barton et al, chem rev, 104 4867 (2004) (special …bip.cnrs-mrs.fr/bip06/pdf/leger_ise2010.pdf ·...
TRANSCRIPT


Marseille
Nice
Southampton
FR
UK
Laboratory of Bioenergetics
and Engineering of Proteins
CNRS, Marseille
Slides can be downloaded at
http://bip.cnrs-mrs.fr/bip06/publications

Barton et al, Chem Rev, 104 4867 (2004) (special issue on Batteries and Fuel cells)

I. Background: the structures of redox enzymes and their cofactors
II. Electrodes for direct electron transfer (ET) to proteins and enzymes
III. « Non catalytic » voltammetry
• at slow scan rates: measuring reduction potentials
• at fast scan rates: measuring rates of interfacial ET and coupled chemical events
IV. Catalytic voltammetry for measuring the enzyme’s activity & studying the mechanism
• principle
• learning about the mechanism from steady-state measurements
• deviation from steady-state
V. Using redox enzymes
• Limitations
• Fuel cells and photoelectrochemical cells
• Heterogenesous catalysts based on conducting particles
• Biosensors

I) Background: structural biology
Proteins (= polypeptides) are linear polymers made of
hundreds of α-aminoacids joined by peptides bonds

There are 21 natural α-aminoacids
which differ by their side chain « R »average Mw of an amino acid = 130 Da
Mw of a protein in the range 10-1000 kDa
Amino Acid Short name Abbrev. Side chain R Hydrophobic pKa Polar
Alanine A Ala -CH3 X - -Cysteine C Cys -CH2SH X 8.18 -Aspartic acid D Asp -CH2COOH - 3.90 X
Glutamic acid E Glu -CH2CH2COOH - 4.07 X
Phenylalanine F Phe -CH2C6H5 X - -
Glycine G Gly -H X - -
Histidine H His -CH2-C3H3N2 - 6.04 X
Isoleucine I Ile -CH(CH3)CH2CH3 X - -
Lysine K Lys -(CH2)4NH2 - 10.54 X
Leucine L Leu -CH2CH(CH3)2 X - -
Methionine M Met -CH2CH2SCH3 X - -
Asparagine N Asn -CH2CONH2 - - X
Proline P Pro -CH2CH2CH2- X - -
Glutamine Q Gln -CH2CH2CONH2 - - X
Arginine R Arg -(CH2)3NH-C(NH)NH2 - 12.48 X
Serine S Ser -CH2OH - - X
Threonine T Thr -CH(OH)CH3 - - X
Selenocysteine U Sec -CH2SeH X 5.73 -
Valine V Val -CH(CH3)2 X - -
Tryptophan W Trp -CH2C8H6N X - -
Tyrosine Y Tyr -CH2-C6H4OH - 10.46 X
http://en.wikipedia.org/wiki/Proteinogenic_amino_acid

The sequence of aminoacids in each protein
(also called primary structure)
is defined by the DNA (by the sequence of the gene)
Example: the sequence of the small protein of the enzyme « hydrogenase »ltakhrpsvvwlhnaectgcteaairtikpyidalildtisldyqetimaaageaae
aalhqalegkdgyylvvegglptidggqwgmvaghpmiettkkaaakakgiicigtc
sayggvqkakpnpsqakgvsealgvktinipgcppnpinfvgavvhvltkgipdlde
ngrpklfygelvhdncprlphfeasefapsfdseeakkgfclyelgckgpvtynncp
kvlfnqvnwpvqaghpclgcsepdfwdtmtpfyeqg
Site-directed mutants are proteins in which one or several
amino acids have been changed, using genetic
engineering
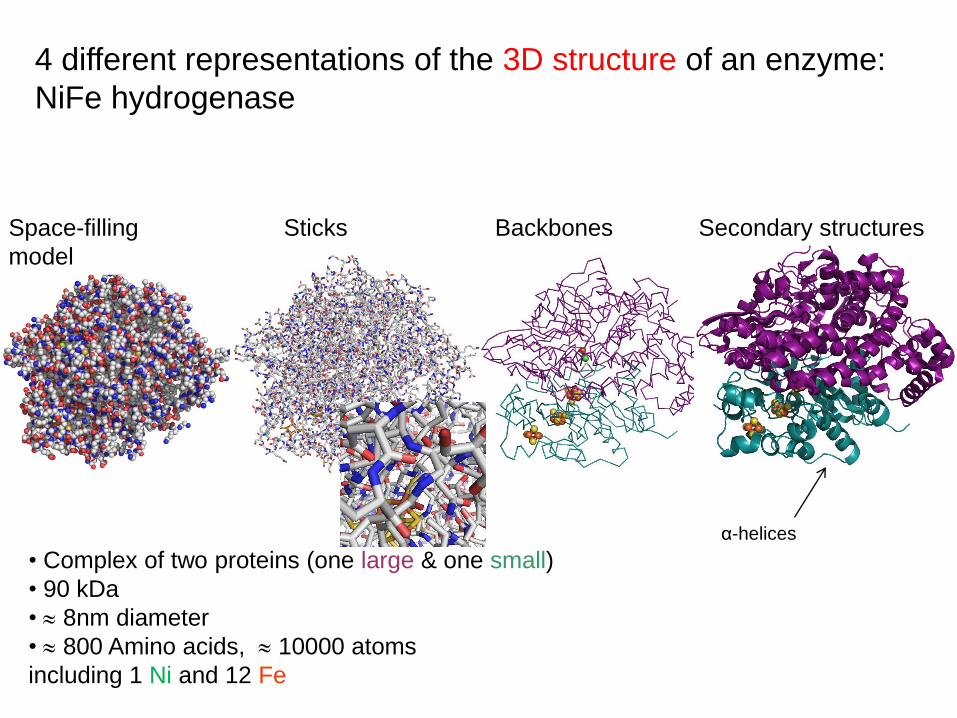
4 different representations of the 3D structure of an enzyme:
NiFe hydrogenase
Space-filling
model
Sticks Backbones Secondary structures
• Complex of two proteins (one large & one small)
• 90 kDa
• 8nm diameter
• 800 Amino acids, 10000 atoms
including 1 Ni and 12 Fe
α-helices

The protein may bind redox cofactors (= prosthetic groups)
required for the protein’s biological activity
FAD Heme Copper site FeS cluster

The protein may bind redox cofactors (= prosthetic groups)
required for the protein’s biological activity
Organic cofactors (Mw < 1 kDa), e.g NAD(P), quinones,
FAD (flavin)
e.g. in the enzyme « glucose oxidase »
quinone (oxidized) /
semiquinone (half reduced) /
hydroquinone (2-electron reduced)

ISE 2010: electrochemistry of cytochromes, see the talks of
Sean J. Elliott, Tuesday, Symp 3, 11:20, Yoon-Bo Shim, Tuesday, symp3, 17:20
Franziska Wegerich, Thursday, symp 11, 15:20, G. Almeida, Monday, symp 3, 11:20
Organic cofactors with bound metal ionsHemes = Fe bound to porphyrin,
in proteins (e.g. cytochrome c, hemoglobin) or enzymes (e.g.
peroxydases)
here the heme of a c-type cytochrome
FeIII/II
His
Met

Inorganic cofactors, e.g. copper sites
in proteins (e.g. azurin) or enzymes (e.g. laccase, billirubin oxidase)
here the type 1 (« blue ») copper site of azurin
His
His
Cys
Met
CuII/I
ISE 2010: electrochemistry of multicopper oxidases, see the talks of
Y. Beyl, Wednesday, Symp 14, 09:40, P. Atanassov, Wednesday, Symp 14, 10:40,
S. Shleev, Monday, Symp 3, 11:00

Inorganic cofactors, e.g. FeS clusters
in proteins « ferredoxins » and enzymes (e.g. hydrogenases)
here a typical [4Fe4S] cluster, the most common FeS cluster
Also [2Fe2S], [3Fe4S] and also much more complex clusters…
[4Fe4S]2+ /+
(2Fe3+,2Fe2+) : [4Fe4S]2+
(Fe3+,3Fe2+): [4Fe4S]+
Cys
Cys
Cys
Cys
4 Fe ions

The biological redox scale at pH 7
(in OEC)
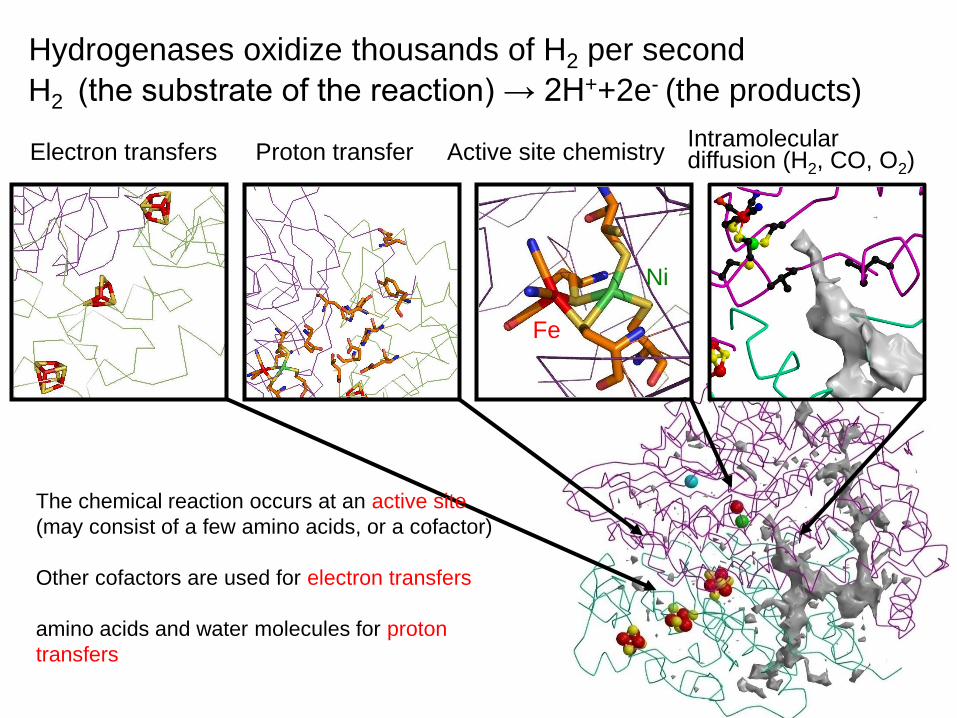
Hydrogenases oxidize thousands of H2 per second
H2 (the substrate of the reaction) → 2H++2e- (the products)
Active site chemistryProton transferElectron transfersIntramolecular diffusion (H2, CO, O2)
Ni
Fe
The chemical reaction occurs at an active site
(may consist of a few amino acids, or a cofactor)
Other cofactors are used for electron transfers
amino acids and water molecules for proton
transfers

Take home message 1
Redox enzymes are large, complex
and very efficient molecular catalysts

Efficient interfacial electron transfer requires that the active
site or at least one redox cofactor is exposed at the surface
of the protein
II) Electrodes for adsorbing redox proteins and enzymes.Case study: adsorbing hydrogenases
Connection with electrodes at the distal cluster
ISE 2010: see the talk of Elisabeth Lojou: Monday, symp 3, 14:40

Examples of electrodes for wiring hydrogenases
1) Au covered with carbon black
INTRODUCTION: « The investigation of enzymes as catalysts for
electrochemical processes is a novel area of chemical enzymology. (…)
Besides, it is also of interest to study electrochemically the mechanism of the
redox enzyme action ».
ISE 2010: see the talk of Arkady Karyakin, Wednesday, Symp 3, 10:00
Yaropolov et al, Bioelectrochem Bioenerg. 12 267 (1984)

Amstrong et al., Chem. Soc. Rev. 26 169 (1997)
Blanford et al., J. Solid. State Electrochem. 10 830 (2006)
SEM image after abrasion Formation of a proteic filmGraphene layers
2H+
electrons
H2
1μm
Examples of electrodes for wiring hydrogenases
2) Pyrolytic graphite “edge”

2H+
electrons
H2
H2ase de A. vinosum adsorbed on stationary PGE electrode,
under 1 atm. Ar
Pershad et al, Biochemistry 38 8992 (1999)
Examples of electrodes for wiring hydrogenases
2) Pyrolytic graphite “edge”
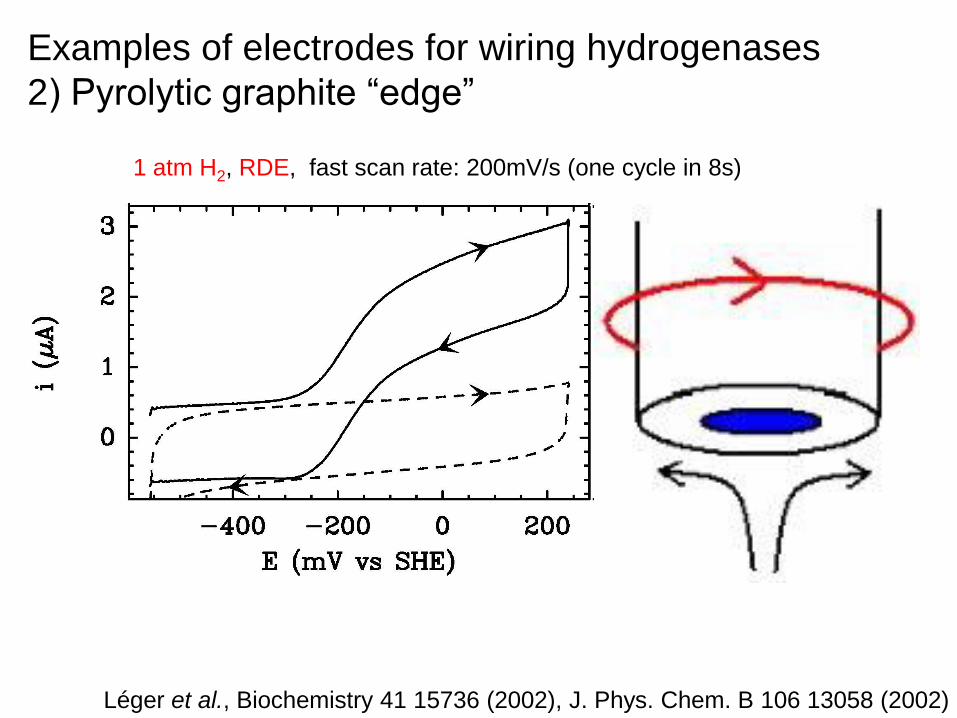
1 atm H2, RDE, fast scan rate: 200mV/s (one cycle in 8s)
Léger et al., Biochemistry 41 15736 (2002), J. Phys. Chem. B 106 13058 (2002)
Examples of electrodes for wiring hydrogenases
2) Pyrolytic graphite “edge”

1atm H2, RDE, slow scan rate: 0.3mV/s (this cycle in 1h30)
Jones et al., J. Am. Chem. Soc. 125 8505 (2003)
Fourmond et al., J. Am. Chem. Soc. 132 4242 (2010)
Examples of electrodes for wiring hydrogenases
2) Pyrolytic graphite “edge”
Inactive
Oxidation
Reduction
Active

Glutamate residues near the surface
exposed 4Fe4S cluster (carboxylates)
Favors favorable orientation
Rüdiger et al., J. Am. Chem. Soc. 127 16008 (2005)
NO2
NO2
NO2
Allongue et al.,
J. Am. Chem. Soc. 119 201
(1997)
Protein SurfaceCarbodiimide
Amide bond
Examples of electrodes for wiring hydrogenases
3) Covalent linkage on functionalized graphite
Carbodiimide coupling for covalent attachment
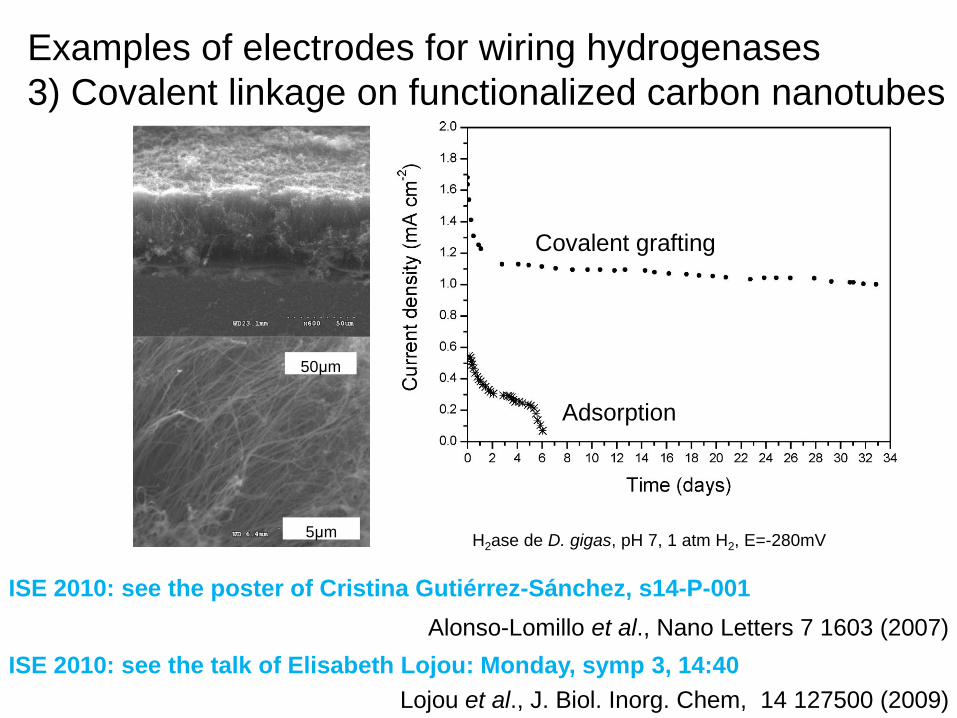
Examples of electrodes for wiring hydrogenases
3) Covalent linkage on functionalized carbon nanotubes
Alonso-Lomillo et al., Nano Letters 7 1603 (2007)
Adsorption
Covalent grafting
H2ase de D. gigas, pH 7, 1 atm H2, E=-280mV
50μm
5μm
ISE 2010: see the poster of Cristina Gutiérrez-Sánchez, s14-P-001
Lojou et al., J. Biol. Inorg. Chem, 14 127500 (2009)
ISE 2010: see the talk of Elisabeth Lojou: Monday, symp 3, 14:40
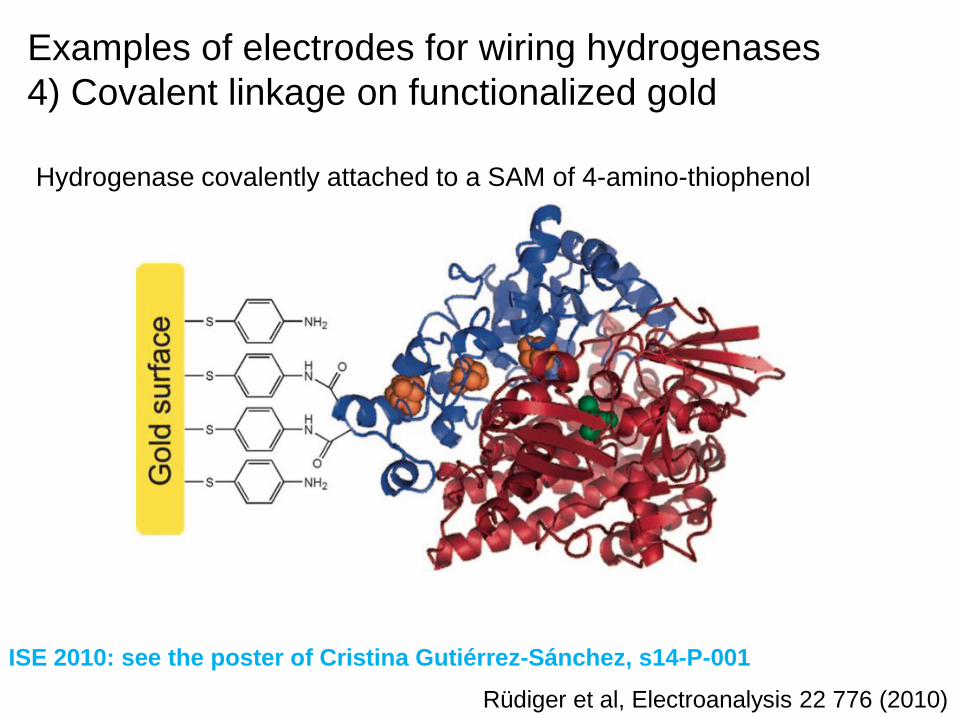
Rüdiger et al, Electroanalysis 22 776 (2010)
Hydrogenase covalently attached to a SAM of 4-amino-thiophenol
ISE 2010: see the poster of Cristina Gutiérrez-Sánchez, s14-P-001
Examples of electrodes for wiring hydrogenases
4) Covalent linkage on functionalized gold

Examples of electrodes for wiring
hydrogenases
5) SAM on Au
→ ready for in situ spectroscopy
D. vulg
aris
Miy
azaki H
2ase, pH
5.5
, 1m
V/s
Millo et al, J. Phys. Chem. B 113 15344 (2009)
Self assembled
monolayer
(SAM) of thiols
HS-(CH2)6-NH2
Ar
H2

Hoeben et al., Langmuir, 24 5925 (2008)
Au + PM + H2ase A. vinosum,
pH 9, 1mV/s
0.13 - 1.3 pmol/cm2
(from kcat en solution)
Mica + Au(111) + PM + H2ase A. vinosum
Image AFM (tapping mode, in air)
0.23 pmol/cm2
PM
Examples of electrodes for wiring hydrogenases
6) Gold nanoelectrode covered with polymyxin
ISE 2010: see the poster of H.A. Heering s14-P-026

Hoeben et al, ACS NANO 12 2497 (2008)A vinosum H2ase, pH 6, 1.5mV/s
Si + 500nm SiO2 + 30nm Au
+ 300nm PMMA
100x100 nm2
Examples of electrodes for wiring hydrogenases
6) Gold nanoelectrode covered with polymyxin
→ studying hydrogenase at the single molecule level
(nearly there)
ISE 2010: see the poster of H.A. Heering s14-P-026
(1 atm Ar)

hydrophobic physiological substrate, interacts with T1 copper site
Blanford et al, Chem Comm 1710 (2007)
Mono- vs multilayers: see M. Sosna et al, Phys. Chem. Chem. Phys 12 10018 (2010)
An electrode for wiring laccasesReviews: M. Sosna et al, Phys. Chem. Chem. Phys 12 10018 (2010), and also Shleev et al, Biosens Bioelectron 20 2517 (2005)
7) anthracene mono(?)layer on PGE or GC
ISE 2010: talks of T. Ruzgas, Mon, Symp 3, 14:00, & S. Shleev, Mon, Symp 3, 11:00

8) Electrodes for plugging into
heme-enzymes, e.g. horseradish peroxydase (HRP)
L. Gorton et al, Anal. Chim. Acta 400 91 (2000)
H. Zimmermann et al, Chem. Eur. J. 6 592 (2000)
FAD- and NADH-dependent enzymes: Willner et al, J. Am. Chem. Soc. 118 10321 (1996)
Kayats et al, J. Am. Chem. Soc. 124 14724 (2002)

9) Using natural or engineered surface exposed
cysteins or histidines for attachment on goldhere horseradish peroxidase
Kartashov et al, PCCP 12 10098 (2010)
Hasan et al, J. Biol. Inorg. Chem. 11 651 (2006)
ISE 2010: see the poster of E. Ferapontova s03-P-047

Take home message 2
Adsorbing the protein or enzyme may be easy,
You must make sure that it is not denatured
by the proximity of the electrode surface
- redox proteins:is the reduction potential consistent with independent determinations (if
available, e.g. potentiometric titrations followed by spectroscopy)
- redox enzymes: simply add the substrate!
is the enzyme still able to catalyse the physiological reaction at a
significant rate, and in a reasonable range of electrode potential?

Part III.
Non catalytic voltammetry (redox proteins and enzymes)
Slow scan voltammetry to measure reduction potentials
Fast scan voltammetry to measure rates of interfacial ET,
Fast scan to measure rates of reactions coupled to ET

Voltammetry of a diffusive redox species
Bard and Faulkner, Electrochemical methods, Wiley, 2004

Example: measuring the reduction potential of a cytochrome
Rhodopseudomonas
palustris
cytochrome c2
~100 residues,
1 surface exposed
heme,
Mw=14kDa
A single redox transition
FeIII/FeII
Geremia et al, Protein Sc. 11 6 (2002)

Example: measuring the reduction potential of a cytochrome
Rhodopseudomonas
palustris
cytochrome c2
at a 4-mercaptopyridine surface-
modified Au electrode
Peak separation
59±2 mV
C=0.2mM,
V=0.5mL,
that is 100nmol of
protein (1.5mg)
Battistuzzi et al, Biochemistry 36 16247 (1997)

Voltammetry of an adsorbed redox species
Bard and Faulkner, Electrochemical methods, Wiley, 2004

Example : a type 1 « blue » copper site
Pseudomonas
aeruginosa azurin
~120 residues,
1 surface exposed
copper site
Mw=14kDa
a single redox transition
CuII/CuI

Example : a type 1 « blue » copper site
Pseudomonas
aeruginosa azurin
adsorbed on PGE
CV recorded at
ν=50mV/s
Jeuken et al. J. Phys. Chem. B. 106 2304 (2002)
ISE 2010: see the talk of Lars Jeuken, Monday, Symp 3, 17:20

The case of a single two-electron redox couple (e.g. FAD)3 redox states O (ox), I (intermediate) and R (reduced)
E0O/I - E0
I/R = 0.4 to -.2 V
Plichon and Laviron, J. Electroanal. Chem. 71 143 (1976)

Example: the 2-electron signal of the flavin in
glucose oxidase
Aspergillus niger
glucose oxidase
~600 residues,
70kDa
1 buried FAD
4 glycosylation sites
16-25 wt %
pdb 1CF3 , Wohlfahrt et al, Acta Crystallogr. D. 55 969 (1999)
ISE 2010: see the talk of Nicolas Mano: Monday, Symp. 3, 10:00

ISE 2010: see the talk of Nicolas Mano: Monday, Symp. 3, 10:00
Example: the 2-electron signal of the flavin in
glucose oxidase
Courjean et al, Angew. Chem. Int. Ed. 48 5897 (2009)
(Adsorbed enzyme catalyses glucose oxidation)
Aspergillus niger
glucose oxidase
~600 residues,
70kDa
1 buried FAD
at a glassy carbon electrode
after enzymatic
deglycosylation
CV recorded at ν=20mV/s
WHH=52mV
hence 2-electron
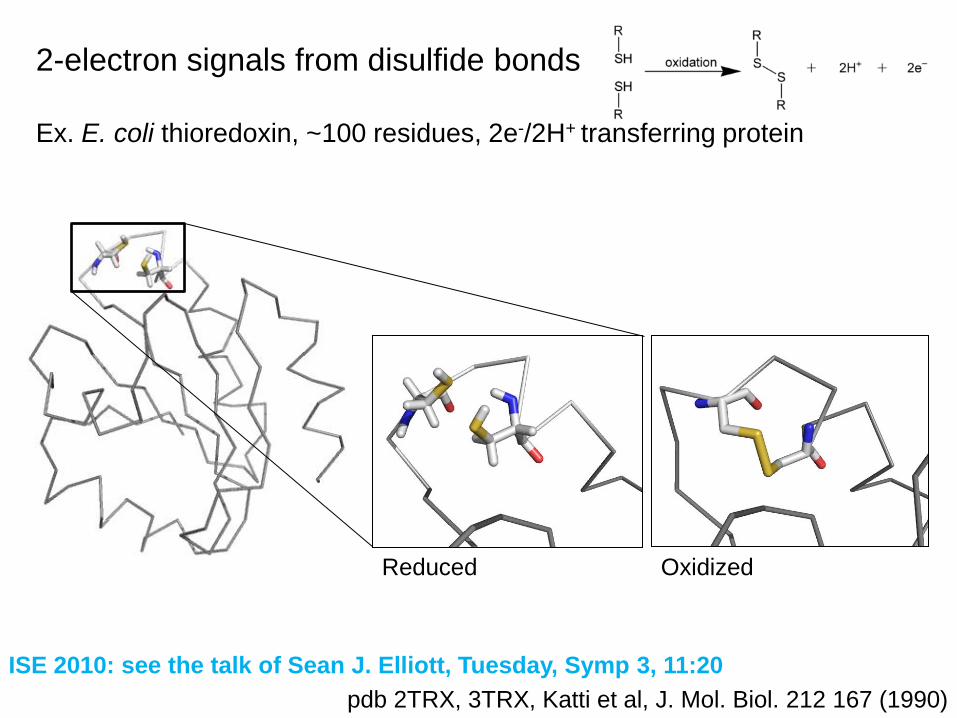
2-electron signals from disulfide bonds
Ex. E. coli thioredoxin, ~100 residues, 2e-/2H+ transferring protein
pdb 2TRX, 3TRX, Katti et al, J. Mol. Biol. 212 167 (1990)
OxidizedReduced
ISE 2010: see the talk of Sean J. Elliott, Tuesday, Symp 3, 11:20

2-electron signals from disulfide bonds
Comparison of various thioredoxins adsorbed on PGE
ISE 2010: see the talk of Sean J. Elliott, Tuesday, Symp 3, 11:20
Chobot et al, Angew. Chem. Int. Ed. 46 4145 (2007)
-60mV/pH

Azodobacter vinelandii
ferredoxin I
~ 100 residues
2 redox centers
[3Fe4S] +/0 and 0/2-
and [4Fe4S] 2+/+
(3 redox couples)
Schipke et al, Biochemistry 38 8228 (1999)
Example: a ferredoxin containing [3Fe4S] and [4Fe4S] clusters

Example: a ferredoxin containing [3Fe4S] and [4Fe4S] clusters
Azodobacter vinelandii
ferredoxin I
~ 100 residues
2 redox centers
[3Fe4S] +/0 and 0/2-
and [4Fe4S] 2+/+
(3 redox couples)
Here at a PGE electrode
Armstrong et al, Biochem. J. 264 265 (1989)

Iverson et al, Science 284 1961 (1999)
Example
E. coli fumarate
reductase,
2 proteins600+200 residues
Mw=100kDa
1FAD,
[2Fe2S]
[4Fe4S]
[3Fe4S] surface exposed
In the case of large enzymes, non catalytic signals are small

In the case of large enzymes, non catalytic signals are small
Léger et al, Biochemistry 2001
Example
E. coli fumarate
reductase,
2 proteins600+200 residues
Mw=100kDa
1FAD,
[2Fe2S]
[4Fe4S]
[3Fe4S] surface exposed
Here at a PGE electrode

pdb 3ABG, Mizutani et al, Acta Crystallogr. F, 66 765 (2010)
Example
Billirubin
oxidoreductase
500 residues
1 trinuclear copper site
(active site for O2
reduction)
1 surface exposed
type-I copper site
(for electron transfer)
In the case of large enzymes, non catalytic signals are small

Ramirez et al, Biochim Biophys Acta, 1777 1364 (2008)
ISE 2010: see the talk of Sergey Shleev, Monday, Symp 3, 11:00
Example
Billirubin
oxidoreductase
500 residues
1 surface exposed
type-I copper site
(for electron transfer)
1 trinuclear copper site
(active site for O2
reduction)
In the case of large enzymes, non catalytic signals are small
here on bare spectrographic graphite

Slow scan voltammetry to measure reduction potentials
The relation with protonation and ligand binding
Fast scan voltammetry to measure rates of interfacial ET,
Measuring rates of chemical reactions coupled to ET
Part III.
Non catalytic voltammetry (redox proteins and enzymes)

Reduction potentials are affected by ligand binding
ligand = proton, substrate, inhibitor, or even the apo-protein
one-electron one-proton coupled electron transfer:
« square scheme » and the corresponding pourbaix diagram
n-electron reduction coupled to the uptake of m protons:

Example: the effect of pH on the reduction potential of
the type-1 copper site in azurin
Jeuken et al., J. Biol. Inorg. Chem. 4 257 (1999)
ISE 2010: see the talk of Lars Jeuken, Monday, Symp 3, 17:20

Example: complete pourbaix diagram of a
« Rieske-type » 2Fe2S ferredoxin
Zu et al, JACS 123 9906 (2001)

Slow scan voltammetry to measure reduction potentials
The relation with protonation and ligand binding
The relation with electron transfer rates
Fast scan voltammetry to measure rates of interfacial ET,
Measuring rates of chemical reactions coupled to ET
Part III.
Non catalytic voltammetry (redox proteins and enzymes)

Intramolecular electron transfer chain in redox enzymes
are sometimes « roaller coasters »
E.g. NiFe hydrogenase:
E0+50mV
E0-350mV
E0-350mV[4Fe4S]2+/+
[4Fe4S]2+/+
[3Fe4S]+/0
Page et al, Nature 402 47 (1999)

Electron transfer betwen two distant redox centers

Marcus’ « inverted region » is very important in biology
Dutton et al, J. Bioenerg. Biomemb. 27 263 (1995)

Interfacial electron transfer, betwen a metallic electrode and a redox center

Electron transfer betwen a metallic electrode and a redox center
Chidsey, Science 251 919 (1991), plotted by Jeuken in Biochim Biophys Acta 1604 67 (2003)
η=E-E0 (V)
(ferrocene terminated thiols self-assembled on Au)

Slow scan voltammetry to measure reduction potentials
The relation with protonation and ligand binding
The relation with electron transfer rates
Fast scan voltammetry to measure rates of interfacial ET,
Measuring rates of chemical reactions coupled to ET
Part III.
Non catalytic voltammetry (redox proteins and enzymes)

ΔEp=0 at small scan rate
(ideally, but residual
small peak separation is
common)
ΔEp is all the greater than
ν is large and/or k0 small
The peaks separate
when
NB: since ΔEp depends on
log(ν/k0), only the order of
magnitude of k0 can be
determined
Measuring k0 from peak splitting against scan rate
Laviron, JEAC 101 19 (1979)
Hirst et al, Anal. Chem. 70 5062 (1998)

Reported values of k0 range from…
very small (~ 0.1s-1) or slower
to very large (up to ~ 10 000 s-1)
Exemple
c555m=
type-1cytochrome
from A. aeolicus.
has a flexible tail
(62 residues)
with terminal cysteine
c555s = soluble version,
no tail
Baymann et al, FEBS Letters, 539 91 (2003)
SAM of hexanethiol on gold

Reported values of k0 range from…
very small (~ 0.1s-1) or slower
to very large (up to ~ 10 000 s-1)
Baymann et al, FEBS Letters, 539 91 (2003)
Two different attachements of the same cytochrome c555
c555m = with the tail attached to Au
c555s = no tail (cleaved before adsorption)
0.01 0.1 1 10 100 1000
0.1
0.2
0.3 c555s
c555m
peak p
ositio
n (
mV
)
scan rate (V/s)
@ 8V/s

Slow scan voltammetry to measure reduction potentials
The relation with protonation and ligand binding
The relation with electron transfer rates
Fast scan voltammetry to measure rates of interfacial ET,
Electronic coupling with the electrode
Reorganization energies and gated electron transfers
Measuring rates of chemical reactions coupled to ET
Part III.
Non catalytic voltammetry (redox proteins and enzymes)

How k0 depends on distance from the
electrode
k0 measured by electrochemistry or spectroelectrochemistry in: Song et al, J. Phys. Chem. 97 6564 (1993)
Murgida et al, JACS 123 4062 (2001), Chi et al, J. Phys. Chem. B 105 3088 (2001)
Reviewed in: Jeuken, Biochim Biophys Acta 1604 67 (2003)
Murgida et al, ChemPhysChem 11 1225 (2010)

How k0 depends on distance from the electrode
Conformational rearrangement or thermal fluctuations
may limit the rate of (i.e. « gate ») interfacial ET
Reviewed in: Jeuken, Biochim Biophys Acta 1604 67 (2003)
Chi et al, PNAS 102 16203 (2005)
Murgida et al, ChemPhysChem 11 1225 (2010)
ISE 2010: see the talk of Q. Chi, Thursday, Symp 3, 17:00

Upper limit to ET rate at short distance or high overpotential
decreases the apparent value of λ, when the data are
analyzed with the Marcus model
Jeuken, Biochim Biophys Acta 1604 67 (2003), and refs therein
Jeuken et al, J. Phys. Chem. B, 106 2304 (2002)

Slow scan voltammetry to measure reduction potentials
The relation with protonation and ligand binding
The relation with electron transfer rates
Fast scan voltammetry to measure rates of interfacial ET,
Electronic coupling with the electrode
Reorganization energies and gated electron transfers
An evidence for kinetic dispersion
Measuring rates of chemical reactions coupled to ET
Part III.
Non catalytic voltammetry (redox proteins and enzymes)

Davis et al, J. Phys. Chem. B, 110 20651 (2006)
Salverda et al, Angew. Chem. Int. Ed. 49 1 (2010)
Azurin adsorbed on Au + SAM of hexanethiol
The protein is labeled with a dye
whose fluorescence is quenched
by nonradiative Energy Transfer (FRET)
when the Cu site is oxidized
Fluorescence detected using a TIRF microscope
ISE 2010: see the poster of H A Heering, s14-P-026
An evidence for kinetic dispersion

Salverda et al, Angew. Chem. Int. Ed. 49 1 (2010)ISE 2010: see the poster of H A Heering, s14-P-026
An evidence for kinetic dispersion

Slow scan voltammetry to measure reduction potentials
The relation with protonation and ligand binding
The relation with electron transfer rates
Fast scan voltammetry to measure rates of interfacial ET,
Electronic coupling with the electrode
Reorganization energies and gated electron transfers
An evidence for kinetic dispersion
Measuring rates of chemical reactions coupled to ET
e.g. protonations
Part III.
Non catalytic voltammetry (redox proteins and enzymes)

Proton transfer is a fundamental process in bioenergetics
Respiratory
enzyme
ATP
Synthase
Reduced substrate O2
H+
H+
ATP + H2O ADP + Pi
Peter Mitchell (1920-1992, Nobel 1978): ATP synthesis uses a gradient of proton concentration
« Chemiosmotic theory » of oxidative phosphorylation
electronsprotons
Inner mitochondrial membrane
driving force for
ATP synthesis:
[H+] gradient
Nicholls and Ferguson « Bioenergetics 3 » Academic Press (2002)
Hydrolysis ΔrG<<0

Proton transfer to the buried [3Fe4S] cluster of a ferredoxin
The pH-dependence of the reduction potential of the [3Fe4S]+/0
demonstrates proton coupled ET, pKred ~ 7.5
The [3Fe4S] cluster is not exposed to solvent
Surface exposed glutamateChen et al, Nature 405 814 (2000)
Camba et al, Biochemistry 42 10589 (2003)
Hirst et al, J. Am. Chem. Soc. 120 7085 (1998)

Proton transfer to the buried [3Fe4S] cluster of a ferredoxin
Molecular dynamics simulations show that the surface exposed glutamate
can approach the clusterChen et al, Nature 405 814 (2000)
Camba et al, Biochemistry 42 10589 (2003)
Hirst et al, J. Am. Chem. Soc. 120 7085 (1998)

Camba et al, Biochemistry 42 10589 (2003)
Chen et al, Nature 405 814 (2000)
Hirst et al, J. Am. Chem. Soc. 120 7085 (1998)
Properties of the D15E mutant
Proton transfer to the buried [3Fe4S] cluster of a ferredoxin
koff (s-1) kon (s
-1M-1)
WT 300 8 109
D15E 2.5 2 107
D →E

Part IV.
Catalytic voltammetry (enzymes)
Using direct electrochemistry to measure the enzyme’s activity
(turnover rate)
and learning about all aspects of the catalytic mechanism:
• active site chemistry
• interfacial electron transfer
• intramolecular electron transfer
• slow (in)activation processes
• substrate transport along substrate channels
• …

H+ H2
Reduced MV
(blue)
Oxidized methyl
viologen
(color-free)
electrons
Solution assay of the activity (=turnover rate) of
hydrogenase
Easy experiment but:
- Characteristic time of the
measurement: 10-20 minutes (steady
state conditions)
- The driving force is not usually varied

H+ H2
Current = [enzyme] x turnover
Reference electrode
Working electrode
Counter electrode
H2
Rotating disc electrode
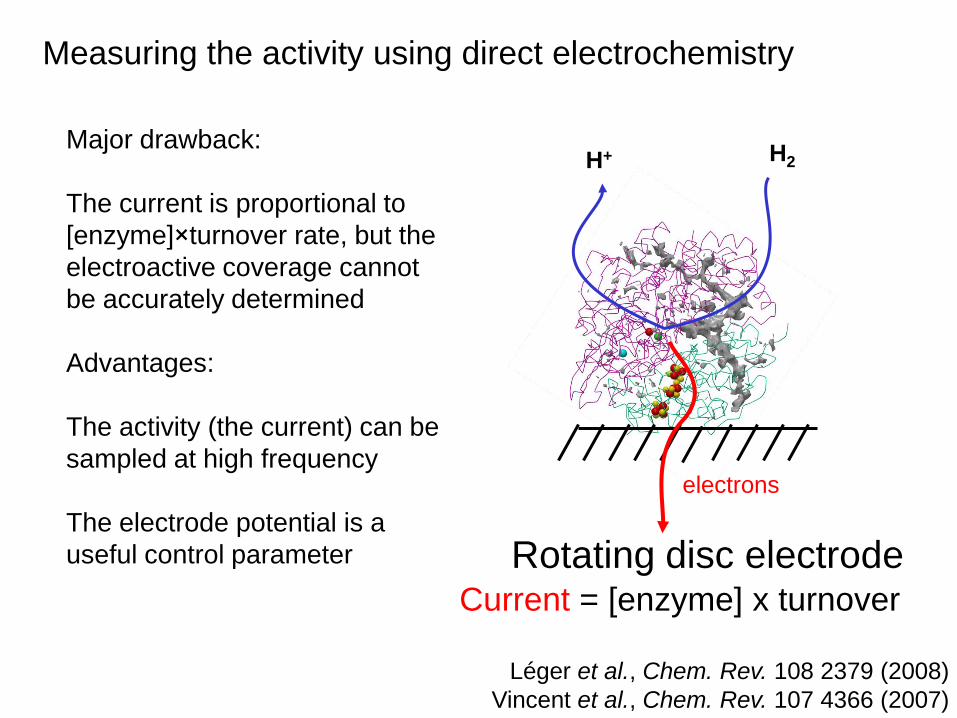
Measuring the activity using direct electrochemistry
electrons
RDE
Léger et al., Chem. Rev. 108 2379 (2008)
Vincent et al., Chem. Rev. 107 4366 (2007)
H+ H2
Current = [enzyme] x turnover
Rotating disc electrode
Léger et al., Chem. Rev. 108 2379 (2008)
Vincent et al., Chem. Rev. 107 4366 (2007)
electrons
Major drawback:
The current is proportional to
[enzyme]×turnover rate, but the
electroactive coverage cannot
be accurately determined
Advantages:
The activity (the current) can be
sampled at high frequency
The electrode potential is a
useful control parameter
Measuring the activity using direct electrochemistry

The dependance of activity on electrode potential
may be complex, we must make it informative
Taken from: Léger et al, Biochemistry 40 11234 (2001), Heffron et al, Biochemistry 40 3117 (2001), Fourmond et al, JACS. 132 4848 (2010)
Hudson et al, JACS 127 6977 (2005), Fourmond et al, J. Phys. Chem. B 114 3341 (2010), Léger et al, J. Phys Chem. B 106 13058 (2002)
Fumarate reductase
oxidizing succinate (top)
reducing fumarate
(bottom)
DMSO (top) and
nitrate (bottom)
reductasesHydrogenases
100mV
All at RDE,
rotating at a high
rate ( > 3 krpm)

Succinate oxidation by E. coli fumarate reductase (at RDE)
electrode
Fumarate
Current = turnover rate
Succinate
Léger et al., Biochemistry 40 11234 (2001)

Michaelis-Menten kinetics, Michaelis constant Km
Henri, C.R. Acad. Sc.135 916 (1902)
Michaelis et al., Biochemisches Zeitschrift 49 333 (1913)
Michaelis-Menten equation
[Succinate]
[Succinate] / Km

Michaelis-Menten kinetics for i
lim
Léger et al., Biochemistry 40 11234 (2001)
Succinate oxidation by E. coli fumarate reductase (at RDE)

The dependence of current on E gives the original information: the reduction potential of the AS under turnover conditions
Léger et al., Biochemistry 40 11234 (2001)
Succinate oxidation by E. coli fumarate reductase (at RDE)
Varying the concentration of succinate
at constant pH

Pourbaix and pourbaix-like diagrams for the change in
E0(FAD) against pH and [succinate] give
acidity and dissociation constants of the active site
Léger et al., Biochemistry 40 11234 (2001)
(@ 1mM [succinate]) (@ pH 7)

In many cases, the current does not reach a plateau at
high overpotential but rather varies in proportion to E
Léger et al, J. Phys Chem. B 106 13058 (2002)
Reveals distribution of orientations
(and thus interfacial ET rates)
Hydrogen oxidation by A. vinosum NiFe
hydrogenase, 3krpm, pH7, 1 bar H2

fitdata
Léger et al, section 2.2.5.4 in Chem Rev 108 2379 (2008)
and J. Phys Chem. B 106 13058 (2002)
Modelisation:
1) choose model for interfacial ET
2) current as a function of k0
3) distribution of orientations
4) distribution of k0
5) current integrated across all values of k0

The case of slow intramolecular ET
(between electron relay and active site)
Exemple of enzyme
with a single relay:
sulfite oxidase
Heme
+ Mo active site
oxidizes sulfite to sulfate
Kisker et al, Cell 91 973 (1997)

The position of the wave when the enzyme has one redox relay
kf
kb ele
ctr
odeAS Rk-1
k1
Léger et al, J. Am. Chem. Soc. 128 180 (2006)

The model predicts a normal current equations but with
reduction potential that takes apparent values
(if k0
very fast)
kithe slowest intramolecular ET rate cst
Léger et al, J. Am. Chem. Soc. 128 180 (2006)
Fast ET Slow ET

Application to sulfite oxidase (one heme mediates electrons from Mo active
site, where sulfite SO32- is oxidized to sulfate SO4
2-)
Using kcat
=95 s-1 , one finds ki=350 s-1
Flash photolysis experiments: 800 s-1
(Enemark 1993)
Léger et al, J. Am. Chem. Soc. 128 180 (2006)
Voltammetry: catalytic wave
whose position does not depend much
on pH
Elliott et al, J. Am. Chem. Soc. 124 11612 (2003)E1

Simple diagnosis of slow interprotein ET in cytc/cyt
cc oxidase
complex adsorbed on SAM/Au
Haas et al, J. Phys. Chem. B 105 11351 (2001)
Cytochrome c oxidase
cytochrome c

Fast scan rate: 500mV/s
Slow scan rate: 5mV/s
The current (activity) drops at high
potential (formation of an oxidized
inactive state called NiB) and it is
recovered on the return scan
(reductive reactivation)
at pH 7, 1bar H2, 40oC, RDE
Oxidative inactivation of NiFe hydrogenase,
and the meaning of the « switch potential »
Inactive (“NiB”)
Oxidation
Reduction
Active

E0 values: M. Pandelia et al., Unpublished .
Voltammetry of A. aeolicus NiFe hydrogenase

E0 values: M. Pandelia et al., Unpublished . Fourmond et al, JACS 132 4848 (2010)
Voltammetry of A. aeolicus NiFe hydrogenase

Fourmond et al, JACS 132 4848 (2010)
Chronoamperometry for studying the inactivation mechanism

Fourmond et al, JACS 132 4848 (2010)
Oxidative inactivation of NiFe hydrogenase,
and the meaning of the « switch potential »

Example of complex catalytic wave shape: nitrate reduction by nitrate reductase,
has 2 hemes and 1 [4Fe4S] cluster
and one Mo active site where nitrate is reduced to nitrite
Fourmond et al, Biochemistry 49 2424 (2010)
The electrochemical behavior is mirrored in solution assays where the driving force
provided by reduced MV decreases as a function of time

Frangioni et al, JACS 126 1328 (2004)
Bertrand et al, JPCB 111 10300 (2007)
The electrochemical waveshape may result from the
competition between multiple reaction pathways

The steady-state catalytic signal for nitrate reduction is complex
In addition, strong hysteresis at high nitrate concentration
Fourmond et al, J. Phys. Chem. B 112 15478 (2008)
Fourmond et al, J. Phys. Chem. B 114 3341(2010)

Fourmond et al, J. Phys. Chem. B 114 3341(2010)
Evidence for substrate inhibition at moderately low potential
both in chronoamperometry and solution assays
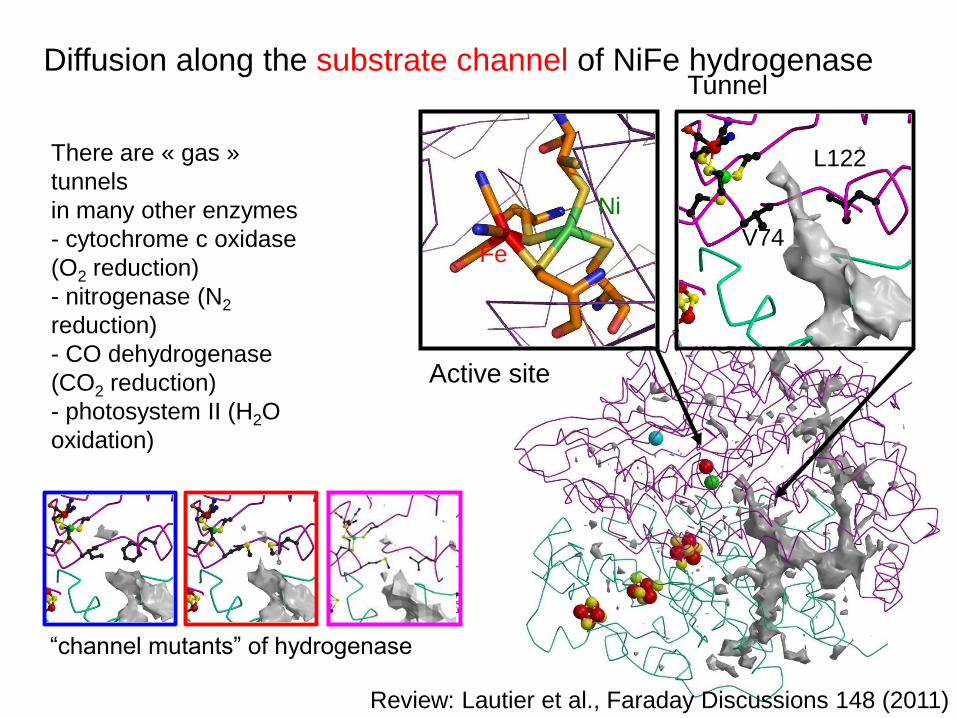
Diffusion along the substrate channel of NiFe hydrogenase
Active site
Tunnel
L122
V74
Ni
Fe
There are « gas »
tunnels
in many other enzymes
- cytochrome c oxidase
(O2 reduction)
- nitrogenase (N2
reduction)
- CO dehydrogenase
(CO2 reduction)
- photosystem II (H2O
oxidation)
“channel mutants” of hydrogenase
Review: Lautier et al., Faraday Discussions 148 (2011)

H2
CO
Kinetics of reaction with CO:
Chronoamperometry at constant electrode potential
Cell flushed with H2
Léger et al, J. Am. Chem. Soc. 126 12162 (2004)

H2
CO
instant inhibition
Kinetics of reaction with CO:
Chronoamperometry at constant electrode potential
Cell flushed with H2, CO injected
Result obtained
with the WT enzyme
Léger et al, J. Am. Chem. Soc. 126 12162 (2004)

H2
Kinetics of reaction with CO:
Chronoamperometry at constant electrode potential
Cell flushed with H2, CO injected, and then flushed away by H2
instant inhibition, reversible
Léger et al, J. Am. Chem. Soc. 126 12162 (2004)
Result obtained
with the WT enzyme

CO
H2
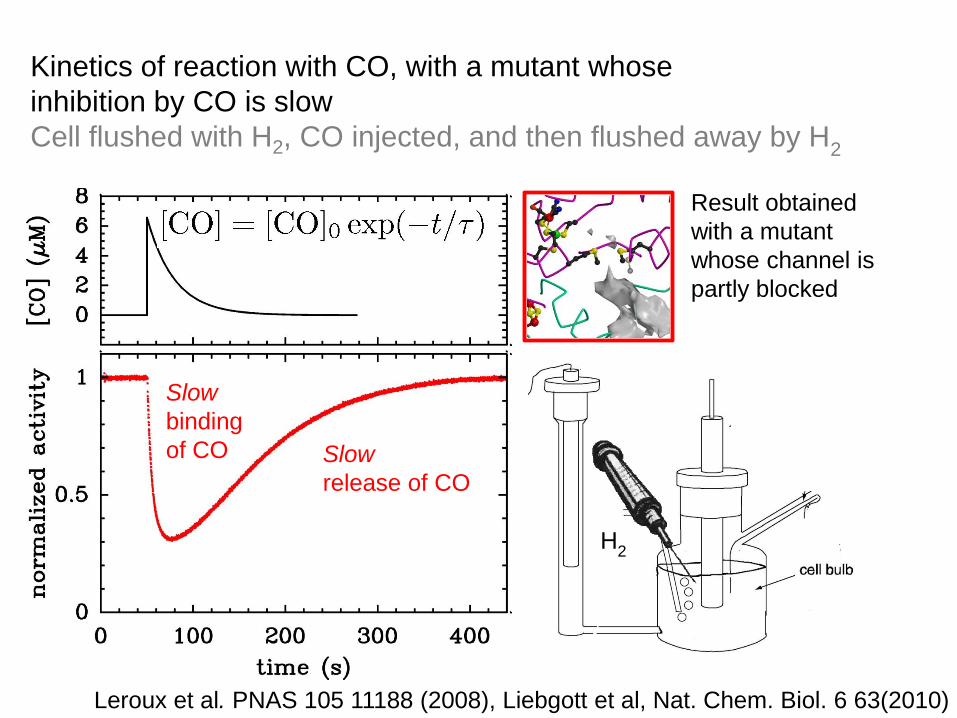
Kinetics of reaction with CO, with a mutant whose
inhibition by CO is slow
Cell flushed with H2, CO injected, and then flushed away by H2
Slow inhibition
→ slow binding of CO
Result obtained
with a mutant
whose channel is
partly blocked
Leroux et al. PNAS 105 11188 (2008), Liebgott et al, Nat. Chem. Biol. 6 63(2010)
H2
Kinetics of reaction with CO, with a mutant whose
inhibition by CO is slow
Cell flushed with H2, CO injected, and then flushed away by H2
Slow
release of CO
Slow
binding
of CO
Result obtained
with a mutant
whose channel is
partly blocked
Leroux et al. PNAS 105 11188 (2008), Liebgott et al, Nat. Chem. Biol. 6 63(2010)

Kinetics of reaction with CO, with a mutant whose
inhibition by CO is slow
Cell flushed with H2, CO injected, and then flushed away by H2
Result obtained
with a mutant
whose channel is
partly blocked
Fit returns
Almeida et al, FEBS Letts 581 284 (2007)
Leroux et al. PNAS 105 11188 (2008), Liebgott et al, Nat. Chem. Biol. 6 63(2010)
and
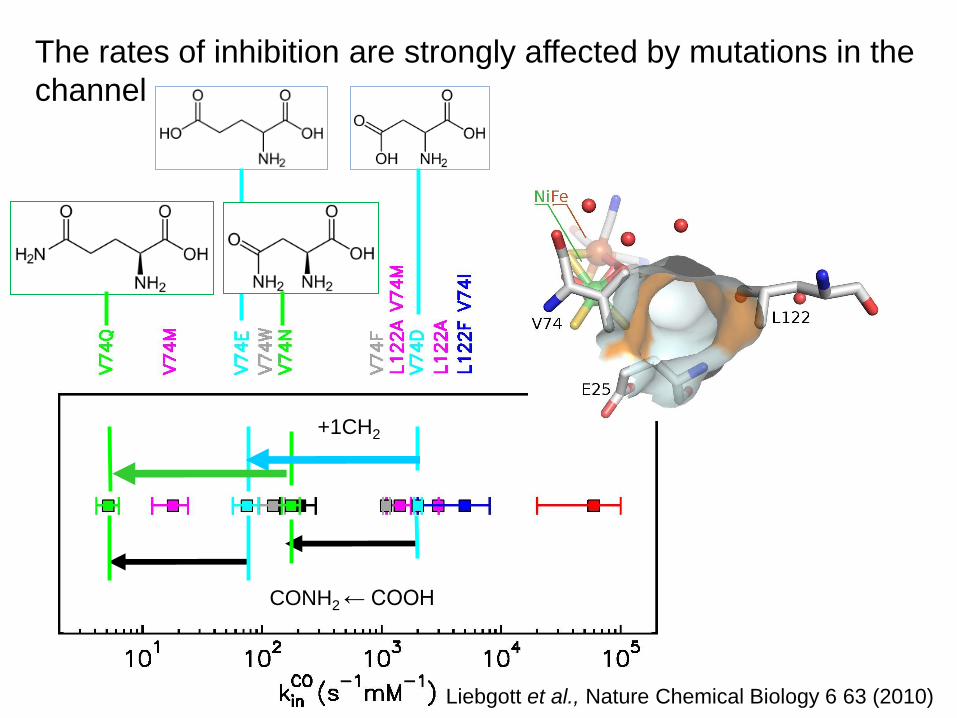
The rates of inhibition are strongly affected by mutations in the
channel
Liebgott et al., Nature Chemical Biology 6 63 (2010)
CONH2 ← COOH
+1CH2

Take home message 3
Electrochemistry has ben used to study all aspects
related to protein function
Active site redox chemistry, substrate binding,
Electron transfer (interfacial, intramolecular),
Proton transfer,
Substrate channels,
(in)activation,
Inhibitition, …
Léger and Bertrand, Chem. Rev. 108 2379 (2008)

Part V.
Using redox enzymes in biotech devices
Intrinsic limitations of DET-based electrodes
Examples of applications:
Biofuel cells (using hydrogenases, GOX, laccases, billirubin oxidases)
Photoelectrochemical fuel cells (hydrogenases)
Particles coupling two redox enzymes
Biosensors (nitrite reductase)

Barton et al, Chem Rev, 104 4867 (2004) (special issue on Batteries and Fuel cells)
Fuel cells and photoelectrochemical cells:
primary challenge is increased biocatalytic power
An ideal monolayer of a typical enzyme
100nm2 cross section, 500 electrons /s → 80μA/cm2
Practical fuel cells operating at > 10mA/cm2
would require thousands of layers
(that is loading in the mg/cm2 range, assuming MW=100kDa)→ mediated electron transfer is more promissing in this respect
Some devices that take advantage of the high selectivity
of the enzymes for their substrates.

1842 1969 2007
E.g. H2/O2 fuel cell
October 2007
special issue on H2
and hydrogenases

Comparison between Pt and hydrogenase adsorbed on
graphite as H2 fuel cell anode catalysts
1/ω1/2 (rpm ½)
Jones et al. Chem Comm. 866 (2002)
A. vinosum H2ase
H2 oxidation at RDE, E=.24V/SHE
C. acetobutylicum H2ase
Oxidation/production of H2 on RD graphite E
Hambourger et al. JACS 130 2015 (2008)
1 bar H2, pH 7, 50mV/s, 3000rpm1 bar H2, pH 7, 45oC, E=0.24V vs SHE
ISE 2010: see the talk of K. Vincent, Thursday, Symp 14, 09:40
Hydrogenase
Pt

H2/O2 fuel cell: avoiding short-circuit requires expensive
membranes
Using selective enzyme electrodes:
Hydrogenase at the anode (H2 oxidation)
laccase or billirubin oxidase at the cathode (O2 reduction)
to design membraneless fuel cells

Vincent et al. Chem. comm. 5033 (2006) & PNAS 102 16951 (2005)
A membraneless H2/O2 biofuel cell using
a hydrogenase that is able to oxidize traces of H2 in air
and a fungal laccase, both connected to PGE
ISE 2010: see the talk of K. Vincent, Thursday, Symp 14, 09:40

Hambourger et al. JACS 130 2015 (2008), Photochem. Photobiol. 81 1015 (2005)
Renewable hydrogen production, in a photoelectrochemical cell
that uses the FeFe hydrogenase from C. acetobutylicum
(instead of Pt)

Kamitaka et al, Phys.Chem.Chem.Phys 9 1793 (2007)
A one-compartment fructose/dioxygen non-mediated biofuel cell
based on fructose dehydrogenase and laccase, up to 0.8 mW/cm2

Enzymatic catalysis on conducting particles
Vincent et al, Nat Chem Biol 3 761 (2007)
Reisner et al, JACS 131 18457 (2009)
H2 as electron source
Photoproduction of H2
~5μm2

Bullen et al, Biosensors and bioelectronics, 21 2015 (2006)
No need to reach high current densities to design biosensors

Silveira et al, Biosensors and bioelectronics 25 2026 (2010)
ISE 2010: see the talk of M. Gabriela Almeida: Monday, symp 3, 11:20
A non-mediated nitrite biosensorbased on penta-heme nitrite reductase NO2
- + 6e- + 8H+ → NH4+ + 2H2O
+ 1 buried
active site heme
4 electron-transfer hemes
2 of which surface exposed

Silveira et al, Biosensors and bioelectronics 25 2026 (2010)
ISE 2010: see the talk of M. Gabriela Almeida: Monday, symp 3, 11:20
A non-mediated nitrite biosensorbased on penta-heme nitrite reductase
encapsulated in a porous silica matrix on graphite
@E= -.5 -.7 and -.9V/AgCl
At -.9V, linear response
in the range 0.12μM to 50 μM nitrite, stable for 2 weeks

Further readings
Willner and Katz, « Integration of layered redox proteins and conductive supports
for bioelectronic applications » Angew Chem Int. Ed. 39 1180 (2000)
Cracknell, Vincent and Armstrong « Enzymes as working or inspirational
electrocatalysts for fuel cells and electrolysis » Chem. Rev. 108 2439 (2008)
Léger and Bertrand « Direct electrochemistry of redox enzymes as a tool for
mechanistic studies » Chem. Rev. 108 2379 (2008)

A free software to analyse electrochemical data
Available for download at : http://bip.cnrs-mrs.fr/bip06/software.html
Works on Mac OS X (one-click installer available) or Linux. Not Windows.
Good for : noise filtering, baseline subtractions, fitting
Fourmond et al, Bioelectrochem. 76 141 (2009)

D. Fructosovorans: Sébastien Dementin, Pierre Pol Liebgott, Marc Rousset
A. Aeolicus: Pascale Infossi, E. Lojou, M.Thérèse Giudici-Orticoni
EPR: Bénédicte Burlat, Emilien Etienne, Bruno Guigliarelli,
Electrochemistry: Pierre-Pol Liebgott, Fanny Leroux, Vincent Fourmond,
Carole Baffert, Pierre Ezzano, Patrick Bertrand
C. acetobutylicum: Thomas Lautier, I. Meynial, Ph. Soucaille (INSA Toulouse)
R. sphaeroides: David Pignol, Monique Sabaty, Pascal Arnoux
Crystallography: Anne Volbeda, Juan Fontecilla Camps (CEA, UJF)
H/D kinetics: Laurent Cournac (CEA, Cadarache)
FTIR: Antonio De Lacey (D. fructosovorans, CSIC, Madrid),
FTIR, EPR: Maria Pandelia, Wolfgang Lubitz (A. Aeolicus, Max Planck, G)
Collaborators
Slides can be downloaded at http://bip.cnrs-mrs.fr/bip06/publications