biol. rev. (2010), 85, pp. 55–110. doi:10.1111/j.1469-185x
TRANSCRIPT

Biol. Rev. (2010), 85, pp. 55–110. 55doi:10.1111/j.1469-185X.2009.00094.x
The origin and early evolution of dinosaurs
Max C. Langer1∗, Martin D. Ezcurra2, Jonathas S. Bittencourt1
and Fernando E. Novas2,3
1Departamento de Biologia, FFCLRP, Universidade de Sao Paulo; Av. Bandeirantes 3900, Ribeirao Preto-SP, Brazil2Laboratorio de Anatomia Comparada y Evolucion de los Vertebrados, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Avda.
Angel Gallardo 470, Cdad. de Buenos Aires, Argentina3CONICET (Consejo Nacional de Investigaciones Cientıficas y Tecnicas); Avda. Rivadavia 1917 - Cdad. de Buenos Aires, Argentina
(Received 28 November 2008; revised 09 July 2009; accepted 14 July 2009)
ABSTRACT
The oldest unequivocal records of Dinosauria were unearthed from Late Triassic rocks (approximately 230 Ma)accumulated over extensional rift basins in southwestern Pangea. The better known of these are Herrerasaurus
ischigualastensis, Pisanosaurus mertii, Eoraptor lunensis, and Panphagia protos from the Ischigualasto Formation, Argentina,and Staurikosaurus pricei and Saturnalia tupiniquim from the Santa Maria Formation, Brazil. No uncontroversialdinosaur body fossils are known from older strata, but the Middle Triassic origin of the lineage may be inferredfrom both the footprint record and its sister-group relation to Ladinian basal dinosauromorphs. These includethe typical Marasuchus lilloensis, more basal forms such as Lagerpeton and Dromomeron, as well as silesaurids: a possiblymonophyletic group composed of Mid-Late Triassic forms that may represent immediate sister taxa to dinosaurs.The first phylogenetic definition to fit the current understanding of Dinosauria as a node-based taxon solelycomposed of mutually exclusive Saurischia and Ornithischia was given as ‘‘all descendants of the most recentcommon ancestor of birds and Triceratops’’. Recent cladistic analyses of early dinosaurs agree that Pisanosaurus mertii
is a basal ornithischian; that Herrerasaurus ischigualastensis and Staurikosaurus pricei belong in a monophyletic Her-rerasauridae; that herrerasaurids, Eoraptor lunensis, and Guaibasaurus candelariensis are saurischians; that Saurischiaincludes two main groups, Sauropodomorpha and Theropoda; and that Saturnalia tupiniquim is a basal member ofthe sauropodomorph lineage. On the contrary, several aspects of basal dinosaur phylogeny remain controversial,including the position of herrerasaurids, E. lunensis, and G. candelariensis as basal theropods or basal saurischians,and the affinity and/or validity of more fragmentary taxa such as Agnosphitys cromhallensis, Alwalkeria maleriensis,Chindesaurus bryansmalli, Saltopus elginensis, and Spondylosoma absconditum. The identification of dinosaur apomorphiesis jeopardized by the incompleteness of skeletal remains attributed to most basal dinosauromorphs, the skulls andforelimbs of which are particularly poorly known. Nonetheless, Dinosauria can be diagnosed by a suite of derivedtraits, most of which are related to the anatomy of the pelvic girdle and limb. Some of these are connected to theacquisition of a fully erect bipedal gait, which has been traditionally suggested to represent a key adaptation thatallowed, or even promoted, dinosaur radiation during Late Triassic times. Yet, contrary to the classical ‘‘compet-itive’’ models, dinosaurs did not gradually replace other terrestrial tetrapods over the Late Triassic. In fact, theradiation of the group comprises at least three landmark moments, separated by controversial (Carnian-Norian,Triassic-Jurassic) extinction events. These are mainly characterized by early diversification in Carnian times, aNorian increase in diversity and (especially) abundance, and the occupation of new niches from the Early Jurassiconwards. Dinosaurs arose from fully bipedal ancestors, the diet of which may have been carnivorous or omnivo-rous. Whereas the oldest dinosaurs were geographically restricted to south Pangea, including rare ornithischiansand more abundant basal members of the saurischian lineage, the group achieved a nearly global distribution bythe latest Triassic, especially with the radiation of saurischian groups such as ‘‘prosauropods’’ and coelophysoids.
Key words: Dinosauria, Dinosauromorpha, Triassic, phylogeny, evolution, biogeography, Herrerasauria.
∗ Address for correspondence: Departamento de Biologia, Faculdade de Filosofia, Ciencias e Letras de Ribeirao Preto, Universidadede Sao Paulo; Av. Bandeirantes 3900, 14040-901, Ribeirao Preto-SP, Brazil (Tel: +55 16 3602 3844; Fax: +55 16 3602 4886;E-mail: [email protected])
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

56 Max C. Langer and others
CONTENTS
I. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56(1) Historical background on early dinosaurs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56(2) The dinosauromorph radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
II. Phylogeny and Systematics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59(1) What makes a dinosaur? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60(2) Phylogenetic definitions: naming early dinosaurs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
III. Dinosaur ‘‘Trail Blazers’’ in Space, Time, and Evolutionary Context . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67(1) The oldest dinosaurs and the rocks that contain them . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67(2) The evolutionary tree of early dinosaurs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71(3) Geographical distribution of basal dinosaurs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75
IV. Ecology of the Dinosaur Radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77(1) The Triassic scene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77(2) Lucky break? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 78(3) Of legs and teeth: insights on the palaeobiology of early dinosaurs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82
V. Outcomes of a Radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84(1) Early ornithischian evolution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84(2) Early sauropodomorph evolution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87(3) Early theropod evolution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90
VI. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 93VII. Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94
VIII. Appendix 1. Institutional Abbreviations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94IX. References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94
I. INTRODUCTION
Dinosaurs originated in the Triassic period, and theLate Triassic represents the first 30 of the 165 millionyears of their ‘‘non-avian’’ history on Earth. Yet, ofthe 500-700 ‘‘well established’’ dinosaur genera (Wang& Dodson, 2006; Olshevsky, 2007), only about 30(approximately 5%) were excavated from Triassic rocks,and the diversity/diversification of the group is mainlyconcentrated in the Jurassic (Rauhut, 2005b; Lloyd et al.,2008) and/or Cretaceous (Wang & Dodson, 2006) periods.This is especially the case if one accepts the inference ofWang & Dodson (2006) that the Late Triassic representsthe best sampled subperiod of the entire Mesozoic in termsof documented dinosaur diversity. Indeed, dinosaurs arerare in most Triassic fossil assemblages in which they occur,although by the end of the period they were already dominantmembers of various palaeocommunities.
Triassic dinosaurs were mostly bipedal, and not exception-ally large. The basal-most forms were probably omnivorous,but predatory and probably herbivorous dinosaurs alsooccurred during Late Triassic times. This includes Her-
rerasaurus ischigualastensis, a top predator up to 4 m long(Novas, 1997a), and Riojasaurus incertus, a plant-eater of aboutthree tons (Seebacher, 2001). In taxonomic terms, most Tri-assic dinosaurs are regarded as members of one of the threemajor lineages of the group: Theropoda, Sauropodomorpha,and Ornithischia. Yet, despite representing well-known taxa,other Triassic dinosaurs have a debated phylogenetic posi-tion. This is particularly the case of the herrerasaurs, whichwere placed basal to the Ornithischia-Saurischia dichotomy,
nested within Theropoda, or regarded as non-eusaurischansaurischians.
Appealing inferences on dinosaur palaeobiology canbe drawn from eggs and nestlings (Bonaparte & Vince,1979; Moratalla & Powell, 1994), monospecific assemblages(Coombs, 1990; Schwartz & Gillette, 1994), visual-display-related morphological features (Vickaryous & Ryan, 1997),and ‘‘stomach contents’’ (Novas, 1997a; Nesbitt et al., 2006)of Triassic dinosaurs. Yet, the most debated aspect of earlydinosaur macroevolution corresponds to their first radiation,and various scenarios were invoked to explain the rise ofthe clade in a time interval during which most terrestrialtetrapods suffered important diversity losses. In fact, by LateTriassic times, dinosaurs arose and took their first stepsalong the evolutionary road, and the investigation of theirobscure origins is crucial for the understanding of dinosaurinterrelationships and palaeobiology as a whole.
(1) Historical background on early dinosaurs
Research on early dinosaurs can be said to have startedwith the work of the German palaeontologist Friedrich vonHuene, and his descriptions of Saltopus elginensis Huene,1910 (Fig. 1A), and Spondylosoma absconditum Huene, 1942.These two forms have completely different provenances,coming respectively from the Elgin area, in NorthernScotland, and Rio Grande do Sul, in South Brazil, butshare curious similarities. Both were regarded as saurischiandinosaurs by Huene (1910, 1942) and were found in depositsconsidered the oldest dinosaur-bearing rocks known at thetime. Huene (1932, 1942) identified various other putativeTriassic dinosaurs as equivalent in age to either Saltopus orSpondylosoma, but most of these were shown to have doubtful
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 57
dinosaur affinities (Galton, 1985b; Benton, 1986b; Galton &Walker, 1996; Benton et al., 2000; Rauhut & Hungerbuhler,2000; Parker et al., 2005; Nesbitt, Irmis & Parker, 2007).Notable exceptions are Thecodontosaurus antiquus (Benton et al.,2000) and the material Cope (1889) originally assigned toCoelophysis bauri (Nesbitt et al., 2007), but these came fromstrata currently considered younger (Benton et al., 2000;Langer, 2005b; Nesbitt et al., 2007). Indeed, the older age ofboth the ‘‘Stagonolepis-beds’’ of Elgin (Huene, 1908) and the‘‘Rio do Rasto’’ [sic] beds at Chiniqua (Huene & Stahlecker,1931) was corroborated by recent work. The LossiemouthSandstone Formation has been dated as Carnian (Benton &Walker, 1985), whereas the Dinodontosaurus Assemblage-Zoneof the Santa Maria Formation is considered of Ladinianage (Langer et al., 2007c); or early-middle Carnian, followingrecent modifications on the Late Triassic time-scale (Muttoniet al., 2004) and the corrections on the radiometric dating ofthe Ischigualasto Formation (Furin et al., 2006).
Although the ages of the Lossiemouth Sandstone andSanta Maria formations were more securely established,the dinosaur affinities of Saltopus elginensis and Spondylosoma
absconditum are still debated (Rauhut & Hungerbuhler, 2000;Galton, 2000; Langer, 2004). This is in part due to thepoor preservation of the specimens, which do not allow acomprehensive assessment of their morphological features.Therefore, it was not until Reig (1963) placed Herrerasaurus
ischigualastensis (Fig. 1B) and Ischisaurus cattoi within Saurischiathat unequivocal early dinosaurs were known to science. Thedescribed specimens were collected in 1961 from depositsof the Ischigualasto Formation, San Juan province, north-western Argentina, which have yielded remains attributableto dinosaurs since the late 1950s (Reig, 1963). With thediscovery, in 1962, of the ornithischian Pisanosaurus mertii
Casamiquela, 1967 (Fig. 1C), in that same stratigraphic unit,the presence of both main dinosaur lineages (i.e. Ornithischiaand Saurischia), in the Triassic of South America wasconfirmed. Another important ‘‘early dinosaur’’ study ofthe time was the description of Staurikosaurus pricei Colbert,1970 (Fig. 1D). Its type and only specimen, discovered in
1936 in the Santa Maria beds of South Brazil, was the firstconsensual early dinosaur to be collected.
While the 1980s were quiet times regarding the studyof early dinosaurs, mainly witnessing the description ofincomplete specimens (Galton, 1985b, 1986; Novas, 1986;Chatterjee, 1987; Murry & Long, 1989), the early ninetiescame with new and exciting discoveries. These include theunearthing, also from the Ischigualasto Formation, of a newbasal dinosaur still to be fully described, Eoraptor lunensis
(Sereno et al., 1993; Sereno, 2007b), and of further materialof Herrerasaurus ischigualastensis (Sereno & Novas, 1992, 1993;Novas, 1993; Sereno, 1993). In the late nineties, a new seriesof discoveries in Rio Grande do Sul, South Brazil, enlargedthe knowledge of early dinosaur diversity. The then basal-most member of the sauropodomorph lineage, Saturnalia
tupiniquim (Langer et al., 1999; Langer, Franca & Gabriel,2007b; Langer, 2003), was unearthed from the Hyperodapedon
Assemblage-Zone of the Santa Maria Formation, whereasthe overlying Caturrita Formation yielded the saurischianGuaibasaurus candelariensis (Bonaparte, Ferigolo & Ribeiro,1999; Bonaparte et al., 2007). Since the beginning of thiscentury, some putative basal dinosaurs have been described(Fraser et al., 2002; Ferigolo & Langer, 2007; Nesbitt et al.,2007; Nesbitt & Chatterjee, 2008; Martinez & Alcober, 2009;Ezcurra, 2008), while the validity of others was evaluated inthe light of new evidence (Remes & Rauhut, 2005; Yates,2007b). More importantly, different evolutionary scenarioswere proposed based on independent cladistic analyses, e.g.Langer & Benton (2006), Ezcurra (2006), Sereno (2007b),Irmis et al. (2007a), which attempted to sum up informationin order to understand better the interrelationships of earlydinosaurs.
(2) The dinosauromorph radiation
For most of the last century, it was accepted that dinosaursarose from ‘‘thecodont’’ precursors, either as a monophyleticgroup or, more frequently (Fig. 2), in the form of inde-pendent lineages (Huene, 1956; Colbert, 1964; Charig,
Fig. 1. Early images depicting some of the oldest putative dinosaurs. (A) Drawing of the slabs containing Saltopus elginensis, fromHuene (1910). (B) Skeletal reconstruction of Herrerasaurus ischigualastensis as mounted in 1965 for exhibition at the UniversidadNacional de Tucuman, from Bonaparte (1997). (C) Skeletal reconstruction of Pisanosaurus mertii, from Bonaparte (1997). (D) Skeletalreconstruction of Staurikosaurus pricei, from Colbert (1970). Scale bars: A = 5 cm; B-D = 10 cm.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

58 Max C. Langer and others
Fig. 2. Schemes of archosaur evolution depicting a polyphyletic Dinosauria. (A) Modified from Krebs (1974). (B) After Thulborn(1975).
Attridge & Crompton, 1965; Romer, 1966). ‘‘Thecodonts’’,as composed of non-crown-group archosaurs, and basalmembers of both the bird and crocodile lines, are cur-rently regarded as a paraphyletic group (Currie & Padian,1997b; Benton, 2004). In his seminal paper on dinosaurphylogeny, Gauthier (1986) applied the name OrnithosuchiaHuene, 1908, to designate a group composed of dinosaurs,pterosaurs (including Scleromochlus), ornithosuchids, Euparke-
ria (questionably), and ‘‘Lagosuchus’’, a small archosaur fromthe Middle Triassic of Argentina (Romer, 1971; Bonaparte,1975; Sereno & Arcucci, 1994). That clade was supposed togroup all archosaurs that share a closer affinity to birds (withinDinosauria) than to crocodiles, which were placed in its sis-ter group Pseudosuchia (Parrish, 1997; Senter, 2005). Morerecent work, however, excluded both Euparkeria (Benton &Clark, 1988; Sereno, 1991a; Juul, 1994; Benton, 2004) andornithosuchids (Sereno, 1991a; Juul, 1994; Benton, 2004)from Ornithosuchia, restricting the inclusivity, and perhapsworthiness (Taylor, 2007) of the name. Indeed, alternativenames were later proposed for the bird line of Archosauria,e.g. Avemetatarsalia Benton, 1999; Panaves Gauthier & DeQueiroz, 2001. The inclusivity of this group could be evenmore reduced considering the labile position of pterosaurs,sometimes regarded as basal archosaurs (Bennett, 1996) oreven outside Archosauria (Peters, 2000; Sobral & Langer,2008). In this scenario, the non-dinosaur members of thebird-lineage of Archosauria would only include Scleromochlus
taylori (a putative sister taxon to Pterosauria) from the LateTriassic of Elgin (Sereno, 1991a; Benton, 1999) and theso-called basal dinosauromorphs.
The name Dinosauromorpha was coined by Benton(1985) to include dinosaurs, birds, and ornithosuchids, butredefined by Sereno (1991a) to its current understanding,which excludes ornithosuchids. The basal (non-dinosaurian)
members of the group (Romer, 1971, 1972a, b; Arcucci,1987) were for a long time known only from the MiddleTriassic Chanares Formation of Argentina (Rogers et al.,2001). These small, gracile forms were grouped within‘‘Pseudosuchia’’, but were soon recognized to have somebearing on the origin of dinosaurs (Romer, 1972a, b), whichbecame evident with the works of Bonaparte (1975), Sereno& Arcucci (1993, 1994), and Novas (1996). Those authorsidentified typical dinosaur hind-limb traits on these taxa,including a distally tapering fibula, an anterior ascendingprocess in the astragalus, a reduced calcaneum, a longermetatarsus with reduced outer elements, and a straightmetatarsal V with reduced articulation area on the outersurface of the lateral distal tarsal (see also Irmis et al.,2007a; Brusatte et al., 2009). The taxonomy of the Chanaresdinosauromorphs has always been subject to some debate(Bonaparte, 1975, 1995; Sereno & Arcucci, 1994; Arcucci,1987, 1998, 2005), and five names entered the literature:Lagerpeton chanarensis Romer, 1971; Lagosuchus talampayensis
Romer, 1971 (nomen dubium; Sereno & Arcucci, 1994);Marasuchus lilloensis (Romer, 1972b, gen. Sereno & Arcucci,1994); Lewisuchus admixtus Romer, 1972a (Arcucci, 1997); andPseudolagosuchus major Arcucci, 1987.
Recent discoveries (Fraser et al., 2002; Dzik, 2003; Irmiset al., 2007a; Ferigolo & Langer, 2007) and interpretations(Novas & Ezcurra, 2005; Ezcurra, 2006; Nesbitt et al., 2007)suggest that basal dinosauromorphs were both more diversein terms of anatomy and inferred habits, and more widelyspread chronologically and geographically. Dromomeron romeri
and D. gregorii (Irmis et al., 2007a; Nesbitt et al., 2009) wererecognized in the Norian of western North America, whichalso yielded Eucoelophysis baldwini. The latter taxon, firstdescribed as a theropod dinosaur (Sullivan & Lucas, 1999),was reassigned to a non-dinosaur dinosauriform position,
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 59
as either the sister taxon to Dinosauria (Ezcurra, 2006) orforming a group with Silesaurus opolensis (Irmis et al., 2007a).The latter form, collected in Carnian deposits of Poland(Dzik, 2003; Dzik & Sulej, 2007), provided the greatestbreakthrough in the recent study of dinosaur origins. Its longfore limbs suggest that the animal was at least facultativelyquadrupedal, while the edentulous front tip of its lower jawapparently bore a corneous beak. This atypical set of traitsrevealed an unsuspected morphological diversity, hinting athow incomplete was, and certainly still is, our knowledge ofthe early stages of dinosauromorph evolution. In addition,the record of Silesaurus opolensis extended the range of basaldinosauriforms into the Late Triassic of Europe, a possibilityonly hinted at before on the basis of controversial British taxasuch as Saltopus elginensis (Rauhut & Hungerbuhler, 2000) andAgnosphitys cromhallensis (Fraser et al., 2002). Further, sincethe description of Silesaurus opolensis, newly and previouslydescribed Norian forms have been considered closely relatedto the taxon. This is the case for Sacisaurus agudoensis Ferigolo& Langer, 2007, from the Caturrita Formation of SouthBrazil, and a set of North American specimens (Nesbittet al., 2007), including material assigned to an unnamedSilesaurus-like form from the Petrified Forest Member, ChinleFormation, of New Mexico, and part of the original materialof Technosaurus smalli Chatterjee, 1984, from the Bull CanyonFormation, Texas (Irmis et al., 2007b). The latter taxonhas been previously assigned to Ornithischia (Weishampel& Witmer, 1990; Sereno, 1991b; Hunt & Lucas, 1994),while Sacisaurus agudoensis might provide evidence that evenSilesaurus opolensis represents a basal member of that dinosaurclade (Ferigolo & Langer, 2007).
The more complete non-dinosaurian dinosaurormorphsform a series of outgroups to Dinosauria, and they giveclues about the origin of the clade (Ezcurra, 2006; Langer &Benton, 2006; Yates, 2007a; Irmis et al., 2007a; Brusatteet al., 2009). The long-held hypothesis of a more basalposition for Lagerpeton chanarensis (Novas, 1992b; Sereno& Arcucci, 1993) was confirmed by independent studies(Irmis et al., 2007a; Brusatte et al., 2009), which allocatedthe genus Dromomeron as its sister taxon (Fig. 3A). BothLagerpeton and Dromomeron lack several apomorphic featuresof Dinosauriformes such as a reduced medial lamina on thepubis, an antitrochanter expanding into the ilium, a lessertrochanter on the proximal femur, and a distal tibia bearinga lateral groove and a squared distal articulation (Irmis et al.,2007a; Brusatte et al., 2009). Within Dinosauriformes, moststudies (Novas, 1992b, 1996; Ezcurra, 2006; Irmis et al.,2007a; Brusatte et al., 2009) place Marasuchus lilloensis as thebasalmost member of the clade (Fig. 3A). More derived formsinclude Pseudolagosuchus major (Novas, 1992b, 1996) and itspossible senior synonym Lewisuchus admixtus (Arcucci, 1998,2005). Along with the identification of further dinosauriformsof equivalent grade (Dzik, 2003; Ezcurra, 2006), twoalternative phylogenetic scenarios were proposed (Fig. 3A).Irmis et al. (2007a) suggested that Eucoelophysis and Silesaurus
form the sister clade to Dinosauria, which may also includePseudolagosuchus according to Nesbitt et al. (2007, p. 214).
Ezcurra (2006), on the other hand, placed all these taxa ina fully pectinated grade, where Pseudolagosuchus, Silesaurus,and Eucoelophysis, are respectively closer to Dinosauria.A somewhat intermediate view was adopted by Brusatteet al. (2009), in which Pseudolagosuchus has a basal position,and Lewisuchus forms, with other taxa, a more restricted sisterclade to dinosaurs (Fig. 3A). In any case, all or some of theseforms share with dinosaurs a number of apomorphic traitsabsent in Marasuchus, e.g. longer pubic shaft; femur withangular greater trochanter, ‘‘spike-like’’ lesser trochanter,and prominent trochanteric shelf; distal tibia with laterallyexpanded outer malleolus; astragalus with pyramid-shapedanterior ascending process; and sigmoidal metatarsal IVwith deeper distal articular surface (Novas, 1996; Irmis et al.,2007a; Brusatte et al., 2009).
Regardless of their status as a clade or ‘‘grade’’, these morederived basal dinosauromorphs fill a gap (between Marasuchus
lilloensis and dinosaurs) in archosaur evolution. Moreimportantly, they fill that gap with the unsuspected diversityof forms that have been informally called ‘‘silesaurids’’.This group may just include Silesaurus, and forms such asSacisaurus and Technosaurus, which share with the Polish taxondental/jaw features possibly related to a more herbivorousdiet (Ferigolo & Langer, 2007; Irmis et al., 2007b), but it couldalso encompass Lewisuchus, Pseudolagosuchus, and Eucoelophysis.Although the basis for this assignment lies on shared traitsof the postcranium, there is no positive evidence that anyof these forms was a facultative/full quadruped as Silesaurus.Yet, ‘‘herbivorous’’ teeth have been tentatively referred toEucoelophysis (Irmis et al., 2007a). The record of ‘‘silesaurids’’and of the species of Dromomeron suggests that an extensiveradiation of non-dinosaurian dinosauromorphs preceded theLate Triassic dinosaur diversification, and that parallel to thefirst radiation of dinosaurs, that grade continued to flourishafter the Ladinian (Irmis et al., 2007a), extending their rangeinto the northern part of west Pangea (Fig. 3B).
II. PHYLOGENY AND SYSTEMATICS
The name Dinosauria was erected by Owen (1842) toinclude three large terrestrial forms which he believed tocompose a distinct group of extinct reptiles (Torrens, 1992;Padian, 1997a). In the following years, a sound conceptof Dinosauria was established by the proposition of severalclassification schemes (Cope, 1866; Huxley, 1870; Marsh,1882; Seeley, 1888). At that time, major taxa such asSauropoda and Theropoda (Marsh, 1878, 1881), as well asSaurischia and Ornithischia (Seeley, 1888) were proposed.These names gained acceptance in the 20th Century (Huene,1932; Romer, 1956) and still represent the major dinosaursubdivisions as currently understood (Fig. 4). However, formost of the last century these different dinosaur groups, andeven some of their subgroups, were believed to have hadindependent origins (Fig. 2) from ‘‘thecodont’’ precursors(Huene, 1914, 1956; Colbert, 1964; Charig et al., 1965;Romer, 1966; Reig, 1970; Krebs, 1974; Thulborn, 1975;
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

60 Max C. Langer and others
Fig. 3. Time-calibrated phylogenies and distribution of non-dinosaur Dinosauromorpha. (A) Recently proposed phylogenetichypotheses; dotted lines indicate ghost lineages; names applied as in Table 1. Position of Pseudolagosuchus in the phylogeny of Irmiset al. (2007a) inferred from Nesbitt et al. (2007). (B) Geographic occurrence of taxa on a Late Triassic map redrawn from Blakey(2006). Black silhouettes adapted from various sources.
Cruickshank, 1975, 1979). The monophyly of Dinosauria wassuggested by Bakker & Galton (1974) and Bonaparte (1975,1976), firmly established by various pioneering cladisticworks (Paul, 1984; Gauthier & Padian, 1985; Cooper, 1985;Brinkman & Sues, 1987), especially that of Gauthier (1986),and represents a consensual hypothesis nowadays (Novas,1989; 1996; Sereno et al., 1993; Sereno, 1999; Langer &Benton, 2006; Irmis et al., 2007a).
(1) What makes a dinosaur?
Even if the monophyly of Dinosauria is consensuallyaccepted, the issue of which morphological traits characterizethe group continues to be debated (Novas, 1996; Langer &Benton, 2006; Sereno, 2007b). Several putative dinosaur
apomorphies were proposed in a variety of studies dealingwith the phylogeny of the group, which frequently divergeupon the distribution of these same characters. This isepitomized by the continuing quarrel over one of thediagnostic features mentioned by Owen (1842) in the originalproposition of the name: the number of vertebrae thatcompose the dinosaur sacrum. In the following text, mostrecent reviews of early dinosaur phylogeny (Novas, 1996;Sereno, 1999, 2007a; Fraser et al., 2002; Benton, 2004;Langer & Benton, 2006; Ezcurra, 2006; Yates, 2007a, b;Irmis et al., 2007a; Brusatte et al., 2008a) are compared andevaluated, in a search for the set of traits that typicallycharacterize the group. Obviously, a key point to set thediagnosis of Dinosauria is to determine whether some ofthe so-called basal dinosauromorphs actually belong to the
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 61
Fig. 4. Generalized phylogeny depicting the position of Dinosauria and its main groups within Archosauria. Dotted lines indicatemajor contentious placement of taxa; arrows indicate stem-based taxa; black circles indicate node-based taxa; names applied as inTable 1; black silhouettes adapted from various sources.
group. As reviewed by Langer & Benton (2006, pp. 316-317),various putative dinosaur apomorphies are seen in Silesaurus
opolensis. These might represent true dinosaur apomorphiesif the taxon is considered to represent a basal ornithischian(Ferigolo & Langer, 2007). Yet, current orthodoxy pointstowards the basal, non-dinosaurian position of Silesaurus, andthis hypothesis of relationships represents the template basedon which the unique dinosaur traits are discussed below.
Novas (1996) and Sereno (1999) respectively listed 17 and18 characters as diagnostic for Dinosauria, while a modifiedversion of one of their characters (presence of three or moresacral vertebrae) is the sole dinosaur apomorphy proposedby Fraser et al. (2002). Langer & Benton (2006) criticallyassessed these characters, questioning the apomorphic statusof several of them. Features related to the cranial anatomy(Sereno & Novas, 1993) are particularly problematic becausemost basal dinosaurs and, especially, basal dinosauromorphslack good skull material. Indeed, traits such as the lack of thepostfrontal bone, although typically absent in non-dinosaurarchosaurs and present in dinosaurs (see Irmis et al., 2007a,char. 14), can not be considered an unambiguous dinosaurapomorphy (Langer & Benton, 2006) given its equivocaloccurrence in most forms placed at the very origin of thegroup. The same applies to other putative apomorphies ofthe dinosaur skull, such as the dorsal overlap of the transverseflange of the pterygoid by the ectopterygoid, and the lateralexposure of the quadrate head (Langer & Benton, 2006); seealso Brusatte et al. (2008a, chars 10, 14, 38, 40, 67). Thestatus of other putative apomorphies of the dinosaur skull isdependent on the position of Silesaurus opolensis, the cranialmaterial of which is reasonably complete (Dzik, 2003; Dzik& Sulej, 2007). If not considered a dinosaur, some of its
cranial traits, e.g. frontal participating in the supratemporalfossa, are dismissed as dinosaur apomorphies. Yet, if itsless consensual position as a basal ornithischian is accepted,these same traits continue potentially to represent dinosaursynapomorphies. On the contrary, plesiomorphic traits inthe skull of Silesaurus such as a large post-temporal fenestrasupports its non-dinosaurian affinity, and helps to define areduced foramen-sized aperture (Fig. 5B) as apomorphic fordinosaurs (Irmis et al., 2007a, char. 21). Other cranial features(Langer & Benton, 2006, char. 12; Ezcurra, 2006, chars 4,20; Yates, 2007a, chars 26, 29; Irmis et al., 2007a, chars 2,25) suggested to represent possible dinosaur apomorphies,pending the criteria used for character optimization, have anerratic distribution among basal dinosaurs, and should notbe considered a priori diagnostic traits of the group. A likelydinosaur apomorphy, related to the axial skeleton (Fig. 5C), isthe presence of epipophyses on the cranial cervical vertebrae(Novas, 1996; Langer & Benton, 2006; Yates, 2007a; contra
Ezcurra, 2006). This feature was previously considered asaurischian apomorphy, but more recently was recorded inbasal ornithischians (Novas, 1996; Langer & Benton, 2006;Butler, Smith & Norman, 2007). Other putative apomorphiesof the dinosaur vertebral column listed by Yates (2007a, chars129, 142) have an inconsistent distribution, and should notbe a priori considered as such.
As mentioned earlier, the increase in the number ofvertebrae that forms the dinosaur sacrum (from two tomore than two) continues to be listed as an apomorphy ofthe group (Novas, 1996; Sereno, 1999; Fraser et al., 2002;Ezcurra, 2006). Recently, as discussed by Langer & Benton(2006), two main strategies of coding characters related tothis transformation have been employed; but see Novas
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

62 Max C. Langer and others
Fig. 5. The dinosaur Plateosaurus engelhardti. (A) Skeletal reconstruction (from Yates, 2003a), with indications of the better knownapomorphic traits of Dinosauria. (B) Occipital view of the skull (from Galton, 1985a) indicating (1) a foramen-sized post-temporalfenestra. (C) Lateral view of a cervical vertebra, indicating (2) the presence of epipophyses. (D) Caudal view of the left humerus,indicating (3) a long deltopectoral crest. (E) Lateral view of the left ilium, indicating (4) an open acetabulum and (5) an arched dorsalmargin. (F) Cranial view of the left femur, indicating (6) a femoral head inturned and distinctly offset from the shaft and (7) anasymmetrical forth trochanter. (G) Proximal view of the left astragalus, indicating (8) an acute anteromedial corner, (9) a broaderascending process, and (10) a reduced fibular articulation. (H) Cranial view of the distal tarsals, indicating (11) a proximally flatlateral distal tarsal. All figured material refers to the mounted skeletons (GPIT I and III) of the ‘‘Sauriersaal’’ at Institut fur Geologieund Palaontologie, Tubingen (Weishampel & Westphal, 1986), except: B = SMNS 12949. Scale bars: A = 1 m; B-E, G-H = 5 cm;F = 10 cm.
(1996) for a combined approach. Some (Fraser et al., 2002;Rauhut, 2003; Ezcurra, 2006; Irmis et al., 2007a) adopteda topographic criterion, simply considering the number ofsacral vertebrae, while others (Sereno et al., 1993, Sereno,1999; Langer, 2004; Langer & Benton, 2006, Yates, 2007a)attempted to recognize whether trunk or caudal elementshave been incorporated into the sacrum. Evidence fora two-vertebrae sacrum within basal dinosaurs is limited,and restricted to incomplete specimens (Langer & Benton,2006; Yates, 2007a; Sereno, 2007b). On the contrary, thesacrum of Silesaurus is clearly composed of three sacralvertebrae (Dzik & Sulej, 2007). Accordingly, based on thecurrent evidence, and considering Silesaurus as closely relatedbut outside Dinosauria, the statement that dinosaurs areapomorphic in having a sacrum composed of more thantwo vertebrae is misleading. A more detailed approach thatattempts to recognize trunk or tail additions to the sacrummay provide further information. In a few basal dinosaurs, i.e.Saturnalia tupiniquim, Herrerasaurus ischigualastensis, Staurikosaurus
pricei, Guaibasaurus candelarienesis, and Eoraptor lunensis, the twoprimordial sacral vertebrae are readily recognized basedon their much larger rib articulations. Other vertebraemay be incorporated into the sacrum from either thetrunk (Herrerasaurus, Eoraptor) or the caudal (Staurikosaurus,Saturnalia) series, but none has a conspicuous sacral rib,
compared to the primordial elements. Such a robust thirdelement is known in Silesaurus opolensis, and we agree withDzik & Sulej (2007) that it is borne by a trunk vertebraadded to the sacrum. Among the major dinosaur groups, alltheropods and ornithischians have trunk vertebrae added tothe sacrum, as is also the case in sauropodomorphs, exceptfor Plateosaurus (Yates, 2003c) and, possibly, Thecodontosaurus
(Yates, 2007a). Accordingly, even if a trunk vertebra addedto the sacrum is seen in most basal dinosaurs, the presenceof this character in Silesaurus dismisses its apomorphic statusfor the group. On the other hand, the incorporation ofa caudal vertebra to the dinosaur sacrum seems morerestricted, absent in various basal forms (i.e. Herrerasaurus,
Eoraptor) and most basal sauropodomorphs (Yates, 2007a).Indeed, the presence of caudosacral vertebrae is also notaccepted as a dinosaur apomorphy. It is evident that weare dealing with a highly homoplastic character, possiblyaffected by frame shift phenomena (Galton & Upchurch,2000). It is also of misleading codification if one considers theambiguous condition of vertebrae that bore small transverseprocesses/ribs that attach to the ilium and/or other sacraltransverse processes/ribs; compare Herrerasaurus in Novas(1993) and Sereno (2007b). The increase in the numberof sacral vertebrae is, generally speaking, surely a typicaldinosaur trait. Yet, until more information, possibly derived
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 63
from better preserved specimens of key taxa, is available,the number of sacral vertebrae, and also the incorporationof either trunk or caudal elements in the sacrum cannot beunambiguously defined as dinosaur apomorphies. Besides,Langer & Benton (2006) considered a dorsally expandedcranial margin of the first primordial sacral rib as apomorphicfor dinosaurs. Similarly, this condition was also recognized inSilesaurus (ZPAL Ab III/404/3), and can not be considered adinosaur apomorphy in the phylogenetic framework adoptedhere.
Few characters of the pectoral girdle and limb have beenconsidered apomorphic for dinosaurs. This may indicate thatthese parts of the dinosaur skeleton are not very modifiedrelative to the basic archosaur condition. Yet, it may alsoreflect the lack of knowledge regarding these anatomicalelements, especially the forearm and hand, in the outgroupsto Dinosauria. This is particularly the case with the char-acters related to the reduction of the outer digits of thedinosaur manus (Gauthier & Padian, 1985; Novas, 1996;Sereno, 1999). Indeed, dinosaur digit IV is always sube-qual to or shorter than metatarsal III and never possessesmore than three phalanges, none of which is an ungual(Langer & Benton, 2006). In addition, almost no dinosauris known to possess more than two phalanges in manualdigit V. On the contrary, manual digits IV and V of otherarchosauromorphs are elongated elements with three ormore phalanges. More recently, Butler et al. (2007) claimedthat an enlarged grasping manus (with elongated pre-ungualphalanges, prominent dorsal extensor pits and proximalintercondylar processes), previously considered typical of Her-
rerasaurus ischigualastensis and theropods (Sereno et al., 1993;Sereno, 1999), may also be apomorphic for dinosaurs, dueto its occurrence in basal ornithischians (Eocursor parvus andheterodontosaurids). However, the manus is unknown innon-dinosaur dinosauromorphs, and it is ambiguous at whichpoint of basal dinosauromorph evolution these modificationsoccurred. Likewise, although no sternal plates have been rec-ognized in basal dinosauromorphs, this may simply representa preservation bias (Padian, 1997b), and their occurrence aspaired ossifications (Sereno, 1999) can not be regarded as atrustworthy dinosaur apomorphy.
In fact, the single feature of the pectoral skeleton acceptedby most previous studies as apomorphic for Dinosauriaappears to be a long deltopectoral crest (Fig. 5D), whichextends for more than 30-35% of the humeral length.Besides, as noted by several authors (Yates, 2007a; Irmiset al., 2007a; Brusatte et al., 2008a), contrasting with that ofpseudosuchians and Silesaurus opolensis, the deltopectoral crestof dinosaurs is subrectangular, rather than subtriangular orrounded. Yet, although lacking its proximal margin, thedeltopectoral crest of Marasuchus lilloensis (PVL 3871) seemsof the subrectangular type (Bonaparte, 1975), implying amore inclusive distribution for that trait. Likewise, a shorterforearm relative to the humerus can not be accepted a priori
as a dinosaur apomorphy (Irmis et al., 2007a), given thata plesiomorphic longer forearm is retained in Herrerasaurus
ischigualastensis and Eoraptor lunensis (Langer et al., 2007b).
Most novel traits of the early dinosaur skeleton are seenin the pelvic girdle and limb. These were often relatedto the acquisition of an improved bipedal gait (Bakker &Galton, 1974), as typical of most basal members of thegroup. Further, some authors, e.g. Bakker (1971) and Charig(1972, 1984), have suggested that these traits represent keyfeatures that allowed, or even promoted, dinosaur radiationin Late Triassic times, while most other archosaurs were indecline. Regardless of their evolutionary consequences (seeSections IV.2,3), it is true that the dinosaur pelvic girdle andlimb bear various apomorphic traits. Indeed, about half ofthe features presented by Novas (1996) and Sereno (1999)as diagnostic for dinosaurs are related to those elements(exclusive of the sacrum), and similar ratios are seen in otherrecent works: four out of 11 in Langer & Benton (2006);seven out of 11 in Ezcurra (2006); eigth out of 15 in Yates(2007a); and 10 out of 14 in Irmis et al. (2007a). Obviously, thefact that these anatomical parts are relatively well known inbasal dinosauromorphs facilitates the recognition of dinosaurapomorphies.
Regarding the pelvic girdle, a perforated acetabulum(Bakker & Galton, 1974; Novas, 1996; Ezcurra, 2006; Yates,2007a), better described as a straight to concave ventralacetabular margin of the ilium (Langer & Benton, 2006; Irmiset al., 2007a; Brusatte et al., 2008a), stands in most recent revi-sions as a valid synapomorphy of Saurischia plus Ornithischia(Fig. 5E), but that is not the case of a brevis fossa/shelf in theiliac postacetabular ala (Novas, 1996; Sereno, 1999; Fraseret al., 2002; Benton, 2004; Yates, 2007a). Whereas a shelfis also present in Marasuchus (Fraser et al., 2002; Langer &Benton, 2006; but see Novas, 1996), a fossa is not onlyseen in some basal dinosauriforms (e.g. Silesaurus), but is alsolacking in herrerasaurids (Novas, 1992b, 1993, 1996; Langer& Benton, 2006). More recently, Ezcurra (2006) proposeda straight to convex dorsal margin of the ilium (Fig. 5E)as a dinosaur apomorphy. Indeed, basal dinosaurs lack adorsally excavated ilium, which seems to be typical of basaldinosauromorphs (Sereno & Arcucci, 1993, 1994), althoughnot well preserved in some (e.g. Silesaurus; ZPAL AbIII/361).On the contrary, other recently proposed apomorphies ofthe dinosaur ilium are either highly homoplastic (‘‘longpreacetabular process’’; Yates, 2007a) or define a moreinclusive clade (‘‘acetabular antitrochanter present’’; Irmiset al., 2007a), i.e. Dinosauriformes (see Sereno & Arcucci,1994). Irmis et al. (2007a) also suggested that a transverselycompressed distal pubis is a dinosaur apomorphy, reversedin sauropodomorphs, but the definition and distribution ofthis feature is not so straightforward, as extensively discussedby Langer & Benton (2006, p. 338).
Irmis et al. (2007a, char. 73) newly proposed that the pubicprocess of the dinosaur ischium is apomorphic, because‘‘separated from the ilial peduncle’’. In fact, the surfaceconnecting the iliac and pubic articulations of the ischiumis simply excavated in many basal dinosaurs, especiallytheropods (Coelophysis rhodesiensis, QVM QG 1; Liliensternus
liliensterni, MB R 2175) and ornithischians (Scelidosaurus
harrisoni, BMNH1111; Scutellosaurus lawleri, UCMP 130580;
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

64 Max C. Langer and others
Butler et al., 2007). On the contrary, in forms suchas Marasuchus lilloensis (PVL 3870) and Silesaurus opolensis
(ZPAL AbIII 1228, 404/1) that excavation does notreach the medial-most margin of the ischium, so that amedially displaced sheet of bone remains, filling the spacebetween pubic process and iliac peduncle. This condition isreminiscent of more basal archosaurs, in which the ischiumcontributes significantly to the composition of the medial wallof a non-perforated acetabulum. Herrerasaurus ischigualastensis
(PVL 2566) retains a much reduced medial sheet of bone, sothat the acetabular surface of the ischium can be consideredfully excavated, i.e. bearing the dinosaur apomorphy asdefined by Irmis et al. (2007a). On the contrary, the conditionamong sauropodomorphs is variable (Yates, 2003c); e.g. inSaturnalia tupiniquim (MCP 3846-PV), although an extensiveantitrochanter disrupts the clear observation of the character(but see Liliensternus liliensterni, MB R 2175), the medial sheet ofbone occupies the space between that structure and the pubicarticulation. Accordingly, the status of the character definedby Irmis et al. (2007a) awaits further investigation. Otherpreviously proposed apomorphies of the dinosaur ischiuminclude the presence of a reduced medioventral lamina(Novas, 1996; Langer & Benton, 2006) and a proximaldorsolateral sulcus (Yates, 2007a). Yet, both features areclearly present in Silesaurus (ZPAL AbIII 361, 404/1), sothat their status as apomorphic for dinosaurs depends on thecontentious position of that taxon.
The femur is possibly the most scrutinized bone in the studyof early dinosaurs, with more than ten different charactersfound as apomorphic for the group in the phylogenies revisedhere. An inturned and subrectangular femoral head, that isdistinctly set from the shaft, has been considered amongthe typical traits of dinosaurs by Bakker & Galton (1974)and Gauthier (1986). Yet, this general state was poorlydismembered into distinct and well-defined phylogeneticcharacters, in order to evaluate the apomorphic condition ofeach. Sereno (1999) defined an angular ‘‘greater trochanter’’(i.e. nearly straight angle between the proximal articulationand the long axis of the shaft) as a dinosaur apomorphy,but that trait was also recognized in basal dinosauromorphs(e.g. Pseudolagosuchus major, PULR 53; Ezcurra, 2006). Thisstructures a subrectangular femoral head, if the latter isdistinctly offset from the shaft, as diagnostic of dinosaurs(Ezcurra, 2006, char. 231; Irmis et al., 2007a, char. 81;Brusatte et al., 2008a, char. 132). That condition appearsalong with an inturned femoral head (Fig. 5F), which can bealso considered a dinosaur apomorphy.
Irmis et al. (2007a) claim that the femoral head of dinosaursapomorphicaly bears a ligament sulcus and an asymmetricalfossa articularis antitrochanterica, but these traits have alsobeen recorded in other basal dinosauromorphs (Novas, 1996;Ezcurra, 2006). Likewise, the apomorphic condition of areduced medial tuberosity (Novas, 1996; Sereno, 1999)and a prominent lesser trochanter (Novas, 1996) havebeen dismissed by most recent studies (Langer & Benton,2006; Ezcurra, 2006; Irmis et al., 2007a). Other features ofthe femoral head were considered apomorphic reversals of
Dinosauria (Ezcurra, 2006, char. 232; Irmis et al., 2007a,char. 85; Brusatte et al., 2008a, char. 135), but dependon character optimization. Besides, although reversed intheropods, the presence of an asymmetrical fourth trochanter(Fig. 5F) appears as a valid dinosaur apomorphy in mostrecent reviews (Langer & Benton, 2006; Ezcurra, 2006;Irmis et al., 2007a), and has been recently recorded also inGuaibasaurus candelariensis (Bonaparte et al., 2007; contra Langer& Benton, 2006) and Chindesaurus bryansmalli (GR 226; contra
Yates, 2007a).Previously defined tibial traits such as the presence of a
cnemial crest (Novas, 1996; Sereno, 1999) and a transverselyexpanded distal articulation (Novas, 1996; Benton, 2004)are no longer believed to represent dinosaur apomorphies,given their erratic distribution among basal dinosaurs anddinosauromorphs (Langer & Benton, 2006; Ezcurra, 2006;Irmis et al., 2007a; Brusatte et al., 2008a). Similarly, becausealso seen in Silesaurus, a descending process of the tibia thatcaudally overlaps the ascending process of the astragalusis also not regarded apomorphic for dinosaurs. Accordingto Yates (2007a), a sub-quadratic distal tibia and a thinnerfibula may represent dinosaur apomorphies, because thereverse condition is seen in Silesaurus. Yet, the record ofthe dinosaur condition in Marasuchus lilloensis jeopardizesthat assumption. Accordingly, no unambiguous apomorphyis currently referred to the dinosaur pelvic epipodium. Inaddition, Ezcurra (2006) considered, under DELTRANoptimization, a tibia subequal to the femur as apomorphic fordinosaurs. Although the contrary was described for Silesaurus
opolensis (Dzik, 2003), a longer tibia is not only typical ofbasal dinosauromorphs (Sereno & Arcucci, 1993; 1994;Pesudolagosuchus major, PVL 4629), but was also retained inbasal ornithischians (Santa Luca, 1980; Butler et al., 2007).Indeed, among basal dinosaurs, only saurischians consistentlybear a subequal or longer femur; but see Staurikosaurus pricei
(Colbert, 1970).The tarsal joint has also been the source of several
anatomical traits believed to characterize dinosaurs. Yet,this is not the case of an astragalar ascending process and alateral articulation between the calcaneum and the astragalaranterolateral process (Sereno, 1999), which were recentlyidentified in other basal Dinosauromorpha (Novas, 1996;Langer & Benton, 2006; Brusatte et al., 2008a). Yet, a reducedfibular articulation (Langer & Benton, 2006; Brusatte et al.,2008a), a broader ascending process (Yates, 2007a, char.314), and an acute anteromedial corner (Irmis et al., 2007a)apparently stand as apomorphies of the dinosaur astragalus(Fig. 5G). On the contrary, some putative apomorphiesof the dinosaur calcaneum, such as a concave fibulararticulation (Novas, 1996) and a rudimentary medial process(Sereno, 1999) have an erratic distribution among basaldinosauromorphs, and can not be unambiguously consideredas such (Langer & Benton, 2006; Ezcurra, 2006; Irmis et al.,2007a; Brusatte et al., 2008a). On the other hand, as faras the condition in the outgroups to Dinosauria can beaccessed, a proximally flat lateral distal tarsal (Novas, 1996;Langer & Benton, 2006; Brusatte et al., 2008a) stands as a
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 65
unique trait of the group (Fig. 5H). The single apomorphy ofthe dinosaur metatarsus proposed in the discussed studiesof early dinosaur phylogeny is the so-called ‘‘sigmoid’’metatarsal IV (Sereno, 1999), a condition given by thelateral displacement of the distal part of the bone (Novas,1996; Brusatte et al., 2008a). This condition is, however, alsoseen in some basal dinosauromorphs (Novas, 1996; Ezcurra,2006), and disregarded as a dinosaur apomorphy.
(2) Phylogenetic definitions: naming early dinosaurs
With the advent of Phylogenetic Nomenclature (De Queiroz& Gauthier, 1990, 1992, 1994), systematists acquired anunprecedented tool to define taxon names in explicitphylogenetic context, setting their composition accordingto given hypotheses. A drawback of this revolution wasthe inflation of phylogenetic definitions for various names(Benton, 2000), as readily recognized in a brief inspectionof Paul Sereno’s webpage TaxonSearch. Indeed, when dealingwith these names, authors currently have to state whichof the available definitions is adopted to translate theminto the phylogenetic nomenclature system. The priorityissue is expected to be settled with the publication ofthe ‘‘companion volume’’ of the PhyloCode (Cantino & DeQueiroz, 2007). Yet, before this volume is published and,more importantly, accepted by the scientific community asthe ‘‘Systema Naturae’’ of phylogenetic definitions, thesewill no doubt continue to proliferate in an unordered way.In the following paragraphs, the phylogenetic definitionspertinent to the discussion of dinosaur origins are treated inhistorical order and, in an attempt to emulate the ‘‘Principleof Priority’’ (ICZN, 1999), those first proposed, with smallmodifications added if absolutely required, are listed inTable 1 and employed throughout the text.
Because Jacques Gauthier was involved in the studyof archosaurs, including dinosaurs, he presented somephylogenetic definitions for related groups (Gauthier &Padian, 1985; Gauthier, 1986) even before the publication ofthe paper that set the theoretical foundation of PhylogeneticNomenclature (De Queiroz & Gauthier, 1990). Gauthier& Padian (1985) provided a phylogenetic definition forOrnithosuchia, while Gauthier (1986) explicitly definedSaurischia and Theropoda. Problematic aspects of thesedefinitions include the use of supraspecific and/or informalspecifiers (e.g. birds, archosaurs, crocodiles, dinosaurs,sauropodomorphs, Ornithischia) and their choice based onthe phylogenetic orthodoxy of the time. Instead, we believethat, for the sake of precision, newly proposed phylogeneticdefinitions should use minimal groups as specifiers, andfor historical coherence rely, as much as possible, on taxamentioned in the original definition of the names. In any case,because first published, those definitions are adopted herefor the names in question (Table 1). Alternative phylogeneticdefinitions for Saurischia (Padian & May, 1993; Padian,1997d; Sereno, 1998; Holtz & Osmolska, 2004; Langer,2004) just replace specifiers, either because these are morespecific (Padian, 1997d; Sereno, 1998) or are quoted in theoriginal proposition of the name (Langer, 2004). Yet, based
on current phylogenetic hypotheses, these circumscribe thesame set of taxa as Saurischia sensu Gauthier (1986). Similarly,alternative specifiers in later definitions of Theropoda aremore specific (Currie, 1997) and either more highly nested(Sereno, 1998) or first named (Padian, Hutchinson & Holtz,1999; Holtz & Osmolska, 2004). Again, their use does notchange the inclusivity of the group as defined by Gauthier(1986).
Further phylogenetic definitions pertinent to the discussedgroups were proposed by Sereno (1991a), Novas (1992b),and Padian & May (1993). Sereno (1991a) gave node-based definitions for Ornithodira Gauthier, 1986, andDinosauromorpha Benton, 1985. These had to be slightlymodified (Table 1) to fit the logical basis of PhylogeneticNomenclature and the updated taxonomy of Sereno &Arcucci (1994), but substitute definitions (Benton, 2004)are redundant. Especially problematic are the stem-based definitions of Dinosauromorpha (Sereno, 1991a,2005; Benton, 2004) that use pterosaurs as the externalspecifier, given the uncertain phylogenetic position of thesereptiles. In their current understanding, Ornithodira andDinosauromorpha differ only by the inclusion of Scleromochlus
taylori and possibly pterosaurs in the former. The leastinclusive Dinosauriformes was node-based defined whenfirst named by Novas (1992b). This was modified (Table 1)to fit the taxonomy of Sereno & Arcucci (1994), but equallyrequires no substitute definitions (Benton, 2004).
Apart from the equivocal list of taxa presented by Gauthier(1986, p. 44; see Padian, 1997a), Novas (1992b) provided thefirst phylogenetic definition of Dinosauria as ‘‘the commonancestor of Herrerasauridae and Saurischia + Ornithischia,and all of its descendants’’. This is in agreement withthe taxonomic orthodoxy of the time (Gauthier, 1986;Brinkman & Sues, 1987; Benton, 1990; but see Gauthieret al., 1989), according to which: (1) saurischians plusornithischians form a clade, contrary to the traditional viewthat these arose independently from ‘‘thecodont’’ precursors;(2) herrerasaurids, conventionally regarded as saurischiandinosaurs (Reig, 1963; Colbert, 1970), are basal to thatclade. Indeed, in order to keep herrerasaurids as dinosaurs,Novas (1992b) used the former as an internal specifier ofthe latter. By contrast, Padian & May (1993) explicitlyrestricted the use of Dinosauria to the clade composed ofSaurischia and Ornithischia, exclusive of ‘‘Herrerasaurus andits allies’’. Despite the ‘‘priority’’ of Novas (1992b), the latterconcept gained almost unconditional acceptance since (e.g.Sereno, 1998, 2005; Fraser et al., 2002) and is employedhere (Table 1). In any case, these alternate definitionsonly circumscribe different groups if herrerasaurids areplaced outside the Saurischia + Ornithischia dichotomy,a hypothesis not supported by most recent studies (seebelow). Other authors (Holtz in Padian, 1997a; Olshevsky,2000; Clarke, 2004) attempted phylogenetically to defineDinosauria using taxa included in the original propositionof the name. In this case, the best option may be using allnames mentioned by Owen (1842) in a node-based fashion,and to define Dinosauria as ‘‘the most recent common
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

66 Max C. Langer and others
Table 1. Phylogenetic definition of names relevant in the context of early dinosaur evolution.
Name Phylogenetic definition
ORNITHODIRAGauthier, 1986
‘‘Pterosauria, Scleromochlus, Dinosauromorpha (including birds), and all descendantsof their most recent common ancestor’’ modified from Sereno (1991a),node-based
DINOSAUROMORPHABenton, 1985
‘‘Lagerpeton chanarensis, Marasuchus lilloensis, Pseudolagosuchus major, Dinosauria (inc.Aves), and all descendants of their most recent common ancestor’’ modified fromSereno (1991a); node-based
DINOSAURIFORMESNovas, 1992b
‘‘The most recent common ancestor of Marasuchus lilloensis, Dinosauria, and all taxastemming from it’’ modified from Novas (1992b); node-based
SILESAURIDAEnew name
‘‘All archosaurs closer to Silesaurus opolensis, than to Heterodontosaurus tucki andMarasuchus lilloensis’’; stem-based
DINOSAURIAOwen, 1842
‘‘All descendants of the most recent common ancestor of birds and Triceratops’’Padian & May (1993); node-based
ORNITHISCHIASeeley, 1888
‘‘Dinosaurs closer to Triceratops than to birds’’ Padian & May (1993); stem-based
GENASAURIASereno, 1986
‘‘Thyreophora and Cerapoda and all descendants of their common ancestor’’Currie & Padian (1997a); node-based
NEORNITHISCHIACooper, 1985
‘‘All genasaurs closer to Triceratops than to Ankylosaurus’’ Sereno (1998); stem-based
THYREOPHORANopcsa, 1915
‘‘All genasaurs closer to Ankylosaurus than to Triceratops’’ Sereno (1998); stem-based
SAURISCHIASeeley, 1888
‘‘Birds and all dinosaurs that are closer to birds than they are to Ornithischia’’Gauthier (1986); stem-based
HERRERASAURIAGalton, 1985b
‘‘All dinosaurs that share a more recent common ancestor with Herrerasaurus thanwith Liliensternus and Plateosaurus’’ Langer (2004); stem-based
HERRERASAURIDAEBenedetto, 1973
‘‘Herrerasaurus, Staurikosaurus, their most recent common ancestor, plus all itsdescendants’’ modified from Novas (1992b); node-based
EUSAURISCHIAPadian et al. 1999
‘‘The least inclusive group of Saurischia, containing Cetiosaurus and Neornithes’’Langer (2004); node-based
SAUROPODOMORPHAHuene, 1932
‘‘The clade including the most recent common ancestor of Prosauropoda andSauropoda and all of its descendants’’ Salgado et al. (1997); node-based
MASSOPODAYates, 2007a
‘‘The most inclusive clade containing Saltasaurus loricatus but not Plateosaurus
engelhardti’’ Yates (2007a); stem-basedSAUROPODIFORMES
Sereno, 2005‘‘The least inclusive clade containing Mussaurus patagonicus Bonaparte & Vince,
1979, and Saltasaurus loricatus Bonaparte & Powell, 1980’’ Sereno (2005);node-based
SAUROPODAMarsh, 1878
‘‘The most recent common ancestor of Vulcanodon karibaensis and Eusauropoda andall of its descendants’’ Salgado et al. (1997); node-based
THEROPODAMarsh, 1881
‘‘Birds and all saurischians that are closer to birds than they are tosauropodomorphs’’ Gauthier (1986); stem-based
NEOTHEROPODABakker, 1986
‘‘Coelophysis, Neornithes, their most recent common ancestor and all descendants’’Sereno (1998); node-based
COELOPHYSOIDEANopcsa, 1928
‘‘All ceratosaurs closer to Coelophysis than to Carnotaurus’’ Sereno (1998);stem-based
AEROSTRAPaul, 2002
‘‘Ceratosaurus nasicornis, Allosaurus fragilis and all the descendants of their most recentcommon ancestor’’ modified from Ezcurra & Cuny (2007); node-based
ancestor of Megalosaurus, Iguanodon, and Hylaeosaurus, and all itsdescendants’’. Again, according to the current phylogenetichypotheses, this definition circumscribes the same set of taxaas that of Padian & May (1993).
Novas (1992b) also proposed a node-based definition forHerrerasauridae, ‘‘emended’’ by Novas (1997a). Yet, bothdefinitions are incomplete and a modified version of them isemployed here (Table 1). There is no good reason to replacethat definition with a stem-based Herrerasauridae (Sereno,1998; Benton, 2004), especially because this is equivalent toHerrerasauria (see below). Further, Padian & May (1993)
provided a stem-based definition for Ornithischia, in afashion that matches its mutual exclusivity in relation toSaurischia sensu Gauthier (1986). Subsequent definitions usemore specific (Sereno, 1998) and also more ‘‘traditional’’(Weishampel, 2004; Norman, Witmer & Weishampel, 2004a)specifiers, but are equally inclusive based on currentphylogenies. Although the use of taxa mentioned in theproposal of Saurischia (e.g. Allosaurus, Camarasaurus) andOrnithischia (e.g. Stegosaurus, Iguanodon) may have been moredesirable, all the previous definitions successfully translateSeeley’s (1888) dichotomous understanding of Dinosauria
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 67
into the Phylogenetic Nomenclature system. Likewise, itcould also be argued that the use of apomorphy-baseddefinitions for Saurischia and Ornithischia better representsthat original proposition, given that the groups were definedon a character basis, i.e. opisthopubic and propubic pelves.Yet, this is problematic because only the ornithischian pelvicconstruction is apomorphic, whereas saurischians retain thegeneral morphology seen in more basal archosaurs.
Salgado, Coria & Calvo (1997) first proposed aphylogenetic definition for Sauropodomorpha (Table 1).Their node-based definition preceded that (stem-based)given by Upchurch (1997b) by a couple of months, butboth suffer from using Sauropoda and Prosauropoda asinternal specifiers. Subsequent proposals attempt to replacethose taxa by more specific, and deeply nested specifiers ineither a node- (Sereno, 1998) or stem- (Galton & Upchurch,2004; Sereno, 2007a) based fashion. Although lower rankspecifiers are desirable, the same level of precision can beachieved using higher taxa that are, in turn, defined withdirect reference (or by typification) to those minimal groups.Moreover, the adequacy of an either stem- or node-basedSauropodomorpha (Upchurch, Barrett & Galton, 2007) isminor in face of the primacy of the definition provided bySalgado et al. (1997).
More recently, Langer (2004) defined a stem-basedHerrerasauria Galton, 1985b, and a node-based EusaurischiaPadian, Hutchinson & Holtz, 1999. The former group ispotentially equivalent to Herrerasauridae sensu Sereno (1998),but the node-based original definition of Herrerasauridae isemployed here. In that context, Herrerasauria (Table 1) canallocate dinosaurs closely related to, but outside the cladecomposed of Herrerasaurus plus Staurikosaurus. Eusaurischia,on the other hand, was first proposed to designate the cladecomposed of Sauropomorpha plus Theropoda (Padian et al.,1999). This is as inclusive as the stem-based Saurischia undercertain phylogenetic schemes (Novas, 1996; Sereno, 1999),but excludes basal forms such as Eoraptor and herrerasaursin alternative frameworks (Langer, 2004; Ezcurra, 2006)and remains a potentially useful name (Table 1). Finally,Silesauridae is here defined as a stem-based taxon thatincludes all archosaurs closer to Silesaurus opolensis than toMarasuchus lilloensis and Heterodontosaurus tucki. The latterform was chosen to represent Dinosauria because of itscompleteness (Santa Luca, 1980) and basal position withinOrnithischia (Butler et al., 2007), a group to which Silesaurus
has been tentatively related (Ferigolo & Langer, 2007).
III. DINOSAUR ‘‘TRAIL BLAZERS’’ IN SPACE,TIME, AND EVOLUTIONARY CONTEXT
(1) The oldest dinosaurs and the rocks that containthem
For most of the last century, except in a few importantcases (Huene, 1926; Colbert, 1989; Sereno & Novas, 1992;Sereno et al., 1993), the knowledge of Triassic dinosaurs was
based on incomplete and/or fragmentary skeletal remains.In the last decade, however, various studies (e.g. Rauhut& Hungerbuhler, 2000; Langer, 2004; Parker et al., 2005;Ezcurra, 2006; Nesbitt et al., 2007) revised those early records,questioning the dinosaur affinity of several of them. On theother hand, the discovery of a variety of more completebasal dinosaurs (e.g. Langer et al., 1999; Bonaparte et al.,1999, 2007; Yates & Kitching, 2003; Butler et al., 2007; Pol& Powell, 2007a, b; Martinez & Alcober, 2009; Ezcurra,2008), allowed a more reliable picture to emerge. As detailedbelow, this accounts for the possible, but poorly supportedMiddle Triassic origin of the group, its first radiation duringthe Carnian, and the full establishment of the main dinosaurgroups from the Norian onwards.
Usually, the oldest dinosaurs (Galton, 2000; Langer,2004) are considered as coming from the Ischigualastianbeds (Langer, 2005a) of northwestern Argentina and southBrazil (Fig. 6). These respectively include the IschigualastoSequence, Ischigualasto-Ischichuca depocenter, BermejoBasin (Stipanicic & Marsicano, 2002; Currie et al., 2009),and the Santa Maria Supersequence, Parana Basin (Zerfasset al., 2003), the continental sedimentation of which filledextensional rift basins related to the Gondwanides orogenesis(Zerfass et al., 2004). Early works dated the Ischigualastoand Santa Maria formations as Middle Triassic (Romer,1960, 1962; Reig, 1961, 1963), but a Late Triassic age,first proposed by Bonaparte (1966), has been supported bymost recent biostratigraphic studies (Ochev & Shishkin, 1989;Lucas, 1998; Langer, 2005a, b). This was corroborated by theradiometric dating of the ‘Herr Toba’ bentonite (Fig. 6C),at the base of the Ischigualasto Formation (Rogers et al.,1993), that provided a 40Ar/39Ar age of 227 ± 0.3 Mya. Yet,following the discrepancy between U-Pb and 40Ar/39Ar dates(Schoene et al., 2006) and other comparative parameters,Furin et al. (2006) recalculated a date of 230.3-231.4 ± 0.3Mya. This corresponds to the late Ladinian in most timescales(Ross, Baud & Manning, 1994; Remane et al., 2000; Ogg,2004; Ogg, Ogg & Gradstein, 2008), but recent works(Muttoni et al., 2001, 2004; Gallet et al., 2003; Kent, Muttoni& Brack, 2006; Kozur & Weems, 2007) assigned older agesfor the Carnian boundaries. In that context, and consideringthe sedimentation rate of comparable rift basins (Rogerset al., 1993; Currie et al., 2009), the dinosaur-rich sites of thelower third of the Ischigualasto Formation can be placed inthe latest Carnian. Yet, the middle third of that stratigraphicunit, that also yielded dinosaur remains, may rest withinthe middle Norian. This was recently corroborated by thedating of another bentonite, from above the middle sectorof the Ischigualasto Formation (Currie et al., 2009), whichprovided a 40Ar/39Ar age of 217.0 ± 1.7 Ma (Shipman,2004), recalculated as 219.4-220.4 ± 1.7 Mya (M. Ezcurra,personal observations).
Ischigualastian dinosaurs (Fig. 6C) include Herrerasaurus
ischigualastensis, along with its possible synonyms Ischisaurus
cattoi and Frenguellisaurus ischigualastensis (Novas, 1993), Eoraptor
lunensis (Sereno et al., 1993), and Panphagia protos (Martinez& Alcober, 2009), from the lower third of the Ischigualasto
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

68 Max C. Langer and others
Fig. 6. Tectonic and sedimentary settings of southwestern Pangea during the Middle and Late Triassic, with emphasis on the SouthAmerican dinosaur-bearing sequences (Zerfass et al., 2004; Veevers, 2005). (A) Idealized east-west cross section from Santa Mariaintraplate rift to the Cuyo back-arc rift and Gondwanides orogen. (B) Palaeogeographic reconstruction; note that the extensionalbasins are perpendicular to the transtensional stresses. Abbreviations as follows: SLV, Sierra de la Ventana; CFB, Cape Fold Belt.Gondwanides orogen in grey. (C) Stratigraphic charts of the Bermejo and Parana Basins, depicting the dinosauromorph/putativedinosaur record. Fm., Formation; HAZ, Hyperodapedon Acme Zone according to Langer et al. (2007c); Mys, million years beforerecent. Asterisks indicate possibly coeval faunas in which the dicynodont Jachaleria occurs.
Formation, and Pisanosaurus mertii (Bonaparte, 1976) from themiddle third of that stratigraphic unit (Rogers et al., 1993),as well as Staurikosaurus pricei (Colbert, 1970) and Saturnalia
tupiniquim (Langer et al., 1999) from the Hyperodapedon
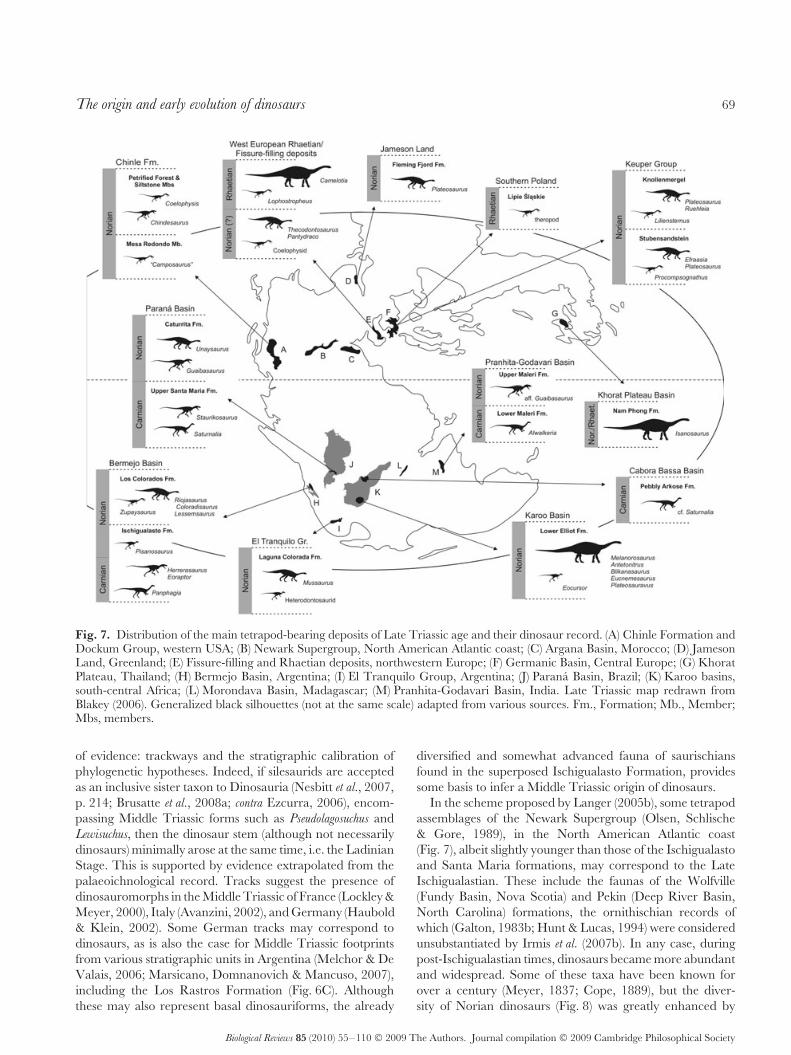
Assemblage-Zone of the Santa Maria Formation (Langeret al., 2007b). More recently, the discoveries of two newherrerasaurids (Martinez & Alcober, 2007; Ezcurra & Novas,2008), a Saturnalia-like animal (Ezcurra & Novas, 2008;Ezcurra, 2008), and a probable basal theropod (Martinez,Sereno & Alcober, 2008) have been announced from theIschigualasto Formation. Outside South America, dinosaursof similar age are much less conspicuous (Fig. 7). Thesemainly include fragmentary remains from Gondwanan areassuch as the possible record of Saturnalia in the Pebbly ArkoseFormation (Cabora Bassa Basin), lower Zambezi Valley,Zimbabwe (Raath, 1996; Langer et al., 1999), and partof the specimens attributed to Alwalkeria maleriensis, fromthe Lower Maleri Formation (Pranhita-Godavari Basin), incentral Peninsular India (Chatterjee, 1987; Remes & Rauhut,2005). The record of dinosaurs in other coeval deposits
such as the Timesgadiouine Formation (Argana Basin),in Morocco (Jalil, 1996, Gauffre, 1993), and the Isalo IIbeds (Morondava Basin), in Madagascar (Flynn et al., 1999),has been dismissed (Jalil & Knol, 2002; Flynn et al., 2008).According to Langer (2005b) the Ischigualastian can betraced into northern Pangea to encompass the LossiemouthSandstone Formation, in northern Scotland. Yet, the onlyputative dinosaur from those strata, Saltopus elginensis, hasdoubtful affinities to the group (Rauhut & Hungerbuhler,2000; Langer, 2004).
All dinosaur osteological records from pre-Ischigualstianstrata have been questioned, including Spondylosoma abscon-
ditum (Galton, 2000; Langer et al., 2007c), from the SantaMaria 1 sequence in south Brazil (Fig. 6C). Further occur-rences of the group in strata of equivalent age, mainlybased on fragmentary European specimens (Huene, 1932),have also been dismissed (Benton, 1986b; Norman, 1990;Galton & Walker, 1996; Rauhut & Hungerbuhler, 2000).On the other hand, suggestions that dinosaurs were alreadypresent in Middle Triassic times are backed up by two lines
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society
The origin and early evolution of dinosaurs 69
Fig. 7. Distribution of the main tetrapod-bearing deposits of Late Triassic age and their dinosaur record. (A) Chinle Formation andDockum Group, western USA; (B) Newark Supergroup, North American Atlantic coast; (C) Argana Basin, Morocco; (D) JamesonLand, Greenland; (E) Fissure-filling and Rhaetian deposits, northwestern Europe; (F) Germanic Basin, Central Europe; (G) KhoratPlateau, Thailand; (H) Bermejo Basin, Argentina; (I) El Tranquilo Group, Argentina; (J) Parana Basin, Brazil; (K) Karoo basins,south-central Africa; (L) Morondava Basin, Madagascar; (M) Pranhita-Godavari Basin, India. Late Triassic map redrawn fromBlakey (2006). Generalized black silhouettes (not at the same scale) adapted from various sources. Fm., Formation; Mb., Member;Mbs, members.
of evidence: trackways and the stratigraphic calibration ofphylogenetic hypotheses. Indeed, if silesaurids are acceptedas an inclusive sister taxon to Dinosauria (Nesbitt et al., 2007,p. 214; Brusatte et al., 2008a; contra Ezcurra, 2006), encom-passing Middle Triassic forms such as Pseudolagosuchus andLewisuchus, then the dinosaur stem (although not necessarilydinosaurs) minimally arose at the same time, i.e. the LadinianStage. This is supported by evidence extrapolated from thepalaeoichnological record. Tracks suggest the presence ofdinosauromorphs in the Middle Triassic of France (Lockley &Meyer, 2000), Italy (Avanzini, 2002), and Germany (Haubold& Klein, 2002). Some German tracks may correspond todinosaurs, as is also the case for Middle Triassic footprintsfrom various stratigraphic units in Argentina (Melchor & DeValais, 2006; Marsicano, Domnanovich & Mancuso, 2007),including the Los Rastros Formation (Fig. 6C). Althoughthese may also represent basal dinosauriforms, the already
diversified and somewhat advanced fauna of saurischiansfound in the superposed Ischigualasto Formation, providessome basis to infer a Middle Triassic origin of dinosaurs.
In the scheme proposed by Langer (2005b), some tetrapodassemblages of the Newark Supergroup (Olsen, Schlische& Gore, 1989), in the North American Atlantic coast(Fig. 7), albeit slightly younger than those of the Ischigualastoand Santa Maria formations, may correspond to the LateIschigualastian. These include the faunas of the Wolfville(Fundy Basin, Nova Scotia) and Pekin (Deep River Basin,North Carolina) formations, the ornithischian records ofwhich (Galton, 1983b; Hunt & Lucas, 1994) were consideredunsubstantiated by Irmis et al. (2007b). In any case, duringpost-Ischigualastian times, dinosaurs became more abundantand widespread. Some of these taxa have been known forover a century (Meyer, 1837; Cope, 1889), but the diver-sity of Norian dinosaurs (Fig. 8) was greatly enhanced by
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

70 Max C. Langer and others
Fig. 8. Skeletal reconstructions (from various sources), at approximately the same scale, of selected Carnian and Norian dinosaurs,partially depicting the Late Triassic diversity of the group. Scale bar (lower left ) = 1 m.
last-decade discoveries, especially from South America andSouth Africa, as outlined below.
Post-Ischigualastian dinosaur faunas in South Americainclude those of the Los Colorados, Laguna Colorada, andCaturrita formations (Langer, 2005a). The latter strati-graphic unit, in south Brazil (Fig. 6C), has yielded thesaurischian Guaibasaurus candelariensis (Bonaparte et al., 1999,2007), as well as the ‘‘prosauropod’’ Unaysaurus tolentinoi
(Leal et al., 2004). ‘‘Prosauropods’’ are well known in the LaEsquina fauna of the Los Colorados Formation (Bonaparte,1972). That stratigraphic unit covers the Ischigualasto For-mation in northwestern Argentina (Fig. 6C), and includesRiojasaurus incertus (Bonaparte & Plumares, 1995), Coloradis-
aurus brevis (Bonaparte, 1978), and Lessemsaurus sauropoides (Pol& Powell, 2007a), along with theropods (Bonaparte, 1972)such as Zupaysaurus rougieri (Arcucci & Coria, 2003; Ezcurra& Novas, 2007a). In Patagonia, the Laguna Colorada For-mation (El Tranquilo Group) has yielded the ‘‘prosauropod’’Mussaurus patagonicus (Bonaparte & Vince, 1979; Pol & Powel,2007b) as well as a heterodontosaurid ornithischian (Baez& Marsicano, 2001). Other dinosaur-bearing gondwanandeposits of similar age (Fig. 7) include the Lower Elliot For-mation (Stormberg Group, Karoo Basin), in South Africa
(Knoll, 2005), and the Upper Maleri Formation, in penin-sular India. The latter, along with the overlying LowerDharmaram Formation, has yielded a diversified, but stillundescribed fauna of basal saurischians (Kutty & Sengupta,1989; Novas et al., 2006), which may include a Guaibasaurus-like form (Kutty et al., 2007). Basal sauropodomorphs arealso well known in the Lower Elliot Formation, whereMelanorosaurus readi, Antetonitrus ingenipes, Blikanasaurus cromp-
toni, Eucnemesaurus fortis, Plateosauravus cullingworthi, and a yetunnamed form (Yates, 2003a, 2007a, b, 2008; Yates &Kitching, 2003) were recorded along with the ornithischianEocursor parvus (Butler et al., 2007) and possible theropod teeth(Ray & Chinsamy, 2002).
In North Pangea, various Norian faunas of Europe andNorth America yielded dinosaur records (Fig. 7). Theseinclude the rich prosauropod fauna of the German Keuper,where Efraasia minor occurs in the Middle Stubensand-stein (Lowenstein Formation) of Baden-Wurttemberg, alongwith Procompsognathus triassicus and other possible theropods(Hungerbuhler, 1998; Rauhut & Hungerbuhler, 2000; Yates,2003a). Specimens/species attributed to Plateosaurus aremuch more widespread both geographically and stratigraph-ically (Yates, 2003c; Moser, 2003; Weishampel et al., 2004),
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 71
also occurring in the overlying Knollenmergel (Trossin-gen Formation, and related stratigraphic units) of Baden-Wurttemberg, Bavaria, Lower Saxony, and Saxony-Anhalt,as well as in putative coeval faunas from France, Switzerland,and Greenland (Jenkins et al., 1994; Galton & Upchurch,2004). In addition, the Thuringian Knollenmergel hasyielded the ‘‘prosauropod’’ Ruehleia bedheimensis and the thero-pod Liliensternus liliensterni (Rauhut & Hungerbuhler, 2000;Galton, 2001). The ‘‘lower’’ fissure-filling deposits of theBritish Isles (southwest England and south Wales) are alsofrequently regarded as Norian in age (Fraser, 1994; Ben-ton & Spencer, 1995), although they might well spreadinto the late Carnian and/or Rhaetian (Benton et al., 2000).Among these, the Pant-y-ffynnon site, in south Wales, isbetter known for its dinosaur fauna, which includes thebasal sauropodomorph Pantydraco caducus (Kermack, 1984;Yates, 2003b; Galton, Yates & Kermack, 2007) and a smalltheropod possibly related to Coelophysis/Syntarsus (Rauhut &Hungerbuhler, 2000). P. caducus was previously assigned tothe genus Thecodontosaurus, the type species of which (T.
antiquus) is also known from various other putatively coevalfissure-filling deposits (Benton & Spencer, 1995; Bentonet al., 2000). In addition, the Cromhall Quary, in Avon, hasyielded the specimens assigned to Agnosphitys cromhallensis, thedinosaur affinity of which is controversial (Fraser et al., 2002;Langer, 2004; Yates, 2007a). Perhaps, the youngest dinosaur-bearing deposits of the European Triassic are the Rhaetianbeds of Normandy (northern France), Somerset-Avon (south-west England), Mid-Glamorgan (south Wales), and Belgium.These include the indeterminate theropod ‘‘Zanclodon’’ cam-
brensis (Rauhut & Hungerbuhler, 2000; Galton, 2005a), thecoelophysoid Lophostropheus airelensis (Ezcurra & Cuny, 2007),sauropodomorphs like Camelotia borealis (Storrs, 1994; see alsoGodefroit & Knoll, 2003), and the very unlikely record ofa stegosaur (Galton, 2005a; Irmis et al., 2007b). In addi-tion, a possible theropod has been recovered recently fromthe Rhaetian beds of Lipie Slaskie, Poland (Dzik, Sulej& Niedzwiedzki, 2008). Isolated ‘‘dinosaur’’ teeth, mainlyassigned to Ornithischia, have also been reported exten-sively from Norian-Rhaetian European strata (Weishampelet al., 2004), none of which was recently confirmed (Butler,Porro & Heckert, 2006; Irmis et al., 2007b). The ‘‘Eurasian’’record of Norian-Raethian dinosaurs (Fig. 7) is completed bythe basal sauropodomorphs of the Nam Phong Formation,Thailand, that include Isanosaurus attavipachi (Buffetaut et al.,1995, 2000).
The record of Triassic dinosaurs in western USA wasrecently reviewed by Nesbitt et al. (2007; see also Parkeret al., 2005; Ezcurra, 2006; Nesbitt & Chatterjee, 2008).No compelling evidence of either sauropodomorphs orornithischians was found, and only coelophysoids werepositively identified, along with putative basal saurischians(herrerasaurs) and basal theropods (Fig. 7). Given thatthe Santa Rosa Formation ‘theropod’ (Heckert, Lucas &Sullivan, 2000) was considered an indeterminate archosaur(Nesbitt et al., 2007), the oldest dinosaur from westernUSA, and possibly the oldest known neotheropod so far
is ‘‘Camposaurus arizonensis’’, an indeterminate coelophysoidfrom the Placerias Quarry (Bluewater Creek Member, base ofthe Chinle Formation), northern Arizona (Hunt et al., 1998).Younger records of coelophysoids include Coelophysis bauri
(Colbert, 1989; Colbert et al., 1992; ICZN, 1996; Spielmannet al., 2007), the material described by Cope (1889) andPadian (1986), as well as other specimens (Ezcurra, 2006;Irmis et al., 2007a; Spielmann et al., 2007), along with some ofthose attributed to Gojirasaurus quayi (Carpenter, 1997; Nesbittet al., 2007). All these come from Norian deposits referredto the Chinle Formation (Petrified Forest Member and‘‘siltstone member’’), in central New Mexico and Arizona,and the Bull Canyon Formation (Dockum Group), in eastNew Mexico and west Texas (Nesbitt et al., 2007). Amongnon-theropod dinosaurs, whereas Caseosaurus crosbyensis (Huntet al., 1998) was regarded as an indeterminate dinosauriform(Nesbitt et al., 2007), putative herrerasaurs occur in thePetrified Forest Member (Chindesaurus bryansmalli) in Arizona,as well as in the Bull Canyon Formation, which also yieldeda putative basal theropod (Nesbitt et al., 2007; Nesbitt &Chatterjee, 2008). In terms of age, except for those of thePlacerias Quarry, all reliable dinosaur occurrences in theTriassic of western North America are considered youngerthan the Blue Mesa Member of the Chinle Formation, inArizona (Nesbitt et al., 2007), which has been radiometricallydated as 219.2 ± 0.7 Myr (Irmis & Mundil, 2008).
In conclusion, although a Middle Triassic (Ladinian) originof dinosaurs might be hypothesized, the oldest definitiverecords of the group date from about 230 million years ago.This corresponds to the Carnian stage of the Late Triassic.Radiometric dating of different levels of the IschigualastoFormation, Argentina (Rogers et al., 1993; Shipman, 2004)suggests that after about 20 million years, i.e. within the latestTriassic, a more diverse (Fig. 8), and specially more abundantand widespread dinosaur fauna was already present (Benton,1983a; Ezcurra & Novas, 2008), as represented by the LosColorados Formation and correlated assemblages from otherparts of the world (Fig. 7).
(2) The evolutionary tree of early dinosaurs
‘‘Early dinosaurs’’ are broadly understood here as all putativerepresentatives of the group collected from Ischigualastianstrata, as well as younger dinosaurs, the position of whichwithin Ornithischia, Theropoda, or Sauropodomorpha, isyet to be firmly established (Table 2). These include reason-ably well-known forms such as Herrerasaurus ischigualastensis,Pisanosaurus mertii, Staurikosaurus pricei, Eoraptor lunensis, Saturna-
lia tupiniquim, and Panphagia protos, as well as more fragmentarytaxa (Huene, 1910, 1942; Chatterjee, 1987; Long & Murry,1995; Bonaparte et al., 1999; Fraser et al., 2002; Langer,2004; Nesbitt et al., 2007). Pisanosaurus has always been con-sidered an ornithischian dinosaur (Thulborn, 1971; Galton,1972; Bonaparte, 1976), while Herrerasaurus and Staurikosaurus
were assigned into the base of Saurischia by pre-cladisticworks (Reig, 1963; Benedetto, 1973; Galton, 1977), althoughmore specific affinities to either sauropodomorphs (Reig,1970; Colbert, 1970; Van Heerden, 1978) or theropods
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

72 Max C. Langer and others
Table 2. Taxonomic assignment of ‘‘early dinosaurs’’, as recently given by different authors.
Taxon Proposed affinity
Agnosphitys cromhallensis Non-dinosaur; Fraser et al. (2002)Dinosauria (partim); nomen dubium; Langer (2004)Basal Theropoda; Yates (2007a)Basal Sauropodomorpha (Guaibasauridae); Ezcurra (2008)
Aliwalia rex Eucnemesaurus fortis; Yates (2007a)Alwalkeria maleriensis Basal Saurischia (partim); Remes & Rauhut (2005)Caseosaurus crosbyensis Dinosauriformes Nesbitt et al. (2007)Chindesaurus bryansmalli Herrerasauridae Irmis et al. (2007a)
Basal Theropoda; Yates (2007a)Basal Saurischia (partim); Nesbitt et al. (2007)
Eoraptor lunensis Basal Theropoda; Sereno (1999); Ezcurra (2006)Basal Saurischia; Langer (2004); Yates (2005)
Guaibasaurus candelariensis Basal Saurischia (Guaibasauridae); Bonaparte et al. (2007)Basal Theropoda; Yates (2007a), Langer et al. (2007a)Basal Sauropodomorpha; Ezcurra (2008)
Herrerasauridae Basal Theropoda; Novas (1996); Sereno (1999)Non-dinosaur; Fraser et al. (2002)Basal Saurischia, Langer (2004); Yates (2005); Ezcurra (2006)
Herrerasaurus ischigualastensis Herrerasauridae Novas (1992b)Panphagia protos Sauropodomorpha Martinez & Alcober (2009)Pisanosaurus mertii Ornithischia Sereno (1999); Butler et al. (2007)Saltopus elginensis Dinosauriformes Rauhut & Hungerb uhler (2000); Langer (2004)Saturnalia tupiniquim Stem-Sauropodomorpha; Langer & Benton (2006)
Basal Saurischia (Guaibasauridae); Bonaparte et al. (2007)Sacisaurus agudoensis cf. Ornithischia; Ferigolo & Langer (2007)
Non-dinosaur; Brusatte et al. (2008a)Silesaurus opolensis cf. Ornithischia; Ferigolo & Langer (2007)
Non-dinosaur; Langer & Benton (2006)Spondylosoma absconditum Non-dinosaur; Galton (2000)
cf. Herrerasauridae; Langer (2004)Staurikosaurus pricei Herrerasauridae Novas (1992b)Teyuwasu barberenai Dinosauria (partim); nomen dubium; Langer (2004)
(Galton, 1973; Bakker & Galton, 1974) were also claimed.On the contrary, early cladistic studies (Fig. 9) depictedStaurikosaurus and Herrerasaurus basal to the Ornithischia+Saurischia dichotomy (Gauthier, 1986; Brinkman & Sues,1987; Benton, 1990; Novas, 1992b), thus outside Dinosauriaon its emerging monophyletic understanding (Gauthieret al., 1989), whereas contemporaneous studies never ques-tioned the ornithischian affinity of Pisanosaurus (Novas, 1989;Sereno, 1991b). These investigations set the basis to futureresearch on basal dinosaur phylogeny, accepting the groupas a monophyletic entity solely composed of Ornithischiaand Saurischia, the latter including equally monophyleticSauropodomorpha and Theropoda. Besides, Novas (1992b)placed Herrerasaurus and Staurikosaurus into a monophyleticHerrerasauridae, a hypothesis almost never contested since.
During the early nineties, new discoveries from theIschigualasto Formation, including almost complete skele-tons of Herrerasaurus ischigualastensis (Sereno & Novas, 1992,1993; Novas, 1993, Sereno, 1993) and the first record ofEoraptor lunensis (Sereno et al., 1993), were announced alongwith a new hypothesis of basal dinosaur relationships. Thiswas advocated based on independent numerical analysesperformed by Sereno et al. (1993; see also Sereno, 1999)
and Novas (1996) that found nearly identical results (Fig. 9).The Herrerasauridae was depicted as the sister-taxon ofNeotheropoda, while Eoraptor was considered the basal-mosttheropod. Apomorphic traits supporting the theropod affin-ity of Eoraptor and Herrerasauridae were given as includingcaudally curved tooth crowns not expanded at the base, abroad axial intercentrum, elongated humerus and manus,deep extensor pits on the distal end of metacarpals I–III,and narrow metacarpal IV, as well as by typical preda-tory adaptations shared by herrerasaurids and theropods(Fig. 10), e.g. intramandibular joint, craniomandibular jointat about the same level as the tooth rows, and manualdigits II and III with elongated penultimate phalangesand strongly curved unguals with enlarged flexor tubercles(Langer & Benton, 2006; Sereno, 2007b; but see Butler et al.,2007). More recently, Chindesaurus bryansmalli was describedas a herrerasaurid (Long & Murry, 1995; Novas, 1997a;Sereno, 1999), a phylogenetic hypothesis accepted by mostauthors up to the late nineties. However, the suggestions thatChindesaurus forms a clade with either Herrerasaurus (Novas,1997a) or Staurikosaurus (Sereno, 1999) were not supported byrecent studies (Langer, 2004; Nesbitt et al. 2007; Bittencourt
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 73
Fig. 9. Main alternative phylogenetic hypotheses depicting the interrelationships of ‘‘early dinosaurs’’, modified from the citedsources. Arrows indicate stem-based taxa and black circles node-based taxa. Names applied as in Table 1, not as in the referredpublications. Abbreviations as follows: O, Ornithischia; T, Theropoda; H, Herrerasauridae; E, Eusaurischia.
& Kellner, 2009), which place the North American taxonbasal to the more ‘‘typical’’ herrerasaurids.
Around the turning of the 20th Century, the descriptionof new basal dinosauriforms such as Saturnalia tupiniquim andAgnosphitys cromhallensis, led to the proposal of novel phylo-genetic hypotheses of basal dinosaur relationships (Fig. 9).Langer et al. (1999) described Saturnalia as the basal-mostsauropodomorph, within alternative phylogenetic arrange-ments depicting Herrerasaurus and Staurikosaurus as eithersaurischians basal to the Theropoda+Sauropodomorphadichotomy, or as a monophyletic sister taxon to Dinosauria.Fraser et al. (2002) described Agnosphitys as the sister taxonto Dinosauria, favouring a position of Herrerasaurus outsidethat clade, but these results were not replicated by anyquantitative analyses performed since then.
In a comprehensive analysis of basal theropod phylogeny,Rauhut (2003) recovered Eoraptor lunensis and herrerasauridsas basal theropods, as first proposed by Sereno & Novas(1992). However, most subsequent studies, including somefocused on basal theropods (Yates, 2005; Smith et al., 2007),contradicted that hypothesis of basal dinosaur relation-ships. Yates (2003b) conducted a cladistic study of basalsauropodomorphs, and found a new phylogenetic arrange-ment among saurischians where herrerasaurids were con-sidered the sister group of all other components of theclade, termed Eusaurischia by Padian et al. (1999). Yates(2003b) also found Saturnalia tupiniquim as the most basalsauropodomorph, as previously claimed by Langer et al.(1999) and mainly accepted since. New comprehensive anal-yses by Langer (2004; see also Langer & Benton, 2006) inde-pendently came to similar results (Fig. 9). These corroborated
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

74 Max C. Langer and others
Fig. 10. Selected anatomical features of Herrerasaurus ischigualastensis, depicting a combination of apomorphic traits shared withNeotheropoda (underlined) and plesiomorphic states relative to the Eusaurischia condition (non underlined). Asterisks indicate traitsalso seen in basal ornithischians according to Butler et al. (2007). Skeletal reconstruction based on Sereno (1993). Scale bar = 10 cm.
the position of herrerasaurids as basal saurischians, addingEoraptor lunensis as the sister taxon to Eusaurischia, andGuaibasaurus candelariensis as a basal theropod. Indeed, sev-eral eusaurischian apomorphies are lacking in herrerasaurids(Fig. 10) and/or Eoraptor, as exemplified by a short caudoven-tral premaxillary process, a nasal that possesses a caudolateralprocess and forms part of the dorsal border of the antor-bital fossa, caudal cervical vertebrae longer than cranialtrunk vertebrae, a large medial-most distal carpal, a stoutmetacarpal I with lateral distal condyle distally expanded,a long metacarpal II relative to metacarpal III, and anexpanded distal end of the ischium (Langer & Benton, 2006).
Subsequent studies broadly agree with the above scenario(Irmis et al., 2007a; Nesbitt & Chatterjee, 2008; Martinez& Alcober, 2009), but differ in minor details (Fig. 9). Ina study focused on the non-dinosaurian affinity of theputative coelophysoid Eucoelophysis baldwini (Sullivan & Lucas,1999), Ezcurra (2006) placed herrerasaurids as non-theropodsaurischians, but Eoraptor as a basal theropod, sister taxonof Neotheropoda. In a study of basal sauropodomorph phy-logeny, Upchurch et al. (2007) placed both Herrerasauridaeand Eoraptor as non-theropod saurischians, but consideredthe former group as the sister taxon of Eusaurischia. Yates(2007a, b) expanded his previous studies, adding Agnosphitys
cromhallensis, Guaibasaurus candelariensis, Chindesaurus bryansmalli,and Eoraptor lunensis to an analysis of basal sauropodomorphs.The former three taxa were found as basal theropods, withGuaibasaurus as the sister taxon of a clade including Chin-
desaurus plus Neotheropoda, and Agnosphitys as the mostbasal theropod. Yet, the theropod affinity of Chindesaurus
was challenged by Irmis et al. (2007a), who supported itsmore traditional relation to Herrerasaurus, both lying basal tothe sauropodomorph/theropod dichotomy, as also suggested
by Langer (2004) and Nesbitt et al. (2009). More recently, thediversity of basal members of the sauropodomorph lineagewas increased by the discovery of Panphagia protos (Martinez& Alcober, 2009) and the undescribed sister-taxon of Satur-
nalia tupiniquim (PVSJ 845; Ezcurra, 2008), both from theIschigualasto Formation, of Argentina. Further, Ezcurra(2008) also included Agnosphitys and Guaibasaurus in thatdinosaur lineage. Indeed, Bonaparte et al. (2007) has alreadyproposed a close relation between Guaibasaurus and Saturnalia,forming Guaibasauridae at the base of Saurischia. However,Langer (2004) suggested the theropod, possibly coelophysoid,affinity of Guaibasaurus (see also Upchurch et al., 2007; Langer,Bittencourt & Schultz, 2007a; Bittencourt, 2008).
Several putative basal dinosaurs were never included innumerical phylogenetic analyses, and their affinities areopen to scrutiny. Langer (2004) offered a comprehensivesummary of these records, but this has to be updated withnew information available since. Remes & Rauhut (2005)reassessed the affinity of Alwalkeria maleriensis, first described asa basal theropod (Chatterjee, 1987; Norman, 1990), but laterregarded as a dinosaur of uncertain (Novas, 1989, 1997a)or eusaurischian (Langer, 2004) affinities. Those authorsfound that the holotype represents a chimera, includingpseudosuchian and possible prolacertiform material, but alsosaurischian specimens. Aliwalia rex Galton, 1985b, on theother hand, previously regarded as a herrerasaurid (Galton,1985b; Paul, 1988) or a dinosaur of dubious affinities (Sues,1990; Galton & Van Heerden, 1998; Langer, 2004) wasshown to represent a junior synonym of Eucnemesaurus fortis,therefore a basal sauropodomorph (Yates, 2007a). Nesbittet al. (2007) recently reviewed the status of the isolatedilium previously assigned to Chindesaurus bryansmalli (Long& Murry, 1995), which constitutes the holotype of Caseosaurus
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 75
crosbyensis (Hunt et al., 1998) and considered the materialundiagnostic above Dinosauriformes. Other putative earlydinosaurs such as Saltopus elginensis, Spondylosoma absconditum,and Teyuwasu barberenai, have not been studied recently.Indeed, their uncertain affinities as proposed by Langer(2004) are provisionally accepted here (Table 2).
In conclusion, recent cladistic analyses of basal dinosaurrelationships agree in various aspects, which are accepted bymost of the authors mentioned above: (1) dinosaurs representa monophyletic group exclusive of forms such as Lager-
peton chanarensis, Marasuchus lilloensis, Pseudolagosuchus major,and Silesaurus opolensis; (2) Dinosauria is composed of twomain lineages, Saurischia and Ornithischia; (3) Pisanosaurus
mertii is a basal ornithischian; (4) Herrerasaurus ischigualastensis
and Staurikosaurus pricei belong into a monophyletic Her-rerasauridae; (5) Eoraptor lunensis, Guaibasaurus candelariensis,and herrerasaurids are saurischians; (6) Saurischia includestwo main groups, Theropoda and Sauropodomorpha; and(7) Saturnalia tupiniquim and Panphagia protos are basal membersof the sauropodomorph lineage.
On the contrary, several aspects of basal dinosaur phy-logeny remain controversial. These include the positionof herrerasaurids, Eoraptor lunensis, and Guaibasaurus cande-
lariensis as basal theropods or basal saurischians, and theaffinity and/or validity of various more fragmentary taxasuch as Agnosphitys cromhallensis, Alwalkeria maleriensis, Chinde-
saurus bryansmalli, Saltopus elginensis, Spondylosoma absconditum,and Teyuwasu barberenai. Other equally incomplete formshave been more thoughtfully studied, but while the affinitiesof Aliwalia rex are better understood, Caseosaurus crosbyen-
sis continues to be problematic. In a reappraisal of themethodologies employed in recent analyses of basal dinosaurrelationships, Sereno (2007b) highlighted that the lack ofconsensus regarding the same phylogenetic problematicsis mainly due to differences in character/character-statechoice and codification among authors. Indeed, it seems thatmore comprehensive studies, discussing these methodologi-cal issues, are necessary to achieve a better understanding ofthe phylogenetic relationships of basal dinosaurs. This is, inturn, essential to recognize the patterns leading to their earlyradiation and success during post-Triassic times.
(3) Geographical distribution of basal dinosaurs
The earliest records of dinosauromorphs and dinosauriformsbased on body fossils, and also most of the trustworthyrecords of the earliest dinosaurs come from southern SouthAmerica, especially Argentina (e.g. Bonaparte, 1975; Sereno& Novas, 1992; Sereno & Arcucci, 1993, 1994; Novas,1992b, 1996; Langer et al., 1999; Galton, 2000; Rogers et al.,2001; Langer, 2004; Ferigolo & Langer, 2007; Martinez &Alcober, 2009). However, relatively few sites representativeof terrestrial ecosystems of that time are known (Hammer,Collinson & Ryan, 1990; Lucas, 1998; Rogers et al., 2001;Weishampel et al., 2004) and no biogeographic hypothesisconcerning the area of origin of the dinosaurian cladescan be robustly tested (Parker et al., 2005). Nonetheless,whereas almost all south-Pangean tetrapod-bearing deposits
of Carnian age (Langer, 2005b) bear undisputed, even ifinconspicuous dinosaur records, the north-Pangean scenariois rather different, with no dinosaur positively identified incoeval tetrapod assemblages. Accordingly, an admittedlytentative scenario can be drawn, hinting at a southernPangean origin of dinosaurs.
Obviously, any biogeographical picture of dinosaur originshas to be backed up by the current phylogenetic hypothe-ses depicting the relationships of the basal members ofthe group and its sister taxa. Accordingly, recent finds ofbasal dinosauromorphs in Europe (Fraser et al., 2002; Dzik,2003) and North America (Ezcurra, 2006; Irmis et al., 2007a;Nesbitt & Chatterjee, 2008) came with new phylogeneticproposals, and indicate that those animals had a broadergeographical and chronostratigraphic distribution than pre-viously thought. Hypotheses that support an inclusive cladeof basal dinosauriformes (i.e. Silesauridae) as the sister taxonto Dinosauria (Nesbitt et al., 2007; Brusatte et al., 2008a)face the problem of a Ladinian ghost-lineage of ‘‘stem-dinosaurs’’ (Fig. 3A), but are roughly in agreement withthe southern origin scenario. Spondylosoma absconditum, fromsouth Brazil, could fill that temporal gap, but its atypicalmorphology and uncertain affinity (Langer, 2004) preventfurther scrutiny. The alternative pectinate topology (Ezcurra,2006) overcomes the ghost-lineage problem, but suggests thatnorth Pangean taxa represent the immediate outgroups toDinosauria (Fig. 3A), jeopardizing the ‘‘out of south Pangea’’model of dinosauromorph/dinosauriform/dinosaur radi-ation. The record of Ladinian-Carnian ‘‘dinosauri-form/dinosaur’’ footprints in various parts of the world (Mel-chor & De Valais, 2006; Thulborn, 2006; Marsicano et al.,2007) also hints at a broader distribution of these basal forms.
Late Triassic dinosaur records as a whole include bodyfossils from Europe, North and South America, India,Africa, and East Asia (Weishampel et al., 2004), as wellas putative tracks from Australia (Thulborn, 2000, 2006).This is congruent with the geographic configuration of thetime (Fig. 7), when the Pangea Supercontinent and the lackof extensive oceanic barriers would favour biotic expansion(Shubin & Sues, 1991). Indeed, several non-dinosaur tetrapodclades also achieved a widespread distribution during theLate Triassic and Early Jurassic (Benton, 1993), but itis important to note that no dinosaur clade had a trulyglobal distribution during Late Triassic times, especiallyin the Carnian Stage (Nesbitt et al., 2007), even if onlythe areas with tetrapod-bearing sites are considered. Thebiogeographic patterns of early dinosaur radiation are, infact, better analyzed having the proposed subdivisions of thegroup as a template.
The osteological record of Triassic ornithischians (Fig. 7)is restricted to three Norian forms: the South African Eocursor
parvus (Butler et al., 2007), an unnamed heterodontosauridfrom Patagonia (Baez & Marsicano, 2001), and Pisanosaurus
mertii, from northwestern Argentina (Casamiquela, 1967;Bonaparte, 1976), the latter of which may come from signifi-cantly older deposits. Various other remains, mostly isolated
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

76 Max C. Langer and others
teeth, from either Europe (Godefroit & Cuny, 1997; Gode-froit & Knoll, 2003) or North America (Chatterjee, 1984;Hunt, 1989; Hunt & Lucas, 1994; Heckert, 2002, 2004)had been assigned to the group. Along with footprints fromNorth America, Europe, and southern Africa, these mayhint at a broader Norian-Rhaetian geographical distribu-tion of ornithischians. Yet, neither the isolated teeth northe footprints can be unequivocally assigned to the group(Parker et al., 2005; Butler et al., 2006; Irmis et al., 2007b).Accordingly, as discussed by Irmis et al. (2007b), ornithis-chians do not seem to have been very diverse or abundantthrough the Triassic, and certain hypotheses of relationship(Sereno, 1991b, 1999; Xu et al., 2006) imply large gaps intheir fossil record. On the contrary, the usually acceptedbasal position of Pisanosaurus is in accordance with its olderage, as is the possible basal position of heterodontosaurids(Butler, Upchurch & Norman, 2008) and Eocursor (Butleret al., 2007) in relation to other ornithischians. The suggestedheterodontosaurid affinity of Pisanosaurus (Bonaparte, 1976;Galton, 1986; Crompton & Attridge, 1986; Butler et al., 2008)implies a minimal ghost-lineage for Genasauria sensu Butleret al. (2008), but is also in general agreement with a southPangean origin of ornithischians. Indeed, Laurasian occur-rences of the group can not be confirmed before the EarlyJurassic (Irmis et al., 2007b), when basal thyreophorans occurin North America, Europe, and Asia (Norman, Witmer &Weishampel, 2004a; Irmis & Knoll, 2008). A different pictureemerges with the tentative placement of Sacisaurus agudoensis
and especially Silesaurus opolensis as the most basal ornithis-chians (Ferigolo & Langer, 2007), but this hypothesis is stillto be backed up by numerical phylogenetic analyses. In anycase, the poorly documented early history of ornithischiansprevents any accurate biogeographic approach. Accordingto Irmis et al. (2007b), possible explanations for their rarityin Late Triassic rocks (e.g. sample bias, differential environ-mental occupation, systematic imprecision) are inconclusive.
Triassic saurischians have a much broader geographicdistribution (Rauhut & Hungerbuhler, 2000; Langer, 2004;Nesbitt et al., 2007). Indeed, basal members of the group,and putative members of the theropod and sauropodomorphlineages occur as body fossils in various Carnian bedsknown from south Pangea (South America, southern Africa,and India) as well as in Norian-Rhaetian deposits ofall continents except Australia and Antarctica (Fig. 7).However, most records of ‘‘basal saurischians’’ are, infact, records of saurischians of uncertain affinities, andonly Eoraptor lunensis and herrerasaurs have been, undercertain phylogenetic hypotheses, positively placed basal toEusaurischia. Well-known herrerasaurids are restricted tothe South American Carnian (Langer, 2004; Bittencourt& Kellner, 2009), including Herrerasaurus ischigualastensis andStaurikosaurus pricei. The clade remains unidentified in therelatively well-known post-Ischigualastian deposits of thatcontinent, hinting at its restricted stratigraphic distribution.Yet, herrerasaurids have been identified in Norian bedsof western USA (Long & Murry, 1995; Hunt et al., 1998;Irmis et al., 2007a; Nesbitt & Chatterjee, 2008), which would
represent the younger records of the group. In addition,given that the affinity of Chindesaurus bryansmalli to eitherof the South American herrerasaurids has been questioned(Langer, 2004; Bittencourt & Kellner, 2009), that NorthAmerican herrerasaur would most probably represent theremnant of a lineage parallel to the typical members of thegroup, and not a more derived outcome of that radiation.On the contrary, the latter seems to be the case for thespecimen described by Nesbitt & Chatterjee (2008), whichbears herrerasaurid apomorphies.
If herrerasaurids and/or Eoraptor lunensis are treated astheropods, the group would fit the ‘‘out of South Pangea’’radiation pattern, with a well-known record of basal formsin the Carnian of South America. In the alternativearrangement (Langer, 2004), the oldest theropod, i.e. thecoelophysoid ‘‘Camposaurus arizonensis’’ (Nesbitt et al., 2007),would not only come from North America, but also fromNorian-age deposits. If not filled by herrerasaurids and/orEoraptor this stratigraphic gap in theropod distribution isunexpected, given the occurrence of basal members ofthe sauropodomorph lineage in the Carnian (Langer et al.,1999; Ezcurra, 2008; Martinez & Alcober, 2009), andthe abundance of both saurischian groups later in theTriassic (Tykoski & Rowe, 2004; Galton & Upchurch, 2004).Indeed, mainly represented by coelophysoids (but see Nesbitt& Chatterjee, 2008), theropods become abundant duringNorian-Rhaetian times (Fig. 7), with body fossils recordedin North America (Jenkins et al., 1994; Nesbitt et al., 2007),Europe (Rauhut & Hungerbuhler, 2000; Ezcurra & Cuny,2007), Argentina (Arcucci & Coria, 2003; Ezcurra & Novas,2007a), India (F. E. Novas, personal observations), andperhaps South Africa (Ray & Chimsamy, 2002). The possibletheropod affinity (Yates, 2007a, b) of controversial Noriantaxa from Europe (Agnosphitys cromhallensis), North America(Chindesaurus bryansmalli), Brazil (Guaibasaurus candelariensis),and India (aff. Guaibasaurus) does not significantly change thisdistribution pattern. In comparison to the more abundantsauropodomorphs, osteological records of theropods arelacking in Norian-Rhaetian deposits from southwest Asia(Nam Phong Formation), suggesting that Pangean far-eastwas first reached by the herbivorous/omnivorous branch ofSaurischia. Yet, this fossil assemblage is imperfectly known(Buffetaut et al., 2000), and the absence of theropods mightsimply represent a circumstantial sample bias. The scarcity oftheropod body fossils of Late Triassic age in southern Africa issomewhat filled by ichnological evidence (Ellenberger, 1974;Olsen & Galton, 1984; Raath et al., 1990), and also inferredfrom their well-known Early Jurassic record (Raath, 1969;Bristowe & Raath, 2004; Yates, 2005). Possible theropodfootprints are also known form Norian-Rhaetian deposits ofother parts of the world (Gatesy et al., 1999; Haubold & Klein,2002; Demathieu & Demathieu, 2004; Thulborn, 2006), butseveral of them have been questioned (King & Benton,1996; Marsicano et al., 2007; Lucas, 2007; Nesbitt et al.,2007), given their possible assignment to non-dinosauriandinosauromorphs. In any case, the overall record leads to ascenario of low abundance of Carnian theropods, followed
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 77
by a significant Norian radiation, when the group occupiedmost parts of Pangea.
Sauropodomorphs are surely the most abundant dinosaurgroup of Triassic times. Basal members of the lineage, i.e.Saturnalia tupiniquim and its allies, come from Ischigualastianbeds of South America (Langer et al., 1999; Da Rosa et al.,2006; Ezcurra, 2008; Martinez & Alcober, 2009), and pos-sibly southern Africa (Raath, 1996). This record could beenhanced by the ‘‘prosauropods’’ of the Lower Maleri For-mation referred to by Kutty & Sengupta (1989), but thesehave not been mentioned in more recent studies (Kutty et al.,2007), which refer the Upper Maleri ‘‘prosauropods’’ to aff.Guaibasaurus. Assuming the above identifications as correct,a southern radiation of ‘‘Saturnalia-like’’ forms may have pre-ceded the Norian diversification of true sauropodomorphs.The basal-most members of that group, Pantydraco caducus
and Thecodontosaurus antiquus (Yates, 2007a, b), come fromfissure-filling deposits of England and Wales of alleged Car-nian age, but this occurrence better fits the much broaderdistribution of later sauropodomorphs (Fig. 7). Indeed, theNorian-Rhaetian record of the group excludes only Antarc-tica, Australia, and continental North America (Vickers-Richet al., 1999; Galton & Upchurch, 2004; Nesbitt et al., 2007).The former two areas, however, lack well-sampled tetrapodfaunas of that age, and the absence of sauropodomorphscould represent a sampling bias. Indeed, the group can besaid to have had a nearly global Norian distribution, but thiswas not uniform through time and space. Most of the basal,non-Plateosauria (sensu Yates, 2007a) taxa were recordedin Europe (Yates, 2003b,c; Galton & Upchurch, 2004;Galton, 2007), while more derived forms are widespread(Yates, 2007a, b; Galton & Upchurch, 2004; Leal et al.,2004; Pol & Powell, 2007a). In this context, the lack ofsauropodomorphs in the Norian of continental North Amer-ica (Nesbitt et al., 2007), though not in Greenland (Jenkinset al., 1994), is intriguing. Indeed, this seems to represent atrue biogeographic pattern, given the abundance of well-sampled tetrapod-bearing deposits of that age in the region.The occurrence of the latest basal dinosauromorphs (Irmiset al., 2007a) and probable herrerasaurs (Nesbitt & Chatter-jee, 2008) in those faunas may also result from the causes thatdrove this pattern. As for basal sauropods, previous studiessuggested that their distribution was initially restricted tosoutheastern Asia, expanding throughout Pangea by theLate Triassic-Early Jurassic (Gillette, 2003). Yet, recent phy-logenetic hypotheses (Yates, 2007a, b; Smith & Pol, 2007;Upchurch et al., 2007) have positioned south Pangean formslike Antetonitrus ingenipes, Blikanasaurus cromptoni, Lessemsaurus
sauropoides, and Melanorosaurus readi, close to, or at the baseof Sauropoda. This suggests a wider distribution of earlymembers of the group, a pattern that seems to fit bet-ter the footprint record (Wilson, 2005). In any case, mostTriassic ichnological evidence of sauropodomorphs was con-sidered poorly substantiated (Lockley et al., 1994; Rainforth,2002, 2003).
IV. ECOLOGY OF THE DINOSAUR RADIATION
(1) The Triassic scene
During the Triassic, following the Late Permian maximumcoalescence of Pangea, most continental areas remainedforming a single landmass (Scotese, 2002; Golonka, 2002;Blakey, 2006). Towards the end of the period, major rift zonesstarted to develop, especially along the Atlantic marginsof North America and North Africa, accounting for theseparation between Laurasia and Gondwana (LeTourneau& Olsen, 2003; Golonka, 2007). Besides, the climateexperienced a trend towards higher instability comparedto Paleozoic settings (Holser & Magaritz, 1987; Kent &Muttoni, 2003). The Triassic palaeoclimate was reviewedin various landmark publications (Tucker & Benton, 1982;Hallam, 1985; Parrish, 1993; Crowley, 1994; Golonka &Ford, 2000), which suggest a warm period, when polar icecaps were absent (Frakes, Francis & Syktus, 1992). Further,a latitudinal zonation seems to have been present, with anarid equatorial/tropical belt, a seasonally humid temperatezone, and mainly humid higher latitudes (Hallam, 1985; butsee Fraser, 2006). In the second half of the period, a highlyseasonal (monsoonal) humid climate prevailed over variousparts of the supercontinent (Parrish, 1993). Triassic biotasreflect the transitional nature of the time interval (Anderson &Anderson, 1993; Fraser, 2006), particularly when terrestrialtetrapod faunas are considered. The period starts with theimpoverished remaining diversity of the end-Permian massextinction (Benton, 2003), ending up with an essentiallymodern fauna, that includes the first representatives of thechelonian, lepidosaur, crocodilian, avian (in the form ofdinosaurs), and mammal lineages.
Part of the Triassic tetrapod diversity was inheritedfrom the Permian, when dicynodonts and limnarchiantemnospondyls reached their climax (King, 1988; Milner,1993). These groups experienced a later diversification withinthe Triassic, along with the first radiation of lineages of latestPermian origin, like procolophonoids (Spencer & Benton,2000), ‘‘protorosaurs’’ (Dilkes, 1998), archosaurs (Gower &Sennikov, 2000), and cynodonts (Botha, Abdala & Smith,2007). These tetrapod groups diversified through the EarlyTriassic, composing the core of the Middle Triassic pre-dinosaur terrestrial palaeocomunities; the ‘‘Kannemeyeroidepoch’’ of Ochev & Shishkin (1989). An example of suchfaunas is known from the Chanares Formation, Argentina(Bonaparte, 1982; Rogers et al., 2001) that is dominatedby herbivorous cynodonts (Massetognathus) and dicynodonts(Dinodontosaurus), along with predatory cynodonts (Chiniquodon)and archosaurs (proterochampsids and ‘‘rauisuchians’’).Nearly coeval faunas were recorded in Brazil, Russia, andSouthern Africa, further including a variety of limnarchians,procolophonids, and rhynchosaurs (Ochev & Shishkin, 1989;Lucas, 1998; Abdala & Ribeiro, 2003). In addition, aspreviously discussed, the Chanares fauna also includes thehighest diversity of basal dinosauromorphs (Romer, 1971,1972a, b; Bonaparte, 1975; Sereno & Arcucci, 1993, 1994).
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

78 Max C. Langer and others
The oldest known dinosaurs are recorded in a particularfaunal context, in which rhynchosaurs, especially thegenus Hyperodapedon, became dominant primary consumersof various terrestrial faunas worldwide (Romer, 1962;Benton, 1983b). These were recorded in the Hyperodapedon
Assemblage-Zone of the Santa Maria Formation (Fig. 11A),the lower part of the Ischigualasto Formation, theLossiemouth Sandstone Formation, and the Lower MaleriFormation (Langer, 2005b). Apart from rhynchosaurs andthe first dinosaurs, these faunas collectively encompasslimnarchian temnospondyls (Marsicano, 1999; Sengupta,2003), various archosaurs such as proterochampsids (Price,1946; Sill, 1967), ‘‘rauisuchians’’ (Huene, 1942; Alcober,2000), poposauroids (Alcober & Parrish, 1997), aetosaurs(Heckert & Lucas, 2002), phytosaurs (Chatterjee, 1978),crocodylomorphs (Bonaparte, 1982; Ezcurra, Lecuona &Irmis, 2008), and ornithosuchids (Benton & Walker, 1985),carnivorous (Martinez, May & Forster, 1996; Bonaparte& Barberena, 2001; Abadala & Gianinni, 2002) andherbivorous (Bonaparte, 1962; Chatterjee, 1982; Hopson,1985) cynodonts; as well as dicynodonts (Cox, 1965).A similar faunal content was recorded in putatively coevalfaunas that lack rhynchosaurs and dinosaurs such as thatof Krasiejow, Poland (Dzik & Sulej, 2007), and the base ofthe Irohalene Member (Timesgadiouine Formation, ArganaBasin), Morocco (Jalil, 1996). In fact, apart from theappearance of some archosaur groups, and the dominance ofrhynchosaurs, the oldest dinosaur-bearing terrestrial faunasare not significantly different from those of Middle Triassicage.
Dinosaurs remain inconspicuous in younger faunas ofNorian age, as seen at the base of the Los ColoradosFormation (Caselli, Marsicano & Arcucci, 2001) and theCaturrita Formation (Langer et al., 2007b), in South America,and in some possibly coeval North American fossil assem-blages (Langer, 2005b), i.e. Sanfordian faunas of the NewarkSupergroup; Camp Springs Member, Dockum Group; andPopo Agie Formation (Huber, Lucas & Hunt, 1993; Lucas,1998). These faunas include metoposaurids, procolophonids,sphenodontians, Hyperodapedon, dicynodonts, traversodontidand mammal-like cynodonts, as well as various pseudo-suchians (aetosaurs, phytosaurs, poposauroids, and possible‘‘rauisuchids’’). Later Norian deposits include a greater num-ber of dinosaur records, within a slightly dissimilar faunalcontext; the ‘‘Prosauropod’’ Empire of Benton (1983a).As recorded from the top of the Los Colorados Forma-tion, the fauna of La Esquina (Bonaparte, 1982; Caselliet al., 2001) includes some of the oldest turtles, alongwith crocodyliforms, ‘‘remaining’’ pseudosuchian lineages(aetosaurs, ornithosuchids, ‘‘rauisuchians’’), and mammal-like cynodonts (Fig. 11B). Putatively coeval faunas of otherparts of the world, especially South Africa (Anderson, Ander-son & Cruickshank, 1998), Europe (Benton, 1994a), andNorth America (Long & Murry, 1995), further includevarious temnospondyls and phytosaurs, inconspicuous dicyn-odonts (Dzik et al., 2008), the latest traversodontids (Hopson,1984), as well as the first mammals (Lucas & Luo, 1993).
(2) Lucky break?
Palaeoecological aspects of the early radiation of dinosaursand its correlation to Late Triassic extinctions andcorresponding biotic/environmental changes have beenaddressed by various classical and more recent studies(Colbert, 1958; Benton, 1983a; Charig, 1984; Olsen et al.,2002; Tanner, Lucas & Chapman, 2004; Brusatte et al.,2008a, b). Focus has been given to two inferred massextinction events, at the Carnian-Norian and Triassic-Jurassic boundaries, and two alternative scenarios for therise of the dinosaurs, the so-called ‘‘competitive’’ and‘‘opportunistic’’ models. Studies from the mid-late 20thCentury postulated that the replacement of various tetrapodgroups, notably pseudosuchians and therapsids, by dinosaurswas a long-term affair driven by competition during theLate Triassic (Cox, 1967; Charig, 1980; Bonaparte, 1982).Its outcome would have been the dominance of dinosaursover terrestrial ecosystems from Norian/Jurassic onwards,thanks to their ‘‘superiority’’ relative to the outcompetedcontemporary tetrapods, pushed to extinction. Usually, theimproved locomotory capability of the fully erect, bipedalearly dinosaurs was considered the most notable advantage ofthe group (Charig, 1972, 1984), but their inferred advancedphysiology has also been mentioned (Bakker, 1971). Fromthe 1980s onwards, Benton (1983a, 1984, 1991) advocatedan alternative model, based on which the Triassic radiationof dinosaurs was faster, opportunistically occupying adaptivezones emptied by the extinction of rhynchosaurs, therapsids(dicynodonts and some cynodonts), and pseudosuchians(phytosaurs, aetosaurs, rauisuchians). More recently, Brusatteet al. (2008a) demonstrated that Norian pseudosuchiansoccupyied more morphospace and showed similar ratesof character evolution compared to dinosauromorphs/dinosaurs. Indeed, this dismisses the classical ‘‘competitive’’model, based on which those archosaurs were graduallyreplaced by dinosaurs. Yet, the scenario seems to be morecomplex in terms of patterns and timing of biotic turnovers,as discussed below.
Of the two proposed Late Triassic extinction events(Benton, 1986a, 1997), the end-Triassic is much betterdocumented in the literature than the end-Carnian, whichis often contested as minor or non-existent (Olsen & Sues,1986; Olsen, Shubin & Anders, 1987; Hallam, 1990; Fraser& Sues, 1994; Hunt et al., 2002). Classical studies revealthe final demise of conodonts and a severe reductionin the diversity of sponges, scleractinian corals, molluscs(ammonoids, gastropods, and bivalves), and brachiopods inthe sea (Hallam, 1981; Raup & Sepkoski, 1982; Sepkoski,1982, 1990), along with extinctions of insects (Benton, 1989)and tetrapods on land (Olsen & Sues, 1986; Benton, 1994b).Causes proposed for the end-Triassic mass extinction rangefrom sea level change (Hallam, 1990), to the impact ofone or more extraterrestrial bolides (Olsen et al., 2002)and the establishment of the Central Atlantic MagmaticProvince (Marzoli et al., 1999). The latter two might haveled to an increase in the levels of atmospheric CO2, and so‘‘greenhouse’’ warming (McElwain, Beerling & Woodward,
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 79
Fig. 11. Reconstruction of two dinosaur-bearing fossil assemblages of the South American Late Triassic. (A) Alemoa fauna (SantaMaria Formation), Carnian of south Brazil, depicting from left to right the aetosaur Aetosauroides sp.; the rhynchosaur Hyperodapedonmariensis; the stem-sauropodomorph Saturnalia tupiniquim (group on background); the cynodont Prozoostrodon brasiliensis (in front), andthe herrerasaurid Staurikosaurus pricei. (B) La Esquina fauna (Los Colorados Formation), Norian of northwestern Argentina, depictingon the left (from back to front), a group of the sauropodomorph Riojasaurus incertus; the ‘‘rauisuchid’’ Fasolasuchus tenax, and thecynodont Chaliminia musteloides; on the right (from back to front), the crocodyliform Hemiprotosuchus leali, and the basal theropodZupaysaurus rougieri subduing the ornithosuchid Riojasuchus tenuiceps. Drawings by Jorge Blanco.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

80 Max C. Langer and others
Fig. 12. Distribution of medium to large sized terrestrial amniotes along the Late Triassic and Early Jurassic and the rise of dinosaurs.Timeline from Gallet et al. (2003). Distribution of carnivorous (grey columns) and herbivorous/omnivorous (black columns) tetrapodsmodified from Benton (1994a), according to Dilkes (1998), Abdala & Giannini (2002), Abdala & Ribeiro (2003), Thulborn & Turner(2003), Lucas & Tanner (2005), Langer et al. (2007c), and Brusatte et al. (2008a). 1, Saturnalia tupiniquim; 2, Herrerasaurus ischigualastensis;3, Guaibasaurus candelariensis; 4, Eocursor parvus; 5, Plateosaurus engelhardti; 6, Liliensternus liliensterni; 7, Massospondylus carinatus; 8, Vulcanodonkaribaensis; 9, Heterodontosaurus tucki; 10, Dilophosaurus wetherilli; 11, Scelidosaurus harrisoni. Silhouettes (roughly at the same scale) adaptedfrom various sources. Mys, million years before recent; Rhaet., Rhaetian.
1999). Yet, Tanner et al. (2004; see also Bambach, Knoll &Wang, 2004) compiled evidence to reject what they call ‘‘themyth of a catastrophic extinction at the Triassic-Jurassicboundary’’. Indeed, as already hinted by some (Benton,1994b; Cuny, 1995; Ezcurra & Cuny, 2007), although variouswell-known Triassic amniote groups have no Jurassic record,some might have gone extinct before the Triassic upperboundary (Fig. 12).
The pioneering studies of Benton (1983a, 1986a, 1989,1994b), which first challenged the long-term ‘‘competitive’’model of dinosaur radiation, also advocated that the mainturnover of terrestrial faunas occurred at the Carnian-Norian, rather than at the Triassic-Jurassic boundary.This would have been characterized by the extinction ofvarious tetrapod groups, connected to climatic/floral changes(Tucker & Benton, 1982). Problems related to that modelinclude: (1) new data suggest the survival, at least untilthe initial stages of the Norian, of taxa believed to havegone extinct at the end of the Carnian (Fig. 12); (2) lackof synchronicity between the climatic/ floral changes ofnorth and south Pangea. Indeed, dicynodonts occur inNorian faunas of South (Langer et al., 2007c) and NorthAmerica (Long & Murry, 1995), in the latest Triassic ofPoland (Dzik et al., 2008), and perhaps in much younger
assemblages as well (Thulborn & Turner, 2003). In addition,the Caturrita Formation, of south Brazil, has yielded thelatest (Norian) remains of proterochampsid archosaurs andrhynchosaurs (Langer et al., 2007c), while lagerpetonidsand herrerasaurids were recorded in the Norian of USA(Irmis et al., 2007a; Nesbitt & Chatterjee, 2008; Nesbittet al., 2009). The diversity of chiniquodontid cynodonts, onthe other hand, has been reduced to a single genus ofAnisian-Carnian distribution in South America and southernAfrica (Abdala & Giannini, 2002; Abdala & Smith, 2009).Accordingly, its absence in Norian strata does not representthe demise of a well-established lineage, as is also thecase of single-genus ‘‘families’’ such as Pisanosauridae andScleromochlidae (Benton, 1994b). On the other hand, itis important to stress that rhynchosaurs and dicynodontsget less common in Triassic faunas after the Carnian. Thisabundance shift, rather than their extinction, could indeedprovide some evidence for a biological crisis at the Carnian-Norian boundary. In the marine realm, classical studiessuggested that invertebrate extinctions were not conspicuousat the end of the Carnian (Simms & Ruffell, 1990), onlyfew groups suffering a moderate loss of diversity (Schafer& Fois, 1987; Smith, 1988). Yet, more recent data onthe so-called Reingraben Turnover suggest that a major
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 81
restructuring of marine ecosystems occurred at the Carnian-Norian boundary (Hornung & Brandner, 2005; Stanley,2006; Hornung, Krystyn & Brandner, 2007b).
Despite Cornet & Olsen’s (1990) statement againstsignificant floristic changes across the Carnian-Norianboundary, this is assumed in a climate-driven fashion by thebiotic substitution model of Benton (1983a). Most authorsagree on the occurrence of a monsoonal humid phase(Fig. 12), minimally affecting the northern Tethyan realmduring the mid-late Carnian (Simms, Ruffel & Johnson,1994; Hornung et al., 2007a). Yet, although a humid phasewas also recorded in the Late Triassic of other parts ofthe world, it seems to have lasted until later within theperiod in those areas. Prochnow et al. (2006) suggest thatapproximately during the deposition of the Petrified ForestMember of the Chinle Formation, presently considered ofNorian age (Nesbitt et al., 2007), western North Americawas experiencing a period of increasing humidity. Similarly,the Santa Maria Supersequence, in south Brazil, showsthe progressive replacement of an ephemeral anastomosedfluvial-lacustrine system by a perennial braided fluvial systemthroughout the Carnian and Norian (Holz & Scherer, 2000;Zerfass et al., 2003), whereas the Norian upper third of theIschigualasto Formation bears evidence of a humidity peak,based on plant taphonomy and the occurrence of argiliticpalaeosols (Colombi & Parrish, 2008). Besides, Dicroidium-floras occur through the entire sequence, as is also the casein other parts of south Pangea (White, 1990; Anderson &Anderson, 1993), while bennettitaleans and conifers onlydominate after the Norian (Fig. 12).
What seems to occur during the Norian is a rise in theabundance of dinosaurs (Fig. 12). These are inconspicuousin Carnian faunas, representing about 5% of tetrapodfossils collected in the Hyperodapedon Assemblage-Zone of theSanta Maria Formation (Azevedo, Schultz & Barberena,1990; Langer et al., 2007c) and the lower third of theIschigualasto Formation (Bonaparte, 1982; Rogers et al.,1993). Carnivorous dinosaurs were proportionally moreabundant, representing nearly 40% of all terrestrial meat-eaters, and half of the medium- to large-sized predatorsof the latter assemblage (Rogers et al., 1993). By contrast,dinosaurs represent from 25 to 60% of the terrestrialtetrapods of classical Norian faunas (Benton, 1983a), notablyin South Africa (Kitching & Raath, 1984), Argentina(Bonaparte, 1982), and Europe (Benton, 1994a). This is betterdocumented for saurischians, especially with the notableradiation of ‘‘prosauropods’’. However, dinosaurs are stillminor components of possibly older Norian faunas such asthat of the Caturrita Formation (Langer et al., 2007c), wherethey represent about 15% of the known diversity. Althoughthe total number of dinosaurs registered in the Norian is atleast three times higher than in the Carnian, Ezcurra & Novas(2008) emphasize that dinosaur diversity in the IschigualastoFormation surpasses that of most Norian stratigraphic units(Fig. 7). This may imply that their radiation in the latterstage resulted in a more abundant and disparate (Brusatteet al., 2008b), but not necessarily more diverse, dinosaur
fauna, leaving the greater total diversity of Norian dinosaursto be explained based on the existence of more tetrapod-bearing sites of that age. Yet, various main dinosaur groupsappear to have originated and/or radiated during the Norian,as is the case for heterodontosaurids, coelophysoids, and‘‘prosauropods’’.
Moreover, as discussed by Novas (1997b; contra Benton,1983a), the Late Triassic dinosaur rise did not occur inan empty ecospace. Instead, dinosaurs radiated during theIschigualastian despite the high diversity and abundanceof other tetrapods. Especially in the case of carnivores,it is difficult to deny a degree of overlap in the use offood resources by sympatric species of similar size (Glen &Dickman, 2008); whereas modern analogues suggest thatfeeding niche overlap is less significant among herbivores(Plumptre, 1996; Begon, Harper & Townsend, 1996, p. 778).In this context, it is possible to envisage strictly herbivorousTriassic dinosaurs, which were not many (see SectionIV.3), using plant resources not previously exploited infull. Yet, counterevidence is given by the non-overlap ofthe rhynchosaur bearing Norian assemblages with youngerfaunas of that age (Fig. 12), in which dinosaurs are moreabundant (Langer, 2005a). In this context, the Norianradiation of herbivorous dinosaurs could be linked toecological release, given the extinction of rhynchosaurs (seealso Brusatte et al., 2008b). The corollary, as suggestedby Novas (1997b), is that competitive pressure of non-dinosaur herbivores may account for the low abundanceof herbivorous dinosaurs in the Carnian.
Whatever the physical causes of the end-Triassic event,direct ‘‘extermination’’ is unlikely to have been the onlyagent of the extinctions. Instead, the exacerbation ofbiotic interactions (including competition) in a changingenvironment probably also played a major role. In thatcontext, the already abundant and diverse dinosaurs mayhave had the key adaptations to succeed and expand,within the shifting environment that drove various othertetrapods towards extinction. This is the second explanationoffered by Brusatte et al. (2008a) accounting for the extinctionof pseudosuchians, and not dinosaurs, at the Triassic-Jurassic boundary. It implies a circumstantial ‘‘superiority’’of dinosaurs, while historical burden enforces the reference toa fully erect gait (Charig, 1972) and/or advanced physiology(Bakker, 1971; Ward, 2006) within the set of dinosaur‘‘advantages’’. This explanation is preferred here, given thatthe first is based on ‘‘chance’’ (Brusatte et al., 2008a), whichjust reflects cases ‘‘when our knowledge does not sufficefor prediction’’ (Popper, 1959). In fact, as acknowledgedby Popper (1959): we may not infer from the fact that anevent is chance-like that its elements are ‘due to chance’.This is because ‘‘chance’’ and lack of knowledge producethe same signatures, and can not be set apart in practice. Inour understanding, opportunistic and competitive scenariosof dinosaurs rise are not mutually exclusive, and competitionmay have played an important role in that radiation episode.Not as a long-term affair, but triggered by the physical eventsultimately linked to the end-Triassic extinction.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

82 Max C. Langer and others
Although the Early Jurassic total diversity of dinosaurgenera is not significantly higher than the Late Triassic(Wang & Dodson, 2006), it is usually accepted that anotherpulse of dinosaur radiation followed the Triassic-Jurassicextinction (Olsen et al., 2002). This is better measured by theorigin of certain lineages and exploitation of new ecologicalroles (Fig. 12). Among saurischians, large carnivores (Ezcurra& Novas, 2007a) and large quadruped herbivores (Yates &Kitching, 2003) were already known in the latest Triassic.In fact, the record of Zupaysaurus (Ezcurra & Novas, 2007a),and studies of Brusatte et al. (2008b), falsifies the hypothesisof theropod size increase due to ecological release afterthe Triassic-Jurassic boundary (Olsen et al., 2002; Lucas &Tanner, 2005). Yet the Early Jurassic saw the radiationof the Dilophosaurus-clade (Smith et al., 2007) and furtheracquisition of typical graviportal traits among sauropods(Barrett & Upchurch, 2007). The radiation of ornithischianswas more notable, with heterodontosaurids attaining theirdiversity peak and the origin of both neornithischians andthyreophorans (Butler et al., 2007). The latter representsthe debut of quadrupedal armoured forms, within amorphospace previously unoccupied by dinosaurs.
In conclusion, the radiation of dinosaurs comprises at leastthree landmark moments (Fig. 12), mainly characterizedby early diversification (Carnian); increase in diversityand, especially, abundance (Norian); and occupation ofnew niches (Early Jurassic). As previously mentioned, theCarnian diversification did not occur in an empty ecospace,but despite the abundance and diversity of contemporarytetrapods. The Norian increase in dominance might beconnected to climatic/ floristic changes and to the extinctionof herbivorous forms such as rhynchosaurs, but the timing ofthese events needs further investigation. The subtle Jurassicdiversification, on the other hand, seems to have occurred inthe aftermath of an extinction event (Brusatte et al., 2008a).Indeed, this might be an example of opportunistic radiationinto released ecospace (Benton, 1983a; Olsen et al., 2002;but see Brusatte et al., 2008b). Obviously, this does not fitthe notion that the ‘‘end-Triassic’’ tetrapod extinctions werescattered over the end of the period (Tanner et al., 2004).Indeed, the lack of various tetrapod groups in Rhaetian bedsand the less than expected dinosaur diversity increase in theEarly Jurassic seem to justify this latter scenario, but the‘‘diversity loss’’ of dinosaurs and other tetrapods during theRhaetian is likely to be due to sampling bias (Ezcurra &Cuny, 2007). In any case, post-Triassic tetrapod biodiversitycan not be understood as the outcome of a single event, butseems modeled by long-term coexistence of different groupsduring the Late Triassic. Punctual events of environmentalchange may have enhanced interaction among lineages,leading to the extinction of some terrestrial forms. This wasprobably topped by a final historical contingency at theTriassic-Jurassic boundary, when dinosaur circumstantial‘‘superiority’’ set the frame for the next 135 million years ofarchosaur evolution.
(3) Of legs and teeth: insights on the palaeobiologyof early dinosaurs
Non-crown-group archosaurs were all quadrupedal andcarnivorans (Charig, 1972; Parrish, 1986; Brusatte et al.,2008a). This general pattern was retained in basal membersof the crocodile-line (Bonaparte, 1984; Sereno, 1991a),although recent discoveries of bipedal (Nesbitt, 2007) andherbivorous/omnivorous (Parker et al., 2005) pseudosuchiansnotably amplified the disparity of these archosaurs. Thebasalmost dinosaurs, instead, were all bipedal, but it is notclear if this was also the case for more basal dinosauromorphs,and several instances of reversion to full or facultativequadrupedalism are known within the group (Padian, 1997c).The feeding habit of basal dinosaurs is even more difficult toassess. Basal dinosauromorphs have a rather unspecializeddentition, but typical carnivores and herbivores occur earlyin dinosaur evolution (Barrett, 2000).
As seen in the previous section, bipedalism is often consid-ered a key dinosaur feature that, along with a fully erect pos-ture, favoured the radiation of the group during Late Triassictimes. Yet, supporters of a polyphyletic origin of dinosaurssuggested that some quadrupedal lineages, particularlysauropods (Charig et al., 1965), have never had bipedal ances-tors, evolving directly from quadrupedal basal archosaurs(Fig. 2). The general acceptance of dinosaur monophyly, andthe identification of its sister taxa within gracile, and presum-ably bipedal Middle Triassic archosaurs (Gauthier, 1986) set-tled the new orthodoxy of originally bipedal dinosaurs, whichwas in turn challenged by more recent data. This includes thediscovery of a potentially quadrupedal basal dinosauromorph(Dzik, 2003), and new ichnological (Haubold & Klein, 2002)and biomechanical (Fechner, 2006) interpretations that hintat higher degrees of quadrupedalism among dinosaur precur-sors. It is consensual that herrerasaurs, basal theropods, andbasal ornithischians were fully bipedal (Carrano, 2000; But-ler et al., 2007), but the condition among sauropodomorphsis less clear (Cooper, 1981; Barrett & Upchurch, 2007). Inany case, only if all evidence in favour of basal dinosauro-morph/dinosaur quadrupedalism is accepted, and optimizedon a favourable phylogenetic framework (Fig. 13) does aquadrupedal/facultative bipedal origin of dinosaurs emergeas parsimoniously as a fully bipedal origin. Otherwise, the lat-ter hypothesis is always favoured if herrerasaurs are regardedas basal saurischians (Yates, 2003b; Langer, 2004; Ezcurra,2006; Irmis et al., 2007a). In fact, a fully quadrupedal dinosaurorigin is consistently ruled out, given that basal members ofthe sauropodomorph lineage, even if capable of walking onall fours, must have relied on bipedalism for higher speedlocomotion (Christian & Preuschoft, 1996; Upchurch, 1997a;Langer, 2003). Actually, this may be the case also of Silesaurus
opolensis, the unusual slender fore limbs of which may nothave endured the same amount of stress as the hind limbs did(see Farina, 1995). Evidently, more detailed biomechanicalstudies of basal dinosauromorphs are needed to recognize ifdinosaurs originated from facultative or fully bipedal ances-tors. At the moment, this is hampered by the scarce material
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 83
Fig. 13. Single hypothesis of basal dinosaur/dinosauromorph relationships in which the recognition of Lagerpeton, Silesaurus, andsauropodomorphs as quadrupedal/facultative bipedal (grey silhouettes) allows an equally parsimonious reconstruction of this gait,relative to a fully bipedal gait (white silhouettes), as ancestral to Dinosauria. White rectangles represent acquisition of bipedalism oneither a quadrupedal/facultative bipedal (C = convergences) or fully bipedal (A = apomorphy) scenario of dinosaur origins. Greyrectangles (R = reversions) represent acquisition of facultative bipedalism/quadrupedalism on a fully bipedal scenario of dinosaurorigins. Silhouettes adapted from various sources.
assigned to most of these forms, specially regarding their forelimb anatomy.
Classical scenarios of dinosaur dietary evolution (Chariget al., 1965) depict independent origins of the major groupsfrom carnivorous ‘‘thecodonts’’. In fact, also in the cladisticparadigm, typical ‘‘carnivorous teeth’’, i.e. pointed, caudallycurved, labiolingually flattened, unexpanded at the base, andwith finely serrated/denticulate keels, are usually acceptedas plesiomorphic for dinosaurs (Gauthier, 1986; Langer &Benton, 2006, but see Barrett, 2000). Indeed, although moreconical in likely piscivorous forms (Sill, 1967; Hungerbuhler,2000), teeth of basal archosaurs (Ewer, 1965; Gower, 2003)and pseudosuchians (Walker, 1964; Gower, 1999; Nesbitt,2003) mainly fit into that pattern. Triassic exceptionsare pseudosuchians that bear modified dentitions towardsomnivory/herbivory/scavenging (Walker, 1961; Parker et al.,2005). Given that known basal dinosauromorph teeth are alllabiolingually flattened, there is a good case that the abovedescribed pattern is indeed ancestral to the group as a whole.Dinosaur dentitions are, however, more heterogeneous.
Until recently, the dentition of non-dinosaur dinosauro-morphs was inferred from the rather fragmentary tooth-bearing bones referred to Lewisuchus admixtus (Romer, 1972a)and Marasuchus lilloensis (Bonaparte, 1975). The isolated par-tial jaw of Lewisuchus may not belong to the taxon, giventhat it was found disarticulated in a concretion with otherarchosaur taxa and seems larger in relation to the holotypeskeleton (PULR 01). The maxilla of Marasuchus (PVL 3870)has been consensually attributed to the taxon (Sereno &Arcucci, 1994), but its teeth are not well preserved enoughfor an accurate inference of its diet. Most of the crowns fitinto the plesiomorphic pattern described above, but morecaudal elements seem slightly distally expanded at the base,making them somewhat ‘‘leaf-shaped’’ (Bonaparte, 1975).Yet, the lack of further dental modifications (see Barrett,2000) precludes the assignment of an alternative diet to
Marasuchus, which probably fed on a variety of small ani-mals. Lately, however, putative basal dinosauromorphs withherbivorous adaptations have been discovered (Dzik, 2003;Ferigolo & Langer, 2007; Irmis et al., 2007a). Silesaurus opolen-
sis and Sacisaurus agudoensis bear an edentulous beak in thelower jaw, plus dental features usually associated with amore herbivorous diet in ‘‘prosauropods’’ and ornithischians(Galton, 1984; Crompton & Attridge, 1986; Sereno, 1991b),but lately given as evidence of omnivory (Barrett, 2000; Irmiset al., 2007b). Accordingly, although a full herbivore ancestryof dinosaurs can be dismissed, there is some evidence that astrictly carnivorous origin was also not the case.
Barrett (2000) comprehensively reviewed early dinosaurfeeding habits, remaining unsure about the ancestraldiet of the group. There is full agreement, however,on the carnivorous habits of theropods and herrerasaurs(Barrett, 2000; Bittencourt, 2008), while basal ornithischians(Irmis et al., 2007b) and typical ‘‘prosauropods’’ (Barrett& Upchurch, 2007) were most probably omnivorous. Thecondition in some small-sized basal saurichians such asEoraptor lunensis and Saturnalia tupiniquim is more uncertain.Eoraptor bore ‘‘leaf-shaped’’ rostral teeth, but its diet was mostprobably still based on small animals. On the other hand,most teeth of Saturnalia are ‘‘leaf shaped’’, and this animalis frequently referred to as bearing herbivorous adaptations(Barrett & Upchurch, 2007). However, the retention of finelyserrated tooth keels (Yates, 2003b) suggests that Saturnalia
was more carnivorous than any basal sauropodomorph.The optimization of dental patterns into current
hypotheses of early dinosaur phylogeny reveals variousalternative scenarios. When forms with less modifieddentition such as Marasuchus and Eoraptor are consideredcarnivores, the reconstruction of the ancestral dinosaurfeeding habit is ambiguous in most cases. Yet, if Eoraptor
plus herrerasaurs are considered basal theropods (Sereno,1999), and Silesaurus opolensis and Sacisaurus agudoensis placedin the sister clade to Dinosauria, then omnivory can be
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

84 Max C. Langer and others
Fig. 14. Hypotheses of basal dinosaur/dinosauromorph rela-tionships in which either omnivory (A) or carnivory (B) isunambiguously reconstructed as ancestral to Dinosauria. Whiteand black silhouettes/circles, respectively, indicate omnivorousand carnivorous taxa/optimizations. Silhouettes adapted fromvarious sources.
unambiguously regarded as plesiomorphic for the group(Fig. 14A). On the contrary, if Silesaurus and Sacisaurus areconsidered basal ornithischians (Ferigolo & Langer, 2007),and either Eoraptor or herrerasaurs basal saurichians (Langer& Benton, 2006), then dinosaurs are plesiomorphicallycarnivores (Fig. 14B). Yet, it is important to stress that theplesiomorphic dinosaurs tooth morphology does not strictlycompare to that of the typically carnivorous herrerasaursand theropods, or to the omnivorous/herbivorous pattern ofornithischians and ‘‘prosauropods’’. Teeth of basal forms asMarasuchus, Saturnalia, and Eoraptor, are less specialized, andthis was most probably also the case for the dinosaur commonancestor, whether it was a strict carnivore or able to includeplant material in its diet. In this context, each major dinosaurgroup seems to have independently acquired its typicaldental traits, in some cases along with a significant increasein size. Indeed, the omnivorous/herbivorous dentition ofbasal ornithischians and sauropodomorphs are sufficientlydifferent to preclude a common origin (Barrett, 2000). On thecontrary, the carnivorous teeth of herrerasaurs and theropodsare primary homologous (Bittencourt, 2008), and can be usedas evidence for the nesting of the former within the lattergroup. Alternatively, they might have arisen independently,as adaptations to the predatory habits of these animals(Langer & Benton, 2006).
The skin of basal dinosaurs has always been thought asscaly, but this view was recently challenged by the discovery
of Tianyulong confuciusi (Zheng et al., 2009), which providesthe second evidence of an ornithischian with integumen-tary filaments (see also Mayr et al., 2002). Its placementwithin Heterodontosauridae (Zheng et al., 2009), i.e. the sis-ter clade of all other ornithischians except Pisanosaurus (seeSection V.1), shows that at least some very basal ornithis-chians bore integumentary filaments. Accordingly, if theseare actually homologous to the ‘‘protofeathers’’ of theropoddinosaurs, the most recent common ancestor of saurischi-ans and ornithischians would also likely bear this kind ofepidermal coverage. Witmer (2009) is cautious about thedermal or epidermal origin of the integumentary structuresof Tianyulong, and their homology with those of theropods.Yet, further research demonstrating the epidermal origin ofthose filaments, a hypothesis currently supported by theirhollow structure (Zheng et al., 2009), would suggest that earlydinosaurs also bore integumentary filaments. If it was suffi-ciently abundant, the coverage could play a thermoregulatoryfunction (insulation), probably implying a higher thermalinertia (Regal, 1975, 1985; Unwin, 1998; Wu et al., 2004).
V. OUTCOMES OF A RADIATION
(1) Early ornithischian evolution
Although the concept of Ornithischia was defined only sometwenty years later (Seeley, 1888), the first dinosaur classifi-cation schemes already congregated most members of theclade we now know within a ‘‘natural group’’, e.g. OrthopodaCope, 1866. Indeed, ornithischian monophyly remains one ofthe few uncontroversial issues in dinosaur taxonomy (Nopcsa,1923, 1928; Huene, 1956; Romer, 1956, 1966; Steel, 1969;Thulborn, 1971, 1972; Galton, 1972; but see Maryanska,1977), having been fully corroborated by cladistic studies(Sereno, 1999; Butler, 2005; Butler et al., 2007, 2008). Tradi-tional ornithischian diagnostic traits such as the opisthopubicpelvis (Seeley, 1888) and the predentary bone (Marsh, 1894)can not be unambiguously considered apomorphic for thegroup, because their occurrence in the suggested basalmostornithischian, Pisanosaurus mertii, is equivocal (Butler et al.,2008). Yet, various other features have been accepted asdiagnostic for the clade, most of which correspond to modi-fications of the teeth and tooth-bearing bones, related to theacquisition of a more herbivorous diet. These include thepresence of a buccal emargination on the maxilla, into whichcheek musculature may have attached, and several changes inthe shape and arrangement of the teeth, as recently reviewedby Irmis et al. (2007b) and Butler et al. (2008).
Five main ornithischian groups are traditionallyrecognized: Stegosauria, Ankylosauria, Ornithopoda, Pachy-cephalosauria, and Ceratopsia (Thulborn, 1972), but the rela-tionships among these lineages remained ambiguous until thefirst cladistic analyses of the group were performed (Sereno,1984, 1986; Norman, 1984; Maryanska & Osmolska, 1985;Cooper, 1985). Among these, the view advocated by Sereno(1986, 1991b, 1999) was highly influential for nearly two
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 85
decades, during which few other phylogenetic studies focus-ing on the basal radiation of Ornithischia were performed.Sereno (1998, 1999) recognizes two main ornithischian splits:Thyreophora and Neornithischia (Table 1). The formerincludes Stegosauria and Ankylosauria, whereas Ceratop-sia and Pachycephalosauria compose Marginocephalia, thesister clade to Ornithopoda within Neornithischia. This gen-eral scheme is accepted by most recent studies (Xu et al.,2006; Butler et al., 2007, 2008), but details of basal formsnesting within each major group remain controversial.
Sereno (1986) combined neornithischians and thyreo-phorans within Genasauria, which encompasses the bulkof the Ornithischia. The first phylogenetic definition ofthe name (Table 1) employed the term Cerapoda in aconnotation equivalent to Neornithischia sensu Sereno (1999).Indeed, Cerapoda was later defined by Weishampel (2004)as ‘‘genasaurs more closely related to Triceratops thanto Ankylosaurus’’. The term was, however, more recentlyabandoned in favour of Neornithischia (Sereno, 1997,1998, 1999, 2005), or used in a radically different node-based concept (Buchholz, 2002; Barrett, Butler & Knoll,2005a; Butler et al., 2008). Originally, non-Genasauriaornithischians included only Lesothosaurus dignosticus, fromthe Early Jurassic Upper Elliot Formation, southern Africa(Thulborn, 1970, 1972; Santa Luca, 1984; Knoll & Battail,2001; Knoll, 2002a, b, c, 2005; Butler, 2005), and Pisanosaurus
mertii (see Section III.1). Despite its plesiomorphic postcranialanatomy, the latter taxon has for a long time been regardedas the sole unequivocal Triassic ornithischian, based on traitsof its partial skull and teeth (Sereno, 1991b; Butler et al., 2007;Irmis et al., 2007b). Historically, Pisanosaurus was related to‘‘fabrosaurids’’ (Thulborn, 1971, 1972), heterodontosaurids(Charig & Crompton, 1974; Bonaparte, 1976; Cooper,1985; Weishampel & Weishampel, 1983; Weishampel, 1984;Crompton & Attridge, 1986), and ‘‘hypsilophodontids’’(Galton, 1972, 1986; Colbert, 1981), but later acceptedas the most basal ornithischian (Novas, 1989; Weishampel& Witmer, 1990; Sereno, 1991b; Butler, 2005; Butler et al.,2007, 2008; but see Norman et al., 2004a). More recently, anon-Genasauria position was also inferred for the only othertwo ornithischian taxa with a Triassic record (Fig. 15A),Heterodontosauridae (Butler et al., 2008) and Eocursor parvus
(Butler et al., 2007). This is partially based on the retentionof several features otherwise atypical for the group suchas a long hand with extensor pits on the metacarpalsand phalanges, longer penultimate phalanges, and stronglyrecurved unguals with prominent flexor tubercles (Butleret al., 2007). Heterodontosaurids are more extensively knownfrom Early Jurassic strata (Fig. 15B), and were traditionallyplaced as the basalmost clade of Ornithopoda (Sereno, 1984,1986; Norman, Witmer & Weishampel, 2004b), althoughunorthodox alternative placements were also proposed inthe cladistic paradigm (Norman, 1984; Cooper, 1985;Maryanska & Osmolska, 1985; Buchholz, 2002; You, Xu& Wang, 2003; Xu et al., 2006). The group is minimallycomposed of Heterodontosaurus tucki, from the Early JurassicUpper Elliot (Santa Luca, 1980) and Clarens (Crompton &
Charig, 1962) formations of South Africa, and Abrictosaurus
consors, from the former unit (Thulborn, 1974). Other post-Triassic heterodontosaurids may include Lycorhinus angustidens
(Gow, 1975), also from the Upper Elliot Formation, unnamedforms from the Kayenta and Clarens formations (Irmis &Knoll, 2008), as well as younger records (Norman & Barrett,2002; Galton, 2005b; Zheng et al., 2009).
From the 1980s onwards, several putative ornithischians(e.g. Galtonia gibbidens, Technosaurus smalli, Revueltosaurus
callenderi, R. hunti, Lucianosaurus wildi, Pekinosaurus olseni,Tecovasaurus murryi, Protecovasaurus lucasi, Crosbysaurus harrisae)were reported from Late Triassic assemblages of NorthAmerica (Hunt, 1989; Hunt & Lucas, 1994; Hunt et al.,1998; Heckert, 2002, 2004) and Europe (Godefroit &Cuny, 1997; Cuny et al., 2000; Galton, 2005b) mostlybased on isolated teeth. Yet, recent studies demonstratedthat the trustworthy Triassic record of the group is muchmore restricted. Parker et al. (2005) reported the first non-dental material of R. callenderi, showing that it represents apseudosuchian rather than an ornithischian. Those authorsalso recognized notable convergences in the dental anatomyof ornithischians and some non-dinosaur archosaurs, e.g.low triangular tooth crowns with expanded base and carinaecomposed of large denticles. Indeed, in a comprehensivereview of the Triassic ornithischian record, Irmis et al. (2007b)reinterpreted most of those isolated teeth as indeterminatearchosauriforms. More recently, Ferigolo & Langer (2007)proposed the ornithischian affinity of the purported basaldinosauromorphs Silesaurus opolensis and Sacisaurus agudoensis
(Fig. 15A). This was partially based on the suggestedhomology of the synapomorphic ornithischian predentarybone to the edentulous tip of the lower jaw seen in bothtaxa, which is formed by independent ossifications in thelatter form. Yet, that proposition was not originally backedup by a numerical cladistic study, and further analysesfailed to recover that hypothesis of relationships (Langer &Benton, 2006; Ezcurra, 2006; Irmis et al., 2007a; Brusatteet al., 2008a).
Until the early eighties, most authors accepted the ‘‘fab-rosaurids’’ as a natural group of basal ornithischians, fre-quently depicted at the stem of either Ornithopoda or a moreinclusive group of non-thyreophoran taxa (Galton, 1978;Norman, 1984). Yet, it became increasingly evident that ‘‘fab-rosaurids’’ congregated a paraphyletic array of early, small-bodied forms (Weishampel & Witmer, 1990; Sereno, 1991b).In his phylogenetic studies, Sereno (1984, 1986, 1991b, seealso Buchholz, 2002; Xu et al., 2006) considered the archety-pal ‘‘fabrosaurid’’ Lesothosaurus dignosticus as the sister taxonto Genasauria. More recently, however, this view was chal-lenged by studies that placed that form within Genasauria(Butler et al., 2007), either as a neornithischian (Butler,2005) or a thyreophoran (Butler et al., 2008). In the lat-ter case, Lesothosaurus would represent the only thyreophoranto lack the typical cortical remodeling of cranial elementsand osteoderms covering the dorsum of the body (Butleret al., 2008). That genus was also reported from the Early
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

86 Max C. Langer and others
Fig. 15. Phylogenetic relationships and distribution of basal ornithischians. (A) Time-calibrated phylogeny depictingheterodontosaurids as the most basal clade of Ornithischia, based on Butler et al. (2007, 2008); dotted lines indicate uncertain positionof Silesaurus and Sacisaurus according to Ferigolo & Langer (2007). (B) Geographic occurrences of Late Triassic (black squares) andEarly Jurassic (white squares) taxa on a Late Triassic map redrawn from Blakey (2006). (C) More conventional phylogeny depictingheterodontosaurids as ornithopods, based on Sereno (1999); composition of Heterodontosauridae according to Butler et al. (2008).Black silhouettes (roughly at the same scale) adapted from various sources, names applied as in Table 1. In the cladograms, node-and stem -based taxa are respectively indicated by black circles and curved lines.
Jurassic La Quinta Formation, in western Venezuela (Rus-sell et al., 1992), but the specimens can only be referred toindeterminate non-cerapodan ornithischians (Barrett et al.,2008). Other taxa previously regarded as ‘‘fabrosaurids’’(Galton, 1978; Peng, 1997) have been considered of inde-terminate affinity or placed within genasaurian subgroups(Sereno, 1991b; Norman et al., 2004b, c; Butler, 2005).
Early Jurassic ornithischians other than Lesothosaurus dig-
nosticus have a less debated phylogenetic position (Fig. 15).
Scutellosaurus lawleri, from the Kayenta Formation of west-ern USA (Colbert, 1981; Rosenbaum & Padian, 2000),Emausaurus ernsti, from Mecklemberg, Germany (Haubold,1991), and the genus Scelidosaurus, known from the LowerLias of Dorset, England (Owen, 1861, 1863; Martill, Batten& Loydell, 2000; Norman, 2001) and possibly also from theKayenta Formation (Padian, 1989), are consensually con-sidered the basalmost thyreophorans (Sereno, 1999; Butleret al., 2008). Early Jurassic thyreophorans are otherwise only
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 87
known from the Lufeng Formation of China (Irmis & Knoll,2008). This includes the type specimens of Bienosaurus lufen-
gensis (Dong, 2001) and Tatisaurus oehleri (Norman, Butler &Maidment, 2007), both of which lack autapomorphic featuresand may represent nomina dubia (Irmis & Knoll, 2008). Exceptfor Lesothosaurus, heterodontosaurids, and thyreophorans,Stormbergia dangershoeki, from the Upper Elliot Formation (But-ler, 2005; but see Knoll et al., 2009), is the sole well knownEarly Jurassic ornithischian. Indeed, although much richerthan during Triassic times, Early Jurassic ornithischian fau-nas are still poorly diverse. Stormbergia dangershoeki seems torepresent the most basal Neornithischia, lacking typical fea-tures otherwise common to the group such as an elongatedprepubic process (Butler, 2005; Butler et al., 2007, 2008).
The divergence time of Ornithischia has been traditionallyconsidered as Late Triassic, given the presence of Pisanosaurus
mertii and various saurischian dinosaurs in South Americandeposits of that age. The phylogenetic hypothesis of Butleret al. (2007) better fits the stratigraphic data, restrictingthe Triassic record of ornithischians to non-Genasauriataxa. Instead, both traditional (Sereno, 1999) and moreunusual (Buchholz, 2002; Xu et al., 2006) arrangementsrequire the existence of long ghost-lineages for Ornithopoda,Marginocephalia, and Thyreophora. The possible positionof two South African forms at the base of both Thyreophoraand Neornithischia (Butler et al., 2008) suggests that the originand early diversification of ornithischians is to be foundin southwestern Gondwana (Rauhut & Lopez-Arbarello,2008), where the Late Triassic record of the group isconcealed (Fig. 15C). On the contrary, the bulk of EarlyJurassic thyreophorans occur in North Pangea, particularlyconsidering the uncertain affinity of the ankylosaur reportedfrom the Kota Formation of India (Nath, Yadagiri & Moitra,2002; Rauhut & Lopez-Arbarello, 2008).
The slightly more conspicuous Middle Jurassic record ofornithischians includes the first stegosaurs and ankylosaurswithin Thyreophora and an array of basal neornithischians.The Lower Shaximiao Formation of China (Peng et al., 2005)has yielded stegosaurs (Maidment & Wei, 2006) and neor-nithischians (Barrett et al., 2005a), while the former groupwas recorded along with a possible pachycephalosaur fromthe Balabansai Formation of Kirghizia (Averianov, Martin& Bakirov, 2005; Averianov, Bakirov & Martin, 2007). InEurope, Middle Jurassic ornithischians are represented bybasal ornithopods (Galton, 1980; Evans & Milner, 1994,Kriwet, Rauhut & Gloy, 1997; Ruiz-Omenaca, Suberbiola& Galton, 2005), stegosaurs (Galton & Powell, 1983, Galton,1990), and ankylosaurs (Galton, 1983a). Indeed, it was notuntil the Late Jurassic and Early Cretaceous that ornithis-chian faunas became abundant, with diverse thyreophorans,ornithopods, ceratopsians, and pachycephalosaurs, especiallyin Laurasian assemblages.
(2) Early sauropodomorph evolution
The sauropodomorph lineage includes dinosaurs with asmall skull, a distally broad humerus, a relatively shorthind limb, plus a series of other skeletal modifications
that support their monophyly in the cladistic paradigm(Sereno, 1999; Yates, 2003b; 2007a; Yates & Kitching, 2003;Langer & Benton, 2006; Upchurch et al., 2007). On thecontrary, as comprehensively reviewed by Sereno (2007a;see also Upchurch, 1997a; Galton & Upchurch, 2004),classical studies (Huene, 1929, 1956; Romer, 1956; Colbert,1964) frequently allocated the basal sauropodomorphsknown as ‘‘prosauropods’’ within a grade of saurischiansfrom which both theropods and sauropods arose. Thename Prosauropoda Huene, 1920, was coined in thatcontext, in reference to a group lying at the base of the‘‘herbivorous-omnivorous’’ branch of Pachypodosauria that,as the name implies, gave rise to sauropods. This includedThecodontosaurus, Plateosaurus, Sellosaurus, and Anchisaurus, butalso Poekilopleuron ‘‘as a blind side stem’’ (Huene, 1920). Onthe contrary, forms at some point regarded as members ofthe sauropodomorph lineage such as Paleosaurus (Benton et al.,2000) and Gresslyosaurus (Yates, 2007a) were allocated to thetheropod-related branch (Huene, 1920). As often mentioned(Benton, 1986b), this was in part due to the mistakenassociation of carnivorous teeth with other ‘‘prosauropod’’skeletal remains (Huene, 1932; Young, 1951). Charig et al.(1965; see also Galton, 1971, 1973) may be said to havesettled the current concept of ‘‘Prosauropoda’’, congregatingvarious early saurischians of the sauropodomorph lineage. Asproperly put by Sereno (2007a), ‘‘prosauropods’’ representedthe first grand radiation of dinosaurs sharing minimalmorphological coherence. Basal sauropodomorphs radiatedrelatively fast during Late Triassic times, becoming thedominant terrestrial herbivores/omnivores from Norian toEarly Jurassic landscapes (Upchurch, 1997a).
Sereno (2007a) selected from the universe of basalsauropodomorphs a subset of five taxa termed ‘‘core-prosauropods’’, which should carry the name Prosauropoda,if found to represent a monophylum exclusive of sauropods.A comparable application is seen in Yates & Kitching (2003)and Upchurch et al. (2007). However, the first phylogeneticdefinition of the name was proposed under the orthodoxy ofa monophyletic ‘‘Prosauropoda’’ as to include ‘‘Thecodon-tosauridae, Plateosauridae (Anchisauridae), Melanosauridae[sic], and all Sauropodomorpha closer to them than toSauropoda’’ (Upchurch, 1997a; Sereno, 2005). Based on thetype-genera of those family rank names, this definition is notapplicable to the current framework of sauropodomorph evo-lution, since a clade that includes Thecodontosaurus, Plateosaurus,Anchisaurus, and Melanorosaurus also includes sauropods. Con-sidering the criteria adopted here (Section II.2), that defi-nition has precedence over following ones that arbitrarilyselect either a single (Sereno, 1998; Galton & Upchurch,2004) or various ‘‘prosauropods’’ (Sereno, 2007a) as inter-nal specifiers. Indeed, the definition of Upchurch (1997a)is to be kept and applied under an eventually revitalizedframework of ‘‘prosauropod monophyly’’, while newly pro-posed names should be used to designate major subgroupsof Sauropodomorpha, as seen in Yates (2007a) and Smith &Pol (2007).
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

88 Max C. Langer and others
Early evolutionary studies of basal sauropodomorphs(Charig et al., 1965; Galton, 1971, 1976) broadly dis-criminate three main ‘‘prosauropod’’ groups: one includ-ing more gracile forms, the so-called ‘‘narrow-footedprosauropods’’, termed either Thecodontosauridae orAnchisauridae; a group of more ‘‘typical’’, Plateosaurus-relatedforms; and a group of bulky quadrupeds, frequently termedMelanorosauridae. The sauropod affinity of the latter groupwas often advocated (Colbert, 1964; Cooper, 1981; Bona-parte, 1986), hinting at ‘‘prosauropod’’ paraphyly. Indeed,the first cladistic approach to sauropodomorph evolutionreproduced that scheme (Gauthier, 1986), allocating The-
codontosaurus antiquus and Efraasia minor at the base of theclade, and the ‘‘broad-footed’’ forms, especially Riojasaurus
incertus, closer to Sauropoda. The succeeding cladistic studies(Sereno, 1989, 1999; Galton, 1989; Benton et al., 2000;Galton & Upchurch, 2004), however, failed to recovera similar paraphyletic array of ‘‘prosauropods’’. Instead,they pointed towards a different picture, in which most, ifnot all ‘‘prosauropods’’ represented the monophyletic sis-ter taxon to Sauropoda. This scheme is reminiscent ofpre-cladistic approaches (Charig et al., 1965; Cruickshank,1975; Van Heerden, 1978) that partially relied on the sup-posed irreversibility of some features of the ‘‘prosauropod’’foot, and on the uniqueness of their hand, in order todiscard their bearing on the origin of sauropods (but seeYates, 2003b; Sereno, 2007a). More recently, ‘‘prosauro-pod’’ monophyly was deemed an analytical artifact derivedfrom poor taxon sampling, that overlooked basal and near-sauropod sauropodomorphs (Yates, 2003b; see also Sereno,2007a). Indeed, most recent studies (Yates & Kitching, 2003;Pol, 2004; Smith & Pol, 2007; Yates, 2007a, b), includ-ing those performed by previous proposers of ‘‘prosauropodmonophyly’’ (Upchurch et al., 2007; Sereno, 2007a) tendto agree that at least some forms previously assigned to‘‘Prosauropoda’’ are basal to the bulk of sauropodomorphs,and that others are closely related to the sauropod radiation(Fig. 16A). Evidently, these hypotheses are not fully congru-ent with one another, but important common points areseen, as outlined below.
The most recently proposed basal sauropodomorphphylogenies (Pol, 2004; Yates, 2007a, b; Upchurch et al.,2007) agree that the Late Triassic Saturnalia tupiniquim,Pantydraco caducus, Thecodontosaurus antiquus, and Efraasia minor
are amongst the most basal members of the lineage, whereasPanphagia protos may be the basal-most member (Martinez& Alcober, 2009). Upchurch et al. (2007) also includedMussaurus patagonicus in that basal grade, but the taxonwas given a more derived position by Pol (2004), Pol &Powell (2005, 2007b), and Sereno (2007a) based on first-handexamination of a more complete set of specimens. Such abasal position was also inferred, in an admittedly tentativefashion, to the newly described Pradhania gracilis from theEarly Jurassic Upper Dharmaram Formation, India (Kuttyet al., 2007). The relative positions of the other forms arenearly consensual, with Saturnalia basal to Thecodontosaurus
and Pantydraco (but see Galton & Upchurch, 2004), and
Efraasia closer to other sauropodomorphs (Fig. 16A). Thesetaxa represent the first radiation of the sauropodomorphlineage, retaining various morphological traits of theirbasal saurischian/basal dinosauriform precursors (Barrett& Upchurch, 2007) such as the smaller size (adults are lessthan 4 m in length) and, at least facultative, bipedality. Othersimplesiomorphies include a relatively long hand, a partiallyclosed acetabulum, a distal femur lacking a well-developedextensor depression, metatarsals I and II closely appressed,plus several other skeletal features (Yates, 2003b; Yates &Kitching, 2003; Smith & Pol, 2007; Upchurch et al., 2007).In addition, some of these forms possess novel herbivorousadaptations such as a higher number of coarsely denticulatedteeth, although they might have retained an omnivorousdiet (Barrett, 2000). The geographic distribution of thesebasal sauropodomorphs, along with that of slightly morederived forms (Pol, 2004; Yates, 2007a), suggests an initialradiation of the clade restricted to western Pangea (Fig. 16C).Considering the node-based definition of Sauropodomorphaand the (by typification) inclusion of Thecodontosaurus as aninternal specifier of Prosauropoda, Langer (2002) proposedthat Saturnalia should be excluded from Sauropodomorpha,and considered instead as a taxon on its stem lineage.
The relationships of sauropodomorphs more derived thanEfraasia minor are far from consensual. In fact, severalpossible arrangements recently have been proposed (seevarious articles in Barrett & Batten, 2007; especially Sereno,2007a). In most of them, however, a relatively stable setof taxa is placed closely related to traditional sauropods(Fig. 16A), minimally including Norian-Rhaetian forms suchas Camelotia borealis, Melanorosaurus readi, Blikanasaurus cromptoni,and Lessemsaurus sauropoides, as well as Antetonitrus ingenipes,which was already first described as a sauropod. This roughlycorresponds to the classical radiation of ‘‘melanorosaurids’’,composed of usually larger (6.5-10 m), more herbivorous‘‘prosauropods’’ (Barrett, 2000). Most of these may haveadopted a fully quadrupedal gait (Yates & Kitching, 2003),although facultative bipedality is still suggested for severalforms (Barrett & Upchurch, 2007). These taxa share arange of traits with eusauropods, including short and highdorsal centra, an increased number of sacral vertebrae, alonger manual digit I with a straighter ungual, broader non-terminal manual phalanges, and a straighter femur, ellipticalin cross section and bearing distally displaced lesser andfourth trochanters (Yates & Kitching, 2003; Yates, 2007a,Upchurch et al., 2007; Pol & Powell, 2007a). Some authors(Yates & Kitching, 2003; Smith & Pol, 2007; Yates, 2007a, b)include all or some of these forms within Sauropoda, given thestem-based definition of the taxon as to include forms closerto Saltasaurus loricatus than to the archetypal ‘‘prosauropod’’Plateosaurus engelhardti (Wilson & Sereno, 1998, Sereno, 1999;Upchurch et al., 2007), or alternative arbitrary attemptsto mimic more traditional/current placement of formseither within or outside the group (Yates, 2007a; Sereno,2007a). However, Sauropoda was first phylogeneticallydefined by Salgado et al. (1997) in a node-based fashion(Table 1) that may exclude forms regularly assigned to the
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society
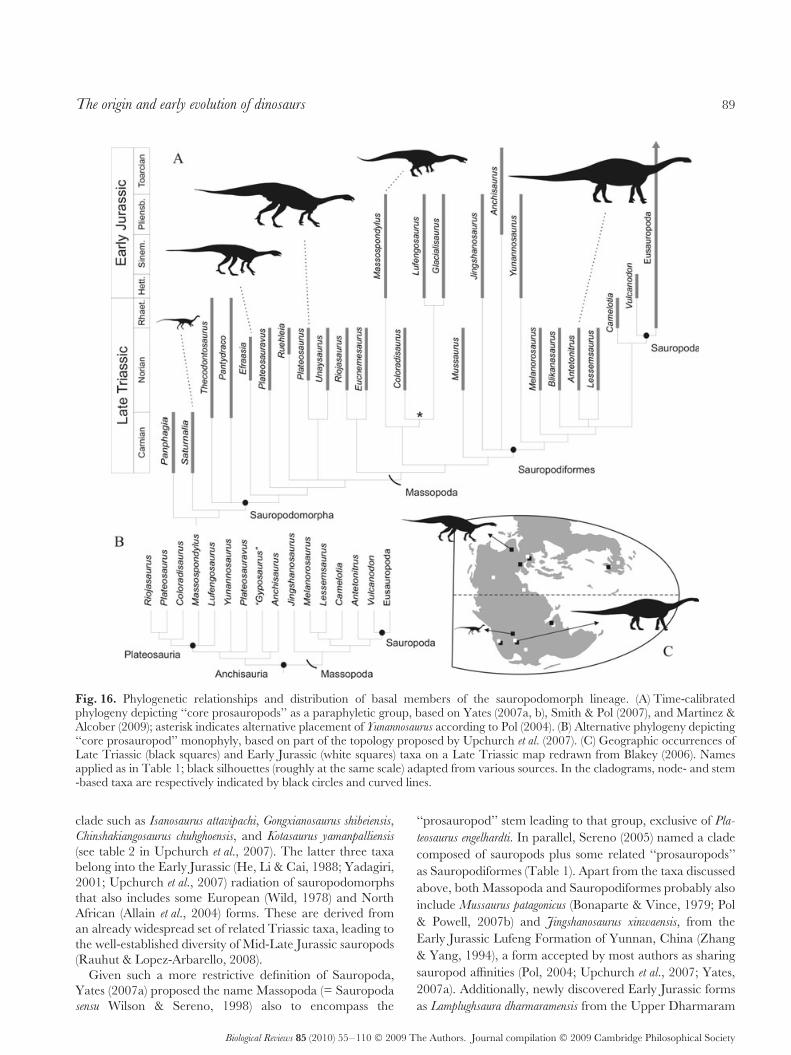
The origin and early evolution of dinosaurs 89
Fig. 16. Phylogenetic relationships and distribution of basal members of the sauropodomorph lineage. (A) Time-calibratedphylogeny depicting ‘‘core prosauropods’’ as a paraphyletic group, based on Yates (2007a, b), Smith & Pol (2007), and Martinez &Alcober (2009); asterisk indicates alternative placement of Yunannosaurus according to Pol (2004). (B) Alternative phylogeny depicting‘‘core prosauropod’’ monophyly, based on part of the topology proposed by Upchurch et al. (2007). (C) Geographic occurrences ofLate Triassic (black squares) and Early Jurassic (white squares) taxa on a Late Triassic map redrawn from Blakey (2006). Namesapplied as in Table 1; black silhouettes (roughly at the same scale) adapted from various sources. In the cladograms, node- and stem-based taxa are respectively indicated by black circles and curved lines.
clade such as Isanosaurus attavipachi, Gongxianosaurus shibeiensis,Chinshakiangosaurus chuhghoensis, and Kotasaurus yamanpalliensis
(see table 2 in Upchurch et al., 2007). The latter three taxabelong into the Early Jurassic (He, Li & Cai, 1988; Yadagiri,2001; Upchurch et al., 2007) radiation of sauropodomorphsthat also includes some European (Wild, 1978) and NorthAfrican (Allain et al., 2004) forms. These are derived froman already widespread set of related Triassic taxa, leading tothe well-established diversity of Mid-Late Jurassic sauropods(Rauhut & Lopez-Arbarello, 2008).
Given such a more restrictive definition of Sauropoda,Yates (2007a) proposed the name Massopoda (= Sauropodasensu Wilson & Sereno, 1998) also to encompass the
‘‘prosauropod’’ stem leading to that group, exclusive of Pla-
teosaurus engelhardti. In parallel, Sereno (2005) named a cladecomposed of sauropods plus some related ‘‘prosauropods’’as Sauropodiformes (Table 1). Apart from the taxa discussedabove, both Massopoda and Sauropodiformes probably alsoinclude Mussaurus patagonicus (Bonaparte & Vince, 1979; Pol& Powell, 2007b) and Jingshanosaurus xinwaensis, from theEarly Jurassic Lufeng Formation of Yunnan, China (Zhang& Yang, 1994), a form accepted by most authors as sharingsauropod affinities (Pol, 2004; Upchurch et al., 2007; Yates,2007a). Additionally, newly discovered Early Jurassic formsas Lamplughsaura dharmaramensis from the Upper Dharmaram
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

90 Max C. Langer and others
Formation (Kutty et al., 2007), and an undescribed Argen-tinean taxon (Pol & Garrido, 2007) may also belong to thesauropod stem. Other Early Jurassic forms such as Yunan-
nosaurus huangi, also from the Lufeng Formation (Young,1942), and the ‘‘prosauropod’’ from the Portland Forma-tion of eastern North America (Anchisaurus polyzelus sensu
Yates, 2004; Ammosaurus major sensu Sereno, 2007a), havehighly controversial affinities, and two antipodal scenarioshave been presented (but see Pol, 2004). They may lineup with other sauropod-related forms (Yates, 2007a, b) orbelong to a clade of typical ‘‘prosauropods’’ (Upchurch et al.,2007). There is limited evidence in favour of a monophyleticgroup of ‘‘core prosauropods’’ (Sereno, 2007a), minimallyencompassing Plateosaurus engelhardti, Riojasaurus incertus, Mas-
sospondylus carinatus, Lufengosaurus huenei, and Coloradisaurus brevis
(Fig. 16B). Potential apomorphic traits of this clade includemodifications in the neurovascular foramina of the maxilla,the manus (carpal II does not completely cover the proxi-mal end of metacarpal II, metacarpal V bears an expandedproximal end divided into two articular surfaces, first pha-lanx of manual digit I twisted by at least 60◦), and foot(metatarsal IV with expanded proximal end). On the con-trary, Pol (2004) and Yates (2007a, b) split that group intoa successive array of three to five lineages on the stem toSauropodiformes sensu Sereno (2007a), where forms relatedto Plateosaurus and Riojasaurus are considered more basal(Fig. 16A). Depending on the position of these forms relativeto other sauropodomorphs is the application of names suchas Plateosauria and Anchisauria, e.g. compare Yates (2007a)and Upchurch et al. (2007).
Either as a clade or grade, ‘‘core prosauropods’’ mayrepresent a biological unit, playing similar roles in the LateTriassic-Early Jurassic ecosystems in which they occurred.Barrett & Upchurch (2007) reviewed the palaeobiology ofthese forms, highlighting some of their ecological adaptations.These include larger size (2.5-10 m) compared to more basalsauropodomorphs, and greater reliance on an herbivorousdiet, although facultative omnivory was not discarded. ‘‘Coreprosauropods’’ were also capable of bipedal locomotion(Cooper, 1981; Christian & Preuschoft, 1996; Bonnan& Senter, 2007), although larger forms were probablyobligatory quadrupeds. This is the case for Riojasaurus incertus,previously connected to the origin of sauropods (Bonaparte,1972; Gauthier, 1986). As a whole, that clade/gradecongregates a relatively high diversity of Late Triassic forms,including Ruehleia bedheimensis and the species of Plateosaurus
in Europe/Greenland, Riojasaurus incertus and Unaysaurus
tolentinoi in South America, as well as Eucnemesaurus fortis
and Plateosauravus cullingworthi in South Africa. Other ‘‘core-prosauropods’’ may fit into Massospondylidae, a clade thatincludes the Triassic Coloradisaurus brevis (Yates & Kitching,2003), the Early Jurassic Glacialisaurus hammeri (Smith & Pol,2007) from the Hanson Formation, Antarctica, and a possibleset of Chinese forms (see Pol, 2004), minimally includingLufengosaurus huenei from the Lufeng Formation (Barrett,Upchurch & Xiao-Lin, 2005b). Its type genus Massospondylus
(M . carinatus) is known from the Upper Elliot Formation
(Cooper, 1981; Gow, Kitching & Raath, 1990; Sues et al.,2004, Reisz et al., 2005) and other stratigraphic units insouthern Africa (Cooper, 1981; Galton & Upchurch, 2004),but not in North America (Attridge, Crompton & Jenkins,1985; Sues et al., 2004). More recently, its sister taxon,Adeopapposaurus mognai, was described from the Lower JurassicCanon del Colorado Formation, Argentina (Martinez, 2009).
(3) Early theropod evolution
The name Theropoda was coined by Marsh (1881) asa new suborder of carnivorous dinosaurs, but its statusas a ‘‘natural group’’ was rejected for the first half ofthe last century. At the time, gracile members of thegroup, the so-called ‘‘coelurosaurs’’, were considered, alongwith ‘‘prosauropods’’, the ‘‘basal stock’’ from which bothsauropods and derived theropods evolved (Huene, 1914,1920, 1932; Romer, 1956). Theropod monophyly was hintedat by Matthew & Brown (1922), firmly established by Col-bert (1964), in an arrangement widely accepted since (Chariget al., 1965; Romer, 1966; Colbert & Russell, 1969; Ostrom,1978; Steel, 1970; Bakker & Galton, 1974), and corrobo-rated by pioneering phylogenetic studies (Thulborn, 1984;Gauthier, 1986; Novas, 1992b; Holtz, 1994; Sereno, 1999).Indeed, taxa consensually assigned to the group share a seriesof typical traits, e.g. promaxillary foramen; well-developedpneumatization in cervical and cranial trunk vertebrae;manus with reduced metacarpal I, slender metacarpal III,and reduced/absent digits IV and V; ilium with promi-nent supracetabular crest and preacetabular ala; tibia withmarked cnemial and fibular crests; transversely compressedcalcaneum; foot with reduced outer digits (Rauhut, 2003;Ezcurra & Cuny, 2007; Ezcurra & Novas, 2007a).
As discussed in Section III.2, the most contentiousaspect of early theropod evolution is the possible nestingof various Triassic forms within the group (Fig. 9). Thisis particularly the case fot Eoraptor lunensis (Sereno et al.,1993) and herrerasaurs (Sereno & Novas, 1992), but alsofor other taxa such as Guaibasaurus candelariensis (Langer et al.,2007a), Agnosphitys cromhallensis, and Chindesaurus bryansmalli
(Yates, 2007a). Indeed, these forms apart, the Noriancoelophysoid ‘‘Camposaurus arizonensis’’ represents the oldesttheropod (Hunt et al., 1998; Nesbitt et al., 2007), which wouldmake Theropoda the only major dinosaur lineage lacking awell-defined Ischigualastian record. Moreover, based on thecurrent knowledge of theropod diversity, and not consideringthe above-mentioned taxa as members of the group, thestem-based Theropoda would be as inclusive as the node-based Neotheropoda (Table 1). This name was first usedby Bakker (1986) to combine theropods more derived than‘‘podokesaurids’’, but phylogenetically defined by Sereno(1998) as a more inclusive group. Yet, Neotheropoda remainsa useful name under alternative arrangements (Fig. 9) and/orif new forms are found to belong to its stem (see Nesbitt &Chatterjee, 2008; Martinez et al., 2008). Although a lessinclusive Neotheropoda (Padian et al., 1999; Wilson et al.,2003) seems more useful in the current orthodoxy, and moreproperly translates the original meaning of the name (Bakker,
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 91
1986), the definition proposed by Sereno (1998) has historical‘‘priority’’.
Another controversial aspect of early theropod evolution isthe possible monophyly of the oldest neotheropods, groupedwithin Ceratosauria and/or Coelophysoidea. Indeed, the firstcladistic analyses of theropod relationships identified twomain neotheropod lineages, Ceratosauria and Tetanurae(Gauthier, 1986; Rowe, 1989; Rowe & Gauthier, 1990).In turn, Ceratosauria was divided into two branches, theLate Triassic-Early Jurassic Coelophysoidea (i.e. Coelophysis,Dilophosaurus, and their kin) and the Jurassic-CretaceousNeoceratosauria, including Ceratosaurus and abelisauroids(Novas, 1992a; Sereno, 1997, 1999; Holtz, 2000; Coria &Salgado, 2000). This arrangement was accepted during mostof the 1990s, but the vast majority of more recent studiesconsider neoceratosaurs more closely related to tetanuransthan to coelophysoids, challenging the monophyly ofthe traditional Ceratosauria (Carrano, Sampson & Forster,2002; Carrano, Hutchinson & Sampson, 2005; Rauhut,2003; Sereno, Wilson & Conrad, 2004; Yates, 2005; Ezcurra,2006; Ezcurra & Novas, 2007a; Ezcurra & Cuny, 2007; Smithet al., 2007; Carrano & Sampson, 2008; but see Tykoski &Rowe, 2004; Tykoski, 2005; Allain et al., 2007). In fact,Ceratosauria was node-based defined by Rowe & Gauthier(1990) as ‘‘including Ceratosaurus nasicornis, Dilophosaurus
wetherilli, Liliensternus liliensterni, Coelophysis bauri, Syntarsus
rhodesiensis, Syntarsus kayentakatae, Segisaurus halli, Sarcosaurus
woodi, and all other taxa stemming from their most recentcommon ancestor’’, based on a phylogenetic frameworkin which these forms compose a monophylum exclusiveof tetanurans. On the contrary, in the current orthodoxy(Fig. 17), Ceratosauria would point to the same node asNeotheropoda, circumscribing a much more inclusive group,and is not employed here. Instead, the term AverostraPaul, 2002, as phylogenetically defined by Ezcurra & Cuny(2007), designates the clade composed of Tetanurae plusNeoceratosauria (Table 1).
Early phylogenetic studies grouped all Triassic andEarly Jurassic neotheropods within the Coelophysoideaclade. This included Dilophosaurus wetherilli, as the sistertaxon to Liliensternus liliensterni plus Coelophysidae (Rowe& Gauthier, 1990; Holtz, 1994). The latter group isminimally composed of Late Triassic (Colbert, 1989; Rauhut& Hungerbuhler, 2000) and Early Jurassic (Raath, 1969;Rowe, 1989) Coelophysis-‘‘Syntarsus’’-related forms (Paul, 1988;Bristowe & Raath, 2004), but may also include Segisaurus
halli and Procompsognathus triassicus (Sereno, 1997, 1999;Tykoski & Rowe, 2004; Knoll, 2008). With the additionof newly described forms, such as Zupaysaurus rougieri andLophostropheus airelensis, various cladisitic analyses (Tykoski &Rowe, 2004; Carrano et al., 2005; Tykoski, 2005; Ezcurra,2006; Ezcurra & Novas, 2007a; Ezcurra & Cuny, 2007)also recovered a monophyletic Coelophysoidea sensu lato
(Fig. 17B). As such, the group can be diagnosed by severalcraniomandibular features such as a flexible articulationbetween premaxilla and maxilla marked by a prominentsubnarial diastema, an expanded rostral end of the dentary,
reduced serrations on premaxillary teeth, and enlarged (fang-like) teeth in the rostral portion of the dentary, as well asby a peculiar pattern of femoral dimorphism (Tykoski &Rowe, 2004; Ezcurra & Novas, 2007a, Ezcurra & Cuny,2007). However, the monophyletic status of that groupwas questioned by Rauhut (2003), who found Dilophosaurus
more closely related to tetanurans and neoceratosaurs thanto coelophysids. Along with other forms, those taxa sharecranial details such as a lacrimal fenestra, a dorsoventrallyelongated orbit, and a reduced tooth count occupyinga shorted portion of the maxilla, modifications in theretroarticular process of the lower jaw, and a higher astragalarascending process (Smith et al., 2007). A similar hypothesis ofrelationship was advocated by Yates (2005), which consideredDracovenator regenti as closely related to Dilophosaurus wetherilli.Smith et al. (2007) also found other Early Jurassic taxasuch as Cryolophosaurus ellioti and ‘‘Dilophosaurus’’ sinensis
as members of such a ‘‘Dilophosaurus clade’’ (Fig. 17A),partially characterized by the presence and/or differentialconfiguration of the dorsal crests of the skull. Yates (2005)also assigned Zupaysaurus to that clade, but the phylogeneticposition of that taxon is highly controversial within non-averostran neotheropods (Carrano et al., 2005; Smith et al.,2007; Ezcurra & Novas, 2007a). Clearly, as stem-based-defined by Sereno (1998), the inclusivity of Coelophysoideais dependent on the adopted evolutionary framework. Itmay include only small to medium-sized forms similarto Coelophysis-‘‘Syntarsus’’ (Fig. 17A), or also congregate anEarly Jurassic radiation of large-bodied Dilophosaurus-liketaxa (Fig. 17B).
Neotheropods experienced a rapid diversification duringthe Norian-Rhaetian and Early Jurassic, achieving a broaddistribution over west Pangea (Fig. 17C). This early radiationwas mostly represented by sensu lato ‘‘coelophysoids’’, i.e.non-averostran neotheropods. Their Norian representativesinclude small to medium-sized forms reported from westernUSA (Colbert, 1989; Carpenter, 1997; Nesbitt et al., 2007)and Europe (Sereno & Wild, 1992; Rauhut & Hungerbuhler,2000; Allen, 2004), most of which are also sensu sticto
Coelophysoidea (Fig. 17B), and perhaps also larger formssuch as Zupaysaurus rougieri (Arcucci & Coria, 2003).Close to the Triassic-Jurassic boundary, theropod remainsbecome scarce and basically only Lophostropheus airelensis
is known (Ezcurra & Cuny, 2007; but see Dzik et al.,2008). Tetanurans previously have been reported from LateTriassic outcrops, but these records are not conclusive. Thesupposed bird Protoavis texensis (Chatterjee, 1991) has beenrecently reinterpreted as a chimaera (Nesbitt et al., 2007),with some elements of ‘‘coelophysoid’’ affinities, but notof tetanuran nature. Otherwise, ‘‘Zanclodon’’ cambrensis wasassigned to Tetanurae (Holtz, Molnar & Currie, 2004),but the available material does not differ from those ofnon-averostran theropods.
During the Early Jurassic (Fig. 17C), sensu sticto coelo-physoids continue to be well represented in western USA,including Segisaurus halli from the Navajo Sandstone (Camp,1936; Carrano et al., 2005) and ‘‘Syntarsus’’ kayentakatae from
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society
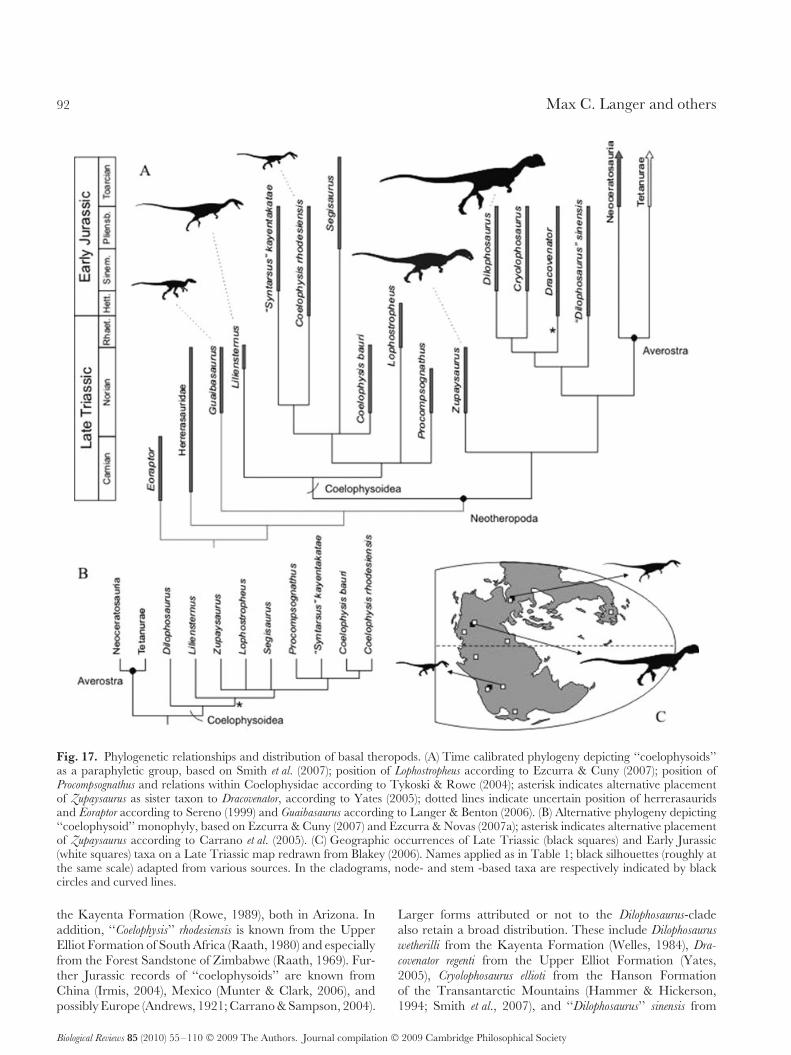
92 Max C. Langer and others
Fig. 17. Phylogenetic relationships and distribution of basal theropods. (A) Time calibrated phylogeny depicting ‘‘coelophysoids’’as a paraphyletic group, based on Smith et al. (2007); position of Lophostropheus according to Ezcurra & Cuny (2007); position ofProcompsognathus and relations within Coelophysidae according to Tykoski & Rowe (2004); asterisk indicates alternative placementof Zupaysaurus as sister taxon to Dracovenator, according to Yates (2005); dotted lines indicate uncertain position of herrerasauridsand Eoraptor according to Sereno (1999) and Guaibasaurus according to Langer & Benton (2006). (B) Alternative phylogeny depicting‘‘coelophysoid’’ monophyly, based on Ezcurra & Cuny (2007) and Ezcurra & Novas (2007a); asterisk indicates alternative placementof Zupaysaurus according to Carrano et al. (2005). (C) Geographic occurrences of Late Triassic (black squares) and Early Jurassic(white squares) taxa on a Late Triassic map redrawn from Blakey (2006). Names applied as in Table 1; black silhouettes (roughly atthe same scale) adapted from various sources. In the cladograms, node- and stem -based taxa are respectively indicated by blackcircles and curved lines.
the Kayenta Formation (Rowe, 1989), both in Arizona. Inaddition, ‘‘Coelophysis’’ rhodesiensis is known from the UpperElliot Formation of South Africa (Raath, 1980) and especiallyfrom the Forest Sandstone of Zimbabwe (Raath, 1969). Fur-ther Jurassic records of ‘‘coelophysoids’’ are known fromChina (Irmis, 2004), Mexico (Munter & Clark, 2006), andpossibly Europe (Andrews, 1921; Carrano & Sampson, 2004).
Larger forms attributed or not to the Dilophosaurus-cladealso retain a broad distribution. These include Dilophosaurus
wetherilli from the Kayenta Formation (Welles, 1984), Dra-
covenator regenti from the Upper Elliot Formation (Yates,2005), Cryolophosaurus ellioti from the Hanson Formationof the Transantarctic Mountains (Hammer & Hickerson,1994; Smith et al., 2007), and ‘‘Dilophosaurus’’ sinensis from
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 93
the Lufeng Formation of China (Hu, 1993). In addition,the oldest averostrans are known from Early Jurassic assem-blages. This includes Berberosaurus liassicus, a neoceratosaurfrom Morocco (Allain et al. 2007; but see Xu et al., 2009),and the very dubious record of a therizinosauroid jaw in theLufeng Formation (Zhao & Xu, 1998; Xu, Zhao & Clark,2001; Rauhut, 2003). In any case, as sister taxon to Neo-ceratosauria, a tetanuran ghost lineage might be inferred forthe latest Early Jurassic (Fig. 17A). Further Early Jurassictheropods were recorded from the La Quinta Formation ofVenezuela (Moody, 1997) based on teeth that can not beallocated in a less inclusive clade.
Middle Jurassic dinosaur-bearing assemblages are rare(Rauhut & Lopez-Arbarello, 2008), but the available datashow that the composition of theropod faunas changeddrastically relative to those of Late Triassic and EarlyJurassic age. ‘‘Coelophysoids’’ disappear completely from thefossil record, and tetanurans became the dominant forms.Apart from indeterminate theropod remains from NorthAfrica (Monbaron, Russell & Taquet, 1999) and Madagascar(Flynn et al., 2006), Middle Jurassic forms include a probablebasal neoceratosaur from Australia (Long & Molnar, 1998;Rauhut, 2005a), and basal tetanurans form Argentina(Rauhut, 2005a), Europe, and China (Holtz et al., 2004;Smith et al., 2007). On the other hand, neoceratosaurs arebetter known from Late Jurassic and especially Cretaceousdeposits (Carrano & Sampson, 2008; Xu et al., 2009).
VI. CONCLUSIONS
(1) The oldest unequivocal records of Dinosauria are ofLate Triassic age (approximately 230 Mya). These wereunearthed from rocks accumulated over extensional riftbasins in Argentina, Brazil, Zimbabwe, and India. Thebetter known early dinosaurs are Herrerasaurus ischigualastensis,Pisanosaurus mertii, Eoraptor lunensis, and Panphagia protos fromthe Ischigualasto Formation, northwestern Argentina, andStaurikosaurus pricei and Saturnalia tupiniquim from the SantaMaria Formation, south Brazil. Other dinosaur records ofequivalent age are either more fragmentary or of dubiousaffinities, hinting at a south Pangea origin of the group. Nouncontroversial dinosaur body fossils are known from olderstrata, but a possible Middle Triassic origin of the lineagemay be inferred from both the footprint record and its sistergroup relation to Ladinian basal dinosauromorphs.
(2) Dinosauria is by definition a monophyletic group that,in the present orthodoxy, combines saurischians and ornithis-chians to the exclusion of other major archosaur groups suchas pterosaurs, phytosaurs, and crocodylomorphs. The firstphylogenetic definition to fit the current understanding ofDinosauria as a node-based taxon solely composed of mutu-ally exclusive Saurischia and Ornithischia was given as ‘‘alldescendants of the most recent common ancestor of birdsand Triceratops’’. This definition should be followed untilmore specific provisions are given by the PhyloCode.
(3) Dinosaurs are nested within the bird-line of archosaursalong with pterosaurs (possibly), Scleromochlus taylori, and basalDinosauromorpha, the phylogeny of which is in state offlux. It includes the archetypal Marasuchus lilloensis, a diver-sity of more basal forms such as Lagerpeton and Dromomeron,as well as silesaurids: a possibly monophyletic group thatcombines Mid-Late Triassic basal dinosauromorphs thatmay represent sister taxa to Dinosauria. Recent cladisticanalyses of basal dinosaur relationships agree in variouskey points: (1) Pisanosaurus mertii is a basal ornithischian;(2) Herrerasaurus ischigualastensis and Staurikosaurus pricei belongin a monophyletic Herrerasauridae; (3) Guaibasaurus candelar-
iensis, Eoraptor lunensis, and herrerasaurids are saurischians;(4) Saurischia includes two main groups, Sauropodomor-pha and Theropoda; and (5) Saturnalia tupiniquim is a basalmember of the sauropodomorph lineage, a position alsoinferred for the recently described Panphagia protos. On thecontrary, several aspects of basal dinosaur phylogeny remaincontroversial, including the position of silesaurids as basalornithischians or non-dinosaur dinosauromorphs; the posi-tion of herrerasaurids, Eoraptor, and Guaibasaurus as basaltheropods or basal saurischians; and the affinity and/or valid-ity of various fragmentary taxa such as Agnosphitys cromhallensis,Alwalkeria maleriensis, Chindesaurus bryansmalli, Saltopus elginensis,Spondylosoma absconditum, and Teyuwasu barberenai.
(4) The identification of dinosaur apomorphies ishampered by the incompleteness of the skeletal remainsattributed to most basal dinosauromorphs, the skull and forelimb of which are particularly poorly known. Nonetheless,to the exclusion of silesaurids, Dinosauria can be diagnosedby a suite of derived traits, namely: foramen-sized post-temporal fenestra; epipophyses on cranial cervical vertebra;long deltopectoral crest; open acetabulum; arched dorsaliliac margin; femoral head inturned and distinctly offset fromthe shaft; asymmetrical fourth trochanter; astragalus withacute anteromedial corner, broad ascending process, andreduced fibular articulation; and proximally flat lateral distaltarsal. On the contrary, long-standing dinosaur apomorphiessuch as the absence of a postfrontal, the presence of morethan two sacral vertebrae, reduced manual digits IV andV, modified ‘‘lesser trochanter’’, and metatarsals II and IVsubequal in length do not unambiguously diagnose the group.The prevalence of dinosaur diagnostic traits related to thepelvic girdle and limb may reflect the better preservationof these structures in their sister taxa, but may also suggestthat these anatomical parts suffered most of the changes seenin the early dinosaur skeleton. Some of these traits can berelated to the acquisition of an erect bipedal gait, which hastraditionally been suggested to represent a key adaptationthat allowed, or even promoted, dinosaur radiation in theLate Triassic.
(5) Contrary to the classical ‘‘competitive’’ model,dinosaurs did not gradually replace other terrestrial tetrapodsover the Late Triassic. Yet, opportunistic and competitivescenarios are not mutually exclusive, and species interactionmay have played a partial role in the rise of dinosaurs,which can be said to have consisted of three landmark
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

94 Max C. Langer and others
moments, separated by controversial (Carnian-Norian,Triassic-Jurassic) extinction events. The Carnian earlydiversification did not occur in an empty ecospace but despitethe abundance and diversity of contemporary tetrapods.The Norian increase in dominance might be connected toclimatic/floristic changes and to the extinction of herbivorousforms such as rhynchosaurs, but the timing of these eventsneeds further investigation. The subtle Jurassic diversificationseems to have occurred in the aftermath of an extinctionevent, and might be an example of opportunistic radiationinto released ecospace, with the origin of neornithischiansand thyreophorans, the heterodontosaurid diversity peak,the rise of the Dilophosaurus-clade, and further acquisition oftypical graviportal traits among sauropods.
(6) It is traditionally believed that dinosaurs arose frombipedal and carnivorous forms, but evidence gatheredfrom newly discovered basal dinosauromorphs indicatethat quadrupedalism and omnivory/herbivory can not bediscarded as possible ancestral traits of the group. At themoment, however, most evidence points towards a fullybipedal origin of dinosaurs. On the contrary, dependingon the accepted hypothesis of relationships, the ancestraldinosaur diet can be reconstructed as either carnivorousor omnivorous. In any case, the plesiomorphic toothmorphology of dinosaurs does not strictly compare to thetypical carnivorous or omnivorous/herbivorous patterns ofmore derived members of the group. Indeed, each majordinosaur group seems to have independently acquired itstypical set of dental traits.
(7) The phylogenetic relationships of the basal membersof each major dinosaur group have recently been reeval-uated in the light of new evidence. Among ornithischians,unorthodox placements inside and outside Genasauria wereproposed for Lesothosaurus diagnosticus and Heterodontosauri-dae, respectively. Within saurischians, both ‘‘Prosauropoda’’and ‘‘Ceratosauria’’ were regarded as paraphyletic in theirbroader understanding. Yet, ‘‘core prosauropods’’ and coelo-physoids may still represent smaller clades at the base ofSauropodomorpha and Theropoda, respectively.
(8) Whereas the oldest dinosaurs were geographicallyrestricted to south Pangea, including rare ornithischiansand more abundant basal members of the saurischianlineage, the group achieved a nearly global distributionby Norian/Rhaetian times, especially with the radiationof saurischian groups such as ‘‘prosauropods’’ and coelo-physoids. This suggests an ‘‘out of south Pangea’’ model ofdinosaur radiation, but no model is better than the evidenceupon which it stands. In this case, the evidence is restrictedto a handful of fossils from scattered areas around the world,and more prospection work is needed in order to build a morereliable scenario of dinosaur origins and basal radiation.
VII. ACKNOWLEDGEMENTS
We thank the following for permission to examine specimensmentioned in this paper and help during this work: Angela
Milner and Sandra Chapman (BMNH); Rainer Schoch andRupert Wild (SMNS); Ricardo Martinez and Oscar Alcober(UNSJ); Dave Unwin (MB); Pat Holroyd, Randy Irmis,and Kevin Padian (UCMP); Michael Maisch (GPIT); JaimePowell and Judith Babot (PVL); Mike Raath (QVM); JerzyDzik and Tomasz Sulej (ZPAL); Maria Claudia Malabarba(MCP); Jorge Ferigolo and Ana Maria Ribeiro (FZB/RS);and Emilio Vaccari (PULR). Aspects of this work were fundedby grants from the Brazilian agencies CNPq, CAPES, andFAPESP (to M.C.L. and J.S.B.), Samuel Welles Fund–UCBerkeley (to M.D.E.), Agencia Nacional de PromocionCientıfica y Tecnica (to F.E.N.), and Santander Bank (toJ.S.B.). Comments by Mike Benton and an anonymousreviewer were much appreciated. This is contribution No.15 of Laboratorio de Paleontologia, FFCLRP-USP.
VIII. APPENDIX 1. INSTITUTIONALABBREVIATIONS
BMNH, Natural History Museum, London, UK; GPIT,Institut fur Geologie und Palaontologie, Tubingen, Ger-many; MB, Humboldt Museum fur Naturkunde, Berlin,Germany; MCP, Museu de Ciencias e Tecnologia, PUCRS,Porto Alegre, Brazil; PULR, Museo de Ciencias Naturales,Universidad Nacional de La Rioja, La Rioja, Argentina;PVL, Fundacion ‘‘Miguel Lillo’’, San Miguel de Tucuman,Argentina; PVSJ, Museo de Ciencias Naturales, UniversidadNacional de San Juan, San Juan Argentina; QVM, NationalMuseum of Natural History, Harare, Zimbabwe; SMNS,Staatliches Museum fur Naturkunde, Stuttgart, Germany;UCMP, University of California Museum of Paleontology,Berkeley, California, USA; ZPAL, Institute of Paleobiologyof the Polish Academy of Science, Warsaw, Poland.
IX. REFERENCES
Abdala, F. & Giannini, N. P. (2002). Chiniquodontid cynodonts:systematic and morphometric considerations. Palaeontology 45,1151–1170.
Abdala, F. & Ribeiro, A. M. (2003). A new traversodontidcynodont from the Santa Maria Formation, Ladinian-Carnian,of southern Brazil, with a phylogenetic analysis of Gondwanantraversodontids. Zoological Journal of the Linnean Society 139,529–545.
Abdala, F. & Smith, R.M.H. (2009). A Middle Triassic cynodontfauna from Namibia and its implications for the biogeography ofGondwana. Journal of Vertebrate Paleontology, in press.
Alcober, O. (2000). Redescription of the skull of Saurosuchus galilei
(Archosauria: Rauisuchidae). Journal of Vertebrate Paleontology 20,302–316.
Alcober, O. & Parrish, J. M. (1997). A new poposaurid fromthe Upper Triassic of Argentina. Journal of Vertebrate Paleontology
17, 548–556.Allain, R., Aquesbi, N., de Jax, J., Meyer, C., Mon-
baron, M., Montenat, C., Richir, P., Rochdy, M., Rus-sell D. & Taquet, P. (2004). A basal sauropod dinosaur fromthe Early Jurassic of Morocco. Comptes Rendus Palevol 3, 199–208.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 95
Allain, R., Tykoski R. S., Aquesbi, N., Jalil, N.-E., Mon-baron, M., Russell, D. & Taquet, P. (2007). An abelisauroid(Dinosauria: Teropoda) from the Early Jurassic of High AtlasMountains, Morocco, and the radiation of ceratosaurs. Journal of
Vertebrate Paleontology 27, 610–624.Allen, D. (2004). The phylogenetic status of Procompsog-
nathus revisited. Journal of Vertebrate Paleontology 24(3, suppl.),34A.
Anderson, J. M. & Anderson, H. M. (1993). Terrestrial floraand fauna of the Gondwana Triassic: Part 1-occurrences. In The
Nonmarine Triassic (eds. S. G. Lucas & M. Morales). New Mexico
Museum of Natural History & Sciences Bulletin 3 3–12.Anderson, J. M., Anderson, H. M. & Cruickshank, A.R.I.
(1998). Late Triassic ecosystems of the Molteno/Lower Elliotbiome of southern Africa. Palaeontology 41, 387–421.
Andrews, C. W. (1921). On some remains of a theropodousdinosaur from the Lower Lias of Barrow–on–Soar. Annals and
Magazine of Natural History 8, 570–576.Arcucci, A. B. (1987). Un nuevo Lagosuchidae (Thecodon-
tia–Pseudosuchia) de la Fauna de los Chanares (Edad ReptilChanarense, Triasico medio), La Rioja, Argentina. Ameghiniana
24, 89–94.Arcucci, A. B. (1997). Dinosauromorpha. In Encyclopedia of
Dinosaurs (eds. P. J. Currie and K. Padian), pp. 179–184.Academic Press, San Diego.
Arcucci, A. B. (1998). New information about dinosaur precursorsfrom the Triassic Los Chanares Fauna, La Rioja, Argentina.Journal of African Earth Sciences 27, 9–10.
Arcucci, A. B. (2005). Una reevalucion de los dinosauriomorfosbasales y el origen de Dinosauria. In Boletim de Resumos do
II Congresso Latino-americano de Paleontologia de Vertebrados (eds.A. W. A. Kellner, D. D. R. Henriques & T. Rodrigues), p. 33.Museu Nacional, Rio de Janeiro, Serie Livros 12.
Arcucci, A. B. & Coria, R. A. (2003). A new Triassic carnivorousdinosaur from Argentina. Ameghiana 40, 217–228.
Attridge, J., Crompton, A. W. & Jenkins Jr., F. A. (1985). Thesouthern African Liassic prosauropod Massospondylus discoveredin North America. Journal of Vertebrate Paleontology 5, 128–132.
Avanzini, M. (2002). Dinosauromorph tracks from the MiddleTriassic (Anisian) of the Southern Alps I (Valle di Non-Italy).Bollettino della Societa Geologica Italiana 41, 37–40.
Averianov, A. O., Bakirov, A. A. & Martin, T. (2007). Firstdefinitive stegosaur from the Middle Jurassic of Kyrgyzstan.Palaontologische Zeitschrift 81, 440–446.
Averianov, A. O., Martin, T. & Bakirov, A. A. (2005).Pterosaur and dinosaur remains from the Middle Jurassic Bal-abansai Svita in the Northern Fergana Depression, Kyrgyzstan(Central Asia). Palaeontology 48, 135–155.
Azevedo, S. A. K., Schultz, C. L. & Barberena, M. C. (1990).Novas evidencias biostratigraficas e paleoecologicas na analise deevolucao explosiva dos rincossauros do Triassico. Paula-Coutiana
4, 23–33.Baez, A. M. & Marsicano, C. A. (2001). A heterodontosaur
ornithischian dinosaur from the Upper Triassic of Patagonia.Ameghiana 38, 271–279.
Bakker, R. T. (1971). Dinosaur physiology and the origin ofmammals. Evolution 25, 636–658.
Bakker, R. T. (1986). The dinosaur heresies. Bath Press, Avon.Bakker, R. T. & Galton, P. M. (1974). Dinosaur monophyly and
a new class of vertebrates. Nature 248, 168–172.
Bambach, R. K., Knoll, A. H. & Wang, S. (2004). Origination,extinction, and mass depletions of marine diversity. Paleobiology
30, 522–542.Barrett, P. M. (2000). Prosauropod dinosaurs and iguanas:
speculations on the diets of extinct reptiles. In Evolution of herbivory
in terrestrial vertebrates. (ed. H.-D. Sues), pp. 42–78. CambridgeUniversity Press, Cambridge.
Barrett. P. M. & Batten, D. J. (eds.) (2007). Evolution andPalaeobiology of Early Sauropodomorph Dinosaurs. Special Papers
in Palaeontology 77.Barrett, P. M., Butler, R. J. & Knoll, F. (2005a). Small-
bodied ornithischian dinosaurs from the Middle Jurassic ofSichuan, China. Journal of Vertebrate Paleontology 24, 823–834.
Barrett, P. M., Butler, R. J., Novas, F. E., Moore-Fay, S. C.,Moody, J. M., Clark, J. M. & Sanchez-Villagra, M. R.(2008). Dinosaur remains from the La Quinta Formation (Loweror Middle Jurassic) of the Venezuelan Andes. Palaontologische
Zeitschrift 82, 163–177.Barrett, P. M. & Upchurch, P. (2007). The evolution of
herbivory in sauropodomorph dinosaurs. In Evolution and
Palaeobiology of Early Sauropodomorph Dinosaurs (eds. P. M. Barrettand D. J. Batten). Special Papers in Palaeontology 77 91–112.
Barrett, P. M, Upchurch, P. & Xiao-lin W. (2005b). Cra-nial osteology of Lufengosaurus huenei Young (Dinosauria:Prosauropoda) from the Lower Jurassic of Yunnan, People’sRepublic of China. Journal of Vertebrate Paleontology 25, 806–822.
Begon, M., Harper, J. L. & Townsend, C. R. (1996). Ecology.
Third Edition. Blackwell Science.Benedetto, J. L. (1973). Herrerasauridae, nueva familia de
Saurısquios Triasicos. Ameghiniana 10, 89–102.Bennett, S. C. (1996). The phylogenetic position of the
Pterosauria within Archosauromorpha. Zoological Journal of the
Linnean Society 118, 261–308.Benton, M. J. (1983a). Dinosaur success in the Triassic: a
noncompetitive ecological model. The Quarterly Review of Biology
58, 29–55.Benton, M. J. (1983b). The age of the rhynchosaurs. New Scientist
98, 9–13.Benton, M. J. (1984). Dinosaurs’ lucky break. Natural History 93,
54–59.Benton, M. J. (1985). Classification and phylogeny of diapsid
reptiles. Zoological Journal of the Linnean Society 84, 97–164.Benton, M. J. (1986a). More than one event in the late Triassic
mass extinction. Nature 321, 857–861.Benton, M. J. (1986b). The late Triassic reptile Teratosaurus - a
rauisuchian, not a dinosaur. Palaeontology 29, 293–301.Benton, M. J. (1989). End-Triassic. In Palaeobiology, a synthesis (eds.
D. E. G. Briggs and P. R. Crowther), pp. 194–198. BlackwellScience, Oxford.
Benton, M. J. (1990). Origin and interrelationships of dinosaurs.In The Dinosauria (eds. D. B. Weishampel, P. Dodson andH. Osmolska), pp. 11–30. University of California Press,Berkley.
Benton, M. J. (1991). What really happened in the Late Triassic?Historical Biology 5, 263–278.
Benton, M. J. (ed.) (1993). The fossil record 2. Chapman and Hall,London.
Benton, M. J. (1994a). Late Triassic terrestrial vertebrate extinc-tions: stratigraphic aspects and the record of the Germanic Basin.Paleontologia Lombarda 2, 19–38.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

96 Max C. Langer and others
Benton, M. J. (1994b). Late Triassic to Middle Jurassic extinctionsamong continental tetrapods: testing the pattern. In In the shadow
of the dinosaurs, early Mesozoic tetrapods (eds. N. C. Fraser and H.-D. Sues), pp. 366–397 Cambridge University Press, Cambridge.
Benton, M. J. (1997). II. Extinction, Triassic. In Encyclopedia
of Dinosaurs (eds. P. J. Currie and K. Padian), pp. 230–236Academic Press, San Diego.
Benton, M. J. (1999). Scleromochlus taylori and the origin of dinosaursand pterosaurs. Philosophical Transactions of the Royal Society of London
B 354, 1423–1446.Benton, M. J. (2000). Stems, nodes, crown clades, and rank-free
lists: is Linnaeus dead? Biological Reviews 75, 633–648.Benton, M. J. (2003). When life nearly died: the greatest mass extinction
of all time. Thames & Hudson, London.Benton, M. J. (2004). Origin and relationships of Dinosauria. In
The Dinosauria, second edition (eds. D. B. Weishampel, P. Dodsonand H. Osmolska), pp. 7–19. University of California Press,Berkley.
Benton, M. J. & Clark, J. M. (1988). Archosaur phylogenyand the relationships of the Crocodylia. In The Phylogeny and
Classification of the Tetrapods. Vol. 1: Amphibians and Reptiles (ed. M. J.Benton), pp. 295–338 Clarendon Press, Oxford.
Benton, M. J., Juul, L., Storrs, G. W. & Galton, P. M.(2000). Anatomy and systematics of the prosauropod dinosaurThecodontosaurus antiquus from the Upper Triassic of southernEngland. Journal of Vertebrate Paleontology 20, 77–108.
Benton, M. J. & Spencer, P. S. (1995). Fossil Reptiles of GreatBritain. Chapman and Hall, London.
Benton, M. J. & Walker, A. D. (1985). Palaeoecology, taphon-omy and dating of Permo-Triassic reptiles from Elgin, north-eastScotland. Palaeontology 28, 207–234.
Bittencourt, J. S. (2008). Triassic dinosaurs from Brazil: speciesand clades of a diversified saurischian fauna. In Libro de Resumenes,
III Congreso Latinoamericano de Paleontologia de Vertebrados (eds.J. O. Calvo, R. J. Valieri, J. D. Porfiri and D. dos Santos), p. 24.Universidade Nacional del Comahue, Neuquen, Argentina.
Bittencourt, J. S. & Kellner, A. W. A. (2009). The anatomyand phylogenetic position of the Triassic dinosaur Staurikosaurus
pricei Colbert, 1970. Zootaxa 2079, 1–56.Blakey, R. (2006). Mollewide plate tectonic maps. http://
jan.ucc.nau.edu/∼rcb7/mollglobe.html.Bonaparte, J. F. (1962). Descripcion del craneo y mandıbula
de Exaeretodon frenguellii Cabrera, y su comparacion con Diade-modontidae, Tritylodontidae y los cinodontes sudamericanos.Publicaciones del Museo Municipal de Ciencias Naturales y Tradicional de
La Plata 1, 135–202.Bonaparte, J. F. (1966). Cronologıa de algunas formaciones
triasicas argentinas. Basada en restos de tetrapodos. Revista de
la Asociacion Geologica Argentina 21, 20–38.Bonaparte, J. F. (1972). Los tetrapodos del sector superior de
la Formacion Los Colorados, La Rioja, Argentina (TriasicoSuperior). I Parte. Opera Lilloana 22, 1–183.
Bonaparte, J. F. (1975). Nuevos materiales de Lagosuchus talam-
payensis Romer (Thecodontia - Pseudosuchia) y su significado enel origen de los Saurischia. Chanarense inferior, Triasico Mediode Argentina. Acta Geologica Lilloana 13, 5–90.
Bonaparte, J. F. (1976). Pisanosaurus mertii Casamiquela and theorigin of the Ornithischia. Journal of Paleontology 50, 808–820.
Bonaparte, J. F. (1978). Coloradia brevis n. g. et n. sp. (Saurischia,Prosauropoda), dinosaurio Plateosauridae de la Formacion los
Colorados, Triasico Superior de La Rioja, Argentina. Ameghiniana
15, 327–332.Bonaparte J. F. (1982). Faunal replacement in the Triassic of
South America. Journal of Vertebrate Paleontology 2, 362–371.Bonaparte, J. F. (1984). Locomotion in rauisuchid thecodonts.
Journal of Vertebrate Paleontology 3, 210–218.Bonaparte, J. F. (1986). The early radiation and phylogenetic
relationships of the Jurassic sauropod dinosaurs, based onvertebral anatomy. In The beginning of the age of dinosaurs
(ed. K. Padian), pp. 247–258 Cambridge University Press,Cambridge.
Bonaparte, J. F. (1995). Origen de los dinosaurios. Anales de la
Academia Nacional de Ciencias Exactas Fısicas y Naturales de Buenos Aires
47, 77–88.Bonaparte, J. F. (1997). El Triasico de San Juan - La Rioja, Argentina
y sus dinosaurios. Museo Argentino de Ciencias Naturales, BuenosAires.
Bonaparte J. F. & Barberena, M. C. (2001). On two advancedcarnivorous cynodonts from the Late Triassic of southern Brazil.Bulletin of the Museum of Comparative Zoology 156, 59–80.
Bonaparte J. F., Brea G., Schultz C. L. & Martinelli A. G.(2007). A new specimen of Guaibasaurus candelariensis (basalSaurischia) from the Late Triassic Caturrita Formation ofsouthern Brazil. Historical Biology 19, 73–82.
Bonaparte, J. F., Ferigolo, J. & Ribeiro, A. M. (1999). A newEarly Late Triassic saurischian dinosaur from Rio Grande do SulState, Brazil. National Science Museum Monographs 15, 89–109.
Bonaparte, J. F. & Plumares, J. A. (1995). Notas sobre elprimer craneo de Riojasaurus incertus (Dinosauria, Prosauropoda,Melanorosauridae) del Triasico Superior de La Rioja, Argentina.Ameghiniana 32, 341–349.
Bonaparte, J. F. & Powell, J. E. (1980). A continental assem-blage of tetrapods from the Upper Cretaceous beds ofEl Brete, northwestern Argentina (Sauropoda–Coelurosauria–Carnosauria–Aves). Memoires de la Societe geologique de France 139,19–28.
Bonaparte, J. F. & Vince, M. (1979). El hallazgo del primernido de dinosaurios triasicos (Saurischia, Prosauropoda) Triasicosuperior de Patagonia, Argentina. Ameghiniana 16, 173–182.
Bonnan, M. F., & Senter, P. (2007). Were the basalsauropodomorph dinosaurs Plateosaurus and Massospondylus habit-ual quadrupeds? Special Papers in Palaeontology 77, 139–155.
Botha, J., Abdala, F. & Smith, R. (2007). The oldest cyn-odont: new clues on the origin and early diversificationof the Cynodontia. Zoological Journal of the Linnean Society
149, 477–492.Brinkman, D. B. & Sues, H.–D. (1987). A staurikosaurid dinosaur
from the Upper Triassic Ischigualasto Formation of Argentinaand the relationships of the Staurikosauridae. Palaeontology 30,493–503.
Bristowe, A. & Raath, M.A. (2004). A juvenile coelophysoidskull from the early Jurassic of Zimbabwe, and the synonymy ofCoelophysis and Syntarsus. Palaeontologia Africana 40, 31–41.
Brusatte, S. L., Benton, M. J., Ruta, M. & Lloyd, G. T.(2008a). Superiority, competition, and opportunism in theevolutionary radiation of dinosaurs. Science 321, 1485–1488.
Brusatte, S. L., Benton, M. J., Ruta, M. & Lloyd, G. T.(2008b). The first 50 Myr of dinosaur evolution: macroevolu-tionary pattern and morphological disparity. Biology Letters 4,733–736.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 97
Brusatte, S. L., Benton, M. J., Desojo, J. B. & Langer, M. C.(2009). The higher-level phylogeny of Archosauria (Tetrapoda:Diapsida). Journal of Systematic Palaeontology, in press.
Buchholz, P. W. (2002). Phylogeny and biogeography of basalOrnithischia. In The Mesozoic in Wyoming (ed. D. E. Brown),pp. 18–34 Tate Geological Museum, Casper.
Buffetaut, E., Martin, V., Sattayarak, N. & Suteethorn,V. (1995). The oldest known dinosaur from Southeast Asia: aprosauropod from the Nam Phong Formation (Late Triassic) ofnortheastern Thailand. Geological Magazine 132, 739–742.
Buffetaut, E., Suteethorn, V., Cuny, G., Tong, H., LeLoeuff, J., Khansubha, S. & Jongautchariyakul, S.(2000). The earliest known sauropod dinosaur. Nature 407, 72–74.
Butler, R. J. (2005). The ‘fabrosaurid’ ornithischian dinosaurs ofthe Upper Elliot Formation (Lower Jurassic) of South Africa andLesotho. Zoological Journal of the Linnean Society 145, 175–218.
Butler, R. J., Porro, L. B. & Heckert, A. B. (2006). A supposedheterodontosaurid tooth from the Rhaetian of Switzerland and areassessment of the European Late Triassic record of Ornithischia(Dinosauria). Neues Jahrbuch fur Geologie und Palaontologie, Monatshefte
10, 613–633.Butler, R. J., Smith, R. M. H. & Norman, D. B. (2007).
A primitive ornithischian dinosaur from the Late Triassic ofSouth Africa, and the early evolution and diversification ofOrnithischia. Proceedings of the Royal Society B 274, 2041–2046.
Butler, R., Upchurch, P. & Norman, D.B. (2008). Thephylogeny of the ornithischian dinosaurs. Journal of Systematic
Palaeontology 6, 1–40.Camp, C. L. (1936). A new type of small bipedal dinosaur from the
Navajo Sandstone of Arizona. University of California Publications on
Geological Sciences 24, 39–56.Cantino, P.D. & de Queiroz, K. (2007). International Code
of Phylogenetic Nomenclature. Version 4. http://www.ohiou.edu/phylocode/.
Carpenter, K. (1997). A giant coelophysoid (Ceratosauria)theropod from the Upper Triassic of New Mexico, USA. Neues
Jahrbuch fur Geologie und Palaontologie Abhandlungen 205, 189–208.Carrano, M. T. (2000). Homoplasy and the evolution of dinosaur
locomotion. Paleobiology 26, 489–512.Carrano, M.T., Hutchinson, J.R. & Sampson, S.D. (2005).
New information on Segisaurus halli, a small theropod dinosaurfrom the Early Jurassic of Arizona. Journal of Vertebrate Paleontology
22, 510–534.Carrano, M. T. & Sampson, S. D. (2004). A review of coelo-
physoids (Dinosauria: Theropoda) from the Early Jurassic ofEurope, with comments on the late history of the Coelo-physoidea. Neues Jahrbuch fur Geologie und Palaontologie Monatshefte
2004, 537–558.Carrano, M. T. & Sampson, S. D. (2008). The phylogeny of
Ceratosauria (Dinosauria: Theropoda). Journal of Systematic
Palaeontology 6, 183–236.Carrano, M. T., Sampson, S. D. & Forster, C. A. (2002).
The osteology of Masiakasaurus knopfleri, a small abelisauroid(Dinosauria: Theropoda) from the Late Cretaceous of Madagas-car. Journal of Vertebrate Paleontology 22, 510–534.
Casamiquela, R. M. (1967). Un nuevo dinosaurio ornitischioTriasico (Pisanosaurus mertii; Ornithopoda) de la FormacionIschigualasto, Argentina. Ameghiniana 4, 47–64.
Caselli, A.T., Marsicano, C.A. & Arcucci, A.B. (2001).Sedimentologıa y paleontologıa de la Formacion Los Colorados,
Triasico superior (provincias de La Rioja y San Juan, Argentina).Revista de la Asociacion Geologica Argentina 56, 173–188.
Charig, A. J. (1972). The evolution of the archosaur pelvisand hind-limb: an explanation in functional terms. In Studies
in Vertebrate Evolution (eds. K. A. Joysey and T. S. Kemp),pp. 121–155 Oliver & Boyd, Edinburgh.
Charig, A.J. (1980). Differentiation of lineages among Mesozoictetrapods. Memoir de la Societe Geologique de France 139, 207–210.
Charig, A.J. (1984). Competition between therapsids andarchosaurs during the Triassic Period: a review and synthesisof current theories. Symposia of the Zoological Society of London 52,597–628.
Charig, A. J. & Crompton, A. W. (1974). The alleged synonymyof Lycorhinus and Heterodontosaurus. Annals of the South African Museum
64, 167–189.Charig, A. J., Attridge, J. & Crompton, A. W. (1965). On the
origin of the sauropods and the classification of the Saurischia.Proceedings of the Linnean Society of London 176, 197–221.
Chatterjee, S. (1978). A primitive parasuchid (phytosaur) reptilefrom the Upper Triassic Maleri Formation of India. Palaeontology
21, 83–127.Chatterjee, S. (1982). A new cynodont reptile from the Triassic
of India. Journal of Paleontology 56, 203–214.Chatterjee, S. (1984). A new ornithischian dinosaur from the
Triassic of North America. Naturwissenschaften 71, 630–631.Chatterjee, S. (1987). A new theropod dinosaur from India with
remarks on the Gondwana-Laurasia connection in the LateTriassic. In Gondwana Six: Stratigraphy Sedimentology and Paleontology
(ed. G. D. Mckenzie).Geophysical Monography 41 183–189.Chatterjee, S. (1991). Cranial anatomy and relationships of a
new Triassic bird from Texas. Philosophical Transactions of the Royal
Societyof London B 332, 277–342.Christian, A. & Preuschoft, H. (1996). Deducing the body
posture of extinct large vertebrates from the shape of the vertebralcolumn. Palaeontology 39, 801–812.
Clarke, J. A. (2004). Morphology, phylogenetic taxonomy, andsystematics of Ichthyornis and Apatornis (Avialae: Ornithurae).Bulletin of the American Museum of Natural History 286, 1–179.
Colbert, E. H. (1958). Triassic tetrapod extinction at the end ofthe Triassic Period. Proceedings of the National Academy of Sciences 44,973–977.
Colbert, E. H. (1964). Relationships of the saurischian dinosaurs.American Museum Novitates 2181, 1–24.
Colbert, E. H. (1970). A saurischian dinosaur from the Triassicof Brazil. American Museum Novitates 2405, 1–39.
Colbert, E. (1981). A primitive ornithischian dinosaur from theKayenta Formation of Arizona. Bulletin of the Museum of Northern
Arizona 53, 1–61.Colbert, E. H. (1989). The Triassic dinosaur Coelophysis. Bulletin of
the Museum of Northern Arizona 57, 1–160.Colbert, E. H., Charig, A. J., Dodson, P., Gillette, D. D.,
Ostrom, J. H., & Weishampel, D. (1992). Coelurus bauri Cope1887 (currently Coelophysis bauri; Reptilia, Saurischia): proposedreplacement of the lectotype by a neotype. Bulletin of Zoological
Nomenclature 49, 276–279.Colbert, E. H. & Russell, D. A. (1969). The small Cretaceous
dinosaur Dromaeosaurus. American Musem Novitates 2380, 1–49.Colombi, C. E. & Parrish, J. T. (2008). Late Triassic Environ-
mental Evolution in Southwestern Pangea: Plant Taphonomy ofthe Ischigualasto Formation. Palaios 23, 778–795.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

98 Max C. Langer and others
Coombs, W. P. (1990). Behaviour patterns of dinosaurs. In The
Dinosauria (eds. D. B. Weishampel, P. Dodson andH. Osmolska), pp. 32–42 University of California Press,Berkley.
Cooper, M. R. (1981). The prosauropod dinosaur Massospondylus
carinatus Owen from Zimbabwe: its biology, mode of life andphylogenetic significance. Occasional Papers of the National Museums
and Monuments of Rhodesia 6, 689–840.Cooper, M. R. (1985). A revision of the ornithischian dinosaur
Kangnasaurus coetzeei Haughton, with a classification of theOrnithischia. Annals of the South African Museum
95, 281–317.Cope, E. D. (1866). Remarks on dinosaur remains from New Jersey.
Proceedings of the Academy of Natural Sciences of Philadelphia 1866,275–279.
Cope, E. D. (1889). On a new genus of Triassic Dinosauria. The
American Naturalist 23, 626.Coria, R. A. & Salgado, L. (2000). A basal Abelisauria Novas
1992 (Theropoda–Ceratosauria) from the Cretaceous ofPatagonia, Argentina. Gaia 15, 89–102.
Cornet, B. & Olsen, P. E. (1990). Early to Middle Carnian (Triassic)
flora and fauna of the Richmond and Taylorsville basins, Virginia
and Maryland, U.S.A. Virginia Museum of Natural History,Martinsville.
Cox, C. B. (1965). New Triassic dicynodonts from South America,their origins and relationships. Philosophical Transactions of the Royal
Society of London B 248, 457–516.Cox, C. B. (1967). Changes in terrestrial vertebrate faunas during
the Mesozoic. In The Fossil Record (ed. W. B. Harland),pp. 71–89 Geological Society, London.
Crowley, T.J. (1994). Pangean climates. In: Pangea: Paleoclimate,
Tectonics, and Sedimentation during Accretion, Zenith and Breakup of a
Supercontinent (ed. G.D. Klein). Geological Society of AmericaSpecial Papers 288 25–39.
Crompton, A. W. & Attridge, J. (1986). Masticatory apparatusof the larger herbivores during Late Triassic and Early Jurassictimes. In The beginning of the age of dinosaurs (ed. K. Padian),pp. 223–236 Cambridge University Press, Cambridge.
Crompton, A. W. & Charig, A. J. (1962). A new ornithis-chian from the Upper Triassic of South Africa. Nature
196, 1074–1077.Cruickshank, A. R. I. (1975). The origin of sauropod dinosaurs.
South African Journal of Science 71, 89–90.Cruickshank, A. R. I. (1979). The ankle joint in some early
archosaurs. South African Journal of Science 75, 168–178.Cuny, G. (1995). French vertebrate faunas and the Triassic-
Jurassic boundary. Palaeogeography, Palaeoclimatology, Palaeoecology
119, 343–358.Cuny, G., Hunt, A. P., Mazin, J-M. & Rauscher, R. (2000).
Teeth of enigmatic neoselachian sharks and an ornithischiandinosaur from the uppermost Triassic of Lons-le-Saunier (JuraFrance). Palaontologische Zeitschrift 74, 171–185.
Currie, B. S., Colombi, C. E., Tabor, N. J., Shipman, T. C. &Montanez, I. P. (2009). Stratigraphy and architecture of theUpper Triassic Ischigualasto Formation, Ischigualasto ProvincialPark, San Juan, Argentina. Journal of South American Earth Sciences
27, 74–97.Currie, P. J. (1997). Theropoda. In Encyclopedia of Dinosaurs
(eds. P. J. Currie and K. Padian), pp. 731–737 Academic Press,San Diego.
Currie, P. J. & Padian, K. (1997a). Genasauria. In Encyclopedia
of Dinosaurs (eds. P. J. Currie and K. Padian), p. 271. AcademicPress, San Diego.
Currie, P. J. & Padian, K. (1997b). Thecodontia. In Encyclopedia
of Dinosaurs (eds. P. J. Currie and K. Padian), p. 728. AcademicPress, San Diego.
Da Rosa, A. A. S. Leal, L. A. Boelter, R. A. & Dambros, C. S.(2006). Um novo Sauropodomorpha para o Triassico Superiordo sul do Brasil. Revista Ciencia e Natura Edicao Especial 31.
de Queiroz, K. & Gauthier, J. (1990). Phylogeny as a centralprinciple in taxonomy: phylogenetic definitions of taxon names.Systematic Zoology 39, 307–322.
de Queiroz, K. & Gauthier, J. (1992). Phylogenetic taxonomy.Annual Review of Ecology and Systematics 23, 449–480.
De Queiroz, K. & Gauthier, J. (1994). Toward a phylogeneticsystem of biological nomenclature. Trends in Ecology and Evolution
9, 27–31.Demathieu, G. & Demathieu, P. (2004). Chirotheria and other
ichnotaxa of the European Triassic. Ichnos 11, 79–88.Dilkes, D. W. (1998). The Early Triassic rhynchosaur Mesosuchus
browni and the interrelationships of basal archosauromorphreptiles. Philosophical Transactions of the Royal Society of London B
353, 501–541.Dong, Z.-M. (2001). Primitive armored dinosaur from the Lufeng
Basin, Yunnan. In Mesozoic Vertebrate Life (eds. D. H. Tanke &K. Carpenter), pp. 237–242 Indiana University Press, Bloom-ington.
Dzik, J. (2003). A beaked herbivorous archosaur with dinosauraffinities from the Early Late Triassic of Poland. Journal of Vertebrate
Paleontology 23, 556–574.Dzik, J. & Sulej, T. (2007). A review of the early triassic krasiejow
biote from Silesia, Poland. Paleontologia Polonica 64, 3–27.Dzik, J., Sulej, T. & Niedywiedzki, G. (2008). A dicynodont-
theropod association in the latest Triassic of Poland. ACTA
Paleontologica Polonica 53, 733–7381.Ellenberger, P. (1974). Contributiona la classification des pistes
de vertebres du Trias: les types du Stormberg d’Afrique duSud (IIeme Partie, le Stormberg superieur - I - Le biome de lazone B/1 ou niveau de Moyeni: ses biocenoses). Palaeovertebrata
(Memoire Extraordinaire) 104, 1–143.Evans, S.E. & Milner, A.R. (1994). Middle Jurassic microver-
tebrate lssemblages from the British Isles. in In the Shadow of
the Dinosaurs, Early Mesozoic Tetrapods (eds. N. C. Fraser and H.-D. Sues), pp. 303–321 Cambridge University Press, Cambridge.
Ewer, R. F. (1965). The anatomy of the thecodont reptile Euparkeria
capensis Broom. Philosophical Transactions of the Royal Society of London
B 248, 379–435.Ezcurra, M. D. (2006). A review of the systematic position of the
dinosauriform archosaur Eucoelophysis baldwini from the UpperTriassic of New Mexico, USA. Geodiversitas 28, 649–684.
Ezcurra, M. D. (2008). A new early dinosaur from the CarnianIschigualasto Formation (Nw Argentina) and the origin ofdinosaurs. In: Libro de Resumenes, III Congreso Latinoamericano
de Paleontologia de Vertebrados (eds. J. O. Calvo, R. J. Valieri,J. D. Porfiri and D. dos Santos), p. 87. Universidade Nacionaldel Comahue, Neuquen, Argentina.
Ezcurra, M. D. & Cuny, G. (2007). The coelophysoid Lophostro-
pheus airelensis, gen. nov.: a review of the systematics of ‘‘Lilienster-
nus’’ airelensis from the Triassic-Jurassic outcrops of Normandy(France). Journal of Vertebrate Paleontology 27, 73–86.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 99
Ezcurra, M. D, Lecuona, A. & Irmis, R. B. (2008). A reviewof the archosaur Trialestes romeri (Suchia, Crocodylomorpha)from the Upper Triassic Ischigualasto Formation of Argentina.In: Libro de Resumenes, III Congreso Latinoamericano de Paleontologia
de Vertebrados (eds. J. O. Calvo, R. J. Valieri, J. D. Porfiri andD. dos Santos), p. 88. Universidade Nacional del Comahue,Neuquen, Argentina.
Ezcurra, D. M. & Novas, F. E. (2007a). Phylogenetic relation-ships of the Triassic theropod Zupaysaurus Rougieri from NWArgentina. Historical Biology 19, 35–72.
Ezcurra, D. M. & Novas, F. E. (2007B). New dinosaur remains(Saurischia: Herrerasauridae) from the Ischigualasto Formation(Carnian) of NW Argentina. Ameghiniana 44, 17R.
Ezcurra, M. & Novas, F. E. (2008). A review of the dinosaurdiversity of the Ischigualasto Formation (Carnian, NWArgentina): insights on early dinosaur evolution. In Boletim de
Resumos, VI Simposio Brasileiro de Paleontologia de Vertebrados (eds.M. C. Langer, J. S. Bittencourt and M. C. Castro). Paleontologia
Em Destaque Edicao Especial 88–89.Farina, R. A. (1995). Bone strength and habits in large glyptodonts.
Lethaia 28, 189–196.Fechner, R. (2006). Evolution of bipedality in dinosaurs. Journal of
Vertebrate Paleontology 26(3, Suppl.), 60A.Ferigolo, J. & Langer, M. C. (2007). A late Triassic dinosauri-
form from south Brazil and the origin of the ornithischianpredentary bone. Historical Biology 19, 23–33.
Flynn, J. J., Fox, S. R., Parrish, J. M., Ranivoharimanana,L. & Wyss, A. R. (2006). Assessing diversity and paleoecology ofa Middle Jurassic microvertebrate assemblage from Madagascar.New Mexico Museum of Natural History & Science Bulletin 37, 476–489.
Flynn, J., Nesbitt, S., Parrish, M., Ranivoharimanana, L.& Wyss, A. (2008). A new species of basal archosauromorph fromthe Late Triassic of Madagascar. Journal of Vertebrate Paleontology
28(3, Suppl.), 78A.Flynn, J. J., Parrish, J. M.; Rakotosamimanana, B.; Sim-
son, W. F.; Whatley, R. L. & Wyss, A. R. (1999). A Triassicfauna from Madagascar, including early dinosaurs. Science 286,763–765.
Frakes, L. A, Francis, E. & Syktus, J. I. (1992). Climate modes of
the Phanerozoic; the history of the Earth’s climate over the past 600 million
years. Cambridge Univesity Press. Cambridge.Fraser, N. C. (1994). Assemblages of small tetrapods from British
Late Triassic fissure fillings. In In the shadow of the dinosaurs, early
Mesozoic tetrapods (eds. N. C. Fraser and H.-D. Sues), PP. 214–226Cambridge University Press, Cambridge.
Fraser, N. C. (2006). Dawn of the Dinosaurs: Life in the Triassic.Indiana University Press, Bloomington.
Fraser, N. C. & Sues, H.-D. (1994). Comments on Benton’s‘‘Late Triassic to Middle Jurassic extinctions among continentaltetrapods’’. In In the shadow of the dinosaurs, early Mesozoic tetrapods
(eds. N. C. Fraser and H.-D. Sues), pp. 398–400 CambridgeUniversity Press, Cambridge.
Fraser, N. C., Padian, K., Walkden, G. M. & Davis, A. L. M.(2002). Basal dinosauriform remains from Britain and thediagnosis of the Dinosauria. Palaeontology 45, 78–95.
Furin, S., Preto, N., Rigo, M., Roghi, G., Gianolla, P.,Crowley, J. L., & Bowring, S. A. (2006). High-precision U-Pbzircon age from the Triassic of Italy: implications for the Triassictime scale and the Carnian origin of calcareous nannoplanktonand dinosaurs. Geology 34, 1009–1012.
Gallet, Y., Krystyn, L., Besse, J. & Marcoux, J (2003).Improving the Upper Triassic Numerical time scale from cross-correlation between Tethyan marine sections and the continentalNewark basin sequence. Earth and Planetary Science Letters 212,255–261.
Galton, P. M. (1971). The prosauropod dinosaur Ammosaurus, thecrocodile Protosuchus, and their bearing on the age of the Navajosandstone of northeastern Arizona. Journal of Paleontology 45,781–795.
Galton, P. M. (1972). Classification and evolution of ornithopoddinosaurs. Nature 239, 464–466.
Galton, P. M. (1973). On the anatomy and relationships of Efraasia
diagnostica (Huene) n. gen., a prosauropod dinosaur (Reptilia:Saurischia) from the Upper Triassic of Germany. Palaontologische
Zeitschrift 47, 229–255.Galton, P. M. (1976). Prosauropod dinosaurs (Reptilia:
Saurischia) of North America. Postilla 169, 1–98.Galton, P. M. (1977). On Staurikosaurus pricei, an early saurischian
dinosaur from the Triassic of Brazil, with notes on theHerrerasauridae and Poposauridae. Palaontologische Zeitschrift 51,234–245.
Galton, P. M. (1978). Fabrosauridae, the basal family ofornithischian dinosaurs (Reptilia: Ornithischia). Palaeontologische
Zeitschrift 52(1/2): 138–159.Galton, P. M. (1980). European Jurassic ornithopod dinosaurs
of the families Hypsilophodontidae and Camptosauridae.Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen
160, 73–95.Galton, P. M. (1983a). Armored dinosaurs (Ornithischia: Anky-
losauria) from the Middle and Upper Jurassic of England. Geobios
13, 825–837.Galton, P. M. (1983b). The oldest ornithischian dinosaur in
North America from the Late Triassic of Nova Scotia, NCand PA. Geological Society of America, Abstracts with programs
15, 122.Galton, P. M. (1984). Cranial anatomy of the prosauropod
dinosaur Plateosaurus from the Knollenmergel (Middle Keuper,Upper Triassic) of Germany. I. Two complete skulls fromTrossingen/Wurtt, with comments on the diet. Geologica et
Palaeontologica 18, 139–171.Galton, P. M. (1985a). Cranial anatomy of the prosauropod
dinosaur Plateosaurus from the Knollenmergel (Middle Keuper)of Germany. II. All the cranial material and details of soft-partanatomy. Geologica et Palaeontologica 19, 119–159.
Galton, P. M. (1985b). The poposaurid thecodontian Teratosaurus
suevicus v. Meyer, plus referred specimens mostly based onprosauropod dinosaurs, from the Middle Stubensandstein (UpperTriassic) of Nordwurttemberg. Stuttgarter Beitrage zur Naturkunde B
116, 139–171.Galton, P. M. (1986). Herbivorous adaptations of Late Triassic
and Early Jurassic dinosaurs. In The beginning of the age of
dinosaurs (ed. K. Padian), pp. 203–221 Cambridge UniversityPress, Cambridge.
Galton, P. M. (1989). Prosauropoda, the basal Sauropodomor-pha. In The age of dinosaurs (eds. K. Padian and D. J. Chure),pp. 80–84 The Paleontological Society, Knoxville, Short Coursesin Palaeontology 2.
Galton, P. M. (1990). Stegosauria. In The Dinosauria (eds.D. B. Weishampel, P. Dodson and H. Osmolska), pp. 435–455University of California Press, Berkley.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

100 Max C. Langer and others
Galton, P. M. (2000). Are Spondylosoma and Staurikosaurus (SantaMaria Formation, Middle-Upper Triassic, Brazil) the oldestknown dinosaurs? Palaontologische Zeitschrift 74, 393–423.
Galton, P. M. (2001). Prosauropod dinosaurs from the UpperTriassic of Germany. In Actas de las I Jornadas Internacionales sobre
Paleontologıa de Dinosaurios y su Entorno (ed. Colectivo Arqueologico-Paleontologico de Salas), pp. 25–92 Junta de Castilla y Leon,Burgos, Spain.
Galton, P. M (2005A). Bones of large dinosaurs (Prosauropodaand Stegosauria) from the Rhaetic Bone Bed (Upper Triassic) ofAust Cliff, Southwest England. Revue de Paleobiologie 24, 51–74.
Galton, P. M. (2005B). Teeth of Ornithischian Dinosaurs (MostlyOrnithopoda) from the Morrison Formation (Upper Jurassic)of the Western United States. In Horns and Beaks. Ceratopsian
and Ornithopod Dinosaurs (Ed. K. Carpenter), PP. 17–48 IndianaUniversity Press, Bloomington.
Galton, P. M. (2007). Notes on the remains of archosaurianreptiles, mostly basal sauropodomorph dinosaurs, from the1834 fissure fill (Rhaetian, Upper Triassic) at Clifton in Bristol,southwest England. Revue de Paleobiologie 26, 505–559.
Galton, P. M. & Powell, H. P. (1983). Stegosaurian dinosaursfrom the Bathonian (Middle Jurassic) of England, the earliestrecord of Stegosauridae. Geobios 16, 219–229.
Galton, P. M. & Upchurch, P. (2000). Prosauropod dinosaurs:homeotic transformations (‘‘frame shifts’’) with third sacral as acaudosacral or a dorsosacral. Journal of Vertebrate Paleontology 20(3,SUPPL.), 43A.
Galton, P. M. & Upchurch, P. (2004). Prosauropoda. p. 232–258. In The Dinosauria, second edition (eds. D. B. Weishampel,P. Dodson and H. Osmolska), PP. 232–258 University ofCalifornia Press, Berkley.
Galton, P. M. & Van Heerden, J. (1998). Anatomy of theprosauropod dinosaur Blikanasaurus cromptoni (Upper Triassic,South Africa), with notes on the other tetrapods from the lowerElliot Formation. Palaontologische Zeitschrift 72, 163–177.
Galton, P. M. & Walker, A. (1996). Supposed prosauropoddinosaurs from Middle Triassic of England referred toParasauchia and Dinosauriformes. Neues Jarhbuch fur Geologie und
Palaontologie Monatshefte 1996, 727–738.Galton, P. M., Yates, A. M. & Kermack, D. (2007). Pantydraco
n. gen. for Thecodontosaurus caducus Yates, 2003, a basalsauropodomorph dinosaur from the Upper Triassic or LowerJurassic of South Wales, UK. Neues Jahrbuch fur Geologie und
Palaontologie Abhandlungen 243, 119–125.Gatesy, S. M., Middleton, K. M., Jenkins, F. A. & Shu-
bin, N. H. (1999). Three dimensional preservation on footmovements in Triassic theropod dinosaurs. Nature 399, 141–144.
Gauffre, F.-X. (1993). The prosauropod dinosaur Azendohsaurus
laaroussi from the Upper Triassic of Morocco. Palaeontology 36,897–908.
Gauthier, J. (1986). Saurischian monophyly and the origin ofbirds. In The origin of Birds and the Evolution of Flight (ed. K. Padian).Memoir of the California Academy of Science 8 1–55.
Gauthier, J., Cannatella, D., de Queiroz, K., Kluge, A. G.& Rowe, T. (1989). Tetrapod phylogeny. In The hierarchy of life:
molecules and morphology in phylogenetic analysis (eds. B. Fernholm,K. Bremer & H. Jornvall), PP. 337–353 Elsevier, Amsterdam.
Gauthier, J. & de Queiroz, K. (2001). Feathered dinosaurs,flying dinosaurs, crown dinosaurs, and the name ‘‘Aves’’. In New
Perspectives on the Origin and Early Evolution of Birds: Proceedings of the
International Symposium in Honor of John H. Ostrom (eds. J. Gauthier &L.F. Gall), PP. 7–41 Peabody Museum of Natural History, NewHaven.
Gauthier, J. & Padian, K. (1985). Phylogenetic, functional, andaerodynamic analyses of the origin of birds and their flight. InThe Beginning of Birds (eds. M. K. Hecht, J. H. Ostrom, G. Viohland P. Wellnhofer), PP. 185–197 Freunde des Jura Museums,Eichstatt.
Gillette, D. D. (2003). The geographic and phylogenetic positionof sauropod dinosaurs from the Kota formation (Early Jurassic)of India. Journal of Asian Earth Sciences 21, 683–689.
Glen, A. S. & Dickman, C. R. (2008). Niche overlap betweenmarsupial and eutherian carnivores: does competition threatenthe endangered spotted-tailed quoll? Journal of Applied Ecology 45,700–707.
Godefroit, P. & Cuny, G. (1997). Archosauriform teeth from theupper Triassic of Saint-Nicolas-de-Port (northeastern France).Palaeovertebrata 26, 1–34.
Godefroit, P. & Knoll, F. (2003). Late Triassic dinosaur teethfrom southern Belgium. Comptes Rendus Palevol 2, 3–11.
Golonka, J. (2002). Plate-tectonic Maps of the Phanerozoic.In Phanerozoic Reef Patterns (eds. W. Kiessling, E. Flugel andJ. Golonka). Society for Sedimentary Geology Special Publication 7221–75.
Golonka, J. (2007). Late Triassic and Early Jurassic palaeogeog-raphy of the world. Palaeogeography, Palaeoclimatology, Palaeoecology
244, 297–307.Golonka, J. & Ford, D. W. (2000). Pangean (Late Carboniferous-
Middle Jurassic) paleoenvironment and lithofacies. Palaeogeogra-
phy, Palaeoclimatology, Palaeoecology 161, 1–34.Gow, C. E. (1975). A new heterodontosaurid from the Red Beds
of South Africa showing clear evidence of tooth replacement.Zoological Journal of the Linnean Society of London 57, 335–339.
Gow, C. E., Kitching, J. W. & Raath, M. A. (1990). Skulls ofthe prosauropod dinosaur Massospondylus carinatus Owen in thecollections of the Bernard Price Institute for PalaeontologicalResearch. Palaeontologia Africana 27, 45–58.
Gower, D. J. (1999). The cranial and mandibular osteology of anew rauisuchian archosaur from the Middle Triassic of southernGermany. Stuttgarter Beitrage zur Naturkunde B 280, 1–49.
Gower, D. J. (2003). Osteology of the early archosaurian reptileErythrosuchus africanus Broom. Annals of the South Africa Museum 110,1–88.
Gower, D. J. & Sennikov A. G. (2000). Early archosaursfrom Russia. In The age of dinosaurs in Russia and Mon-
golia (eds. M. J. Benton, M. A. Shishkin, D. M. Unwin andE. N. Kurochkin), pp. 140–159 Cambridge University Press,Cambridge.
Hallam, A. (1981). The end–Triassic bivalve extinction event.Palaeogeography, Palaeoclimatology, Palaeoecology 35, 1–44.
Hallam, A. (1985). A review of Mesozoic climates. Journal of the
Geological Society 142, PP. 433–445.Hallam, A. (1990). The end–Triassic mass extinction event.
In Global catastrophes in earth history: an interdisciplinary conference
on impacts, volcanism, and mass mortality (eds. V. L. Sharptonand P. D. Ward). Geological Society of America Special Paper 247,577–583.
Hammer, W. R., Collinson, J.W. & Ryan, W.J. (1990). A newTriassic vertebrate fauna from Antarctica and its depositionalsetting. Antarctic Science 2, 163–169.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 101
Hammer, W. R. & Hickerson, W. J. (1994). A crested theropoddinosaur from Antarctica. Science 264, 828–830.
Haubold, H. (1991). Ein neuer Dinosaurier (Ornithischia,Thyreophora) aus dem unteren Jura des nordlichen Mitteleuropa.Revue de Paleobiologie 9, 149–177.
Haubold, H. & Klein, H. (2002). Chirotherien und Grallatoridenaus der Unteren bis Oberen Trias Mitteleuropas und dieEntstehung der Dinosauria. Hallesches Jahrbuch fur Geowissenschaften
B24, 1–22.He, X., Li, K. & Cai, K. (1988). [The Middle Jurassic Dinosaurian
Fauna from Dashanpu, Zigong, Sichuan: sauropod dinosaurs.Vol. 4, Omeisaurus tanfuensis.] Sichuan Scientific and TechnologicalPublishing House, Chengdu. (in Chinese with English summary).
Heckert, A. B. (2002). A revision of the upper Triassicornithischian dinosaur Revueltosaurus, with a description of anew species. New Mexico Museum of Natural History & Science Bulletin
21, 253–267.Heckert, A. B. (2004). Late Triassic microvertebrates from the
lower Chinle Group (Otischalkian-Adamanian: Carnian), south-western USA. New Mexico Museum of Natural History & Science
Bulletin 27, 1–170.Heckert, A. B. & Lucas, S. G. (2002). South American occur-
rences of the adamanian (Late Triassic: latest Carnian) indextaxon Stagonolepis (Archosauria: Aetosauria) and their biochrono-logical significance. Journal of Paleontology 76, 852–863.
Heckert, A. B., Lucas, S. G. & Sullivan, R. M. (2000). Triassicdinosaurs of New Mexico. New Mexico Museum of Natural History &
Science Bulletin 17, 17–26.Holser, W. T. & Magaritz, M. (1987). Events near the Permian-
Triassic boundary. Modern Geology 11, 155–180.Holtz, T.R. (1994). The phylogenetic position of Tyrannosauri-
dae: implications for theropod systematics. Journal of Paleontology
68, 1110–1117.Holtz, T. R. (2000). A new phylogeny of the carnivorous
dinosaurs. Gaia 15, 5–61.Holtz, T. R. & Osmolska, H. (2004). Saurischia. In The
Dinosauria, second edition (eds. D. B. Weishampel, P. Dodson andH. Osmolska), pp. 21–24 University of California Press, Berkley.
Holtz, T. R., Molnar, R. E. & Currie, P. J. (2004). BasalTetanurae. In The Dinosauria, second edition (eds. D. B. Weishampel,P. Dodson and H. Osmolska), pp. 71–110 University of Califor-nia Press, Berkley.
Holz, M. & Scherer, C. (2000). Sedimentological and paleon-tological evidence of paleoclimatic change during the SouthBrazilian Triassic: the register of a global trend towardsa humid paleoclimate. In Epicontinental Triassic, Vol. 3 (eds.G. H. Bachmann and I. Lerche). Zentralblatt fur Geologie und
Palaontologie (Teil I) 11–12 1589–1609.Hopson, J. A. (1984). Late Triassic traversodont cynodonts from
Nova Scotia and southern Africa. Palaeontologia Africana 25,181–201.
Hopson, J. A. (1985). Morphology and relationships of Gomphodon-
tosuchus brasiliensis von Huene (Synapsida, Cynodontia, Trity-lodontoidea) from the Triassic of Brazil. Neues Jahrbuch fur Geologie
und Palaontologie Monatshefte 1985, 285–299.Hornung, T. & Brandner, R. (2005). Biochronostratigraphy of
the Reingraben Turnover (Hallstatt Facies Belt): Local blackshale events controlled by regional tectonics, climaticchange andplate tectonics. Facies 51, 460–479.
Hornung, T., Brandner, R., Krystyn, L., Joachimski, M. M.& Keim, L. (2007a). Multistratigraphic constraints on the NWTethyan ‘‘Carnian Crisis’’. In The Global Triassic (eds. S. G. Lucasand J. A. Spielmann). New Mexico Museum of Natural History &
Science Bulletin 41 59–67.Hornung, T., Krystyn, L. & Brandner, R. (2007b). A Tethys-
wide mid-Carnian (Upper Triassic) carbonate productivity crisis:Evidence for the Alpine Reingraben Event from Spiti (IndianHimalaya)? Journal of Asian Earth Sciences 30, 285–302.
Hu, S. (1993). A new Theropoda (Dilophosaurus sinensis sp. nov.)from Yunnan, China. Vertebrata PalAsiatica 31, 65–69.
Huber, P., Lucas, S.G. & Hunt, A.P. (1993). Vertebratebiochronology of the Newark Supergroup Triassic, easternNorth America. In The Nonmarine Triassic (eds. S. G. Lucas andM. Morales). New Mexico Museum Natural History Science Bulletin 3179–186.
Huene, F. von (1908). Die Dinosaurier der europaischen Triasfor-mation, mit Berucksichtigung der ausser–europaischen Vorkom-nisse (part). Geologie und Palaontologie Abhandlungen 6(1, suppl.),345–419.
Huene, F. von (1910). Ein primitiver Dinosaurier aus der mittlerenTrias von Elgin. Geologie und Palaontologie Abhandlungen 8, 317–322.
Huene, F. von (1914). Beitrage zur Geschichte der Archosaurier.Geologie und Palaontologie Abhandlungen (Neue Folge) 13, 1–53.
Huene, F. von (1920). Bemerkungen zur Systematik und Stammes-geschichte einiger Reptilien. Zeitschrift fur Induktive Abstammungslehre
und Verebungslehre 24, 162–166.Huene, F. von (1926). Vollstandige Osteologie eines Plateosauri-
den aus dem schwabischen Keuper. Geologie und Palaeontologie
Abhandlungen (Neue Folge) 15, 139–179.Huene, F. von (1929). Kurze Ubersicht uber die Saurischia und
ihre naturlichen Zusammenhange. Palaontologische Zeitschrift 11,269–273.
Huene, F. von (1932). Die fossile Reptil-Ordnung Saurischia,ihre Entwicklung und Geschichte. Monographien zur Geologie und
Palaontologie 4, 1–361.Huene, F. von (1942). Die fossilen Reptilien des Sudamerikanischen Gond-
wanalandes. C. H. Becksche Verlag, Munich.Huene, F. von (1956). Palaontologie und Phylogenie der niederen Tetrapo-
den. Gustav Fischer Verlag, Jena.Huene, F. von & Stahlecker, R. (1931). Geologische Beobach-
tungen in Rio Grande do Sul. Neues Jahrbuch fur Geologie, Mineralogie
und Palaontologie Abhandlungen Abteilung B 65, 1–82.Hungerbuhler, A. (1998). Taphonomy of the prosauropod
dinosaur Sellosaurus, and its implications for carnivore faunasand feeding habits in the Late Triassic. Palaeogeography, Palaeocli-
matology, Palaeoecology 143, 1–29.Hungerbuhler, A. (2000). Heterodonty in the European phy-
tosaur Nicrosaurus kapffi and its implications for the taxonomicutility and functional morphology of phytosaur dentitions. Journal
of Vertebrate Paleontology 20, 31–48.Hunt, AP. (1989). A new ?ornithischian dinosaur from the Bull
Canyon Formation (Upper Triassic) of east central New Mexico.In: The Dawn of the Age of Dinosaurs in the American Southwest (eds.S. G. Lucas and A. P. Hunt), pp. 355–358 New Mexico Museumof Natural History, Albuquerque.
Hunt, A. P. & Lucas, S. G. (1994). Ornithischian dinosaursfrom the Upper Triassic of the United States. In In the
shadow of the dinosaurs, early Mesozoic tetrapods (eds. N. C. Fraser
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

102 Max C. Langer and others
and H.– D. Sues), pp. 227–241. Cambridge University Press,Cambridge.
Hunt, A. P., Lucas, S. G., Heckert, A. B., Sullivan, R. M. &Lockley, M. G. (1998). Late Triassic dinosaurs from the westernUnited States. Geobios 31, 511–531.
Hunt, A. P., Lucas, S. G., Heckert, A. B. & Zeigler, K.(2002). No significant nonmarine Carnian-Nnorian (LateTriassic) extinction event: evidence from Petrified For-est National Park. http://gsa.confex.com/gsa/2002AM/finalprogram/abstract_42936.htm
Huxley, T. H. (1870). On the Classification of the Dinosauria withobservations on the Dinosauria of the Trias. Quarterly Journal of
the Geological Society 26, 32–51.ICZN (1996). Opinion 1842: Coelurus bauri Cope, 1887 (currently
Coelophysis bauri; Reptilia, Saurischia): lectotype replaced by aneotype. Bulletin of Zoological Nomenclature 53, 142–144.
ICZN (1999). International Code of Zoological Nomenclature, fourth edition.International Trust for Zoological Nomenclature, London.
Irmis, R. B. (2004). First report of Megapnosaurus (Theropoda:Coelophysoidea) from China. PaleoBios 24(3): 11–18.
Irmis, R. B. & Knoll, F. (2008). New ornithischian dinosaurmaterial from the Lower Jurassic Lufeng Formation ofChina. Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen
247, 117–128.Irmis, R. B. & Mundil, R. (2008). New age constraints from the
Chinle Formation revise global comparisons of Late Triassic ver-tebrate assemblages. Journal of Vertebrate Paleontology 28(3, suppl.),95A.
Irmis, R. B., Nesbitt, S. J., Padian, K., Smith, N. D., TurnerA. H., Woody D. & Downs, A. (2007a). A Late Triassicdinosauromorph assemblage from New Mexico and the riseof dinosaurs. Science 317, 358–361.
Irmis, R. B., Parker, W. G., Nesbitt, S. J. & Liu, J. (2007b).Early ornithischian dinosaurs: the Triassic record. Historical Biol-
ogy 19, 3–22.Jalil, N.E. (1996). Les Vertebres permiens et triasiques de la
Formation d’Argana (Haut Atlas occidental): liste fauniquepreliminaire et implications stratigraphiques. In Le Permien et le
Trias du Maroc: etat des Connaissances (ed. F. Medina), pp. 227–250.Universite Semlalia, Marrakech.
Jalil, N. E. & Knoll, F. (2002). Is Azendohsaurus laaroussii (Carnian,Morocco) a dinosaur? Journal of Vertebrate Paleontology 22(3, suppl.),70A.
Jenkins, F. A., Jr., Shubin, N. H., Amaral, W. W., Gatesy,S. M., Schaff, C. R., Clemmensen, L. B., Downs, L. B.,Davidson, A. R., Bonde, N., & Osbraeck, F. (1994). LateTriassic continental vertebrates and depositional environmentsof the Fleming Fjord Formation, Jameson Land, east Greenland.Meddelelser om Grønland, Geoscience 32, 1–25.
Juul,L. (1994). The phylogeny of basal archosaurs. Paleontologia
Africana 31, 1–38.Kent, D.V. & Muttoni, G. (2003). Mobility of Pangea: implica-
tions for Late Paleozoic and Early Mesozoic paleoclimate. In The
Great Rift Valleys of Pangea in Eastern North America, Volume 1. Tectonics,
Structure, and Volcanism (eds. P. M. LeTourneau and P. E. Olsen),pp. 11–20 Columbia University Press, New York.
Kent, D.V., Muttoni, G. & Brack, P. (2006). Reply to ‘Dis-cussion of ‘‘Magnetostratigraphic confirmation of a much fastertempo for sea–level change for the Middle Triassic Latemarplatform carbonates’’ by D.V. Kent, G. Muttoni, and P. Brack
[Earth Planet. Sci. Lett. 228 2004 369–377]’ by L. Hinnov. Earth
and Planetary Science Letters 243, 847–850.Kermack, D. (1984). New prosauropod material from South Wales.
Zoological Journal of the Linnean Society 92, 67–104.King, G. M. (1988). Anomodontia. In Handbuch der Palaoherpetologie,
Teil 17 (ed. O. Kuhn), pp. 1–84. Fischer-Verlag, Stuttgart.King, M. J. & Benton, M. J. (1996). Dinosaurs in the Early and
mid Triassic? - The footprint evidence from Britain. Palaeogeogra-
phy, Palaeoclimatology, Palaeoecology 122, 213–225.Kitching, J. W. & Raath, M. A. (1984). Fossils from the Elliott
and Clarens Formations (Karoo sequence) of the northeasternCape, Orange Free State and Lesotho and a suggested biozona-tion based on tetrapods. Palaeontologia Africana 25, 111–125.
Knoll, F. (2002a). Les Fabrosauridae Galton, 1972 (Dinosauria:Ornithischia): repartition geographique et stratigraphique,systematique et phylogenie. PhD thesis, Museum Nationald’Histoire Naturelle, Paris, France, 243 pp.
Knoll, F. (2002b). Nearly complete skull of Lesothosaurus
(Dinosauria: Ornithischia) from the Upper Elliot Formation(Lower Jurassic: Hettangian) of Lesotho. Journal of Vertebrate Pale-
ontology 22, 238–243.Knoll, F. (2002c). New skull of Lesothosaurus (Dinosauria: Ornithis-
chia) from the Upper Elliot Formation (Lower Jurassic) ofsouthern Africa. Geobios 35, 595–603.
Knoll, F. (2005). The tetrapod fauna of the Upper Elliot andClarens formations in the main Karoo Basin (South Africa andLesotho). Bulletin de la Societe Geologique de France 176, 81–91.
Knoll, F. (2008). On the Procompsognathus postcranium (Late Tri-assic, Germany). Geobios 41, 779–786.
Knoll, F. & Battail, B. (2001). New ornithischian remains fromthe Upper Elliot Formation (Lower Jurassic) of Lesotho and strati-graphical distribution of southern African fabrosaurids. Geobios
34, 415–421.Knoll, F., Padian, K. & de Ricqles, A. (2009). Ontogenetic
change and adult body size of the early ornithischian dinosaurLesothosaurus diagnosticus: Implications for basal ornithischian tax-onomy. Gondwana Research doi: 10.1016/j.gr.2009.03.010.
Kozur, H.W. & Weems, R. (2007). Upper Triassic conchostra-can biostratigraphy of the continental rift basins of EasternNorth America: it’s importance for correlating Newark super-group events with the Germanic basin and the internationalgeologic time scale. In The Global Triassic (eds. S. G. Lucas andJ. A. Spielmann). New Mexico Museum of Natural History & Science
Bulletin 41, 137–188.Krebs, B. (1974). Die Archosaurier. Naturwissenschaften 61, 17–24.Kriwet, J., Rauhut, O. W. M. & Gloy, U. (1997). Microverte-
brate remains (Pisces, Archosauria) from the Middle Jurassic(Bathonian) of southern France. Neues Jahrbuch fur Geologie und
Palaontologie Abhandlungen 206, 1–28.Kutty, T. S., Chatterjee, S., Galton, P. M. & Upchurch, P.
(2007). Basal sauropodomorphs (Dinosauria: Saurischia) fromthe Lower Jurassic of India: their anatomy and relationships.Journal of Paleontology 81, 1218–1240.
Kutty, T. S. & Sengupta, D. P. (1989). The Late Triassic For-mations of the Pranhita-Godavary Valley and their vertebratefaunal succession - a reappraisal. Indian Journal of Earth Sciences 16,189–206.
Langer, M. C. (2002). Is Saturnalia tupiniquim really asauropodomorph? Resumenes del I Congreso Lationoamericano de Pale-
ontologia de Vertebrados, pp. 38–39 Santiago de Chile, Chile.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 103
Langer, M. C. (2003). The sacral and pelvic anatomy of thestem-sauropodomorph Saturnalia tupiniquim (Late Triassic, Brazil).Paleobios 23(2): 1–40.
Langer, M. C. (2004). Basal Saurischia. In The Dinosauria, sec-
ond edition (eds. D. B. Weishampel, P. Dodson and H. Osmolska),pp. 25–46 University of California Press, Berkley.
Langer, M. C. (2005a). Studies on continental Late Triassic tetra-pod biochronology. I. The type locality of Saturnalia tupiniqium
and the faunal succesion in south Brazil. Journal of South American
Earth Sciences 19, 205–218.Langer, M.C. (2005b). Studies on continental Late Triassic
tetrapod biochronology. II. The Ischigualastian and a Car-nian global correlation. Journal of South American Earth Sciences
19, 219–239.Langer, M. C., Abdala, F., Richter, M. & Benton, M. J.
(1999). A sauropodomorph dinosaur from the Upper Triassic(Carnian) of souther Brazil. Comptes Rendus de l’Academie des Sciences
IIA 329, 511–517.Langer, M. C. & Benton, M. J. (2006). Early dinosaurs:
a phylogenetic study. Journal of Systematic Palaeontology
4, 309–358.Langer, M.C., Bittencourt, J.S. & Schultz, C.L. (2007a).
The inclusivity and phylogenetic position of Guaibasaurus can-
delariensis: a basal dinosaur from the Late Triassic of Brazil.Journal of Vertebrate Paleontology 27(3, suppl.), 103A–104A.
Langer, M. C., Franca, M. A. G., & Gabriel, S. (2007b).The pectoral girdle and forelimb anatomy of the stem-sauropodomorph Saturnalia tupiniquim (Late Triassic, Brazil). InEvolution and palaeobiology of early sauropodomorph dinosaurs (eds.P. M. Barrett and D. J. Batten). Special Papers in Palaeontology 77,113–137.
Langer, M. C., Ribeiro, A. M., Schultz, C. L., andFerigolo, J. (2007c). The continental tetrapod–bearing Tri-assic of South Brazil. In The Global Triassic (eds. S. G. Lucas andJ. A. Spielmann). New Mexico Museum of Natural History & Science
Bulletin 41 201–218.Leal, L. A., Azevedo, S. A. K., Kellner, A. W. A. & Da
Rosa, A. A. S. (2004). A new early dinosaur (Sauropodomor-pha) from the Caturrita Formation (Late Triassic), Parana Basin,Brazil. Zootaxa 690, 1–24.
LeTourneau, P. M. & Olsen, P. E. (eds.) (2003). The Great Rift
Valleys of Pangea in Eastern North America, vol. 1-2. Columbia Uni-versity Press, New York.
Lloyd, G.T., Davis, K.E., Pisani, D., Tarver, J.E., Ruta, R.,Sakamoto, M., Hone, D.W.E., Jennings, R. & Benton, M.J.(2008). Dinosaurs and the CretaceousTerrestrial Revolution. Pro-
ceedings of the Royal Society of London B 275, 2483–2490.Lockley, M. & Meyer, C. (2000). Dinosaur tracks and other fossil
footprints of Europe. Columbia University Press, New York.Lockley, M.G., Lucas, S.G., Hunt, A.P. & Gaston, R. (2004).
Ichnofaunas from the Triassic-Jurassic boundary sequences ofthe Gateway area, Western Colorado: implications for fau-nal composition and correlations with other areas. Ichnos
11, 89–102.Long, J. & Molnar, R. E. (1998). A new Jurassic theropod
dinosaur from Western Australia. Records of the Western Australian
Museum 19, 121–129.Long, R. A. & Murry, P. A. (1995). Late Triassic (Carnian and
Norian) tetrapods from the Southwestern United States. New
Mexico Museum of Natural History & Science Bulletin 4, 1–254.
Lucas, S. G. (1998). Global Triassic tetrapod biostratigraphy andbiochronology. Palaeogeography, Palaeoclimatolology, Palaeoecology 143,347–384.
Lucas, S. G. (2007). Tetrapod footprint biostratigraphy andbiochronology. Ichnos 14, 5–38.
Lucas, S. G. & Hancox, P. J. (2001). Tetrapod-based correlationof the nonmarine Upper Triassic of southern Africa. Albertiana
25, 5–9.Lucas, S. G. & Luo, Z. (1993). Adelobasileus from the Upper Tri-
assic of West Texas: the oldest mammal. Journal of Vertebrate
Paleontology 13, 309–334.Lucas, S. G. & Tanner, L. H. (2005). Late Triassic extinction
events. Albertiana 31, 31–40.Maidment, S. C. R. & Wei, G. (2006). A review of the
Late Jurassic stegosaurs (Dinosauria, Stegosauria) fromthe People’s Republic of China. Geological Magazine
143, 621–634.Marsh, O. C. (1878). Principal characters of American Jurassic
dinosaurs I. American Journal of Science 16, 411–416.Marsh, O. C. (1881). Principal characters of American Jurassic
dinosaurs V. American Journal of Science 16, 411–416.Marsh, O. C. (1882). Classification of the Dinosauria. American
Journal of Sciences 23, 81–86.Marsh, O. C. (1894). The typical Ornithopoda of the American
Jurassic. American Journal of Sciences 48, 85–90.Marsicano, C. A. (1999). Chigutisaurid amphibians from the
upper Triassic of Argentina and their phylogenetic relationships.Palaeontology 42, 1–21.
Marsicano, C. A., Domnanovich, N. S. & Mancuso, A. C.(2007). Dinosaur origins: evidence from the footprint record.Historical Biology 19, 83–91.
Martill, D. M., Batten, D. J. & Loydell, D. K. (2000). A newspecimen of the thyreophoran dinosaur cf. Scelidosaurus with softtissue preservation. Palaeontology 43, 549–559.
Martınez, R. N. (2009). Adeopapposaurus mognai, gen. et sp. nov.(Dinosauria: Sauropodomorpha), with comments on adaptationsof basal Sauropodomorpha. Journal of Vertebrate Paleontology 29,142–164.
Martinez, R. N. & Alcober, O. A. (2007). Um nuevo her-rerasaurido (Dinosauria, Saurischia) de la FormacionIschigualasto (Triasico Superior, Carniano). Ameghiniana
44(4, suppl.), 28R–29R.Martinez, R. N. & Alcober, O. A. (2009). A basal
sauropodomorph (Dinosauria: Saurischia) from the IschigualastoFormation (Triassic, Carnian) and the early evolution ofSauropodomorpha. PLoS ONE 4, 1–12.
Martinez, R. N., May, C. L. & Forster, C. (1996). A newcarnivorous cynodont from the Ischigualasto Formation (LateTriassic) with comments on eucynodont phylogeny. Journal of
Vertebrate Paleontology 16, 271–284.Martinez, R. N., Sereno, P. C. & Alcober, O. A. (2008). A new
basal theropod from the Ischigualasto Formation of San Juanprovince, Argentina. In: Libro de Resumenes, III Congreso Latinoamer-
icano de Paleontologia de Vertebrados (eds. J. O. Calvo, R. J. Valieri,J. D. Porfiri and D. dos Santos), p. 153. Universidade Nacionaldel Comahue, Neuquen, Argentina.
Maryanska, T. (1977). Ankylosauridae (Dinosauria) from Mon-golia. Palaeontologia Polonica 37, 85–151.
Maryanska, T. & Osmolska, H. (1985). On ornithischian phy-logeny. Acta Palaeontologica Polonica 30, 137–150.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

104 Max C. Langer and others
Marzoli, A., Renne, P. R., Piccirillo, E. M., Ernesto, M.,Bellieni, G. & de Min, A. (1999). Extensive 200-million-year-old continental flood basalts of the central Atlantic Magmaticprovince. Science 284, 616–618.
Matthew, W. D. & Brown, B. (1922). The family Deinodon-tidae, with notice of a new genus from the Cretaceousof Alberta. Bulletin of the American Museum of Natural History
46, 367–385.Mayr, G., Stefan Peters, D., Plodowski, G. & Vogel, O.
(2002). Bristle-like integumentary structures at the tailof the horned dinosaur Psittacosaurus. Naturwissenschaften
89, 361–365.McElwain, J. C., Beerling, D. J. & Woodward, F. I. (1999).
Fossil plants and global warming at the Triassic-Jurassic bound-ary. Science 285, 1386–1390.
Melchor, R. N. & de Valais, S. (2006). A review of Triassic tetra-pod track assemblages from Agentina. Palaeontology 49, 355–379.
Meyer, H. von (1837). Mitteilung an Prof. Bronn (Plateosaurus
engelhardti). Neues Jarhbuch fur Mineralogie, Geognoise, Geologie und
Petrefakten-Kunde 1837, 1–317.Milner, A. R. (1993). Amphibian-grade Tetrapoda. In The Fossil
Record 2 (ed. M. J. Benton), pp. 663–677 Chapman and Hall,London.
Monbaron, M., Russell, D. A. & Taquet, P. (1999). Atlasaurus
imelakei n. g., n. sp., a brachiosaurid-like sauropod from the Mid-dle Jurassic of Morocco. Comptes Rendus de l’Academie des Sciences de
Paris IIA 329, 519–526.Moody, J. M. (1997). Theropod teeth from the Jurassic of
Venezuela. Boletın de la Sociedad Venezolana de Geologos 22(2): 37–42.Moratalla, J. J. & Powell, J. E. (1994). Dinosaur nesting pat-
terns. In Dinosaur eggs and babies (eds. K. Carpenter, K. F. Hirsch,and J. R. Horner), pp. 37–46 Cambridge University Press, NewYork.
Moser, M. (2003). Plateosaurus engelhardti Meyer, 1837 (Dinosauria:Sauropodomorpha) aus dem Feuerletten (Mittelkeuper; Ober-trias) von Bavaria. Zitteliana 24, 3–186.
Munter, R. C. & Clark, J. M. (2006). Theropod dinosaurs fromthe Early Jurassic of Huizachal Canyon, Mexico. In Amniote
paleobiology: perspectives on the evolution of mammals, birds, and reptiles
(eds. M. T. Carrano, T. J. Gaudin, R. W. Blob and J. R. Wible),pp. 53–75 University of Chicago Press, Chicago.
Murry, P.A. & Long, R.A. (1989). Geology and paleontology ofthe Chinle Formation, Petrified Forest National Park and vicinity,Arizona and a discussion of vertebrate fossils of the southwesternUpper Triassic. In Dawn of the Age of Dinosaurs in the American South-
west (eds. S. G. Lucas and A. P. Hunt), pp. 29–64 New MexicoMuseum of Natural History, Albuquerque.
Muttoni, G., Kent, D. V., Di Stefano, P., Gullo, M.,Nicora, A., Tait, J. & Lowrie, W. (2001). Magnetostratig-raphy and biostratigraphy of the Carnian/Norian bound-ary interval from the Pizzo Mondello section (SicaniMountains, Sicily). Palaeogeography, Palaeoclimatology, Palaeoecology
166, 383–399.Muttoni, G., Kent, D.V., Olsen, P.E., Di Stefano, P.,
Lowrie, W., Bernasconi, S.M. & Hernandez, F.M. (2004).Tethyan magnetostratigraphy from Pizzo Mondello (Sicily)and correlation to the Late Triassic Newark astrochrono-logical polarity time scale. Geological Society of America Bulletin
116, 1043–1058.
Nath, T. T., Yadagiri, P. & Moitra, A. K. (2002). First recordof armoured dinosaur from the Lower Jurassic Kota Forma-tion, Pranhita-Godavari Valley, Andrah Pradesh. Journal of the
Geological Society of India 59, 575–577.Nesbitt, S. J. (2003). Arizonasaurus and its implications for
archosaur divergence. Proceedings of the Royal Society of London B
270(suppl.), S234–S237.Nesbitt, S. J. (2007). The anatomy of Effigia okeeffeae (Archosauria,
Suchia), theropod-like convergence, and the distribution ofrelated taxa. Bulletin of the American Museum of Natural History
302, 1–84.Nesbitt, S.J., Turner, A.H., Erickson, G.M., & Norell, M.A.
(2006). Prey choice and cannibalistic behaviour in the theropodCoelophysis. Biology Letters 2, 611–614.
Nesbitt, S. J. & Chatterjee, S. (2008). Late Triassic dinosauri-forms from the Post Quarry and surrounding areas, west Texas,U.S.A. Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen 249,143–156.
Nesbitt, S. J., Irmis, R. B. & Parker, W. G. (2007). A critical re-evaluation of the Late Triassic dinosaur taxa of North America.Journal of Systematic Palaeontology 5, 209–243.
Nesbitt, S. J., Irmis, R. B., Parker, W. G., Smith, N. D.,Turner, A. H. & Rowe, T. (2009). Hindlimb osteology anddistribution of basal dinosauromorphs from the Late Triassic ofNorth America. Journal of Vertebrate Palaeontology 29, 498–516.
Nopcsa, F. (1915). Die Dinosaurier der siebenburgischen Lan-desteile Ungarns. Mitteilung des Jahrbuches der Konigliche Ungarische.
Geologische Reichsanstalt 23, 1–26.Nopcsa, F. (1923). Die Familien der Reptilien. Fortschritte der Geologie
und Paleontologie 2, 1–210.Nopcsa, F. (1928). The genera of reptiles. Palaeobiologica 1,
163–188.Norman, D. B. (1984). A systematic reappraisal of the reptile order
Ornithischia. In Third Symposium on Mesozoic Terrestrial Ecosys-
tems, Short Papers (eds. W.-E. Reif and F. Westphal), pp. 157–162Attempto Verlag, Tubingen, Germany.
Norman, D. B. (1990). Problematic Theropoda: ‘‘Coelurosaurs’’.In The Dinosauria (eds. D. B. Weishampel, P. Dodson andH. Osmolska), pp. 280–305 University of California Press,Berkley.
Norman, D. B. (2001). Scelidosaurus, the earliest complete dinosaur.In The Armored Dinosaurs (ed. K. Carpenter), pp 3–24. IndianaUniversity Press, Bloomington.
Norman, D. B. & Barrett, P. M. (2002). Ornithischian dinosaursfrom the Lower Cretaceous (Berriasian) of England. In Life and
Environments in Purbeck Times (eds. A. Milner and D. J. Batten).Special Papers in Palaeontology 68, 161–189.
Norman, D. B., Witmer, L. M. & Weishampel, D. B. (2004a).Basal Ornithischia. The Dinosauria, second edition (eds. D. B.Weishampel, P. Dodson and H. Osmolska), pp. 325–334 Uni-versity of California Press, Berkley.
Norman, D. B., Witmer, L. M. & Weishampel, D. B. (2004b).Basal Ornithopoda. The Dinosauria, second edition (eds.D. B. Weishampel, P. Dodson and H. Osmolska), pp. 393–412University of California Press, Berkley.
Norman, D. B., Witmer, L. M. & Weishampel, D. B. (2004c).Basal Thyreophora. The Dinosauria, second edition (eds.D. B. Weishampel, P. Dodson and H. Osmolska), pp. 335–342University of California Press, Berkley.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 105
Norman, D. B., Butler, R. J. & Maidment, S. C. R. (2007).Reconsidering the status and affinities of the ornithischiandinosaur Tatisaurus oehleri Simmons, 1965. Zoological Journal of
the Linnean Society 150, 865–874.Novas, F. E. (1986). Un probable teropodo (Saurischia) de la For-
macion Ischigualasto (Triasico superior), San Juan, Argentina. InIV Congreso Argentino de Paleontologıa y Estratigrafıa. 2: 1–6 Mendoza,Argentina.
Novas, F. E. (1989). The tibia and tarsus in Herrerasauridae(Dinosauria, incertae sedis) and the origin and evolution of thedinosaurian tarsus. Journal of Paleontology 63, 677–690.
Novas, F. E. (1992a). La evolucion de los dinosaurios carnıvoros. InLos dinosaurios y su entorno biotico (eds. J. L. Sanz and A. Buscalioni),pp. 125–163 Actas II Curso dePaleontologıca en Cuenca. Ayun-tamiento deCuenca, Spain.
Novas, F. E. (1992b). Phylogenetic relationships of the basaldinosaurs, the Herrerasauridae. Palaeontology 35, 51–62.
Novas, F. E. (1993). New information on the systematics andpostcranial skeleton of Herrerasaurus ischigualastensis (Theropoda:Herrerasauridae) from the Ischigualasto Formation (Upper Tri-assic) of Argentina. Journal of Vertebrate Paleontology 13, 400–423.
Novas, F. E. (1996). Dinosaur monophyly. Journal of Vertebrate Pale-
ontology 16, 723–741.Novas, F. E. (1997a). Herrerasauridae. In Encyclopedia of Dinosaurs
(eds. P. J. Currie and K. Padian), pp. 303–311 Academic Press,San Diego.
Novas, F. E. (1997b). South American Dinosaurs. In Encyclope-
dia of Dinosaurs (eds. P. J. Currie and K. Padian), pp. 678–689Academic Press, San Diego.
Novas, F. E. & Ezcurra, M. D. (2005). The evolutionary radia-tion of Triassic dinosauriforms. Ameghiniana 42, 36–37R.
Novas, F. E., Chatterjee, S., Haro, J. A. & Kutty, T. S.(2006). New primitive dinosaurs for the Late Triassic of India.Ameghiniana 43, 48R.
Ochev, V.G. & Shishkin, M. A. (1989). On the principles ofglobal correlation of the continental Triassic on the tetrapods.Acta Palaeontologica Polonica 34, 149–173.
Ogg, J. G. (2004). Status of division of the International GeologicalTime Scale. Lethaia 37, 183–199.
Ogg, J.G., Ogg, G., & Gradstein, F. M. (2008). The Concise Geo-
logic Time scale. Cambridge University Press, Cambridge.Olsen, P. E & Galton, P. M. (1984). A review of the reptile and
amphibian assemblages from the Stormberg Group of southernAfrica with special emphasis on the footprints and the age of theStormberg. Palaeontologia Africana 25, 87–110.
Olsen, P. E., Kent, D. V., Sues, H.-D., Koeberl, C.,Huber, H., Montanari, A., Rainforth, E. C., Fowell,S. J., Szajna, M. J. & Hartline, B. W. (2002). Ascent ofdinosaurs linked to an iridium anomaly at the Triassic-Jurassicboundary. Science 296, 1305–1307.
Olsen, P. E., Schlische, R.W. & Gore, P. J. W. (1989). Field
Guide to the Tectonics, stratigraphy, sedimentology, and paleontology of the
Newark Supergroup, eastern North America. International GeologicalCongress, Guidebooks for Field Trips.
Olsen, P.E., Shubin, N.H. & Anders, M.H. (1987). New EarlyJurassic tetrapod assemblages constrain Triassic-Jurassic tetrapodextinctionevent. Science 237, 1025–1029.
Olsen, P.E. & Sues, H.-D. (1986). Correlation of continental LateTriassicand Early Jurassic sediments, and patterns of the Tri-assic–Jurassic tetrapod transition. In The beginning of the age of
dinosaurs (ed. K. Padian), pp. 321–351 Cambridge UniversityPress, Cambridge.
Olshevsky, G. (2000). An annotated checklist of dinosaur speciesby continent. Mesozoic Meanderings 3, 1–157.
Olshevsky, G. (2007). The dinosaur genera list. http://www.polychora.com/dinolist.html
Ostrom, J. H. (1978). The osteology of Compsognathus longipes Wag-ner. Zitteliana 4, 73–118.
Owen, R. (1842). Report on British fossil reptiles. Part II.Reports of the British Association for the Advancement of Science
11, 60–204.Owen, R. (1861). A monograph of the fossil Reptilla of the Lias For-
mations. I Scelidosaurus harisonii. Palaeontographical Society Monographs
13, 1–14.Owen, R. (1863). A monograph of the fossil Reptilia of the Liassic
formations. Part II. Scelidosaurus harrisonii. Palaeontographical Society
Monographs 14, 1–26.Padian, K. (1986). On the type material of Coelophysis Cope
(Saurischia: Theropoda) and a new specimen from the Petri-fied Forest of Arizona (Late Triassic: Chinle Formation). In The
beginning of the age of dinosaurs (ed. K. Padian), pp. 45–60 CambridgeUniversity Press, Cambridge.
Padian, K. (1989). Presence of the dinosaur Scelidosaurus indicatesJurassic age for the Kayenta Formation (Glen Canyon Group,northern Arizona). Geology 17, 438–441.
Padian, K. (1997a). Dinosauria: Definition. In Encyclopedia of
Dinosaurs (eds. P. J. Currie and K. Padian), pp. 175–179 Aca-demic Press, San Diego.
Padian, K. (1997b). Pectoral girdle. In Encyclopedia of Dinosaurs
(eds. P. J. Currie and K. Padian), pp. 530–536 Academic Press,San Diego.
Padian, K. (1997c). Quadrupedality. In Encyclopedia of Dinosaurs
(eds. P. J. Currie and K. Padian), pp. 619–626 Academic Press,San Diego.
Padian, K. (1997d). Saurischia. In Encyclopedia of Dinosaurs (eds.P. J. Currie and K. Padian), pp. 647–653 Academic Press,San Diego.
Padian, K. & May, C. L. (1993). The earliest dinosaurs. New Mexico
Museum of Natural History & Science Bulletin 3, 379–381.Padian, K., Hutchinson, J. R. & Holtz, T. R. (1999). Phylo-
genetic definitions and nomenclature of the major taxonomiccategories of the carnivorous Dinosauria (Theropoda). Journal of
Vertebrate Paleontology 19, 69–80.Parker, W. G., Irmis, R. B., Nesbitt, S. J., Martz, J. W. &
Browne, L. S. (2005). The Late Triassic pseudosuchian Revuel-
tosaurus callenderi and its implications for the diversity of earlyornithischian dinosaurs. Proceedings of the Royal Society of London B
272, 963–969.Parrish, J. M. (1986). Locomotor adaptations in the hindlimb and
pelvis of the Thecodontia. Hunteria 1, 1–36.Parrish, J. M. (1997). Archosauria. In Encyclopedia of Dinosaurs
(eds. P. J. Currie and K. Padian), pp. 20–24 Academic Press,San Diego.
Parrish, J. T. (1993). Climate of the supercontinent Pangea. Jour-
nal of Geology 101, 215–233.Paul, G. S. (1984). The segnosaurian dinosaurs: relics of the
prosauropod-ornithischian transition? Journal of Vertebrate Pale-
ontology 4, 507–515.Paul, G. S. (1988). Predatory Dinosaurs of the World: A Complete Illus-
trated Guide. Simon and Schuster, New York.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

106 Max C. Langer and others
Paul, G. S. (2002). Dinosaurs of the air. The Johns Hopkins UniversityPress, Baltimore.
Peng, G. (1997). Fabrosauridae. In Encyclopedia of Dinosaurs (eds.P. J. Currie and K. Padian), pp. 237–240 Academic Press, SanDiego.
Peng, G., Ye, Y., Yuhui, G., Shu, C. & Jiang, S. (2005). [Jurassic
dinosaur faunas in Zigong]. Zigong Dinosaur Museum, Zigong. (inChinese).
Peters, D. (2000). A reexamination of four prolacertiforms withimplications for pterosaur phylogenesis. Rivista Italiana di Paleon-
tologia e Stratigrafia 106, 293–336.Plumptre, A. J. (1996). Modelling the impact of large herbivores
on the food supply of mountain gorillas and implications formanagement. Biological Conservation 75, 147–155.
Pol, D. (2004). Phylogenetic relationshps of basal Sauropodomorpha. Ph.D
thesis, Columbia University, New York, USA 303 pp.Pol, D. & Garrido, A. C. (2007). Um nuevo dinosaurio
sauropodomorfo basal de la Formacion Las Leoneras (JurasicoInferior), Chubut, Argentina. Ameghiniana 44(4, suppl.), 64R.
Pol, D. & Powell, J. E. (2005). Anatomy and phylogenetic rela-tionships of Mussaurus patagonicus (Dinosauria, Sauropodomorpha)from the Late Triassic of Patagonia. In Boletim de Resumos
do II Congresso Latino-americano de Paleontologia de Vertebrados
(eds. A. W. A. Kellner, D. D. R. Henriques & T. Rodrigues),pp. 208–209 Museu Nacional, Rio de Janeiro, Serie Livros 12.
Pol, D. & Powell, J. E. (2007a). New information on Lessem-
saurus sauropoides (Dinosauria: Sauropodomorpha) from the UpperTriassic of Argentina. In Evolution and palaeobiology of early
sauropodomorph dinosaurs (eds. P. M. Barrett and D. J. Batten). Spe-
cial Papers in Palaeontology 77, 223–243.Pol, D. & Powell, J. E. (2007b). Skull anatomy of Mussaurus patag-
onicus (Dinosauria: Sauropodomorpha) from the Late Triassic ofPatagonia. Historical Biology 19, 125–144.
Popper, K. (1959). The Logic of Scientific Discovery. Hutchinson, Lon-don.
Price, L.I. (1946). Sobre um novo pseudossuquio do TriassicoSupeiror do Rio Grande do Sul. Boletim da Divisao de Geologia e
Mineralogia 120, 1–38.Prochnow, S. J., Nordta, L. C., Atchleya, S. C., Hudecc,
M. R. (2006). Multi-proxy paleosol evidence for Middle andLate Triassic climate trends in eastern Utah. Palaeogeography,
Palaeoclimatology, Palaeoecology 232, 53–72.Raath, M. (1969). A new coelurosaurian dinosaur from the Forest
Sandstone of Rhodesia. Arnoldia 4, 1–25.Raath, M. (1980). The theropod dinosaur Syntarsus (Saurischia:
Podokesauridae) discovered in South Africa. South African Journal
of Science 76, 375–376.Raath, M. (1996). Earliest evidence of dinosaurs from central
Gondwana. Memoirs of the Queensland Museum 39, 703–709.Raath, M. A., Kitching, J. W., Shone, R. W. & Rossouw,
G. J. (1990). Dinosaur tracks in Triassic Molteno sediments:the earliest evidence of dinosaurs in South Africa. Palaeontologia
Africana 27, 89–95.Rainforth, E. C. (2002). The pangean ichnites Otozum and Pseu-
dotetrasauropus: an example of relationships among and betweentetrapod ichnotaxa skeletal taxa. Journal of Vertebrate Paleontology
26(3, suppl.), 98A.Rainforth, E. C. (2003). Revision and re-evaluation of the
Early Jurassic dinosaur ichnogenus Otozoum. Palaeontology
46, 803–838.
Rauhut, O. W. M. (2003). The interrelationships and evolution ofbasal theropod dinosaurs. Special Papers in Palaeontology 69, 1–213.
Rauhut, O. W. M. (2005a). Osteology and relationships of a newtheropod dinosaur from the Middle Jurassic of Patagonia. Palaeon-
tology 48, 87–110.Rauhut, O. (2005b). Saurischian diversity and radiations in the
early Mesozoic. In Boletim de Resumos do II Congresso Latino-
americano de Paleontologia de Vertebrados (eds. A. W. A. Kellner,D. D. R. Henriques & T. Rodrigues), p. 216. Museu Nacional,Rio de Janeiro, Serie Livros 12.
Rauhut, O. W. M. & Hungerbuhler, A. (2000). A review ofEuropean Triassic theropods. Gaia 15, 75–88.
Rauhut, O. W. M. & Lopez-Arbarello, A. (2008). Archosaurevolution during the Jurassic: a southern perspective. Revista de la
Asociacion Geologica Argentina 63, 557–585.Raup, D. M. & Sepkoski Jr., J. J. (1982). Mass extinction in the
marine fossil record. Science 215, 1501–1503.Ray, S. & Chinsamy, A. (2002). A theropod tooth from the Late
Triassic of southern Africa. Journal of Bioscience 27, 295–298.Regal, P. (1975). The evolutionary origin of feathers. Quarterly
Review of Biology 50, 35–66.Regal, P. (1985). Common Sense and reconstructions ofthe biol-
ogy of fossils: Archaeopteryx and feathers. In The beginnings of birds
(eds. M. K. Hecht, J. H. Ostrom, G. Viohl, & P. Wellnhofer),pp. 67–74 Freunde des Jura-Museums, Eichstatt.
Reig, O. A. (1961). Acerca de la posicion sistematica de la familiaRaiusuchidae y del genero (Reptilia, Thecodontia). Revista del
Museo Municipal de Ciencias Naturales y Tradicional de La Plata 1(3),73–113.
Reig, O. A. (1963). La presencia de dinosaurios saurisquios en los‘‘Estratos de Ischigualasto’’ (Mesotriasico Superior) de las provin-cias de San Juan y La Rioja (Republica Argentina). Ameghiniana
3, 3–20.Reig, O. A. (1970). The Proterosuchia and the early evolution of
the archosaurs, an essay about the origin of a major taxon. Bulletin
of the Museum of Comparative Zoology 139, 229–292.Reisz, R. R., Scott, D., Sues, H-D., Evans, D. C. & Raath,
M. A. (2005). Embryos of an Early Jurassic prosauropod dinosaurand their evolutionary significance. Science 309, 761–764.
Remane, J., Cita, M. B., Decourt, J., Bouysse, P., Repetto,F.L. & Faure-Muret, A. (2000). International stratigraphicchart. In Palaeobiology II (eds. D.E.G. Briggs and P. R. Crowther).Blackwell Science, London.
Remes, K. & Rauhut, O. (2005). The oldest Indian dinosaurAlwalkeria maleriensis Chaterjee revised: a chimera includingremains of a basal saurischian. In Boletim de Resumos do II Congresso
Latino-americano de Paleontologia de Vertebrados (eds. A. W. A. Kellner,D. D. R. Henriques & T. Rodrigues), p. 218. Museu Nacional,Rio de Janeiro, Serie Livros 12.
Rogers, R., Arcucci, A.B., Abdala, F., Sereno, P. C.,Forster, C. A. & May, C. L. (2001). Paleoenvironment andtaphonomy of the Chanares Formation tetrapod assemblage(Middle Triassic), northwestern Argentina: spectacular preserva-tion in volcanogenic concretions. Palaios 16, 461–481.
Rogers, R. R., Swisher III, C. C., Sereno, P. C., Forster,C. A. & Monetta, A. M. (1993). The Ischigualasto tetrapodassemblage (Late Triassic) and 40Ar/39Ar calibration of dinosaurorigins. Science 260, 794–797.
Romer, A. S. (1956). Osteology of the Reptiles. University of ChicagoPress, Chicago.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 107
Romer, A. S. (1960). Vertebrate-bearing continental Triassic stratain Mendoza region, Argentina. Bulletin of the Geological Society of
America 71, 1279–1293.Romer, A. S. (1962). La evolucion explosiva de los Rhynchosaurios
del Triasico. Revista del Museo Argentino de Ciencias Naturales, Ciencias
Zoologicas 8(1), 1–14.Romer, A. S. (1966). Vertebrate Paleontology. University of Chicago
Press, Chicago. p. 468.Romer, A.S. (1971). The Chanares (Argentina) Triassic reptile
fauna. XI. Two but new incompletely known long-limbed Pseu-dosuchians. Breviora 378, 1–10.
Romer, A. S. (1972a). The Chanares (Argentina) Triassic reptilefauna. XIV. Lewisuchus admixtus a further thecodont from theChamares Bed. Breviora 390, 1–13.
Romer, A. S. (1972b). The Chanares (Argentina) Triassic reptilefauna. XV. Further remains of the thecodonts Lagerpeton andLagosuchus. Breviora 390, 1–7.
Rosenbaum, J. N. & Padian, K. (2000). New material of thebasal thyreophoran Scutellosaurus lawleri from the KayentaFormation (Lower Jurassic) of Arizona. PaleoBios
20(1), 13–23.Ross, C.A., Baud, A. & Manning, M. (1994). Project Pangea
time scale. In Pangea: Paleoclimate, Tectonics, and Sedimentation during
Accretion, Zenith, and Breakup of a Supercontinent (ed. G. D. Klein).Geological Society of America Special Papers 288.
Rowe, T. (1989). A new species of the theropod dinosaur Syntarsus
from the Early Jurassic Kayenta Formation of Arizona. Journal of
Vertebrate Paleontology 9, 125–136.Rowe, T. & Gauthier, J. A. (1990). Ceratosauria. The Dinosauria
(eds. D. B. Weishampel, P. Dodson and H. Osmolska), pp.151–168 University of California Press, Berkley.
Ruiz-Omenaca, J. I., Suberbiola, X. P. & Galton, P.M.(2005). Callovosaurus leedsi, the earliest dryosaurid dinosaur(Ornithischia: Euornithopoda) from the Middle Jurassic of Eng-land. In Horns and Beaks: Ceratopsian and ornithopod dinosaurs (ed.K. Carpenter), pp. 3–16 Indiana University Press, Bloomington.
Russell, D.E., Rivas, O. O., Battail, B. & Russell, D.A.(1992). Decouverte de vertebres fossils dans la Formation deLa Quinta, Jurassique du Venezuela Occidental. Comptes Rendus
de l’Academie des Sciences IIA 314, 1247–1252.Salgado, L., Coria, R. A. & Calvo, J. O. (1997). Evolution of
titanosaurid sauropods. I. Phylogenetic analysis based on thepostcranial evidence. Ameghiniana 34, 3–32.
Santa Luca, A. P. (1980). The postcranial skeleton of Heterodon-
tosaurus tucki (Reptilia, Ornithischia) from the Stormberg of SouthAfrica. Annals of South African Museum 79, 159–211.
Santa Luca, A. P. (1984). Postcranial remains of Fabrosauridae(Reptilia: Ornithischia) from the Stormberg of South Africa.Palaeontologia Africana 25, 151–180.
Schafer, P. & Fois, E. (1987). Systematics and evolution of Tri-assic Bryozoa. Geologica et Palaeontologica 21, 173–225.
Schoene, B., Crowley, J. L., Condon, D. J., Schmitz, M. D.& Bowring, S. A. (2006). Reassessing the uranium decay con-stants for geochronology using ID-TIMS U Pb data. Geochimica et
Cosmochimica Acta 70, 426–445.Schwartz, H. L. & Gillette, D. D. (1994). Geology and taphon-
omy of the Coelophysis quarry, Upper Triassic Chinle Formation,Ghost Ranch, New Mexico. Journal of Paleontology 68, 1118–1130.
Scotese, C. R. (2002). Paleomap Project. http://www.scotese.com.
Seebacher, F. (2001). A new method to calculate allometric length-mass relationships of dinosaurs. Journal of Vertebrate Paleontology 21,51–60.
Seeley, H. G. (1888). On the classification of the fossil animalscommonly named Dinosauria. Proceedings of the Royal Society of
London 43, 165–171.Sengupta, D. P. (2003). Triassic temnospondyls of the Pranhita-
Godavari Basin, India. Journal of Asian Earth Sciences 21, 655–662.Senter, P. (2005). Phylogenetic taxonomy and the names of the
major archosaurian (Rreptilia) clades. PaleoBios 25, 1–7.Sepkoski Jr., J. J. (1982). Mass extinctions in the Phanerozoic
oceans: a review. In Geological Implicationsof Impacts of Large Asteroids
and Comets on theEarth (eds. L. T. Silver and P. H. Schultz). The
Geological Society of America Special Paper 190, 283–289.Sepkoski Jr., J. J. (1990). The taxonomic structure of periodic
extinction. In Global Catastrophes in Earth History (eds. L. Sharptonand P. D. Ward). The Geological Society of America SpecialPaper 247 33–44.
Sereno, P. C. (1984). The phylogeny of the Ornithischia: a reap-praisal. In Third Symposium on Mesozoic Terrestrial Ecosystems, Short
Papers (eds. W.-E. Reif and F. Westphal), pp. 219–226 AttemptoVerlag, Tubingen, Germany.
Sereno, P. C. (1986). Phylogeny of the bird-hipped dinosaurs.National Geographic Research 2, 234–256.
Sereno, P. C. (1989). Prosauropod monophyly and basalsauropodomorph phylogeny. Jounal of Vertebrate Paleontology 9(3,suppl.), 38.
Sereno, P. C. (1991a). Basal archosaurs: phylogenetic relation-ships and functional implications. Memoir of the Society of Vertebrate
Paleontology 2, 1–53.Sereno P. C. (1991b). Lesothosaurus, ‘‘fabrosaurids,’’ and the early
evolution of Ornithischia. Jounal of Vertebrate Paleontology 11,168–197.
Sereno, P. C. (1993). The pectoral girdle and forelimb of thebasal theropod Herrerasaurus ischigualastensis. Jounal of Vertebrate
Paleontology 13, 425–450.Sereno, P. C. (1997). The origin and evolution of dinosaurs. Annual
Review of Earth & Planetary Sciences 25, 435–489.Sereno, P. C. (1998). A rationale for phylogenetic definitions, with
application to the higher-level taxonomy of Dinosauria. Neues
Jahrbuch fur Geologie und Palaontologie Abhandlungen 210, 41–83.Sereno, P. C. (1999). The evolution of dinosaurs. Science 284,
2137–2147.Sereno, P. C. (2005). TaxonSearch: Database for Suprageneric Taxa &
Phylogenetic Definitions. http://www.taxonsearch.org/Sereno, P.C. (2007a). Basal Sauropodormorpha: historical and
recent phylogenetic hypothesis, with comments on Ammosaurus
major (Marsh, 1889). In Evolution and palaeobiology of early
sauropodomorph dinosaurs (eds. P. M. Barrett and D. J. Batten). Spe-
cial Papers in Palaeontology 77, 261–289.Sereno, P.C. (2007b). The phylogenetic relationships of
early dinosaurs: a comparative report. Historical Biology
19, 145–155.Sereno, P. C. & Arcucci, A. B. (1993). Dinosaurian precursors
from the Middle Triassic of Argentina: Lagerpeton chanarensis.Journal of Vertebrate Paleontology 13, 385–399.
Sereno, P. C. & Arcucci, A. B. (1994). Dinosaurian precursorsfrom the Middle Triassic of Argentina: Marasuchus lilloensis, gen.nov. Journal of Vertebrate Paleontology 14, 53–73.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

108 Max C. Langer and others
Sereno, P. C., Forster, C. A., Rogers, R. R. & Monetta,A. M. (1993). Primitive dinosaur skeleton from Argentina andthe early evolution of the Dinosauria. Nature 361, 64–66.
Sereno, P. C. & Novas, F. E. (1992). The complete skull andskeleton of an early dinosaur. Science 258, 1137–1140.
Sereno, P. C. & Novas, F. E. (1993). The skull and neck of thebasal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate
Paleontology 13, 451–476.Sereno, P. C. & Wild, R. (1992). Procompsognathus: theropod, ‘‘the-
codont’’ or both? Journal of Vertebrate Paleontology 12, 435–458.Sereno, P. C., Wilson, J. A. & Conrad, J. L. (2004). New
dinosaurs link southern landmasses in the Mid-Cretaceous. Pro-
ceedings of the Royal Society of London B 271, 1325–1330.Shipman, T.C. (2004). Links between sediment accumulation rates
and the development of alluvial architecture: Triassic Ischigualasto
Formation, northwestern Argentina. Unpublished Ph.D. Thesis. Uni-versity of Arizona, Tucson-AZ. 243 p.
Shubin, N. H. & Sues, H.-D. (1991). Biogeography of Early Meso-zoic continental tetrapods: patterns and implications. Paleobiology
17, 214–230.Sill, W. D. (1967). Proterochampsa barrionuevoi and the early evolu-
tion of the Crocodilia. Bulletin of the Museum of Comparative Zoology
35, 414–456.Simms, M.J. & Ruffell, A.H. (1990). Climatic and biotic change
in the Late Triassic. Journal of the Geological Society of London 147,321–327.
Simms, M. J., Ruffel, A. H. & Johnson, A. L. A. (1994). Bioticand climatic changes in the Carnian of Europe and adjacentareas. In In the shadow of the dinosaurs, early Mesozoic tetrapods (eds. N.C. Fraser and H.-D. Sues), pp. 352–365 Cambridge UniversityPress, Cambridge.
Smith, A. B. (1988). Echinoid evolution from the Triassic to earlyLiassic. Cahiers de l’Universite Catholique de Lyon Series des Sciences 3,79–117.
Smith, N. D. & Pol, D. (2007). Anatomy of a basalsauropodomorph dinosaur from the Early Jurassic Hanson For-mation of Antarctica. Acta Palaeontologica Polonica 52, 657–674.
Smith, N. D., Makovicky, P. J., Hammer, W. R. & Cur-rie, P. J. (2007). Osteology of Cryolophosaurus ellioti (Dinosauria:Theropoda) from the Early Jurassic of Antarctica and implica-tions for early theropod evolution. Zoological Journal of the Linnean
Society 151, 377–421.Sobral, G. & Langer, M. (2008). A supertree approach to prolac-
ertiform phylogeny. Journal of Vertebrate Paleontology 26(3, suppl.),145A.
Spencer, P. S. & Benton, M. J. (2000). Procolophonids fromthe Permo-Triassic of Russia. In The age of dinosaurs in Rus-
sia and Mongolia (eds. M. J. Benton, M. A. Shishkin, D. M. Unwinand E. N. Kurochkin), pp. 160–176 Cambridge University Press,Cambridge.
Spielmann, J. A., Lucas, S. G., Rinehart, L. F., Hunt, A. P.,Heckert, A. B. & Sullivan, R. M. (2007). Oldest records ofthe Late Triasssic theropod dinosaur Coelophysis bauri. In The
Global Triassic (eds. S. G. Lucas and J. A. Spielmann). New Mexico
Museum of Natural History & Science Bulletin 41, 384–401.Stanley, G. D. (2006). Photosymbiosis and the evolution of mod-
ern coral reefs. Science 312, 857–858.Steel, R. (1970). Saurischia. In Handbuch der Palaoherpetologie, Teil
14 (ed. O. Kuhn), pp. 1–88 Fischer-Verlag, Stuttgart.
Stipanicic, P.N. & Marsicano, C.A. (2002). Lexico estratigrafico
de la Argentina: Triasico, vol. 3. Asociacion Geologica Argentina,Buenos Aires.
Storrs, G. W. (1994). Fossil vertebrate faunas of the BritishRhaetian (Latest Triassic). In Vertebrate Palaeobiology (eds.M. J. Benton and D. B. Norman). Zological Journal of the Linnean
Society 112, 217–259.Sues, H-D. (1990). Staurikosaurus and Herrerasauridae. In
The Dinosauria (eds. D. B. Weishampel, P. Dodson andH. Osmolska), pp. 143–147 University of California Press,Berkley.
Sues, H.-D., Reisz, R. R., Hinic, S. & Raath, M. A. (2004). Onthe skull of Massospondylus carinatus Owen, 1854 (Dinosauria:Sauropodomorpha) from the Elliot and Clarens formations(Lower Jurassic) of South Africa. Annals of Carnegie Museum 73,239–257.
Sullivan, R. M. & Lucas, S. G. (1999). Eucoelophysis baldwini, anew theropod dinosaur from the Upper Triassic of New Mexico,and the status of the original types of Coelophysis. Journal of Vertebrate
Paleontology 19, 81–90.Tanner, L. H., Lucas, S. G. & Chapman, M. G. (2004). Assess-
ing the record and causes of Late Triassic extinctions. Earth-Science
Reviews 65, 103–139.Taylor, M. (2007). Phylogenetic definitions in the pre-PhyloCode
era; implications for naming clades under the PhyloCode. Paleo-
Bios 27, 1–6.Thulborn, R. A. (1970). The skull of Fabrosaurus australis, a Triassic
ornithischian dinosaur. Palaeontology 13, 414–432.Thulborn, R. A. (1971). Origins and evolution of ornithischian
dinosaurs. Nature 234, 75–78.Thulborn, R. A. (1972). The post-cranial skeleton of the Trias-
sic ornithischian dinosaur Fabrosaurus australis. Palaeontology 15,29–60.
Thulborn, R. A. (1974). A new heterodontosaurid dinosaur (Rep-tilia: Ornithischia) from the Upper Triassic Red Beds of Lesotho.Zoological Journal of the Linnean Society 55, 151–175.
Thulborn, R. A. (1975). Dinosaur polyphyly and the classifi-cation of archosaurs and birds. Australian Journal of Zoology
23, 249–270.Thulborn, R. A. (1984). The avian relationships of Archaeopteryx,
and the origin of birds. Zoological Journal of the Linnean Society 82,119–158.
Thulborn, R. A. (2000). Australia’s earliest theropods: footprintevidence in the Ipswich coal measures (Upper Triassic) of Queens-land. Gaia 15, 301–311.
Thulborn, T. (2006). On the tracks of the earliest dinosaurs: impli-cations for the hypothesis of dinosaurian monophyly. Alcheringa
30, 271–311.Thulborn, T. & Turner, S. (2003). The last dicynodont: an Aus-
tralian Cretaceous relict. Proceedings of the Royal Society of London B
270, 985–993.Torrens, H. (1992). When did the dinosaur get its name? New
Scientist 4, 40–44.Tucker, M.E. & Benton, M.J. (1982). Triassic environments,
climates and reptile evolution. Palaeogeography, Palaeoclimatology,
Palaeoecology 40, 361–379.Tykoski R. (2005). Osteology, ontogeny, and relationships of the coelo-
physoid theropods. PhD thesis, The University of Texas, Austin,USA 553 pp.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

The origin and early evolution of dinosaurs 109
Tykoski, R.S. & Rowe, T. (2004). Ceratosauria. In The
Dinosauria, second edition (eds. D. B. Weishampel, P. Dodson andH. Osmolska), pp. 47–70 University of California Press, Berkley.
Unwin, D.M. (1998). Feathers, filaments and theropod dinosaurs.Nature 391, 119–120.
Upchurch, P. (1997a). Prosauropoda. In Encyclopedia of Dinosaurs
(eds. P. J. Currie and K. Padian), pp. 599–607 Academic Press,San Diego.
Upchurch, P. (1997b). Sauropodomorpha. In Encyclopedia of
Dinosaurs (eds. P. J. Currie and K. Padian), pp. 658–660 Aca-demic Press, San Diego.
Upchurch, P., Barrett, P.M. & Galton, P.M. (2007). A phy-logenetic analysis of basal sauropodomorph relationships: impli-cations for the origin of sauropod dinosaurs. In Evolution and
palaeobiology of early sauropodomorph dinosaurs (eds. P. M. Barrett andD. J. Batten). Special Papers in Palaeontology 77 57–90.
Van Heerden, J. (1978). Herrerasaurus and the origin of the sauro-pod dinosaurs. South African Journal of Science 74, 187–189.
Veevers, J. J. (2005). Updated Gondwana (Permian–Cretaceous)earth history of Australia. Gondwana Research 9, 231–260.
Vickaryous, M. K. & Ryan, M. J. (1997). Ornamentation. InEncyclopedia of Dinosaurs (eds. P. J. Currie and K. Padian),pp. 448–493 Academic Press, San Diego.
Vickers-rich, P., Rich, T. H., McNamara, G. C. & Mil-ner, A. (1999). Is Agrosaurus macgillivrayi Australia’s oldestdinosaur? Records of the Western Australian Museum 57(suppl.),191–200.
Walker, A. D. (1961). Triassic reptiles from the Elgin area:Stagonolepis, Dasygnathus and their allies. Philosophical Transactions of
the Royal Society of London B 244, 103–204.Walker, A. D. (1964). Triassic reptiles from the Elgin area: Ornitho-
suchus and the origin of carnosaurs. Philosophical Transactions of the
Royal Society of London B 248, 53–134.Wang, S. C. & Dodson, P. (2006). Estimating the diversity of
dinosaur. Proceedings of the National Academy of Sciences of the United
States of America 103, 13601–13605.Ward, P. D. (2006). Out of Thin Air: Dinosaurs, Birds, and Earth’s
Ancient Atmosphere. Joseph Henry Press, Washington, DC.Weishampel, D. B. (1984). Interactions between Mesozoic plants
and vertebrates fructifications and seed predation. Neues Jarhbuch
fur Geologie und Palaontologie Abhandlung 167, 224–240.Weishampel, D. B. (2004). Ornithischia. In The Dinosauria, second
edition (eds. D. B. Weishampel, P. Dodson and H. Osmolska),pp. 323–324 University of California Press, Berkley.
Weishampel, D. B., Barrett, P. M., Coria, R. A.,Le Loeuff, J., Xu, X., Zhao, X., Sahni, A., Gomani,E. M. P., Noto, C. R. (2004). Dinosaur distribution. In The
Dinosauria, second edition (eds. D. B. Weishampel, P. Dodsonand H. Osmolska), pp. 517–606 University of California Press,Berkley.
Weishampel, D. B. & Weishampel, J. B. (1983). Annotated local-ities of ornithopod dinosaurs: Implications to Mesozoic paleobio-geography. Mosasaur 1, 43–87.
Weishampel, D. B. & Westphal, F. (1986). Die Plateosaurier vonTrossingen. Ausstellungs Katalog der Universitat Tubingen 19, 1–27.
Weishampel, D. B. & Witmer, L. M. (1990). Lesothosaurus,Pisanosaurus, and Technosaurus. In The Dinosauria (eds. D. B.Weishampel, P. Dodson and H. Osmolska), pp. 416–425 Uni-versity of California Press, Berkley.
Welles, S.P. (1984). Dilophosaurus wetherilli (Dinosauria,Theropoda): osteology and comparisons. Palaeontographica
Abteilung A 185, 85–180.White, M. E. (1990). The Flowering of Gondwana. Princeton Univer-
sity Press, Princeton.Wild, R. (1978). Ein Sauropoden-Rest (Reptilia, Saurischia) aus
dem Posidonienschiefer (Lias, Toarcium) von Holzmaden.Stuttgarter Beitrage zur Naturkunde B 41, 1–15.
Wilson, J. A. (2005). Integrating ichnofossil and body fossil recordsto estimate locomotor posture and spatiotemporal distribution ofearly sauropod dinosaurs: a stratocladistic approach. Paleobiology
31, 400–423.Wilson, J. A. & Sereno, P. C. (1998). Early evolution and higher-
level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleon-
tology, Memoir 5, 1–68.Wilson, J. A., Sereno, P. C., Srivastava, S., Bhatt, D. K.,
Khosla, A. & Sahni, A. (2003). A new abelisaurid (Dinosauria,Theropoda) from the Lameta Formation (Cretaceous, Maas-trichtian) of India. Contributions of the Museum of Palaeontology of the
University of Michigan 31, 1–42.Witmer, L.M. (2009). Dinosaurs: Fuzzy origins for feathers. Nature
458, 293–295.Wu, P., Hou, Plikus, M., Hughes, M., Scehnet, J., Suk-
saweang, S., Widelitz, R.B., Jiang, T.-X. & Chuong, C.-M.(2004). Evo-Devo of amniote integuments and appendages. Inter-
national Journal of Developmental Biology 48, 249–270.Xu, X., Zhao, X. & Clark, J. M. (2001). A new therizinosaur
from the Lower Jurassic Lower Lufeng Formation of Yunnan,China. Journal of Vertebrate Paleontology21, 477–483.
Xu, X., Forster, C. A., Clark, J.M. & Mo, J. (2006). A basalceratopsian with transitional features from the Late Juras-sic of northwestern China. Proceedings of the Royal Society B
273, 2135–2140.Xu, X., Clark, J. M., Mo, J., Choiniere, J., Forster, C. A.,
Erickson, G. M., Hone, D. W. E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C-k., Han, F-l., & Guo, Y. (2009). A Jurassic ceratosaur from China helpsclarify avian digital homologies Nature 459, 940–944.
Yadagiri, P. (2001). The osteology of Kotasaurus yamanpalliensis, asauropod dinosaur from the early Jurassic Kota Formation ofIndia. Journal of Vertebrate Paleontology 21, 242–252.
Yates, A.M. (2003a). A definite prosauropod dinosaur from theLower Elliot Formation (Norian: Upper Triassic) of South Africa.Palaeontologia Africana 39, 63–68.
Yates, A. M. (2003b). A new species of the primitive dinosaur,Thecodontosaurus (Saurischia: Sauropodomorpha), and its implica-tions for the systematics of early dinosaurs. Journal of Systematic
Palaeontology 1, 1–42.Yates, A. M. (2003c). The species taxonomy of the
sauropodomorph dinosaurs from the Lowenstein Formation(Norian, Late Triassic) of Germany. Palaeontology 46, 317–337.
Yates, A. M. (2004). Anchisaurus polyzelus (Hitchcock): The small-est known sauropod dinosaur and the evolution of gigantismamongst sauropodomorph dinosaurs. Postilla 230, 1–58.
Yates, A. M. (2005). A new theropod dinosaur from the EarlyJurassic of South Africa and its implication for the early evolutionof theropods. Palaeontologia Africana 41, 105–122.
Yates, A. M. (2007a). Solving a dinosaurian puzzle: the identity ofAliwalia rex Galton. Historical Biology 19, 93–123.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society

110 Max C. Langer and others
Yates, A. M. (2007b). The first complete skull of the Tri-assic dinosaur Melanorosaurus Haughton (Sauropodomorpha:Anchisauria). Special Papers in Palaeontology 77, 9–55.
Yates, A. M. (2008). A second specimen of Blikanasaurus
(Dinosauria: Sauropoda) and the biostratigraphy of the lowerElliot Formation. Palaeontologia Africana 43, 39–43.
Yates, A. M. & Kitching, J. W. (2003). The earliest knownsauropod dinosaur and the first steps towards sauropodlocomotion. Proceedings of the Royal Society of London B
270, 1753–1758.You, H., Xu, X. & Wang, X.-L. (2003). A new genus of Psitta-
cosauridae (Dinosauria:Ornithopoda) and the origin and earlyevolution of marginocephalian dinosaurs. Acta Geologica Sinica 77,15–20.
Young, C.-C. (1942). Yunnanosaurus huangi Young (gen. etsp. nov.), a new Prosauropoda from the Red Beds atLufeng, Yunnan. Bulletin of the Geological Society of China
22, 63–104.
Young, C.-C. (1951). The Lufeng saurischian fauna in China.Palaeontologia Sinica C 13, 1–96.
Zerfass, H., Lavina, E.L., Schultz, C.L., Garcia,A.G.V., Faccini, U.F. & Chemale Jr., F. (2003). Sequencestratigraphy of continental Triassic strata of southernmost Brazil:a contribution to Southwestern Gondwana palaeogeography andpalaeoclimate. Sedimentary Geology 161, 85–180.
Zerfass, H., Chemale Jr., F., Schultz, C.L. & Lavina, E.L.(2004). Tectonics and sedimentation in Southern South Americaduring Triassic. Sedimentary Geology 166, 265–292.
Zhang Y. & Yang Z. (1994). A new complete osteology of Prosauropoda
in Lufeng Basin, Yunnan, China: Jingshanosaurus. Yunnan PublishingHouse of Science and Technology, Kunming.
Zhao X.-J. & Xu X. (1998). The oldest coelurosaurian. Nature 394,234–235.
Zheng, X.-T., You, H.-L., Xu, X. & Dong, Z.-M. (2009). AnEarly Cretaceous heterodontosaurid dinosaur with filamentousintegumentary structures. Nature 458, 333–336.
Biological Reviews 85 (2010) 55–110 © 2009 The Authors. Journal compilation © 2009 Cambridge Philosophical Society