, P., Hall, M., Realff, M.J., Lee, J.H., Bommarius, A.S., 2010. Multivariatestatistical analysis of X-ray data from cellulose: a new method to determinedegree of crystallinity and predict hydrolysis rates. Bioresour. Technol. 101,4461–4471.
Beckham, G.T., Matthews, J.F., Bomble, Y.J., Bu, L.T., Adney, W.S., Himmel, M.E.,Nimlos, M.R., Crowley, M.F., 2010. Identification of Amino Acids Responsible forProcessivity in a Family 1 Carbohydrate-Binding Module from a FungalCellulase. J. Phys. Chem. B 114, 1447–1453.
Brennan, T.C.R., Datta, S., Blanch, H.W., Simmons, B.A., Holmes, B.M., 2010. Recoveryof sugars from ionic liquid biomass liquor by solvent extraction. Bioenergy Res.3, 123–133.
Cardona, C.A., Quintero, J.A., Paz, I.C., 2010. Production of bioethanol from sugarcanebagasse: status and perspectives. Bioresour. Technol. 101, 4754–4766.
Carrard, G., Koivula, A., Soderlund, H., Beguin, P., 2000. Cellulose-binding domainspromote hydrolysis of different sites on crystalline cellulose. Proc. Natl Acad.Sci. USA 97, 10342–10347.
Ciolacu, D., Kovac, J., Kokol, V., 2010. The effect of the cellulose-binding domainfrom Clostridium cellulovorans on the supramolecular structure of cellulosefibers. Carbohydr. Res. 345, 621–630.
Davies, G., Henrissat, B., 1995. Structures and mechanisms of glycosyl hydrolases.Structure 3, 853–859.
Din, N., Gilkes, N.R., Tekant, B., Miller, R.C., Warren, A.J., Kilburn, D.G., 1991. Non-hydrolytic disruption of cellulose fibers by the binding domain of a bacterialcellulase. Bio-Technology 9, 1096–1099.
Esteghlalian, A.R., Srivastava, V., Gilkes, N.R., Kilburn, D.G., Warren, R.A.J., Saddler,J.N., 2001. Do cellulose binding domains increase substrate accessibility? Appl.Biochem. Biotechnol., 575–592.
Gao, P.J., Chen, G.J., Wang, T.H., Zhang, Y.S., Liu, J., 2001. Non-hydrolytic disruptionof crystalline structure of cellulose by cellulose binding domain and linkersequence of cellobiohydrolase I from Penicillium janthinellum. Acta Biochim.Biophys. Sin. 33, 13–18.
Gilkes, N.R., Henrissat, B., Kilburn, D.G., Miller, R.C., Warren, R.A.J., 1991. Domains inmicrobial beta-1,4-glycanases – sequence conservation, function, and enzymefamilies. Microbiol. Rev. 55, 303–315.
Hall, M., Bansal, P., Lee, J.H., Realff, M.J., Bommarius, A.S., 2010. Cellulosecrystallinity – a key predictor of the enzymatic hydrolysis rate. FEBS J. 277,1571–1582.
Himmel, M.E., Ding, S.Y., Johnson, D.K., Adney, W.S., Nimlos, M.R., Brady, J.W., Foust,T.D., 2007. Biomass recalcitrance: engineering plants and enzymes for biofuelsproduction. Science 315, 804–807.
Jeihanipour, A., Karimi, K., Taherzadeh, M.J., 2010. Enhancement of ethanol andbiogas production from high-crystalline cellulose by different modes of NMOpretreatment. Biotechnol. Bioeng. 105, 469–476.
Jeoh, T., Michener, W., Himmel, M.E., Decker, S.R., Adney, W.S., 2008. Implications ofcellobiohydrolase glycosylation for use in biomass conversion. Biotechnol.Biofuels 1.
Jervis, E.J., Haynes, C.A., Kilburn, D.G., 1997. Surface diffusion of cellulases and theirisolated binding domains on cellulose. J. Biol. Chem. 272, 24016–24023.
Kraulis, P.J., Clore, G.M., Nilges, M., Jones, T.A., Pettersson, G., Knowles, J.,Gronenborn, A.M., 1989. Determination of the 3-dimensional solutionstructure of the C-terminal domain of cellobiohydrolase-I from Trichodermareesei – a study using nuclear magnetic-resonance and hybrid distancegeometry dynamical simulated annealing. Biochemistry 28, 7241–7257.
Lehtio, J., Sugiyama, J., Gustavsson, M., Fransson, L., Linder, M., Teeri, T.T., 2003. Thebinding specificity and affinity determinants of family 1 and family 3 cellulosebinding modules. Proc. Natl Acad. Sci. USA 100, 484–489.
Lemos, M.A., Teixeira, J.A., Mota, M., Gama, F.M., 2000. A simple method to separatecellulose-binding domains of fungal cellulases after digestion by a protease.Biotechnol. Lett. 22, 703–707.
Lemos, M.A., Teixeira, J.A., Domingues, M.R.M., Mota, M., Gama, F.M., 2003. Theenhancement of the cellulolytic activity of cellobiohydrolase I andendoglucanase by the addition of cellulose binding domains derived fromTrichoderma reesei. Enzyme Microb. Technol. 32, 35–40.
Linder, M., Teeri, T.T., 1996. The cellulose-binding domain of the majorcellobiohydrolase of Trichoderma reesei exhibits true reversibility and a highexchange rate on crystalline cellulose. Proc. Natl Acad. Sci. USA 93, 12251–12255.
Linder, M., Mattinen, M.L., Kontteli, M., Lindeberg, G., Stahlberg, J., Drakenberg, T.,Reinikainen, T., Pettersson, G., Annila, A., 1995. Identification of functionallyimportant amino-acids in the cellulose-binding domain of Trichoderma-Reeseicellobiohydrolase-I. Protein Sci. 4, 1056–1064.
Lynd, L.R., Weimer, P.J., van Zyl, W.H., Pretorius, I.S., 2002. Microbial celluloseutilization: fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 66, 506.
Lynd, L.R., Laser, M.S., Brandsby, D., Dale, B.E., Davison, B., Hamilton, R., Himmel, M.,Keller, M., McMillan, J.D., Sheehan, J., Wyman, C.E., 2008. How biotech cantransform biofuels. Nat. Biotechnol. 26, 169–172.
Ouajai, S., Shanks, R.A., 2006. Solvent and enzyme induced recrystallization ofmechanically degraded hemp cellulose. Cellulose 13, 31–44.
Pinto, R., Carvalho, J., Mota, M., Gama, M., 2006. Large-scale production of cellulose-binding domains. Adsorption studies using CBD-FITC conjugates. Cellulose 13,557–569.
Pinto, R., Amaral, A.L., Ferreira, E.C., Mota, M., Vilanova, M., Ruel, K., Gama, M., 2008.Quantification of the CBD-FITC conjugates surface coating on cellulose fibres.Bmc Biotechnol. 8, 2–4.
Sinnott, M.L., 1998. The cellobiohydrolases of Trichoderma reesei: a review ofindirect and direct evidence that their function is not just glycosidic bondhydrolysis. Biochem. Soc. Trans. 26, 160–164.
Stahlberg, J., Johansson, G., Pettersson, G., 1991. A new model for enzymatic-hydrolysis of cellulose based on the 2-domain structure of cellobiohydrolase-I.Bio-Technology 9, 286–290.
Takashima, S., Ohno, M., Hidaka, M., Nakamura, A., Masaki, H., 2007. Correlationbetween cellulose binding and activity of cellulose-binding domain mutants ofHumicola grisea cellobiohydrolase 1. FEBS Lett. 581, 5891–5896.
Wang, L.S., Zhang, Y.Z., Gao, P.J., 2008. A novel function for the cellulose bindingmodule of cellobiohydrolase I. Sci. China Ser. C-Life Sci. 51, 620–629.
Wormald, P., Wickholm, K., Larsson, P.T., Iversen, T., 1996. Conversions betweenordered and disordered cellulose. Effects of mechanical treatment followed bycyclic wetting and drying. Cellulose 3, 141–152.
Xiao, Z.Z., Gao, P.J., Qu, Y.B., Wang, T.H., 2001. Cellulose-binding domain ofendoglucanase III from Trichoderma reesei disrupting the structure ofcellulose. Biotechnol. Lett. 23, 711–715.
Yu, H.B., Guo, G.N., Zhang, X.Y., Yan, K.L., Xu, C.Y., 2009. The effect of biologicalpretreatment with the selective white-rot fungus Echinodontium taxodii onenzymatic hydrolysis of softwoods and hardwoods. Bioresour. Technol. 100,5170–5175.
Zhang, Y.H.P., Lynd, L.R., 2004. Toward an aggregated understanding of enzymatichydrolysis of cellulose: noncomplexed cellulase systems. Biotechnol. Bioeng.88, 797–824.
Zhao, X., Rignall, T.R., McCabe, C., Adney, W.S., Himmel, M.E., 2008. Molecularsimulation evidence for processive motion of Trichoderma reesei Cel7A duringcellulose depolymerization. Chem. Phys. Lett. 460, 284–288.
Zhao, H., Jones, C.I.L., Baker, G.A., Xia, S., Olubajo, O., Person, V.N., 2009.Regenerating cellulose from ionic liquids for an accelerated enzymatichydrolysis. J. Biotechnol. 139, 47–54.