barab-marszak, 2006
TRANSCRIPT

8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 1/9
Effect of tumor necrosis factor alpha and infliximab on apoptosisof B lymphocytes infected or not with EpsteineBarr virus
Fanny Baran-Marszak a,b,*, Christelle Laguillier a, Ibtissam Youlyouz c, Jean Feuillard c,Xavier Mariette a,d, Remi Fagard b, Martine Raphael a
a INSERM E109, CHU Bicetre, Assistance Publique-Hopit aux de Paris, Universite Paris 11 , 942 75 Le Kremlin Bicetre, Franceb Universite Paris13, EA3406, 93009 Bobigny, France
c UMR CNRS 6101, Facul te de Medecine, CHU Dupuytren, 87000 Limoges , Franced Service de Rhumatologie, CHU Bicetre, Assistance Publique-Hop itaux de Paris , Univers ite Paris 11, 94275 Le K remlin Bicetre, France
Received 1 June 2005; received in revised form 10 March 2006; accepted 19 March 2006
Abstract
Chronic inflammation and immunosuppressive therapies increase the risk of non-Hodgkin’s lymphoma associated or not with EpsteineBarr
virus (EBV) infection. A possible link between infliximab treatment and increased risk of lymphoma has been suggested. Indeed, infliximab
induces apoptosis of monocytes and activated T lymphocytes, but its effect on B lymphocytes infected or not with EBV is unknown. Secreted
tumor necrosis factor (TNF) a and the expression level of TNF receptor 1 (TNFR1) and TNFR2 were compared in EBV-positive and negative B-
cell lines. The impact of TNFa and infliximab on apoptosis of EBV-positive cells was analyzed regarding the activity of NF-kB. Increased ex-
pression of TNFa in EBV-positive cells suggested that infliximab could affect their survival. However, TNFa or infliximab incubation had no
effect on apoptosis of EBV-positive cells. Loss of NF-kB activity sensitized lymphoblastoid cell lines to TNFa-induced apoptosis, but no direct
effect of infliximab on apoptosis was detected. On the basis of our in vitro data, neither TNFa nor infliximab has a direct effect on apoptosis of B
lymphocytes and EBV-positive cell lines. Thus, if an increased incidence of lymphoma were induced by TNFa blockers, it would not involvea direct effect on B cells but rather an impaired immune surveillance by T cells.
Ó 2006 Elsevier Ltd. All rights reserved.
Keywords: Tumor necrosis factor a; Infliximab; B lymphocyte; Apoptosis; EpsteineBarr virus
1. Introduction
The incidence of lymphoma is increased in patients with
autoimmune diseases such as Sjogren’s syndrome, Hashimo-
to’s disease and rheumatoid arthritis (RA) [1e
6]. Most of
this increased risk is linked to the persistent activation of
autoimmune B lymphocytes, as in Sjogren’s syndrome [7], or
to the inflammatory activity of the disease, as in RA [8]. Inter-
estingly, the incidence of lymphoma is also increased in pa-
tients with non-autoimmune chronic inflammatory diseasessuch as pyothorax [9] or Crohn’s disease (CD) [1].
The increased risk of lymphoma in patients with autoim-
mune or inflammatory diseases may be also related to immu-
nosuppressive treatment. The two classical drugs that are
thought to increase this risk of lymphoma are methotrexate
(MTX) and azathioprine. In fact, a few cases of Epsteine
Barr virus (EBV)-associated lymphomas have been reported
with MTX treatment for autoimmune diseases [10,11], which
regressed after withdrawal of the drug, similar to what is ob-
served in EBV-associated lymphoproliferative disorders in
Abbreviations: RA, rheumatoid arthritis; CD, Crohn’s disease; MTX,
methotrexate; EBV, EpsteineBarr virus; TNF, tumor necrosis factor; NF-kB,
nuclear factor kB; LCL, lymphoblastoid cell line; LMP1, latent membrane
protein 1; PBMC, peripheral blood mononuclear cells; NGFR, nerve growth
factor receptor; I-kB, inhibitor of NF-kB; TNFR, TNF receptor; ELISA, en-
zyme-linked immunosorbent assay; PBS, phosphate-buffered saline.
* Corresponding author at: Service d’Hematologie biologique, Hopital Avic-
enne, 125 route de Stalingrad, 93000 Bobigny, France. Tel.: þ33 1 48 95 56
46; fax: þ33 1 48 95 56 28.
E-mail address: [email protected] (F. Baran-Marszak).
1043-4666/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.cyto.2006.03.005
www.elsevier.com/locate/issn/10434666
Cytokine 33 (2006) 337e345

8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 2/9
immunodeficient patients [12]. Thus, in RA with lymphoma,
MTX could be beneficial, resulting from efficient control of
the disease, or deleterious, leading to increased risk of EBV-
associated lymphoproliferations. This latter effect seems mar-
ginal, since in two large prospective studies, treatment with
MTX did not significantly increase the risk of non-Hodgkin’s
lymphoma with RA [6,13], although in one of these studies therisk of Hodgkin’s lymphoma (usually 10 times less frequent
than non-Hodgkin’s lymphoma) was increased in RA patients
treated with MTX.
Tumor necrosis factor a (TNFa) blockers are a new class of
therapeutic agents that are efficient in the treatment of refrac-
tory RA, spondylarthropathies and CD. TNFa has a dual role
and, depending on the cellular context, can induce cell survival
by activating nuclear factor kB (NF-kB) or trigger apoptosis
by activating caspases [14]. An increased secretion of TNFa
is observed in B lymphocytes upon EBV infection but lympho-
blastoid cell lines (LCLs) are resistant to TNFa-induced
apoptosis [15] because the viral latent membrane protein 1
(LMP1) protects against it.Infliximab is a chimeric monoclonal antibody that binds
specifically to human TNFa and neutralizes its biologic prop-
erties, but some effects of infliximab cannot be explained by
the neutralization of soluble TNFa alone [16]. For instance,
it induces the rapid suppression of mucosal inflammation in
CD and the specific increase of apoptosis in T lymphocytes
of the lamina propria in vivo and in vitro. Infliximab also in-
duces apoptosis in synovial monocytes/macrophages in RA
[17] and in CD through a caspase-dependent pathway [18].
Results of a few studies suggested an increased incidence
of lymphoma in RA patients treated with TNFa blockers
[19,20]. However, to date it has not been possible to discrim-inate between an increased incidence of lymphoma due to the
TNFa blockers themselves and an increased incidence because
most severe forms of RA are treated with TNFa blockers.
Such increased incidence, which is not established at the pres-
ent time, could be due either to a defect of immunosurveil-
lance of EBV-infected B cells by T cells or to a direct effect
of TNFa blockers on B cells infected or not by EBV. The pos-
sibility of an increased risk of lymphoma is one of the major
issues concerning the safety of long-term TNFa blocker use.
Only a few data are available concerning the action of
TNFa blockers on B cells infected or not with EBV. The pur-
pose of this work was to determine whether TNFa affects ap-
optosis in EBV-infected B lymphocytes, in Burkitt cell lines
infected or not with EBV and in B cells isolated from periph-
eral blood monoclonal cells (PBMCs), and whether infliximab
has a direct effect on apoptosis in these different types of B
cells in vitro.
2. Materials and methods
2.1. Cell culture
The LCL PRI and the Burkitt lymphoma cell line BL2 and
its EBV-infected counterpart BL2.B98.5 were grown at 37 C
in humidified 5% CO2 air in RPMI 1640 medium containing
GlutaMAX (GibcoBRL, Life Technologies, Cergy-Pontoise,
France) supplemented with 10% decomplemented fetal calf
serum (Dutscher, Brumath, France), 100 U/ml penicillin,
10 mg/ml streptomycin (GibcoBRL), 1 mM sodium pyruvate
(GibcoBRL), MEM vitamins 100Â (GibcoBRL) and 5 mg/ml
plasmocin (Cayla InvivoGen, Toulouse, France). Cells were
treated with TNFa (Invitrogen, Carlsbad, CA, USA), 10 ng/ ml for 30 min or 24 h, and infliximab (RemicadeÒ, Scher-
ing-Plough, Levallois-Perret, France), 5 mg/ml for 24 h.
2.2. Plasmid and transfection
The VINL Ik-Ba32/36a episomal vector was derived from
the previously described CKR 516 [21] vector by replacing the
enhanced green fluorescent protein-inducible marker with the
truncated version of the nerve growth factor receptor (NGFR)
lacking the cytoplasmic domain. This vector contains the
EBNA1 gene to ensure episomal replication, a bidirectional
tetracycline-inducible promoter driving the expression of two
independent cDNAs: Ik-Ba32/36a and NGFR, as a markerof induction. The expression of NGFR was determined by
flow cytometry and used to select cells with magnetic beads.
The induction of Ik-Ba32/36a was verified by Western
blotting.
A total of 1 mg vector/106 cells was transfected by double-
pulse electroporation (first pulse: 750 V, 25 mF and 201 U; sec-
ond pulse: 125 V, 3000 mF and 99 U). After 3 days of culture,
hygromycin-resistant cells (stable transfectants) were selected
(three weeks) by the use of 3 ml/ml hygromycin (Hygromycin B,
Sigma, Saint-Quentin Fallavier, France).
2.3. Selection of induced cells
Stably transfected cells were induced with 0.6 mg/ml doxy-
cyline for 24 h (Doxycycline, AP-HP, France) and labeled with
anti-low affinity NGFR (LNGFR) coupled with magnetic
micro beads for separation by the ‘‘MACSelectÔ LNGFR
System’’ procedure (Miltenyi Biotech, Paris, France). The in-
duced expression of LNGFR was assessed by flow cytometry.
CD19 quiescent B lymphocytes were separated from PBMCs
using the same method with anti-CD19.
2.4. Flow cytometry
Induced cells were washed in phosphate buffered saline
(PBS; BioMerieux, Marcy l’Etoile, France), incubated with
10 ml PE-labeled NGFR antibody (BD Pharmingen, Morangis,
France) and counted using a Coulter EPICS XL (Beckman
Coulter, Villepinte, France).
To measure the expression of TNFR1 and TNFR2, cells
were washed in PBS, incubated with 5 ml FITC-labeled
TNFR1 (FAB225F) and TNFR2 (FAB226F) antibodies
(R&D Systems Inc., Lille, France) and counted as described.
To measure the apoptosis rate, cells were washed in PBS,
resuspended in Annexin V binding buffer, incubated with
5 ml FITC-labeled Annexin V antibody (BD Pharmingen)
and 5 ml propidium iodide and counted as described.
338 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345
8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 3/9
2.5. Western blotting
Viable cells were counted by the use of the trypan-blue
method. Total protein extracts were obtained as follows:
5 million cells were resuspended in lysis buffer containing
0.01% bromophenol blue (Sigma), 50 mM TriseHCl, pH 6.8, 2%
sodium dodecyl sulfate (Sigma), 2% glycerol (Sigma) and 2%2-mercaptoethanol (Sigma). The lysates were sonicated, boiled
and stored atÿ80 C. The extracts were separated on 10% poly-
acrylamide denaturing gel and transferred to nitrocellulose
membranes (Hybond-C Extra, Amersham Pharmacia Biotech,
Orsay, France). After transfer, the membranes were stained
with Ponceau-red (Sigma) to check that equal amounts of protein
were present in each lane. Membranes blocked for 2 h with 1%
non-fat milk (Regilait, Saint-Martin Belle Roche, France) in
Tris-buffered saline (TBS; 20 mM NaCl, 500 mM TriseHCl,
pH 8.0; Bio-Rad) were then incubated overnight with the first
antibody in TBS containing 1% non-fat milk. The antibodies
were anti-I-kBa (MAD 10B I-kBa, Dr R. Hay, University of
St Andrews, St Andrews, UK) at 1/50. After three washes inTBS and 1% non-fat milk, the corresponding horseradish
peroxidase-conjugated secondary anti-mouse antibodies (Bio-
Rad) at 1/5000, were added for 1 h. After being washed for 2 h
in TBS1% non-fat milk and0.1% Tween (Sigma), themembranes
underwent chemiluminescence (Lumiglo, Santa Cruz, CA, USA)
and autoradiography (Biomax, Kodak, Rochester, NY, USA).
2.6. Electrophoresis mobility shift assay (EMSA)
Nuclear and cytosolic proteins were extracted as described
previously [22]. Twenty micrograms of nuclear protein extract,
in 5 ml extraction buffer C (20 mM Hepes, 25% glycerol,
0.5 M NaCl, 1.5 mM MgCl2) were mixed with 1 ml of an-
nealed 32P-labeled oligonucleotide containing the kB site
(Genset, Paris, France) [23]. EMSA was performed as previ-
ously described [22]. The DNA-protein complexes were sepa-
rated on 6% non-denaturing polyacrylamide gel in 0.25ÂTriseBorateeEDTA buffer (Bio-Rad) by migration at 250 V.
The gel was dried and exposed to a phosphor-imaging screen
(Packard Instruments, Meriden, CT, USA). The radioactive
signal was visualized by the use of a phosphor system analyzer(Cyclone, Packard Instruments).
0
52
0
1
0,0
128
0100 101 102 103 104
128
0
10,0
20,0
30,0
40,0
50,0
60,0
medium PRI BL2 BL2B95.8
A
B
T N F α s
e c r e t i o n p g / m l
r e l a t i v e c e l l s n
u m b e r
log of fluorescence intensity
for TNFR1
100 101 102 103 104
log of fluorescence intensity
for TNFR2
BL2
BL2
B95.8PRI
BL2
BL2
B95.8PRI
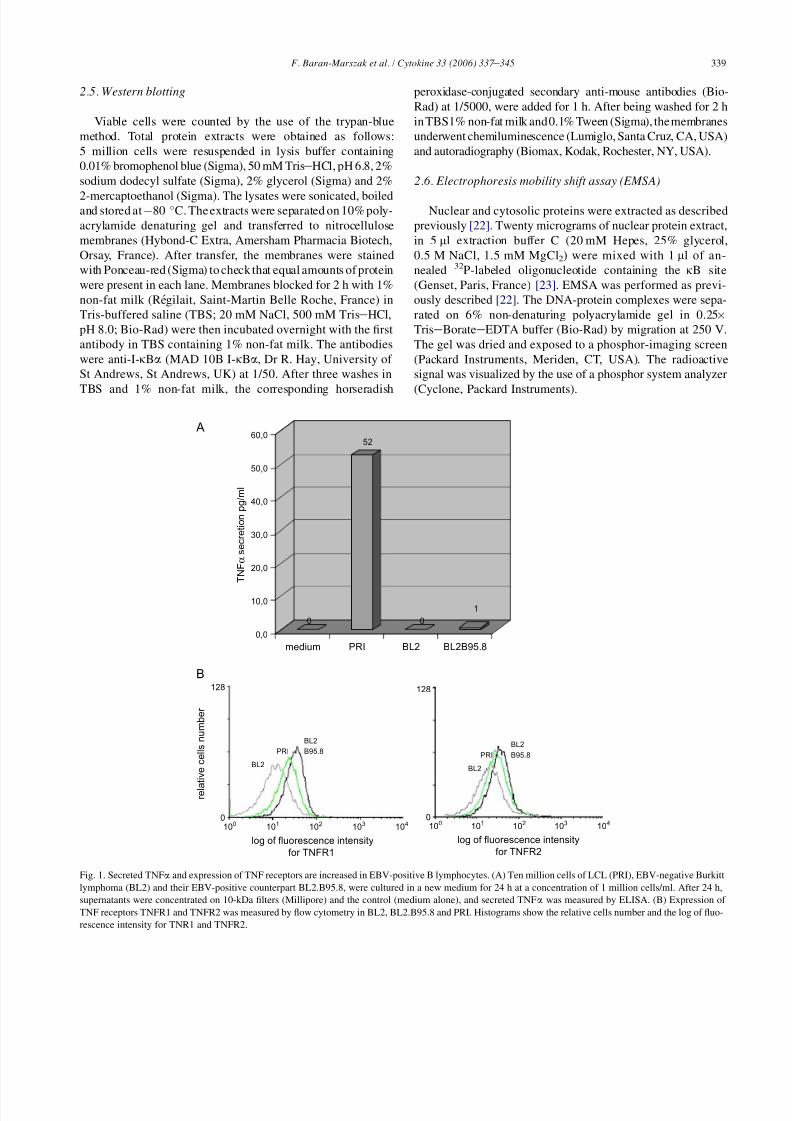
Fig. 1. Secreted TNFa and expression of TNF receptors are increased in EBV-positive B lymphocytes. (A) Ten million cells of LCL (PRI), EBV-negative Burkitt
lymphoma (BL2) and their EBV-positive counterpart BL2.B95.8, were cultured in a new medium for 24 h at a concentration of 1 million cells/ml. After 24 h,
supernatants were concentrated on 10-kDa filters (Millipore) and the control (medium alone), and secreted TNFa was measured by ELISA. (B) Expression of
TNF receptors TNFR1 and TNFR2 was measured by flow cytometry in BL2, BL2.B95.8 and PRI. Histograms show the relative cells number and the log of fluo-
rescence intensity for TNR1 and TNFR2.
339 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345

8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 4/9
2.7. Quantification of cytokines in cell culture
supernatants by ELISA
Ten million cells were washed and grown in 15 ml com-
plete medium culture for 24 h. Supernatants were concentrated
to 500 ml with a centrifugal filter (cut-off:10 kDa; Amicon Ul-
tra-15 Centrifugal Filter Devices, Millipore, Bedford, MA,USA). Secreted TNFa was measured by ELISA (Quantikine
ELISA kit, R&D Systems). The optical density was deter-
mined by the use of a spectrophotometer (Multiskan EX,
Thermo, Cergy-Pontoise, France) set to 450 nm and was ana-
lyzed with use of the Ascent Software (Thermo). Duplicate
concentrations were averaged.
2.8. Quantitative RT e PCR
Total RNA was extracted from sorted NGFR-positive and
-negative cells by the use of the Qiagen kit following the rec-
ommendations of the manufacturer. We defined as referenceRNA a pool of RNAs extracted from different tonsils, lymph
nodes and spleens with benign reactive follicular hyperplasia.
RNA levels for the TNFa gene were quantified in parallel in
the different RNA extracts and in the RNA pool on an ABI
PRISM 7000 automat by the use of the TaqManR ‘‘Assay on
demandÔ’’ gene expression reference system (Applied Bio-
system, website: http//www.appliedbiosystem.com) (product
reference: Hs00174128-m1). The Abl1 gene was used as a ref-
erence gene for the control of amplification (product refer-
ence: Hs00245443-m1). Reverse transcription of 2 mg of
total RNA involved the use of the Archive kit RT from Ap-
plied Biosystems in a final volume of 50 ml. From this, a vol-
ume of 1.25 ml of cDNA was used for gene amplification. All
these steps were performed following the recommendations of
the manufacturer. The relative expression levels of the genes
were calculated as previously reported [24], with Abl1
mRNA expression used for normalization.
3. Results
3.1. TNFa, TNFR1 (p55) and TNFR2 (p75) are
differentially expressed in EBV-positive B-cell lines
The expressions of TNFa and its receptors were measured
in the LCL PRI and BL cell lines infected (BL2.B95.8) or not
(BL2) with EBV. The secretion of TNFa was measured by
ELISA in cell culture supernatants, and the expression of
TNFR1 and TNFR2 was determined by flow cytometry.TNFa secretion was elevated in PRI LCL cells (52 pg/ml)
(Fig. 1A). In contrast with BL2 cells, which did not release
any detectable amount of TNFa, BL2 B95.8 cells secreted
1 pg/ml of TNFa after 24 h of culture in a new medium. Al-
though low, this amount of TNFa was sufficient to induce
IkBa degradation in BL2 cells (see Fig. 5A, lane 3). The ex-
pression of the TNF receptors TNFR1 and TNFR2 was high
in all EBV-positive cells (Fig. 1B), but in our experiments it
was higher in BL2B95.8 than in PRI cells. PRI cells that se-
creted the highest levels of TNFa were subsequently used to
A
I-kBα
Ponceau red
15
7.8
0,0
5,0
10,0
15,0
B
I-kBm- +
- +
I-kBm - + - +
NGFR
FSC
0
0,5
1
1,5
2
2,5
C
I-kBm
2.3
1.3
T N F α
r e
l a t i v
e
e x p r e
s s i
o n
l e v
e l
T N F α s
e c r e t i o n p g / m l
Fig. 2. Specific inhibition of NF-kB reduces TNFa secretion. (A) LCL PRI cells were stably transfected with a tetracycline-inducible vector construct with the
cDNAs coding for I-kBa mutated on serine 32 and 36 (I-kBm) and NGFR as a marker of induction. Expression of NGFR revealed by flow cytometry and
I-kBa by Western blotting was induced after 0.6 mg/ml doxycycline treatment for 24 h. (B) After 24 h of induction, NGFR-positive cells were magnetically selected.
NGFR-positive cells and NGFR-negative cells were cultured for 24 h, supernatants were then concentrated, and secreted TNF a was measured by ELISA. (C)
mRNA expression levels of IkBm selected positive and negative cells were analyzed by quantitative RTePCR. The relative expression level of TNFa mRNA
was calculated with Abl1 mRNA expression used for normalization.
340 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345
8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 5/9
analyze the regulation of the expression of the TNFa gene and
the effects of TNFa and infliximab on apoptosis.
3.2. The expression of TNFa is under the control of
NF-k B in LCLs
To specifically inhibit the transcriptional factor NF-kB, themutated form of I-kBa (mutated on serines 32 and 36) was
over-expressed in PRI cells. Cells were stably transfected
with the NGFR/mutated I-kBa-inducible vector. Inducibility
by doxycycline was verified by flow cytometry and Western
blotting (Fig. 2A). The number of NGFR-positive cells varied
from 30% to 60% after 24 h of induction by doxycycline; cells
were therefore selected in order to analyze homogeneous
populations of NGFR-positive cells. Secretion of TNFa was
measured by ELISA in the supernatants of the selected NGFR-
positive and NGFR-negative cells. The secretion of TNFa was
reduced from 15 to 7.8 pg/ml in NGFR-positive selected cells
(Fig. 2B) and the relative TNFa mRNA expression level was
twofold lower after inhibition of NF-kB (Fig. 2C).
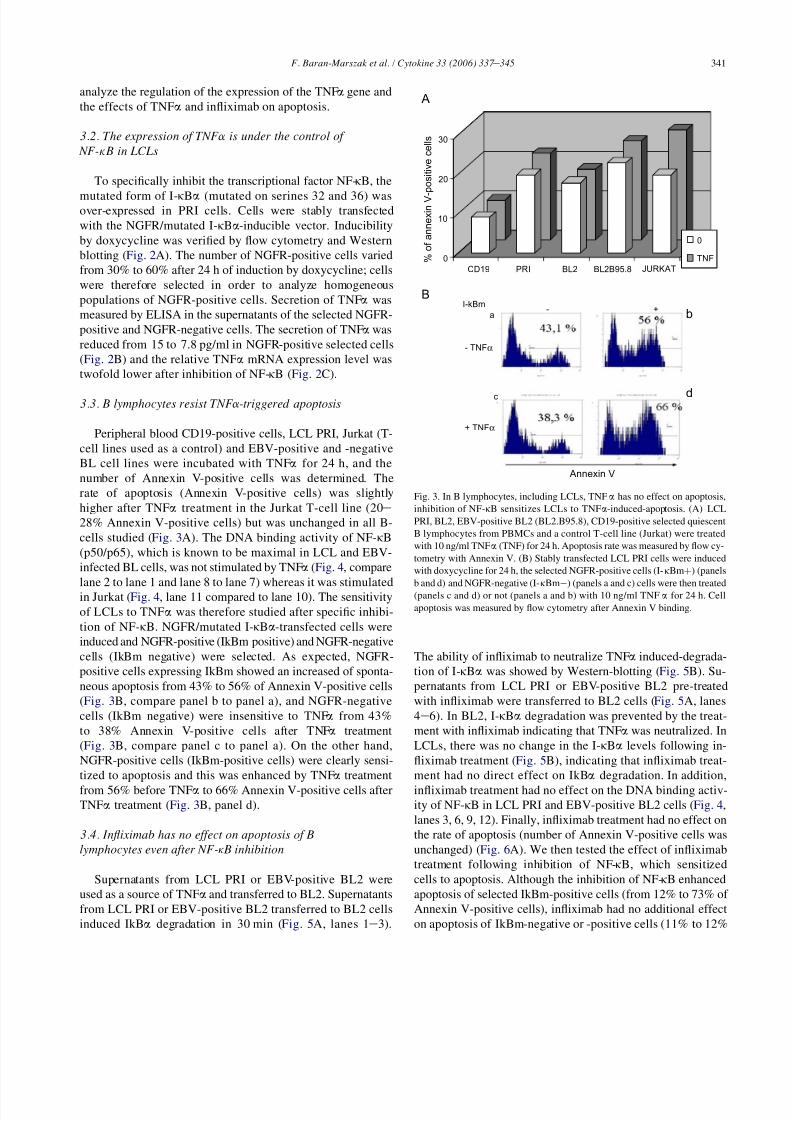
3.3. B lymphocytes resist TNFa-triggered apoptosis
Peripheral blood CD19-positive cells, LCL PRI, Jurkat (T-
cell lines used as a control) and EBV-positive and -negative
BL cell lines were incubated with TNFa for 24 h, and the
number of Annexin V-positive cells was determined. The
rate of apoptosis (Annexin V-positive cells) was slightly
higher after TNFa treatment in the Jurkat T-cell line (20e
28% Annexin V-positive cells) but was unchanged in all B-
cells studied (Fig. 3A). The DNA binding activity of NF-kB
(p50/p65), which is known to be maximal in LCL and EBV-infected BL cells, was not stimulated by TNFa (Fig. 4, compare
lane 2 to lane 1 and lane 8 to lane 7) whereas it was stimulated
in Jurkat (Fig. 4, lane 11 compared to lane 10). The sensitivity
of LCLs to TNFa was therefore studied after specific inhibi-
tion of NF-kB. NGFR/mutated I-kBa-transfected cells were
induced and NGFR-positive (IkBm positive) and NGFR-negative
cells (IkBm negative) were selected. As expected, NGFR-
positive cells expressing IkBm showed an increased of sponta-
neous apoptosis from 43% to 56% of Annexin V-positive cells
(Fig. 3B, compare panel b to panel a), and NGFR-negative
cells (IkBm negative) were insensitive to TNFa from 43%
to 38% Annexin V-positive cells after TNFa treatment
(Fig. 3B, compare panel c to panel a). On the other hand,
NGFR-positive cells (IkBm-positive cells) were clearly sensi-
tized to apoptosis and this was enhanced by TNFa treatment
from 56% before TNFa to 66% Annexin V-positive cells after
TNFa treatment (Fig. 3B, panel d).
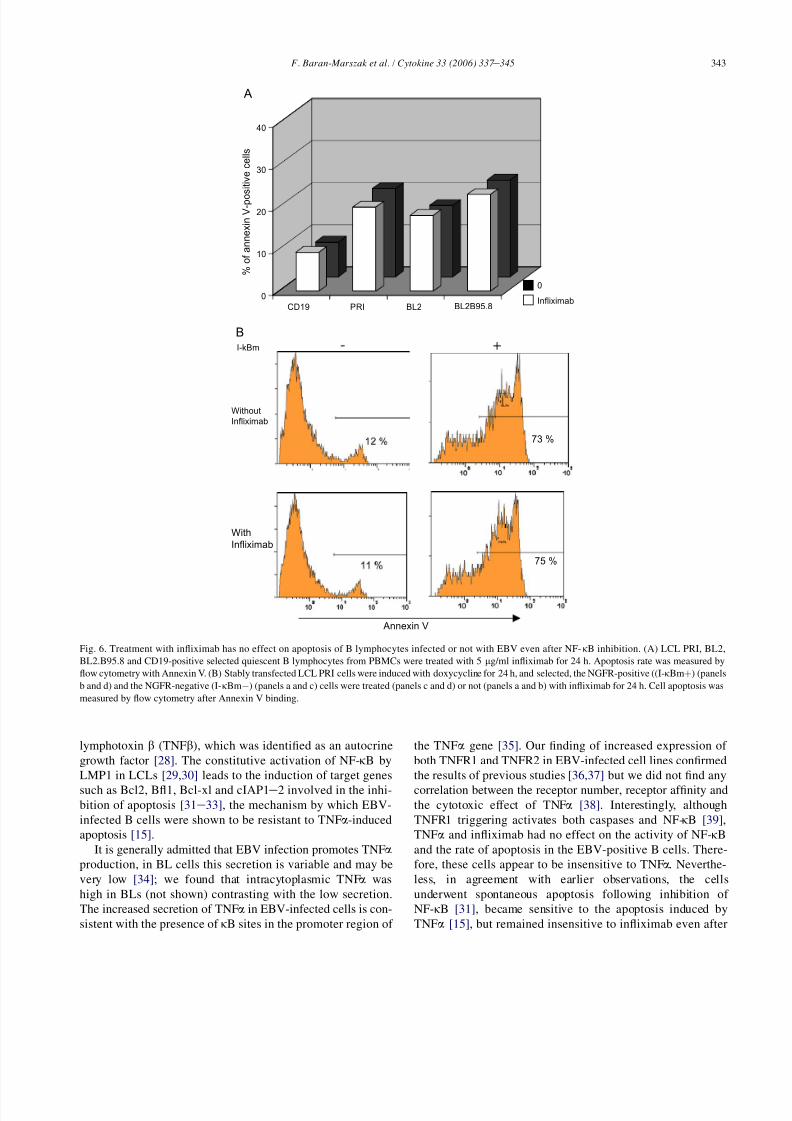
3.4. Infliximab has no effect on apoptosis of B
lymphocytes even after NF-k B inhibition
Supernatants from LCL PRI or EBV-positive BL2 were
used as a source of TNFa and transferred to BL2. Supernatants
from LCL PRI or EBV-positive BL2 transferred to BL2 cells
induced IkBa degradation in 30 min (Fig. 5A, lanes 1e
3).
The ability of infliximab to neutralize TNFa induced-degrada-
tion of I-kBa was showed by Western-blotting (Fig. 5B). Su-
pernatants from LCL PRI or EBV-positive BL2 pre-treated
with infliximab were transferred to BL2 cells (Fig. 5A, lanes
4e6). In BL2, I-kBa degradation was prevented by the treat-
ment with infliximab indicating that TNFa was neutralized. In
LCLs, there was no change in the I-kBa levels following in-
fliximab treatment (Fig. 5B), indicating that infliximab treat-
ment had no direct effect on IkBa degradation. In addition,
infliximab treatment had no effect on the DNA binding activ-
ity of NF-kB in LCL PRI and EBV-positive BL2 cells (Fig. 4,
lanes 3, 6, 9, 12). Finally, infliximab treatment had no effect on
the rate of apoptosis (number of Annexin V-positive cells was
unchanged) (Fig. 6A). We then tested the effect of infliximab
treatment following inhibition of NF-kB, which sensitized
cells to apoptosis. Although the inhibition of NF-kB enhanced
apoptosis of selected IkBm-positive cells (from 12% to 73% of
Annexin V-positive cells), infliximab had no additional effect
on apoptosis of IkBm-negative or -positive cells (11% to 12%
B
0
10
20
30
CD19 PRI BL2 BL2B95.8 JURKAT
0
TNF
A
% o
f a n n e x i n V - p o
s i t i v e c e l l s
I-kBm
- TNFα
+ TNFα
b
d
- +a
c
Annexin V
Fig. 3. In B lymphocytes, including LCLs, TNFa has no effect on apoptosis,
inhibition of NF-kB sensitizes LCLs to TNFa-induced-apoptosis. (A) LCL
PRI, BL2, EBV-positive BL2 (BL2.B95.8), CD19-positive selected quiescent
B lymphocytes from PBMCs and a control T-cell line (Jurkat) were treated
with 10 ng/ml TNFa (TNF) for 24 h. Apoptosis rate was measured by flow cy-
tometry with Annexin V. (B) Stably transfected LCL PRI cells were induced
with doxycycline for 24 h, the selected NGFR-positive cells (I-kBmþ) (panels
b and d) and NGFR-negative (I-kBmÿ) (panels a and c) cells were then treated
(panels c and d) or not (panels a and b) with 10 ng/ml TNFa for 24 h. Cell
apoptosis was measured by flow cytometry after Annexin V binding.
341 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345

8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 6/9
for IkBm-negative cells treated with infliximab, and 75%
to 73% for IkBm-positive cells treated with infliximab)
(Fig. 6B).
4. Discussion
In this study, we demonstrated that neither TNFa nor inflix-
imab had an effect on apoptosis in B cells infected or not with
EBV. The increased production of TNFa by EBV-positive
cells, particularly LCLs, suggested a possible effect of inflix-
imab on these cells. However, our observation that infliximab
had no effect on apoptosis of LCLs, even when cells were
sensitized to apoptosis by inhibition of NF-kB suggests that
TNFa had no autocrine action in these cells and that inflixi-
mab had no TNFa-independent effect.Previous studies have demonstrated that exogenous TNFa
increases DNA synthesis and immunoglobulin production by
mitogen-activated human B cells [25]. In addition, TNFa is
an autocrine growth factor in B-cell chronic lymphocytic leu-
kemia [25,26]. Finally, TNFa is one of the earliest genes tran-
scribed after antigen stimulation of B cells and is not sufficient
for but augments anti-Ig or anti-CD40þ interleukin-4-induced
B-cell proliferation [27]. In EBV-infected B cells, TNFa had
no effect on apoptosis, although it could compete with
p50/p65
p50/p50
000 Inf
6 7 8 9 10 11 12
tnf 0Inf Inf Inf tnf tnf tnf
PRI BL2 BL2B95.8 Jurkat
21 3 4 5
Fig. 4. TNFa or infliximab have no effect on the DNA binding activity of NF-kB in B lymphocytes. LCL PRI (lanes 1, 2 and 3), BL2 (lanes 4, 5 and 6), BL2.B95.8
(lanes 7, 8 and 9) and Jurkat cells (lanes 10, 11 and 12) were treated with TNFa (tnf) (lanes 2, 5, 8, 11) or infliximab (inf) (lanes 3, 6, 9, 12) for 24 h, and NF-kB
activity (p50/p65) was analyzed by EMSA.
A
B
1-kBα
Ponceau red
I-kBα
Ponceau red
Infliximab
1 2 3 4 5 6
BL2 BL2
0 P R I s u
p
B L 2 B 9 5
. 8 s u p
0 P R I i n f l i x
i m a b
p r e t r e
a t e
d s
u p
B L 2 B 9 5
. 8 i n
f l i x i m
a b
p r e t r e
a t e
d s
u p
0
Fig. 5. Infliximab neutralizes the secreted TNFa but has no effect on the TNF-induced I-kBa degradation in EBV-infected B cells. (A) Supernatants from EBV-
positive cells (PRI sup or BL2.B95.8 sup) were transferred to EBV-negative BL cells (BL2) for 30 min. The total protein extracts were separated on gel and I-kBa
degradation was analyzed by Western blotting. BL2 incubated with: LCL PRI supernatant (lane 2), BL2.B95.8 supernatant (lane 3), infliximab-pretreated LCL PRI
supernatant (lane 5), infliximab-pretreated BL2.B95.8 supernatant (lane 6). (B) Absence of a direct effect of infliximab. LCL PRI cells were treated with infliximab
for 24 h and expression of I-kBa was analyzed by Western blotting on total protein extracts.
342 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345
8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 7/9
lymphotoxin b (TNFb), which was identified as an autocrine
growth factor [28]. The constitutive activation of NF-kB by
LMP1 in LCLs [29,30] leads to the induction of target genes
such as Bcl2, Bfl1, Bcl-xl and cIAP1e2 involved in the inhi-
bition of apoptosis [31e33], the mechanism by which EBV-
infected B cells were shown to be resistant to TNFa-induced
apoptosis [15].
It is generally admitted that EBV infection promotes TNFa
production, in BL cells this secretion is variable and may be
very low [34]; we found that intracytoplasmic TNFa was
high in BLs (not shown) contrasting with the low secretion.
The increased secretion of TNFa in EBV-infected cells is con-
sistent with the presence of kB sites in the promoter region of
the TNFa gene [35]. Our finding of increased expression of
both TNFR1 and TNFR2 in EBV-infected cell lines confirmed
the results of previous studies [36,37] but we did not find any
correlation between the receptor number, receptor affinity and
the cytotoxic effect of TNFa [38]. Interestingly, although
TNFR1 triggering activates both caspases and NF-kB [39],
TNFa and infliximab had no effect on the activity of NF-kB
and the rate of apoptosis in the EBV-positive B cells. There-
fore, these cells appear to be insensitive to TNFa. Neverthe-
less, in agreement with earlier observations, the cells
underwent spontaneous apoptosis following inhibition of
NF-kB [31], became sensitive to the apoptosis induced by
TNFa [15], but remained insensitive to infliximab even after
0
10
20
30
40
CD19 PRI BL2 BL2B95.8
% o
f a n n e x i n V - p o s i t i v e c e l l s
Annexin V
I-kBm
Without
Infliximab
A
B
With
Infliximab
0
Infliximab
11 %
12 %
75 %
73 %
Fig. 6. Treatment with infliximab has no effect on apoptosis of B lymphocytes infected or not with EBV even after NF-kB inhibition. (A) LCL PRI, BL2,
BL2.B95.8 and CD19-positive selected quiescent B lymphocytes from PBMCs were treated with 5 mg/ml infliximab for 24 h. Apoptosis rate was measured by
flow cytometry with Annexin V. (B) Stably transfected LCL PRI cells were induced with doxycycline for 24 h, and selected, the NGFR-positive ((I-kBmþ) (panels
b and d) and the NGFR-negative (I-kBmÿ) (panels a and c) cells were treated (panels c and d) or not (panels a and b) with infliximab for 24 h. Cell apoptosis was
measured by flow cytometry after Annexin V binding.
343 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345

8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 8/9
the inhibition of NF-kB. Thus confirming the absence of a di-
rect effect of infliximab on LCLs. In addition, we ruled out the
possibility of increased apoptosis following a direct interaction
of infliximab with transmembrane TNFa as suggested in T
lymphocytes [16]. After neutralization of TNFa with inflixi-
mab no effect on apoptosis of LCLs was observed confirming
that TNFa had no autocrine role in EBV-infected cells. Thus,in comparison with its ability to induce apoptosis in T cells of
the lamina propria in vivo and in vitro in CD, of blood mono-
cytes in CD and RA and of synovial monocytes/macrophages
in RA, infliximab is unable to increase apoptosis in B cells,
which demonstrates that the action of TNFa blockers may
vary in different cellular types.
The possible increased risk of lymphoma with TNF antag-
onists is one of the major issues concerning the long-term
safety of TNFa blockers. Even if a few studies suggest that
the incidence of lymphoma is increased in RA patients treated
with TNFa blockers [20,21], it is impossible to determine if
this incidence is linked with anti-TNFa-treated RA being
more active or corresponds to excessive risk induced by theTNFa blocker itself. If an increased incidence of lymphoma
was induced by TNF blockers, it would not involve, on the ba-
sis of our in vitro data, a direct effect of the treatment of B
cells infected or not with EBV but rather an impaired immune
surveillance by T cells.
Acknowledgments
This work was supported in part by a grant from Schering-
Plough. C.L. was supported by the Fondation pour la re-
cherche medicale (FRM).
References
[1] Bernstein CN, Blanchard JF, Kliewer E, Wajda A. Cancer risk in patients
with inflammatory bowel disease: a population-based study. Cancer
2001;91:854e62.
[2] Georgescu L, Quinn GC, Schwartzman S, Paget SA. Lymphoma in pa-
tients with rheumatoid arthritis: association with the disease state or
methotrexate treatment. Semin Arthritis Rheum 1997;26:794e804.
[3] Kamel OW, Holly EA, van de Rijn M, Lele C, Sah A. A population
based, case control study of non-Hodgkin’s lymphoma in patients with
rheumatoid arthritis. J Rheumatol 1999;26:1676e80.
[4] Kamel OW, van de Rijn M, Hanasono MM, Warnke RA. Immunosup-
pression-associated lymphoproliferative disorders in rheumatic patients.Leuk Lymphoma 1995;16:363e8.
[5] Kumar S, Fend F, Quintanilla-Martinez L, Kingma DW, Sorbara L,
Raffeld M, et al. EpsteineBarr virus-positive primary gastrointestinal
Hodgkin’s disease: association with inflammatory bowel disease and
immunosuppression. Am J Surg Pathol 2000;24:66e73.
[6] Mariette X, Cazals-Hatem D, Warszawki J, Liote F, Balandraud N,
Sibilia J. Lymphomas in rheumatoid arthritis patients treated with meth-
otrexate: a 3-year prospective study in France. Blood 2002;99:3909e15.
[7] Martin T, Weber JC, Levallois H, Labouret N, Soley A, Koenig S, et al.
Salivary gland lymphomas in patients with Sjogren’s syndrome may fre-
quently develop from rheumatoid factor B cells. Arthritis Rheum
2000;43:908e16.
[8] Baecklund E, Ekbom A, Sparen P, Feltelius N, Klareskog L. Disease ac-
tivity and risk of lymphoma in patients with rheumatoid arthritis: nested
case-control study. BMJ 1998;317:180e
1.
[9] Nakatsuka S, Yao M, Hoshida Y, Yamamoto S, Iuchi K, Aozasa K. Pyo-
thorax-associated lymphoma: a review of 106 cases. J Clin Oncol
2002;20:4255e60.
[10] Kamel OW, van de Rijn M, Weiss LM, Del Zoppo GJ, Hench PK,
Robbins BA, et al. Brief report: reversible lymphomas associated with
EpsteineBarr virus occurring during methotrexate therapy for
rheumatoid arthritis and dermatomyositis. N Engl J Med 1993;328:
1317e21.
[11] Kamel OW, Weiss LM, van de Rijn M, Colby TV, Kingma DW, Jaffe ES.
Hodgkin’s disease and lymphoproliferations resembling Hodgkin’s dis-
ease in patients receiving long-term low-dose methotrexate therapy.
Am J Surg Pathol 1996;20:1279e87.
[12] Starzl TE, Nalesnik MA, Porter KA, Ho M, Iwatsuki S, Griffith BP, et al.
Reversibility of lymphomas and lymphoproliferative lesions developing
under cyclosporin-steroid therapy. Lancet 1984;1:583e7.
[13] Wolfe F, Michaud K. Lymphoma in rheumatoid arthritis: the effect of
methotrexate and anti-tumor necrosis factor therapy in 18,572 patients.
Arthritis Rheum 2004;50:1740e51.
[14] Natoli G, Costanzo A, Guido F, Moretti F, Levrero M. Apoptotic, non-
apoptotic, and anti-apoptotic pathways of tumor necrosis factor signalling.
Biochem Pharmacol 1998;56:915e20.
[15] Asso-Bonnet M, Feuillard J, Ferreira V, Bissieres P, Tarantino N,
Korner M, et al. Relationship between IkappaBalpha constitutive expres-
sion, TNFalpha synthesis, and apoptosis in EBV-infected lymphoblastoid
cells. Oncogene 1998;17:1607e15.
[16] ten Hove T, van Montfrans C, Peppelenbosch MP, van Deventer SJ. In-
fliximab treatment induces apoptosis of lamina propria T lymphocytes
in Crohn’s disease. Gut 2002;50:206e11.
[17] Catrina AI, Trollmo C, af Klint E, Engstrom M, Lampa J, Hermansson Y,
et al. Evidence that anti-tumor necrosis factor therapy with both etaner-
cept and infliximab induces apoptosis in macrophages, but not lympho-
cytes, in rheumatoid arthritis joints: extended report. Arthritis Rheum
2005;52:61e72.
[18] Lugering A, Schmidt M, Lugering N, Pauels HG, Domschke W,
Kucharzik T. Infliximab induces apoptosis in monocytes from patients
with chronic active Crohn’s disease by using a caspase-dependent path-
way. Gastroenterology 2001;121:1145e57.
[19] Feltelius N, Fored CM, Blomqvist P, Bertilsson L, Geborek P,Jacobsson LT, et al. Results from a nationwide postmarketing cohort
study of patients in Sweden treated with etanercept. Ann Rheum Dis
2005;64:246e52.
[20] Fleischmann R, Yocum D. Does safety make a difference in selecting the
right TNF antagonist? Arthritis Res Ther 2004;6:S12e8.
[21] Baran-Marszak F, Feuillard J, Najjar I, Le Clorennec C, Bechet JM,
Dusanter-Fourt I, et al. Differential roles of STAT1{alpha} and STAT1
{beta} in fludarabine-induced cell cycle arrest and apoptosis in human
B cells. Blood 2004;104:2475e83.
[22] Feuillard J, Gouy H, Bismuth G, Lee LM, Debre P, Korner M. NF-kappa
B activation by tumor necrosis factor alpha in the Jurkat T cell line is in-
dependent of protein kinase A, protein kinase C, and Ca(2þ)-regulated
kinases. Cytokine 1991;3:257e65.
[23] Wu F, Garcia J, Mitsuyasu R, Gaynor R. Alterations in binding charac-
teristics of the human immunodeficiency virus enhancer factor. J Virol1988;62:218e25.
[24] Gabert J, Beillard E, van der Velden VH, Bi W, Grimwade D,
Pallisgaard N, et al. Standardization and quality control studies of
‘real-time’ quantitative reverse transcriptase polymerase chain reaction
of fusion gene transcripts for residual disease detection in leukemia -
a Europe Against Cancer program. Leukemia 2003;17:2318e57.
[25] Jelinek DF, Lipsky PE. Regulation of human B lymphocyte activation,
proliferation, and differentiation. Adv Immunol 1987;40:1e59.
[26] Heslop HE, Bianchi AC, Cordingley FT, Turner M, Chandima W, De
Mel CP, et al. Effects of interferon alpha on autocrine growth factor
loops in B lymphoproliferative disorders. J Exp Med 1990;172:
1729e34.
[27] Boussiotis VA, Nadler LM, Strominger JL, Goldfeld AE. Tumor necrosis
factor alpha is an autocrine growth factor for normal human B cells. Proc
Natl Acad Sci USA 1994;91:7007e
11.
344 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345

8/8/2019 Barab-Marszak, 2006
http://slidepdf.com/reader/full/barab-marszak-2006 9/9
[28] Estrov Z, Kurzrock R, Pocsik E, Pathak S, Kantarjian HM, Zipf TF, et al.
Lymphotoxin is an autocrine growth factor for EpsteineBarr virus-
infected B cell lines. J Exp Med 1993;177:763e74.
[29] Laherty CD, Hu HM, Opipari AW, Wang F, Dixit VM. The Epsteine
Barr virus LMP1 gene product induces A20 zinc finger protein expres-
sion by activating nuclear factor kappa B. J Biol Chem 1992;267:
24157e60.
[30] Lam N, Sugden B. CD40 and its viral mimic, LMP1: similar means to
different ends. Cell Signal 2003;15:9e16.
[31] Cahir-McFarland ED, Davidson DM, Schauer SL, Duong J, Kieff E. NF-
kappa B inhibition causes spontaneous apoptosis in EpsteineBarr virus-
transformed lymphoblastoid cells. Proc Natl Acad Sci USA
2000;97:6055e60.
[32] Chen C, Edelstein LC, Gelinas C. The Rel/NF-kappaB family directly
activates expression of the apoptosis inhibitor Bcl-x(L). Mol Cell Biol
2000;20:2687e95.
[33] Chen F, Castranova V, Shi X. New insights into the role of nuclear factor-
kappaB in cell growth regulation. Am J Pathol 2001;159:387e97.
[34] Klein SC, Kube D, Abts H, Diehl V, Tesch H. Promotion of IL8, IL10,
TNF alpha and TNF beta production by EBV infection. Leukemia Res
1996;20:633e6.
[35] Goldfeld AE, Strominger JL, Doyle C. Human tumor necrosis factor al-
pha gene regulation in phorbol ester stimulated T and B cell lines. J Exp
Med 1991;174:73e81.
[36] Baran-Marszak F, Fagard R, Girard B, Camilleri-Broet S, Zeng F,
Lenoir GM, et al. Gene array identification of Epstein Barr virus-regu-
lated cellular genes in EBV-converted Burkitt lymphoma cell lines.
Lab Invest 2002;82:1463e79.
[37] Gibbons DL, Rowe M, Cope AP, Feldmann M, Brennan FM. Lympho-
toxin acts as an autocrine growth factor for EpsteineBarr virus-trans-
formed B cells and differentiated Burkitt lymphoma cell lines. Eur J
Immunol 1994;24:1879e85.
[38] Munker R, DiPersio J, Koeffler HP. Tumor necrosis factor: receptors on
hematopoietic cells. Blood 1987;70:1730e4.
[39] Wajant H, Pfizenmaier K, Scheurich P. Tumor necrosis factor signaling.
Cell Death Differ 2003;10:45e65.
345 F. Baran-Marszak et al. / Cytokine 33 (2006) 337 e 345