biogeography and ecology of sand-dwelling noctuids (noctuidae: lepidoptera) in … ·...
TRANSCRIPT

Israel Journal of entomology, Vol. 43, 2013, pp. 33-50
Biogeography and ecology of sand-dwelling noctuids (Noctuidae: Lepidoptera) in Israel.
KravchenKo vasiliy D.1, Pstygo irina2, sPeiDel Wolfgang3 anD Müller günter c.4
1 Department of Zoology, George S. Wise Faculty of Life Sciences, Tel Aviv University, Tel Aviv 69978, Israel
2 Faculty of Foreign Languages and Professional Communications, Ulyanovsk State University, 432017, Russia
3 Museum Witt, Tengstr. 33, D-80796 München, Germany 4 Department of Microbiology and Molecular Genetics,
IMRIC, Kuvin Centre for the Study of Infectious and Tropical Diseases, Faculty of Medicine, Hebrew University, Jerusalem 91120, Israel
ABSTRACT
All together 44 species of Noctuidae are specific to 5 sandy areas in Israel: Arava Valley, Rotem-Yamin Plain, Uvda Valley, Western Negev and Coastal Sand Dunes. The areas are different in temperature and moisture regimes and belong to different biogeographical districts in Israel (Mediterranean, Irano-Turanian and Saharo-Sindian). More than half of the species (26/44) are found only in one of these sandy areas. The highest number of these species is recorded from the sands of the Arava Valley (18 species) including all 8 Israeli species of Anumeta, no unique species were found in the Uvda Valley. Most of the sand-dwelling species are univoltine. On inland sands (Arava, Rotem, Uvda) they fly in November or February until April. On the Coastal Dunes and on adjacent sands of Western Negev the species fly in January because of warm nights (mean minimal temperature per month is 10.1ºC) compared to inland areas (mean minimal temperature of 4ºC). According to their general distribution pattern, most of the species inhabit-ing inland sands belong to eremic groups. Species inhabiting Coastal Sand Dunes and sands of the Western Negev are all circum-Mediterranean. The reasons for the association of these noctuids with sandy soils are discussed.
KEYWORDS: sand-dwelling, desert, sands, dunes, psammophilous, Noc-tuidae, Lepidoptera, Israel, biogeography.
INTRODUCTION
Sand covers about 20% of Earth's deserts (online at: http://earthsky.org/earth/how-did-the-sand-in-the-desert-get-there). Sandy areas are home for many species of

34 KravchenKo et al. Isr. J. entomol.
plants, animals, including many insects. For example, many plant species were de-scribed as specific to sandy areas in Greece (Sykora et al., 2003), in United Kingdom (UK) (Rodwell, 2000), and in Finland (Hellemaa, 1998). Many insects of different orders also occur only on sandy soils. In Norway 181 species of Coleoptera, 119 spe-cies of Hymenoptera, and 65 Lepidoptera species have been found to be specific to sand dunes (Ødegaard et al., 2010). Lepidoptera that are specific to sandy soils often comprise an essential part of the total number of species collected. On Coastal Sand Dunes in Cornwall (UK), for example, among 331 species of macro-Lepidoptera, 32 (9.7%) were specific to sand (Spalding, 1995). In the Vakhsh valley (Tajikistan) 28.9% of Lepidoptera species occurred only on sand (Shetkin, 1960; 1965), in the desert of western Uzbekistan this value is even higher 32% (Falkovich, 1968; 1979).
Biotopes characterized by sandy soils, especially Coastal Sand Dunes, are under strong pressure from development and recreation (Doody, 1991; Kutiel, 2001; Mun et al., 2011). Therefore, sand-dwelling (psammophilous) species are often at risk of extinction and included in the regional “red lists” (threatened species) of nature pro-tection organizations. In Norway nearly 16% of all the threatened and near threatened species on the Red List are inhabitants of sandy areas (Ødegaard et al., 2010).
Israel is situated at a crossroads of four biogeographical regions: Mediterranean, Irano-Turanian, Saharo-Sindian and in Sudanian penetration (Fig. 1) (Zohary, 1966; Furth, 1975). Sandy areas in Israel are common and present in all the biogeographical regions. Species assemblages and the ecology of psammophilous plants in Israel were repeatedly discussed by botanists (Orshan and Zohary, 1963; Danin, 1978; Danin & Nokrian, 1991; Danin, 1996); however, there are no publications devoted to any group of sand inhabiting insects in the region. In this paper we consider species com-position, distribution and the seasonal dynamic of flight of sand-dwelling noctuids in Israel.
Noctuidae is the most species-rich family of Lepidoptera, with 549 species docu-mented in Israel (Kravchenko et al., 2007a; 2007b). In recent years the classification of Noctuidae has been the subject of permanent classification changes. For this study we follow the recent classification of Fibiger and Lafontaine, 2005; Fibiger and Hack-er, 2005; Lafontaine and Schmidt 2010; Zahiri et al., 2012).
MATERIALS AND METHODS
Classification of biogeographical categories applied in this survey follows that for the Lepidoptera of Levant (Kravchenko et al., 2007a; 2007b) and Saudi Arabia (Wiltshire, 1994). The biogeographic term “Eremic” applied to species inhabiting the southern Palearctic desert belt from the African Atlantic coastal desert, through the Sahara to deserts of central Asia and northern India. Species are classified into four sub-groups, e.g. West-Eremic, East-Eremic, Central-Eremic and Pan-Eremic. We consider the cir-cum-Mediterranean distribution a special group. Information about the hostplants, distribution of species within Israel and their phenology are taken from the literature mentioned above and from our records for desert noctuids (Kravchenko et al., 2006).

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 35
Method of collecting. Since 1986, in the context of an Israeli-German project for the study of the Israeli Lepidopteran fauna, a network of permanent automatic light traps (220V 20W black light UVB & UVC tubes) was maintained. Traps were relo-cated on an annual basis. Results were published in two monographs (Kravchenko et al., 2007a; 2007b).
Terminology. Some species occur exclusively on sand, while others prefer san-dy soils. The terms psammophilous and ammophilous are used for sand-adapted and sand-associated terrestrial arthropods, respectively (Acorn, 2011). We apply the term psammophilous to species collected exclusively on sand and ammophilous for species in which more than 75% of the specimens were collected on sand. The term Levant is an imprecise term that denotes the area roughly bounded on the north by the Taurus Mountains, on the south by the Arabian Desert, and on the west by the Mediterranean Sea, while on the east it extends into Upper Mesopotamia (online at: http://www.gla-phyridae.com/Biogeografia/NEL.html).
Description of sandy areas. We collected Noctuidae in five relatively isolated sandy areas that vary in temperature/moisture regimes (Tab. 1) (Jaffe, S., 1988; Stern et al., 1986; Danin, 1988) and belong to different biogeographical regions (Fig 1). The Arava Valley is a penetration of Ethiopian (Sudanian) elements into the Levant (Shmi-da and Aronson, 1986). This represents an extreme desert with average annual precip-itation of less than 25 mm and an average daily maximum temperature in August (the hottest month) of 38ºC. The Northern Negev includes the Rotem-Yamin Sands and is in the Irano-Turanian region with a mean maximum temperature in August of 32ºC. The Southern Negev and Uvda Valley is a Saharo-Arabian region with an average daily maximum of temperature in August ranging from 34º-36ºC. Sands of Western Negev in the Saharo-Arabian region has an average maximum temperature in August of 34ºC. Coastal Sand Dunes in Israel extend along the shore of the Mediterranean Sea for about 190 km. This Mediterranean region has an average daily maximum tempera-ture in August ranging from 28º-30ºC.
Tab. 1Meteorological characteristics of the sandy areas.
Sandy areas Annual precipita-tion in mm
Mean temperature in coldest month (January) in ºC
Lowest Highest
Arava < 25 4 25
Rotem-Yamin 100-200 7 17.5
Uvda Valley ~40 4.2 22.1
Western Negev 200-300 7.3 23.6
Coastal Sand Dunes 400-700 10.1 22.4
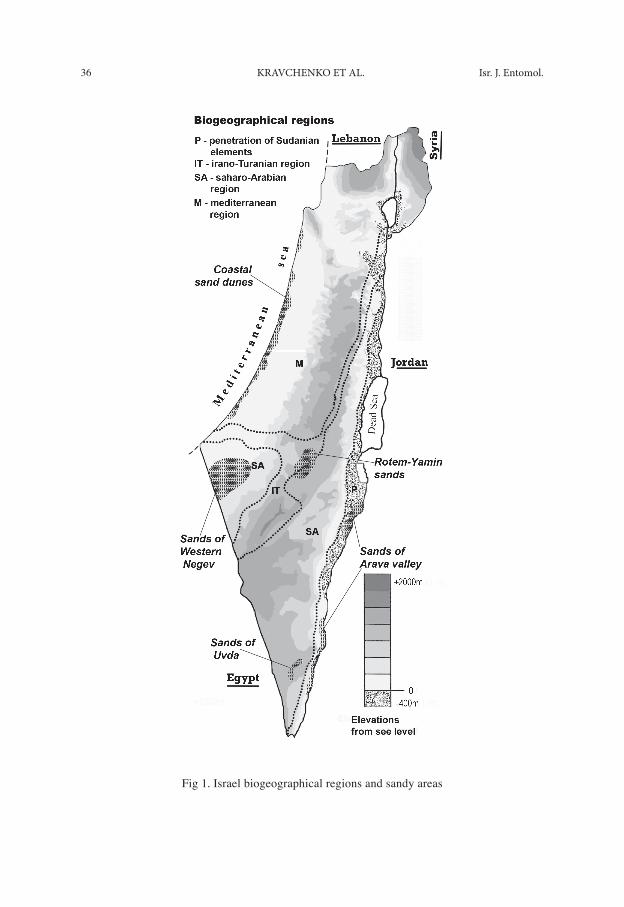
36 KravchenKo et al. Isr. J. entomol.
Fig 1. Israel biogeographical regions and sandy areas

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 37
RESULTS
Faunistic survey of sand-dwelling Noctuidae.
Family ErebidaeSubfamily ErebinaeTribe Toxocampini
1. Anumeta spilota ErshovDistribution: Pan-Eremic. From the Western Sahara to the Levant, Central Asia, Pa-kistan, and India. In Israel: Arava Valley.Period of flight: multivoltine, flying from March to September with the greatest abun-dance from March to May; larvae observed in May.Host-plants: in Israel, Calligonum comosum L. (Polygonaceae); in Turkmenistan, Calligonum spp.
2. Anumeta henkei (Staudinger)Distribution: East-Eremic. European part of southern Russia, Central Asia, Levant, Iran, Iraq, and Afghanistan. In Israel: Arava Valley.Period of flight: multivoltine, flying from March to October with the greatest abun-dance from March to May.Host-plants: in Israel and elsewhere unknown. Congeners on Calligonum spp.
3. Anumeta atrosignata (Walker)Distribution: East-EremicEast-Eremic. India, Iran, Iraq, Arabian Peninsula, and Le-vant. In Israel: Arava Valley.Period of flight: multivoltine, March to July with the greatest abundance from March to May.Host-plants: in Israel, Calligonum comosum; elsewhere unknown.
4. Anumeta straminea (A. Bang-Haas)Distribution: West-Eremic. Throughout the Sahara and the Arabian desert to Bahrain, northern Oman, and the Levant. In Israel: Arava Valley.Period of flight: univoltine, winter species, flying from December to March with the greatest abundance in January.Host-plants: in Israel unknown; in Saudi Arabia on Calligonum spp.
5. Anumeta arabiae WiltshireDistribution: Endemic to Saudi Arabia and the Arava. In Israel: Arava Valley.Period of flight: bivoltine, spring species, flying from January to April with the great-est abundance from January to February and in April.Host-plants: unknown. Congeners on Calligonum spp.

38 KravchenKo et al. Isr. J. entomol.
6. Anumeta asiatica Wiltshire
Distribution: East-Eremic. Southwestern Iran, Iraq, Levant, Kuwait, Saudi Arabia, Oman, and United Arab Emirates. In Israel: Arava Valley.Habitat: deserticolous, psammophilous.Period of flight: multivoltine, summer species, flying from May to August with the greatest abundance in May.Host-plants: in Israel, Calligonum comosum; elsewhere unknown.
7. Anumeta hilgerti Rothschild
Distribution: West-Eremic. From the deserts of North Africa, including the central Sahara, to the Arabian Peninsula, and the Levant. In Israel: Arava Valley.Period of flight: univoltine, spring species, February to May with the greatest abun-dance in March. Host-plants: unknown. Congeners on Calligonum spp.
8. Anumeta cestis (Ménétriès)
Distribution: Pan-Eremic. From the Sahara to the Levant, Central Asia, Kazakhstan, S.Urals. In Israel: Arava Valley.Period of flight: univoltine, spring species, February to May with the greatest abun-dance in March. Host-plants: unknown. Congeners on Calligonum spp.
Tribe Melipotini
9. Drasteria oranensis Rothschild
Distribution: West-Eremic. From Algeria to Libya, Egypt, Levant, and Saudi Arabia. In Israel: on sands of Rotem-Yamin. Period of flight: so far collected only in March and in April; in Tunisia June to Sep-tember.Host-plants: in Israel Helianthemum sp.(Cistaceae); in North Africa and Saudi Ara-bia, Calligonum comosum.
10. Drasteria kabylaria (A. Bang-Haas)
Distribution: West-Eremic. From the western and central Sahara to the Arabian Peninsula, Iran, and the Levant. In Israel: on all sandy areas except Coastal Sand Dunes.Period of flight: bivoltine, March to May and October.Host-plants: in Israel Helianthemum lippii (L.)

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 39
Tribe Ophiusini
11. Cerocala sana Staudinger
Distribution: West-Eremic. Turkey, Iraq, Iran, Levant, Egypt, Libya, and Algeria. In Israel: all sandy areas.Period of flight: probably bivoltine, flying from October through winter to April with the greatest abundance in November and March.Host-plants: in Israel unknown, in Saudi Arabia monophagous on Helianthemum ka-hiricum Delile and H. lippii (L.) (Cistaceae).
Tribe Anydrophilini
12. Anydrophila stuebeli (Calberla)
Distribution: West-Eremic. Central Arabia, United Arab Emirates, and Levant. In Israel: Arava Valley.Period of flight: univoltine, spring species, March to May.Host-plants: in Israel, Rumex and Calligonum spp. (both Polygonaceae), elsewhere unknown.
Subfamily AcontiinaeTribe Acontiini
13. Acontia biskrensis (Oberthür)
Distribution: West- and Central-Eremic. From the Canary Islands through North Af-rica and the Sahara to the Arabian Peninsula, Levant, Iraq, and Iran. In Israel: Arava Valley and Dead Sea Area.Period of flight: in Israel univoltine, spring species, March to April.Host-plants: unknown.
Tribe Armadini
14. Armada nilotica A. Bang-Haas
Distribution: probably endemic to southeast Mediterranean coast. Described from the coastal dunes of El-Arish oasis in Egypt. In Israel: sands of western Negev.Period of flight: univoltine, collected so far only from March to May; in Egypt (Sinai) from May to June.Host-plants: in Israel unknown; in Sinai, Heliotropium luteum Poir and H. arbain-ense Fresen..
15. Tarachephia hueberi (Ershov)
Distribution: East-Eremic. Iraq, Iran, Levant, Trans-Caspian region, Uzbekistan, Turkmenistan, and Afghanistan. In Israel: Arava Valley and Dead Sea Area.

40 KravchenKo et al. Isr. J. entomol.
Period of flight: univoltine, spring species observed so far only in March; in Central Asia flying from March to May.Host-plants: in Israel unknown; in Central Asia, larvae found in May on leaves of Arnebia decumbens (Boraginaceae).
Subfamily Metoponiinae
16. Epharmottomena eremophila (Rebel,)
Distribution: West-Eremic. From Morocco and the western Sahara to the Levant. In Israel: Western Negev.Period of flight: probably bivoltine, September through winter to April with the greatest abundance in October and March.Host-plants: unknown.
17. Iranada turcorum (Zerny)
Distribution: West-Eremic. Egypt, Levant, southwestern Iran, Iraq, and Oman. In Israel: Coastal Sand Dunes and western Negev.Period of flight: in Israel univoltine, spring species, January to March.Host-plants: unknown.
Subfamily Cuculliinae
18. Cucullia macara Rebel
Distribution: West-Eremic. Egypt, Arabian Peninsula and Levant. In Israel: all sandy areas except Coastal Sand Dunes.Period of flight: in Israel univoltine, winter species, December to February.Host-plants: unknown. Congeners on Scrophularia spp. (Scrophulariaceae).
19. Cucullia strigicosta Boursin
Distribution: Iranian. Iraq, Turkey, Azerbaijan, Iran and Levant. In Israel: Western Negev.Period of flight: in Israel univoltine, winter species, January to March.Host-plants: in Israel unknown; in Iraq, probably Scrophularia spp. (Wiltshire, 1957).
20. Cucullia macewani Wiltshire
Distribution: Probably endemic to coasts of the Arabian Peninsula and east Mediter-ranean. Saudi Arabia, Yemen, and Levant. In Israel: Coastal Sand Dunes.Period of flight: in Israel univoltine, winter species, January to February.Host-plants: unknown. Congeners on Scrophularia spp. and Verbascum spp. (both are Scrophulariaceae).

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 41
Subfamily Oncocnemidinae
21. Lithophasia quadrivirgula (Mabille)
Distribution: West-Eremic. Throughout North Africa from Morocco to Egypt, also Levant and Iraq. In Israel: all sandy areas.Period of flight: in Israel univoltine, winter species, November to February with the greatest abundance in December.Host-plants: unknown.
22. Calophasia barthae Wagner
Distribution: north (Circum-)Mediterranean. Southern part of Balkans, Turkey and Levant. In Israel: sands of Western Negev.Period of flight: in Israel univoltine,, spring species, collected so far only in March.Host-plants: unknown.
23. Calophasia angularis (Chrétien)
Distribution: Pan-Eremic. From the western Sahara and Morocco throughout North Africa to Saudi Arabia, the Levant, Iraq and Turkmenistan along the Kopet-Dagh Mts. In Israel: all sandy areas except Coastal Sand Dunes.Period of flight: in Israel univoltine, early spring species, February to March.Host-plants: unknown.
24. Cleonymia chabordis (Oberthür)
Distribution: West- and Central-Eremic. North Africa, Saudi Arabia, Levant, Iraq and Iran. In Israel: all sandy areas.Period of flight: in Israel univoltine, early spring species, January to April.Host-plants: in Israel, Helianthemum stipulatum, elsewhere unknown.
25. Metopoceras omar (Oberthür)
Distribution: Pan-Eremic. From northwestern Africa to the Levant, Sicily and south-ernmost Italy, the Arabian Peninsula, coasts of the Caspian Sea, Turkmenistan, the Near and Middle East. In Israel: all sandy areas.Period of flight: in Israel univoltine, early spring species, January to April with the highest rate of occurrence from February to March.Host-plants: unknown.
26. Metopoceras kneuckeri (Rebel)
Distribution: Pan-Eremic. From northwestern Africa across the Arabian Peninsula to Pakistan. In Israel: all sandy areas except Coastal Sand Dunes.Period of flight: in Israel univoltine, spring species, March to April.Host-plants: unknown.

42 KravchenKo et al. Isr. J. entomol.
Subfamily NoctuinaeTribe Caradrinini
27. Scythocentropus eberti HackerDistribution: probably endemic to sands of the Arava Valley. Collected in all sandy areas of Arava Valley, also on Jordanian side the Arava.Period of flight: in Israel univoltine, autumn species, October to November with the highest rate of occurrence in November. Host-plants: unknown.
28. Scythocentropus inquinata (Mabille)Distribution: Pan-Eremic. Morocco, Algeria, Tunisia, western and central Sahara, Levant, Arabian Peninsula, Iraq, Iran, Turkey, Pakistan and northern India. In Israel: all sandy areas.Period of flight: in Israel univoltine, autumn species, September to December with the highest rate of occurrence in November.Host-plants: unknown.
29. Diadochia stigmatica WiltshireDistribution: West-Eremic. Western Saudi Arabia and Levant. In Israel: Arava Valley.Period of flight: in Israel univoltine, autumn species, November.Host-plants: in Israel unknown; larvae of Central Asian congeners develop from April to May on Salsola arbuscula Pallas and S. richteri (Moq.) (Amaranthaceae).
Tribe Xylenini
30. Polymixis epiphleps (Turati & Krüger)Distribution: south (Circum-)Mediterranean. Libya and Levant. In Israel: Western Negev and Coastal Sand Dunes.Period of flight: in Israel univoltine, winter species, November to January with the highest rate of occurrence in December.Host-plants: unknown.
31. Mniotype compitalis (Draudt)Distribution: south (Circum-)Mediterranean. Coastal Libya, Egypt and Levant. In Israel: Coastal Sand Dunes.Period of flight: in Israel univoltine, winter species, November to December.Host-plants: unknown.
32. Boursinia lithoxylea (A. Bang-Haas)Distribution: West-Eremic. North Africa, western Arabian Peninsula, and Levant. In Israel: Arava Valley.

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 43
Period of flight: in Israel univoltine, autumn species, November.Host-plants: in Israel unknown; elsewhere, Retama raetam. (Fabaceae).
Tribe Hadenini
33. Odontelia daphnadeparisae Krachenko, et al.Distribution: East-Eremic. Arabian Peninsula, Turkmenistan, Afghanistan, Iran and Levant. In Israel: Arava Valley.Period of flight: in Israel univoltine, winter species, December to March with the highest rate of occurrence in February.Host-plants: unknown. Larvae in Israel were found living subterranean in sand dunes with Haloxylon persicum Bunge (Amaranthaceae)..
Tribe Leucaniini
34. Leucania punctosa (Treitschke)Distribution: Mediterranean-Iranian. From Morocco to Libya, southern Europe, Tur-key, Armenia, Levant, Iran, Iraq and Turkmenistan. In Israel: Western Negev and on Coastal Sand Dunes.Period of flight: in Israel univoltine, autumn species, November to December.Host-plants: in Israel, Desmostachya bipinnata Stapf (Poaceae=Gramineae); in Eu-rope, undefined Poaceae (Gramineae).
Tribe NoctuiniSubtribe Agrotina
35. Euxoa oranaria (A. Bang-Haas)Distribution: West-Eremic. Iberian Peninsula, one record in Sicily, also Morocco, Al-geria, Saudi Arabia and Levant. In Israel: all sandy areas except Coastal Sand Dunes.Period of flight: in Israel univoltine, autumn species, November.Host-plants: unknown.
36. Euxoa anarmodia (Staudinger)Distribution: West-Eremic. From Algeria to Tunisia, Libya, Egypt and Levant. In Israel: all sandy areas.Period of flight: in Israel univoltine, autumn species, October to December with the greatest abundance in November.Host-plants: in Israel and elsewhere unknown; in captivity reared on Hyoscyamus muticus L. (Solanaceae).
37. Euxoa canariensis (Rebel)Distribution: Pan-Eremic. From the Canary islands throughout the arid and semi-arid

44 KravchenKo et al. Isr. J. entomol.
parts of North Africa to the Arabian Peninsula, the Levant, Iran, and Afghanistan. In Israel: all sandy areas except the Coastal Plain.Period of flight: in Israel univoltine, spring species, March to May.Host-plants: unknown; in captivity reared on Atriplex halimus L. (Chenopodiaceae).
38. Agrotis alexandriensis Bethune-BakerDistribution: south (Circum-)Mediterranean. From Tunisia to Egypt and Levant. In Israel: Coastal Sand Dunes.Period of flight: in Israel collected so far only in October, possibly univoltine, autumn species; in North Africa bivoltine, flying in April and November.Host-plants: unknown.
39. Agrotis herzogi RebelDistribution: Pan-Eremic. North Africa, Arabian Peninsula, Levant, Iran, Pakistan and western India. Migrating to southern Europe, found in Sicily and Greece. In Isra-el: Arava Valley.Period of flight: in Israel probably bivoltine, October to April with the greatest abun-dance in November and March.Host-plants: in Israel, Atriplex leucoclada Boiss. (Chenopodiaceae); elsewhere un-known.
40. Agrotis haifae StaudingerDistribution: West-Eremic. North Africa, Arabian Peninsula and Levant. In Europe restricted to one locality in northern Greece. In Israel: Arava Valley.Period of flight: in Israel univoltine, autumn species, October to April with the great-est abundance in November.Host-plants: in Israel and elsewhere unknown.
41. Agrotis sardzeana BrandtDistribution: Pan-Eremic. North Africa, Arabian Peninsula, Levant, Iran, Pakistan and eremic parts of India. In Israel: Arava Valley.Period of flight: in Israel univoltine, autumn species, October to December with the greatest abundance in November.Host-plants: unknown. In captivity reared on Atriplex halimus L.
42. Agrotis pierreti (Bugnion)Distribution: West- and Central-Eremic. Southeastern Spain, North Africa, Iraq, southern Iran and Levant. In Israel: all sandy areas.Period of flight: in Israel univoltine, autumn species, October to November with the greatest abundance in November.Host-plants: unknown.

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 45
43. Agrotis lasserei (Oberthür)
Distribution: Pan-Eremic. Widespread throughout the eremic zone of the Palearctic region. From southeastern Spain and the western Sahara to Turkey, the Levant, Iraq, Iran, and Turkmenistan. In Israel: all sandy areas except Coastal Sand Dunes.Period of flight: in Israel univoltine, autumn species, October to November.Host-plants: unknown.
44. Agrotis boetica (Boisduval)
Distribution: south (Circum-)Mediterranean. Southern Iberian Peninsula, North Afri-ca (Morocco, Algeria and Tunisia), and Levant. In Israel: Western Negev and Coastal Sand Dunes.Period of flight: in Israel univoltine, winter species, November to December.Host-plants: in Israel unknown; in Spain on Lithospermum spp (Boraginaceae), Echi-nops spp (Asteraceae) and Astragalus spp, (Fabaceae).
Species composition on different sandy areas
More than half of the species (26/44 = 59.1%) occurred only in a single sandy area (Table 1). There are 18 species found exclusively in the sands of the Arava Valley in-cluding all 8 Israeli species of Anumeta. One species (Drasteria oranensis) has been found only in Rotem-Yamin sands. In the Uvda Valley no unique species has been found. Four species are found exclusively in the sands of the western Negev, and three species in Coastal Sand Dunes. Seven species inhabit all the sandy areas other than the Coastal Sand Dunes. Four species were common for the Coastal Sand Dunes and the Sands of the Western Negev and another seven species were found in all sandy areas.
Biogeographical composition
According to their general distribution pattern, most of the species inhabiting inland sands belong to the eremic group. There are 14 West-Eremic species; 5 East-Eremic; 5 Central-Eremic, 3 West- and Central-Eremic and 10 Pan-Eremic. Most of these species are widespread on sands in the Palearctic desert belt, e.g. 11 of them are a common component of sand dwelling Lepidoptera in central Asian deserts (Falkov-ich, 1979).
Species inhabiting the Coastal Sand Dunes and sands of the Western Negev are all circum-Mediterranean. There are four south Circum-Mediterranean species inhabiting coastal sands of the African Mediterranean coast: M. compitalis, A. alexandriensis, P. epiphleps, A. boetica. Also one Mediterranean-Iranian species: L. punctosa maintains a preference for sandy areas throughout its distribution (Hacker, 1999; 2001).
The endemic group includes four species with different types of distribution. Ar-mada nilotica is known only from the type locality (Egypt, El-Arish oasis, Coastal Sand Dunes) and from the adjacent part of the Western Negev in Israel; A. arabia was

46 KravchenKo et al. Isr. J. entomol.
Table 2Species assemblages according to their preference for different sandy areas

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 47
described from Saudi Arabia and recorded from a few localities along the Arava Val-ley; C. macewani is probably an inhabitant of coastal dunes of the Arabian peninsula and east-Mediterranean; and S. eberti is probably endemic to the Arava Valley sands, recorded both in the Israeli and Jordanian part of this valley.
Host plants
Host plants for most species are unknown. Some noctuid genera can be associated with host plant genera. For example, desert plants of the genus Calligonum are known to be hosts for species of Anumeta. Larvae of Cucullia usually develop on Scrophu-laria spp. Larvae of Noctuinae are known to be polyphagous on roots and stems of various grasses.
Periods of flight
The only multivoltine species among the sand-dwelling noctuids were species of An-umeta, flying from February to July with 2-3 generations. Other species are univoltine and fly during a 2-3 week period once a year. On inland sands (Arava, Uvda, Rotem) species fly during two periods (Fig. 2), one in November (autumnal species) and an-other from February to April (spring species). In the Coastal Sand Dunes and adjacent Sands of the Western Negev, sand-dwelling species were collected only in winter (December and January).
Fig 2. Periods of flight of the sand-dwelling species in different sandy areas

48 KravchenKo et al. Isr. J. entomol.
DISCUSSION
Aridisation and sand formation are inextricably linked. If there is no vegetation or water to hold the soil, the wind takes away the clay's particles and dried organic mat-ter. The rest is desert sand (Lancaster, 2009). Probably sandy areas have become an independent center of speciation inside the desert; therefore, the vast majority of the sand dwelling species in Israel has an Eremic type of distribution.
Only seven species were found in all of the sandy areas, others showed prefer-ence for different sandy areas. The primary differences of species composition in biogeographical areas and in periods of seasonal flight are between species living in the inland and coastal sands. Species restricted to coastal dunes and to sands of the Western Negev belong to a Circum-Mediterranean type of distribution and fly in the winter. The minimal winter temperatures and moisture are different in coastal and in-land areas. The coastal areas get more moisture (Table 1) in the form of precipitation and from fog (Goldreich, 2003). Relatively high winter temperatures allow moths to initiate flight activity in the winter in contrast to the Arava Valley, where the winter temperature often drops to 4ºC at night (Table 1).
Conformity of monophagous species to sandy soils most probably is a result of host plant specialization to sandy soils. Sand-dwelling species probably become host specific because of the low diversity of plants occurring on sandy soils. Mechanical characteristics of sand must be specific in order for the cutworms to be able to inhabit these types of habitats. In desert Lepidoptera larvae hide in the soil during hot and dry days and come out to feed only at night. Therefore, the relationship between soil type and species composition of Lepidoptera in the desert is derived not only through the relation of soil type and host plant, but also through the suitability of the soil as a shelter for the larva (Falkovich, 1968). Pupation of most sand-dwelling Lepidoptera also takes place in the soil. Sand-dwelling Lepidoptera are specifically adapted to successfully emerge from the pupa and make their way to the surface. That success depends on: a) hatching is under the control of a diurnal rhythm, allowing hatching only in the most humid night hours (for example Jacobson, 1970; Peng-Fei, et al., 2007); and b) many desert Lepidoptera possess special morphological structures for successful squeezing out that are strong spines on the tibia, or frontal tubercles of different shapes in order to push the soil (Falkovich, 1968). Psammophilous species of Euxoa without a frontal tubercle have less difficulty emerging from the soil as the non-psammophilous species (Lafontaine, 1981).
ACKNOWLEDGEMENTS
We thank all our colleagues and the many generous Israeli citizens who helped with this survey. We are grateful to the Israel Nature and Parks Authority (INPA), who sup-plied the collecting permits, especially to Dr. Reuven Ortal. Prof. Zoya Yefremova is thanked for the useful comments early draft of this manuscript.

VOL. 43, 2013 BIogeography and ecology of sand-dwellIng noctuIds 49
LITERATUREAcorn, J.H. 2011. Sand Hill Arthropods in Canadian Grasslands, 2: 25-43. In: Floate, K. D.
(ed.) Arthropods of Canadian Grasslands: Inhabitants of a Changing Landscape. Ed. by K. D. Floate. Biological Survey of Canada.
Danin, A. 1978. Plant species diversity and plant succession in a sandy area in the Northern Negev. Flora. 167: 409-422.
Danin, A. 1988. Flora and vegetation of Israel and adjacent areas. Pp. 129-157. In: Yom-Tov Y. and Tchernov J. (eds.), The Zoogeography of Israel. The Distribution and Abundance at a Zoogeographical Crossroad. Dr. W. Junk Publishers, Dordrecht, 600pp.
Danin, A. and Nokrian, R. 1991. Dynamics of dune vegetation in the southern coastal area of Israel since 1945. Documentes Phytosociologique. 13: 281-296.
Danin, A. 1996. Plants of desert dunes. Springler-Verlag, Berlin Heidelberg. 177 pp.Doody, P. (ed.). 1991. Sand dune inventory of Europe. – EUCC, Leiden/JNCC, Peterborough,
80 pp.Falkovitch, M.I. 1968. Food relationships of desert Lepidoptera in Central Asia. 21th Annual
Meeting dedicated to the memory of N.A. Kholodkovsky. Nauka, Leningrad: 53-89 (in Russian).
Falkovich, M.I. 1979. Seasonal development of the desert Lepidoptera of Soviet Central Asia and a historical analysis of the development of the lepidopteran fauna. Entomological Re-view 58 (2): 260-281 (in Russian).
Fibiger, M. and Hacker H. 2005: Systematic list of the Noctuoidea of Europe (Notodontidae, Nolidae, Arctiidae, Lymantriidae, Erebidae, Micronoctuidae, and Noctuidae). Esperiana 11: 93-205.
Fibiger, M. and Lafontaine J.D. 2005. A review of the higher classification of the Noctuoidea (Lepidoptera) with special reference to the Holarctic fauna. Esperiana 11: 7-92.
Furth, D.G. 1975. Israel, a great biogeographic crossroad. Discovery. 11: 3-13.Goldreich, Y. 2003. Israel Climatology: observation and research. Part I. Evaporation, humidi-
ty, cloudiness, fog, dew and visibility. pp. 270.Hacker, H.H. 1999. Systematic list of the Lepidoptera of the Arabian Peninsula with a survey of
the spread with special reference to the fauna of Yemen. Esperiana 7: 15-237.Hacker, H.H. 2001. Fauna of the Nolidae and Noctuidae of the Levante with description and
taxonomic notes. P.1-315. Esperiana 8: 1-942.Hellemaa, P. 1998. The development of coastal dunes and their vegetation in Finland. Fennia
(Helsinki) 176:1. Jacobson, L.A. 1970. Laboratory ecology of the red-backed cutworm, Euxoa ochrogaster (Lep-
idoptera: Noctuidae). The Canadian Entomologist 102(1): 85-89.Jaffe, S., 1988. Climate of Israel. Pp. 79-95. In: Yom-Tov, Y. and Tchernov, E, (eds.). The Zoo-
geography of Israel. The Distribution and Abundance at a Zoogeographical Crossroads 79-95. Kluwer Academic Publishers, Netherlands, 600 pp.
Kravchenko, V.D., Hausmann, A., and Müller, G.C. 2006. Deserticolous Noctuidae from Israel: New host plant records and description of larval habitats (Lepidoptera: Noctuidae). Mittei-lungen der Münchner Entomologischen Gesellschaft. 96: 27-40.
Kravchenko, V.D., Fibiger, M., Hausmann A., and Müller, G.C. 2007a. Erebidae. In: Müller, G.C., Kravchenko, V.D., Hausmann, A., Speidel, W., Mooser, J., and Witt, T.J. (eds.).The Lepidoptera of Israel, Vol. 1, Pensoft Series Faunistica 62, 1-168 pp.
Kravchenko, V.D., Fibiger, M., Hausmann, A., and Müller, G.C. 2007b. Noctuidae. In: Müller,

50 KravchenKo et al. Isr. J. entomol.
G.C., Kravchenko, V.D., Hausmann, A., Speidel, W., Mooser, J., and Witt, T.J. (eds.).The Lepidoptera of Israel, Vol. 2, Pensoft Series Faunistica 63, 1-400 pp.
Kutiel, P. 2001. Conservation and management of the Mediterranean Coastal Sand Dunes in Israel. Journal of Coastal Conservation 7: 183-192
Lafontaine, J.D. 1981. Classification and phylogeny of the Euxoa detersa group (Lepidoptera: Noctuidae). Quaestiones Entomologicae 17:1-120.
Lafontaine, J.D. and Schmidt, C. 2010. Annotated check list of the Noctuoidea (Insecta, Lepi-doptera) of North America north of Mexico. ZooKeys. Special Issue 40: 1-239.
Lancaster, N. 2009. Aeolian features and processes. Pp. 1-26. In: Young, R. and Norby, L. (eds.). Geological monitoring.The Geological Society of America.
Mun, S., Gallego Ferna, J. B., Dellafiore, C.M. 2011. Dune Vulnerability in Relation to Tourism Pressure in Central Gulf of Cádiz (SW Spain), a Case study. Journal of Coastal Research 27 (2): 243-251.
Ødegaard, F., Brandrud, T.E., and Pedersen, O. 2010. Sandy areas. Pp. 79-88. In: Kålås, J.A., Henriksen, S., Skjelseth, S., and Viken, Å. (eds.). Environmental conditions and impacts for Red List species. Norwegian Biodiversity Information Centre, Trondheim. Pp. 1-135.
Orshan, G. and Zohary, M. 1963. Vegetation of the sand deserts in the Western Negev of Israel. Plant Ecology. 11(3): 112-120
Lu, P.-F. Qiao, H.-L. Wang, X.-P., Xi-Qiao Wang, X.-Q. and Lei, C.-L. 2007. The emergence and mating rhythms of the legume pod borer, Maruca vitrata (Fabricius, 1787) (Lepidop-tera: Pyralidae). Pan-Pacific Entomologist 83(3): 226-234.
Rodwell, J.S. (Ed.). 2000. Maritime communities and vegetation of open habitats, Vol. 5. Brit-ish plant communities. Cambridge University Press, 1-512 pp.
Shetkin, U.L. 1960. Higher Lepidoptera of Vakhsh Valley (Tajikistan). Part I. Rhopalocera and Geterocera. Proceedings of Institute of Zoology and Parazitology of Tajikistan 19: 1-303
Shetkin, U.L. 1965. Higher Lepidoptera of sands of Vakhsh Valley, Publishing house of Tajiki-stan Akademy of Science, Dushanbe, 1-194 pp.
Shmida, A. and Aronson, J.A. 1986. Sudanian elements in the flora of Israel. Annals of the Missouri Botanical Garden. 73 (1): 1-28
Spalding, A. 1995. The moths at Carlyon Bay, Cornwall recorded 1989-1993. British Journal of Entomology and Natural History 8: 61-71.
Stern, E., Gradus, Y., Meir, A., Krakover, S., and Tzoar, H. (eds.). 1986. Atlas of the Negev. Keter Publishing House, Jerusalem (in Hebrew and English). Pp. 1-44
Sykora, K.V., Babalonas D. and Papastergiadou, E.S., 2003. Strandline and sand-dune vege-tation of coasts of Greece and of some other Aegean countries. Phytocoenologia 33 (2-3): 409-446.
Wiltshire, E.P. 1957. The Lepidoptera of Iraq. Nicholas Kaye Ltd., London, 162 pp.Wiltshire, E.P. 1994. An illustrated, annotated catalogue of the Macro-Heterocera of Saudi Ara-
bia. Fauna of Saudi Arabia 11: 91-250.Zahiri, R., Holloway, J. D., Kitching, I. J., Lafontaine, J. D., Mutanen, M., and Wahlberg, N.
2012. Molecular phylogenetics of Erebidae (Lepidoptera, Noctuoidea). Systematic Ento-mology 37(1): 102-124.
Zohary, M. 1966. Flora Palaestina, Part I, text. The Israel Academy of Sciences and Human-ities, Jerusalem, 364 pp.