australian society for reproductive biology … · programme guide hour 0830 monday, august 26...
TRANSCRIPT


announcement . • •
I
I AUSTRALIAN SOCIETY FOR REPRODUCTIVE BIOLOGY
SEVENTEENTH ANNUAL CONFERENCE
Adelaide, Australia
AUGUST 25-28, 1985
ANNUAL CONFERENCE 1986
1st-3rd September, 1986
at University of Queensland
Abstracts should be prepared by May 1986
Local Organiser: Jim Cummins (07) 377 3733
PROGRAMME
AND
A B S T R ACT S 0 F PAP E R S
Copyright Australian Society for Reproductive Biology, 1985
CONTENTS
Personnel I IAcknowledgements IIIProgramme Guide IVProgramme V-XVIIAuthors XVIII-XVIVAbstracts 1-116Minutes of 16th AGM 1984

AUSTRALIAN SOCIETY FOR REPRODUCTIVE BIOLOGY THE AUSTRALIAN SOCIETY FOR REPRODUCTIVE BIOLOGY
August, 1985wishes to thank the following fortheir support of the 1985 meeting
OFFICE
Chairman
Secretary
BEARERS
DR J .K. FINDLAY
DR J. CUMMINS
ANSETT·AIRLINES
ORGANON AUSTRALIA
S.A. DIAGNOSTIC SUPPLIES
IMMUNONUCLEAR AUSTRALIA
BIOMEDIX AUSTRALIA
Treasurer DR C. NANCARROW
Committee Members DR B.G. MILLER
DR P. TEMPLE-SMITH
The contents of these proceedings have not been edited by theSociety or the Editor and are reproduced as submitted.Responsibility for accuracy of the communications rests withthe authors.
DR G. EVANS
DR J. RODGERMaterial in these proceedings may not be reproduced withoutpermission of the society.
DR J. FALCONER Price of Proceedings to non-members. $A10.00 plus postage.
PROF. D.R. LINDSAY
PROGRAMME COM MIT TEE
ill ,
Ii!JaI
p
LEGEND
N
1'RAA~ I;
/.i'..
B c.: 0
4 10 .
tt7.
, ~:
J. OLIVER
C. ROEGER
P. QUINN
COM MIT TEE
DR N.A. ADAMS
DR P. WILLIAMSON
DR N. BRUCE
DR 1. PURVIS
DR J. YOVICH
o R G A N I SIN G
S. JUDD (CONVENOR)
C. WEBSTER (SAPMEA)
R. BURNET
L 0 CAL
Committee Members
Chairman
II III

PROGRAMME GUIDE
Hour
0830
Monday, August 26
Session 1:Chairman, Dr C.D.MathewsFERTILITY & INFERTILITY
Abst. 1 - 6
Oral presentations
Tuesday, August 27
Session 5:Chairman, Dr H.W. BakerTHE MALE
Abst. 46 - 51
Oral presentations
Wednesday, August 28
Robinson Symposium:Chairma~ Dr J. Findlay
0830 - 0915Dr R. Ortavant
0915 - 1000Prof. N°. W. Moore
AUSTRALIAN SOCIETY FOR REPRODUCTIVE BIOLOGY
PROGRAMME
Sunday, August, 25th
1000 TEA 1800 Welcoming Reception at ConstitutionalMuseum, North Terrace. (Site No.5).
1030 Session 2:Chairman, Prof. R.G.WalesIN VITRO STUDIES &PITUITARY
Abst. 7 - 12
Oral presentations
Session 6:Chairman, Dr. B. BindonTHE OVARY
Abst. 52 - 57
Oral presentations
1030 - 1100Prof. D.R. Lindsay
1100 - 1145Dr R.J.Scaramuzziand Dr J.F. Smith
1145 - 1215Prof.T.J.Robinson
1215 - 1300DISCUSSION
Registration from 1800.
Monday, August, 26th
Session 1: FERTILITY & INFERTILITY. Oral presentations
S. JunkP. MatsonF. O'HalloranJ.L. Yovich
C.D. NancarrowJ.D. MurrayR.J. ScaramuzziJ.T. MarshallI.G. Hazeltona.H. HoskinsonM.P. Boland
L.J. WiltonP.D. Temple-SmithH. W. G. BakerD.M. de Kretser
Horace Lamb Theatre
I.F. DavisD.J. KertonR.A. Parr
v
Fertilization and embryo development in androstenedione-immune sheep.
Chairman, Dr C.D. Mathews
Epididymal necrozoospermia: A newlydefined cause of asthenozoospermiaand its potential treatment.
The use of immunobeads to detecthuman anti-spermatozoal antibodies.
Number of spermatozoa and ovulationrate affect fertility and prolificacy of sheep.
A.W.N. CameronC.M. OldhamI.J. FairnieE.J. KeoghD.R. Lindsay
Reinnervation of the rat vas deferens R.M. DeGarisfollowing vasovasostomy after J.N. Pennefathervasectomy
Progesterone treatments to improvefertility after uterine AI in ewes
Title
6
4
3
5
2
Abst.No.-1-0830
0845
z-1
0900,-0915
10930
~?
0945
Abst. 111 - 116
Oral presentation
Session 9: COMPARATIVEREPRODUCTIVE BIOLOGYProfessor R.V. Short
Junior Scientist AwardPresented
IV
TEA
ASRB AGM
Abst. 58 - 104
Oral presentations
Abst. 105 - 110
LUNCH
Session 7(A) MALEREPRODUCTION: Dr E.J.Keogh&Prof. T. Glover7(B) THE OVARY:Dr L.Cahi1l7(C)*COMPARATIVE REPRODUCTIVE BIOLOGY: Dr M.Renfree
Session 8:Dr C. NancarrowTHE UTERUS & THE EMBRYO
Poster Discussion
HARRISON LECTURE
Dr Michael Berridge
Session 4:Chairman, Prof. D.M. deKretserTHE TESTIS & SPERMATOZOA
Abst. 13 - 39
Session 3(A) FERTILITY &INFERTILITY: Prof. T.J.Robinson,3(B) IN VITRO STUDIES &PITUITARY: Dr P. Quinn3(C) *SEASONALITY: Dr R.Seamark
Abst. 40 - 45
Poster Discussion
Oral presentations
GODING LECTURE
Professor B. Setchell
1400
Posters located throughout Meeting for viewing and discussion in The Crit Library.Oral presentations in the Horace Lamb Theatre.Goding and Harrison Lectures in the Flentje Theatre.
*In Upper Refectory.
1300
1730
1530
1600
1200

Session 2:
1030 7
IN VITRO STUDIES AND PITUITARY
Chairman, Prof. R.G. Wales
Morphology of pre-ovulatory humanoocytes.
Oral presentations
Horace Lamb Theatre
G.M. JonesA. LopataL. ChiappazzoY. du PlessisH. BourneD. Levran
16
17
Failure of testicular vein ligationto increase fertility in men withvaricoceles.
Cyclic changes in the sialic acidcontent of cervical mucus.
H.W.G. BakerH.G. BurgerD.M. de KretserB. HudsonG.C. RennieW.G.E. Straffon
P. LutjenJ. HoyF. Martinez
1200-1300
Effects of enzyme inhibitors on hamster J.M. Cumminsgamete interactions. R. Yanagimachi
1045
II~/
1100I
1115
1130
~
1145
8
9
10
11
12
Male Factor Patients FertilizationRates in IVF
On the molecular basis of humansperm-egg recognition
B-Endorphin in ovine pituitary portalblood during suckling.
Platelet derived growth factors arenecessary for mouse embryo implantation in-vitro
JAMES GODING MEMORIAL LECTURE
Movement of substances and fluids inthe testes.
C.A. YatesA.O. TrounsonD.M. de Kretser
P. LutjenM. de Witt
K. GordonM.B. RenfreeR.V. ShortI.J. Clarke
C. O'Neill
Flentje Theatre
Prof. B. Setchell
18
19
20
21
22
The reproductive performance afterexposure to oestrogenic pastures ofewes immunized against androstenedioneor oestrone.
Gonadotrophin responses to LHRH inKoonoona and Booroola-Koonoona firsteros ram lambs: influence of LHRHdose and age.
The effects of different patternsof insemination, and of numbers ofmotile sperm inseminated, on theefficienty of AID treatment.
Clover infertility in ewes: Is there agenetic component to its development?
The association between occupationalgroup and sperm concentration ininfertile men.
R.I. CoxD.L. LittleS.K. Walker
D.O. KleemanM.J. D'OcchioS.K. WalkerC. PapachristoforouD.H. SmithB.P. Setchell
B. GodfreyC.D. Mathews
K.P. CrokerT.J. JohnsonR.J. Lightfoot
J. HendersonH.W.G. BakerG.C. Rennie
1400-1530
13
14
15
Session 3(A) FERTILITY& INFERTILITY: Poster discussion
Chairman, Prof. T.J. Robinson
Penile erection in the dog
Further observations on F geneprogeny testing using response toPMSG in prepubertal ewe lambs
Oral antifertility activity ofgossypol steroidal analogues andcyproterone acetate in male rats
VI
The erit Library
C.J. CaratiK.E. CreedE.J. Keogh
LoR. PiperB.M. BindonS.K. WalkerJ.R.W. WalkleyD. Phillips
I.G. WhiteR. VishwanathP.D. Brown-WoodmanD. RidleyM.A. Swan
If23
24
Lupin feeding and fertility in theMerino ewe.
Use of the glucose test to improvebreeding flock productivity.
VI I
M.B. NottleP.I. HyndB.P. SetchellR.F. Seamark
R.A. ParrM.A. MilesP.J. Langdon

Session 3(C) SEASONALITY: Poster Discussion
1400-1530
25
26
27
I'}
28
29
30
31
32
33
1400-1530
34
35
Session 3(B) IN VITRO STUDIES [.PITUITARY: Poster Presentation
Chairman, Dr P. Quinn
Effect of growth factors on the rateof mouse embryo development in-vitro.
Long-term oestrogen-progestagentreatment does not increase theinhibitory effects of oestradiol ongonadotrophin secretion in ovariectomised ewes.
A transient elevation of prolactinsecretion following hypothalamopituitary disconnection in the ewe.
Oestrogen and progesterone stop GnRHpulses in ovariectomized anestrousewes.
Effect of addition and removal ofglycerol and method of transfer onviability of sheep embryos.
Effect of type of storage vessel andmethod of glycerol removal on viabilicy of frozen-thawed sheep embryos.
Temperature requirements for sucrosedilution of glycerol from frozenthawed mouse embryos.
Visualization of pronuclei and cultureof one-cell embryos of sheep.
Electron microscopic histochemistry ofthe matrix of the mouse cumulusoocyte complex.
Chairman, Dr R. Seamark
Effect of long daylength onreproduction in Javanese thin-tail ewes.
The effect of melatonin, progesteroneand PMSG on the ovulatory and matingresponse of Romney ewe lambs tosexually active rams.
VIII
The Crit Library
C.M. CaroA. Trounson
Z.C. ZhangP.J. WrightJ.A. BurdenI.J. Clarke
G.B. ThomasJ.T. CumminsLynda Cavanagh1.J. Clarke
I.J. ClarkeF.J. KarschJ.T. Cummins
J.R. HuntonW.M.C. MaxwellJ.P. Ryan
J.P. RyanW.M.C. MaxwellJ.R. Hunton
A. SzellJ.N. Shelton
S.K. WalkerP. QuinnR.J. AshmanD.H. Smith
R. I. WebbJ.M. Cummins
The Cri t Library
I.C. FletcherI. Gede Putu
R.W. MooreP.R. LynchC.M. MillerR.A. S. WelchP. Haszard
((
1600
1615
1630
1645
1700
1715
36
37
38
39
40
42
43
44
45
Effect of melatonin on postpartumanoestrus in beef cows.
Tropical rusa deer - a long daybreeder in Victoria.
Seasonal LH profiles inganglionectomised Merino crossbredewes.
Induction of ovulation in maresduring seasonal anoestrus by infusionof gonadotrophin releasing hormone(GnRH).
Session 4: THE TESTIS &SPERMATOZOA: Oral Presentations
Chairman, Prof. D.M. de Kretser.
Sexual activity and seminal characteristics in rams treated withepidermal growth factor.
Daily sperm production of matureMerino rams immunized early in lifewith an inhibin-enriched fractionfrom bovine follicular fluid.
Motility characteristics of ramspermatozoa extracted withTriton X-IOO.
Is Leydig cell heterogeneity in~ an artifact?
Seminal transferrin, an index ofSertoli cell function: possibleclinical value.
Sertoli cell influence on myoidcell growth and/or proliferation.
IX
P.H. SharpeD.R. GiffordP.F. FlavelM.B. NottleD.T. Armstrong
Simone van MourikT. Stelmasiak
S.A. CutlerG. EvansR.J. Scaramuzzi
J. HylandP.J. Wright1.J. ClarkeR.S. CarsonD.A. LangsfordL.B. Jeffcott
Horace Lamb Theatre
P.E. MattnerB.W. BrownB.A. Panaretto
S.A.R. Al-ObaidiB.M. BindonT. O'SheaM.A. Hillard
R. VishwanathM.A. SwanI.G. White
A.a. LawsN.G.M. WrefordD.M. de Kretser
Liu DeyiE.J. CooperH.W.G. Baker
E.W. ThompsonJ.S. ElkingtonA.W. Blackshaw

Tuesday, August 27
Session 5: THE MALE: Oral presentations 1200-1300
KEITH HARRISON MEMORIAL LECTURE
Cell membrane phospholipid metabolism:mediators of hormone action.
~/( Jl-ft. f.e.el./e-.
Flentje Theatre -
11030 52
?J1045 53
/' 1100 54
1115 55
0900 48(,0915 49
j.)
0930 50
100945 51
tv
Session 7(A) MALE REPRODUCTION: poster Discussion
Chairmen: Dr E.J. Keogh & Prof. T. Glover. The Crit Library
/
D.E. Brooks
C. PholpramoolJ.L. ZuppB.P. Setchell
D.B. GallowayZhang Zhong Cheng
S. SalamonW.M.C. MaxwellG. Evans
O. AyodejiH.W.G. Baker
E. Herrera CastanedaP.D. Temple-smith
D. StevensonG.F. KingA.R. Jones
T.A. ReidA.R. JonesI.G. White
M.K. HayesD.E. Brooks
A.W.N. CameronA.J. TilbrookD.R. LindsayE.J. KeoghI.J. Fairnie
Purification and properties of a22-kDa protein present in testicularand epididymal fluids and. on thesurface of spermatozoa.
Sperm motility and morphology inspermatic granuloma of the epididymalhead (spermiostasis).
The motility of epididymal spermatozoain the bull.
Fertility of ram semen frozen-storedfor 16 years.
Is there a specific abnormality ofsperm morphology in men withvaricoceles?
In vivo and in vitro effects of humanchorionic gonadotropin (hCG).
Inhibition of glyceraldehyde phosphatedehydrogenase in boar spermatozoa by(S)_3_Chlorolactaldehyde.
The effect of (S)-a-Chlorohydrin onthe metabolic activity of ejaculatedram spermatozoa.
Hormones effects on protein synthesisand secretion by Sertoli cells in thedeveloping bovine testis.
The effect of testicular weight andinsemination technique on fertility.
58
,.g 59
60
~r
61
62
63
64
65
66
67
1400-1530
S. MaddocksJ. CormackB.P. Setchell
R.C. Fry1.J. ClarkeL.P. Cahill
B.J. WaddellN.W. BruceA.M. Dharmarajan
W.R. GibsonP.J. Roche
R.J. FaircloughL.D. StaplesJ.D. 0' Shea
Horace Lamb Theatre
A.M. Simpson1.G. White
R.D. AllenT.K. Roberts
A. StojanoffH. BourneR.V. Hyne
Y. ChandrasekharM.J. D'O,<chioB.P. Setchell
Horace Lamb Theatre
H. IshidaG.P. RisbridgerD.M. de Kretser
Blood flow in the ovary and oviductof the rat after sympathetic denervation.
Chairman, Dr H.W. Baker
Ovarian secretion of progestinsduring late pregnancy in the rat.
The failure of thyroid allograftsin the ovine testis.
Ovarian ~xytocin and progesteronesecretion and degranulation of lutealcells in sheep given exogenous prostaglandins.
Variation in the effect of a nongonadotrophic Leydig cell stimulating factor in testicular interstitial fluid (IF) after exposureof the testis to a single episodeof heat treatment.
Measurement and manipulation ofcytoplasmic free calcium of ram andboar sperm.
Isolation of the guinea pig spermacrosome and characterization of itsenzymic content
Immunosuppressive and cytotoxiceffects of seminal plasma.
The influence of the pituitary onovarian compensation followingunilateral ovariectomy in sheep.
Pubertal development in ram lambswith induced hyperthyroidism orreceiving maintenance feedings
Session 6: THE OVARY: Oral presentationsChairman, Dr B. Bindon
47
460830
0845
1130 56
1145 57
Vascular growth in developing lutealtissue.
Flow rate and composition of ovarianlymph in the day 16 pregnant rat.
x
K. I. MackinG.T. Meyer
A.M. Dharmarajan,N.W. BruceH.J. McArdle
68 The effects of ethylene dimethanesulphonate on adult rat leydig cells.
XI
A.E. JacksonP.C. O'LearyM.M. AyresD.M. de Kretser
'vi

69 Is there a paracrine regulator of G.P. Risbridgertesticular Leydig cell steroidogenesis? J.A.Muir
G. Jenkinn.M. de Kretser
1400-1530
Sessiqn 7eB) THE OVARY: Poster Discussion
Chairman, Dr L. Cahill The Crit Library
Pubertal development in bull calves M.J. D'Occhioactively immunized against testosterone n.R. Giffordand oestradiol-17 S R.M. Hoskinson
T. WeatherlyP.C. FlavelP. E. MattnerB.P. Setchell
Inverse relationship between liveweight N.R. Adamsand uterine oestrogen receptors in A.J. Ritarovariectomized ewes.
An interaction between progesterone P.J. Wrightpretreatment and stage of the anoestrous K.E.Davisseason the ovulatory response of ewes J.D. O'Sheato bolus GnRH treatment. J.A. Burden
70
71
72
73
74
75
76
77
The unilateral cryptorchid rat model:Evidence for local as well as humoralcontrol of Leydig cell recoveryfollowing Ens administration.
The effects of ethylene dimethanesulphonate (EDS) on the bilaterallycryptorchid rat testis
Acute gonadotrophin responses tocastration in prepubertal and pubertalbulls.
Response of the epididymis in ram lambsto oestradiol-17S
Extracellular matrix modulation ofproteoglycan production in Sertolicell-myoid cell co-cultures.
Seminiferous tubule fluid productionin potassium-depleted rats.
Endocrine variation between crossbredMerino rams with and without a copyof the Booroola ! gene.
XII
P. O'LearyA.E. JacksonG.P. Risbridgern.M. de Kretser
D.M. de KretserA. E. JaCKsonP.C. O'LearyS.A. Averill
D.R. GiffordM.J. D'OcchioT. WeatherlyB.P. Setchell
C. PapachristoforouM.J. n'OcchioD. HorsefallW. TilleyB.P. Setchell
E.W. ThomsonJ.S.H. ElkingtonA.M. Blackshaw
C.L. AuC. WangJ.P. QiuP.Y.D. Wong
I.W. PurvisB.M. BindonT.N. EdeyM.A. HillardL.R. Piper
If
lb.
78
79
80
81
82
83
84
85.
86
87
88
Ovarian compensatory hypertrophy inunilaterally ovariectomised anDestrousewes.
NADPH stimulation of aromataseactivity in ovine granulosa cells.
A comparison of luteal progesteroneand oxytocin release and theirsubcellular storage.
Superovulation and embryo recoveryin Awassi fat tail =~eep.
Plasma oxytocin concentration inheifers at midcycle following acloprostenol stimulas.
Do small luteal cells differentiateinto large luteal cells during theovine oestrous cycle?
Estimated utero-ovarian productionrates of steroids in unmated and inpregnant gilts between days 9 &17after oestrus/coitus.
Oxygen consumption by the ovary of thepregnant rat.
Effects of pyruvate and lactate onin vitro development of I-cell pigembryos.
XIII
B.K. CampbellR.J. ScaramuzziJ.A. DowningG. Evans
L.A. HutchinsonS.M. CampoR.S. CarsonJ.K. Findlay
G. JenkinG.E. RiceG.D. Thorburn
W. M. C. MaxwellJ.P. RyanR.G. CaseyR.P. LewerA. Louca
S. MockJ. ParsonsM. GardnerS. McPheeR.J. Fairclough
J.D. O'SheaR.J. RodgersP.J. Wright
B.A. StoneO.M. PetruccoP. QuinnR.F. Seamark
R. SwannN. Bruce
A. MichalskaB.A. StoneP. QuinnR.J. Ashman

Development of sheep embryos in vitro B.G. Millerin a medium supplemented with~ifferent N.W. Mooreserum fractions. P.A. Batt
rb
89
90
91
92
93
94
95
96
97
98
99
Biological activity of cronolone inthe sheep, mouse and rabbit.
Structural and functional involutionand regeneration of the uterus ofpost-partum ewes.
Protein secretion by endometrialepithelial cells in early pregnancyin the ewe.
Oxytocin binding in sheep endometriumduring the oestrous cycle and earlypregnancy.
NA+-Glycine co-transport inpFeimplantation mouse embryos.
The progesterone receptor antagonistRU 486 stimulates prostaglandinproduction in human endometrial cells.
Characterization of the mouse blastocyst glucose transporter.
The intercotyledonary endometrium ofthe ewe during pregnancy: Progesteronecontent, protein secretion and ultrastructure.
Phagocytic properties of murinetrophoblast in vitro.
Effect of time of transfer on survivalof embryos in ovariectomized ewes.
XIV
x. ZhangG.M. StoneB.G. Miller
A.H. WilliamsP.J. WrightJ.D. O'SheaI.J. Clarke
L.A. SalamonsenS.D. PhilpottB. DoughtonJ.K. Findlay
S.D. PhilpottB. DoughtonL.A. SalamonsenJ.K. Findlay
J.G. HobbsP.L. Kaye
D.L. HealyR.W. KellyM.J. CameronLT. CameronD.T. Baird
H. GardnerP.L. Kaye
Y. TangG.M. Stonex. ZhangB.G. Miller
B.L. DrakeJ.C. Rodger
N.W. MooreB.G. Miller
102
104
105
106
107
108
·109
Session 7(C) COMPARATIVE REPRODUCTIVEBIOLOGY: Poster Discussion
Chairman:
Withdrawn
Structural features of the epididymisin a dasyurid marsupial.
Protein secretion by the epididymisMacropus eugenii.
rian function and female sexutivity during the breeding season
greyheaded flying fox Peropushalus.
pig
on embryo
Response to Hypoxemia after administration of nerve growth factor antibodies in foetal lambs.
XV
JonesWorrardSetchell
L. MartinP. Towers
C.D. MatthewsK. PorterI. CookeM. GuerinD. Bigham
B.A. StoneP. QuinnR.F. SeamarkS. Deam
N.K. KhuranaR.G. Wales
R.G. WalesC.L. Cuneo
B.M. BindonL.R. Pl.perY.C. CurtisR.D. Nethery
V.G. HanfH.R. TinnebertL. MettlerT.K. Roberts
SchuijersWalkerBrowneThorburn

Session 7(C) COMPARATIVE REPRODUCTIVEBIOLOGY: Poster Discussion
1400-1530 Chairman: Dr M. Renfree
The Crit LibrarySymposium:BREEDING
Wednesday, August 28
PROSPECTS FOR CONTROLLED SHEEPA TRIBUTE TO T. J. ROBINSON. Horace Lamb Theatre
100 WithdrawnChairman: Dr J. ,~Q.l*&y,
Dr R.J. Scaramuzzi and Dr J.F. Smith.
Prof. D.R. Lindsay
The speakers will be as follows
Prof. N.W. Moore
Dr R. Ortavant
1030-1100
1100-1145
0915-1000
0830-0915
D.A. TaggartP.D. Temple-Smith
Structural features of the epididymisin a dasyurid marsupial.
R.C. JonesK.C. WorrardB.P. Setche11
Ovarian function and female sexual L. Martinreceptivity during the breeding season P. Towersin the greyheaded flying fox Pteropuspo1iocephalus.
Protein secretion by the epididymisof the tammar, Macropus eugenii.
103
102
101
104 The establishment of a common marmosetcolony (Ca1lithirix Jacchus) andobservations on the circadian restactivity cycle using a microcomputer.
C.D. MatthewsK. PorterI. CookeM. GuerinD. Bigham
1145-1215
1215-1300
Prof. T.J. Robinson
DISCUSSION
Session 8: THE UTERUS & THE EMBRYO: Oral Presentations
Chairman, Dr C. Nancarrow Horace Lamb Theatre
Session 9:BIOLOGY,
COMPARATIVE REPRODUCTIVEOral Presentations Horace Lamb Theatre~
1615 106
1600 105
1630 107Aberrant spermiogenesis in a finchhybrid.
J.C. RodgerT.P. FletcherC.H. Tyndale-Biscoe
P. TowersL. Martin
P.A. HufA.R. BourneT.G. Watson
M.A. Swan
L.A. HindsJ.D. HarderC.A. HornC.H. Tyndale-Biscoe
M.A. McGuckinA.W. Blackshaw
Prof. R.V. Short
Active anti-paternal immunization doesnot affect the success of marsupialpregnancy.
PMSG induced ovulation in the flyingfox Pteropus scapulatus.
Progesterone and oestradiol inperipheral and utero-ovarian venousplasma during the peripartum periodin the tammar.
Identification of C19-and CZ1steroid conjugates ~n the male lizardTiliqua Rugosa.
Photoperiod influences reproductionin the male fruit bat, Pteropuspoliocephalus.
Chairman:
116
115
114
113
112
111
1515
1500
1430
1415
1400
1445
2..7
1
R.G. WalesC.L. Cuneo
B.M. BindonL.R. PiperY.C. CurtisR.D. Nethery
V.G. HanfH.R. TinnebertL. MettlerT.K. RobertsJ.A. SchuijersD.W. WalkerC.A. BrowneG.D. Thorburn
B.A. StoneP. QuinnR.F. SeamarkS. Deam
N.K. KhuranaR.G. Wales
Early pregnancy factor followingtherapeutic abortion.
Response to hYpoxemia after administration of nerve growth factor antibodies in foetal lambs.
Incorporation of labelled amino acidsby sheep embryos.
Release of steroids by cultured pigblastocyts.
Fremartins in Booroo1a Merinopopulations.
Effect of uterine proteins on embryometabolism.
110
109
108
1715
1700
1645
XVI XVII

Author Page No. AuthorPage No.
Adams ,N.R. -------------------------------------- 78
Batt, P.A. ------------------------------------- 89
Bigham, D. ------------------------------------- 104
Bindon, B.M. ---------------------------- 14,41,77 ,108
Blackshaw, A. W. ---------------------------- 45,75,114
Boland, M.P. ----------------------------------- 6
Bourne, A. R. ----------------------------------- III
Bourne, H. ----------------------------------- 7,48
Brooks, D.E. --------------------------------- 59,67
Brown, B.W. ------------------------------------ 40
Brown-Woodman, P.D. ----------------------------- 15
Browne, C.A. ---------------------------------- 110
Bruce, N.W. ------------------------------- 53,57,86
Burden, J.A. --------------------------------- 26,87
Burger, H.G. ----------------------------------- 16
Cahill, L.P. ----------------------------------- 52
Cameron, A.W.N. -------------------------------- 2,58
Cameron, I.T. ---------------------------------- 95
Cameron, M.J. ---------------------------------- 95
Campbell, B.K. ---------------------------------- 79
Campo, S.M. ------------------------------------ 80
Carati, C.J. ----------------------------------- 13
Caro, C.M. ------------------------------------- 25
Carson, R. S. --------------------------------- 39,80
Casey, R.G. ------------------------------------ 82
Castaneda, E. Herrera ---------------------------- 64
27
46
65
7
19,46,71,73,74
1
87
-------------------------------
----------------------------------
__________---------------------- 10,33________________________________ 27,28
107___________________________________ 108
38
------------------------------------------------------------------------
-----------------------------~-------
--------------~---------------------
------------------------------------
--------------------------------------
duPlessis, Y.
Doughton, B.
Downing, J.A. ---------------------------------
Drake, B.L. ------------------------------------
de Kretser, D.M. -------------- 4,8,16,43,47,68,69,70,72
de Witt, Minnie ----------------- 9
Deam, S.--------------------------------------- 105DeGaris, R.M. 3
DeYi, Liu ------------------------- 44Dharmarajan, A.M. 53,57
_________________________________ 92,93
79
98
7
77
Davis, I.F.
Davis, K.E.
Cutler, S.A. ----------------------------------
D'Occhio, M.J.-------------------------
Curtis, Y.C.
Cummins, J .M.
Cummins, J. T.
Cuneo, C.L.
Elkington, J. S.H. -------- 45,75
Evans, G. .:.______________________ 38,62,79
Fairclough, R.J. 55,83
Fafrnie, I.J. ------------------------- 2,58
Findlay, J .K. -------------------- 80,92,93
Flavel, P. F. ----------------------- 36,71
Cooke, I.
Cooper, E.J. -----------------------------------
Cormack, J.
Cox, R.I. -------------------------------------
Creed, K.E. ------------------------------------
Croker, K.P. -----------------------------------
Cheng, Zhang Zhong ----------------""':'-------------
Chiappazzo, L.----------------------------------
Clarke, I.J. ------------------- 11 ,26 ,27 ,28,39,52,91104
44
51
18
13
21
Chandrasekhar, Y.
Cavanagh, Lynda ---------------------------------
41
50
36
32,88
76
72
63
68
95
Ashman, R.J----------------------------------
Au, C.L. --------------------------------------
Averill, S.A. ---------------------------------
Ayodeji, O. ------------------------------------
Ayres, M.M. -----------------------------------
Baird, D. T. ------------------------------------
Al-Obaidi, S.A.R. ------------------------------
Allen, R.D.-------------------------------------
Armstrong, D. T. ---------------------------------
. .Baker, H.W~G. --------------------------
XVIII XIX

Author Page No. Author Page No.
6
57
1
106
61
19
24
39
43
7
82
9,17
35
56
51
21.82
2,58
18
7
82
69.81
21
-------------------------------------
Maddocks. S. ----------------------------------Marshall. J.T.----------------------------------
Martin. L ----------------------------------- 103.115
Martinez, Francesca ----------------------------- 17
5
McArdle. H.J.-----------------------------------
Mackin. Karen 1.
Lutjen, P.
Lynch. P.R.
Matson. P. -----,--------------------------------
Matthews. C.D. -------------------------------- 20.104
Mattner. P.E. --------------------------------- 40,71
Maxwell, W.M.C. -------------------------- 29.30,62.82
Louca. A. --------------------------------------
Lightfoot. R.J. ------------------------------
Lindsay. D.R. -------------------------------
Little. D.L. ----------------------------------
Lopata. A. -------------------------------------
Levran. D.
Lewer. R.P.
Kleemann. D.D.---------------------------------Langdon. P.J. ---------------------------------
Langsford. D.A. --------------------------------
Laws. A.D. -------------------------------------
King. G.F.
Khurana. N.K.
Jenkin. G.
Kerton, D.J.
Jones. A.R. ---------------------------------- 60.61
Jones. G.M. ------------------------------------ 7
Jones. R. C. ------------------------------------ 102
Junk. S. --------------------------------------- 5Karsch. F.J. ----------------------------------- 28.
Kaye, P.L. ----------------------------------- 94.96
Kelly. R.W. ------------------------------------ 95
Keogh. E.J. -------------------------------- 2.13.58
Johnson, T.J.-----------------------------------
34
116
52
65
96
83
34
54
Gardner. H.
Gardner. M.
Hayes. M.K. -----------------------------------
Hazelton. I.G. ---------------------------------
Healy. D.L. ---.--------.-------------------------
Gede Putu, I. ---------------------------------
Gibson. W.R. -----------------------------------
Gifford. D. R. ----------------------------- 36.71.73
Godfrey. B. ------------------------------------ 20
Gordon. K. ------------------------------------- 11
Guerin, M. ------------------------------------- 104
Hanf, V.G. ------------------------------------- 109
Harder. J.D • .,..---------------------------------- 113
Haszard. P. ------------------------------------ 35
59
6
95
22Henderson. J.
Fletcher. I.C. -'--------------------------------
Fletcher. T.P.----------------------------------
Fry. R.C. -------------------------------------
Galloway, D.B.----------------------------------
Hillard. M.A. -------------------------------- 41.77
Hinds. L.A. ------------------------------------- 113
Hobbs, J .G. ------------------------------------ 94
Horn. C.A. ------------------------------------- 113
Horsefall. D. ---------------------------------- 74
Hoskinson. R. M. ------------------------------- 6.71
Hoy. Julie ------------------------------------- 17
Hudson. B. ------------------------------------- 16
Huf. P.A. -------------------------------------- III
Hunton, J. R. --------------------------------- 29.30
Hutchinson. L.A. -------------------------------- 80
Hyland. J. H. ----------------------------------- 39
Hynd, P. I. -------------------------------------, 23
Hyne. R. V. ------------------------------------- 48
Ishida. H. --------------------.----------------- 47
Jackson, A.E. ------------------------------ 68.70.72
Jeffcott. L. B. ---------------------------------- 39
xxXXI

Author Page No. Author Page No.
McGuckin. M.A. ---------------------------------- 114
McPhee, S. ------------------:-------------------- 83
Mettler. L. ------------------------------------ 109
Meyer, Geoffrey T. ------------------------------ 56
Michalska, A. ----...,,------------------------------ 88
.Miles, M.A. ------------------------------------ 24
Miller, B.G. ---------------------------- 89.90.97,99
Miller. C.M. ----------------------------------- 35
Mo~, S. --------------------------------------- 83
Moore. N.W. ---------------------------------- 89,99
Moore, R. W. ------------------------------------ 35
Muir, J .A. ------------------------------------- 69
Murray, J. D. ----------------------------------- 6
Nancarrow, C.D. --------------------------------- 6
Nethery, R.D. ---------------------------------- 108
Nottle, M.B. --------------------------------- 23,36
o'Halloran, F. ---------------------------------- 5
O'Leary, P.C. ----------------------------- 68,70,72
a 'Neill, C. ------------------------------------ 12
O'Shea, J.D. ---------------------------- 55,84,87,91
O'Shea. T. ------------------------------------- 41
Oldham, C.M. ----------------------------------- 2
Panaretto, B.A. --------------------------------- 40
Papachristoforou, C. --------------------------- 19.74
Parr, R.A. ----------------------------------- 1,24
Parsons, J. ------------------------------------ 83
Pennefather, J .N. ------------------------------- 3
Petrucco, a.M. ---------------------------------- 85
Phillips, D. ----------------------------------- 14
Philpott, Susan D. ---------------------------- 92, 93
Pholpramool, C. --------------------------------- 66
Piper. L. R. ------------------------------- 14,77 .108
Porter, K. ------------------------------------- 104
Purvis, I. W. ----------------------------------- 77
Qiu, J.P. -------------------------------------- 76
Quinn, P. ------------------------------- 32,85.88,105
XXII
Reid, T.A. ------------------------------------- 60
Renfree, M.B. ---------------------------------- .11
Rennie. G. C. --------------------------------- 16,22
Rice, G.E. ------------------------------------- 81
Ridley, D. ------------------------------------- 15
Risbridger, G.P. --------------------------- 47,69,70
Ritar, A.J. ------------------------------------ 78
Roberts, T.K. -------------------------------- 50,109
Roche, P.J. ------------------------------------ 54
Rodger, J .C. --------------------------------- 98,116
Rodgers. R.J. ---------------------------------- 84
Ryan, J. P. -------------------------------- 29,30,82
Salamon, S. ------------------------------------ 62
Salamonsen, L~A. ------------------------------ 92,93
Scaramuzzi, R.J. ---------------------------- 6.38,79
Schuijers, J .A. --------------------------------- 110
Seamark, R. F. ------------------------------ 23,85,105
Setchell, B.P. -------------- 19,23,46,51,66,71,73,74,102
Sharpe, P. H. ----------------------------------- 36
Shelton, J.N. ---------------------------------- 31
Short, R. V. ------------------------------------ 11
Simpson, A.M. ---------------------------------- 49
Smith, D.H. ---------------------------------- 19,32
Staples, L. D. ---------------------------------- 55
Stelmasiak, T. --------------------------------- 37
Stevenson, D. ---------------------------------- 61
Stojanoff, A. ---------------------------------- 48
Stone, B.A. ------------------------------- 85,88,105
Stone, G.M. ---------------------------------- 90.97
Straffon, W.G.E. -------------------------------- 16
Swan, M.A. -------------------------------- 15,42,112
Swann, R. ------..:.------------------------------- 86
Szell, A. -------------------------------------- 31
Taggart, D.A. ---------------------------------- 101
Temple-Smith, P.D. --------------------------- 4,64,10
Tang.Y ------------------------------------------97
XXIII
Yanagimachi, R. --------------------------------- 10
Yates, C.A. ------------------------------------ 8
Yovich, J .L. -----------------------------"':'----- 5
Zhang, X. ------------------------------------ 90,97
Zhang, Z.C. ------------------------------------ 26
Zupp, J .L. ------------------------------------- 66
Thompson, E.W. -------------------------------- 45,75
Thorburn, G.D. -------------------------------- 81,110
Tilbrook, A.J. ---------------------------------- 58
Tilley, W. ------------------------------------- 74
Tinneberg, H.R. --------------------------------- 109
Towers, P ----------------------------------- 103,115
Trounson, A.D. --------------~----------------- 9,25
Tyndale-Biscoe, C.H. -------------------------- 113,116
Van Mourik, S----------------------------------- 37
Vishwanath, R. ---------------------------------- 42
Waddell, B.J. ---------------------------------- 53
Wales, R.G. --------------------------------- 106,107
Walker, D.W. ----------------------------------- 110
Walker, S.K. ---------------------------- 14,18,19,32
Walkley, J .R.W. --------------------------------- 14
Wang, C.--------------------------------------- 76
Watson, T.G. ----------------------------------- III
Weatherly, T. -------------------------------- 71,7
Webb, R. I. ------------------------------------- 33
Welch, R.A. S. ---------------------------------- 35
White, I.G. ----------------------------- 15,42,49,60
Williams, A.H. ---------------------------------- 91
Wilton, L.J. ----------------------------------- 4
Wishwanath, R. ---------------------------------- 15
Wong, P. Y. D. ----------------------------------- 76
Worrard, K. C. ---------------------------------- 102
Wreford, N.G.M. --------.,.------------------------ 43
Author
Thomas, G.B.
Wright, P.J.
Page No.
27
26,39,84,87,91
PROGESTERONE THERAPY TO IMPROVE FERTILITY AFTER UTERINE A.I. IN EWES.
I.F. Davis, D.J. Kerton and R.A. Parr.
Department of Agriculture and Rural Affairs, Vic.Animal Research Institute, Werribee, 3030.
A technique for intrauterine insemination of ewes has beendeveloped (1) and using this technique it is possible to reduce thesperm dose to 12 x 106 sperm in fresh semen (2). The experimentreported here examined the effect of hormonal treatments on thefertility of ewes subjected to uterine A.I.
Three hundred Merino ewes had intravaginal 'Controlled InternalDrug Release' (CIDR) devices, which contained 9% progesterone,inserted for 12 days. At CrDR removal the ewes were divided intofour equal groups. These were a control group which had nofurther hormonal treatments, a group which received an injection(s.c.) of 400 i.u. of PMSG (Folligon, Intervet) at CrDR removal, agroup which received an injection (i.m.) of 25 u.g. of GnRE(Fertagyl, Intervet) at A.I. and a fourth group which had anotherCIDR inserted from days 10 to 16 after removal of the synchronizingCIDR. Ewes were joined with 4% of vasectomized rams and checked foroestrus on the evening before, and again in the morning and earlyafternoon on the day of A.I. Ewes which had shown oestrus weredrafted into separate groups after each observation and wereinseminated in order of oestrus detection. Semen was collected byartificial vagina and used within two hours of collection.Insemination of 0.05 of semen, diluted with UBT milk and containing10 x 106 sperm, into each uterine horn was carried out 50-56 h afterCIDR removal. Foetuses were counted by ultra-sound scanning 70 daysafter A.I. Data were examined by analysis of Chi-square.
Table 1. Fertility after hormone treatment.Treatment Lamb % Pregnancy rate % Foetuses per ewe %Control 54a 51 a 106aProgesterone 75b 60a 126bPMSG 69b 58a 121 bGnRB 56a 50a 111 aa, b f data with unlike superscripts differ, P<0.05.
Treatment of ewes with PMSG or progesterone during Days 10-16increased lambing percentage and the number of foetuses per ewepregnant. Injection of GnRB had no beneficial effect. The studyconfirms an earlier report (3) that progesterone therapy for 6 daysafter mating increases lambing rate probably by a reduction inembryo mortality. Mild superovulation with PMSG has a similareffect, possibly brought about by an increase in the amount ofof luteal tissue.
(1) Killeen, I.D. and Caffery, G.J., Aust.Vet.J. 22 : 95 (1982).(2) Davis, I.F., Kerton, D.J., McPhee, S.R., Grant, I. and Cahill,
L.P. Proc. Aust •. Soc. Reprod. BioI. 16 : 101 (1984).(3) Peterson, A.J., Barnes, D., Shanley, ~ and Welsh, R.A.S.
Proc. N.Z., Endocr. Soc. ~ 13 (1984).
XXIV
2
NUMBER OF SPERMATOZOA AND OVULATION RATE AFFECT FERTILITYAND PROLIFICACY OF SHEEP
A.W.N. rameron+, C.M. Oldham+, I.J. Fairnie¢, E.J. Keogh* and D.R.Lindsay
+Departments of Animal Science asd *Clinical Biochemistry, Universityof Western Australia, Perth and Muresk College, Northam, W.A.
Sperm transport may be improved in ewes given exogenous oestrogen(1), and peak plasma levels of oestradiol-17B vary with ovarianfollicular development (2), hence increases in ovulation rate mightimprove sperm tansport. Thus we examined whether the maximum level offertility was higher in ewes having more than 1 ovulation and whetherthe optimum dose of spermatozoa was lower in such ewes.
Mature Merino ewes were artificially inseminated in Februarywith either 25, 50, 100, 200 or 400 million sperm. Ewes were derivedfrom flocks which were a) supplemented with 750 g per head per day oflupin seed for at least 8 days before insemination b) similarlysupplemented with lupins as well as immunized against androstenedione,6-7 and 3-4 weeks before insemination, using Fecundin (Glaxo) and c)untreated control ewes. The number of ovulations for each ewe wasdetermined by laparoscopy 5 days after insemination and the number offoetuses counted using realtime ultrasound scanning 36-45 days afterinsemination. Data were analysed by fitting generalized linear models.
The mean ovulation rates were 1.41, 1.73 and 1.11 for the lupin,lupin-fecundin and control flocks respectively. The pregnancy rate(table 1) was increased in ewes with more than 1 ovulation (P<0.05),and reduced in ewes treated with Fecundin (P<0.05), neither of whicheffects were dependent on the number of sperm. Fertility varied withnumber of sperm (P<0.05).
Table 1. The percentage of ewes pregnant (n) .Ovulation rate: 1 >1No sperm (millions): 25 50 100 200 400 25 50 100 200 400Ewe groupControl 38 45 44 33 38 40 67 67 60 100
(48 ) (51 ) (48) (40) (48) (5 ) (6) (3 ) (10) (3 )Lupin 19 48 52 52 53 39 71 44 50 50
(32 ) (25 ) (27) (29) (40) (18) (24) (16 ) (20 ) (12 )Fecundin-lupins 33 33 30 44 30 54 49 36 50 44
(24) (27) (20 ) (25) (20) (38) (39) (37) (39 ) (40)
Both reducing the number of sperm inseminated or treatment withFecundin reduced the proportion of twin-ovulating pregnant ewes thatbore twins (P<O.Ol and P<0.05 respectively).
We conclude that fertility increases with ovulation rate butthis is not due to improved sperm transport. Although Fecundinincreases ovulation rates fertility is reduced in treated ewes.Finally the number of sperm inseminated affects prolificacy, presumablyby affecting the level of embryonic wastage.
(1) Hawk, H.W and Cooper, B.S. (1975) J. Anim. Sci. 11: 1400-1406.(2) Evans, G. and Robinson, T.J. (1980) J. agric. Sci., Camb. 94: 69-88
3
REINNERVATION OF THE RAT VAS DEFERENS FOLLOWINGVASOVASOSTOMY AFTER VASECTOMY
R.M. DeGaris and J.N. Pennefather
Department of Pharmacology, Monash University,Clayton, Victoria, 3168.
A rapid decline in catecholamines and loss of response to stimulation of intramural nerve terminals occurs in the epididymal (proximal)segment of the rat vas deferens following vasectomy by medial transection (1-3). Reinnervation does not occur during the four weekperiod following surgery, and is minimal 3 months later. Segments oftransected vasa deferentia have now been examined after vasovasostomyto gauge the degree of reinnervation associated with return of fertility.
Long-Evans rats (200-300 g) were unilaterally or bilaterallyvasectomised. Four weeks later the transected halves of one vas deferensin each animal were rejoined. All surgery was conducted using halothaneanaesthesia. Eight weeks later rats were killed by cervical dislocationand epididymal and prostatic halves of each vas deferens were examinedfor catecholamine fluorescence or set up in vitro for recording forcedeveloped in response to electrical field stirr-ulation (10 pulses, 60V,lms, 0.1-50 Hz) as described previously (4). Use of unilaterally andbilaterally vasectomised rats permitted comparisons between epididymalsegments from rejoined vasa deferentia with corresponding segments fromunoperated and transected vasa deferentia respectively.
Epididymal segments from rejoined vasa deferentia exhibited morecatecholamine fluorescence than transected segments but less than unoperated segments. Noradrenaline was equipotent in contracting epididymal segments isolated from unoperated and rejoined vasa deferentia. Forcedeveloped in response to field stimulation in segments from rejoined vasadeferentia was significantly less than that developed by unoperated segments (t-tests, P<0.05 10d.f.); differences were greatest above 2 Hz. Incontrast, catecholamine fluorescence in prostatic segments and theirresponses to noradrenaline and to stimulation were relatively unaffectedby the operative procedures. To determine when fertility was restored,six rats bilaterally vasovasostomised four weeks after bilateral vasectomy were mated with females of known fertility 4-8 weeks after vasovasostomy. pregnancies with successful outcome resulted from matingswith five of the operated males.
If the rates of reinnervation of the vas deferens are similar afterunilateral and bilateral vasovasostomy, these findings indicate thatcomplete noradrenergic reinnervation of the rat vas deferens is notessential for return of fertility after vasectomy and subsequent vasovasostomy .
(1) Norberg, K.A.; Rimky, P.L. and ungenstedt, U. Experientia Zl:392-394(1967)
(2) DeGaris, R.M., Hartley, M.L., Leedham, J.A. and Pennefather, J.N.CEn. Exp. Pharmacol. Physio). 10: 703-704 (1983)
(3) DeGaris, R.M. and Pennefather, 'J":'"N. Proc. Aust. Physiol. Pharmacal.Soc. 16: 54 P (1985)
(4) Pennefather, J.N., Vardolov, L. and Heath, P. Clin.Exp. Pharmacal.Physiol. ~: 451-462 (1974).

4
EPIDIDYMAL NECROZOOSPERMIA: A NEWLY DEFINED CAUSE OF ASTHENOZOOSPERMIAAND ITS POTENTIAL TREATMENT
L.J. Wilton, P.D. Temple-Smith, H.W.G. Baker * and D.M. de Kretser.
Department of Anatomy, Monash University and *M.R.C. Prince Henry'sHospital, Melbourne.
. ~fter production in the testes, human sperm require several days inep1d1dymal passage ~nd storage prior to ejaculation. Although it hasbeen sugge:t:d that d1sturbances in epididymal transit could cause poorsperm mot1l1~Y. (asthenozoospermi~. AZS) and consequent infertility nosuch abnorma11t1es have been descr1bed. This report documents for thefir~t time, human AZS due to abnormalities in the e;ididymalenv:ronment and provides methods for improving sperm motility in thesepat1ents.
During electron microscopic (EM) assessment of the sperm from 13AZS men, 3 were identified with severe AZS and necrozoospermia (deadsperm). EM showed that the majority of ejaculated sperm wered:generate~ with. disrupted cell membranes, acrosomal caps andm1tochondr1al cr1stae, and poorly defined axonemal microtubules. Twoo~ th:se patients (J.M. and S.G.) were available for further study.B10ps1es revealed that testicular sperm from these men weremor~hologically normal i~dicating that sperm degeneration occurreddU~1~g transit through the reproductive tract, possibly duringep1d1dymal passage and storage or upon mixing with the seminal plasmaat ejaculation.
The possible presence of a factor toxic to sperm in the seminalplasma of J.M. and S.G. was tested by placing normal sperm in thepa~ients: se~inal plasma and measuring the change in sperm vitality (%a11ve) w1th t1me. Over 44 hours there was no difference in vitalitybetween ~ormal sperm in their own seminal plasma or in that of J.N. orS.G. show1ng that seminal plasma from these patients was normal.
Frequent ejaculations are known to exhaust the extra-gonadal spermreserves a~d decrease t~e.time sperm spend in the epididymis. In anattempt to 1mprove the mot1l1ty of ejaculated sperm, J.M and S.G. wereasked to collect t~o ejacul~tes per day (approximately 12 hrs. apart)for.4~5 days •. ~he f1rst spec1men each day was analysed for spermm~t1l1ty~ mot111t~ index and vitality and processed for EM. ExhaustiveeJa:ulat10n (EE) 1mproved all three semen analysis parameters in thesepat1ents (see tabl:).. ~ showed an increa:ed number of sperm hadacrosomal caps and d1st1ngu1shable axonemal microtubules.
EE DAY 1 2 3 4 5J.M. % MOTILE SPERM 12 7 17 17 17
MOTILITY INDEX 17 8 23 25 31% LIVE SPERM 17 9 30 34 35
S.G. % MOTILE SPERM 2 6 13 ISMOTILITY INDEX 3 6 21 17% LIVE SPERM 9 18 30 27The ~esult~ :tro~gly suggest that this previously undescribed
cause of 1nfert1l1ty 1S of epididymal origin. A distinguishing featurefrom mos~ ~ther types of AZS is the concommitant necrozoospermia. Theter~ .ep1d1dymal necro~oospermia is therefore proposed. Although spermmot1l1ty after EE rema1ned below normal, it increased sufficiently forthese me~ to be considered for IVF by timing the EE so that maximumsperm mot1lity coincides with ovulation in the female partner.
THE USE OF IMMUNOBEADS TO DETECT HU~~N ANTI-SPER~TOZOAL ANTIBODIES
S. Junk, P. Matson, F. O'Halloran and J.L. Yovich*
PIVET Laboratory, Perth, W.A. and >}University of Western Australia.
Infertile couples were routinely screened for the presence of antispermatozoal antibodies (ASAB). This was done by an indirect methodusing immunobeads (BioRad Lab. Pty. Ltd., Australia) specific for IgG,IgA and IgM (1,2).
A cross-sectional study showed that ASABs were detected in the serum of8/95 (8.4%) men with the class of antibody being IgG in 2/8 (25%), IgAin 1/8 (12.5%) and both IgA and IgG in 5/8 (62.5%). All of the men withASAB in the blood had corresponding antibodies in the seminal plasma.Interestingly, there were an additional 6 men who gave negative serumresults but had either IgA (n = 4) or both IgA and IgG (n = 2) in thesemen. No IgM antibodies were detected in either the serum or the semen.Seven of the men were included in the IVF program and overall afertilization rate of 18/35 (51.4%) oocytes was obtained. The rate washigher for IgA antibody in semen (11/13, 84.6%) compared to combined IgAand IgG antibody (4/17, 23. 5%; p < 0.001). The postcoital test (PCT)was performed in six couples in which the male partner had either IgA(n = 1) or IgA and IgG (n = 5) in semen, and all six had a negative PCT.
Analyses of the blood of 105 women gave positive results in 9 (8.6%),with the estimated classes being IgA in 6/9 (66.7%) both IgA and IgGin 2 (22.2%) and a combination of IgA, IgG and IgM in 1 (11.1%).Those women who had antibodies of the IgA class only (n = 4) gave apostive PCT in two instances and the other two couples gave a negativePCT. The second group of women (n = 2) had both IgA and IgG antibodiesand both gave a negative PCT. Six of the women were included in the IVFprogram and achieved an overall fertilization rate of 18/30 (60.0%) ofoocytes with 3/6 women conceiving following embryo transfer.
The immunobead test appears to provide a useful screening technique forASAB and allows classification of the antibody type. There appears tobe a relationship between antibody' type in males and the likelihood offertilization. r
(1) Clarke GN, Stojanoff A, Cauchi MN, McBain JC, Speirs AL andJohnston WIH. Am. J. Reprod. Immunol. 5:61 (1984).
(2) Clarke GN, Hsieh C, Koh SH and Cauchi MN. Am. J. Reprod. Immunol.5: 179 (1984).

6
FERTILIZATION AND EMBRYO DEVELOPMENT IN ANDROSTENEDIONE-IMMUNE SHEEP
C.D. Nancarrow, J.D. Murray, R.J. Scaramuzzi, J.T. Marshall,I.G. Hazelton, R.M. Hoskinson and M.P. Boland
CSIRO, Division of Animal Production, Prospect, N.S.W.
Controlled androstenedione-immunization of ewes with Fecundin(Glaxo, Aust.) results in an increased ovulation rate of 40-80% butonly a 15-30% increase in lambing rate. This reproductive wastage isgreater in Merino ewes than cross-bred ewes. We have carried out twoexperiments in 1984 and 1985 to examine this loss in Merinos.
Ewes were immunized twice with Fecundin, 4 weeks apart, theboos ter being given 14 days before expected rna ting (1 0% rams) at thesecond oestrus following intravaginal sponge removal. Control eweswere untreated. Ovulation (OR) and recovery rates (RR) of eggs andembryos were obtained at 2, 13 and 24-32 days (Expt 1) and 2, 9 and13 days (Expt 2) after mating. Fertilization rates (FR) wereassessed by cleavage stage or presence of sperm; embryos wereclassed as abnormal if development was significantly retarded;length measurements (L) were made on day 9 and 13 embryos and day 13embryonic discs (ED); day 25 embryos (less membranes) were weighed(Wt) (Table 1). Statistical analysis (P) was by X20r ANOVA.
TABLE 1. Effect of androstenedione immunization on OR, RR{%), FR{%),normal embryos{%) and embryonic development. (Number of ewes/embryos).
EXPERIMENT 1 EXPERIMENT 2Day Control Immune P Control Immune P
OR 1 .42{ 169) 2.16{ 179) <.01 1.46( 160) 2.25(145) <.01RR 2 89.3 (57) 60.5 (60) <.001 91.6 (50) 76.0 (46) <.02FR 2 86.7 78.2 .25 89.2 83.6 .88RR 9 77.9 (50) 64.7 (44) >.05Normal 9 88.3 63.6 <.01RR 13 77.5 (54) 56.4 (52) <.01 91.8 (60) 68.8 (55) <.001Normal 13 92.7 63.6 <.001 96.2 76.1 <.001RR 25 70.3 (50) 41.2 (39) <.001Normal 25 96.2 96.4 .57
L (\.lm) 9 260 (51 ) 203 (43) <.001L (mm) 13 8.52 (42) 3.54 (24) <.05 6.16 (75) 2.91 (70) <.001ED{\.Im) 13 331 (65) 248 (49) <.001Wt{mg) 25 211 (14) 167 ( 10) <.05
Fecundin t::-eatment caused at 53% increase in OR but RR of eggsand embryos were decreased at all times. At 24-32 days after mating ahigher proportion of control ewes were pregnant (P<.05). FR wereunaffected. The percentage of abnormal embryos collected at days 9and 13 were higher dnd the rates of embryonic deve lopmen t s lower today 25. We conclude that immunological perturbation to the normalsteroidogenic pathways involving androstenedione reduces ovum captureby the fimbria and interferes with embryogenesis. The Fecundintreated ewe provides a model for investigation of the steroidrequirements of early pregnancy.
MORPHOLOGY OF PRE-OVULATORY HUMAN OOCYTES
G.M.Jones, A.Lopata, L.Chiappazzo, Y.duPlessis, H.Bourne and D.Levran.
Department of Obstetrics &Gynaecology, University of Melbourne, andReproductive Biology Unit, Royal Women's Hospital, Carlton, Victoria.
The aim of this study was to evaluate the meiotic maturity and thestatus of the cytoplasm and vitelline membrane of oocytes that werebeing pro,' .... .:ed by ovarian hyperstimulation in an in vitro fertilizationprograrnm_' .
Pre-cJuiatv'y oocytes were obtained from women treated with acombination of ~Iomiphene citrate, human menopausal gonadotrophin andhuman chcrinnic gonadotrophin (HCG) as previously described (1). Whenfour or more oocytes were obtained from a patient, one or more oocyteswere rand~nly selected for the morphological study. All of the oocyteswere enC1JS2d by a cumulus mass and were fixed in 3.5% glutaraldehydein cacodylate buffer within 30 minutes of aspiraton of their follicles.Oocytes were post-fixed in 1% osmium tetroxide, dehydrated and embeddedin Spurr resin. Serial 0.5 ~m sections were mounted on glass.slidesand stained with 1% methylene blue. In some oocytes, ultrath1nsections were cut and mounted on copper grids, doubly stained withuranyl acetate and lead citrate for electron microscopic examination.
Forty-six oocytes were collected and sectioned from pole to pole.Of these, 71.7% were at Metaphase II (MIl), 15.2% were at Metaphase I(MI) or Telophase I (TI) and 13.0% contained a germinal vesicle (GV).Of the 33 oocytes that were at MIl, 12 had recently completed the firstmeiotic division as indicated by the presence of a mid-body close tothe polar body.
The stage of maturity was not related to the number of eggsco11 ec ted per pa ti en t. However, a11 the imma ture (GV) oocytes wereobtained from follicles <4.5 ml in volume.
It was found that many of the oocytes showed varying degrees ofvacuolation (5% - 45%) of the cytoplasm and clumping of organelles (5%_ 85%). The oocytes were also noted to have a reduced complement ofcortical granules, confirmed by electron microscopy.
The number of follicles aspirated per patient, number of eggscollected fertilization rate, embryo development success rate andimplantation rate, were not significantly different between patients inthe study group and those in a control group. The eggs chosen for thestudy were therefore representative of the eggs being produced~by thestimulation programme. It was concluded that 34 hours after HcGinjection was adequate for the majority of oocytes to reach MIl, andthat one third of these had only recently completed the first meioticdivision. It was not clear whether the GV oocytes were derived fromunresponsive or atretic smaller follicles « 20 mm diameter). Thesignificance of vacuolation of the cytoplasm and clumping of theorganelles needs to be further evaluated to determine whether theyrepresent early degenerative changes in the ooycte.
(1) BaylY,C.M., McBain,J.C., Clarke,G.A., Gronow,M.J., Johnston,W.I.H.,Martin,M.J. and Speirs,A.L. Abstracts III World Congress of InVitro Fertilization and Embryo Transfer, p. 106, Helsinki, 1984.

s
MALE FACTOR PATIENTS' FERTILIZATION RATES IN IVF ON THE MOLECULAR BASIS OF HUMAN SPERM-EGG RECOGNITION
C.A.YATES, A.O.TROUNSON & D.M. DE KRETSER Peter Lutjen and Minnie de Witt
Departments of Anatomy andUniversity, Melbourne.
Obstetrics and Gynaecology, Monash Department of Obstetrics and Gynaecology, Monash University,Queen Victoria Medical Centre, Melbourne, Australia 3000.
Sperm bound to zona (mean ± SE)43.4 ± 16.36.5 ± 2.8*
26.2 ± 7.3
E.333025
ControlGDPCDP
* Significantly different from control values (p 0.00001)
The use of the nucleotides guanosine diphosphate (GDP) andcytidine diphosphate as inhibitors of fucosyl and sialyltransferaseactivity respectively has been described elsewhere (4) - Freshlyejaculated and washed spermatozoa were labelled with the fluorescentlabel monobromobimane(5). Unfertilized human oocytes were storedin 0.5M (NH4)2S04 with 1M MgC12 until use (6). Labelled sperm andsalt stored oocytes were placed in in vitro culture 37.5°C for 16 hrin T6 media + 10% fetal calf serum containing 0, 10 roM GDP or 10 ~CDP. Sperm attachment to the zona pe11ucida was assessed byfluorescence microscopy.
TABLE 1. The effect of nucleotide on human sperm-zona binding.Statistical analysis was by the Mann Whitney U Test. Means of9 separate experiments
Cell surface glycosyltransferase enzymes have been implicatedin a number of cellular phenomena. These include cellularrecognition, adhesion and more recently, the processes of mammalianfertilisation (1) . It has also been shown that the fucose richheteropolysaccharide, fucoidin, is a potent inhibitor of spermzona binding in the guinea pig, hamster and human(2). Such a rolefor fucose and its transfer in sperm-zona recognition is furthersuggested by the hiyh fucosyltransferase levels associated withhuman spermatozoa(3 .
The aim of the study was to examine the role of carbohydratetransfer between human spermatozoa and the oocyte zona pellucidathrough feedback inhibition of glycosyltransferase activity.
58.154.254.13.6
53.59.1
41.677.762.671.152.952.7
76
61487
18
48
2419
K.Antisperm antibodiesL.EV patients
Due to the low numbers of sperm (50,000-150,000 sperm/egg)required for in vitro fertilization (IVF), it has been suggested thatIVF could be used as a treatment for male infertility. This studyexamines the fertilization rates of male with subnormal semen in theQueen Victoria-Epworth IVF unit.
Male patients were classified into groups according to themean values of sperm count, motility and morphology from at least 2semen analyses prior to the treatment cycle. Males with subnormal semenwere further classified into groups according to their semen defects,including patients who have had epididymovasostomies (EV) and thus whosesperm is bypassing sections of the epididymis. Procedures for females'treatment cycle and oocyte collection are those described by Trounson etal.,(l). The semen samples were collected 4 hours after oocyte retrievaland treated according to the techniques described by"Mahadevan eta1.,(2). The results are presented in Table 1. "TARLE 1 (n no. of treatment cycles; % fert = % of eggs fertilized)
.!!. % Fert.SINGLE DEFECTSA.O-20% motilityB.20-40% motilitvC.40-60% motility (poor progression)D.>50% abnormal forms£.<5 million sperm/mlF.5-20 million sperm/ml~IULTIPLE DEFECTSC.Low motil.«40%) + Low count «20 mill/ml)11.Low motil.«4G%) + >50% AbnormalsI.Low count «20 mill/ml) + >50% ahnormalsJ.Low count, low motility + abnormal forms
The results show that the fertilization rate while reduced,is at an acceptable level in most cases when compared t~ the normal rate(75%). However, when there is a combination of defects in countmotility and morphology, the fertilization rate is poor (3.6%). This i~also true for the EV patients, where the fertilization rate is very poor(9.1%). These results indicate that, even though the fertilization ratedecreases slightly in most cases, IVF does offer some prospects in maleinfertility.
(1) Trounson, A.G., Leeton, J.F., Wood, C., Webb, J. & Wood, J. (1981)Science 212:381(2) Mahadevan, M.M., Trounson, A.a & Leeton J.F. Fert. Ster. (1983)40: 708 '
The results show that blockage of fucosyltransferase activityresults in a drastic reduction of binding of spermatozoa to thezona pellucida. The decrease associated with a block to siaLyltransfer was not significant.
We conclude that this finding further supports a role forglycosylation of membrane receptors by ectog1ycosyltransferasesin cell-cell interaction, including sperm-zona recognition.
(1) Shur B.D. (1982). In "The Glycoconjugates", Academic Press NY, 145(2) Huang T.T.F. & Yangimachi R. (1984). Exp. Cell Res. 153, 363-373(3) de Witt et al (1984). Proc. Fertil.Soc.Aust. 2:L(4) Scudder P.R. & Chantler E.N. (1981). Biochem.Biophys.Acta
660 : 128-135(5) Huang T.T.F. et al (1984). Bio1.Reprod. 31 : 797-809(6) Yangimachi R. (1979). Fertil.Steri1. 31 : 562-574

10
EFFECTS OF ENZYME INHIBITORS ON HA}fSTER GAMETE INTERACTIONS
J.N. Cummins and R. Yanagimachi
Department of Veterinary Anatomy, University of Queensland,and Department of Anatomy, University of Hawaii.
The site of initiation of the acrosome reaction in the fertilizingmammalian spermatozoon is currently controversial: the traditional viewwas that in most species it occurs in, and is responsible for, the penetration 0f the cumulus oophorus; however a number of recent reports havesuggestvd that it may be initiated by contact with the zona pellucida,and that acr~somal enzymes are irrelevant prior to that stage.
To study this problem freshly ovulated hamster eggs in cumuli werechallenged with spermatozoa under conditions which allowed continuousobservation and examination of the degree of penetration of the cumulusmatrix as well as the state of the acrosome. In vivo capacitated spermatozoa were recovered from the oviducts of females 16-24 hours afternatural mating. In vitro capacitated spermatozoa were obtained using asystem in which the first acrosome reactions and hyperactivated motilitywere seen after 1:45 hours. Gamete interactions were also studied inthe presence of hyaluronidase inhibitors (Na-heparin, 0.5-1.0 mg/ml; Naaurothiomaleate, 0.25 mg/ml) and protease inhibitors (Soybean trypbininhibitor, 1 mg/m1; Benzamidine HCl, 1.2rnM; TLCK, 0.05 roM): these wereadded to uncapacitated and capacitated spermatozoa at various times.
Sperm were examined immediately (30 seconds-5 minutes) after contactwith the cumulus, or after 15-30 minutes of interaction. Only motilespermatozoa were considered at each end-point.
Uncapacitated (less than I-hour preincubated) spermatozoa could notpenetrate the cumulus matrix but usually adhered to the surface singlyor in groups. In vivo capacitated spermatozoa penetrated extremely rapidly, and reached the zona pellucida within a few minutes. Penetrationby in vitro capacitated spermatozoa coincided closely with the first observed acrosome reactions, and the majority of pentrating spermatozoashowed optically modified or absent acrosomal caps: of 287 spermatozoaobserved during penetration only 10(3.5%) appeared to have unmodified,'intact' acrosomal caps. As preincubation times increased, increasingnumbers of spermatozoa were observed which had completed the acrosomereaction in vitro: these attached to the cumulus but were unable to penetrate and did not survive long.
Hyaluronidase inhibitors totally blocked cumulus penetration regardless of the source or treatment of the spermatozoa. Protease inhibitorsdid not prevent capacitated spermatozoa from penetrating provided thespermatozoa were in contact with the inhibitors for less than 30 minutes;longer exposures inhibited penetration even though motility was largelyunaffected and control suspensions treated in parallel could still penetrate.
We conclude that in the hamster, penetration of the cumulus matrixand hence access to the egg for fertilization is accomplished by a phaseof capacitation which involves active release or activity of sperm hyaluronidase, and which precedes final loss of the acrosomal cap. The earlyevents of this sequence involve at least one stage which is susceptibleto protease inhibitors, and this is consistent with current models of themembrane events of the acrosome reaction. Support:UHPS Grant HD-03402,
11
a-ENDORPHIN IN OVINE prrUrrARY PORl'AL BLOOD DURING SUCKLING
*K. Gordon, *M.B. Renfree, +*R.V. Short and I.J. Clarke
Departments of Anatomy*, Physiology+, Monash University andMedical Research Centre, Prince Henry's Hospital, Melbourne
In many mammals the young animal sucking on the teat provides the basisfor a suckling associated suppression of reproductive activity and it has beensuggested that S-endorphin (S-end) may be involved in mediating thisinhibition. In rats, S-end is released by suckling, and intracerebroventricular injection of S-end inhibit LH and increases prolactin (Prl).
Using a previous ly describen method (1) we monitored fluctuations in Send release into the pituitary portal blood of conscious ewes duringlactation, and related these fluctuations to the sucking activity of the lamb.In Group 1 ewes were penned such that their lambs had free access to themthroughout the sampling period. To establish a basF'line S-end release ratebefore the onset of a series of suckling bouts, the Group 2 ewes weresimilarly penned except that the lambs were temp,')rarily exclude r1 from theirdams by a weldmesh ga~e. A third experimental group of non-lactating ewes atvarious stages of the oestrous cycle served as controls.
A pulse of S-end was defined as an increase in release rate of at least25% when compared to the previous sample. Release rate was calculated as thetotal amount of S-end present in the sample per unit time (pg/min). The pulsefrequency was calculated, and the amplitude of each pulse estimated bysubtracting the basal from peak values. In the lactating ewes each pulse wasclassified as due to 1) sucking bout 2) disturbance (e .g. handling theanimal) 3) unknown. There was no significant difference between theampli tudes of pulses associated wi th any of these causes so the results werepooled for comparison with the non-lactating ewes (Table 1).
TABLE 1. Means ± S.E.M. of measured pulse parameters.
GROUP NO. NO. SHEEP PULSE FREQUENCY PULSE AMPLITUDE(pulses/hour) (pg/min)
1 3 4. 26±2.1 2 451.9±57.22 4 2.34±0.24 341.2±39.23 (control) 3 2.13:1:0.64 146.1±16.7
The number of sucking bouts occurring during the collection period wasrecorded and related to whether or not there was an associated peak of S-endrelease. Almost all of the sucking bouts (84%) were followed by a significantrise in pituitary blood S-end release.
The overall mean pulse amplitude of 399.7±31.9 for the sucklinq ewes wassignificantly greater than the mean pulse amplitude of 146.7±16.7 pg/min forthe non-lactating ewes (t-test, P<0.001). There were no significantdifferences in pulse frequency between lactating and non-lactating ewes (MannWhitney U-test).
We conclude that the close association of suckling bouts with subsequentrises in S-end release suggest that S-end may be involved in mediating theinhibitory effects of lactation on reproduction in the ewe.
(1; Clarke, LJ., and Cummins, J.T. Endocr. 111: 1737-1739 (1982).

12 13
PLATELET DERIVED GROWTH FACTORS ARE NECESSARY FOR MOUSEEMBRYO lMPLANTATI ON Ut.:-Y_LIRg
PENILE ERECTION IN THE DOG
~ Carati, K E Creed* and E J Keogh
Department of Obstetrics and Gynaecology, Universityof Sydney, Royal North Shore Hospital, St Leonards NSW
Impotence Study Group of WA, Reproductive Medicine Research Institute,Queen Elizabeth II Medical Centre
*School of Veterinary Studies, Murdoch Uni., Perth WA.
BLASTOCYSTS_1::..;'R;.;..E=..'A:.:.=.-T.:..:M-=E~N-=T U_tJ..Q~.~ G0 I NG 0 UTGRa WT H (% L __!'J_lJ..MJ~_~.& __
10% PRS (V/V) 92 :!: 8 3510% PPS (V/V) 8 .:!: 10 35
1% PRS (from NP mice) 35 .:!: 16 251% PRS (from Day 2 81 :!: 8 25
_ ..____~egnant mice) ----_.._----~--_. __..__ •__._•.. _--"--- --
Table 1 shows that medium supplemented with serumsupports implantation of embryos in-vitro. Whole serumderived from platelet depleted pl~-;-~-;-wa~- not capable ofsupporting significant blastocyst outgrowth. Factorssupplied by platelets appear to have a role in sugportin gblastocyst implantation. They were stable at 56 C butlabile at 70
0C and showed greater activity in pregnancy PRS
compared with non-pregnacy PRS, possibly due to the releaseof platelet factors following early pregnancy associatedplatelet activation.
Platelet activation is an initial maternal response toconception (1) and is caused by the release by thepreimplantation embryo of a soluble platelet activatiingfactor (2). Platelets release an array of biologicallyactive molecules following their activation. The possibilitythat some of these factors playa role in embryo developmentwas investigated.
Blastocyst outgrowth in--~itro_ is considered a model forembryo implantation and was used in this study. Blastocystswere fJushed from the uterus of day 4 pregnant QS mice. Fiveblastocysts per treatment were cultured for 3 days in 20 uldrops of Eagles Minimal Essential Medium under paraffin oiland the proportion of embryos undergoing outgrowth recorded.Medium was supplemented with platelet poor or platelet richserum. Blood from the orbital plexus was mixed with 3.2%(w/v) citrate (9 parts blood:l part citrate) and centrifusedat 2000g for 10 minutes. Plasma was mixed with 10platelets/ml or an equivalent volume of erythrocytes andcoagulation initiated with 2.5 »M CaCl 2/ml of plasma. Theresulting serum was designated platelet poor (PPS) orplatelet rich serum (PRS).
J Physiol 20:372-406 (1896)Acta Physiol Scand 106:199-214 (1979)
1. Langley J N and Anderson H K2. Sjostrand N 0 and Klinge E
The mechanism of penile erection remains a poorly understoodaspect of reproductive physiology, a fact which has hampered advancesin the tredLme~t of impotence. For this reason we have investigatedthe physiology of erection in the dog. The penis receives parasympathet~c iI:nervation from the pelvic nerves and sympatheticinnervation from the sympathetic chain and hypogastric nerves (1,2).We report the effects of electrical stimulation of these nerves.
Twelve mature dogs (10-41 kg, mean 20) were anaesthetised withNembutal and halothane. Through a midline incision the pelvic nerve,hypogastr~c nerve and sympathetic cllain (at level L5) were exposed, cutand mounted on electrodes. They were stimulated individually or incombination. The arterial supply to the penis (dorsal artery) wascannulated for pressure recording (DAP) and close arterial injection ofdrugs. Pressure within the penis (CCP) was monitored via a 21G needleinserted directly into the corpus cavernosum, a part of the penisclosely re~embling that of man. All manipulations were doneunilaterally, on the right side. Systemic blood pressure was monitoredvia the ~arotid artery.
During pelvic nerve stimulation (PNS, 10 Hz) there was animmediate, transient drop in DAP of 10-40% (basal pressure=systemicpressure, 100-160 mmHg). Ten to 30 secs later CCP rose rapidly from abasal pressure of 20 mnHg to reach 80-100% of systemic pressure within60 secs. Elevated CCP was maintained for the duration of PNS (up to 5mins) and the penis was turgid, simulating an erection. When PNS wasstopped CCP decreased to basal pressure within 40-120 secs, often in abiphasic manner. These events were partially blocked by atropine(cholinergic antagonist, 20-40 ug/kg) which caused diminished CCPand/or delayed onset of erection. Propanolol (beta-adrenergicantagonis~, 100 ug/kg) had no effect on the response to PNS.
Sympathetic chain stimulation (SCS, 10 Hz) itself had no effect onthe penis, but it inhibited the effect of PNS. It took 2-4 timeslonger f~r erection to occur, which was then 2-3 times quicker tosubside. A similar effect was observed after administration ofphenylephrine (alpha-adrenergic agonist, 0.5-5 ug/kg). In 1 of 2 dogsthe effect of SCS was blocked by phentolamine (alpha-adrenergicar.tagcnist, 100-200 ug/kg) but propanolol had no effect. Hypogastricnerve stimulation had little or no effect.
Penile erection is inducec by stimulation of the parasympatheticpelvic nerves, partially due to cholinergic activity but notbeta-adrenergic activity. Inhibitory pathways to the erectile processtravel via the sympathetic chain, and are probably alpha-adrenergic innature. There appears to be no role for the hypogastric nerve inpenile erection in the dog.
rich
J. Reprod. Fertil. 73(2): 567-577 (1985).J. Reprod. Fertil. 73(2): 578-585 (1985).
TABLE 1. The effect of platelet poor or plateletserum on mouse blastocyst outgrowth in-vitro.
(1) O'Neill, C.(2) O'Neill, C.

14 15
ORAL ANTIFERTILITY ACTIVITY OF GOSSYPOL, STEROIDAL ANALOGUESAND CYPROTERONE ACETATE IN MALE RATS
suppl
and
COilPOUI/D 8
·"""Im
HOXJ(J -
Int. J. Androl.
COUPOUIID ACYPROTEROIIE ACETATEGOSSYPOL
I.G. White*, R. Vishwafath*, P.O. Brorn-woodman**,D. RIdley and M.A. Swan
+Departments of Veterinary Physiology*, Organic Chemistry,Histology and Embryology , University of Sydney and
Department of Biological Sciences**, Cumberland CollegeLidcombe, N.S.W.
(1) Prasad, M.R.N., and Diczfalusy, E.2:53-70 (1982).
Each compound was administered orally in 2% carboxyl methylcellulose to six male rats for 62 days at a daily dose of 20mg/kg. Onday 55 two female rats were caged with each male. On day 62 the malerols were weighed and killed and the weights of testes, epididymides,adrenals, ventral prostate and seminal vesicles were recorded. Spermwere flushed from the vas deferens and their morphology, motility andoxygen uptake assessed.
The % fertility and mean litter size ~ S.E. for each group were:Conlrol: 100%, 7.03 + 0.9; cyproterone acetate: 50%, 3.58 ~ 1.85;gossYPol: 0%, 0 ~ 0; -compound A: 100%, 6.7 ~ 1.23; Compound B: 0%, 0+ O. Sperm motility and oxygen uptake were significantly reduced bygossypol and Compound B (p < 0.001), and sperm heads were oftendetached from tails. Rat testes treated with compound B containedabnormally large amounts of lipid and resi¢lual bodies and exhibitedvarious abnormalities such as reduplication of the basement lamina andaberrant nuclear condensation of some spermatids. Few maturespermatozoa were present in the epididymis.
Compound B decreased all the organ weights and the weights of theseminel vesicles, ventral prostate and coagulating glands weredecreased by cyproterone acetate (p < 0.01).
Compound B appears to be as active as gossypol and more activethan cyproterone acetate as an antifertility agent, however, it ispossible that at least some of the activity may be due to othersubstances in the preparation.
Gossypol, a lipid soluble polyphenolic constituent of cottonseed, reduces the fertility of men and male rodents on oral administration (1). However it is not free of side effects and, in the hopeof producing more innocuous male antifertility agents, we havesynlhesized two steroidal analoljues (Compounds A and B) and comparedtheir activity with gossypol and the anti-androgen, cyproterone,lcclate.
TABLE 1. OR after PMSG in Booroola x Corriedale ewe lambsRam Geno- Total Number of Ewes Lambs With: Mean Mean
type Ewe Recent Ovulation Rate Ovul. Live(# ) Lambs Ovul's 1 2 3 >3 Rate Weight
1 F+ 28 9 5 2 2 1. 67 26.42 F+ 36 16 5 9 2 1. 81 25.43 F+ 25 16 10 1 5 1. 69 27.24 FF 35 17 2 9 6 2.24 29.25 FF 39 20 4 10 5 1 2.15 27.46 FF 22 12 1 6 3 2 2.50 31.67 FF 26 19 1 8 8 2 2.68 28.9
# Known genotype (Rams 1,7) ; Genotype for rams 2-6 determinedfrom progeny ORs at 15-16 and 18-19 mo in this experiment.
CSIRO, Division of Animal Production, Armidale; Department ofAgriculture, Adelaide and Kybybolite , South Australia.
*L.R.Piper, B.M.Bindon, S.K.Walker, J.R.W.Walkley and D.Phillips
*
The ovine F gene, which has a major deregulatory effect onovulation rate{OR), has no apparent expression in males. Ram genotype must therefore be assessed by progeny test. Recent data indicate that the ov~rian response to PMSG in prepubertal ewe lambsmay be a useful test criterion (1). This paper compares theresponse to PMSG in ewe lamb progeny of FF and F+ rams andcontrasts these with the natural DRs of the same ewes ag~d 18 mo.
In May 1983, 7 Booroola rams were each joined with 50 Corriedale ewes in south east South Australia. Two rams were of knowngenotype (FF and F+), while the other five were expected frompedigree ~cords~o be either FF or F+. In May 1984, the eweprogeny (aged 6-7 mol were injected (s.c.) with 500 i.u. PMSG. ORwas assessed by laparoscopy{local anaesthetic) 90-96 hr later andewes were weighed. DRs were assessed again in February and May1985 when ewes were aged 15-16 and 18-19 mo. After PMSG injection37% of the 211 ewe lambs had corpora lutea judged older thanconsistent with that injection. These, and the 11% anovular ewes,were discarded from the analysis. The mean live weight, and themean and distribution of ovulation rate for the 52 % of ewe lambswith recent ovulations (< day 4) are given in Table 1.
The OR distributions in Table 1 for rams 1 and 7 are clearlydifferent (X~= 9.2; P<.Ol). Those for rams 2 and 3 do not differfrom that for ram 1, but are different (P<.05) from that for ram7. The distributions for rams 4,5 and 6 do not differ from thatfor ram 7 and approach being significantly different {P<.06,P<.15,P<.06 respectively)from that for ram 1. These groupings of rams(1-3 v 4-7) correspond exactly with K locus genotype based onnatural OR and we conclude that the OR response to PMSG in ewelambs is therefore useful for distinguishing FF from F+ males.
FURTHER OBSERVATIONS ON £ GENE PROGENY TESTING USING THE OVARIANRESPONSE TO PMSG IN PREPUBERTAL EWE LAMBS
(1) Oldham,C.M., Gray,S.J., Poindron,p. and Bindon,B.M. (1984) InReproduction in Sheep {Eds D.R.Lindsay and D.T.Pearce)pp.260-261.

16
FAILURE OF TESTICULAR VEIN LIGATION TO INCREASE FERTILITY INMUJ WITH VARICOCELES
H.W.G. Baker, H.G. Burger, D.M. de Kretser, B. Hudson,G.C. Rennie and W.G.E. Straffon
Reproductive Medicine Clinic and Medical Research Centre,Prince Henry's Hospital and Howard Florey Institute of Experimental
Physiology and Medicine, Melbourne.
Surgery is commonly advised for infertile men with varicoceles(1) yet the value of this treatment has not been established unequivocally (2). Therefore we examined fertility and semen analysis resultsin 651 subfertile men with varicoceles and potentially fertile wivesbefore and after testicular vein ligations performed in 283. Semenwas analysed three times before and at approximately 2 month intervalsafter surgery. Life table estimates of proportions of female partnerspregnant in couples after ligation were similar to those in couplesbefore ligation or in whom no surgery was performed: approximately30% by one year and 45% by 2 years. The pregnancy rates were notsignificantly different in the two groups (Table 1).
TABLE 1. Logrctnk test for pregnancy rates before and after testicularvein ligation.
Operation No. of No. of Expected No. RelativeGroup Patients Pregnancies of Pregnancies Pregnancy
Rate
Before 611 151 146 1.03After 283 109 114 0.96
2 = 0.37 NSX
With these numbers, there was a greater than 95% chance offinding as statistically significant at the 5% level, a 25% increasein pregnancy rate after ligation. Cox regression was performed toallow for the effects of other variables which influence pregnancyrates and timing of the operation. Variables in the regression were inorder of signlficance - duration of infertility, pretreatment spermconcentration, previous fertility, size of left varicocele and spermautoimmunity. Testic~lar vein ligation was not significantly relatedto pregnancy rate (X =0.29). Sperm motility increased with time inboth before (4.4 SEM 1.7%) and after (5.4 SEM 1.3%) ligation groups,but there was no significant difference between this or other semenanalysis results in the two groups. These results suggest that testicular vein ligation is not effective in increasing fertility or semenquality in subfertile men with varicoceles.
(l) Cockett, A. T.K., Takihara, H. ,and Cosentino, J.M. Fertil.Steri1. 41: 5-11 (1984).
(2) Vermeulen, A., and Vanderweghe, M. Fertil. Steril. 42: 249-256,(1984) .
17
CYCLIC CHANGES IN THE SIALIC ACID CONTENT OF CERVICAL MUCUS:A ROLE IN THE NORMAL REGULATION OF SPERM TRANSPORT
Peter Lutjen, JU~~HO~ and Francesca Martinez
Department of Obstetrics and Gynaecology, Monash University,Queen Victoria Medical Centre, Melbourne, Australia.
Secretion of cervical mucus is under oestrogenic influence(l) ,altering the chemical and physical properties of the mucus throughout the menstrual cycle. The molecular basis for these changesand their effects on sperm penetration remains unclear. Recently,an association between decreased levels of mucus sialic acid andinfertility has been reported(2). The dim of thp. present study wasto examine the level of sialic acid in cervic<.ll mucus throughout themenstrual cycle and equate any changes to aJ the hormonal profileand b) the ability of spermatozoa to penetrate the mucus.
Cervical mucus samples were collected by aspiration fromnormally cycling women (n=5) throughout a single menstrual cycle.The content of sialic acid and protein in the cervical mucus wasdetermined by methods previously described(2). Plasma levels ofoestrogen (E2), luteinizing hormone (LH) and progesterone weremeasured by conventional radioimmunoassay. Quantitive assessmentof sperm penetration through the mucus samples has also beendescribed elsewhere~3)The concentration of sialic acid is expressedper mg protein of cervical mucin. Statistical analysis was byPearson's Correlation Test.
TABLE 1. Hormonal profile and mucus sialic acid content duringthe periovulatory and ovulatory phase of the menstrual cycle(n=5). Day 0 = day of LH peak (mean ± SD)
Day of cycle LH IU/L E2 pg/ml Sialic acid ~g/mg mucin
-2 10.4 ± 3.0 252 ± 69.1 140.9 ± 82.6-1 35.3 ± 24.3 319.5 ± 92.1 158.7 ± 121.5
0 100.8 ± 14.0 279.6 ± 141.9 226.2 ± 180.31 30.5 ± 17.8 150.5 ± 102.0 133.3 ± 72.52 15.8 ± 5.6 86.8 ± 13.4 132.5 ± 75.5
The results show a significant positive correlation between thesialic acid content of cervical mucus and the levels of plasmaE2(P=0.007) and LH (P=0.011). The increase in the level of mucussialylation was associated with a marked decrease in the proteincontent of the mucus (P=O.OOl). Mucus sialic acid content wasalso correlated with the degree of sperm penetration through themucus (P=0.026), with maximal sperm penetration concomitant withmaximal sialic acid levels.
We conclude that there is a midcycle surge in cervical mucussialic acid levels under the influence of the endogenous hormonelevels, which appears associated with the increased penetration ofspermatozoa through the mucus at this stage of the menstrual cycle.
(1) Carlborg, L. & Gemzell C. (1969). Acta Endocrinol.62:711-720(2) Lutjen, P.J. et al (1985). Gam Res. (in press)(3) Wang, C. et al (1985). Fertil.Steril. (in press)

18 19
THE REPRODUCTIVE PERFORMANCE AFTER EXPOSURE TO OESTROGENIC PASTURES OFEWES IMMUNIZED AGAINST ANDROSTENEDIONE OR OESTRONE
GONADOTROPHIN RESPONSES TO LHRH IN KOONOONA AND BOOROOLA-KOONOONA FIRSTCROSS RAM LAMBS: INFLUENCE OF LHRH DOSE AND AGE
R.I. Cox,* D.L. Littlet and S.K. Walkert D.O. Kleemann~", M.J. D'Occhiot, S.K. Walker;'" C. Papachristoforout,D.H. Smith* and B.P. Setchell*
Study supported in part by a grant from the J.S. Davies Bequest.
*S.A. Department of Agriculture, Turretfield Research Centre, Rosedale,S.A., 5350 and tDepartment of Animal Sciences, Waite Agricultural
Research Institute, Glen Osmond, S.A., 5064.
LltRH ~ (wk)
(ng/l~g BW) 2 4 6 8 10 12
--------K 10 2.5 4.3 4.5 7.6 9.0 10.8
K 100 6.3 15.3 18.0 21.1 25.8 27.0
BK 10 1.8 2.9 3.6 5.8 6.4 7.2
BK 100 4.5 8.8 13.9 19.5 22.9 20.8
In both K and BK ram lambs, the 100 ng dose of LHRH induced agreater (P<O.Ol) LH response than the 10 ng dose across all periods. Kand BK rams showed increased (P<O. 01) LH responses with age to bothdoses of LHRH. There were no differences in LH responses to ~HRHbetween K and BK rams, either with respect to dose of LHRH or perIod.There was lle acute FSH response to LHRH and mean FSH levels across 2 to12 wk did not di ffer between K and BK rams. These results indicatethat during 2 to 12 wk there is a progressive increase in capacity ofthe pituit~ry in ram lambs to release. LH. T~e. data also suggest thatram lambs of genotypes which differ In prohhcacy can show parallelmaturation of the hypothalamo-pituitary unit during the prepubertal
period.
Ponzoni, R.W., Walker, S.K., Walkley, J.R.W. and Fleet, N.R. In:Genetics of Reproduction in Sheep. R. B. Land and D. W. Robinson(eds.), 127-138 (1984).
(1)
The LH response in ram lambs to exogenous LHRH can be influencedby prevailing patterns of gonadotrophin secretion which, during theprepubertal period, undergo dynamic fluctuations. In the presentstudy, LH re~ponses to varying doses of LHRH were determined from 2 to12 \oIk of 1ge for Koonoona (K) rams and Booroola x Koonoona (BK) ramsheterozygous f~r the Booroola high fecundity (F) gene. BK ewesheterozygous for the F gene show an increased ovulation rate comparedwith K ewes and it was therefore of interest to compare LH responses toLHRH in rams of genotypes differing in proli ficacy (1). At intervalsof 2 wk ram lambs were fitted with jugular vein cannulae and over thefollowing three days received successively 1, 10 and 100 ng LHRH/kg BW(i. v. bolus). Serum LH and FSH levels (ng/ml) were measured using RIAand all hormone data were transformed to log10 before ANOVA. Therewere no consistent LH responses to 1 ng/kg LHRH. Results for LHresponses (~LH, ng/ml) to 10 and 100 ng/kg LHRH are presented as meansin the Table (pooled SEM: genotype 2.1; LHRH dose 1.2; age 1.0).
PASTURE IMMUNITY n O.R. % EWES % EWES FOETUSES/EWEGRAZED MARKED PREGNANT (a)JOINED (b) PREGNANT
Trikkalaa a
0.97(1.03a
)ab
86 1. 37 91 85 (86)* 1.13Yarloop 87 1 37a 97 79ab (92) 0.82(1.06a ) 1.04a
• b69~ (89) 0.90(1.19b ) 1.30bcYarloop A 89 2.22
b90
Yarloop E 91 1.94 86 71 (93) 1.03(1.39c ) 1.46c
Immunity A, to androstenedione; E, to oestrone.*Values in parentheses are for 1984 J01n1ng.Groups with different superscripts are different at the P<0.05 level.
TABLE 1. Reproductive performance in ewes after grazing subterraneanclover based pastures over 2 consecutive seasons.
* CSIRO, Division of Animal Production, Prospect, N.S.W.t Department of Agriculture, Adela~de, S.A.
The results (Table 1) show that oestrus was unaffected and O.R.swere elevated in the immune groups. However there was a higher proportion of dry ewes in the immune-groups which did not OCCur in the previous season following only one year of exposure to Yarloop clover.This data indicates that exposure to a second season of oestrogenicpasture led to lower fertility in steroid immune animals.
(1) Little, D.L., Cox, R.I., Walker, S.K. and Flavel, P.F. In"Reproduction in Sheep" (Eds. D.R. Lindsay and D.T. Pearce)pp. 365-367. Australian Academy of Science (1984).
Immunization of ewes against some steroids can increase theirovulation rate (O.R.) and prolificacy; such animals grazed onoestrogenic pasture for a season and then joined on dried-offpasture when it is no longer oestrogenic, showed normal fertility andincreased prolificacy (1). However, since the effects ofphyto-oestrogens can take several seasons to develop, the reproductiveperformance of the ewes previously studied (1) was re-assessed aftergrazing on oestrogenic pastures for a second season.
Four hundred Merino ewes from an area with oestrogen-free pasturewere randomly allocated to 4 treatment groups: 1, control, maintainedon a low-oestrogen pasture (T. subterraneum cv Trikkala); 2,oestrogenic control, grazed on oestrogenic T. subterraneum cv Yarloop;3, oestrone-immune grazed on Yarloop; 4, androstenedione immune grazedon Yarloop. Groups 3 and 4 were initially immunized in March 1983.The ewes were maintained on Yarloop or Trikkala based pastures fromA.pril 1983 for 2 growing seasons. The immune groups were boos ted onDecember 10th 1984 and all ewes were joined as one flock with 3.2%rams 3 weeks later for 8 weeks. Mating records were taken weeklyduring joining. Initial O.R. was determined by laparoscopy on January15-18th, 1985. The ewes were slaughtered in April and numbers offoetuses present were determined.

20 21
THE EFFECTS OF DIFFERENT PATTERNS OF INSEMINATION, AND OF NUMBERS OFMOTILE SPERM INSEMINATED, ON THE EFFICIENCY OF AID TREATMENT CLOVER INFERTILITY IN EWES: IS THERE A GENETIC COMPONENT TO ITS DEVELOPMENT?
B. Godfrey, C.D. Matthews K.P. Croker, T.J. Johnson and R.J. Lightfoot
SignificAntly different from oat control: * P < 0.05; ** P < 0.01; *** P <0.001.
Although the fertility of the Resistant ewes on Dinninup has beenaffected in comparison to the Control ewes on oats, the effect was less thanthat for the Control ewes on Dinninup. This observation suggests that someresistance to the development of clover infertility may be inherited. A moreintense selection based on both ewes and rams grazed on oestrogenic pasturesmay produce a higher level of resistance. The development of resistance couldbe one of the reasons for the disappearance of the major problems of dystociaand uterine prolapse that occurred on clover pastures in the 1940's.
YEAR OATS DINNINUPCONTROL RESISTANT CONTROL RESISTAN'l'
1979 79 83 67 90
1980 58 77 71 76
1981 93 87 ** 70 *75
1982 90 91 ** 65 80
1983 98 96 *** 69 **75
1984 86 82 *** 32 *60
1985 63 85 * 30 36
Department of Agriculture, South Perth, W.A., 6151
In western Australia there are an average of 18% fewer pregnant ewes incommercial flocks grazed on oestrogenic pastures compared with those onnon-oestrogenic pastures (1,2), and in some situations the depression is muchlarger (3). However, some ewes lamb consistently while grazing oestrogenicpastures. An experiment was conducted to determine whether ewe progenyderived from Merino sheep which had lambed each year during grazing ofpastures containing highly oestrogenic cultivars of subterranean clover weremore resistant to exposure to oestrogenic pasture.
The ewes to generate the experimental animals were obtained from 4experiments which examined reproductive performances on a range ofsubterranean clover cultivars varyir.g in oestrogenicity. Resistant ewes hadlambed each year during 4 to 7 years exposure to highly oestrogenic cultivars;unselected ewes (controls) where chosen at random from the ewes which hadgrazed non-oestrogenic pastures in these experiments. All these ewes weretaken to Badgingarra Research Station where a breeding programme was used toproduce 3/8 Resistant and Control ewes for comparison. From 10 months of age(n=63-.66), during the growing season each year, half of each group of ewesgrazed an oat crop (non-oestrogenic) while the remainder grazed on pastureswhich each September contained more than 70% subterranean clover,C.v. Dinninup (1.01% formononetin, range 0.65-1.54% leaf dry weight basis).The fertility of the ewes to 8 years of age is summarised in Table 1.
Percent ewes pregnant (to ewes present).TABLE 1.
MOTILE SPERM x10~-
CYCLES PREGNANCIES % PREGNANTPER INSEMINATION
<10 113 9 8.010-20 236 32 13.620-30 197 37 18.8 I
I
>30 100 22 22.0 I
INSEMINATION CYCLES PREGNANCIES % PREGNANT2 mornings 151 23 15.2%3 mornings 115 18 15.7%pm/am/pm 380 59 15.5%
Department of Obstetrics and Gynaecologyuniversity of Adelaide
Table 1. pregnancy rates in spontaneously ovular cycles with a clear LHsurge
It was concluded that inseminations 9-15 hours apart did not appearto be more effective than daily treatment. However an increased~regn~ncy rate was observed where higher numbers of motile sperm werelnsemlnated.
The mean number of motile sperm per insemination in these 646 cycles was20.2 ± .4 (sem) million. Higher numbers of motile sperm were inseminatedin conc~ptual cycles (23.0 ± 1.0) than in non-conceptual (19.6 ~ .4),Mann-Whltney test p=.0001. The pregnancy rate per cycle improved withincreasing numbers of motile sperm inseminated, X2 test, p=.015 (Table2 )
Different methods of timing of insemination were used in a programof Artificial Insemination by Donor (AID) using frozen semen. In mostc~ses.timing was based on plasma luteinising hormone (LH) levels, in amlnorlty of treatment cycles on cervical mucus assessment. Until 1980treatrn~nt generally consisted of insemination on 2 successive morningsfollowlng the day of the LH surge, or on 3 successive mornings centredon the day of the LH surge. A study was set up, where 3 inseminationsper cycle were performed, late in the afternoon on the day of the LHsurge, followed by morning and afternoon insemination on the next day.The purpose was to inseminate as close as possible to the time ofovulation, the period of survival of thawed sperm in-vivo beingunknown.
In spontaneously ovular cycles with a definite LH surge nodifference in pregnancy rates between the 2 morning methods of timing,and the study method, was observed. (Table 1)
Table 2. Motile sperm per insemination and pregnancy rate
(1) Wroth, R.H. and Lightfoot, R.J. (1976). Proc. Aust. Soc. Anim. Prod.11: 225-228.
(2) Marshall, T., Beetson, B.R. and Lightfoot, R.J. (1976). Proc. Aust.soc. Anim. Prod. 11: 229-232.
(3) Neil, H.G., Lightfoot, R.J. and Fels, H.E. (1974). Proc. Aust. soc.Anim. Prod. 10: 136.
22 23
THE ASSOCIATION BETWEEN OCCUPATIONAL GROUPAND SPERN CONCENTRATION IN INFERTILE HEN
LUPIN FEEDING AND FERTILITY IN THE MERINO EWE
J. Henderson, H.W.G. Baker and G.C. RennieM.B. Nottle, P.I. Hynd, B.P. Setchell and *R.F. Seamark
Division of Biological and Health Sciences, Deakin University,Geelong, Medical Research Centre, Prince Henry's Hospital andHoward Florey Institute of Experimental Physiology andMedicine, Melbourne.
Department of Animal Sciences, Waite Agricultural Research Institute,The University of Adelaide, Glen Osmond, SA 5064; *Department.ofObstetrics & Gynaecology,The University of Adelaide, Queen El~zabeth
Hospital, Woodville, SA 5011.
TABLE 1. Geometric mean sperm concentration in differentoccupational groups (p< 0.01).
The highest sperm concentrations were in the administrativegroup and the lowest in the farming and transport groups.Men in the farming group also reported significantly greaterexposure to heat and chemicals which may contribute to thelower sperm concentrations. The nine occupational groupswere very broad in coverage and lacked sensitivity. Furtherresearch is needed on the mech~~ism of the association betweenoccupational group and sperm concentration.
In the study of the effects of occupational agents on humanreproduction, relatively few investigations have examinedmale infertility. Involuntary infertility affects about10 - 15% of couples (1). A number of substances have beenshown to have a detrimental effect on male fertility egoradiation, ledd and 1,2-dibromo-3-chloropropane (DBCP) (2).Other sUDstarces having a less potent effect may well havebeen overlooked and the high rate of idiopathic maleinfertility (approximately 75%) may be due in part tooccupational exposures.
The present study used data collected at the ReproductiveMedicine Clinic, Prince Henry's Hospital to assess whetheroccupation was associated with results of semen analysis.Occupations were classified into one of nine groups basedon the Yearbook of Australia. A significant association wasfound between sperm concentration and occupational group(Table 1).
1) Oldham, C.M. PhD thesis The Univ. of West. Aust. (1~80). .2) Remsley, J.A., Reis, P.J. and Downes A.M. Aust. J. b~ol. Sc~.
26: 961-72 (1973).3) Hu;';, I.D., Aust. J. Agric. Res. 12: 155-165 (1974).
No. ewes ovulating Ovulation rateTreatment n 0 1 2 (Mean ± SEM)Control 40 2 33 5 1.08 ± 0.07 a
RCOR casein 39 0 21 IS 1.46 ± O.OSbStarch 40 4 29 7 LOS ± 0.08~
COR . / t h 40 2 20 18 1.40 ± 0.09R case~n s arc 40 1 19 20 1.4S + 0.09bLupin )Means with different superscripts differ significantly (P<0.05 •
The results demonstrate that lupin grain and iso-nitrogenoussupplements of HCOH casein initiate similar i~crea~es in OR. Theprovision of extra ME did not increase OR. L~vewe~ght was unchanged bysupp lementat ion.
In conclusion, increases in OR in response t~ short-term lupinfeeding can be explained by increases in post-rum~nal protein supply.
Supplementation with lupin grain six days prior to ovulati~n hasbeen shown to increase fertility in Merino ewes by 30%(1). It ~sunclear whether this increase in ovulatio~ rate (OR) ~s r:lated to asignificant amount of lupin protein escap~ng ~egradat~on ~n the :umenor to increases in metabolisable energy (ME) ~ntake. The f~llow~ng.
experiment was designed to define the relative roles of lup~n prote~n
and ME. . f200 six year old Merino ewes (44:S~0.4 ~g) w:re a~s~gned to one 0
five treatments on the basis of strat~f~ed l~vewe~ght ~.e. 1) control;2) protein: 160 g/ewe/d of formaldehyde treated (HCOH) casein (2); 3)energy: 400 g/ewe/d of wheat starch; 4) protein plus energy: 160 g ofRCOR casein plus 200 g wheat starch/ewe/d and 5) lupi~ 'control': ~OO gof cracked lupin grain/ewe/d. All ewes received a ma~ntenance rat~on
of wheaten hay, 30% of which was pelleted to incorporate the .supplements. Each group gra~ed 0.5 ha of barley stubble. Prote~n
supplements were formulated to provide similar amounts of pro~e~n
post-ruminally (2,3). Supplements of wheat starch, HCOR case~n plusstarch and lupin grain were of equal ME value. Supplemen~s were fedfor 14 days beginning on day 4 of th: oestrous cycle. ~n~mals wereweighed at 14 day intervals. Ovulat~on rate was determ~ned byendoscopy.Table 1 The ovulation rate of ewes fed either the control diet or thecontrol diet supplemented with either HCOR casein, starch, RCOR caseinplus starch or lupin grain.
4.689.554.794.901. 552.242.693.555.13
SPERM CONCENT~\TION (xl0 6/ml)n
422184143115
93116487
9927
OCCUPATIONAL GROUP
ProfessionalAdministrativeClericalSalesFarmingTransportCraftsmenServiceNot employed
(1) Rachootin, P. and Olsen, J. JOM. 25:394-402 (1983).(2) Steeno, a.p. and Pangkahila, A. Andrologia ~:5-22 (1984).

•24 25
USE OF THE GLUCOSE TEST TO IMPROVE BREEDING FLOCK PRODUCTIVITY
R.A. Parr, M.A. Miles and P.J. LangdonAnimal Research Institute, Department of Agriculture
and Rural Affairs, Werribee, Victoria 3030.Centre for Early Human Development, Monash University,
Queen Victoria Medical Centre,Lonsdale Street, Melbourne.
EFFECT OF GROWTH FACTORS ON THE RATE OF MOUSE EMBRYODEVELOPMENT IN-VITRO
C.M. Caro and A. Trounson
Preimplantation mouse and human embryos can develop normally in vitro inprotein-free medium (l,2) but it is known that the clearage rate of mouse emb:y~s
from the 2-cell to blastocyst stages in-vivo is approximately 24 hrs faster than it isin-vitro. In this study we investigated serum replacements in Whittingham's :6medium and their effect on the rate of development of 2-cell mouse embryos 10
vitro. The medium additives studied were Insulin (Collaborative Research Inc.,USA) at a concentration of 10Ilg/ml, and a synthetic serum replacement call~d NuSerum (Collaborative Research Inc.). Nu-Serum is a consistent formulatlOn ofhormones and growth factors. Results (Table 1) show cleavage rates of embr~os
grown in protein-free media (Contro!), media supplemented with 4 m?/ml BovmeSerum Albumin (BSA) plus 10 Ilg/ml Insulin and media supplemented With 10% NuSerum. Two hundred embryos were cultured in each of the Control and twotreatment groups and the stages of development were progressively assessed at 24,48 and 72 hours in vitro.
TABLE 1. The percentage of embryos developing from 2-cells at 24 hr intervals invitro.
2.63.33.5
EWEPRODUCTIVITY+(kg)
LOW -0.4 25/76(33)MED -0.2 23/93(25)HIGH +0.1 22/94(23)
L
GLUCOSE NUTRITION MEAN CONDITION LAMBGROUP GROUP SCORE CHANGE MORTALITY*(%)
Lamb deaths in the peri-natal period are a major problem in manyAustralian sheep flocks. The 'Glucose Test' has recently beenproposed as a means of reducing this loss through strategic feedingand management during late pregnancy (1). The aim of this study wasto examine the influence of nutrition during late pregnancy on lambsurvival and productivity from ewes with low or high blood glucoseconcentrations at mid-pregnancy.
A flock of 400 B.L.xMer. ewes was divided into Low (L) and High(H) glucose groups 87-93 days after a synchronized joining accordingto methods previously described (2). They were then allotted to low,medium or high nutrition groups (mean ME - 8.5, 11.2 and 14.0MJ/kgDM respectively), and were fed accordingly until three weeks postlambing. Lambing was closely supervised and all dead lambs underwentpost-mortem examination.TABLE 1. Change in ewe condition score during treatment period,lamb mortality and productivity from ewes joined.
Culture time (embryo stage) Control BSA + Insulin Nu-Serum N
24 hrs ( % Compacted Morula) 4 18 30 20048 hrs ( % Blastocyst) 21 31 40 20072 hrs { % Expanded Blastocyst 75 82 92 200
Both media supplements significantly increased the rate of embryodevelopment at 24 and 48 hrs. (Insulin, p<O.OOOl at 24 hrs, p<0.05 at ~8 ~~s. NuSerum p<O.OOOl at 24 hrs, p<O.OOOl at 48 hrs). At 72 hrs. there was a slgnlfl~antly
higher proportion of embryos reaching the expanded blastocyst stage only 10 thegroup with Nu-Serum supplement (p<0.005).
Nu-Serum appears to be an effective media supplement for acceleratingembryo development in-vitro. Nu-Serum may be beneficial in the human IVFsystem as a replacement for human or bovine serum with the added ad~antages ofuniformity between batches, stringent quality control and a protem contentapproximately 30% of that in fetal calf serum.
(l) Caro, C.M., and Trounson, A. J. Vitro Fert. Embryo Transfer .!.: 183-187 (1984).(2) Caro, C.M., and Trounson, A. Proc. Aust. Soc. Rep. BioI. .!.§.: 106 (1984).

26 27
A TRANSIENT ELEVATION-OF PROLACTIN SECRETION FOLLOWINGHYPOTIIALAMO - PITUITARY DISCONNECTION IN THE EWE.
(1) Bryant, G.D., Greenwood, F.C., Kann, G., Martinet, J. and Denamur R.J. Endocr. 51:405-406 (1971).
(2) Clarke, I.J.:-Cummins, J.T. and de Kretser, D.M. Neuroendocrinology36:376-384 (1983).
G.B. Thomas, J.T. Cummins+, Lynda Cavanagh and I.J. Clarke.
Medical Research Centre, Prince Henry's Hospital and +Departmentof Neurosurgery, St Vincent's Hospital, Melbourne, Victoria.
21.0 84.3 ± 32.72.6 67.3 ± 28.2
15.3b ,c 44.0 ± 3.525.1c 26.6 ± 3.720.4 59.2 ± 13.916.6c 32.2 ± 5.620.3 60.0 ± 37.130.4 43.6 ± 13.519.6 51.0 ± 24.621.3 62.1 ± 40.214.4 24.1 ± 9.8
b: P < 0.05 compared toc: P < 0.05 compared to SHAM-HPD.
The transient increase in prolactin secretion following HPD contraststo the p~olonged elevation noted in other species after stalk-section.We conclude that, in the ewe, the hypothalamus does exert a tonicinhibitory influence but that the pituitary possesses an autonomouscapacity to secrete prolactin in the absence of putative hypothalamic-releasing-factors.
a: Mean of 10 min x 2h blood sampling.pre-surgical levels in the same sheep.
Early studies showed that pituitary stalk-section in ewes resultedin a decreased level of prolactin (1) due to infarction of the pituitarygland. However, if the pituitary gland was isolated from the hypothalamuswithout vascular damage the level of prolactin on the day after surgerywaS increased (2). Our aim was to define the long-term changes inprolactin secretion in the ewe after hypothalamo-pituitarydisconnection (HPD).
Prior to the experiment eight ewes were housed and handled over 4months (Oct - Jan). On day 0, 4 ewes underwent HPD (2) and 4 ewes weresubjected to a sham surgical procedure (SHAM-HPD). Blood was collected at10 minute intervals for 2 hours on dRyS -50, -1, +1, +7 and +43 viaindwelling jugular cannulae. Single blood samples were also co11ecteg ondays 2, 3, 14, 21, 28 and 35. Plasma was harvested and stored at -20 Cfor prolactin RIA.
Plasma prolactin levels increased 2-fo1d the day after HPD (table 1;P < 0.05) but then slowly decreased to values similar to those seen beforesurgery. Prior to HPD the level of prolactin was not different betweenthe groups, but on days 1, 2 and 7 following HPD the level of prolactinwas higher (p < 0.05) in HPD ewes than in SHAM-HPD ewes.
TABLE 1. Effect of HPD or SHAM-HPD on plasma prolactin (ng/ml:mean±SEM).
DAY OF EXPERIMENT HPD SHAM-HPD
_50a 86.3 ±l a 58.6 ±
+ l a 127.8 ±2 108.3 ±3 93.3 ±7a 114.4 ±
14 100.7 ±21 103.3 ±28 84.1 ±35 91.6 ±43a 67.9 +
82.5(12.7)
0.1 67.3(35.0)
0.3 32.07.1 10.0
Oestradiol/h 1-/';'iT-,-~rr--....,.-~~-+--n':""'--"---!.~-"""---"""'--
LONG-TERM OESTROGEN-PROGESTAGEN TREATMErn DOES NOT INCREASE THEINHIBITORY EFFECTS OF OESTRADIOL ON GONADOTROPHIN SECRETION IN
OVARIECTOMISED EWES
On D7 plasma LH and FSH concentrations were lower P < 0.05 in OSewes (3.7 ± 0.4 ng/ml, 28.3 ± 4.2 ng/ml) than in OV and AV ewes (5.8 ±0.6 ng/ml, 41.3 ± 4.2 ng/ml). Treatment (AV, OV, OS) had asignificant (P < 0.05) effect on the changes in secretion of both LHand FS~ (T~bl e 1). Oestradiol infusion significantly reduced FSHsecret10n 1n each treatment group, but LH secretion only in Group AV(the anovulatory season).
The results show that 1) long-term treatment of ewes with E and Pdoes not increase E negative feedback on gonadotrophin secretion and2) during the ovulatory season E has a greater inhibitory effect onFSH than on LH secretion. The increased E negative feedback inacyclic post-partum ewes is unlikely to be due to the effects ofpregnancy plasma concentrations of E and P, but could reflect theneuroe~docrine effects of suckling, lactation and nutritional loss.1. Wnght, P.J., Geytenbeek, P.E., Clarke, I.J. and Findlay, .J.K.
J. Reprod. Fert. 61, 97-102 (1981).
Z.C. Zhang, P.J. Wright, J.A. Burden and I.J. Clarke+Department of Veterinary Clinical Sciences, Iiniversity of Melbourne,Werribee 3030; +Medical Research Centre, Prince Henry's Hospital,
Melbourne 3003, Victoria.
The inhibitory effect of oestrogen (E) on LH secretion (E negativefeedback) is greater in acyclic post-partum ewes than in cyclicewes(l). We have tested the hypothesis that this increased inhibitoryeffect of E is due to the long-term exposure during pregnancy of thehypothalamo-pituitary axis to progesterone (P) and E.
During the ovulatory season Corriedale ewes (n = 12) were treatedwith oestradiol-17~ (140 ~g) and P (60 mg) administered in vehicle(arachis oil) i.m. daily for 5 weeks (Group OS). Two control groupseach of 12 ewes were similarly treated with vehicle during theovulatory (Group OV) or the anovulatory (Group AV) seasons. At theend of this treatment the ewes were ovariectomised (DO) and on 08 to014 received i.v. continuous infusions of oestradiol-17a (0, 0.1, O.~
~g/h, 4 ewes/group).On 07 and 014 blood samples were taken for LH and FSH determin
ation each 20 min for 6 h. Changes in LH and FSH secretion for eachewe were expressed as the mean hormone concentration in plasma samplestaken on 014 as a %of mean values for samples taken on 07.
TABLE 1. Changes (0/" mean (s.e.m.)) in plasma LH and FSHconcentrations in ovariectomised ewes after oestradiol infusion for 7da s

as
28
OESTROGEN AND PROGESTERONE STOP GnRH PULSES INOVARIECTOMIZED ANESTROUS EWES
I.J. Clarke, F.J. Karsch and J.T. Cummins+
Medical Research Centre, Prince Henry's Hospital and+DepartmentNeurosurgery, St. Vincent's Hospital, Melbourne, Vic.
Previous studies have shown that the negative feedback effects ofoestrogen (E) and progesterone (P) on luteinizing hormone (LH) secretionin sheep are enhanced during anoestrus (1,2). Plasma levels of E and Pthat allow the secretion of LH during the mating season completelyblock LH secretion during the anoestrous season (ibid). Our aim was todetermine wh0ther or not the negative feedback effect of E or P thatis exerted d1.lring anestrus, is due to an effect on GnRH secreti~n.
Anestrous Cnrriedale ewes were ovariectomized (ovx) on Day 0 andreceived either nJ treatment (n=4), a single subcutaneous (s.c.) Eimplant (n=5)(1) or 4 s.c. 'packets· of P (n=5) (2). On Day 8 theplasma P levels in the P treated ewes were 6.2 ± 1.1 ng/ml (s.e.m.).Six days after the commencement of treatment jugular bloed was collectedat 10 min intervals for 6h. On the seventh day, ewes were anaesthetizedand prepared for the collection of hypophyseal portal bloed (3), whichwas collected over 4 (n=l) to 6 (n=13) h on days 8,9 or 10. On thesame day, jugular venous bloed samples were collected at 10 minintervals for 2h, before the portal bloed vessels were punctured.Portal samples were collected over 10 min intervals and jugular sampleswere taken at the end of each 10 min collection peried. After 6h ofportal sampling the sheep were given a 250 ng i.v. bolus of GnRH andjugular samples were collected at 5 min intervals for 20 min. Jugularplasma samples were assayed for LH and portal plasma for GnRH. LH andGnRH inter-pulse intervals (IPI) were calculated as the differencebetween successive peaks in the plasma profiles.
In control sheep the LH pulse IPI was similar to Day 6 (84 ± 6 min;s.e.m.) and on the day of portal sampling (88 ± 13). None of the E orP treated sheep showed LH pulses on either D6 or D8-10. In controls,each LH pulse was concomitant with a GnRH pulse. No GnRH or LHpulses were detected in the E and P treated ewes. LH responses to the250 ng challenge of GnRH were lower in the E (2.6 ± 0.9 ng/ml; P <0.01)and P (3.1 ± 1.1 ng/ml; P < 0.05) treated ewes than in the controls(8.0 ± 1.2 ng/ml).
Thes~ data show that E and P block the secretion of GnRH in ovx:,wes dur~ng anoestrus. Reduced LH responsiveness to a GnRH challenge~n stero~d treated ewes may be due to prolonged GnRH deprivation aswell as dir~ct pitUitary effects of the steroids.
(1) Legan, S.J., Karsch, F.J. & Foster, D.L. Endocrinology 101818 (1977). ---'
(2) Karsch, F.J., Legan, S.J., Hauger, R.L. & Foster, D.L.Endocrinology 101,800 (1977).
(3) Clarke, LJ. &CUmmins, ,T.T. Endocrinology 111,1737 (1982).
£
29
EFFECT OF ADDITION AND REMOVAL OF GLYCEROL AND METHOD OF TRANSFER ONVIABILITY OF SHEEP EMBRYOS.
J.R. HUNTON, W.M.C. MAXWELL and J.P. RYAN.
Animal Breeding and Research Institute, Katanning, W.A.
Frozen storage of mammalian embryos requires the addition of a cryoprotective agent prior to freezing and its removal upon thawing. A stepwise dilution procedure is generally used to remove the cryoprotectantfollowing thaWing of frozen sheep embryos whereas a single dilution in asucrose gradient has proved effective for cattle embryos. The developmentof methods for intrauterine insemination of sheep by laparoscopy hasprovided the possibility of using laparoscopy for transfer of uterinestage sheep embryos.
In the present study, Day 6 embryos (Day 0= day of oestrus) collectedfrom donor Merino ewes were randomly assigned to one of four treatmentgroups: a) No treatment [held in Dulbecco phosphate buffer containing 20%foetal calf serum (DBS)], b) addition of glycerol (cryoprotectant) byplacing sequentially for 10 minutes each in DBS containing 0, 0.35,0.70,1.05 and 1.40M glycerol, followed by removal of glycerol by reversing thestep-wise procedure, c) addition of glycero~ as in ~reatment (b) ~o~lowed
by removal of glycerol by placing embr~os d~~ect~y ~nto DBS co~t~~n~ng
0.5M sucrose for 10 minutes then wash~ng tw~ce ~n DBS, d) add~t~on ?fglycerol as in treatment (b) followed by removal of glycer?l by plac~ng
embryos directly into DBS containing 1.0M sucrose for 10 m~nutes thenwashing twice in DBS. .
Following treatment, the embryos were transferred to the ~ter~~e hornipsilateral to corpora lutea of recipient Merino ewes syr.chron~sed.~n
oestrus with the donors. The embryos were deposited either follow~ng
exposure of the reproductive tract by laparotomy, or via a.l~ngt~ of polyethylene tubing (0.5mm i.d, 0.8mm o.d.) passed through a r~g~d p~pette
into the uterine lumen with the aid of a laparoscope. The ewes wereslaughtered 18 to 100 days after transfer and the number of foetusesrecorded.
TABLE 1. Proportion of embryos developing in relation to method of transfer and method of glycerol removal.
Method Glycerol removalof
transfer a) no treat. b) gly.dil. c) 0.5N suc. d) l.O~l sue.
Laparotomy 20/33 (17) 16/27 (14) 12/14 (7) 11/22 (12)Laparoscopy 5/17 (8) 15/29 (15) 5/12 (6) 7/18 (9)
( ) number of ewes
The addition of glycerol and its removal by step-wise dilution orsucrose gradients is not in itself detrimental to the. viability ~f day 6sheep embryos. The laparoscopic transfer method requ~res less t~me toperform than laparotomy, is less traumatic to the. ewe ~nd may re~uce theincidence of post-operative adhesions. However, ln thlS study hlgherpregnancy rates were obtained when embryos were transferred by laparotomy.
Supported by the Australian Meat Research Committee.

30
EFFECT OF TYPE OF STORAGE VESSEL AND METHOD OF GLYCEROL ~EMOVAL ONVIABILITY OF FROZEN-THAWED SHEEP EMBRYOS.
J.P. Ryan, W.M.C. Maxwell and J.R. Hunton.
Animal Breeding and Research Jnstitute, Katanning, W.A.
Glass ampoules have been the main vessel used for the frozen storageof sheep embryos, and step-wise dilution the means for cryoprotectantremoval from the embryos upon thawing (1). Evidence from work with cattleembryos suggests that plastic straws are suitable storage vessels, andsucrose gradients may be used to remove cryoprotectants prior to transfer(2).
ln the present study, Day 6 embryos (Day O=day of oestrus) werecoll:cted from ~onor Merino ewes and cryoprotectant (glycerol) added byplac~ng sequent~ally for 10 minutec each in Dulbecco phosphate buffercontaining 20% foetal calf serum (DDS) plus 0, 0.35, 0.70, 1.05 and 1.40M glycerol. Embryos were loaded into 1.2ml glass ampoules or 0.25mlplastic straws and cooled at -l DC/min to -6 DC where ice-crystallisationwas induced. They were then cooled at -O.3 DC/min to -30 DC, held at thattemperature for 10 minutes, and plunged into liquid nitrogen (-196 DC).
The ampoules and stra~s were thawed by holding in a water bath at37 DC. Following thawing, the glycerol was removed by either (a) dilutionby a reve~s~l of the step-wise addition procedure, (b) placing embryos inDBS c~n~a~n~ng p.5H sucrose for 10 minutes, or (c) placing embryos in DBSconta~n~ng 1.OM sucrose for 10 minutes. The embryos were transferred atlaparotomy to the uterus of recipient Merino ewes on Day 6 of an inducedoestrous cycle. The recipients were slaughtered 18-100 days after transfer and the number of foetuses recorded.
Table 1. Proportion of frozen-thawed embryos developing in relation totype of storage vessel and method of glycerol removal.
Type of Glycerol removalstoragevessel (a) gly. dil. (b) 0.5M suc. (c) 1.0M suc. Total
ampoule 25/45 (21) 12/39 (20) 5/42 (19) 42/126 (60)straw 26/46 (21) 17/41 (18) 23/48 (18) 66/135 (57)
Total 51/91 (42) 29/80 (38) 28/90 (37) 108/261(117)
number of ewes
The met~od of glycerol removal had no effect on the proportion of embryosdevelop~ng after frozen storage in straws. However, when ampoules werethe st~rage.ves7el, more emb~yos developed when glycerol was removed bystep-w~se d~lut~on than by e~ther sucrose gradient. This suggests thatthe thawing rate may influence the method of glycerol removal chosen foroptimum embryo survival.
(1) Bilton, R.J., and Moore, N.W. Proc. Aust. Reprod. BioI. (1983).(2) Leibo, S.P. Theriogenology 11: 767-790 (1984).
Supported by Australian Meat Research Committee.
•31
TEMPERATURE REQUIREMENTS FOR SUCROSE DILUTION OF GLYCEROL FROMFROZEN-THAWED MOUSE EMBRYOS
A. Szell and J.N.Shelton
Department of Immunology, The John Curtin School of Medical Research,Australian National University, Canberra, A.C.T.
The aim of the present work was to find optimal conditons for sucrosedilution of glycerol from frozen-thawed day 3 mouse embryos. In Exp 1the temperature during sucrose dilution (20 or 36°C) was studiedindependently fr0ffi the temperature of the thawing bath (20 or 36°C) atfour levels of sucrose concentration (0.25, 0.5, 1.0 or 1.5 M). InExp 2 the effect of the time of mixing the freezing medium with thediluent (immediately before freezing or after thawing) wasinvestigated at three levels of sucrose concentration (0.5, 1.0 or 1.5M).
The embryos were loaded into 0.5 ml straws which contained 40 ul offreezing medium (3.0 M glycerol+0.25 M sucrose) separated by an airbubble from 440 ul sucrose diluent. Following 10 min equilibration inthe freezing medium the embryos were frozen by placing the straw intoliquid nitr0gen vapour for 7 to 10 min and then stored in liquidnitrogen. The freezing medium and the diluent were mixed eitherimmediately before freezing (Exp 1 and 2) or after thawing (Exp 2).The straws were thawed in a water bath without agitation. To allowglycerol removal the embryos were left for 10 min at controlledtemperature (Exp 1, 20 or 36°C; Exp 2, 20°C) before being washed inphosphate buffered saline supplemented with 5% foetal calf serum andthen cultured for 2 days in Whitten's medium. The number of embryosdeveloping to blastocyst stage was expressed as the proportion ofthose frozen.
In Exp 1 the thawing temperature did not affect the survival rate(20°C 15% 48/320, 36°C 17.2% 55/320; F<l). The viability was markedlyhigher after removing the glycerol at 20°C (25.3% 81/320) than at 36°C(6.9% 22/320; P<O.OOl). The best survival rate (51% 41/80) wasobtained when the diluent contained 1.0 M sucrose and the glycerol wasremoved at 20°C.
In Exp 2 mixing the diluent and the freezing medium after thawingresulted in substantially higher viability (65% 117/180) than mixingbefore freezing (37% 67/180; P<O.OOl). The effect of sucroseconcentration was significant when the mixing was done before freezing(P<O.OOl), but not when it was done after thawing (P>O.l). Thiscaused an interaction between the two factors (P<0.05). The highestsurvival rate resulted (73% 44/60) when the freezing medium was mixedwith 1.0 M sucrose solution after thawing.
The results show that the temperature should be strictly controlledduring sucrose dilution and suggest that both extracellular andintracellular concentrations of glycerol are important in thecryoprotection of embryos.
M

• Ii
3233
VISUALIZATION OF PRONUCLEI AND CULTURE OF ONE-CELL EMBRYOS OF SHEEP
S.K. Walker, P. Quinn, R.J. Ashman and D.H. Smith*
ELECTRON }1ICROSCOPIC HISTOCHEMISTRY OF THE MATRIX OF THE MOUSE OOCYTECUMULUS COMPLEX
Department of Obstetrics and Gynaecology, University of Adelaide, TheQueen Elizabeth Hospital, Woodville, S.A. 5011 and *Turretfield ResearchCentre, Department of Agriculture, Rosedale, S.A.
R.1.' Webb and J .M. Cummins
Reproductive Biology Croup, Department of Veterinary Anatomy,University of Queensland, St Lucia 4067.
Supported by a grant from Queensland Medical Laboratories.
(1) Thomopolous, C.N., Schulte, B.A. & Spicer, 5.S. (983)Histochemistry ~: 417-431.
(2) Sannes, P.L., Katsuyama, T. & Spicer, S.S. (1978) J. Histochem.Cytochem. ~: 55-61
(3) Talbot, P. (1984) J. expo Zoo1. 229: 309-316.
An electron microscopic histochemical study of the complex carbohydrates of the matrix of the oocyte-cumulus complex (OOC) was undertakento study changes during pre- and post-ovulatory maturation. Mice (C57 xCBA hybrids) were primed with 7iu PMSC and thEm induced t~ ovul~te with7iu HCG given 48 hours later. OOC's were r.ecovercd at varl~us tl~es
before and after ovulation, after killing the mice by cervlcal dlslocation. Specimens were fixed in 2% glutaraldehyde: 4% paraformaldehydein O.lM cacodylate buffer (pH 7.4, 290 mOsmol). A variety of .embeddi~g
media were evaluated: Araldite; Epon; Lowicryl K4M and LR Whlte. Tlnnsections were stained with dialyzed iron or with tannic acid/uranyl acetate. Dialyzed iron is specific for carboxyl and/or sulphate groups ofacidic glycosaminoglycans and glycoconjugatesl1); tannic acid/uranylacetate, is specific for hydroxyl groups of glycoprotein~ (2). .
Pre- and post-ovulatory changes in the OOC result In the productlonand expansion of an extracellular matrix that with tannic acid/uranylacetate staining appears as a network of electron dense granules andfilaments. A positive staining reaction with dialyzed iron confirms forthe first time the presence of acidic glycoconjugates and glycosaminoglycans in the matrix: previous studies using ruthenium red pro~cs~ing
as well as enzymic digestion and biochemical means have merely lndlcatedthe presence of complex carbohydrates such as hyaluronic acid.(3~.Both ,staining procedures indicate that the absolute amount o~ matrlx ~ncreases
as the OOC matures, although shrinkage artifacts - partlcularly In thepost-ovulatory OOC - make quantitation difficult. The carbohydrate .components of the matrix are concentrated primarily in the corona radlata region, and are distributed throughout the rest of the cumulusmatrix in coarse strands.
Of the embedding media assessed so far, Lowicryl K4M and LR Whitegive far superior results to Araldite or Epon. An additional observation in this study was the detection of large numbers of electron lucentvacuoles in the cumulus cells and in the matrix: these were particularlyprevalent during the pre-ovulatory phase of OOC maturation and , as theydid not stain with any of the reagents used here, may represent a noncarbohydrate component of the matrix. The nature of these vacuoles isunder investigation.
Time after median time of ovulationStudy 1 Study 2
2-4h 5-7h 8-10h 14-16h 17-19h 20~22h
The visibility of pronuclei in sheep ova is a major impediment togene microinjection (1). Because pronuclei move within the cell, theremay be a specific time after fertilization when their visibility wouldbe enhanced. This stUdy examined the relationships between time afterOVUlation, pronuclei visibility and the subsequent development of onecell sheep embryos in culture.
Ewes were inseminated with 20xl06 motile sperm/horn 24h afterprogestagen sponge (Sp) removal. Ovulation was synchronized by i.v.administration of GnRH (lOOug gonadorelin/ewe). The median time toovulation in PMSG (Livestock Laboratories pty Ltd, Melbourne) and FSH-P(Burns Biotec, Omaha, USA)- treated ewes was 48h and 49h respectivelyafter Sp removal with 74.2% and 82.3% of all ovulations occurringbetween 47-50h. Single cell embryos were collected between 2-10h(StUdy 1, PMSG treatment) and 14-22h (Study 2, FSH-P treatment) afterthe median time of ovulation and examined by Nomarski optics (x125magnification) (1) immediately after collection, (2) after centrifugationat 70~for 90 seconds if pronuclei were not observed and (3) afterculture in bicarbonate-buffered synthetic oviduct fluid medium for up to10 days.
Results are presented below.
(1) Ward, K.A., Murray, J.D., Nancarrow, C.D., Boland, M.P. and Sutton,R. (1984). In: Reproduction in Sheep. Eds D.R. Lindsay and D.T.Pearce. Australian Academy of Science, Canberra. p279-285.
No. ova 37 33 45 30 29 38Fertile ova (%) 32.4 51.5 70.0 89.3 86.2 88.9% fertile ova** 9.1 23.5 3.6 57.7 72.0 23.5with visible pronuclei (18.2) (58.8) (21.4) (69.2) (76.0) (47.1)No. cleaVing ova 7/11 12/13 19/26 11/23 12/21 8/16No. blastocysts 0/7 0/12 0/19 0/11 1/12 2/8
* includes 45.2% 2-4 cell embryos** centrifuged ova in brackets.Visualization of pronuclei in the 14-16h and 17-19h periods was
SUbstantially higher than values from previous studies (i). These periodsapproximately correspond with the 6h before first cell division asestimated by the appearance of 2-cell embryos in uterine flushings.Centrifugation improved the Visibility of pronuclei partiCUlarly inStudy 1. However, collection between 2-10h after ovulation is not justified as many cells were not fertilized. Techniques for the culture ofsingle-cell embryos may need to be re-assessed given that 37.3% of thefertilized cells failed to divide.

35
Whatawhata Hill Country Research Station and Ruakura Agricultural ResearchCentre, Ministry of Agriculture and Fisheries, Hamilton, New Zealand
THE EFFECT OF MELATONIN, PROGESTERONE AND f'MSG ON THE OVULATORY ANDMATING RESPONSE OF ROMNEY EWE LAMBS TO SEXUALLY ACTIVE RAMS
R.W. Moore, P.R. Lynch, C.M. Miller, R.A.S. Welch and P. Haszard
64 0 0 18 27.98 8 0 0 26.1o 0 0 0 27.4
111212
Ram+PMSGRamNo Ram
TreatmentMelatonin
Ram+PMSG 13 8 0 ~8 0 27.SNo Prog. Ram 13 0 0 0 0 28.8
No Ram 13 0 0 0 0 27.8
No MelatoninRam+PMSG 14 64 7 21 14 29.1
Prog. Ram 14 7 7 7 7 28.5No Ram 13 0 0 0 14 28.6
Ram+PMSG 12 0 0 0 0 26.1No Prog. Ram 12 0 0 0 0 28.3
No Ram 12 0 0 0 0 29.8
Prog.
Previous experiments have shown that melatonin administered subcutaneously(1) and intravaginally (2) advanced the onset of oestrus in ewe lambs. In thepresent experiment the effect of melatonin, progesterone and PMSG on Uie ovulatoryand mating response of Romney ewe lambs to the introduction of sexually active ramsbefore the normal breeding season was examined. Rams run with oestrCJus e,·;es weremore effective in stimulating ovulations in c":Jo.-;·strou', ewes than rams that have notrecently been sexually active (3).
The experiment was a 2x2x3 factorial with 2 melatonin levels (1 implantcontaining 435 mg melatonin inserted subcutaneously on January 18. 2. no impl~nt) i
2 progesterone levels (l.intravaginal CTDR (4) containing 328 mg pro~ester~ne l.nsertedfor 12 days beginning March 20. 2. no CIDR) i and 3 levels of ewe stl.mulatl.on(1. sexually active ram plus 400 i.u. PMSG on April 1. 2. sexually active ram only3. no rams no PMSG). Mating marks and ovulations (by laparoscopy) were recordedon April 4 and pregnancy by ultra-sonic scanning in late May.
Table 1. Mating and ovulatory activity following treatment, and liveweights April 1.
n %Mated & 9ciMated & %Ovulated& %Pregnant Live weightovulated not ovulated not mated April 1 (kg)
April 1 live weights were too low to expect a high incidew'p, May-June oestrc. !sactivity. With progesterone pre-treatment rams plus PMSG induced 64% of ewes tomate and ovulate irrespective of melatonin treatment. Without progesteronepre-treatment the rams plus PMSG induced 8% of ewes to mate and ovulate and a further38% to ovulate without mating in the melatonin implanted group. In contrast therewas no mating or ovulations in similarly treated ewes without melatonin implants.Melatonin probably induced ovulations before April 1 which gave these ewes someprogesterone pre-treatment.
(1) Moore R.W., Miller C.M., Lynch P.R., Welch R.A.S., Barnes D.R. and Hockey H.U.P.Proc. NZ. Soc. Anim. Prod 44:21-23 (1985)
(2) Nowak Rachel and Rodway, R~. J.Steroid Biochemistry 20:146 (1984)(3) Knioht T.W. Proe. NZ Soc. Anim. Prod. 45 (In press). (1985)(4) wel;h R.A.S., Andrewes W.D., Barnes D.R. Bremner K., and Harvey T.G.
Proc. Tenth Int. Congress on Anim. Reprod. and A.I.l:354 (1984)
EFFECT OF LONG DAYLENGTH ON REPRODUCTION IN JAVANESE THIN-TAIL EWES
Balai Penelitian Ternak, P.O. Box 123, Bogor, Indonesia
34
I.C. Fletcher and I. Gede Putu
Javanese thin-tail (JTT) ewes in their near-equatorial environmentare capable of breeding at any time of year with average inter lambingintervals of only 7 months (I). Their reproductive response to changesin daylength was investigated comparatively with that of BorderLeicester x Merino (BLM) ewes as a preliminary appraisal of the possible contributio~ of the JTT genotype to increased lambing frequency intemperate reg ior,s.
Five mature JTT ewes taken at random from a group of 16 and 5 BLMewes which had been imported from southern Australia some 2~ yearspreviously were housed as separate groups in ventillated light-proofrooms at ambient temperature and relative humidity. They wereSUbjected sequentially to 8h light: 16h dark (short day) for 3 months,then to 16h light: 8h dark (long day) for 6 months, and again to shortday for a further 3 months. The remaining 11 JTT ewes were housed inan uncontrolled environment. All ewes were offered a maintenance dietso that their body weights did not fluctuate widely from a mean ofabout 30kg for JTT and 45kg for BLM ewes.
Each group of ewes was run continuously with a vasectomized ram,using a total of 4 rams rotated through the 3 ewe groups and a restingpen (uncontrolled environment) at fortnightly intervals. Oestrus wasrecorded daily. Ovulation was recorded by laparoscopy at the end ofthe first short day period (May 1982), at the middle and end of thelong day p~riod (Aug and Nov), and at the end of the second short dayperiod (Feb 1983). Jugular blood samples for plasma progesterone assaywere taken 3 times per week from BLM and JTT ewes SUbjected to daylength cha.nge.
JTT ewes in the uncontrolled environment showed oestrus at regularintervals of about 17 days and showed no marked variation in meanovulation rate between (May 1.91), Aug (l.82), Nov (l.90) and Feb(1.80). ~ll JTT ewes subjected to daylength change and all BLM eweswent into a period of anoestrus, beginning 103±14 (mean ± s.e.m.) and39±8 days respectively after the change from short to long daylength,and continuing until 49±6 and 49±5 days respectively after theSUbsequent change back to short daylength. During anoestrus, all JTTewes continued to show regular cyclical changes in plasma progesteroneconcentration with a period of about 17 days, whereas all BLM ewesshowed a prolonged period of unchanging basal plasma progesteroneconcentrations. Mean ovulation rates in JTT and 8LM ewes respectivelywere 1.80 and 1.80 (May), 1.25 and 1 (Aug), 1.00 and 0.40 (Nov), and1.50 and 1.60 (Feb).
Although JTT ewes were affected to a lesser extent than BLM ewes,the fact that they nevertheless went into anoestrus after exposure tolong daylength indicates that their capacity to breed throughout theyear in the west Java environment would be reduced or lost at higherlattitudes.
(I) Obst, J.M., Boyes, T. and Chaniago, T. Proc. Aust. Soc. Anim.Prod. ll: 321-324 (1980)

a 1#
3637
EFFECT OF MELATONIN ON POSTPARTUM AN()ESTRUS IN BEEF COWS
Different superscripts indicate significant differences (P<0.05).
* *P.H. Sharpe, D.R. Gifford, P.F. Flavel, M.B. Nottle and D.T. ArmstrongTROPICAL RUSA DEER - A LONG DAY BREEDER IN VICTORIA
Simone Van Mourik and T.Stelmasiak
University of Melbourne, School of Agriculture and Forestry, Parkvi~le, Vic., 3052and Attwood Veterinary Research Laboratory, Department of Agriculture and
Rural Affairs, Westmeadows, Vic., 3047.
In the last century Rusa deer (Cervus rusa timorensis) were ,transferred, from thetropics to Australia, exposing the animals to annual changes In photoperIOd. Rusadeer are non-seasonal breeders in the tropics but show an annual pattern ofreproduction in Victoria with a peak in rutting activity in August followed bybirths in April (1).
Since all other boreal deer species are short-day breeders we investigated theendocrinological basis of the long-day breeding pattern of rusa deer. Bl~od
samples were collected from stags (N=lB) at regular intervals over a 2 year perIodand analyzed by radioimmunoassay for luteining hormone (~H), testoste,ro~e (T)and prolactin (Pr!) concentrations. Besides, the responsIveness of pltultarygonadal axis to LH-RH (Hoechst A.G., West Germany) at different times of theyear was investigated.
LH reached maximal concentrations between January-April and was undetectablefor th~ rest of the year. Highest plasma T concentration was recorded in Au~ust
(39.09 + 22.7 nmol/l) and lowest in November (2.99 ..:!:. 2.3 nmol/l). Concentra~lons
for other months were intermediate between these values. Ma~lmalresponsiveness of the pituitary-gonadal axis to LH-RH, expressed In Tconcentrations, was recorded in August (2).
A similar relationship between fluctuations of these hormones and occurr~nce ofreproductive activity and antler growth has been. report~d for other cer~lds ,(3).However, timing of these events for rusa deer tn ~elatlOn to photopertod IS 6months out of phase as compared with other deer species.
In contrast, prolactin concentrations showed iden~ical ~hang~s over the year a~ inthe other cervids. Prolactin concentrations rose tn sprtng WIth peak levels durtngsummer (79.11 + 14.9 ng/m!) and low concentrations during winter (23.32 ..:!:. 11.6ng/ml\ following the changes in photoperiod (4).
These observations illustrate the differences in sensitivity to photoperiodicstimulation and indicate that the endogenous mechanisms regulating the gonadalfunctions may differ between cervid species.
OvulationControl Melato~~
46 ± 3d
60 ± 3b
oestrus and ovulation (x+SE)Oestrus
Control Melato~
48 ± 4d
61 ± 4b
Days from calving to first
Intervals from calving to first oestrus and ovulationin cows and effect of mcliltonin
TABLE 1.
Multiparous
Primiparous
Dept. of Animal*Sciences, Waite Agricultural Research Institute, GIl;;nOsmond, S.A.; Turretfield Research Centre, Rosedale, S.A.
Mean plasma melatonin concentrations during daytime were significantly higher in treated than in control cows one and two weeksafter melatonin injections (330 and 235 vs 117 and 133 fmol/ml,P<0.05). Daytime concentrations of plasma melatonin in treated cowsone and two weeks after treatment were approximately 200-400 fmol/ml(x ± 1 SE) and were within the lower range of night-time valuesreported for Jersey cows (::) and Friesian heifers (3). Thus thr melatonin treatment raised daytime plasma concentrations of melate I n intothe range of night-time vulues for two weeks and melatonin-tJ l.ltedmUltiparo~s cows showed a delay of two weeks in onset of oestrus.The absence nf an effect of melatonin in primiparous cows could bedue to the already-extended postpartum anoestrus. These resultsSupport the possibil i ty of a F)1c of photoper iod (l) I throughmelatonin secretion, in the onset of postparutm oestrus in cattle.
Intervals from calving to first oestrus may be influenced byphotoperiod (1). The effect of melatonin treatment on intervals fromcalving to first postpartum oestrus and ovulation was determined in 45Shorthorn cows which calved May 8- June 14 on Kangaroo Island. Ovulation was determined from progesterone concentrations in jugular venousblood, collected weekly from June Lo August. Melatonin was suspendedin beef tallow at 50°C and single doses of 500 mg in 14 ml were storedin syringes at 4°C. Syringes were warmed to 43°C prior to s.c. injection into 23 of the cows on June 15 (4 to 38 days postpartum) .
Mean intervals to first oestrus and first ovulation (Table 1) weresisnificantly longer in primip~rous than in multiparous cows. Melatonin treatment caused a significant increase in the intervals tofirst postpartum oestrus and ovulation in multiparous but notprimiparous cows.
(1)
(2)
(3)
Hansen, P.J. and Hauser, E.R. 1984. Theriogenology 22: 1-14.Hartin, T.C., Cunninghilm, N.!". <.lIld Silba, N. 1983.J. Endocrinol. 89: 189-196.Kennaway, D. 1984. Pinedl Rcscc1rch Rcvi":\-IS 2: 113-140.
(l)(2)(3)
(4)
Van Mourik, S., Appl. Anim. Beh. Sci. (in press) (1985)Van Mourik, S. and Stelmasiak, T., Compo Bioc~em. Physiol. (in press) (~985)
BUbenik, G.A., Development of Antlers in Cervldae, Caesar Kleberg Wlldl. Res. Inst.,Kingsville, Texas, USA (1982) , "Van Mourik, S. and Stelmasiak, T., Compo BlOchem. Physlol. (In press) (1985)

a ;
38 39
(1) Turek, F.W. and Campbell, C.S. Biol. Reprod. 20: 35-50 (1979).
SEASONAL LH PROFILES IN GANGLIONECTOMISED MERINO CROSSBRED EWES
S.A. Cutler1
, G. Evans 1 and R.J. scaramuzzi 21Department of Animal Husbandry, University of Sydney, Sydney.
2CSIRO, Division of Animal Production, Prospect, N.S.W.
* Based on entire ewes progesterone levels> 1.0 ng/ml
Sham GNX sheep displayed marked seasonal variation in LH pulseFreq. (P<0.05), Amp. (P<0.05) and Base (P<0.05).
Mean LH profile parameters in GNX ,sheep did not differ betweenbreeding and non-breeding seasons. However, individual GNX sheep didshow. a changing pattern of LH secretion which appeared to have noconslstent pattern when the data were pooled. We conclude that thisfluctuating period of high and low pituitary activity, is independent ofdaylength. Since the mean breeding season nocturnal plasma melatoninlevel in GNX ewes was 38±10 pg/ml compared with 320±60 pg/ml in sham GNXewes (P<O.OS), it appears that the asynchronous LH profiles exhibited inGNX ewes are associated with the inability of the pineal gland torelease melatonin in the dark phase.
* Days from OM!? insertion. + x i sem. a,b p(.05 withm columns.
GnRH infusions resulted in ovulation in the treated mares in a doserelated ma.nner. In addition, LH pulse frequency at days 7 and 21 washigher in the treated mares than the controls. ~an daily plasma LH concentrations increased in treated mares during infusion and ~re associated with increased plasma GnRH concentrations. Surges of plasma LH~re associated with ovulation, although no trends in plasma oestradiolconcentrations could be identified in association with follicular growth.Late anoestrus. Ten mares ~re divided into 2 equal groups. One group(C) was not treated and the other group (T) received 100 ng/kg/h GnRH viasubcutaneous OM!? Blood samples ~re collected every 2 to 5 days andovarian palpations ~re carried out to detect ovulation. Mares ~re
ma.ted to a stallion every other day they sho~d oestrus."l'ilre (xisem) to ovulation in C mares (54 . 8±10 . 8 days) was longer
(p<..Ol) than in T mares (18.6i4.l days). This was reflected in the tilreof conception in C mares (67. 8±16. 0 days) which was 6 ~eks later (p <. 01)than in T mares (26.6i9.7 days).
The results of both experirrents demonstrate that GnRH infusion is auseful rrethod for inducing fertile oestrus in anoestrous mares.
INDUCfION OF OVULATION IN MARES DURING SEASONAL AIDESTRUSBY INFUSION OF mNA.IX:YI'OOPHIN RELEASING HOIM:>NE (GnRH)
J.H. Hyland, P.J. Wright, I.J. Clarke*, R.S. carson+,D.A. Langsford and L.B. Jeffcott
Departrrent of Veterinary Clinical Sciences, University of ~lbourne,
Werribee; *Animal Research Institute, Agriculture Victoria,Werribee and +~dical Research Centre, Prince Henry IS
Hospital, ~lbourne.
We have assessed the value of GnRH treatrrent for the induction ofovulation and fertile oestrus in Standardbred rrares during the mid-(June-July) and late (Sept. -OCt.) anoestrous season. .Mid-anoestrus. Fourteen mares ~re divided into 3 groups and ~re glvensubcutaneous infusions of GnRH at doses of 0 (Controls, n=5), 50 (n=5)or 100 (n=4) ng/kg/h for 28 days via osmotic minipunps (a.1P). Folliculardeveloprent was monitored by ovarian palpation and blood samples ~re
collected every 15, 60 or 180 min bet~en 0700h and l800h daily for 6days before and 28 days after OM!? insertion (Table 1).TABLE 1. Plasma. Luteinizing hormone (LH) pulse frequency, LH pulse amplittrle and ovulation data in mares treated with GnRH during mid-anoestrus
Ovulation LH pulseI
LH pulsedata frequency amplitude
I Day*'+ of(x,/8h) (x,ng/ml)
GnRH Mares Day* Day*I(ng/kg/h) n Ovulating Ovulation 0 7 I 21 0 7 21
0 5 0 - 1.0 1.4a 2.2a 0.98 1.09 1.41
50 5 2 l8.5i1.5 1.0 3.0b 3.8ab 1.26 1.07 1.21
100 4 3 17.3+1.5 1.5 3.0b 4.8b 0.83 1.15 1.63
0.79±0.57
0.24±0.09
1.26±0.27
2.59±0.30
"Non-Breeding" SeasonFreq. Amp. Base
no./8 h ng/ml ng/ml
1.16±0.48
3.51±2.02O.59±0.182.18±O.26
LH pulse profile parameters (~ ± s.e.m.).
* "Breeding" SeasonFreq. Amp. Base
no./8 h ng/ml ng/ml
3.75±1.31
TABLE 1.
GNXn=4SHAMn=6 7.35±0.56 O.92±O.14 1.99±0.30
In sheep, the pineal gland is thought to be involved in the mechanisims by which photoperiod influences reproduction. The pineal gland isinnervated by sympathetic nerves linked to the superior cervicalgan?l~a; sectioning of this nerve supply disrupts the normal secretoryactlvlty of the gland. Melatonin is one of the principal products of thepineal, and is thought to influence secretion of the pituitary gland(1). This experiment investigated the effects of ganglionectomy on theseasonal secretion of luteinising hormone (LH) in Merino crossbred ewes.
Ten ewes were ovariectomised (OVX) and silastic implants containingoestradiol-17B were inserted subcutaneously. Four of these ewes werebilaterally ganglionectomised (GNX) and 6 were sham ganglionectomised.In addition, 8 entire ewes were included.
Animals were maintained with nutrition and temperature constant.They were subjected to an artificial lighting regimen of alternating 16week periods of long days (16L:8D) and short days (8L:16D). Twice duringeach period blood samples were taken from all OVX ewes every 10 min. for8 h. LH levels were measured by RIA using N.I.H. oLH WRR-2-B asstandard. In entire ewes, progesterone was measured in weekly bloodsamples to define "breeding" and "non-breeding" seasons. At the midpointof each season, blood samples were taken from all ewes at 2 hourlylntervals for 24 h to monitor diurnal melatonin concentrations.
LH pulse frequency (Freq.), amplitude (Amp.) and Basal level (Base)from all OVX ewes at a sampling period corresponding to the peak andnadir of ovarian activity in entire ewes are presented in Table 1.

40
SEXUAL ~CTIVITY AND SEMINAL CHARACTERISTICS IN RAMS TREATED WITHEPIDERMAL GROWTH FACTOR
P.E. Mattner, B.W. Brown and B.A. Panaretto
CSIRO, Division of Animal Production, Prospect, N.S.W.
There have been no reports on the fertility of rams treated withepidermal growth factor (EGF). In the present stUdy, the sexualactivity and seminal characteristics of adult rams given a depilatorydose of EGF were monitored prior to and for 10 weeks after treat~ent.
EGF prepared from mouse submaxillary glands (1) was given atapproximar:ely 100 ~g/kg body weight over 24 h as an i.v. infusion insaline tv each of 5, four-year old fine Wool Merino rams. Fivesimilar con~r..)l rams received saline only for the same period. Thesexual activir:y of the rams was monitored with libido tests (2) at2-weekly intervals. Semen was collected by electro-ejaculationweekly and the seminal characteristics were assessed as described by(3). Nigrosin-eosin staining was used to determine the proportionsof live and dead spermatozoa (4).
Sexual activity levels. The mean nUmber of services performedby EGF-treated and control rams did not differ significantly duringeither the pre- or post-treatment periods.
Seminal characteristics. In control rams, the mean percentagesfor morphologically normal spermatozoa, for live spermatozoa and formotile s?ermatozoa ranged from 93.8 S.E.M. 2.3 to 95.8 ± 1.2, 85.6 ±2.3 to 95.2 ± 1.3 and 80.0 ± 4.2 to 92.2 ± 1.2, respectively. InEGF-treated rams there was: (a) No material change in the percentageof morphologically normal spermatozoa. (b) A fall in the percentageof live Spermatozoa from 87.9 ± 3.9 to 69.0 ± 13.0 within 1 week oftrea tment and the mean values remained 12-16% lower than those incontrol rams (p < 0.05) for a further 7 weeks. (c) A markeddepression in the percentage of motile spermatozoa at weeks 6, 7 and8 post-treatment p < 0.01 (43.8 ± 13.0, 38.0 ± 13.8 and 29.0 ± 17.1,respect i vely).
Therefore, it appears that, in rams, large doses of EGF mayinduce a temporary effect at a particular stage of spermatogenesiswhich results in a reduced capacity for flagellation of the tailpiecein otherwise apparently normal spermatozoa appearing in theejaculates 6-8 weeks later. The time of appearance of Suchspermatozoa in the ejaculates suggests that this effect is inducedonly in spermatozoa that are at the spermatocyte stage of developmentat the time of EGF treatment.
(1) Savage, C.R. Jnr., and Cohen, S. J. biol. Chern. 247: 7609-7611( 1972).
(2) Mattner, P.E., Braden, A.W.H., and George, J.M. Aust. J. expoAgric. Anim. Husb. 11: 473-477 (1971).
(3) Mattner, P.E., andfuglmayr, J.K. Aust. J. expo Agric. Anim.Husb. 3.: 78-81 (1962).
(4) Hancock, J.L. J. expo BioI. 29: 445-452 (1952).
41
o RAMS IMMUNIZED EARLY IN LIFEDAILY SPERM PRODUCTION OF MAFTUR~~T~~~I~ROM BOVINE FOLLICULAR FLUIDWITH AN INHIBIN-ENRICHED
Al-Obaidi, B.M. Bindon*, T. O'Shea and M.A. HillardS.A R.
Ph siology University of New England, Armidale N.S.W. andDepartment of . ~. f'A' al Production, Armidale, N.S.W.*C.S.I.R.O., D~v~s~on 0 n~m
W;th an inhibin-enriched fraction from bov~ne fImmunization • bert and increased ovulat~on rate 0
follicular fluid (l-bFF) advanced pu (2)YrespectivelY However, there wasMerino ewe lambs (1) a~d mature e~esl rowth in ram' lambs to 8 months ofno effect of immunizat~on on tes;~cutarfgsuch immunization on adult dailye This study descr~bes the e ec 0ag . ( SP) . these ramssperm production D ~n. s'born in Oct. 1982 were treated as
Three groups of 7 Mer~no ram . f 0 3 mg _ 1 0 mg protein I-bFFI eived 9 inject~ons o. .follows: Group 3 rec I received 3 injections of 0.3 mg Ct ~during 3-39 weeks.of ag~ G~OUPf ~ge. Control group (C) received.9 I / . (.protein I-bFF dur~ng.~ w~e s 0 weeks of age. All I-bFF inject~onsinjections of adjuva~t du~~n~r~~~~'s complete adjuvant. One week afterwere given s.c. and ~.m. ~n 11 rams were bled hourly for 5 h.each injection and at 12.mon~hs ~~rar~Df were recorded at the bleedingLiveweight (LW) and test~s d~~:nd FSH were evaluated on composite samplestimes and at 22 and 29 mo. ust 1984 (age 22 mo.) and March, 1985for each ram each month. In Aug, llected for 11 and 9 days,) 1 24h urine samples were co ( )(age 29 mo. c ean DSP by the method of Lino et al. 3.respectively, to.stud~ avera
g; (T ble 1) significantly increased (P <
Immunizat~on w~th I-bF a d'n season (Aug). There was no0.01) DSP in I rams dU~ing the no:~~r~~e~3ggrOUpsdurin~ the breedingsignificant ditference ~n DSP bet~ 'f' ntly (P < 0 05) elevated in 13season (March). Plasma FSH was s~gn~ ~ca t signific~ntlY different at any
k f age Plasma LH was no (P <rams at 25 wee so. f l'f TD was greater in 13
ramspoint during the first.12 months 0 ~ e.
0.025) in Aug but not ~n Mar~h... arl in life appears to eliminate theImmunization against ~nh~b~n e ~ s The effect could be
. DSP of mature Mer~no ram • fseasonal difference ~n 1 ma FSH the overall means 0
related to prepubertal differences i:n~ ~~57 + 0:03 ng/ml NIAMD-oFSH-RP1,which were 0.55 ±.0.04, 1.02 ± 0.25 _f C I and I respectively.or '3 9
. . ht testis diameter and daily spermTable 1. Mean + S.E. L~vewe~g, ontrol (C) Merino rams.excretion in immunized (13 and 19) an~Dc Daily Sperm SignificanceTreat. Date Days LW excretion 13 vs Cgroup (Age urine (cm) rx 10-6 (t-test)
mo) ColI. (kg) 20 41.23
t± 2.77
19C1319
t 1 (1983) Proc. Aust. Soc. Reprod. BioI. 15:82.(1) AI-Obaidi, S. A. R. .!L.£L.., A t Soc Reprod BioI. 14 :85( ) O'Shea T et al., (1982) Proc. us. . 4'2 . B' F' et al (1967) Nature. 213 (5076): 59(3) L~no, •• __

s
42 43
MOTILITY CHARACTERISTICS OF RAM SPERMATOZOA EXTRACTEDWITH TRITON X-100 IS LEYDIG CELL HETEROGENEITY III VITRO AM ARTIFACT?
R. Vishwanath, M.A. Swan* and I.G. White
Departments of Veterinary Physiology, and *Histology and Embryology,University of Sydney, N.S.W., 2006.
Th~ ability of ATP to restore motility of mammalian 3 ermextracted wl~h the detergent Triton X-100 is well documented (1). ~hiSsystem permlts the study of compounds that do not cross the cellmembrane and have direct effects on the anoneme. The aim of thepresent ,study was to assess the extent of demembranation of ram s ermby varylng concentrations of Triton X-100 and analyse th I p.changes in motility characteristics. e resu tlng
R~m sperm, were extracted for 3 minutes in a medium (2) containinO.O~%, 1% Trlton X-100 and reactivated in the presence of lrnM ATP
g
Motlilty characteristics were analysed by % reactivation (i ' •sperm), stroboscopy and cinemicrography. After fixation ~~. mO~ln~of demembranation was assessed by electron microscopy. ' e ex~en
Sperm extracted with 0.01% Triton X-l00 were only artialldemem~ranat~d and e~te~sive areas of plasma membrane remaine: intact
Y
especlally In the mldplece region. Treatment with 0.1% Triton x-106re~oved ~ll plasma membranes and extracted the mitochondrial membranes~n m~tnx. In the absence of ATP, 16.6% + 0.4 (n=5) of the partially
emem rana ted sperm were motile but sperm-extracted with 0 1% T ' tX-l00 were c~mpletely immotile. On adding ATP, partially dem~mbra~~t~~sperm rea,ctlvated better (81.6% .::t 2.8) than sperm completel deme-branated In 0.1% Triton X-l00 (39.5% + 4.6). Y m
. ,There w~s a significant increase-in beat frequency of the neck and'~~~p~ec~lreglons w~en partially demembranated sperm were treated with
ITa e I). Thls was also reflected in the forward velocity which~ear y d~ubled on addition of ATP. Partially demembranated s erm hadlntact mltochondria that presumably were still able to t ~although their movement differed from that of sperm rea~tf~~t~~ew~TthPexogenous ATP.
A.a. Laws, N.G.M. Wreford and D.M. de Kretser
Anatomy Department, Monash University,Clayton. Victoria 3168
In recent years, several ~roups have claimed to isolate twopopulations of rat Leydig cells (LC) of differing densities andsteroidogenic capacity on density gradients 0,2). The aim of thisstudy was to further characterize these populations using; a combinedcytological and functional approach.
Intertubular cell suspensions were prepared by collag;enasedispersion of decapsulated testes from 5 adult Sprag;ue-Dawley rats(2). The cells obtained from 2.5 testes were fractionated on 0-77%Percl)ll gradients. Cells from the gradients were pooled into 8fr.-1.ctions (F) according to refractive index. The cells were fixed incacodylate-buffered glutaraldehyde for light (LM) and electronmicroscopy (EM) or used to determine testosterone (T) production and hCGbinding by Scatchard analysis. LC's were found in FI-4; macrophages,fibroblasts, endothelial cells and residual bodies resided in F3-4.Germ cells were abundant in F5-7 and damaged cells and membranousmateri.al in F8. Stereological techniques at the LM level were used todetermine the averag;e diameter and number of LC's in FI-4 (Table 1).
Table 1: Average LC diameter (ll m) and re lat i ve number of LC's(expressed as a % of the total number of cells in one fraction) in Fl4. Values represent the mean ± SE for 3 experiments.
Fraction Specific Gravity Diameter (llm) Relative Number (7.)
Number (g;/mI)
1 1.0745-1.0900 11.44 ± 0.41 78.93 ± 3.71
2 1.0590-1.0744 10.14 ± 0.54 42.43 ± 2.93
3 1.0515-1.0589 6.57 ± 0.54** 29.45 ± 1.544 1.0440-1.0514 6.11 ± 0.18** 29.61 ± 2.98
* Significantly increased in the presence of ATP (P < 0.001)
(1) Lindemann, C.B. and Gibbons, J.R.(1975).
J .Cell.BioI. 65: 147-162
(2 ) Ishiguro, K. , Murojushi, H. and Sakai,92:777-782 (1982).
H. J .Cell.Biol. Endocrinology
performing detailedisolated on densityheterogeneous cell
(1980)
These results stress the importance ofcytological evaluation of the types of cellsgradients before drawing conclusions aboutpopulations.
1. Payne, A.H. ,Downing, J.R. and Wong;, K.-L.106: 1424-1429.
** p < 0.01 vs FI-2
LC's in FI-2 responded to a maximum dose of hCG (Pregnyl. Or~anon) withan 11-fold increase in T-production and contained 23.068 receptors perLe. In contrast, LC's in F3-4 responded with only a 2.2-fold increasein T-production and contained 15,419 receptors per "LC". Bindingaffinity (Ka) was similar in Fl-2 and F3-4. The average diameter ofLC's in F3-4 was less than that in FI-2 (Table 1). Lig;ht and electronmicroscopic examination revealed that the majority of the lessfunctional Le's in F3-4 were in fact membrane bound non-nucleated piecesof LC cytoplasm.
REGION BEAT FREQUENCY AMPLITUDE FORWARD VELOCITY(Hz) (pM) (flM/sec)
Neck 8.8 .::t 0.9Midpiece
2.5.:t 0.3 38.9 + 1.48.4.::t 0.7 4.9.::t 0.3
Neck *13.5 .::t 0.9 3.2 .::t 0.3 *66.1 .::t 2.8Midpiece *13.5 .::t 0.8 5.8 2: 0.3
1rnM
TABLE 1: Effect of ATP on motility characteristics (Mean + S E ~-5of partlally demembranated sperm extracted with 0.01% Trito~ X~100. - )
ATP
2. Dehejia, A., Nozu. K., Catt, K.J. and Dufau, M.L. (1982) J. BioI.Chem. 257: 13,781-13,786.

44
SEMINAL 1RANSFERRIN, AN INDEX OF SERTOLI CELL F'I.JOCI'ION.POSSIBLE CLINICAL VALUE '
Liu DeYi, E.J. Cooper and H.W.G. Baker
Medical Research Centre, Prince Henry's Hcspital and Howard FloreyIr~stltute of Experimental Physiology and Medicine, MelJ:xmrne.
Sertoli ~el~s in cUltur~ secrete transferrin (1). Most (/80%) ofthe. tr~sfe~rln In human semlnal plasma is of testicular origin andvarlatlons l~ level~ appear to reflect Sertoli cell function {2,3}. Toa~~~ss the dlagnostlC value of seminal transferrin levels in an infertl lty cllnlc, we performed slngle radial immunodiffusion using prepared plates for low concentrat~on ranges (LC-Partigen) and transferrinstandard (~ mg=35 IU) from Behrlngwerke. As shown in the Tableconcentratlons of transferrin were significantly lower than no~~a~ng~~PS of m~n.wlth azoospermia,resulting from gonadotrophin deficiency,~'d arr semlnlfe~ous tubule fallure or genital tract obstruction (epi-
1 ~a obstructlon, vasectomy and congenital absence of the vas and:e~l~~ veslcles). Within groups the concentrations was inversel; <
Ie a to semlnal volume and expressing results as amour.t per eJ'acuate accentuates the differences. .-
TABLE 1. Seminal transferrin (mean, range) in normal semen anal sis(NO~M! or azoospermia due to gonadotrophin deficiency {GD} rim~seml~lferous tubule failure (PSTF) or genital tract obstru~tion (~T)
tate OORM GD PSTF <:ESTn 9 6 8
Concentration 77 17** 36* 11**(UJ/ml) 31-154 5 29 7 110 31- - 10-64Amoun~ 244 28** 104** 72**(UJ/eJaculate) 123-378 11 46 27 462 6 190
W'l . * **1 COXln test v. normal: p<0.05; p<0.01
Semen from men with gor.adotrophin deficiency being treated withg~r.adotrophlns had hlgher seminal transferrin (70 107 and 308 I
!~~~~~:~:~~l~::;l:~~u~~~t~~iP~:~l t~~~~~;~s~~i~:~1~:~ ~::~~a;~~:~~=. oweve~, both prlmary semlnlferous tubule failure and obstru t'
~~~P~i~~:~a~:~l~a~~e:o~hich~verlap. the normal range. It is poss~~~nof spermat . e subJects.wlth obEtruction or severe defectswi th 1 k ogenfesls may result from lnflammcltion in the genital tract
ea age 0 transfernn from blocrl 1 whten-fold higher than . 'lIP asma ere concentrations are
, In semlna p asma (2) The overlap ak . ,~~kel~ ~hat measur~ment of seminal transfe;rin could be :S~St~Ld~=lnguls those subJects with azoospermia, normal serum FSH an l~
~:~~~~lo~r:~~r:~~~~~~~~.SUChas germ cell arrest from thosedw~th
(1) Mather, J.P., Gunsalus, G.L., Musto N.A. et alchem. 19: 41 (1983). ' . J. Steroid Bio-
(2) Holmes~s.D.. Lipschultz L.r., and Smith, R.G.30: 600 (1982).' Fertil. Steril.
(3) Orlando, C., Caldini, A.L. Barni T et al290 (1985). ',.. Fertil. Steril. 42:
45
SERTOLI CELL INFLUENCE ON I>1YOID CELL GROWTH AND/OR PROLIFERATION
E.W. Thompson, J.SeH. Elkington and A.W. Blackshaw
School of Science, Griffith University, Nathan, Qld., andDepartment of Physiology and Pharmacology, University of Queensland,
St. Lucia, Qld.
The co-culture of myoid cells ~",ith Sertoli cells results in theabolition of the usual myoid celh! requirement of serum for maintenanceand/or proliferation in vitro (1). We have attempted to quantify themyoid cell proliferative response using measurement of total DNA,cellular protein and 3H-thymidine (3H-Thy) incorporation. Furthermore,we have investigated the influence of Sertoli cells on myoid cellfibronectin production, since fibronectin has been implicated in themechanism of cell proliferation in other systems (2). We have reportedthat myoid cells produce fibronectin in the presence of either Sertolicells or serum (3), both of which are permissive for myoid cellproliferation. Parabiotic culture, in which myoid cells and Sertolicells share the same culture medium but are restricted from actualcontact by a plastic partition embedded in silicone grease, has beenused to differentiate between cellular and humoral influences.
Sertoli cells and myoid c~lls were prepared and cultured aspreViously described (3). The co-culture of Sertoli cells (freshlyprepared from 20-22 day old rats) with a monolayer of once passagedmyoid cells prepared 7 days f.oreviously from similarly aged animals, wassynergistic in terms of total DNA and cellular protein. This has beeninterpreted as increased growth in the myoid cell component on thebasis that Sertoli cells from rats of this age do not divide inculture (4).
In another experiment, the incorporation of 3H-Thy (scintillationcounting and autoradiography) by myoid cells freshly prepared from20-22 day old rats was increased approximately 5-fold when cultured inparallel to Sertoli cells in parabiotic culture. Sertoli cellsincorporated significantly more 3H-Thy than myoid cells, but were notaffected by myoid cells in paraculture. The combined 3H-Thyincorporation into both cell types in co-culture was greater than theresponse of cells in parabiotic culture.
These results provide quantitative evidence for myoid cellproliferation in response to Sertoli cells, and indicate that this isat least partially due to secreted factors.
Fibronectin was localized in the parabiotic culture experimentusing indirect immunofluorescence as previously described (2).Fibronectin was present in association with individual myoid cells inpara culture with Sertoli cells, but not in myc~u cells cultured alone.
Fibronectin, either supplied in serum sl~pplemented medium orproduced by myoid cells in response to Ser- -,Ii cell products, may beprerequisite for myoid cell proliferation in vitro.
(1) Tung, P.S. and Fritz, I.B., BioI. of Reprod. 23: 207 (1980).(2) Orly, J. and Sato, G., Cell. 17: 295 (1979).(3) Thompson, E.W., Elkington, J.~H. 2nd Blackshaw, A.W. (1983)
Aust. Soc. Reprod. BioI. Abstracts p. 35.(4) Orth, J.M., Anat. ~ec. 203: 485 (1982).

4647
PUBERTAL DEVELOPNENT IN RAM LAMBS WITH H."DUCEDHYPERTHYROIDISM OR RECEIVING ~~INTENANCE FEED
Y. Chandrasekhar, M.J.D'Occhio and B.P. Setchell
Departmen: of ~nimal Sciences, Waite Agricultural Research Institute,Un1vers1ty of Adelaide, Glen Osmond, South Australia 5064.
~n a re7ent study in ram lambs, induced hyperthyroidism wasassoc1a:ed w1th suppressed body growth and retarded testicularmaturat10n (~). The present study was undertaken to determine ifreduced test1s ~rowth ~n.the above lambs represented a direct responseto a hyperthyro1d cond1t10n, or whether it was related more generallto overall suppression in body growth. Twelve ram lambs (22.0+ 1 1 ~amean ±SEM) were divided into three equal groups and over a per~d' f 8'weeks (16-24 wk of ) . d . 0. age rece1ve one of the follow1ng treatments:(d ~lb fed cont:ols (C), feed intake adjusted to maintain body weight~), hype:thyro1d group, daily SC injections of thyroxine (25 ug/kg BW)
w~th.ad llb feeding (H). During week 7, all animals were bled at 20Hlln 1ntervals for 12 h to charactt!rize patterns of LH secretion. LIIand te~tosterone responses to exogenous LHRH (5 ng/kg B\v) weredeterm1ned at 8 weeks. At completion of the experiment animals werer:turned to pasture and periodic measurements of body w~ight and test'~lze were made ~p :0.44 weeks of age. Data were analysed using ANOVA 1Sand the least slgn1flcant difference test. Increments in body weiaht(kf) from 16-24 wk were lower (P<0.05) in M (0.15±0.03, meant SEM)oand~ 0.20iD.?2) ram: compared with Crams (3.s0±O.90). In contrast,1ncreases 1n test1s volume (cm3) over the same period were similar fC (1~.2±2.6) and M (13.1±3.9) animals, but lower (p<O.Os) in H anima~:(0.6_0.3)' The number of LH pulses/12h and Ulean LH levels (nQ/ml) w .also 10wer.(P 0.05) in H rams (1.3±0.3 and 0.2s±0.OS, respec~ively)elecompared.w1th.C (4.S±0.9 and 0.70tO.20) and M (3.3 t O.s and 0.39±0 ~O)r~m:, Wh1Ch d1d not differ from one another. LH responses to LHRH wereslm1lar fo: the.thr:e groups (peak 1.s8±0.30 ng/ml); however, thecorrespond1ng r1se 1n testosterone (ng/ml) was smaller (P<0.05) in Hrams (0.20±0.01+) than in C (1.52±0 42) and N (1.59+0 43) Af
( . • . -. rams. tertreatment 24-44 wk), ga1ns in body weight (kg) were similar for thethree group: (s.s4±0.s3). However, testis growth (cm3) continued to besuppressed 1n H rams (10.2±2.l) relative to C (s4.2±12.2) and M (45 9+~2.1) rams. These results indicate that body growth is suppressed b~th111 H alld H :am lambs; however, testis gro\.!th is n0H11al in N lambs butsuppressed. l~ H lambs. It can be concluded, therefore, thathyperthyro1d1sm has a direct influence on maturation of thehypotha1am~-pituitarY-t!,olladalaxis. Reduced endogenous LH pulsefrequency 1n H rams, together with normal LH responses to exogenousLHRIl, sugg:st that hYl~erthyroi~ismmay influence the activity ofhy~~t~alam~c and/or 111gher bra1n centres. Transient hyperthyroidism~UI It: o an 1mportant phase of sexual maturat ion may cause permanent1mpa1rment of sexual development.
(1) Chandrasekhar, Y. and Setchell, B.P. (1984)Reprod. BioI. ~: 40 (abstract).
Proc. Aust. Soc.
VARIATION IN THE EFFECT OF A NON~ONADOTROPIC LEYDIG CELL STIMULATINGFACTOR IN TESTICULAR INTERSTITIAL FLUID (IF) AFTER EXPOSURE OF THETESTIS TO A SINGLE EPISODE OF HEAT TREATMENT
H. Ishida, G.P. Risbridger and D.M. de Kretser
Department of Anatomy, Monash University, Clayton Victoria 3168
The interaction between seminiferous tubules and Leydig cells is animportant aspect of the local control of testicular function. Recentlytesticular IF was shown to contain a factor and which may be a localregulator which is not LH but is capable of stimulating Leydig cellsteroidogenesis. The exposure of the testis to a single episode of heattreatment results in a temporary disruption of spermatogenesis withsubsequent recovery of germ cells, and we have shown that the capacityof Leydig cells to produce testosterone in vitro was significantlyincreased two weeks after heat-exposure of the testis, and returned tonormal level 8 weeks after treatment. Attempts to demonstrate changesin the interstitial fluid factor were inconclusive. The aim of thepresent study was to re-investigate, using improved methodology, thechanges of interstitial fluid in terms of both its volume and its effecton Leydig cell steroidogenesis after exposure of the testis to heat.
The scrota of adult rats were immersed in a 43°C waterbath for ISmin and studied I, 2, 4 and 8 weeks. The albumin space within thetestis is approximately equal to the volume of testicular interestitialfluid, because albumin does not penetrate appreciably into theseminiferous tubules. In this study the albumin space was measured 20 hafter injection of l2sI-labelled BSA and the volume of IF calculated.Interstitial fluid, treated with charcoal to remove endogenous steroids,was added to Leydig cell preparations which had been purified ondiscontinuous gradients of Percoll. The effect on testosteroneproduction in the presence of a maximally stimulating dose of hCG wasdetermined for each animal.
The stimulatory effect of interstitial fluid was maximal 2 weeksafter heat-treatment and subsequently declined. These changes ofstimulatory activity of interstitial fluid paralleled the previouslyobserved changes of Leydig cell function and germ cell cyto logy af te rheat-exposure of the testis. The volume of IF was unchanged 2 weeksafter heat-treatment but increased significantly after 4 and 8 weeks.
These data support the theory that a non-gonadotropic local factorin interstitial fluid may be involved in the interaction of seminiferoustubules and Leydig cells. The activity of this factor may be influencedby the state of seminiferous tubules and/or the changes in the volume ofinterstitial fluid.
Table I: Duration of Treatment (weeks)0 I 2 4 S
Vol IF( ]JI, n=6) 203.3±8.7 236.s±17.1 214.3±12.4 2s8.9±16.8** 271. 2±10. 7*To Prod.(ng, n=3) 874±SO.9 930±31. 7 1049±45.l* 876±6l.3 837±36.9
Mean ± SE; *p < 0.05; **p < 0.01

48
ISOLATION OF THE GUINEA PIG SPERM ACROSOMEAND CHARACTERIZATION OF ITS ENZYMIC CONTENT
A. Stojanoff, H. Bourne and R.V. Hyne
University of Melbourne Reproductive Biology Unit,The Royal Women's Hospital, Melbourne, Victoria, 3052.
Since the membrane-bound mammalian acrosome has to date never beenisolated as an organelle, it has not been possible to identify itsspecific content with certainty or study its fusogenic properties. Wereport for the first time a procedure for the isolation, under isoosmotic conditions, of acrosomes from guinea pig spermatozoa. Caudaepididymal spermatozoa were suspended in a medium that simulates thecomposition of the cytosol with respect to the main ions composed of70mM KH2P04, 100mM sucrose, 2mM MgS04, ImM EDTA, ImM p-aminobenzamidine(pAB), ImM phenylmethylsulfonyl fluoride (PMSF), 25mM 2-(N-morpholino)ethane sulfonic acid (MES) and the pH adjusted to 6.2 with NaOH(~30mM Na+). The sperm suspension was homogenized at low speed witha Polytron homogenizer. The suspension was subsequently centrifuged(2000 x g, 10 min, 40C) and the pellet resuspended in a iso-osmoticPercoll solution prepared by mixing 9 parts of Percoll solution(Pharmacia) with 1 part of a solution containing 1.6M sucrose and 0.5M
MES (pH3.9) to give a final pH of 5.6. The acrosomes were then isolatedby loading the sperm homogenate on a discontinuous sucrose gradient of1.90M, 1.95M and 2.00M sucrose and centrifuging at 45,000 x g av., 40Cfor 30 min. The acrosomes, which retain their crescentric morphology,banded at the top of the Percoll layer, while the spermatozoa enteredthe Sucrose gradient. Examination of the sperm homogenate by transmission electron microscopy showed that the large apical segment of theacrosomes were removed and were free of plasma membrane, but bounded bydiscontinuous outer acrosomal membrane. For enzymic studies, the acrosomes were lysed by suspending the acrosomes in a hypertonic mediumcomposed of 50mM sodium citrate, 1m, glycerol (pH 2.5). Significantacrosin activity could be detected if the lysed acrosomes were directlyassayed for esterase activity using N-benzoyl-L-arginine ethyl ester(BAEE) as substrate, however maximal activity was obtained usinga soluble acrosin preparation (initially in a zymogen or proacrosinform) after dialysing the lysed acrosome against ImM HCL (pH 3.0).Maximal acrosin activity measured as ~moles of BAEE hydrolyzed perminute gave a specific activity of 5.4 units/mg protein measured inCa2+-free medium buffered with 40mM Tris (pH 8.0). Ca2+(10mM) caused~ time-lag in the autoactivation of guinea pig proacrosin, but did notaffect the maximal velocity. Mg2+(10mM) caused a 64 percent reductionin the maximal velocity of acrosin. The protease inhibitor pAB(lmM)prevented the autoactivation of the proacrosin, whereas PMSF(lmMj hadno effect. The soluble dialysed acrosome extract also contained highactivity of hyaluronidase of 78 nmoles of N-acetylglucosamine released/min/mg protein, but only trace amounts of acid phosphatase activity(0.014 nmoles of 4-nitrophenol formed/min/mg protein) and no arylsUlphatase (pH S.2) activity could be detected.
These data provide unequivocal evidence for the localization ofacrosin and hyaluronidase within the guinea pig sperm acrosome.
F
49
MEASUREMENT AND MANIPULATION OF CYTOPLASMIC FREECALCIUM OF RAM AND BOAR SPERM
A.M. Simpson and I.G. White
Department of Veterinary Physiology,University of Sydney, N.S.W. 2006
In this study a highly selective fluorescent calcium indi~~tor
louin 2' has been employed to measure the cytoplasmic free -Ca conc~ntrations of ram and boar sperm after manipulation by compoundsthat affect Ca 2+ flux (1).
Ram sperm were collected by electrical ejaculation and boar spermwere obtained from the cauda epididymis after slaughter. Sperm werewashed twice in a HEPES-buffered saline solution containing glucoseand sperm !Tl'·tility was scored by microscopic examination. Sperm wereloaded by incub,,+-ion at 37°C with lmM CaCl and quin 2 acetoxymethylester (quin 2/AM) (2) which hydrolyses in the cytoplasm of cells tr-apping the impermeant quin 2. 2+
Fluorescence levels indicated a normal intracellular rCa 1 ofabout 193 nM for ram sperm ~~d 175 nM for cauda boar sperm. Table1 show2 the cytoplasmic lCa ] in ram and boar sperm after add1ngthe Ca + iocophore A23187, filipin, rotenone, antimyc1n A and carbonylcyanide m-chlorophenylhydrazone (CCP).
TABLE 1. Cytoplasmic free-Ca 2+ concentrations (mean nM + S.E., n=3)of ram and boar sperm.
SPERM TIME CONTROL A23187 FILIPIN ROTENONE ANTIMYCIN CCP(mins) (3.2IJM) (0.2nM) (6IJM) A (8IJM) (2IJM)
RAM 5 193+8 *380.:1:: 11 *360+31 *212+8 *285+11 220.:1:: 1720 193+8 *380+ 11 *360+31 *105+8 210+12 *103+8
BOAR 5 175.:1::8 *420.:1:: 11 *307+38 *195+5 *275+14 213+1420 175+8 *420.:1:: 11 *307+38 *182+9 190+19 *68.:1::9
P < 0.01; significantly different from the control.
Additiun of A23187 immediately increased [ca 2+] which remainedelevated for the 20 mins of the experiment. Apparently this wasnot dependent on the oxidative metabalism of the mitochondria assUbseq~~nt addition of the inhibitor, CCP, to the spe~~ had no effecton lCa 1 in either spec1es. A lar~e lncrease 1n rCa 1 also occurredon adding filipin, due to increased permeability of the sperm membranes. When rotenone, antimycin A and CCP were added t02~uin 2 loadedram and bear sperm there was a transi1~t elevatlon of Ca posslblycaused by release of mi~~chondrial Ca . This was followed by subsequent extrus10n of Ca and return to control levels or ~~low,
most likely due to the operation of the plasma membrane Ca pump.
(1) Simpson, A.M., & White, I.G. (1984) Proc. 16th Annu. Conf.Aust. Soc. Reprod. BioI. Melb: 45.
(2) Tsien, R.Y., Pozzan, T., & Rink, T.J. (1982) J. Cell. BioI.94: 325-334.

50
IMMUNOSUPPRESSIVE AND CYTOTOXIC EFFECTS OF SEMINAL PLASMA
R.D. Allen and T.K. Roberts
Department of Biological Sciences, University of Newcastle.Shortland, NSW. 2308
Seminal plasma is known to contain factors which can inhibit variouslymphocyte functions in vitro (1). These factors have also been shown toact in vivo. Indeed, some speculation exists as to the role of 'thesefactors in the predisposition of male homosexuals to AIDS (3).
We have investigated the immunosuppressive effects of seminal plasmaand have observed a cytotoxic effect of seminal plasma on lymphocytes.In the present study we set out to further investigate the cytotoxiceffect of seminal plasma on lymphocytes and to determine whether thiseffect was involved in the observed immunosuppressive effects of seminalplasma. Since the immunosuppressive properties of seminal plasma havebeen reported to be non-species-specific (4), we studied two systems:-a homologous system of mouse seminal constituents on mouse lymphocytesand a heterologous system of human seminal plasma on mouse lymphocytes.
Human seminal plasma, as well as extracts of mouse seminal vesicles,prostate and epididymis all showed strong inhibition of the response oflymphocytes to Concanavalin A, as determined by 3H- t hymidine incorporation(Table 1).
TABLE 1. Suppression of the response of mouse lymphocytes to Con A byhuman seminal plasma (Husepl, 572 ug/ml), and extracts of mouse seminalvesicles (SV, 775 ug/ml), prostate (Pros, 163 ug/ml) and epididymis (Ep,338 ug/ml). Results are expressed as counts per minute.
Control Husepl SV Pros Ep
cpm 72,901 1,562 14,180 41,733 10,826+ 14,076 + 1,589 + 2,849 + 8,301 + 3,684
%inhibition 97.9** 80.5** 42.8* 85.1**
%~nh~b~t~on-((control-test)/control)xlOO%
*P<O.Ol; **P<O.OOOl by Student's t-test
However, human seminal plasma was found to have a strong cytotoxiceffect on the mouse myeloma line NS-l, while seminal vesicle extracts ofthe mouse had no cytotoxic effect. Human seminal plasma was alsocytotoxic for mouse splenic lymphocytes and, durj~g Con A stimulation oflymphocytes, there was a close inverse correlation between human seminalplasma dose and lymphocyte viability (r=-0.994) and also close correlationbetween viability and the degree of response to Con A (r=0.95 at 6 ug/mlCon A and r=0.991 at 8 ug/ml Con Ai.
It therefore appears that a) human seminal plasma contains a factor~hat is cytoto~ic for lymphocytes, and b) some of the work suggesting~mmunosuppress~on by seminal plasma may be explained by the cytotoxiceffects of seminal plasma. In view of the strong cytotoxic effects ofrelatively low concentrations of human seminal plasma, it is interestingto speculate on the possible effects that the cytotoxic factor may have onlymphocytes in vivo.
(1) Hess, E.V. and Marcus, Z.H. (1980) J. Lab. Clin. Med. 96:577-581.(2) Anderson, D.J. and Tarter, T.R. (1982) J. Imm. 128:535:S3S.(3) James, K. (1984) Nature 309:116. ---(4) Lord, E.M. et a1 (1977) ~Imm. 118:1704-1711.
51
THE FAILURE OF THYROID ALLOGRAFTS IN THE OVINE TESTIS
*S. Maddocks, J. Cormack and B.P. Setchell
Department of Animal Sciences, Waite AgriculturalResearch Institute, University of Adelaide,
Glen Osmond, South Australia 5064.* Department of Radiology, Flinders Medical Centre,
Bedford Park,South Australia 5042.
The mammalian testis has historically been regarded as animmunologically privileged site. We have previously shown t~atallografts of pancreatic islets of Langerhan~ tra~splanted lntothe testes of rats will survive and secrete lnsulln (1). Otherpreliminary experiments in our laboratory have co~firmed thereport of Aron et al (2) that thyroid allografts ln ra~ testeswill survive and concentrate iodine. Because of anatomlcaldifferences, the rodent testis may not be a very suitable modelfor the testes of other species. Consequently, we hav: .investigated the survival of thyroid grafts in the oVln: testlswhich is morphologically more similar to the h~man te~tls.
Eight sexually mature Merino rams were thyroldectomlsed and atthe same time 4 received intratesticular autografts and 4received allografts of one thyroid lobe into one testis with thecontralateral testis serving as a control. Four weeks aftertransplantation all animals received an intravenous bolus ofIodine-131 (1-131) and were castrated 48 hours later. Venousblood and lymph from both testes and jugular blood samples werecollected at the time of castration for comparison of ~hyr~idhormone levels. Each testis was imaged for 1-131 locallsatlonusing a gamma camera. The testes were then .cut into thicksections, and tissue from the transplant slte of the grafted.testis and the same region of the control testis was counted ln agamma counter and then prepared for histology. .
Localised uptake of iodine by the grafts was observed ln thetestes of 3 of the 4 autografted animals. However, none of the 4allografts showed any specific localisation. His~ologi:alexamination revealed characteristic thyroid foll~cles ln ~hetestes of 3 of the 4 autografted animals suggestlng.funct~onalthyroid grafts, ~ut only follicle remnants were eVlde~t ln theother autografted animal and all of the allografted anlmals.Hormone levels are currently being assayed.
The results to date indicate that thyroid allografts do notsurvive in the ovine testis. This implies that theimmunologically privileged status of the t:stiS may not b: ~general mammalian characteristic, but may ln fact be speclflc torodents.
(1) Maddocks,S.; Oliver, J.R. and Setchell, B.P.(1984)I.N.S.E.R.M. 121: 497-502.
(2) Aron, M.; Marescaux, J. and Petrovic, A.(1957)Colloq. Int. C.N.R.S. ]a: 25-33.
S.M. is supported by the Wool Research Trust Fund.

• p52
53
THE INFLUENCE OF THE PITUITARY ON OVARIAN COMPENSATIONFOLLOWING UNILATERAL OVARIECTOMY IN SHEEP. OVARIAN SECRETION OF PROGESTINS DURING LATE PREGNANCY IN THE RAT
R.C. Fry, I.J. Clarke+ and L.P. Cahill B.J. Waddell, N.W. Bruce and A.M. Dharmarajan
Day 22/PNDay 22/AMDay 16
Ovarian secretion rates (~g/h per ovary) of progesterone and20a-OBP in rats at days 16 and 22 of pregnancy (means ± SEM).
Secretion of:1. Ob b
Progesterone 26.9 ± 4.0a 4.5 ± 3.2 ± 0.3ba a20a-OBP 5.6 ± 1. 7a 5.4 ± 1. 3b
18.9 ± 1.2
Total 32.5 + 5.2 9.8 ± 1.2 22.6 ± 1. 4c
Row values without common superscripts differ, P<0.05, one way ANOVA.
Table 1:
1) Wiest, W.G. ~ al. (1968). Endocrinology 82, 844-859.2) Lacy, L.R. ~ al. (1976). Endocrinology 99, 929-934.3) Bruce, N.W. & Meyer, G.T. (1981). J. Reprod. Fert. 61, 419-423.
Department of Anatomy and Human Biology,The University of Western Australia, Nedlands, W.A., 6009.
Ovarian P secretion fell between day 16 and day 22/AM but remainedunchanged during day 22. The rate of 20a-OHP secretion was similarbetween day 16 and day 22/AM, but increased substantially by theafternoon of day 22. Total progestin secretion (p + 20a-OHP) fellsignificantly between day 16 and day 22/AM (70% fall, P<O.OI) butincreased by more than twofold from the morning to the afternoon of day22 (P<O.OI). This final value for total progestin secretion remainedsignificantly lower (31%, P<O.05) than the original day 16 value.
Overall, these res~lts contra-indicate the operation of acatabolic regulation model for the control of ovarian progestinsecretion. Furthermore, the apparently active'stimulation of 20a-OHPsecretion during day 22 raises the p08sibility that this progestin hasa direct biological role at this stage of pregnancy.
Ovarian secretion of progesterone (P) falls prior to parturitionin the rat, and secretion of 20a-hydroxypregn-4-en-3-one (20a-OHP)increases. These patterns of secretion led to proposal of a ~catabolic
regulation model~ for the control of ovarian progestin secretion in therat (1). This model contends that total progestin ~ecretion by theovary remains relatively constant during late pregnancy, but itsprogestational potency is reduced by switching the major secretoryproduct from P to 20a-OHP. However, the model has not been directlytested by measurement of ovarian progestin secretion dur·ing theswitchover period. Furthermore, indirect evidence from peripheralblood concentrations does not support the model (2).
In the present study, an ovarian venous outflow technique (3) wasused to directly measure ovarian progestin secretion rates at day 16 ofpregnancy, the time of maximal P secretion, and in the morning (before14.00 hours) and afternoon (after 14.00 hours) of day 22. Five ratswere examined in each group and mean secretion rates were derived fromeither three or four estimates made in each rat. Results are shown inTable 1.
C.L. perovary in ewesovulating
1.81.8
4/65/6
Treatment Group No. of Ewes. C.L. perovulating ewes ovulating
1. Hypox-entire2. - Hypox-U.L.O.
3. - Intact-entire 6/6 2.3 1.2a~. - Intact-TI.L.O. 6/6 3.0 3.0c
Following unilateral ovariectomy (TI.L.O.) as late as Day 14 ofthe cycle in the sheep compensation occurs in the remaining ovary(1) and the normal ovulation rate per ewe is maintained. A smalltransient elevation in the FSH levels, 5-12 h following TI.L.O. hasbeen observed, suggesting that gonadotrophins may be involved inthis compensatory phenomena. This experiment was conducted tcexamine the role of the pituitary and gonadotrophins in ovariancompensation following TI.L.O.. Progestagen intravaginal sponges (Repromap, Upjohn) werelnserted for 12 days in 24 BL x Mer ewes (4 groups x 6 ewes)during the breeding season. At sponge removal the ewes weredivided into the following groups : Group 1- (hypox-entire)hypophysectomy and 500 iu. PMSG at 0 h, 250 iu. PMSG at 24 hand 750 ~u. HCG at 48 h after sponge removal; Group 2(hypox-TI.L.O.) - same treatment as group 1 plus U.L.O. wasperformed immediately following hypox; Group 3 (Intact-entire)- the same exogenous hormonal treatment as group 1 but thepituitary was left intact and Group 4 (Intact-U.L.O.) - the samehormonal treatment as group 1 with pituitary left intact plusU.L.O. p8rformed at sponge removal. Ovulation rate was recordedby endoscopy 34 halter HCG injection. Analysis of ovulationrate was carried out by 2 way Anova.Table 1. Ovulation rate of ewes 34 h after HCG injection and
following hypophysectomy and TI.L.O.
Victoria~ Department of Agriculture and Rural Affairs,Anlmal Research Institute, Werribee, 3030.
+ M.R.C., Prince Henry's Hospital, Melbourne.
Dlfferent superscripts denote significant differences P<0.05.In hypox ewes there were single or twin ovulations whilst
int~ct ewes with both exogenous and endogenous gonadotrophins hada ~lgher O.R. In both hypox and intact ewes compensation followingTI.~.O. occu:re~ ~uch that.O.R. per ewe was maintained or O.R. perovary w~s slgnlflcantly hlgher (P<0.05) in the remaining ovary.. It lS concluded that compensation following TI.L.O. actslndependently of the pituitary. It is possible that follicle~rowth in~ibito:s found in ovine follicular fluid (3) could belnvolved In an lnter-ovarian mechanism.(1) Land (1973). J.Reprod. Fert. 33, 99-105.(2) Findlay & Cumming (1977). Bio~ Reprod. 17 178-183.(3) Cahill et al., (1984). Proc. Aus. Soc. Rep: BioI. Abstr.

.....
54
BLOOD FLOW IN THE OVARY AND OVIDUcr OF THE RAT AFrERSYMPATHETIC DENERVATION
W.R. Gibson and P.J. Roche
f55
OVARIAN OXYTOCIN AND PROGESTERONE SECRETION AND DEGRANULATION OFLUTEAL CELLS IN SHEEP GIVEN EXOGENOUS PROSTAGLANDINS.
R.J. Fairclough, L.D. Staples and J.D. O'Shea*
Department of Physiology, Monash University, Clayton, Vic. 3168.
. . Sympathetic nerves of the ovary terminate near follicles, interstltlal cells, the margins of corpora lutea (CL), and blood vessels.Those of the oviduct terminate mainly near blood vessels and smoothmuscle ~f the wall of the isthmus. In earlier work we found thatdestru:tlon of. these nerves by freezing did not interfere withovulahon or wlth pregnancy in the rat (1,2). In the present studyb~oorl flow was. measured in denervated ovaries and oviducts of' rat~e1 ther un~ergo1ng oestrous cycles or during pregnancy when corporalutea are large and fully functional.
Unilate~al denervations were done by briefly freezing the ovarianvascular ped1cle and the ovari~n suspensory ligament at metoestrus oron day 7 or 13 of pregnancy. Blood flow was measured either 1 d or 410.d after s?fger~ (always on d 14 of pregnancy in the pregnant rats)u~lng 15)Jm .Co-mlcrospheres ('3) while the rats were anaesthetizedw1th me.thohexlt~ne sodium (Brietal sodium, Eli Lilley), 5Omg/kg andamobarbltal sod1um (Amy tal SOdium, Eli Lilley), 83mg/kg. Denervationwas checked by fluorescence microscopy.
In 15 cyclic rats, the day after surgery, blood flow to thede~ervated oviducts was 88% higher than to contralateral controloVlducts (5.~±0.9 (SEM) v. 2.9±0.7 ml.min- 1 per g; P<0.01, Wilcoxontest). Ovanan blood flow was not affected (12±1 v. 11±1; N.S.). In11 rats at 4-10 d, oviducal blood flow was 42% higher on thedenervated side (2.5±0.5 v. 1.8±0.5; P<0.1) and ovarian blood flow wasnot affected (6.6±1.6 v. 5.9±1.1; N.S.).
In 8 pregnant rats, the day after surgery, blood flow to thedenervated oviducts was increased 5-fold (4.6:1:1.6 v. 0.9±O.1; P<O.01)but that to the ovarian tissues was not affected (CL: 26±3 v. 29:1:2non-luteal.tissue: 8.3±0.9 v. 6.8±1.3; N.S.). In 7 rats at 7 d afte~surgery oV1ducal blood flow remained elevated (4.1±1.6 v. 1.2±O.6;P<0.05) and that ~o the ovarian tissues remained normal (CL: 20:1:4 v.24±1; non-luteal tlssue 4.6±1.0 v. 5.2:1:0.5; N.S.).
It is concluded that hlood flow to the oviduct is normallyregul~ted by neural vasoconstrictor activity. The physiologicalfunctl~ns of the oviduct are not impaired by the increased blood flowfOllo~llng denervation, since normal pregnancies can occur (2). Ourexper~mental procedure does not reveal any neural regulation of bloodflow ln the ovary. -
(1984)
(1)
( 2)
(3)
Wylie, S.N., Roche, P.J. and Gibson, W.R. (1983)Soc. Reprod. BioI. 15: 7.
Roche, P.J., parkingto~ H.C. and Gibson, W.R.Aust. Soc. Reprod. BioI. 16: 30.
Bruce, N.W. (1976) J. Reprod:--Fertil. 46: 359-362.
Proc. Aust.
Proc.
Department of Agriculture and Rural Affairs.* Animal Research Institute, Werribee, 3030 •
Department of Veterinary Preclinical Sciences,University of Melbourne, Parkville.
The nature of the substance(s) stored in densely staining"Secretory" granules of the ovine corpus luteum (CL) is stilluncertain although some authors have suggested previously that thesegranules contain progesterone (1). More recent studies havesuggested that oxytocin is localised in granules of the CL (2). Thepresent study ~as undertaken to determine oxytocin and progesteroneconcentrations in ovarian venous plasma and to observe ultrastructural changes to e~es given exogenous prostaglandin F~ or PGE2.
PGF2u (1 ~g) in 0.5 ml plasma ~as injected into one ovarianartery of t~o e~es ~ith bilateral CL and the ipsilateral ovary of oneunilaterally ovulating e~e. Four unilaterally ovulating ewes weregiven arterial injections of either PGE2 (1 pg, n=2 e~es) or theplasma vehicle (n=2). Ovarian venous blood samples were collected at15, 10 and 5 min before and at 2, 5, 10, 15 min after the injection.The ovaries were then removed, perfused with 3% glutaraldehyde in 0.1M cacodylate buffer and po~t fixed in 1% osmium tetroxide forultrastructural examination. Oxytocin was extracted from plasmausing C-18 Sep-Paks and oxytocin and progesterone were analysedusing radioimmunoassay. In ewes given PGF2a or PGE2, the ipsilateralovarian venous oxytocin concentrations rose sharply from a basalvalue of 6.8 ng/ml to a mean maximum of 54 ng/ml (range 30.5 - 85ng/ml) or 92 ng/ml (range 75.5 - 110 ng/ml) respectively at 10 minpost-injection. Oxytocin concentrations in ovarian venous plasma ofthe four untreated or control CL sho~ed no significant rise with theconcentrations remaining below 7 ng/ml. Ovarian venous plasmaprogesterone concentrations did not show a significant change in thePG-treated and control ewes although there was considerable variationin the basal concentrations (range 1.8 - 3.3 ~g/ml). Secretorygranules in the control CL were distributed singly and uniformly withoccasional evidence of exocytosis of single granules. In PGF2atreated CL, the majority of secretory granules were present inclusters close to the cell membrane and many granules were alsopresent in the extracellular space. Aggregation and exocytosis ofgranules also occurred follo~ing PGE2 treatment although to a lesserextent than that following the PGF~-treatment.
The close association between degranulation and an increase inovarian venous oxytocin concentrations in the PG-treated ewes givesfurther support for the view that the secretory granules of thecorpus luteum are the source of ovarian oxytocin in the ewe and thatoxytocin is released rapidly in response to PGF2a or PGE2' Howeverthe data provide no support for an association between granulerelease and progesterone secretion.(1) Gemmell, R.I., Stacey, B.D. and Thorburn, G.D. BioI. Reprod.
11 : 447. (1974) •(2) Rodgers, R. et al. Endocrinology 113 : 2302 (1983).

56 57
VASQJLAR GROWTH IN DEVELOPING LUTEAL TISSUE.
Karen I. Mackin and Geoffrey I, Meyer.
Department of Anatomy and Human BiologyUniversity of Western AustralIa. Nedlands. 6009
Formation of the corpus luteum (OL) Involves vascularlzatl~ of theruptured follicle and complete luteInIzatIon of the granulosa eel Is. IheaIm of this work was to determIne whether there Is an Interactionbetween these two processes.
The left ovary of rats at 12 hours (n=8 rats). 24 hours (n=9) and 36hours (n=8) of gestation was processed for lIght microscopy. SectIonswere cut through each ovary until a newly-formed CL was recognized; a0.5 um section was taken through the maximum diameter of this OL. Usingstandard stereologlcal methods, regIons of tissue were examined fromeach quadrant of al I OL sections to obtain quantitative morphologicalInformatIon. REGION 1 was adjacent to the outer thecal capsule of theCL. REGION 3 was near the centre of the CL. and REGION 2 was anIntermediate area of luteal tIssue.
Ihere was no evidence that angiogenesis was Inltlafed preferentiallyfrom the rupture point or In regIons Immediately adjacent to stromalvessels. Capillary sprouts entered the ruptured fol I Icle equally fromIts entire circumference. Subsequent development of the vascular spaceInvolved a high level of dIvisIon (Indicated by tritiated-thymIdinelabel lIng) and migratIon, of the endothelial cel Is. The number ofendothelial cel Is within each OL Increased signIfIcantly between 12hours ( 8.8 ± 2.i thousand) and 36 hours (31.4 ± 6.5 thousand).
At 12 hOllrs, cap" larles occupied 8.4 ± 1.8% (mean ± S.E.M.) of theperiphery of the CL (REGION 1); luteal cel Is In this regIon containednumerous lipId droplets and had a cel I volume of 0.65 ± 0.06 pl. Incontrast. the central area of the CL (REGION 3) was avascular; cel Iscontained some lipid but were much smaller (0.45 ± 0.05 pI). But by 2AhQucs. luteal cel I sIze was sImilar throughout the tissue (0.81±O.07.0.7610.07, 0.70±O.06 pi In REGIONS 1-3 respectIvely). This was despItesignificantly less vascularity In the central REGIONS (6.8±1.5, 4.3±1.3,2.2±1.2 %; P<0.05). Similar amounts of lipid droplets were observed Inthese cel Is regardless of theIr position In the tissue. A large centralfluld-fll led antrum had now formed which occupied about 14% of the totalCL volume; this antrum was lined by a squamous-I Ike epithelium. Therewas also much Intercellular fluid between the luteinIzing cel Isparticularly toward the central REGION. ThIs fluId Is presumably plasmafrom the very II Ieaky" vascuf ar sprouts.
By 36 hOllrs, both luteal cell volume (1.12±Q.17, 1.1510.14,1.1610.29 pi) and vascularIty (13.7±2.8, 12.2±3.3, 12.4±5.5 %), weresImIlar throughout the entire CL. The large central antrum was nowabsent but we have histological evIdence to suggest It was Involved Inthe formatIon of a venous outlet.
We conclude that Initially. eel Is Immedlatlely adjacent to newvessels showed advanced luteinIzation. However, the extremepermeabIlity of these sprouts and the resultant extravascular plasmaenables a blood-borne luteInizIng stimulus to promote dIfferentiation ofthose eel Is Isolated from these capIllarIes. This differentiation andthe Increased metabol Ie activity of these eel Is may stimulate completeluteal angIogenesIs.
FLOW RATE AND COMPOSITION OF OVARIAN LYMPH IN THE DAY 16 PREGNANT RAT
A.M. Dharmarajan, N,W. Bruce* and H.J, McArdle*
Department of Anatomy and Human Biology and* The Raine Centre for the Study of Perinatal and Developmental Biology
The University of Western Australia, Nedlands, W,A. 6009.
The mammalian ovary has a remarkably rich blood vascular system anda well developed set of lymph drainage vessels (1). The few studiesavailable on rates of lymph flow from the ovary suggest that they are
. relative to most other organs of the body. Little is known of thehlgh "t· fbiological significance of the ovarian lymph or of the. composl lon .0
lymph relative to ovarian venOUS blood. Such comparlsons can provldeinformation on the mechanisms and significance of transport processes
between luteal cell, interstitial space and b~OOd (2), .The present study was therefore carrled out to examlne rates.of
ovarian lymph and blood flow in five rats at day 16 of pregna~cy, a tlmeof maximal progesterone secretion, Rats were anaesthetlzed an~ amodification of a venous outflow technique was used to collect .ovarlanvenous blood (B) and lymph (L) for a 2 hr period. Both flulds wereanalysed for progesterone (P), 20a-hydroxypregn-4-en-3-one ~2?a-OHP),total protein, transferrin and albumin concentrations. In.ad~ltl~n, SDSgel electrophoresis was carried out to obtain an. initial . lndlcatlon ofpermeability of capillaries to the various proteln.f~actlons. .Table 1: Ovarian blood flow, lymph flow and composltlon of ovarlan
plasma and lymph
Blood Lymph L/B(%)~ **
Flow rate (ul/min) l70± 1.1 ± 0.2
Progestin conc (ng/ml) **Progesterone 2824 ± 469 1049 ± 114 37
± 118 48 **20a-OHP 1107 ± 70 530
Secretion rate (ug/h/ovary)0.5 **
Progestin 22.1 ± 1.7 0.10 ± 0.0145 ± 7 50 **
Total protein (mg/ml) 90 ± 6
Albumin (mg/ml J 37 ± 3 28 ± 6 75 NS
0.6 51 *Transferrin (mg/ml) 4.8 ± 0.6 2.1 ±
Values are means ± SEM: n=5. *, P < 0.05; ** P < 0.01 (paired "t" test)The rate of lymph flow from the ovary was high, (0.6% of the rate of
blood flow) and in agreement with studies on sheep ovaries (1,3~. As aroute for progestin secretion however, th: lymph dr~lnage isinsignificant (0.5%). There were high concentratlons .Of maJor .serum
teins in the lymph but from the electrophoretlc analysls the~:~viest ~olecular weight protein fraction did not appear in the ~ymp~.The most surprising finding was that concentrations of pr~gest~ns.lnlymph were lower than those in ovarian venouS plasma. . ThlS "flnd~ngquestions the view that progestins are transported by slmple dlffUS1~nfrom the luteal cell to the blood and suggest that there lScountercurrent flow between the interstitial space and blood.
1) Morris, B. & Sass, M.B. (1966). Proc.R.Soc.B 164, 577-591.2) Dharmarajan, A.M, et aI, (1985). Am. J. Anat. 172, 87-993) staples, L.D. et al. (1982). J. Reprod. Fert. 6 4, 409-420

F
58
THE EFFECT OF TESTICULAR WEIGHT AND INSEMINATION TECHNIQUE ONFERTILITY
r!~,!j~
II
59
HORMONE EFFECTS ON PROTEIN SYNTHESIS AND SECRETION BY SERTOLICELLS IN THE DEVELOPING BOVINE TESTIS.
A.W.N. Cameron*, A.J. Tilbrook*, D.R. Lindsay*, E.J. Keogh+and I.J. Fairnie¢.
+Departments of *Animal Science agd Clinical Biochemistry, Universityof Western Australia, Perth and Muresk College, Northam, W.A.
M.K. Hayes and D.E. Brooks
Dept. Animal Sciences, Waite Agricultural Research Institute,Glen Osmond, South Australia, 5064.
There were no significant interactions but the difference betwp.eninsemination techniques tended to be greatest in unteased ewes,inseminated with only 50 million sperm (table 2).
(1) Blockey, M.A. deB. (1980) Proc. Aust. Soc. Anim. Prod., 13: 52-53.(2) Kerton, D.J., McPhee, S.R., Davis, I.F., White, M.B., Banfield,J.C. and Cahill, L.P. (1980) Proe. Aust. Soc. Anim. Prod. l2: 701.
In conclusion there is no difference in the quality of semen from ramswith large or small testicles. OTR was better than SID but furtherwork is justified in determining whether this can be overcome byinseminating more sperm or post-inseminal teasing.
Table 2. Percentage of ewes pregnant.Dose of Sperm 250 million 50 millionPost-inseminal teasing Plus Minus Plus MinusInsemination technique OTR 50.7 58.5 47.9 49.0
SID 51.2 50.2 40.8 33.9
thederitheof
AGE OF ANIMAL TESTIS WEIGHT RELATIVE INCOPORATION OF RADIOACTIVITYINTO SECRETED PROTEINS
(months) (gms) TreatmentI II III IV
5 45 100 60 735.5 51 100 135 68 866 73 100 131 89 1276.5 71 100 105 146 151
11 231 100 86 171
I no treatmentj II FSH, 25 mg/mlj III Testosterone, 0.7 ~Mj IV FSH, 25ng/ml; Testosterone, 0.7 ~Mj Insulin, 5 ~g/ml; Transferrin, 5 ~g/mljEpidermal Growth Factor, 10 ng/ml; Retinol 0.35 ~M. All treatments wereduplicated.
Differences in protein secretion between treatments (Table 1) canbe attributed to differences in total synthesis of proteins, as measured by incoporation of radioactivity into ~ytosolic pro~eins. At 5.5and 6 months FSH increased protein synthes1s and secret1on. At 6.5months, an increase occurred with Testosterone treat~e~t. At a~l ages,the effect of hormones was to change the overall rate of prote1~ synthesis and secretion, but no spec1fic qualitative changes were 1nduc~d
in the pattern of secreted proteins. N~ major dif~erenpes were seen 1nthe secreted protein pattern of Serto11 cells der1ved from calves ofdifferent ages.
(1) Hayes, M.K., and Brooks, D.E. Proc. Aust. Soc. Reprod. BioI. ~: 83(1984) .(2) O'Farrell, P.H. J. BioI. Chern. 250: 4007-4021 (1975).
TABLE 1. Effect of hormone treatment and age on incorporation ofradioactivity into secreted proteins.
We have previously reported that there was no difference inpattern of secreted proteins from Sertoli cell-enriched culturesved from immature and mature bovine testes (1). We have expandedstudy to investigate protein synthesis and secretion from testescalves covering the pubertal period from 5 to 11 months of age.
Sertoli cell cultures were successfully established from animalsof 5 to 6.5 and 11 months. Hormones, added to the culture medium, weretested for their effect on incorporation of radioactive methionine intototal and secreted protein and the composition of the radio~ctive proteins was investigated by two dimensional gel electrophores1s (2).
NSP<0.05P<0.05NS
SignificancePregnant46.744.043.047.3
48.9 vs51.3 vs52.7 vs48.3 vs
Percentagepregnant.Table 1. Percentage of ewes
We wished to examine whether in rams, as in bulls (1), lowtesticular weights are associated with poor quality semen. In additionwe examined whether the reduced fertility obtained using 'shot in thedark' (SID) rather than 'over the rail' (OTR) insemination (2) could beovercome using larger numbers of sperm or post-inseminal teasing.
Fifty-eight Merino rams were screened for testicular weight usingan orchidometer, of which the 8 with the smallest testicles (450±23,X±S.E.) and the 8 with the largest (700±46, X±SE) were chosen. Semen,collected from these us~ng an artificial vagina, was diluted to ac~ncentration of 1 x 10 sperm per ml using UHT milk. This was used toinseminate oestrous ewes obtained daily from a flock of 2.5 yo,naturally cyclic Merinos, beginning on the 5/01/85. The ewes wereallocated to a factorial experiment in which they were inseminated witheither 50 or 250 million sperm, obtained from rams with large or smalltesticles, using SID or OTR insemination, with or withoutpost-inseminal teasing. Pregnancy was diagnosed using realtimeultrasound scanning, 35-45 days after insemination. Pregnancy results,obtained for 814 ewes, were subjected to analysis of variance.
The only significant factors were the method of insemination andthe number of sperm inseminated (table 1).
TreatmentLarge vs Small TesticlesOTR vs SID50 vs 250 million spermPost-inseminal teasing (+ or -)

p
60
THE EFFECT OF (S)-a-CHLOROHYDRIN ON THE METABOLIC ACTIVITYOF EJACULATED RAM SPERMATOZOA
T-A. Reidt, A.R. Jones t and I.G. White*
Departments of Biochemistryt and Veterinary Physiology*,The University of Sydney, Sydney, N.S.W. 2006
The mechanism of action of (R,S)-a-chlorohydrin as a maleantifertility agent was first deduced in 1978 (1). Usingejaculated ra~ sperwatozoa it was shown that the compoundinhibited the reaction catalysed by glyceraldehyde phosphatedehydrogenase thereby preventing the use of fructose as a metabolic fuel for the forwation of ATP. As it is now known thatin the sperwatozoa of several species this inhibition is causedby (S)-3-chlorolactaldehyde, a metabolite of (S)-a-chlorohydrin,the effect of these compounds (2) on the metabolic activity ofejaculated ram spermatozoa has been investigated.
In Ih at 340C, O.lrnM (S)-a-chJorohydrin inhibited theoxidative metabolism of fructose (lmM) by 48%; this was increasedto 71% at 0.25rnM. An increase in the concentration of keyglycolytic intermediates (fructose-l,6-bisphosphate and thetriosephosphates) and a decrease in that of lactate confirmedthe action involved inhibition of glyceraldehyde phosphatedehydrogenase. At 10rnM, (R)-a-chlorohydrin had no effect on theoxidative metabolism of fructose nor on the concentration of thekey glycolytic intermediates. In the presence of O.lrnM (S)-achlorohydrin, the oxidation of fructose was unaffected until theincubation had proceeded for 20 win. Thereafter, steadyinhibition occurred reaching 60% of control values after Ih. Asthe degree of inhibition increased so did the concentration ofthe key glycolytic intermediates while that of lactate wasconcomitantly reduced. In the presence of 5rnM (R,S)-3-chlorolactaldehyde the oxidative metabolism of fructose was inhibitedimmediately to the extent of 40-50% of control values after5 min.
While these results are consistent with the specificoxidation of (S)-a-chlorohydrin to (S)-3-chlorolactaldehyde in~itu, one result appears to be at variance with those obtainedpreviously. In the incubations of the spermatozoa of severalother species, the presence of glycerol suppresses the productionof (S)-3-chlorolactaldehyde thereby eliwinating glycolytic inhibition but with ram spermatozoa, up to 50rnM glycerol did not haveany "appdrent" effect on the oxidation of fructose. However, ahigh amount of glycerol kinase activity in ram spermatozoa (3)together with the rapid conversion of glycerol to carbon dioxide(43 nmoles/h/mg protein) indicates that the "apparent" ineffectiveness of glycerol in preventing the formation of (S)-3-chlorolactaldehyde is a reflection of the increase in utilisation ofglycerol at the expense of fructose.
1. Brown-Woodman, P.D.C., Mohri, H., Mohri, T., Suter, D. andWhite, I.G. Biochem. J. 170: 23-37 (1978)
2. Jones, A.R. Aust. J. Bio~Sci. 36: 333-50 (1983)3. Mohri, H. and Masaki, J. J. Reprod. Fertil. 14: 179-94 (1967)
r!II
61
INHIBITION OF GLYCERALDEHYDE PHOSPHATE DEHYDROGENASE IN BOARSPERMATOZOA BY (S)-3-CHLOROLACTALDEHYDE
D. Stevenson, G.F. King and A.R. Jones
Department of Biochemistry, The University of Sydney,Sydney, N.S.W. 2006
(S)-a-Chlorohydrin is an effective antifert~lity agent inthe male of a number of species. This re~po~se ~nvolves.a.
specific two-stage process which occurs w~th~n mature e~~d~dymal
spermatozoa; firstly, the oxidation of (~)-~-~h~orohydr~~ to(S)-3-chlorolactaldehyde and, secondly, ~nh~blt~on by th~s .metabolite of glyceraldehyde phosphate dehydrogenase (l~. W~l~e
the spe~ies-specificityof (S)-a-chlorohydrin as an ant~fert~l~tyagent can be correlated with the ability of s?er~a~o~oa toperform th~ former process, the mechanism of lnh~b~t~on ofspermatozoal glyceraldehyde phosphate dehydrogenase by (S)-3-chlorolactaldehyde is unknown. .
The pure enzyme from rabbit muscle was weakl~ su~cept~b~e
to inhibition by (R,S)-3-chlorolactaldehyde.a~dk~net~c stud~es
have revealed that the inhibition is compet~t~ve (control Krn 102,vmax 11.2; +inhibitor Km 143, vmax 10.8, ~mo17s NADH produced/min/mg protein and ~M, respectively). Isolat~on of glycer~ldehyde phosphate dehydrogenase from boar sperwatozoa ~a7achieved by sonication in a 20mM phosphate buffer contaln~ng
5rnM mercaptoethanol. At a conce~tration o~ 0.33rnM NAD+, the _enzyme exhibited a typical sigmo~dal veloc7t~ v substrate concentration curve showing positive cooperat~v~ty tow~rds dNAD+ (2). Increasing the NAD+ to 1.5rnM overcam7 t~~~ eff7 ct anthe presence of 5rnM (R,S)-3-chlorolactaldehyde ~nhlb~ted ~ts
activity in a competitive, albeit very' weak, manne~ (control Km290 v 0 109' +inhibitor Km 380, vmax 0.116, un~ts as above).
, w~~~ b;ar ~permatozoa were incubated with 5rnM (R,S)-3chlorolactaldehyde for Ih at 340 and ~he gly~e7aldehyde phosphatedehydrogenase subsequently isolated, ~ts act~v~ty.was reduced by35%. However, when the incubation w~s.performed 7n the presenceof 0.5rnM (S)-a-chlorohydrin, the act~v~ty of the ~solated enzymewas reduced by 61%. . .
That glyceraldehyde phosphate ~eh~d70~enase w~th~n boar _spermatozoa is more susceptible to ~nhlb~t~on by (S)-3-chlorolactaldehyde than when it is extracted from the sp~rma~ozoa mayindicate that the enzyme undergoes some change dur~ng ~ts
isolation. The apparent specificity of t~e ?permatozoa~ enz~we
to inhibition in ,~itu.., however, way a.lso ~nd~cate that ~~,.l~ke
spermatozoal lactate dehydrogenase (3), exists as a spec~f~c
iso-enzyme.
1. Stevenson, D. and Jones, A.R. J. Reprod. Fertil. (in press)(1985)
2. Wang, C-S. and Alaupovic, P. Arch. Biochem. Biophys.205: 136-45 (1980)
3. Burgos, C., Gerez de Burgos, N.M., Coronel, C.E. andBlanco, A. J. Reprod. Fertil. ~: 101-6 (1979)
62
lDepartment of Animal Husbandry} University of Sydney} Sydneyand 2Animal Breeding and Research Institute} Katanning} W.A.
FERTILITY OF RAM SEMEN FROZEN-STORED FOR 16 YEARS
S. salamo~l} W.M.C. Maxwel1 2 and G. Evans 1
There was no difference between the fertility for the doses of motile spermatozoa inseminated (X~ = 1.03} N.S.). The mean lambing rate forfrozen-thawed semen (119/193; 61.63%} Test 2) was indistinguishable fromthat of fresh-diluted semen (X; = 0.35} N.S.). These tests} conductedin both W.A. and N.S.W. and using a relatively wide range of motilesperm doses} gave a reliable estimate of fertility. We conclude that theram semen was satisfactorily preserved during the 16 year storageperiod. In addition, these results indicate that acceptable fertilitycan be obtained with frozen-thawed semen using an intra-uterineinsemination dose of only 10 x 106 motile sperm.
(1) Salamon} S. Proc. 9th Int. Congo Anim. Reprod. and A.I. Madrid, 2:420-421 (1980).
(2) Killeen} I.D.} and Caffery} G.J. Aust. vet. J. 59: 95 (1982).
A
MacLeod, J. Seminal Cytology in the Presence of Varicocele.Fertil. Steril. 16 (6), 735-757 (1965).Dubin L. & Amelar, R.D. Varicocele size and results ofvaric~celectomy in selected subfertile men with varicocele.Fertil. Steril. 21 (8), 606-609 (1970).Brown, J.S. Varicocelectomy in the subfertile male: A ten-yearexperience with 295 cases. Fertil. Steril. 27 (9), 1046-1053(1976) .
( 3)
(2)
(1)
TABLE 1. Sperm motility and morphology (mean and range) in infertilemen with and without varicocele.
VARICOCELE CONTROL p*Motility (%) 44 34 0.05
(14 - 73) (11 - 80)
Normal Morphology (%) 52 54 NS(18 - 88) (12 - 86)
Tapering (%) 1.0 0.9 NS(0 - 9) (0 - 6)
Immature (%) 3.1 7.8 0.05(0 - 36) (0 - 70)
* Wilcoxon test; NS - not significant
Control men without varicoceles had significantly lower spermmotility and higher proportions of immature forms than men w~th
varicoceles (Table 1). No significant difference was found 1n proportions of normal oval shaped and taperi~g sperm betwee~ t~e ~roups
with and without varicocele. We do not f1nd a character1st1c stresspattern' of sperm morphology in association with varicoceles.
IS THERE A SPECIFIC ABNORMALITY OF SPERM MORPHOLOGYIN MEN WITH VARICOCELES?
63
O. Ayodeji and H.W.G. Baker
Medical Research Centre Prince Henry's Hospital and Howard FloreyInstitute of Experim~ntal Physiology and Medicine, Melbourne
MacLeodl described a characteristic 'stress pattern' of spermmorphology in men with varicoceles; increas~ taper~ng and. immatureforms. Others believe the 'stress pattern' 1S of d1agnost1c and prog-nostic significance (2,3). . .
To assess the significance of this finding in our 1nfert11e.population, a retrospective study of sperm morphology has been ~arr1ed out.
Fifty men with large left varicoceles and 50 controls w1thGutvaricoceles were matched for age (mean 30, range 23-42 years), lefttesticular volume (19, 10-25 ml), serum FSH level (4:0~ 0.9-19.I.U./l)and sperm concentration (45, 1-392 milli~n/ml). Ind~v1duals w1thknown causes for defective sperm product10n or funct10n such as undescended testes, and sperm autoimmunity were excluded.
r!I
II
26(66.7)
40(51.9)*33(66.0)28(58.3)32(64.0)26(57.8)
eweslambed (%)
39
7750485045
insemination
No. ofinseminated
80402010
40
No. motile sperm/inseminate (x 106 )
slaughter 100 days after( 40 Fresh (Control)
TestNo.
(W.A. )
2 (N.S.W.)
*Pregnant at
Preservation of fertility of semen during a prolonged period ofstorage is of practical importance. To examine the fertility of ramsemen after long-term storage} a semen bank was laid down and fertilitytests were conducted after 3} 5} 7 and 11 years of storage (1). Thiscommunication presents fertility data following insemination of eweswith semen stored for 16 years. Two tests were conducted} the first inWestern Australia (February} 1984) and the second in N.S.W. (May} 1984).
The semen was processed and frozen in pellet form as describedearlier (1). For intra-uterine insemination} the frozen pellets werethawed in dry test tubes held in a water bath at 37oC. In Test 1 theewes received 40 x 106 motile spermatozoa in a 0.08 ml inseminate (0.04ml/uterine horn). The sperm doses used in Test 2 were achieved bydiluting the frozen-thawed and freshly collected semen (control) withinositol (210 mM)-sodium citrate (40 mM) solution to cell concentrationswhich provided the required number of motile spermatozoa in a 0.16 mlinseminate (0.08 ml/uterine horn).
Mature Merino ewes were treated with intravaginal sponges containing 40 mg Cronolone (Searle) for 12 days} and 400 Lu. PMSG (Folligon}Intervet) at sponge withdrawal. Intra-uterine inseminations with theaid of a laparoscope (2) were performed 60-64 h after sponge removal.The results are presented in Table 1.
TABLE 1. Fertility after intra-uterine insemination with semen storedfor 16 years.

6465
SPERM MOTILITY AND MORPHOLOGY IN SPERMATIC GRANULOMAOF THE EPIDIDYMAL HEAD (SPERMIOSTASIS)
D.B. Galloway and Zhang Zhong ChengDepartment of Veterinary Clinical Sciences, University of Melbourne,
Princes Highway, werribee 3030
(1) Blom, E. & Christensen, N.O. Nord. Vet-Med 12: 453-470 (1960).(2) Rao, A.R., Bane, A. & Gustafsson, B.K. Theriogen. 1!: 1-11 (1980).
Naturally occurring cases and the experimental rams showed that adisturbance of epididymal funtion in ruminants can cause reducedmotility and elevations of sperm with mp-t abnormalities. Failure ofproper maturation of the sperm appears to precede degenerativechanges. Observations of an increase in attached cd'~ associated withepididymal dysfunction add to previous work on the origin of spermdefects (2).
In 2 rams A and B experimental spermiostasis was established byligating some efferent tubules. The tissue included in the ligaturewas 2-3 mm in diameter. Spermatic granulomas developed at 3 of the 4sites. A control ram C was sham operated. Semen was collected byelectroejaculation on 3 occasions before the operation and 3 times inweeks 2, 3, 4 and 5 afterwards. For each week means of the totalmidpiece-tail (mp-t) abnormalities in the semen of rams A a~d B werecompared with the means for ram C. Significant increases in thepercentages of mp-t abnormalities were observed in ram A in weeks 4and 5 (27.8 ± 4.7 and 70.0 ± 13.6 respectively cf 6.0 ± 2.5 and 6.6 ±2.3; P < 0.01) and in ram B in weeks 3 and 4 (25.6 ± 2.7 and 26.6 ±10.2 respectively cf 4.9 ± 2.4 and 6.0 ± 2.5; P < 0.01 and P < 0.05).Morphological abnormalities contributing to these increases wereprincipally dd's (to 16% in A, 10% in B), th's (to 32% in A, 57% inB), sbt's (to 22% in A, 4% in B) and dbt's (to 53% in A, 10% in B).For ram C in weeks 2-5 the percentages were (upper limit of the normalrange in parenthesis): dd's 0-4% (4%), th's 0.5-11% (10%), sbt's 0-5%(8%) and dbt's 0-1% (3%). Earliest changes in the semen were anincrease in dd's in both rams in week 3. Motility fell in weeks 4 (A)and 2 (B). A granulomatous reaction, bilateral in A and unilateral inB was palpable by week 3.
In 24 ruminants (4 goats, 11 rams, 9 bulls) with spermiostasis(l)semen was examined at the time of clinical diagnosis. Changes arereported relative to semen laboratory standards. In 8 unilateralcases semen motility and morphology were normal. Four bilateral caseswere aspermic and 3 nearly so, with few immotile sperm or taillessheads present. In 6 bilateral cases motility was low. There werecombinations of increased numbers of sperm with proximal or distalcytoplasmic droplets (cd's), tailless heads (th's) and singly benttails (sbt's). Increased th's was the only change in 3 unilateralcases. Two of the bilaterally affected bulls, confirmed post mortemas early cases, had very low motility and consistently increasedproximal droplets (pd's) in one (all of 33 samples, 4-49%) and distaldroplets (dd's) in the other (9 of 10 samples, 4-52%).
9 ± 9
18 ± 11
61 ± 36*
36 ± 22
102 ± 75
136 ± 52*
8 ± 9
17 ± 11
28 ± 12*
37 ± 27
47 ± 19*
23 ± 18
IN VIVO AND IN VITRO EFFECTS OF HlJKA.N CHORIONIC GONADOTROPIN (heG) ON
11IE I«JTILITY OF DtHA11JRE RABBIT SPERMATOZOAE. Herrera Castaneda and P.o. Temple-Smith
D~pa:tment of Anatomy. Monash University. Clayton. Victoria 3168VarIOus ag~nt,s. including hCG. have been shown to stimulate and
i~prove ,the motIlIty of mammalian spermatozoa. In normal rabbits aSIngle Intravenous dose of hCG enhanced sperm motility in eiaculate!"collec:ed 48 hr later (1) but the cause of tbis effect - was no~determIned. This study examined the direct and indirect effects of hCGo~ the motility of immature spermatozoa from three rabbits withbIlateral vasoepididymal bypasses at the level of thepididymidis. e caput
, ,For in vivo studies. each rabbit was given control iniections ofdIstIlled water on Day 0 (DO) and D20, and 100 IU hCG on Dio and 03:).Blood samples were collected at O. 3, 6. 12. 24. 48 and 72 hr aflpr 00and, ~10 injections and testosterone (T) levels were asses~ed bvradlolm~iltlno~s~ay.. Semen samples were collected from each animal 48 h~after each In:ectlon. Initial motility (7. and index) was determined andeac~ sample was then prepared for in vitro examination (2). Sperm~otIlit~ of eac~ sam~le wa~ assessed after I, 3. 6, 12 and 24 hrIncubatIon. a: 37 C. DIrect In vitro effects of 1, 10 and IDa IU hCG onsperm motIlIty were examined using similar sperm preparation dassessmc:t p:ocedures, and these were compared with sperm motil~~vobservatIons In corresponding controls. -
In vivo studies showed that the initial motility of eiaculatedsperm taken 48 hr after hCG treatment was significantly higher than thof sperm from contr~l ejaculates. Sperm from hCG-treated animals we:~also able, to sustal~ motility (7. and index) at significantly higherlevels durIng 24 hr Incubation in vitro than controls (see table)Table: Effect of in vivo treatment with beG on sperm motility in· vitro
I Motility IndexInitial (0 hr) Final (24 hr) Initial (0 hr) Final (24 hr)Day 0
(control)Day 10(hCG)Day 20(control)Day 30
~hCG? o. 49 ± 26* 26 ± 17* 130 ± 70* 38 ± 28*, SIgnIfIcantly higher (p < 0.05) than corresponding control.
however no signific~nt differences in sperm motility were observedwhen control sperm were Incubated for 24 hr i dO.,
100 IU hC n me 1um contaInIng 1 10or '.0 G suggesting that hCG has no direct effect on s'ermmotILIty. After heG treatment in vivo T lev lIPsig Of' 1 b • e s were e evated
TIl lcant y etween 6 and 72 hr after hCG treatment (p < 0 05) hcompared with control levels. We conclude that the h· d w ensustained '1 0 en ance and
o spe:m mot I 1ty seen after in vivo hCG treatment is caused~lt~er by a dIrect ~ffect of T on sperm or, perhaps more likelv bv'an1nd1rect effect of Increased plasma T on the epididymi 1" '. - .changes in the 1 . 1 0 s resu tlng 1n
. um1na enV1ronment which Support the development of thcapaCIty for sperm motility. e0) Ogawa et aI, 1972, Nature, 238: 270.(2) Hosoi et aI, 1981, ~Reprod., 24: 637

66
THE MOTILITY OF EPIDIDYMAL SPERMATOZOA IN THE BULL
C. Pholpramool*, J.L. Zupp, B.P. Setchell
Department of Animal Sciences, Waite Agricultural Research InstituteUniversity of Adelaide, Glen Osmond SA 5064; *Present address: 'Department of Physiology, Mahidol University, Bangkok, Thailand.
It is believed that most mammalian spermatozoa acquire the potentialfor motility during their passage through the epididymis but themotility of undiluted spermatozoa from the cauda epididymidis in anumber of species is still debatable. Contradictory results have beenreported for the bull. Other investigators have reported negligible ormoderate motility with no progressive movement, but in theseinvestigations, motility of spermatozoa may have been underestimatedbec~use spermatozoa were restricted with no access to oxygen, in glasscap~llary tubes or under cover glasses with no precautions to preventst~c~ing of the spermatozoa to the glass. In the present study, bullep~d~d~al spermatozoa were collected by micropuncture or by retrogradeperfus~on of the ductus deferens and their motility examined in adroplet of undiluted semen under paraffin oil by the method describedpreviously by Turner & Howards (1) and Turner & Giles (2).
. ~nder th:se co~ditions, bull caudal spermatozoa showed vigorousmot~l~:y: T~~s mot~lity was less in samples collected from further upthe ep~d~dym~s, samples from the distal caput showed weak butdetectable motility while those from the proximal and mid-caput werecomplete~y quiescent. Motility of spermatozoa from the distal caputand prox~mal corpus was markedly increased after incubation at 34° or37° C for 1 h, but was depressed by incubation at 25° C. Similar butsmaller effects were observed with spermatozoa collected from the midc~rpus and the mid cauda, except that motility of spermatozoa from them~d corpus was reduced after 1 h incubation at 37° C. The inhibitoryeffect of low temperature was completely reversible. Incubation ofcaudal spermatozoa under anaerobic conditions produced only partial andreversible inhibition of motility. The results suggest that bullepididymal spermatozoa may not be completely quiescent in their nativeenvironment as previously assumed. A video tape will be provided toillustrate these findings.Supported by the J.S. Davies Beef Cattle Bequest.
(1) Turner, T.T. and Howards, S.S. BioI. Reprod. 18: 571-578 (1978)(2) Turner, T.T. and Giles R.D. Gamete Res. i: 283-295 (1981)
67
PURIFICATION AND PROPERTIES OF A 22-kDa PROTEIN PRESENT IN TESTICULARAND EPIDIDYMAL FLUIDS AND ON THE SURFACE OF SPERMATOZOA
D.E. Brooks
Department of Animal Sciences, University of Adelaide, WaiteAgricultural Research Institute, Glen Osmond, S.A.
A 22-kDa protein has been purified from rat epididymal plasma usinga 'Fast Protein Liquid Chromatography' system (Pharmacia Pty Ltd).Polyclonal antibodies against the protein were isolated by affinitychromatography on a column containing the 22-kDa protein coupled toSepharose.
Western blotting procedures established that the protein isuniquely present in testicular and epididymal luminal fluids, but notin a variety of other extracellular secretions (blood, peritonealfluid, saliva, milk, uterine fluid, and secretions from the prostate,seminal vesicle and coagulating gland). Nevertheless, the protein wasfound to exist in all tissues examined as a cytosolic protein. Directsynthesis within particular tissues was established by in vitroincubation with radioactive methionine and separation of the resultantradioactive proteins on two-dimensional polyacrylamide gels .
The 22-kDa protein was detected as a surface component ofspermatozoa by indirect immunofluorescence microscopy and by surfa:eiodination procedures. The protein is the major surface component ofsperm containing sulphydryl groups and disulphide bonds. Theseproperties may be relevant to the maintenance of sperm viability duringepididymal transit and during capacitation.

68 69
Cells+ hCG380.4300.1262.5216.5106.2
TESTICULAR. LEYDIG CELLIS THERE A PARACRIHE REGULATOR. OFSTEROIDOGENESIS?
G.P. Risbridger, J.A. Muir, G. Jenkin and D.M. de Kretser
Department of Anatomy, Monash University, Clayton, V~ctoria 3168
The presence of a paracrine regulatory interaction between theseminiferous tubules and Leydig cells of the testis has been extensivelydocumented. However there is little agreement as to whether or notlocal factors act to inhibit or stimulate, basal or gonadotrophinstimulated testosterone (T) production by crude or purified Leydig cellpreparations in vitro. This study investigates the presence of a factorin rat testicular interstitial fluid (IF) which affects T production andits site of action on the steroidogenic pathway. Testicular rat IF wascollected a~d treated with charcoal to remove endogenous steroids.Crude Leydig cell preparations were prepared by collagenase dispersionof adult rat testes and purified preparations obtained by centrifugationthrough discontinuous Percoll gradients.
IF consistently stimulated both basal and gonadotrophin-stimulqtedT production by both crude and purified rat Leydig cells (Table 1). Thestimulatory activity was not due to non-specific factors which improvedthe responsiveness of Leydig cells, as the addition of increasingvolumes of thoracic duct lymph did not affect T production. Furthermorein the presence of serum (collected under the same conditions as IF) thestimulation activity was unaltered (p > 0.05).
To investigate the site of action of IF on steroid biosynthesis theeffects of IF were determined in the presence or absence of inhibitorsof 3 [3 hydroxysteroid dehydrogenase and 17 a hydroxylase enzymes. Theeffects of hCG and hCG + IF on T and pregnenolone (P) production weresimilar to previous observations. In the presence of the inhibitors, Tproduction was suppressed and hCG stimulated P production; the additionof IF caused a further significant increase in P production (Table 2).
These data therefore demonstrate the presence of a factor in IFwhich is not pituitary LH and is capable of stimulating Leydig cellsteroidogenesis prior to the production of pregnenolone.
Table 1: T Production - ng/106 cells/20 hBy Crude Cells By Purified
Vol IF (lll) + Medium + hCG + Medium25.0 5.29 26.19 59.412.5 4.99 23.52 44.76.2 4.25 18.48 43.83.1 3.55 15.29 30.30.0 2.26 6.75 11.0
Mean of triplicate determinations, SE omitted for clarity
Table 2: Sample T Production (ng) P Productionmedium 220.9 ± 32.1 0.5 ± 0.2
medium + hCG 437.4 ± 17.0 0.7 ± 0.1Medium + inhibitors 3.1 ± 1.1 28.5 ± 1.8
medium + inhibitors + hCG 2.3 ± 0.9 60.9 ± 13.1medium + IF + hCG 1580 ± 62.3 41.7 ± 2.5
medium + IF + inhibitors + hCG 7.6 ± 3.9 1213.9 ± 151.7Mean ± SE (n = 3)
THE EFFECTS OF ETHYLENE DIHETHANE SULPHONATE ON ADULT RAT LEYDIG CELLSA.E. JRckson, P.C. O'Leary, M.M. Ayres and D.M. de Kretser
Department of Anatomy, Monash University, Clayton Victoria 3168Ethylene dimethRne sulphonate (EDS) is a diester of methane
sulphonic acid which has a reversible effect on the fertility of malerats. This study investigated the specific effect on adult rRt Leydigcells, by morphological and biochemical parameters.
Adult male Sprague-Dawley rats were given a single ip injection ofEDS (75 mg/kg) in rnso as vehicle. Control animals were given DMSOalone. Rats (10 per group) were killed at 2, 6, 12, 18 and 24 hours, 3,7,14,21,28,35 and 49 days after in;ection for serum testosterone(T~, FSH and LH levels and hCG binding to testicular homogenates. Threeam.mals per group were perfused with 5% glutaraldehyde and 4%formalde~Yde and testes prepared for light and electron microscopy andthe sectlons used for morphometric analysis of the intertubular tissue.
Testosterone levels were significantly less than DMSO controls (3.3± 0.3 ng/ml) 48 hrs after EDS (0.3 ± 0.1 ng/ml), remained low from 2-14days, returning to normal levels by 21 days. Serum LH levels showed theinverse pattern to testosterone, being elevated at 48 hrs (5.1 ± 1ng/ml; control 1.1 ± 0.1 ng/ml), reaching a peak at 14 days (6.6 ± 0.8ng/ml) and returning to control levels at 28 days. FSH levels wereelevated at 48 hrs (7.5 ± 0.8 ng/ml; control 4.1 ± 0.3 ng/ml) andreturned more slowly to normal at 49 days.
.Light and electron microscopic examination showed marked changes inLeyd~g. cells ~t 24 hours post EDS, with vacuolated cytoplasm andpyknot1C nucle~. Morphometric analysis (Table 1) showed a loss ofLe!dig cell. volume b~t a significant increase in macrophages, many ofWh1Ch contalned Leyd1g cell cytoplasm in lysosomes. No Leydig cellswere detectable by 3 days. From 14 days post EDS there wasreduplication of nuclei of connective tissue cells along the margins ofthe tubules and around the capillaries. "By 21 days these cells haddev~loped features of foetal type Leydig cells with smooth endoplasmicretlculum, mitochondria with tubular cristae and lipid droplets. Thevo:ume ~f Leydig cells increased to reach normal levels by 49 days, bywh~ch tlme they showed the features of normal Leydig cells with abundantsmooth endoplasmic reticulum and mitochondria with lamellar cristae.Parallel hCG binding studies showed that binding was not detectahle by 7days (control 75 ± 19 moles x 10-15 per testis) and gradually recoveredto normal levels by 49 days.
These s:udies i~dicate that EDS destroys Leydig cells and recoveryof T levels ~s as~oclated wit~ restoration of the Leydig cell populationfrom connectlve tlssue cells 1n the intertubular area.
Table: Changes In The Absolute Volume Of Cells In The IntertubularTissue After EnS (microlitres ± SEM)
Days After EnSo 1 7 21 49
40.S1 a±5.4 17.72±2.32 0 lS.02±4.38 32.1Sa±7.36.49±1.03 14.29b±2.1S 4.97±6.22 5.44±1.07 6.15±1.01
Leydig CellsMacrophagesConnectiveTissue Cells 10.40±O.48 14.00±O.86 12.43±0.99 16.73c±3.06 12.09±0.91
asig. dif. from all time points p(O.Ol; bsig • diE. [rom controlsp(O.Ol; Csig. dif. from controls p(O.05

70 71
DEVELOPMENT IN BULL CALVES. ACTIVELY IHMUNIZED AGAINSTPUBERTAL TESTOSTERONE AND OESTRADIOL-17~
42.5 6 4.1 3.1C 7 0.7 14.7
6 3.7 14.415.9 48.2Anti-T 7 1.16 5.3 11.30.8 14.5 44.0Anti-OE z 9
2.0 0.8 4.2Pooled SEM 0.1 0.4
J. Endocr.
Weatherly"· ,
Responses to LHRHllLH L'lTn
and Kinder, J.E.
n
Schanbacher, B.Do, D'Occhio, MoJ.93: 183-192 (1982).
(1)
the testes in prepubertal bulls ish d maturation of dGrowt an . ort (1). The present study examine
dependent on gonadotrophln supp 0 d 1 rated testiculard LH and FSH secretIon, an acce e 0 0 •
whether increase. . oun bulls by active immunIzatIon agalnstgrowth, could be achle~e~ ln y . g ative feedback regulation of
'd h'ch partlclpate ln neg . ysterOl s ~ 1 . Shorthorn bull calves received a .pnrnargonadotrophln secretlon. (O_T' T-human serum albumln (HSA)immunization against testos~eroned oAnt~t)' or oestradiol-17~ (Anti-OE
z;
conj uga te in Freund t s ( co)mp;t: : ;~;abooster inj ections at two monthly
OEz-HSA) at 3 months (mo) 0 .g d HSA emulsified in adjuvant. Bloodintervals. Controls C recelve Ot serum LH FSH and T levels. Body
t ken weekly to monl or , . 0 dsamples were a . were measured monthly. Pl tUl tary anweights and testes dlameters HRH (5 /k BW' i. v. bolus) were
to exogenous L ng g , d ttestes responses All hormone data were transforme 0
ascertained at 10 ~o of o ageANOVA Results for mean serum LH and FSH10g10 before analysls uSIng 10' ~estes diameters at 10 mo, andlevels (ng/ml) from 3 to ~o,
LHRH are presented 1n the Table.responses to, . T t"Gonadotrophlns es lS
LH FSH (mm)
b d responses in Anti-T (717 ± 231, reciprocalThe secondary anti 0 y (12,205 ± 4,366) bull calves indicated
antlObody titre) and Anti-EOz 0 T b lIs had highero in both groups. Antl- u. 0
that immunlty was achieved h A to-OE and C bulls dunng the penod(P <O. 05) LH and FSH levels t an n 1 Z t (P<O 05) than in C
o 0 0 A ti-T bulls was grea er. h3 to 10 mo. Testls Slze ln n t LHRH were similar for t ebulls from 6 rno onwards. LH [f:~sponses A 0 0 -OE bulls showed a greaterthree groups' however, both Antl-T and d nt.l
th~ bulls. These results
' ponse compare Wl 1 0
(P<0.05) testost:roneo re~ bull calves against T resu ts lnindicate that lmmunl~atlon o~on and accelerated testicular growt:-h.increased gonadotrophln se~ret h s which participate in negatlveImmunity early in life agaln~t ormone h to influencing both pubertal
Ode a practIcal approacfeedback may provl d t've performance in bulls~maturation and adult repro uc I
D'Occhio*, D.R. Giffordt, R.M. Hoskinsontt, T~M.J. P.F. Flavelt, P.E. Mattnertt and B.P. Se~chelln
o . Waite Agricultural Research Institute,'·Department of Anlmal sClences'D t t of Agriculture Turretfield
d S A 5064' tSA epar men '. 1Glen Osmon, .. , 's A 5350 and ttCSlRO Division of All1.maResearch Centre, Rosedale, ., , N S W 2149
Production, Prospect, .,. , .
THE UNILATERAL CRYPTORCHID RAT foI>DEL; EVIDENCE FOR LOCAL AS WELL ASHUMORAL CONTROL OF LEYDIG CELL RECOVERY FOLLOWING EDS ADMINISTRATION
P. O'Leary, A.E. Jackson, G.P. Risbridger and D.M. de Kretser
Department of Anatomy, Monash University, Clayton, Victoria 3168
J<:DS has been shown to specifically destroy Leydig cells within 24to 48 hours i.n testes from bilaterally cryptorchid and normal rats.There follows a period of low serum T, decreased testicular hCG-hindingand elevated gonadotrophins, which return to control levels as Leydigcells differenti!lte and repopulate the interstitial space. While therecovery phase occurs over a period of approximately 49 days in thenormal rCit, it is reduced to 28 days in the bilaterally cryptorchidrat. The unilaterally cryptorchid rat model provides an uniqueopportunity to study the response of normal and seminiferous tubuledamaged testes which are simultaneously exposed to the same circulatinglevels of hormones. It has previously been found (1) that there is nosignifiCA."t increase in serum LH and FSH levels until 4 weeks afterinductio,; ()f uni lateral cryptorchidism in adult rats. Therefore 2 weeksafter surgery. ~ive groups of rats were in;ected with either EDS (n = 7)or DMSO vehicle, (n = 7) and sacrificed 3, 7, 14, 21, 28 days later.Blood was collected for T, LH and FSH radioimmunoassay, and the testesprocessed for hCG-binding analysis.
Serum T levels decreased from 3.0 ± 0.6 ng/ml (mean ± SEM) to 0.1ng/ml by ~ days after EOS and remained significantly lower than controlsuntil 14 days. Serum FSH and LH levels were elevated during this periodbut had returned to control levels by 28 days after EOS. In vitro hCGbinding to test icular homogenates showed a different recovery patternbetween the abdominal and scrotal testes after EDS administration.Scrotal testicular hCG binding returned to 32 ± 5% of corresponding DMSOcontrols at 28 days, but hCG binding to abdominal testes was 37 ± 7% ofOMSO controls by 14 days after EDS. Although the hCG-binding did notattain uMSO-control levels within the 28 day study period, we concludethat the recovery of hCG-binding and therefore Leydig cells is due to acombination of local and humoral factors. The accelerated recovery ofthe abdominal testis hCG-binding compared to the scrotal testis,suggests a local mechanism is operating to stimulate Leydig celldifferentiation. The fact that neither scrotal nor abdominal t~sticularhCG binding recovered entirely by 28 days, implies that elevatedgonadotrophins, as found in the bilaterally cryptorchid rat, are alsorequired to complete the recovery of testicular Leydig cells folloWingEDS treatment.
Table 1: beG-binding (aol x 10-15 bound/testis) to scrotaland abdominal testes following a single injection of EDS
Scrotal Testis Abdominal TestisDay EDS DMSO % Control EDS DMSO % Control3 3.1 ± 1.0 152 ± 9 2.0 ± 0.6 0 21 ± 2.2 07 3.0 ± 0.2 186 ± 11 1.6 ± 0.1 3.0 ± 0.5 33 ± 12.0 9.1 ± 1. C;14 67 ± 22 113 ± 10 5.9 ± 1.9 11.0 ± 5.8 30 ± 8.4 37.0 ± 7.321 12.6 ± 2 166 ± 9 7.6 ± 0.5 9.0 ± 2.4 16 ± 1.3 56.0 ± 15.028 59 ± 9 185 ± 5 32.0 ± 5.0 12.0 ± 0.5 27 ± 2.0 44.0 ± 1.8Values represent mean ± SEM (n = 7)
(1) Risbridger,Reprod 24:
G. P., Ke r r , J. B.534-540.
and de Kretser, D.M. (1981 ) BioIf the J S Davies Bequest.Study supported in part by a grant rom ..

73
J. Endocr.and Kinder, J.E.
Day 4 (3) 5 (4) 6 (4) 7 (3) 8 (3) 9 (4) Pooled SEM
Schanbacher, B.D., D'Occhio, M.J.93: 183-192 (1982).
(1)
0.49 0.52 0.37 0.31 0.38 0.18serum LH -3 0.921.34 1.45 0.323 2.43 1.72 1.49 1.852.6 2.7 0.44.3 4.0 4.0 3.6LH pulses 3
20.3 2.718.5 19.6 24.5 22.0serum FSH -3 20.747.2 41.2 5.93 55.1 43.4 45.3 56.1
ACUTE GONADOTROPHIN RESPONSES TO CASTRATION IN PREPUBERTALAND PUBERTAL BULLS
D.R. Gifford~'I', M.J. D'Occhiot, T. Weatherlyt and B.P. Setchellt
~S A Department of Agriculture, Turretfield Research Centre, Rosedalel,
~ . . An' 1 S . Waite AgriculturaS.A., 5350 and tDepartment of ~ma C1ences,Research Institute, Glen Osmond, S.A., 5064.
The transient rise in gonadotrophin secretion which occurs i~ bull1 es during the first six months of life is thought to prov~de an
cadv. . nal which initiates maturation of the immature testes (1).en ocr~ne S1g h . c'ty of theThe present study examined the concept that c anges ~n capa 1 d .hypothalamo-pituitary unit for hormone sy~thesi~ and r:leasfe d~r1~g
development may occur independe~tly ~f sh~fts 1n net:t~:~ F~~ :~r~Accordin ly acute post-castrat~on r1ses 1n serum .determin~d in crossbred bull calves castrated at month~y ~nterv~ls from4 to 9 months (mo) of age. Patterns o.f g.onadotroph1n secret10n werecharacterized by bleeding bulls at 20 m1n ~ntervais ~or ~ ~ ~:r~:y~a~sbefore castration (day -3), and at 10 min 1nterva sd or'
ngRIA and ali
5 and 7 after castration. LH and FSH were measure US1 fhormone data were transformed to 10g10 before ANOVA. R~sUltsb oro~e~serum LH and FSH levels (ng/m~) on days -3 and 3, an num erpulses on day 3, are presented ~n the Table.
Months of Age (number of animals)
. LH levels were higher (P<0.05) in 4 mo oldBefore cast~at10n mean and 9 mo. On day 3 after castration,
bulls compared w1th bulls at 7(p~0 05) number of LH pulses than did 84 rna bulls showed a greater . . ulsatile LH release, there
11 D P 'te differences 1n pand 9 mo bu s. es 1 . LH levels on days 3, 5 or 7.were no differences with age ~n. mean FSH levels with age, either
1 d . fferences 1n mean .There were a so no 1 Its confirm the transient r1sebefore or aft~r castration i The;: r~:~a also indicate that rhythmicin serum LH 1n young bul s. e, t' of castrates is established as
. t' f LH character1s 1C h .pulsat11e secre 10n a 1 f dal steroid feedback bot 1nd fter remova a gona LH
early as three ays alb 11 Age differences in frequency ofprepubertal and puberta u s't t' on suggest that, during earlypulses three days a~ter c~s. tra 1 f the hypothalamic LHRH pulse
t h ges ~n act1V1 y 0developmen, c an . d dently of feedback mechanisms.generator may occur 1n epen
b grant from the J.S. Davies Bequest.Study supported in part y aBio 1
Volume Vith Time After Kl)S ( 1 ± SEM)1 Day 7 Days 21 Days 28 days1.15±O.36 O.14±0.08 9.43±3.46 19.01±1.6
THE EFFECTS OF ETHYLENE DIMETHANE SULPHONATE (EDS) ON THEBILATERALLY CRYPTORCHID RAT TESTIS
D.M. de Kretser, A.E. Jackson, P.C. O'Leary and S.A. AverillDepartment of Anatomy, Monash University, Clayton, Victoria 3168
EDS has a specific destructive effect on adult rat Leydig cellwhich are removed from the intertubular area by macrophages. This isassociated with a decrease in testosterone (T) and a rise in serum FSHand LH for 21-28 days following EDS injection. Recovery of Leydig cellsoccurs from connective tissue cells and is complete by 49 days (1). Ourprevious studies (2) have suggested that local factors produced by theseminiferous tubules may influence Leydig cell function and we have usedcryptorchid rats to investigate the effects of such factors on therestoration of the Leydig cell population after EDS.
Adult male Sprague Dawley rats 4 weeks after the induction ofcryptorchidism, were injected with EDS ip (75 mg/kg) and killed 1, 3, 7,14, 21 and 28 days later. Serum Testosterone (T), FSH and LH weremeasured and the hCG binding capacity was determined in testishomogenates. The testes from three rats in each group were perfusedwith 5% glutaraldehyde and 4% formaldehyde and processed for light andelectron microscopy, the sections being used for morphometric analysis.
Morphometric analysis of light microscopy sections demonstratedthat Leydig cell destruction was advanced by 24 hours. Macrophages wereenlarged and contained Leydig cell cytoplasm. Leydig cells wereundetectable after 24 hours until 7 days when a new population of cellsappeared around blood vessels and along the margins of the tubules(Table 1). Houever, the volume of Leydig cell cytoplasm was still only50i. of control levels by 28 days. Results indicated a significant dropin T levels 24 hours after injection of EDS (0.28 ± 0.07 ng/ml) comparedto DMSO controls (2.8 ± 0.45 ng/ml) but began to recover by 7 daysreaching levels indistinguishable from normal by 14 days (2.08 ± 0.4ng/ml). Serum LH and FSH levels rose dramatically after 24 hrs reachinga peak at 3 days (7.5 ± 0.6 and 16 ± 0.9 ng/ml respectively) compared toDMSO controls (2.6 ± 0.3 and 9.0 ± 0.5 ng/ml respectively). By 14 days,serum LH had returned to DMSO control levels, but FSH remained elevateduntil 21 days. The hCG binding was significantly lower than DMSOconfsols (65 ± 4 moles x 10-15 per testis) at 24 hours (17 ± 3 moles x10- per testis) but gradually recovered after 14 days to return tonormal by 28 days.
These results indicate that the Leydig cell population incryptorchid testes is rapidly restored from connective tissue cells; thereasons for this rapid recovery when compared to the normal situationrequire further study.
Table 1: Changes In CellControl
Leydig Cells 35.77±1.77Connective tissue
cells 9.22±0.44 6.12±0.28 3.84±O.02 6.15±0.35 7.95±0.31Macrophages 6.24±O.25 15.67±2.67 3.89±0.03 6.33±0.64 7.14±0.861. Jackson, E.A., O'Leary, P.C., Ayres, M.M. and de Kretser, D.M.
(1985) The effects of ethylene dimethane sulphonate on adult ratLeydig cells (Abstract), ASRB, 1985.Risbridger, G.P., Kerr, J.B. and de Kretser, D.M. (1981)Reprod..3.::.: 534-540.
2.

74 75
RESPONSE OF THE EPIDIDYMIS IN RAM LAMBS TO OESTRADIOL-17~ EXTRACELLULAR MATRIX MODULATION OF PROTEOGLYCAN PRODUCTIONIN SERTOLI CELL-MYOID CELL CO-CULTURES
C. Papachristoforou*, M.J. D'Occhio*, D. Horsefallt,W. Tilleyt and B.P. Setchell* E.W. Thompson, J.S.H. Elkington and A.W. Blackshaw
*Department of Animal Sciences, Waite Agricultural Research Institute,Glen Osmond, S.A., 5064, and tDepartment of Surgery, Flinders Medical
Centre, Bedford Park, S.A., 5042.
School of Science, Griffith University, Nathan, Qld., andDepartment of Physiology and Pharmacology, University of Queensland,
st. Lucia, Qld.
We have previously reported (1) that the co-culture of Sertolicells (Sc) and myoid cells (MC) from immatur~ rats (20-22 days old) issynergistic in terms of proteoglycan product~on. We report here therelative contributions of each cell type to this synerg~sm and ~heinvolvement of the extracellular matrix (ECM) in these ~ntera7t~ons.ECM was obtained by treatment of the appropriate cell layer w~th 20mMNH~OH for 20 minutes at room temperature. .
Freshly prepared Sc (O.4ml experiment 1, 0.2m! exper~ment 2) ,wereplated onto either confluent, once passaged (s~condary) Mc (exper~ments1 2) or the corresponding Mc ECM (Mrntx, exper~ment 2). Mc werep;epared 7 days previously from 20-22 day old rats and passaged on day4 (4 x 10 5 cells replated/plate). In experiment three, freshlytrypsinised myoid cells (4 x 10 5 cells/plate) were plated onto 4 dayold cultures of Sc (O.4ml) or the corresponding Sc ECM (Smtx). In allcases production of secreted, cellular and ECM glycosaminoglycans(GAGI~) was measured over the next 3 days as previously described ~l).
Secreted GAG levels were elevated in the co-culture of establ~shedSC with freshly passaged Mc (experiment 3) or the co-culture of fewerSc with confluent Mc (experiment 2) but unaffected by co-culture of thehigher number of Sc with confluent Mc (experiment 1). Mrntx ~ad no ,effect on Sc secreted GAG (experiment 2) but Smtx caused an ~ncreas: ~n
Mc secreted GAG production (experiment 3). .co-culture of Mc and Sc enhanced the cellular GAG product~on when
the Mc were confluent (experiment 1 and 2) irrespective of Sc number,but was inhibitory in cultures of Sc with freshly passaged ~c(experiment 3). Smtx decreased production of cellular ~AG ~n Mc(experiment 3) but Mrntx had no effect on Sc GAG product~on
(experiment 2). ,Reduced production of ECM GAGs was achieved w~th the co-culture of
Sc (0.4ml) with either confluent (experiment 1) or f~eshly p~ssaged(experiment 2) Mc. This parameter was not assess~d ~~ exper~~ent 2,but smtx decreased the myoid cell ECM GAG product~on ~n exper~ment,3.
The presence of di-buturyl cyclic AMP «BU)2 cAMP, 0.5mM) dur~ng
the labelling period had no effect on these trends.These results indicate that the effect of co-culture on
proteoglycan production is sensitive.to the relative and/or absoluteproportions of both myoid and Sertol~ cells and ~e cultu:e status ofeach cell type. In terms of proteoglycan ~roduct~on, myo~d cellsappear to be modulated by sertoli cell ECM.
(1) Thompson, E.W., Elkington, J.S.H. and Blackshaw, A.W. (1984),Aust. Soc. Reprod. BioI. Abstracts.
Total1.562. 77"'~
0.27
(g)
Cauda0.871. 34"'~
0.130.10
Corpus0.350.82?'<
Epididymis
Caput0.340.61*
0.06
(g)5.35.7
0.9
Testis
n66
OE2 treatment had no effects on either the weight or histology ofthe testis, but resulted in a significant increase in epididymalweight. Segments of the epididymis showed a differential growthresponse with the greatest increase occurring in the corpus (134%)followed by the caput (79%) and cauda (54%). The increase inepididymal weight was, due mainly to hypertrophy of the fibro-muscularstroma of the duct. OE 2 had no apparent effect on androgen oroestrogen receptors in the testis or caput epididymidis. However,oestrogen receptor levels (fmol/mg protein) in the corpus and cauda ofOE2-inj rams (50, 66 and 97, 145 respectively) tended to be twice thelevels in Crams (16, 28 and 48, 76). A similar trend was observed forandrogen receptors in the corpus and cauda of OE2-inj rams (74, 103 and170, 222) compared with Crams (44, 45 and 90, 132). These resultsindicate that the immature ram epididymis responds to OE2 by undergoingincreased growth. Since OE2 influences both androgen and oestrogenreceptors in the epididymis, normal growth and maturation of this organmay involve an interaction between androgens and oestrogens. Thereappears to be some regionality in the response of the immature ramepididymis to OE2.
Oestrogens, in addition to androgens, may participate in thephysiological control of growth and maturation of the epididymis in anumber of species. Receptors specific for oestradiol-17~ (OE2 ) havebeen demonstrated in epididymal tissues in the rat, rabbit, dog and man(1). The present study examined the effects of OE 2 on growth andhistology of the testis and epididymis in prepubertal rams.Cytoplasmic androgen and oestrogen receptor levels in the above tissuesof control and OE2 treated rams, were also determined. Three-month-oldMerino rams were used. OE2 treatment (OE 2-inj; n=6) consisted of i.m.injections of 5 mg OE2 administered on Mon, Wed and Fri. Control rams(C; n=6) received injections of vehicle alone. During week 8 oftreatment all rams were castrated under general anaesthesia. Afterweighing, the testes and epididymides of 2 rams from each group weresnap-frozen for receptor analysis, whilst tissues from the remainingrams were fixed for histological evaluation. Data were analysed byANOVA and results for tissue weights are presented in the Table (?~,
P<O.Ol).
(1) Orgebin-Crist, M.C., Eller, B.C. and Danzo, B.J. (1983) Endocr.113: 1703.
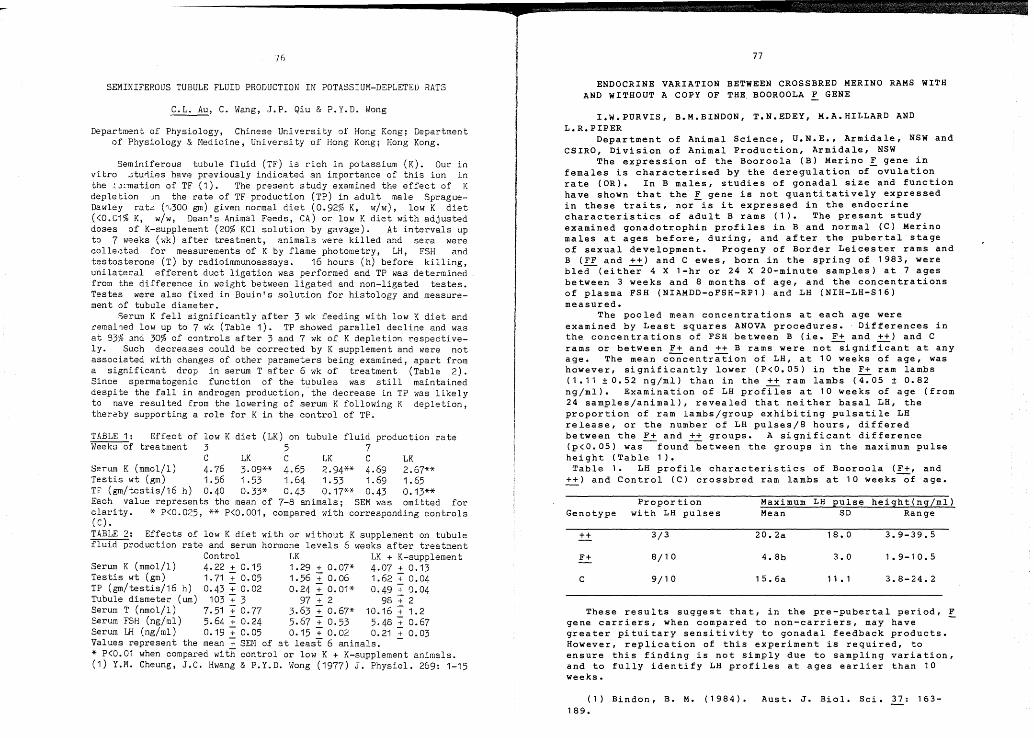
1n 77
SEMINIFEROUS TUBULE FLUID PRODUCTION IN POTASSIUM-DEPLETED RATS ENDOCRINE VARIATION BETWEEN CROSSBRED MERINO RAMS WITHAND WITHOUT A COpy OF TH~ BOOROOLA ~ GENE
C.L. Au, C. Wang, J.P. Qiu & P.Y.D. Ivang
Department of Physiology, Chinese University of Hong Kong; Departmentof Physiology & Medicine, University of Hong Kong; Hong Kong.
Proportion Maximum LH pulse height(ng/ml)Genotype with LH pulses Mean SD Range
++ 3/3 20.2a 18.0 3.9-39.5
F+ 8/10 4.8b 3.0 1.9-10.5
C 9/10 15.6a 11.1 3.8-24.2
These results suggest that, in the pre-pubertal period, Fgene carriers, when compared to non-carriers, may havegreater pituitary sensitivity to gonadal feedback products.However, replication of this experiment is required, toensure this finding is not simply due to sampling variation,and to fully identify LH profiles at ages earlier than 10weeks.
I.W.PURVIS, B.M.BINDON, T.N.EDEY, M.A. HILLARD ANDL.R.PIPER
Department of Animal Science, U.N.E., Armidale, NSW andCSIRO, Division of Animal Production, Armidale, NSW
The expression of the Booroola (B) Merino F gene infemales is characterised by the deregulation of-ovulationrate (OR). In B males, studies of gonadal size and functionhave shown that the F gene is not quantitatively expressedin these traits, nor-is it expressed in the endocrinecharacteristics of adult B rams (1). The present studyexamined gonadotrophin profiles in B and normal (C) Merinomales at ages before, during, and after the pUbertal stageof sexual development. Progeny of Border Leicester rams andB (FF and ++) and C ewes, born in the spring of 1983, wereble~(either 4 X 1-hr or 24 X 20-minute samples) at 7 agesbetween 3 weeks and 8 months of age, and the concentrationsof plasma FSH (NIAMDD-oFSH-RP1) and LH (NIH-LH-S16)measured.
The pooled mean concentrations at each age wereexamined by Least squares ANOVA procedures. Differences inthe concentrations of FSH between B (ie. ~ and ++) and Crams or between F+ and ++ B rams were not significant at anyage. The mean concentration of LH, at 10 weeks of age, washowever, significantly lower (P<0.05) in the F+ ram lambs(1.11 ± 0.52 ng/ml) than in the ++ ram lambs (4:"05 ± 0.82ng/ml). Examination of LH profiles at 10 weeks of age (from24 samples/animal), revealed that neither basal LH, theproportion of ram lambs/group exhibiting pulsatile LHrelease, or the number of LH pulses/8 hours, differedbetween the F+ and ++ groups. A significant difference(p<0.05) was--found~etween the groups in the maximum pulseheight (Table 1).Table 1. LH profile characteristics of Booroola (F+, and
++) and Control (C) crossbred ram lambs at 10 weeks-;f age.
fluid production rate7C LK4·69 2.67**1.69 1.650.43 0.13**
SEM was omitted forcorresponding controls
Seminiferous tubule fluid (TF) is rich in potassium (K). Our invitro 3tu~ies have previously indicated an importance of this ion inthe .~ J:'mation of TF (1). The present study examined the effect of Kdepletion )n the rate of TF production (TP) in ddult male SpragueDawley rat.s (,\,300 gm) given normal diet (0.92% K, w/w), low K diet«0.01% K, w/w, Dean's Animal Feeds, CA) or low K diet with adjusteddoses of K-supplement (20% KCl solution by gavage). At intervals upto 7 weeks (wk) after treatment, animals were killed and sera werecolle0tad for measurements of K by flame photometry, LH, FSH andtestosterone (T) by radioimmunoassays. 16 hours (h) before killing,unilateral efferent duct ligation was performed and TP was determinedfrom the difference in weight between ligated and non-ligated testes.Testes were also fixed in Bouin's solution for histology and measurement of tubule diameter.
Serum K fell significantly after 3 wk feeding with low K diet andremained low up to 7 wk (Table 1). TP showed parallel decline and wasat 33% anti 30% of controls after 3 and 7 wk of K depletion respectively. Such decreases could be corrected by K supplement and were notassociated with changes of other parameters being examined, apart froma significant drop in serum T after 6 wk of treatment (Table 2).Since spermatogenic function of the tubules was still maintaineddespite the fall in androgen production, the decrease in T2 was likelyto have resulted from the lowering of serum K following K depletion,thereby supporting a role for K in the control of TP.
TABLE 1: Effect of low K diet (LK) on tubuleWeeks of trea tment 3 5
C LK C LKSerum K (mmol/l) 4.76 3.09** 4.65 2.94**Testis wt (gm) 1.56 1.53 1.64 1.53Tr (gm/testis/16 h) 0.40 0.33* 0.43 0.17**Each value represents the mean of 7-8 animals;clarity. * P<0.025, ** P<0.001, compared with(C) •TABLE 2: Effects of low K diet with or without K supplement on tubulefluid production rate and serum hormone levels 6 weeks after treatment
Control LK LK + K-supplementSerum K (mmol/l) 4.22 + 0.15 1.29 + 0.07* 4.07 + 0.13Testis wt (gm) 1.71 +0.05 1.56 +0.06 1.62 + 0.04TP (gm/testis/16 h) 0.43 +0.02 0.24 + 0.01* 0.49 + 0.04Tubule diameter (um) 103 + 3 97 + 2 98 + 2Serum T (nmol/l) 7.51 + 0.77 3.63 + 0.67* 10.16 +1.2Serum FSH (ng/ml) 5.64 + 0.24 5.67 + 0.53 5.48 + 0.67Serum LH (ng/ml) 0.19 + 0.05 0.15 ~ 0.02 0.21 ~ 0.03Values represent the mean + SEM of at least 6 animals.* P<0.01 when compared with control or low K + K-supplement animals.(1) Y.M. Cheung, J.C. Hwang & P.Y.D. Wong (1977) J. Physiol. 269: 1-15
(1) Bindon, B. M. (1984). Aust. J. BioL Sci. 37: 163189.

78
TNlTF:RSE RELATIONSHIP BETWEEN LIVEWEIGHT AI-ID UTERINEOESTROGEN RECEPTORS IN OVARIECTOMIZED EWES
N.R. Adams and A.J. Ritar
CSIRO, Division of Animal Production, wembley, WA 6014
The ovulation rate of ewes is strongly correlated with theirliveweight, but the mechanisms involved in this relationship areunknown. It is commonly assumed that enhanced nutrition acts byincreasing the release of gonadotrophins from the pituitary. However,the observations presented here suggest that there may be a differentlink between 1iveweight and the reproductive system.
In the first experiment, 16 4-tooth Merino ewes wereovariectomized and 14 days later a segment of upper uterine hornweighing 0.5 to 1 g was taken under general anaesthesia. Theliveweights of the ewes at this time ranged between 42 and 58 kg.
In the second experiment, 14 mature Merino ewes were fedindiVidually over a 5 month period to achieve liveweights rangingbetween 28 and 58 kg. The ewes were ovariectomized 8 weeks beforeslaughter, at which time the entire uterus was taken, minced andhomogenized.
Sp~cific binding of [3H]-oestradiol in the 100,000 xg supernatantof ~ter1ne homogenate was estimated as described previously (1), usinga s1ngle point assay at 10nM. In the first experiment, specificb;nding of progestagens was determined by a similar method, using 10nMr H]-RS020, with non-specific binding determined in the presence of a100 x excess of progesterone.
In the first study, the mean concentration of cytoplasmicoestrogen receptors was 0.60±0.03 pmo1/mg protein, and that ofprogesterone receptors was 1.11±0.05 pmol/mg protein. Thec~ncen~ration of oestrogen receptors was inversely correlated with thel1vewe1ght (r = -0.52, P < 0.05), but there was no significantcorrelation between progesterone receptors and liveweight (r = -0.27)or between oestrogen and progesterone receptors. In the second studythe mean concentration of cytoplasmic oestrogen receptors was '0.84±0.06 pmol/mg protein. The concentration of receptors was againinversely correlated with liveweight (r = -0.66, P < 0.05).
Oestrogen receptors mediate the effects of oestrogen and one ofthese effects is to decrease ovulation rate. The manner by whichliveweight might influence the concentration of oestrogen receptors inovariectomized ewes is unknown. However, it is probable that thechanges observed above reflect the mechanism by which liveweightinfluences ovulation rate.
(1) Tang, B.Y., and Adams, N.R. J. Endocr. 89: 365-370 (1981).
79
OVARIAN COMPENSATORY HYPERTROPHY IN UNILATERALLYOVARIECTOMISED ANOESTROUS EWES
B.K. Campbell1
, R.J. scaramuzzi 2 , J.A. Downing2 and G. Evans 1
1Department of Animal Husbandry, University of Sydney, Sydney.2CSIRO, Division of Animal Production, Prospect, N.S.W.
The compensatory phenomena displayed by the ovary remaining afterunilateral ovariectomy (ULO) in the cycling ewe is associated with atransient increase in follicle stimulating hormone (FSH) levels 5!-12 hafter ULO( 1). In seasonally anoestrous ewes, however, neithercompensatory hypertrophy (COH) (1) nor changes in FSH levels have beenobserved following ULO. This study examined the COH ability of anoestrous ewes when induced to ovulate by exposure to rams before or afterULO.
In October, 1983, 36 seasonally anoestrous Border Leicester xMerino ewes were allocated to one of 4 groups (n=9). Surgery wasperformed on Groups 1 (ULO) and 2 (Sham) 14 days before ram introduction(RI) and Groups 3 (ULO) and 4 (Sham) 1 day after RIo Since priorprogestagen treatment is necessary for stimulation of normal lutealfunction, intravaginal progestagen sponges (Repromap, Upjohn) wereinserted for 10 days prior to RI. Nine days after RI the animals werekilled and the remaining ovaries recovered. Blood samples were taken at20 min. intervals from 4 h prior to ULO to 12 h after ULO, and twicedaily thereafter, to monitor plasma FSH levels.
Of the 36 ewes exposed to rams, 32 displayed oestrus and ovulatedwith no differences evident between the 4 groups. Ovarian weights,ovulation rates and plasma FSH levels are recorded in Table 1.
TABLE 1. Ovulation rates, ovarian weights and FSH levels (x ± sem) inewes sham-operated or ULO at Day RI-14 (Gps.1&2) or RI+1 (Gps.3&4).
GROUP 1 (ULO) 2 (SHAM) 3 (ULO) 4 (SHAM)Ovulation rate 1. 0 1. 0 1. 3±0. 2 1. 3±0. 2
(CL/Ewe ovulating) (n=7) (n=9) (n=9) (n=7)Ovarian weight 2.0±0.3a 1.2±0.1 1.3±0.1 1.S±0.1
minus CL (g) (n=9) (n=18) (n=9) (n=18)FSH l~vels -4 h-ULO 1.S±0.3 1.6±0.3 0.8±0.1 1.0±0.2(ng/ml; n=9) aJ ULO-4 h 1.9±0.3 1.8±0.3 0.9±0.1 1.0±0.2
e 4 h - 8 h 2. 4±0. 4b 1. 6±0. 3 1. 3±0. 2c 1. O±O. 1.~ 8 h-12 h 2.7±0.4b 1.2±0.2 1.7±0.3c 0.9±0.1
24 h+ 1.6±0.2 1.2±0.3 1.1±0.2 0.9±0.1Significantly different (P<O.OS) (a) from Sham control and (b,c) fromFSH levels before ULO within each group.
Ovulation rates were similar in all groups, while mean ovarianweights were greater than controls (P<0.05) only in ewes ULO 14 daysbefore RI. There was a transient increase (P<O. OS) in FSH levelsbetween 4 and 12 h after ULO, irrespective of time of ULO.
From these data, it can be concluded that (a) anoestrous ewes ULOafter RI do not exhibit COH in terms of ovarian weight, as do ewes ULObefore RI, but are able to adjust with respect to the number of eggsshed, and (b) that ULO in anoestrous ewes is followed by a transientincrease in FSH levels similar to that seen in the cycling ewe.
(1) Findlay, J.K., and Cumming, I.A. Biol. Reprod. 22: 178-183 (1977).

80 In
NADPH STIMULATION OF AROMATASE ACI'IVITY IN OVINE GRANULOSA CELLS
L.A. Hutchinson, S.M. Campo, R.S. Carson and J.K. Findlay
Supported by the Australian Meat Research Committee.
Medical Research Centre, Prince Henry's Hospital, Melbourne, Vic., 3004
A COMPARISON OF LUTEAL PROGESTERONE AND OXYTOCIN RELEASE AND THEIRSUBCELLUAR STORAGE
G. Jenkin, G.E. Rice and G.D. Thorburn.Department of Physiology, Monash University, Clayton, Vic.-3168.
The synthesis and release of steroid hormone is thought to bedirectly coupled to stimulation. An active process of release has,however, been proposed which is consistent with the hypothesis thatprogesterone (P4) is sequested into granules and transported to thecell membrane, from where it is released in conjunction with a P4binding protein. It has also been suggested that the enzyme complex3~-hydroxy-steroid dehydrogenase/~5_~4 isomerase (3~-HSD) ,which is principally microsomal and has a high affinity for P4 , maybe the membrane-bound P4-binding protein. Such a mechaniSm wouldprovide a high capacity for the controlled release of P4 from lutealcells. Recently, immunoreactive oxytocin (OT) has been identified incorpora lutea (CL's) from a number of species. Since neurohypophysialaT is stored and transported in membrane-bound granules, lutealgranules observed throughout the oestrous cycle may contain OT andrepresent endogenous peptide synthesis. To further characterizeluteal processing of P4 and OT, we examined their subcellularlocalization, within ovine CL's, using differential and densitygradient centrifugation. Furthermore, we compared some physical andbioch~mical properties of particle-associated P4 with those of OTisolated from the same tissue. CL's, obtained on day 7 or 8 of theoestrous cycle were homogenized in 0.16 M sucrose and subjected todifferential (900, 10,000, 20,000, 30,000 and 100,000 x g)centrifugation. A significantly (p<O.OOl) lower proportion of lutealP4 was sedimented, during differential centrifugation, than aT (42.1+ 1.6% and 63.0 ~ 2.6%, respectively). This difference wasessentially due to the greater proportion of OT recovered in the10,000 x g pellet, which contained 57.0 ~4.0% of luteal OT but only28. 8 ~ 1 .0% of the P4 (p<O. 02) .
The 10,000 x g pellet was subjected to Percoll density-gradientcentrifugation. P4 and OT were associated with particles whichsedimented to a density of 1.049 - 1.054 glml and 1.054 - 1.061 glml,respectively. 3~-HSD activity was evenly distributed throughout thePercoll gradient. The effects, on aT and P4 release from the regionof maximum enrichment, of incubation pH, osmolality and incubationwith a secretagogue (either 5 mM MgATP or 1 mM CuATP) were examined.OT release was not affected by incubation pH but was stimulated byincubation in medium of low osmolality « 200 mOsm, P < 0.01) and byincubation with either MgATP or CuATP. These data are consistent withthe physico-chemical properties reported for peptide-containingneurohypophysial secretory granules. Upon incubation of particleassociated progesterone in buffered medium at 370 C, 70% of totalP4 was recovered in the incubation medium (compared with 33% foroxytocin). The release of the remaining stable particle-associatedP4 w~s not affected by either incubation pH or osmolality, nor wasit affected by the inclusion of MgATP or CuATP. These data are notconsistent with the suggestion that progesterone is stored, in aprotein-bound form, in luteal secretory granules. They are, however,indicative that oxytocin is compartmentalized in a population ofluteal storage granules.
Tsonis, C.G., Carson, R.S. & Findlay, J.K. J. Reprod. Fert. 72:153-163 (1984).Gore-Langton, R.E. & Darrington, J.H. Mol. Cell. Endocrinol. 22:135-151 (1981).
TABLE. Aromatase activity ig ovine granulosa cells.(Mean ng Estrogen/IO viable cells/hr ± S.E., n=3).
Follicle Diameter NADPH Atretic follicles Healthy follicles
< 3.5mm 0.84 ± 0.04a 0.53 ± 0.06a+ 0.53 ± 0.17 1.02 ± 0.18
> 3.5mm 0.90 ± 0.20a 4.58 ± 0.49b+ 2.09 ± 0.17 27.38 ± 1.71c
Aromatase activity (AA) in sheep ovarian granulosa cells (GC)increases during follicular maturation and decreases during atresia(1). The regulatory mechanisms for this activity are not fullyunderstood.
Because AA is dependent on a labile co-enzyme (NADPH), this studyexamined the effect of NADPH on AA in isolated GC from ovine folliclesat different stages of development and atresia. Follicles were obtainedfrom Merino ewes at late-luteal and early-follicular phases of theoestrous cycle, and assigned to one of 4 groups, according to size andmOrPhological atresia (1). GC were then isolated, pooled within eachof the 4 groups and subjected to a one hour radiometric assay validatedfor AA, in the presence or absence of NAD~H at saturating concentrations(0.5 mM), (2).
In the absence of NADPH, AA was low in GC from all atretic andsmall healthy follicles, whereas AA in GC from large healthy follicleswas 4-10 fold higher (a vs b, P < 0.02), confirming earlier studies (1).NADPH stimulated AA in GC from large healthy follicles by 6-fold(b vs c, P < 0.001), but had no significant effect on AA in GC fromsmall healthy or atretic follicles.
It is concluded that atresia is associated with an absence orinactivation of NADPH-dependent Cytochrome P450 - aromatase protein.Furthermore, the ability of NADPH to stimulate AA in ovine GC fromhealthy follicles in vitro, offers a model to study the regulationof AA.
(1)
(2)

82
SUPEROVULATION AND EMBRYO RECOVERYIN AWASSI FAT TAIL SHEEP
W.M.C. MAXWELL 1, R.J. LIGHTFOOT1
, J.P. RYAN 1, R.G. CASEy 1
,
R.P. LEWER 1 and A. LOUCA 2
Department of Agriculture, Western Australia1 andMinistry of Agriculture and Natural Resources, Nicosea, Cyprus 2
There are no reports on the superovulation and collection of embryosfrom Middle East fat tail sheep breeds. This paper presents the resultsof different superovulation treatments which were applied as part of aprogramme to recover and freeze Awassi embryos in Cyprus for importationinto Australia.
During May 1985, 139 mixed age Awassi ewes were treated with 3 superovulatory regimes: (a) 400 i.u. PMSG (Folligon, Intervet) + 12 mg FSH-P(Burns - Biotec.); (b) 200 i.u. PMSG + 12 mg FSH-P; or (c) 400 i.u. PMSG+ 8 mg FSH-P. Intravaginal sponges (Repromap - Upjohn) were inserted for11 - 15 days. PMSG was administered as a single intramuscular injection48 hr before sponge removal. FSH-P treatment commenced 48 hr beforesponge removal and was administered on 3 successive days as 2 intramuscularinjections per day (at 0800 and 1700 hr) using a decreasing dose of 2, 2;1.5, 1.5; 0.5, and 0.5 mg (8 mg) or 3, 3; 2, 2; 1 and 1 mg (12 mg). Intrauterine insemination by laparoscopy (fresh semen collected by artificialvagina or electroejaculation extended 1:2 with Dulbeccos phosphate buffer)was conducted 24 hr after sponge removal. On day 6 (day 0 = day of oestrus and insemination) ewes were subjected to laparotomy, when number andstatus (normal or regressed) of corpora lutea (CL) were recorded. Theresults are presented in Table 1.
TABLE 1. Ovarian response and recovery of embryos in relation to superovulation treatment.
No. ewes No. ova/embryos (%)Treat. (n) Mean no. regressed
CL CL (%) Recovered Fertilised
a (44) 12.6±1.21 15(34.1) 258(65.8) 149(57.8)b (49) 9.3±0.83 21(42.9) 160(54.4) 86(53.8)c (46) 10.3±0.96 14(30.4) 219(59.8) 115(52.5)
Mean numbers of CL and proportion of eggs recovered were higher forewes treated with superovulation regime (a) than for (b) and (c). Eggfertilisation rates were low due to the use of poor quality semen collectedby electroejaculation from young rams.
The results of this programme have shown that techniques developed inAustralia may be applied to the superovulation and collection of embryosfrom Middle East fat tail sheep.
83
PLASMA OXYTOCIN CONCENTRATIONS IN HEIFERS AT MID CYCLEFOLLOWING A CLOPROSTENOL STIMULUS
S. Mock, J. Parsons, M. Cardner, S. McPhee* and R.J. Fairclough*
Department of Agriculture and Fisheries, Melbourne University*Department of Agriculture and Rural Affairs,
Animal Research Institute, Werribee, 3030.
The finding of high amounts of oxytocin in corpora lutea(CL), of ewes at mid cycle (1) and the observation of activesecretion of oxytocin by the ovary in sheep given a prostaglandinF2a stimulus (2) have provided good evidence that oxytocin isst~red and released from luteal tissue in sheep in~. Also incattle active secretion of oxytocin has been demonstrated duringthe luteal phase of the oestrous cycle (3). However the factor(s)responsible for stimulating ovarian oxytocin release duringluteolysis is still uncertain. The present study was undertakento determine plasma oxytocin concentrations in cows at mid cyclefollowing a PGF2a stimulus.
Six heifer cows at Day 10 of the oestrous cycle were givenan im injection of cloprostenol (500 ug). Blood samples werecollected by venepuncture into heparised vacutainer tubes at 30and 15 min before and at 0, 2, 5, 10, 15, 20, 30 and 60 min afterthe injection.
Oxytocin wac extracted from plasma by eluting from a C-18 SepPak using 75% methanol/water (0.1% TFA), evaporating the extractto dryness under vacuum and then assaying for oxytocin using aspecific radioimmunoassay. The sensitivity of the method was 2pg/ml and the interassay coefficient of variation for the assaywas 12%.
Results showed a marked increase in plasma oxytocin levelsin each of the six heifers. The plasma oxytocin concentrationsincreased from a mean value of 17.2 ± 8 (SD) pg/ml in the3 samples before the PGF2a injection to a mean maximum of35.6 ± 9.8 pg/ml (range 24-50 pg/ml) at 5 min post injection.
These data in heifers are consistent with the earlier resultsin sheep that exogenous PGFs can cause an increase in plasmaoxytocin concentrations at mid cycle.
(1) Wathes, D.C. and Swann, R.W. Nature 297: 225 (1982).(2) Flint, A.P.F. and Sheldrick, E.L. Nature 297: 587 (1982).(3) Schams, D. et al., Acta Endocr. Suppl. 25'6:' 261 (1983).

8485
J. D. O'Shea, R. J. Rodgers and P. J. Wright
DO SMALL LUTEAL CELLS DIFFERENTIATE INTO LARGE LUTEAL tELLSDURING THE OVINE OESTROUS CYCLE?
School of Veterinary Science, University of Melbourne,Parkville, Victoria, 3052
Jugular(N=16)
UOV vein(,tl=ll)
Jugular(,tl=16)
UOV vein(,tl=23)
ESTIMATED UTERO-OVARIAN PRODUCrrON RATES OF STEROIDS IN UNMATED AND INPREGNANT GILTS BETWEEN DAYS 9 & 17 AFTER OESTRUS/COITUS
~~ne~~~~~;----~~~;-~-;~;:;_-3.3~-;-:~;---~7.5 ~--~~~-~~ ± ;~~Preg. sulphate 56.5 ± 18.0 37.3 ± 8.4 124.8 ± 10.5 25.0 ± 1.9Progesterone 95.1 + 31.9 72.7 ± 12.0 409.9 ± 25.4 89.2 ± 6.4DHEA 0.19 +0.02 0.10 ± 0.02 0.60 ± 0.08 0.44 ± 0.10Androstenedione 1.35 ±0.34 0.45 ± 0.04 8.04 ± 0.83 1.15 ± 0.14Testosterone 0.21 + 0.09 0.85 ± 0.18 2.97 ± 0.39 0.84 ± 0.16oestradiol 0.04 ±0.01 0.03 ± 0.01 0.42 ± 0.12 0.03 ± 0.01
~~~~~~--~~~~ct~;~~~~~-(PR) -~~~~ogeste~on~-i~-~~~~d-~~d- in- ma~~dgilts between Days 9 and 17 after oestrus are near 158 and 274 umoles/2~
respectively (2) • As corpora lutea are the principal source of thJ.ssteroid in SCMS, these PRs can be related to differences in progester?neconcentrations between UOV and peripheral plasma (Table 1), to denveconstants (K) which can then be used in the estimation of PRs of othersteroids of mv origin. That is,
PR = K x [V - A] ,where V represents the concentration of progesterone in the UOV venousdrainage and A represents the concentrat~on.of progesterone in the UOVarterial supply (estimated by the level J.n Jugular plasma). .
On the basis of our progesterone data, K values have been derJ.ved formated (K = 854 1/24h) and unmated (K = 7053 1/24h) gilt-:- between Days 11and 17 after oestrus. Using these values ar:d the data. J.n Table I, PRs ofother UOV-derived steroids were then determJ.ned accordJ.ng to ~he aboveformula. Values (umoles/24h) for unmated and pregnant gJ.lts are,respectively; pregnenolone (46, 37), pregnenolone sulphate (135, 85),DHEA (0.63, 0.14), androstenedione (6.35, 5.90), testosterone ~O, 1:80)and oestradiol (0.07, 0.33). These values approximate physJ.ologJ.calrates of steroid replacement for ovariectomized sows between Days 9 and17 after oestrus/coitus.
(1) Stone, B.A. & Seamark, R.F. J. Reprod. Fert. (in press). (1985).(2) Stone, B.A., Johnson, D.W. & Seamark, R.F. Proc. Aust. Soc.
Bgprod. BioI. 14: 99, Abstr. (1983).
------------_.------- --------------------------------------------------
B.A. Stone, a.M. Petrucco, P.QUinn and R.F. Seamark
Deparbnent of (j:)stetrics and Gynaecology, university of Adelaide
Levels of steroid hormones have been determined (by radioimmunoassay ofchranatographed extracts; 1) in peripheral and in utero-ovarian (UOV)venous plasma of gilts between Days 9 and 17 after oestrus. The results(Table 1) indicate release of pregnenolone, pregnenolone sulphate,progesterone, DHEA, androstenedione and oestradiol by the UOV system ofunmated 'and of pregnant gilts during this period. Testosterone appearedto be secreted in the pregnant animals only.
Table 1 : Steroid hormone levels (nM; mean ± s.e.m.) in peripher:al and--rn UOV venous plasma of gilts between Days 9 and 17 after oestrus----------------------------~~;d----------------------~~;d----------
Fitz, LA., Sawyer, H.R. and Niswender, G.D. (1981) Biol.Reprod. 24: Supple 1 54A.O'Shea, rD., ,Rodgers, R.J. and Wright, P.J. (1984) J. Reprod.rert. 2£: 75-85.
On the basis of evidence from counts of small and large lutealcells after enzymatic dispersion of ovine luteal tissue it has beensuggested that small luteal (SL) cells may differentiate into largeluteal (LL) cells during the latter part of the oestrous c:£cle (1).Using morphometric methods on whole luteal tissue, we have tested thissuggestion by investigating whether LL cells increase in number, andSL cells correspondingly decrease in number, between the mid and lateluteal phases of the cycle.
Corpora lutea (CL) were collected from cyclic Corriedale ewes 9and 13 days after the onset of oestrus (.:: 8 and .:: 12 days afterovulation) (n = 5 ewes per group), and fixed by vascular perfusion.Morphometric methods (2) were used on electron micrographs of lutealtissue to determine numbers and sizes of the different cell types.
(1 )
(2)
These data demonstrate that there may be some enlargement of theluteal cells in the latter part of the cycle. However, our findingsprovide no support for the hypothesis that SL cells differentiate intoLL cells, whose numbers were not elevated late in the oestrous cycle.
Total ~umbers of cells per CL were got different (200.8 ! s.e.m.20.6 x 10 vs 212.5 + 23.8 x 10) between 8 and 12 dayspost-ovulation. Nor were there 6 significant diffegences in thenumbers of LL cebls (9.6 ! 0.9 x 10 ~s 7.7 ! 0.4 x 10 ) or SL cells(57.5 + 8.1 x 10 vs 47.5 + 9.1 x 10 ) rjspect1vely. Mean size ofthe LL -cells (15.6 + 1.3 vs -19.65 :!; 0.7 J.lm x 10 ) was greater 12 dayspost-ovulation than-at 8 days (P < 0.05, !-test). 3SL ce1l sizes didnot differ significantly (2.1 ! 0.3 vs 2.7 :!; 0.3 J.lm x 10 ) between 8and 12 days.

86
OXYGEN CONSUMPTION BY THE OVARY OF THE PREGNANT RAT
R. Swann and N. Bruce
Department of Anatomy and Human Biology, University of W.A., NedlandsW.A. 6009.
Evidence for high metabolic activity of luteal tissue includes largenumbers of mitochondria in luteal cells, a high rate of ovarian bloodflow and the secretion of large amounts of steroids per unit mass oftissue (1). Oxygen consumption should further reflect this highmetabolic activity but frequently it has been noted that ovarian venousblood is red suggesting a low rate of abstraction of oxygen. Thequestion then becomes whether the high blood flow is sufficient to stillresult in a high oxygen consumption. Data is available on oxygenconsumption by the intact ovary in the follicular and luteal phase ofwomen (2) and sheep (3) but concomitant rates of steroid secretion werenot recorded.
In the present work, both oxygen consumption and progestin secretionwere measured and compared in 5 rats at day 16 of gestation: a time ofmaximum progesterone secretion. The rats were prepared for ovarianvenous outflow (1) and arterial and venous samples collected for bloodgas analysis and steroid assay. Ovarian blood flow, heart rate,arterial blood pressure, respiration rate and body temperature weremonitored during the experiment. Oxygen content was determined by amOdified micromanometric technique of Van Slyke. Progesterone (P) and20-ahydroxypregn-4-ene-3-one (20a) were measured by RIA.
Oxygen consumption for a single ovary was 22.8 ± 3.8 ml/min per kgovary (mean ± SEM, n = 5). P secretion was 2.3 ± 0.4 nmol/min ; 20a was0.5 ± 0.15 nmol/ mi~ glving a total progestin secretion of 2.8 ± 0.5nmol/min. The amount of oxygen consumed was positively correlated withP, 2Da and total progestin (significant only for total progestins,r=0.85 P<0.05).
The oxygen consumption of the pregnant rat ovary is comparable withthat found in non-pregnant sheep during the luteal phase (14ml/min/kg).It was 7 fold greater, however, than that consumed by ovarian slicesfrom superovulated rats (3ml/min per kg). The rate of progestinsecretion in the latter tissue was less than 5% of the intact ovaryexamined here.
Theoretical consideration of the amount of oxygen consumed in thesynthesis of both of the progestinsfrom cholesterol suggests that3.8ml/min/kg ovary is required to achieve the rate of secretion observed(2.8nmol/mi~). Thus on~y 18% of the total amount consumed by the ovarycan be attrlbuted to thlS step. Since over 80% of the ovarian bloodflow goes to the CL oxygen consumption by other tissues is unlikely tobe significant. The high oxygen consumption found reflects either thehigh cost of maintaining such active steroidogenesis and secretion orthe ~vary is performing some other major oxygen consuming function:Studles are progressing on requirements for 0 tension as distinct fromconsumption for maintainence of normal steroiaogenic rates.
(1) Waddell, B.J. ~~ J. Endocr. 94: 61-67 (1982)(2) Frazer, I. et ~ J. Reprod. Fert. 33: 11-17 (1973).(3) Baird, D.T. ~~ J. Endo. 21: 405=411 (1973).(4) Flint, A.P.F. and Denton, R.M. Biochem. J. ~: 243-254 (1973).
•I
III!~
87
AN INTERACTION BETWEEN PROGESTERONE PRETREATMENT AND STAGE OF THEANOESTROUS SEASON ON THE OVULATORY RESPONSE
OF EWES TO BOLUS GnRH TREATMENT
P.J. Wright, K.E. Davis, J.D. O'Shea and J.A. BurdenSchool of Veterinary Science, University of Melbourne, Werribee 3030,
Victoria.
Ovulation induced in seasonally anoestrous ewes by GnRHadministered in bolus (1) or in pulsatile (2) treatment regimenscommonly results in inadequate corpora lutea that produce littleprogesterone. Progesterone pretreatment results in normal corporalutea in response to pulsatile treatment regimens (2). We haveassessed the influence of progesterone pretreatment on the ovulatoryresponse of ewes to a bolus GnRH treatment regimen.
Seasonally anoestrous Corriedale ewes were studied during early(October) and late (December) anoestrous season. The ewes receivedprogesterone (20mg) or vehicle i.m•• GnRH (3 x 30ug i.v. at 90 min'intervals) was administered 72h later ~Day 0). On Day 9 blood samplesfor progesterone determination were taken, the ewes anaesthetised andthe ovaries removed. Corpora lutea were identified, dissected out andweighed.
TABLE 1. The ovulatory response and luteal function in GnRH-treatedewes pretreated with progesterone or vehicle
Stage of Anoestrous SeasonEarly Late
progesterone Pretreatment+ +
No of ewes 12 12 21 19No of ewes with CL(a) 4 3 14 3
(g±s.d.) (a) 0.2808 0.2771 (b)0.2792 0.2079CL wt
±0.116 ±0.087 ±0.073 ±0.083Plasma progesterone(a) 2.8 2.5 1.5 1.2
(ng/ml) ±0.67 ±0.4 ±0.5 ±0.4(a) Day 9, Day O=day of GnRH administration. (b) n 4, 2CL for 1 ewe.
The results (Table 1) demonstrate that progesterone pretreatmentof ewes treated with bolus GnRH i) increased the ovulatory responseonly in late anoestrus (p<0.001), but ii) did not affect lutealfunction as assessed from luteal weight and plasma progesteroneconcentrations.
These findings suggest that pretreatment of anoestrous ewes withprogesterone promotes luteal function by an interaction with theresponses to pulsatile GnRH treatment and not with the preovulatorysurge release of LH. The basis of the interaction between season andprogesterone pretreatment on ovulation rate is not known.(1) McNeilly, A.S. and Land, R.B. J. Reprod. Fert. 56: 601-609
(1979) •(2) McLeod, B.J., Haresign, W. and Lamming, G.E. J. Reprod. Fert. 65
: 223-230 (1982).
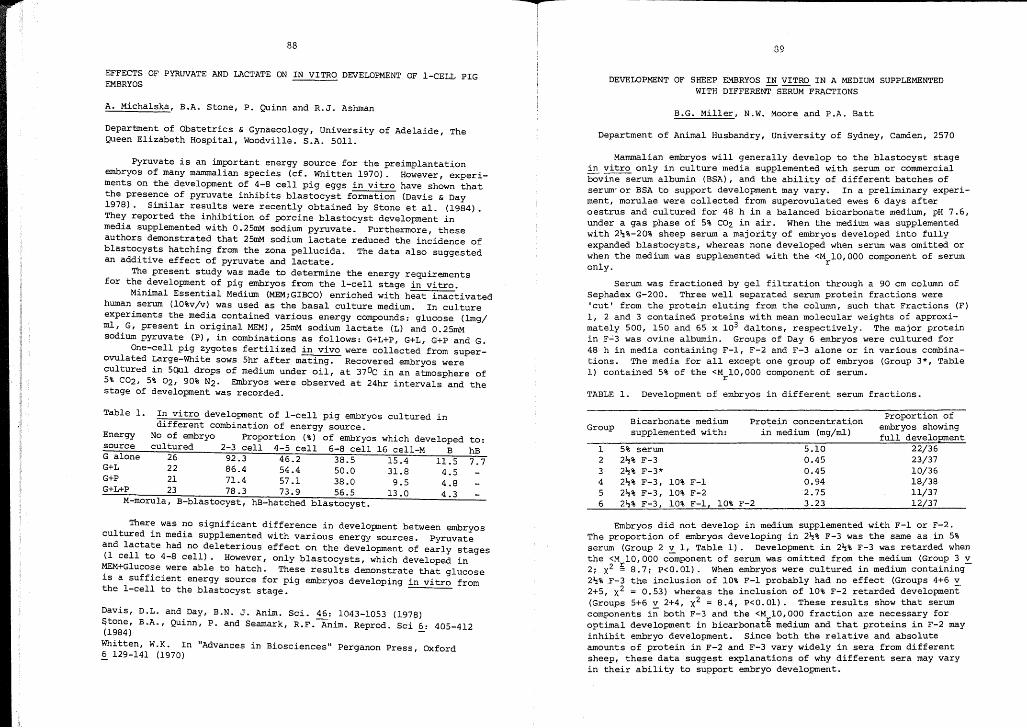
'188 no
UJ
EFFECTS OF PYRUVATE AND LACTATE ON IN VITRO DEVELOPMENT OF I-CELL PIGEMBRYOS DEVELOPMENT OF SHEEP EMBRYOS IN VITRO IN A MEDIUM SUPPLEMENTED----WITH DIFFERENT SERUM FRACTIONS
A. Michalska, B.A. Stone, P. Quinn and R.J. AshmanB.G. Miller, N.W. Moore and P.A. Batt
Department of Obstetrics & Gynaecology, University of Adelaide, TheQueen Elizabeth Hospital, Woodville. S.A. 5011. Department of Animal Husbandry, University of Sydney, Camden, 2570
Mammalian embryos will generally develop to the blastocyst stagein vitro only in culture media supplemented with serum or commercialbovine serum albumin (BSA), and the ability of different batches ofserum/or BSA to support development may vary. In a preliminary experiment, morulae were collected from superovulated ewes 6 days afteroestrus and cultured for 48 h in a balanced bicarbonate medium, pH 7.6,under a gas phase of 5% C02 in air. When the medium was supplementedwith 2~%-20% sheep serum a majority of embryos developed into fUllyexpanded blastocysts, whereas none developed when serum was omitted orwhen the medium was supplemented with the <MrlO,OOO component of serumonly.
Serum was fractioned by gel filtration through a 90 cm column ofSephadex G-200. Three well separated serum protein fractions were'cut' from the protein eluting from the column, such that Fractions (F)1, 2 and 3 contained proteins with mean molecular weights of approximately 500, 150 and 65x 103 daltons, respectively. The major proteinin F~3 was ovine albumin. Groups of Day 6 embryos were cultured for48 h in media containing F-l, F-2 and F-3 alone or in various combinations. The media for all except one group of embryos (Group 3*, Table1) contained 5% of the <MrlO,OOO component of serum.
TABLE 1. Development of embryos in different serum fractions.
Embryos did not develop in medium supplemented with F-l or F-2.The proportion of embryos developing in 2~% F-3 was the same as in 5%serum (Group 2 v 1, Table 1). Development in 2~% F-3 was retarded whenthe <M 10,000 c;mponent of serum was omitted from the medium (Group 3 ~2; X2 ~ 8.7; p<O.Ol). When embryos were cultured in medium containing2~%F-3 the inclusion of 10% F-l probably had no effect (Groups 4+6 v2+5, X2 = 0.53) whereas the inclusion of 10% F-2 retarded development(Groups 5+6 v 2+4, X2 = 8.4, P<O.Ol). These results show that serumcomponents i; both F-3 and the <M 10,000 fraction are necessary foroptimal development in bicarbonat~ medium and that proteins in F-2 mayinhibit embryo development. Since both the relative and absoluteamounts of protein in F-2 and F-3 vary widely in sera from differentsheep, these data suggest explanations of why different sera may varyin their ability to support embryo development.
22/3623/3710/3618/3811/3712/37
Proportion ofembryos showingfull development
5.100.450.450.942.753.23
Protein concentrationin medium (mg/rol)
Bicarbonate mediumsupplemented with:
5% serum2~% F-32~% F-3*2~% F-3, 10% F-l2~% F-3, 10% F-22~% F-3, 10% F-l, 10% F-2
123456
Group
M-morula, B blastocyst, hB-hatched blastocyst.
Pyruvate is an important energy source for the preimplantationembryos of many mammalian species (cf. Whitten 1970). However, experiments on the development of 4-8 cell pig eggs in vitro have shown thatthe presence of pyruvate inhibits blastocyst formation (Davis & Day1978). Similar results were recently obtained by Stone et al. (1984).They reported the inhibition of porcine blastocyst development inmedia supplemented with 0.25mM sodium pyruvate. Furthermore, theseauthors demonstrated that 25mM sodium lactate reduced the incidence ofblastocysts hatching from the zona pellucida. The data also suggestedan additive effect of pyruvate and lactate.
The present stUdy was made to determine the energy requirementsfor the development of pig embryos from the I-cell stage in vitro.
Minimal Essential Medium (MEM;GIBCO) enriched with heat inactivatedhuman serum (lO%v/v) was used as the basal culture medium. In cultureexperiments the media contained various energy compounds: glucose (lmg/rol, G, present in original MEM), 25mM sodium lactate (L) and 0.25mMsodium pyruvate (P), in COmbinations as follows: G+L+P, G+L, G+P and G.
One-cell pig zygotes fertilized in vivo were collected from superovulated Large-White sows 5hr after mating. Recovered embryos werecultured in 5Qul drops of medium under oil, at 37 0C in an atmosphere of5% C02, 5% 02, 90% N2. Embryos were observed at 24hr intervals and thestage of development was recorded.
Table 1. In vitro development of I-cell pig embryos cultured indifferent combination of energy source.
Energy No of embryo Proportion (%) of embryos which developed to:source cultured 2-3 cell 4-5 cell 6-8 cell 16 cell-M B hBG alone 26 92.3 46.2 38.5 15.4 11.5 7.7G+L 22 86.4 54.4 50.0 31.8 4.5G+P 21 71.4 57.1 38.0 9.5 4.8G+L+P 23 78.3 73.9 56.5 13.0 4.3
Ther~ was ~o significant difference in development between embryoscultured ~n med~a supplemented with various energy sources. Pyruvateand lactate had no deleterious effect on the development of early stages(1 cell to 4-8 cell). However, only blastocysts, which developed in~+Gluco~e.were able to hatch. These results demonstrate that glucose~s a suff~c~ent energy source for pig embryos developing in vitro fromthe I-cell to the blastocyst stage.
Davis, D.L. and Day, B.N. J. Anim. Sci. 46: 1043-1053 (1978)qtone, B.A., Quinn, P. and Seamark, R.F.~nim. Reprod. Sci ~: 405-412(1984)
Whitten, W.K. In "Advances in Biosciences" Perganon Press, Oxford~ 129-141 (1970)

90 91
BIOLOGICAL ACTIVITY OF CRONOLONE IN THE SHEEP, MOUSE AND RABBIT
x. Zhang, G.M. Stone* and B.G. Miller
STRUcrURAL AND FUNCTIONAL INVOLUTION AND REGENERATIONOF THE UTERUS OF POST-PARTUM EWES
University of Sydney; Departments of Animal Husbandry, Camden,2570; and Veterinary Physiology*, Sydney, 2006
Rates of dissociation of 3H-progesterone and 3H-cronolone from thereceptor were determined after addition of excess cold steroid. In the~heep progesterone dissociated more rapidly than did cronolone, whereas~n the mouse cronolone bound only weakly to the receptor and dissociatedmore rapidly than progesterone. Scatchard analyses employing 3H-progestero~r and 3H-c~onolone yielded association constants of 0.62 and 0.93 109mo~ '_Iespect~ve~y, for sheep endometrial receptor, and 0.47 and 0.00410 mol , respect~vely, for mouse uterine receptor. The results indicatethat cronolone cannot sustain pregnancies in the mouse because it bindsonly weakly t~ the uterine progesterone receptor. The failure of cronolone to susta~n pregnancies in rabbits appears to have a differenteXPl~natio~, and the ability of cronolone to sustain pregnancies inovar~ectom~zed ewes remains to be determined.
(1) Holst, P.J. and Moore, N.W. J. Endocrin. 41, 255-61 (1968).(2) Shelton, J.N. and Fletcher, I.C. J. Endocrin. 47, 395-6 (1970).
Cronolone (9n-fluoro-llS-hydroxy-17a-acetoxyprogesterone) is usedextensively to regulate ovarian activity in the ewe and is more than 20times as potent as progesterone in bioassays based on the priming ofspayed ewes to exhibit oestrus in response to oestradiol (1). However,cronolone is less potent than progesterone in maintaining pregnancy ~n
rats (2). In an attempt to understand this anomalous behaviour of cronolone, we have examined its ability to sustain pregnancies in ovariectomized mice and rabbits and its binding affinity, in vitro, to highaffinity cytosol progesterone receptors recovered:Erom the endometriumof the sheep and whole uteri of mice and rabbits.
When mice were ovariectomized on Day 6, pregnancy was maintaineduntil Day 16 in 13/15 mice given 1.0 mg progesterone daily, but in 0/10,0/14 and 0/15 mice given 0.1, 1.0 and 2.0 mg respectively of cronolonedaily. Similarly, when rabbits were ovariectomized on Day 10, pregnancywas ~aintain~d until Day 20 in 5/6 rabbits given 6 mg progesterone daily,but ~n 0/6 g~ven each of 0.5 and 6.0 mg cronolone daily. The implantedembryos died promptly after the time of ovariectomy in all mice andrabbits receiving cronolone. To determine in vitro binding affinities,cytosol receptor and various tritiated ligands were incubated for 16hours at 0° in the absence or presence of 4 levels of unlabelled steroid(Table l~. Relative binding affinity was calculated as mol of progesterone wh~ch reduced Bo by 50%/mol of cronolone which reduced Bo by 50%, .x 100 (where Bo = binding of the 3H-ligand to the receptor in the absenceof added, unlabelled steroid) .
Stage PP Oestradiol(days) n Deqree of re-e ithelialization receptorsa
1 2 3 4 (fm/mg protein)10-22 7 4 1 2 0 566.9 ± 109.429-32 9 1 1 4 3 566.4 ± 126.838-41 9 0 1 2 6 673.7 ± 141.048-50 10 0 1 0 9 715.9 ± 113.458--61 8 0 0 1 7 510.0 ± 121.0Control EwesCyclic 5 0 0 0 5 245.8 ± 74.0Anoestrous 8 0 0 0 8 599.4 ± 143.3Oestradiol receptorsa 553.0 786.5 595.9 576.0(fm/mq protein) ±109.9 ±105.6 ±126.9 ± 61.2
A.H. Williams, P.J. wright*, J.D. O'Shea* and I.J. Clarke+Department of Agriculture, Animal Research Institute; *school of
Veterinary Science, University of Melbourne, Werribee 3030; +MedicalResearch Centre, Prince Henry's Hospital, Melbourne 3003, Victoria.
a Mean ± s.e.m.Incomplete structural regeneration of the uterus is a factor
likely to limit fertility in ewes up to 41 days PP (Table 1). Thelack of relationship between ER concentrations and either stage PP ordegree of structural regeneration indicates that inadequacy of uterineER concentrations is not a factor limiting fertility in PP ewes.1. O'Shea, J.D. and Wright, P.J. Cell Tiss. Res. 236:477-485 (1984).2. Findlay, J.K., Clarke, I.J., Swaney, J., colvin:-N. and Doughton,
B. J. Reprod. Fert. 64: 329-339 (1982).
TABLE 1. Re-epithelialisation and oestrogen receptor concentrationsof uteri from PP, and from control cyclic and seasonally anoestrousewes
uteri were removed from Corriedale ewes that were 10-61 days PP,and from control dioestrous and seasonally anoestrous ewes. Sectionswere taken for the determination of cytosolic oestrogen receptor (ER)concentrations (2) and for histological examination (1). From PP ewesthe uterine horn that had borne the foetus was used. The uterinesections included 3-6 caruncles. On histological examination of atleast 3 caruncles the uteri were graded acdording to the degree of reepithelialisation thus:- 1 = re-epithelialiation incomplete, completenecrotic plaques still present on some caruncles, 2 = reepithelialisation incomplete, ± remnants of plaques, 3 = reepithelialization almost complete ± hyaline tissue below theepithelium, 4 = all caruncles fully re-epithelialized.
A study of uterine involution, involving a small number of ewes,showed that the process was completed by 30-40 days post partum(PP)(l). We have assessed aspects of both structural (reepithelialisation of the uterus) and functional (oestrogen receptorconcentrations) involution of the uterus in a larger number of ewes todetermine the likely significance of inadequacy of uterine involutionas a factor limiting fertility in PP ewes.
100205
Rabbitprogesterone
10016
Mouseprogesterone
100126
cronoloneSheep
100155
progesterone
Relative binding affinity of progesterone and cronolone.
Species3H-Ligand
TABLE 1.
ProgesteroneCronolone

92 93
PROTEIN SECRETION BY ENDOMETRIAL EPITHELIAL CELLS INEARLY PREGNANCY IN THE EWE
OXYTCX:IN BINDING IN SHEEP ENDOMETRIUM DuRING THE OESTROUSCYCLE AND EARLY PREGNANCY
L.A. Salamonsen, S.D. Philpott, B. Doughton and J.K. Findlay
Medical Research Centre, Prince Henry's Hospital, Melbourne, Vic, 3004
Susan D. Philpott, B. Doughton, L.A. Salamonsen and J .K. Findlay
Medical Research Centre, Prince Henry's Hospital, Melbourne, 3004
a vs 0, p < 0.05
Oxytocin binding sites were located on epithe~ial cell m~ranes;
mean specific binding ranged from 1.97 to 3.37% uslng S~20~10
epi thelial cells per tube in 3 experiments. However, blndlng wasnegligible using a corresponding number of stromal c~lls.
In conclusion our results support the hypothesls that the concentration of end~etrial oxytocin receptors on epithelial cell membranes is a fact~r limitir:g PGF?a release by the uterus, and thusluteolysis, partlcularly 1n earIy pregnancy.
(1) Roberts, J.S., McCracken, J.A., Gavagan, J.E., and Soloff, M.S.Endocrinology 99: 1107-1114, (1976).
(2) Soloff, M.S. Biochem. Biophys. Res. Commun. 66 ~71-677 (1975).(3) Salamonsen, L.A., 0, Wai Sum, Doughton, B. and Flndlay J.K.
Endocrinology (in press).
Supported by the Australian Wool Research Trust Fund.
PGF is the luteolytic agent responsible for the regression of thecorpus lui:~um in nonpregnant sheep. It has been,postulated t~a~ ~heconcentration of endometrial oxytocin receptors 1S a factor 11m1t1ngPGF release by the uterus, and thus luteolysis (1). .
2a The aims of this study were firstly to describe the changes 1nconcentration of endometrial oxytocin binding sites during the oestrouscycle and early pregnancy in the ewe, and secondly to determine on whichendometrial cell type the binding sites are located. _
Corriedale ewes, mated with either a vasectomized ram (nonpregnant,N=16) or a fertile ram (pregnant, N=6) underwent hysterectomy on Days12 to 15 of the estrous cycle and on Days 13 and 15 ~f pregnan~y (~a~
O=estrus). Endometrial membranes were assayed for [ H]-oxytoc:m ~lnd1ng
activity using a published method (2). For the bindir:g local1satlOnexperiments, ovariectomized ewes (N=3) were treated w1th estrogen andprogesterone implants (E+P-P). Epithelial and3stromal c711s.fr~ theendometrium wre isolated (3) and assayed for [ HJ-oxytoc1n b1nd1ngactivity. . .....
The relative concentration of endometr1al oxytoc1n b1nd1ng slteswas found to vary cyclically with a high concentration ~f s~te~ aPP7aring from Day 14 (p < 0.05) in nonpregnant ewes. Oxytoc1n bmd1ng d1dnot increase in early pregnant ewes (Table 1).
TABLE 1. Specific binding of [3HJ-oxytocin by endometrial membranes(900 ~g protein/tube) from ewes during the estrous cycle (NP) and earlypregnancy (p) (N=2-3/group; mean %± s.d.)
NP 1.26±0 .05 1.26±0 .09a 1.8S±0 .36 5 .0±6 .38P 1.8S±0.80a
14.5 15
6.69±4.63 lS.99±12.68~1.16± 0.15
14DAY 12 13 13.5
There is a direct action of the blastocyst on endometrial proteinsynthesis by endometrial tissue slices (1,2). The aim of this studywas to demonstrate an effect of the blastocyst in vivo and in vitro onsecretion of individual proteins by endometrial epithelial cells.
Epithelial endometrial cells were isolated on day 13 from nonpregnant (NPr) and pregnant ewes (Pr) (3) and cultured (in the presenceor absence of protein secreted by IS-day blastocysts maintained inseparate culture (EM) (4) or from culture3gf skeletal muscle (SM») underoptimiZed conditions in the presence of S-methionine (S-met). Thesecreted proteins were analysed by measuring the incorporation of S-metinto secreted protein and by 2D-SDS-PAGE and autoradiography.
Incorporation of S-met into secreted protein was higher (p < 0.05)for epithelial cells fr~ Pr than from NPr animals [79 ± 18 (N=16)vs 33 ± 8 (N=13) q:rn x 10 per dish :mean ± SEJ confirming previousfindings using whole tissue (1). Secretion from NPr cells was increased[67 ± 34 (N=S): P = O.lJ by 30 ~g EM/ml, whereas 30 ~g SM/ml did notchange secretion by Pr cells [61 ± 8 (N=S)J.
2D-PAGE showed 4 secreted proteins (mol wt range 85 kD-IOO kDpI < 6) which were either absent from or present in only small amountsin secretions from NPr cells on both days of culture. The proportionof these proteins greatly increased in secretions from Pr cells,especially on day 2-3 of cuJture. The presence of EM in NPr culturesenhanced the secretion of these proteins. However, this effect was notseen if EM was heat-treated (8OC/lSmin) or if SM was substituted forEM • These 4 proteins are in a similar mol wt and pI range as thoseinduced in endometrial slices from day 15 NPr ewes by ovine trophoblastprotein I (2).
It is concluded that endometrial epithelial cells from Pr ewesare metabolically more active than those from NPr ewes on Day 13, andthat the blastocyst and its secretions induce the secretion of severalspecific proteins by epithelial cells. The role of these proteins inblastocyst growth and implantation requires further investigation.
Supported by the Australian Wool Research Trust Fund.
(1) Findlay, J .K., Clarke, LJ., Swaney, J., Colvin, N., and Deughton,B. (1982) J. Reprod. Fert. 64: 329-339.
(2) Godkin, J.D., Bazer, F.W. andRoberts , R:M: (1984) Endocrinology114: 120-130.
(3) Salamonsen, L.A., 0, W.S., Doughton , B. and Findlay, J .K.Endocrinology (in press). .
(4) Salamonsen, L.A., Doughton, B. and Findlay, J.K. (1984) InReproduction in Sheep, D.R. Lindsay and D.T. Pearce (eds). pp.115-117. Australian Academy of Science, Canberra.

94 95
NA+-GLYCINE CO-TRANSPORT IN PREIMPLANTATION MOUSE EMBRYOS
Jill G. Hobbs and Peter L. Kaye
THE PRCXiE;;TERONE RECEPTOR ANTAGONIST RU 486 STIMULATESPROSTAGLANDIN PRODUCTION IN HUMAN ENDOMETRIAL CELLS
Department of Physiology and Pharmacology,University of Queensland, st Lucia 4067
Glycine enters embryos of both stages by a combination of a nonsaturable system independent of Na+ and a saturable system dependenton Na+ for activity. The kinetic constants (Table 1) show that Kc thediffusion constant doubles, and both the affinity (1Sn) and maximaluptake rate (Vmax ) of the saturable system increase duringdevelopment, suggesting modifications of the transporter.
The Na+-dependence of the saturable component is best describedby a function in [Na+j2 i.e. a Hill coefficient of 2 at both stages.This means that 2 Na+ are required for each glycine moleculetransported.
We conclude that embryos possess glycine-2 Na+ co-transportsystems. In blastocysts this system may frovide intracellular Na+ forthe activity of trophectodermal Na+/K -ATPases during blast~cystexpansion.
This work was supported by grants from NHMRC and University ofQueensland Special Projects.
D.L. Healy, R.W. Kelly, M.J. Cameron, I.T. Cameron and D.T. Baird
Dept. O&G and M.R.C. Reproductive Biology Unit, Edinburgh, Scotland(D.L.H. Current Address: Medical Research Centre, PHH &
Dept. O&G, QVMC, Monash University, Melbourne.
Menstruation in women and in non,human primates can be inducedby the progesterone receptor antagonist RU 486 (17~ hydroxy-l~-4
dimethylaminophenyl-17~ (1-propynyl)-estra~4,9-dien~3-one:l). AlthoughRU 486 can produce uterine bleeding by a d~rect.act~o~ upon the endo~
metrium, without any requirement for a luteolyt~c a~tl0n, the endocr~ne
basis for this action has not been reported. In th~s study, weexplored in vitro the mechanism of menstruation induced by RU 486.
Endometrium from normal women between days 6 and 25 of theirmenstrual cycle and undergoing 5ubal sterilization (n=~2), waso .collected in Hank's solution (4 C), incubated for 30 m~n at 37 C lncollagenase and DNA'ase and filtered through glass wool and nylon meshto separate stromal from glandular cells. Glandular6cells were treatedfurther with collagenase, dispersed, washed and lxlO glandular orstromal cells per ml aliquots cultured for ~4h in supplem~nted Ml99medium with 100 nM oestradiol (E ) and varylng concentratlons ofprogesterone (P) or RU 486 (10 ~ to 1 uM)'. After i~cubat~on of thecells, aliquot~ of medium (150 ~l) were asplrated, m~~ed w~th an ~alvolume of methyloximating solution (0.12 mM methoxyam~ne hydr~chlor~de
in 1 M sodium acetate, pH 5.6-5.8) and assaYed3by antlser~ ra~sed .against prostaglandin methyl oximes and using H labelled prostagland~ns
which were methyl oximated. ..Endometrial stromal or glandular cells ~ncubated ~n E?t~lone showed
considerable variation in pros6aglandin F2 (PGF2~) produc wn .(stromal: median 0.7 ng per 10 6cells per ~4h, range (0.3-3.4),glandular; median 5.0 ng per 10 cells per 24h, range 1.7~14~7!.
Incubation of stromal cells in the presence of P4 alone slgn~f~cantly
reduced PGF production at 100 nM (p< 0.05) an~ 1.0 uM (p<.0.02). Bycontrast c6~ture of endometrial stromal cells ~n RU 486 st~mulatedPGF pr~uction in a dose-related manner (p< 0.01; 300% of controlva16~ at 1.0 uM). RU 486 in the presence of 100 nM progesterone alsostimulated PGF production at all concentrations tested, although theincrease in pGt~ levels was lower than that seen with RU486 alone.The percentage 6~ live cells at the end of these incuba~i~ns w~s .equivalent between control and RU 486-treated cells. ~~mllar ~n~ubat~on
of glandular cells showed no increase in PGF2~ productIon follow~ng RU486 exposure. .. ,
We conclude (i) RU 486 stimulates PGF2 productlon ~n endometr~al
stromal but not glandular cells (ii) this ~gcrease is'd~se-dependentand competitively inhibited by P and (iii) RU 486 may 1ndu~e
menstruation by stimulating end~etrial stromal prostagland~n release.
(1) Healy, D.L., Baulieu, E.E. & Hodgen, G.D. Fertil. Steril. 40:253(1983) .
1.04 ± 0.062.30 ± 0.13
15 ±35 ±
Vmax (fmol min- 1 e- 1 )
68 ± 17154 ± 37
Table 1.
2-cellblastocyst
Glycine is key metabolite which enters many cells by a specificrestricted glycine-2 Na+ co-transport system. Provision of the Na+critical for Na+/K+-ATPase activity during mouse blastocyst expansionhas 'not been described. Mouse embryos contain high levels of glycine,possess specific glycine transport systems and may have access tohigh glycine concentrations in the luminal fluids. We examined theglcyine dependence of glycine uptake and its requirement for Na+ in 2cell embryos and blastocysts to assess possible contribution ofglycine uptake to Na+ uptake.
Embryos were collected from superovulated mice 48 h or 96 h posthCG by standard techniques. 3H-glyc ine uptake was assessed over10 min at 37°C in media (M2) with glycine concentrations from 690,000 11M. Na+-dependence of 3H-glycine (1 mM) uptake was measureduE.ing M2 medium depleted of Na+ by replacing NaCl and sodium lactatewith choline chloride, NaHC03 with KHC03 and adjusting the pH withKOH. Media with various K+ concentrations were obtained by varyingthe NaCl/choline chloride. Kinetic parameter~ were estimated byfitting functions to the untransformed uptake data by non-linear leastsquares regression using a PDP-11 minicomputer.

97
THE INTERCOTYLEDONARY ENDOMETRIUM OF THE EWE DURING PREGNANCYPROGESTERONE CONTENT, PROTEIN SECRETION AND ULTRASTRUCTURE
Y. Tanga
, G.M. Stonea
, x. zhangb
and B.G. Millerb
University of Sydney: Departments of Veterinary Physi~logy,
Sydney, 2006a
and Animal Husbandry, Camden, 2570
Plasma P Tissue Tissue P Protein Protein(ng/ml) (ng/g) synthesis secretion
(dpm/~g DNA)
Pregnant 29.9 :!:: 2471 .:!:: 424 :!::horn 3.47 214 34
11.2 :!::1611 :!:: 276 :!::2.48 Non-preg- 13.4 :!::
nant horn 1. 91 134 60
J. Anim. Sci. 2l:Suppl. 1, 307.
B.G. et al., (1983) J. Endocrin. 96:137-46.B. G. et al., (1984) Reproduction in Sheep p. 134, Aust.of Science.R.J. et al., (1980)
Miller,Miller,AcademyMoffat,(3 )
(1)
(2 )
Table 1 shows that each parameter measured in the endometrium issignificantly greater in the pregnant horn ,(P < 0.05). In this tissuethere is a significant negative correlat1on (r = 0.61, P < 0.05)between protein secretion and P content. When data for the. pregnanthorn endometrium of animals between days 92-99 are compared w1th.tho~e
for animals between 109 - 124 there is a significant (P < 0.05) r1se 1ntissue P associated with its expected rise ,in plasma. ~ur1ng th~S
period there is no change in protein synthes1s o~ secret1~n. The",eresults indicate that although endometrial prote1n, secretlon dunngpregnancy is P dependant (3) the rate cannot be dlrectly related totissue P concentration at that time.
Protein secretion by the intercotyledonary endometrium risesmarkedly from day 28 of pregnancy to reach a plateau from about day 84until at least day 112. The concomitant rise in t,issue, nuclearprogesterone (P) receptor content suggests the secretion 1S P dependant( 1 ) . Ligation of the non pregnant horn of ewes at days 5-6 ~f pregnancy is followed by the accumulation of a large volume o~ flu1d, r1chin secreted protein (2). It was of interest to exam1ne rates ofprotein synthesis and secretion b~ endometrium, from pregnant andnon-pregnant horns of such animals 1n terms of t1ssue P content, andalso to examine their ultrastructure. of
Hemi-pregnant ewes were killed between days 92, and 124pregnancy, plasma was collected and porti~ns of endometr1um wer~ takenfor estimation of tissue P concentrat1on, prote1n synth~sls andsecretion (1) and electron microscopy. P in plasma and t1ssue wasmeasured by specific RIA.
No consistent differences in the glandular and surface epitheliaare observed between the two horns indicating that ligation does notcause changes in the ultrastructure of thes~ tissues.
TABLE 1. Plasma and tissue P and in vitro protein synthesis andsecretion by endometrium
H. Gardner and P.L. Kaye
CHARACTERIZATION OF THE MOUSE BLASTOCYST GLUCOSE TRANSPORTER
Department of Physiology and Pharmacology, University of Queensland,St Luica, Queensland, 4067
The mechanisms by which glucose enters the mouse blastocystremain virtually undefined despite the fact that glucose is a keymetabolite. Although not required for energy in the pre-compactionmouse embryo, early evidence suggests that glucose is taken up by afacilitated diffusion system (1). Since transporter activity mayregulate glucose metabolism we proceeded to characterize the glucosetransporter in blastocysts using the non-metabolizable analogue 3,0methyl glucose (3OMG) to dissociate transport from metabolism.
Blastocysts collected by standard methods from super-ovulatedmice, 96 h post-hCG, were held in modified M2 containing 5.5 mM Dglucose, and then washed extensively in glucose-free medium beforeuse. Uptake of 3H-30MG at 5.5 mM and 25 mM (1-2 mCi/ml) at 22 and37°C was determined. In all cases the rate of uptake was constant forat least the first 7 min. Therefore uptake over 5 min was selected tocalculate initial uptake rates, for temperature, efflux and diffusionstudies. 30MG uptake was determined at 4-37°C in 5.5 mM (3OMG) and aQ10 of 3 was obtained. This is much higher than expected fordiffusion, but comparable with those characterisitc of facilitatedtransport. Trans-acceleration and efflux are characteristic of acarrier mediated system and were readily demonstrated with nonmetabolizable substrates. Blastocysts were loaded with 3H-30MG for5 min at 37°C, briefly washed, and. transferred to non-radioactivemedium containing 0, 5, 25 and 40 mM 3OMG. The rate of efflux over15 min was determined. When no external 30MG was present the 3H-3OMGcontent of the blastocyst declined rapidly. Increased concentrationsof 30MG in the medium accelerated this efflux. In all cases 95% ofthe 3H-30MG initially present effluxed within 12 min at 370C. At 40Cthis efflux was reduced to the extent that only approximately 5% hadeffluxed after 10 min. The relative contributions of diffusion andfacilitated transport to the total 30MG uptake were determined bycomparing the uptake at 22°C in the presence of cytochalasin-B (aninhibitor of glucose transport) with standard incubation at 40C.Cytochalasin-B inhibits uptake at 22°C by about 20%, and incubation onice shows a similar degree of inhibition.
The results show that blastocysts accumulate 30MG predominantlyby a facilitated system. Diffusion accounts for about 20% of thetotal, which may be significantly different from the situation in precompaction embryos (1). This may reflect either the structuralchanges that Occur with compaction and cavitation, or the greatlyincreased glucose metabolism.
(1) Wales, R.G. & Brinster, R.L. (1968) J. Reppod. Fepc. 15: 415-422.

98
Dept. Immunology, John Curtin School, GPO Box 334, Canberra.
Belinda L. Drake & John C. Rodger
12/172/15
13/1611/17
Proportionof ewes
4444
Maintenance Ptransfer (days)
4624
Age of ewe atPost-oestrus
99
N.W. Moore and B.G. Miller
EFFECT OF TIME OF TRANSFER ON SURVIVAL OF EMBRYOSIN OVARIECTOMIZED EWES
Department of Animal Husbandry, University of Sydney, Camden, 2570
1 - Control2 - Delay transfer3 - Advance transfer4 - Delete part of Maint.P
Embryo transfer has clearly demonstrated the need for close synchrony between the "post-oestrous ages" of embryos and the reproductivetracts to which they are transferred. Embryos will tolerate a degreeof asynchrony when progesterone is given during the first few daysafter oestrus in entire ewes (1) or when the doses of progesteronegiven after an induced oestrus in ovariectomized ewes are varied (2).However, in all studies the effects in the recipient of post-oestrousage and time under the influence of progesterone have been confounded.
In this study progesterone/oestradiol (piE) treated ovariectomizedewes were used and the time of transfer of Day 4 embryos (Day 0 = dayof oestrus) after the induced oestrus was varied (Table 1). otherwise,the regimen of Priming P, Oestrous E and Maintenance P used was thatknown to support the synchronous transfer of Day 4 embryos (2,3). Eachewe received 2 embryos and their survival and development was assessedat slaughter 16-17 days after transfer.
TABLE 1. Proportion of ewes with normal embryos.
(3)(3)
(3)o
On Covers lips
o2.5 - 6
5.3 - 10 (3)
*
Phagocytic Cells
of experiments *not present
Percentage OfIn Suspension19 - 12 (3)#10 - 15 (3)
<1 (3)
2.9 - 4 (3)
o (3)
o (3)
assay (1) #number
Cell TypeMacrophagePMNTrophoblastMacrophagePMNTrophoblast
Staph. aureusParticle
OpsonisedSRBC
§Modified macrophage
At implantation, the invading trophoblast engages in activephagocytosis of maternal uterine tissue and blood cells. However,the role that this activity plays in the establishment of asuccessful pregnancy is not known. Cultures of disaggregatedplacental tissue are frequently used to examine trophoblastimmunobiology, although its phagocytic activity has not beensystematically studied in vitro. This study set out to establish thein vitro phagocytic properties of murine trophoblast cells frominvasive (ectoplacental cone, d7 gestation) and mature (d12-16gestation) placental stages, using cells in suspension, culturedovernight on glass covers lips and, in the case of ectoplacental cone(EPC), embedded in a clot of maternal blood or plasma.
TABLE: 1. Percentage of phagocytic cells in preparations 8fdisaggregated mature placenta after I hr incubation at 37 C §
PHAGOCYTIC PROPERTIES OF MURINE TROPHOB..::.AST IN~
A very small percentage of trophoblast cells «1% total) werecapable of limited phagocytosis in suspension cultures, but therewas no evidence of phagocytosis by trophoblast cultured overnight oncoverslips (Table 1). In contrast, placental cells of the monocytemacrophage lineage were markedly phagocytic when maintained insuspension or cultured overnight on CQverslips. Surprisingly,despite its well established activity in vivo, trophoblast from theinvasive EPC stage did not demonstrate-any phagocytosis when grownon glass covers lips for short (0-48 hrs) or long « 6 days) periods,These cells did, however, phagocytose maternal red blood cells andsperm when invading into blood and plasma clots during the 24 hrincubation period.
Therefore, under appropriate experimental conditions, trophoblastcells will demonstrate phagocytic activity in vitro in a mannerthat reflects their in vivo potential. Further experiments will usethe blood/plasma clot system to investigate the role of trophoblastphagocytosis in the reproductive and transplantation biology ofmurine pregnancy.
(l) Van Furth, R. & Diesselhoff-Den Dulk, M.M.C. (1980) Scand.J o Immunol: 12 265-269.
Delaying the start of Maintenance P by 2 days so that transferswere carried out on the 6th day after oestrus had a marked effect ondevelopment of embryos (Group 2 v 1; X2 = 10.6; P<O.Ol), whereas advancing the start of Maintenance P by 2 days (transfer 2 days afteroestrus - Group 3) had no effect. Embryos do not develop normally ifOestrous E is omitted, or if the start of Maintenance P is advanced by5 days in relation to oestradiol treatment (3). There seems to be alimit of a few days during which the oestradiol stimulated uterus canrespond to progesterone and establish an environment suitable for thesurvival of Day 4 embryos. The omission of the first 2 days of Maintenance P was without effect (Group 4 v 1), suggesting that the initialdoses of progesterone are too small to influence uterine function andthat progesterone treatment for' as little as 24 hours renders theuterus receptive to Day 4 embryos. When the time relationships betweenOestrous E and Maintenance P are satisfied, it appears that the dose ofprogesterone given at around the time of transfer of Day 4 embryos iscritical for their survival.
(1) Lawson, R.A.S. and Cahill, L.P. J. Reprod. Fert. , 67 : 473-475(1983) .
(2) Miller, B.G. and Moore, N.W. Aust. J. BioI. Sci. , 29: 565-573(1976) .
(3) Miller, B.G., Moore, N.W., Murphy, L. and stone, G.M.Aust. J. Biol. Sci. , 30: 279-288 (1977) .

101
irregular lumen.Region Caput Proximal Distal Cauda
Cor1>us Corous
Segment No 1 2 4 5 8 9 11 12
Tubule 90.35 86.04 102.91 92.78 94.94 104.58 137.35 132.70
Diameter (\1) ±5.79 ±7.82 ±8.94 ±II. 25 ±7.48 ±4.91 ±8.20 ±7.56
Epithelial 25.44 25.39 20.54 19.58 26.84 28.24 33.81 32.44
Height (\1) ±2.63 ±3.18 ±2.90 ±2.6 ±4.10 ±2.82 ±27.82 ±26.60
The ep~thelium lining the duct consisted predominantly of principalcells which were generally similar in structure throughout, althoughlarge numbers of granular inclusions occurred in principal cells in thecaput and proximal corpus regions. Stereocilia lined the luminalsurface of these cells except in caudal segments where they werereplaced by a brush border of microvilli. No evidence of cytoplasmicdroplet phagocytosis by principal cells was observed in this species.Basal cells were found throughout the tract but were most abundant inthe distal corpus. Clear cells were also present in low numbe.rs ~utwere most numerous in the caudal region. The few mitochondr~a-r~chcells which were seen in caput and corpus segments were morphologicallysimilar to those in other mammalian species. "
The epididymis of AntechinuB is distinguished from other marsupialsby the unusual structural organization of the duct in the caudal regionand by the absence of cytoplasmic droplet phagocytosis by the prin:ipalcells. It remains to be seen if these differences are characterist~c ofthe Dasyuridae.
STRUCTURAL FEATURES OF THE EPIDIDYMIS IN A DASYURID MARSUPIALD.A. Taggart and P.D. Temple-Smith
Department of Anatomy, Monash University, Clayton, Victoria 3168
Few detailed studies have been made of epididymal structure in"marsupials. However, from these studies the structural organization ofthe marsupial epididymis appears generally similar to that of othermammals, although some structural features are unique to the gr~up.
This study describes the structure of the epididymis in Antechtnusstuaptiia small forest dwelling member of the Dasyuridae, a family inwhich no 'detailed structural observations have been made on epididymis.
Four sexually mature A. Btuaptii males were used in this study.Each was perfused with buffered glutaraldehyde fixative and theepididymides were removed, subdivided into 12 regions, and embedded inplastic for light (LM) and electron microscopy (EM). Thick sections(1 \1) from each region were stained in toluidine blue for 1M. Frompreliminary observations of these sections 8 regions were then selectedfor further examination. Tubule diameters and epithelial heights weredetermined from 50 transverse sections of the duct in each region peranimal using a Leitz ASM. Thin sections were stained in lead and uranylstains for EM studies.
Macroscopically the epididymis of AntechinuB resembled that ofother scrotal mammals and caput, corpus and caudal regions were readilyidentified. Tubule diameter and epithelial height were relativelyconstant throughout the epididymis except for the caudal regions where asignificant increase was observed. In these regions epithelial heightwas extremely variable (see table), and was associated with a narrow and
(1) Sadleir, R.M.F., and Tyndale-Biscoe, C.H. BioL Reprod. 16:605-508 (1977) •
(2) Hinds, L.A., and den Ottolander, R.C. J. Reprod. Fert. 69:631-639 (1983) •
(3) McConnell, S.J. and Tyndale-Biscoe, C.H. J. Reprod. Fert.73: 529-538 (1985).
(4) Bittmann, E.L., and Karsch, F.J. BioI. Reprod. 30: 585-593(1984) .
100
C.H. Tyndale-Biscoe, S.J. McConnell and L.A. Hinds
CSIRO, Division of Wildlife and Rangelands Research, Lyneham, A.C.T.
After its formation in February the corpus luteum (CL) of thetammar wallaby is tonically inhibited until after the summer solsticein December. From the winter solstice in June increasing daylength isthe proximate factor in this inhibition and it can be mimicked by anartificial photoperiod of 15 h Light:9 h Dark (15L:9D) (1). Increasingthe dark phase by 3 h to 12 L:12 D caused reactivation of the CL andembryo with birth 32 d later (I, 2), and a corresponding increase induration of elevated plasma melatonin (3). Daily injections ofmelatonin administered 2.5 h before lights out on a photoregimen of15L:9D, mimicked the light change to 12L:12D and bIrths occurred 32 dlater (3). These results indicated a role for melatonin in photoperiod response of the tammar but did not differentiate between asensitivity to elevated melatonin at a particular time of day (19002200 hrs) from a change in duration of elevated melatonin due tosummation of the exogenous and endogenous melat~nin. We report theresults of 2 experiments designed to differentiate these twohypotheses. To test the sensitive period hypothesis, 8 quiescent nonlactating tammars were exposed to 24L:OD (days 0 to 14), 14L:9D (days15 to 34) then ambient daylength of -13L:IID on day 35. From day 0 to22 the tammars were injected subcutaneously each day at 19.30 h witheither melatonin (400 ng/kg; N=4) or the arachis oil vehicle (N=4).The nocturnal melatonin elevation was abolished by 24L:OD. Themelatonin-treated tammars <jave birth and/or came into oestrus on day45 + 0 (mean + s.e.m.), that is to say, 31 days after the change to15L~9D. Thus-the tammars failed to respond to the melatonininjection alone, but did so when this was combined with the endogenousmelatonin elevation of 15L:9D as previously reported (3). The controltammars remained quiescent until reactivated by the decrease in daylength from 15L:9D to -13L:IID on day 35. To test the durationhypothesis 6 non-lactating quiescent tamrnars were exposed to aphotoregimen of 15L:9D (days -15 to -1), then 12L:12D (days 0 to 24)by extending the dark phase by 3h in the morning. This extended theplasma melatonin profile by 2 - 3h and the tamrnars gave birth on day30.2 ~ 0.2. Together, these results suggest that, as in the sheep(4), the duration of the nocturnal melatonin rise mediates theresponse of the tammar to photoperiod ch~,ge.
RESPONSE OF THE TAMMAR WALLABY TO CHANGE IN DURATION OF DARK PHASEAND/OR MELATONIN INJECTION

102
R.C. Jones, K.C. Worrard and B.P. Setchell
PROTEIN SECRETION BY THE EPIDIDYMIS OF THE TAMMAR, MACROPUS EUGENII
103
OVARIAN FUNCTION AND FEMALE SEXUA~ RECEPTIVITY DURING THE BREEDINGSEASON IN THE GREY HEADED FLYING FOX Ptepopus potioaephatus
Len Martin and Phil Towers
Department of Physiology and Pharmacology,University of Queensland, st Lucia 4067
(1) McGuckin, M.A. & Blackshaw, A.W. (1984) Ppoa. Aust. Soa. Reppod.Bio1,. 16: 23.
(2) Towers, P.A:"" & Martin, L. (1985) Aust. MamT7Yl1,. (in press)(3) Nelson, J.E. (1963) PhD Thesis, University of Queensland.
* mean ± SEM
P. potioaephatus is a seasonal breeder mating in March/April.Mechanisms controlling the breeding season are unknown, but mayinvolve day length. Males show seasonal changes in testis weight/testosterone levels (1) and body odour. In this study females wereshot in the wild. Plasma progesterone (P4) and oestradiol (E2 ) weremeasured by RIA, and uteri/ovaries examined histologically. Antralfollicles were classified as healthy or atretic (pycnotic granulosacells) and measured along 2 axes in the section containing the nucleusof the ovum (2).
In late pregnant/post-partum adults (August-November) no antralfollicles are present. By January-February most ovaries had manyfollicles 200-400 ~m (41 ;I; 6*, n=8) and some of 400-600 ~m (3 ± 0.3,n=6). No sperm were present in the uteri. In March, 13/14 bats hadovaries' with many follicles 200-400 inn (18 ± 2, n=26) 1 8/14 had someof 400-600 ~m (4.4 ;I; 2.4, n=6), and one a single follicle of 680 ~:
11 including the last animal, plus another with only one 200 ]Jmfollicle, had numerous sperm in the tract. No preovulatory follicles(>1500 \lm) or aopporo tutea (CL) were found. Peripheral E2 levelsranged from 5-25 pg/ml (15.0 ± 2.5 pg/ml, n=11), and showed nocorrelation with number/size of healthy antral follicles, or presenceof sperm in the tract. The mean value did not differ from that fornon-pregnant bats throughout the year (12.9 ± 1.2 pg/ml, n=53). Noneof the endometria were oestrogen stimulated. Peripheral P4 levelsshowed no change during the breeding season (March 6.3 ± 1.6 ng/ml,n=191 overall non-pregnant 7.8 ± 1.3 ng/ml, n=42). No pregnant batswere found before April.
OUr data suggest that. female sex hormones are unimportant ingenerating sexual receptivity in P. potiocephatus. In the wild, malesattempt copulation throughout the year, with intromission as late asJune 1 we have found abundant sperm in the tract of a female bearing anadvanced implanted embryo. Perhaps like humans, femaleP. potiocepha1,us are always 'receptive' , with acquiescence andfrequency of copulation determined largely by male drive, perseverenceand possibly pheromones. The absence of preovulatory follicles/CLsfrom females with sperm, suggests that follicle maturation/ovulationare not reflexly induced by a single copulation. Nelson's observation(3) that pairs copulate 'several times a day throughout' March-April,indicate repeated stimulation may be required. However, many femalesare still lactating in March and this may delay ovulation.
Proteinconcentration
mg/ml
Reabsorptionof fluid
%%
SpermatocritsSample Site
Jones, R.C., Hinds, L.A. and Tindale-Biscoe, C.H. (1984). Cell TissueRes. ~, 525-535.
Matsudaira, P.T. and Burgess, D.R. Anal. Biochem. (1978). 87, 386-396.
Rete testis 1.5 ± 0.2 0.9 ± 0.4
Ductus epididymidisProximal initial segment 11. 3 ± 2.5 87 3.0 ± 0.5Distal initial segment 50.8 ± 3.4 78 43.4 ± 7.7Distal middle segment 69.9 ± 4.5 27 32.2 ± 7.0Mid terminal segment 66.9 ± 1.6 0 18.2 ± 2.3Distal terminal segment 61.3 ± 2.6 -9 15.6 ± 2.2
Blood71.6 ± 3.7
(1)
TABLE 1. Concentrations of sperm and protein in the rete testis,epididymal luminal fluids and blood (means ± SE from 6 animals).
Epididymal luminal fluids were collected from tammars (1) anaesethizedwith Inactin (Byk Gulden Pharm, W. Germany) using micropuncture procedures. PAGE was carried out using a denatured linear gradient system (2).
This report shows that there are similarities between the tammar and therat and rabbit in the secretion and reabsorption of proteins in theepididymis.
Dept Biological Sciences, Univ. Newcastle, N.S.T~. and Dept Physiology,Waite Agric. Res. Inst., Univ. Adelaide, S.A.
(2)
Table 1 shows that the concentration of protein in rete testis fluid wasabout 1 % of that in blood, about half the protein leaving the testis isreab~o~b~d by the ductuli efferentes, considerable protein is secreted bythe ~n~t~al segment, and there is a net loss of protein from the middleand terminal segments.
There were 5 bands present in PAGE gels of rete testis fluid which werenot present in gels of blood (bands correspond to molecular weights, MW~f 14,700, 2~,~00, 24,100,43,200 and 44,800). There were 10 bands present~n gels of ep~d~dymal fluids which were not present in samples from bloodor rete testis fluid. Four (MW of 10,900, 12,800, 30,000 and 32,000) weresecreted by all segments of the epididymis, 2 (MW of 18 700 and 21 400) bthp. init~al and middle segments, 2 (MW of 39,800 and 90:600) by th~ middl~and term~nal segments, 1 (MWof 31,300) by only the initial segment and1 (MW of 56,300) by only the terminal segment.

105
B.A. Stone, P. QUinn, R.F. Seamark and S. Deam
RELEASE OF STEROIDS BY CULTtmED PIG BLASTOCYSTS
Heap, R.B., Flint, A.P.F., Hartmann, P.E., Gadsby, J.E., Staples,L.D., Ackland, N. & Hamon, M. J. Endocr. 89: 77P-99P (1981).Flint, A.P.F., Burton, R.D., Gadsby, J.E., Saunders, P.T.K. & Heap,R.B. Ciba Fdn. Symp. 64. Excerpta Medica, Amsterdam, 209-228 (1979).Stone, B.A. & Seamark, R.F. J. ReQrod. Fert. (in press). (1985).Dickman, Z., Dey, S.K. & senGupta, J. vito Horm. 34: 215-242 (1976).
(1)
(2)
(3)(4)
Dep3.rtment of Obstetrics and Gynaecolbgy, University of Adelaide
The cap3.city for dispersed pig blastocyst tissue to synthesize oestrogenfrom C21 and C19 steroids in vitro has been demonstrated (1). However,pig blastocysts cannot synthesize pregnenolone from acetate orcholesterol (2), so the nature of the dependence of porcine blastocystson maternal C21 steroids as precursors for de novo steroid synthesis isuncertain. In view of recent evidence for accumulation of pregnenolonesulphate in uterine fluid of early pregnant sows (3), this. ster~d
conjugate may be an important precursor to ~strogen synt~Sl.S by p~g
blastocysts. This study thus examines secretl.on of sterol.ds by pl.gblastocysts cultured in media supplemented with pregnenolone sulphate.
Embryos were recovered from sows at slaughter, and were cultured in amedium based on MEM, supplemented with 25 mM sodium bicarbonate a:nd 10%(v/v) human serum. In the first experiment, pregnenolone sulphate wasprovided in culture media at final concentrations of 10.4, 12.8, 17.0 a~d
4470 nM. Eleven embryos (5 days post-coitum, p.c.) were cultured for Sl.Xdays in each medium, after which culture media were recovered. Rates ofrelease of pregnenolone and progesterone were calculated from theiraccumulated levels in each culture medium, as determined byradioimmunoassay of chromatographed extracts (3). calculated rates ofrelease of pregnenolone by single blastocysts increased (from 0.43 to 1.7pool/day) as the concentration of pregnenolone sulphate in the culturemedium increased (from 10.4 to 4470 nM), respective rates of release ofprogesterone being 0.07 to 0.38 J?ffiOl/day. . ...
A further experiment examl.ned the effect of l.nhl.bl.tl.on of 3-8 HSD(with Trilostane: 93.6 uM) on the release of six steroids by blastocy~ts
(D:l.y 3-6 p.c.), cultured for four days in a medium supplemented Wl.thpregnenolone sulphate to 1.0 uM. Levels of the steroids wer7 determinedin the recovered media (3). calculated rates of sterol.d release byblastocysts (fool/day: mean ± s.e.m.) cultured in control and inTrilostane-supplemented media were, respectively: pregnenolone (315 ±246,813 ± 679): progesterone (44.5 ± 26.5,1.3 ± 0.7): androstenedione(26.3 ± 20.8,5.9 ± 3.5): testosterone (26.8 ± 10.1,0.0), oestrone (0.8+ 0.4, 1.8 + 0.6), and oestradiol (9.2 ± 3.3,9.6 ± 4.2).- These r;sults verify sulphatase activity in intact pig bIastocysts,and show release of progesterone, andros tenedione, testosterone, oestroneand oestradiol by blastocysts cultured in the presence of pregnenolonesulphate. Lower rates of progesterone release in the .presence ,?fTrilostane confirm earlier evidence for activity of 3-8 HSD l.n early pl.gembryos (4). In view of the abundance of pregnenolone sulphate inuterine fluid of early pregnant sows (3), the results also suggest thatthis C2l steroid sulphoconjugate may be a significant precursor tosteroid synthesis by preimplantation pig embryos in vivo.
THE ESTABLISHMENT OF A COMMON MARMOSET COLONY (CALLITHIRIX JACCHUS)AND OBSERVATIONS ON THE CIRCADIAN REST-ACTIVITY CYCLE USING AMICROCOMPUTER
~·E: Matthews, K. Porter, I. Cooke. M. Guerin, D. Bigham
104
The Department of Obsetrics and Gynaecology University of AdelaideS.A.
The Common Marmoset has been shown to be an excellent model forreproductive and other biomedical studies. It is a prolific breederin captivitiy, has an ovarian cycle of 16 days. a gestation period of140 days, a high frequency of multiple births, no lactationalanoestrus and breeds non seasonally.
Si~ proven breeding pairs of Marmosets were imported from InterSimian (U.K.) in November 1983. The holding conditions included anambient temperature 24°C - 27°C, humidity 45%. 2 x 40 watt fluorescenttubes providing 256 Lux of lighting at cage level on a LD 12:12schedule (0700-1900 hours) with supplementary V·V light 60 mins daily.
Each animal received 15g primate meal (Milling Industries), 15gHarpers Greyhoud chow, 1/3 banana supplemented by 1/8 orange or appledaily. with water ad Lib. Double cages (100x120x160cms) were used forholding family groups, each cage being thoroughly cleaned each weekusing Biogard disinfectant.
During the 18 month period to June 1985 a series of 20 birthstook place including 7 pairs twins, 7 triplets and 4 quadruplets. Thefrequency of quadruplets was 20%, of triplets 35.0% and of twinpregnancies 35.0%.
Twenty five infants were lost at birth or within 24 hoursincluding 3 sets of triplets and the remainder were 3rd and 4thinfants. One infant (from a triplet) was hand reared and a furtheranimal was collaboratively raised with a sibling. The-birth ratio was16:12 males:females. The delivery-delivery interval in 9 animals wasless than 160 days. Ovulation therefore was assumed to take place 1014 days following parturition and during active lactation.
The rest activity cycle of one cage of 6 marmosets was initiallymonitored employing a loud speaker as a microphone together with a twostage amplifier in order to display activity on a CR5 Trilek singlechannel recorder with an intermittent on-off switch to control motoractivity to 40 msec recording every 460 msec. Initial observationsunder the 12:12 L.D. cycle indicate the diurnal nature of the Marmosetstrictly entrained to the LD Zeitgeber with the onset of the activityphase coinciding with artificial dawn and the offset anticipating duskby approximately 1 hour.
More recently a microcomputer based monitoring facility has beenes~ablished allowing the sampling of the rest activity ,of individualanlmals over a 7 day time span before processing the data andgraphical display. The facility has the potential to recordadditional parameters as they are interfaced.
Experiments are now in progress to determine whether the periodof the rest-activity cycle is less than or greater than 24 hours.

106
EFFECT OF UTERINE P~OTEINS ON EMBRYO METABOLISM
N.K. Khurana and R.G. Wales
School of Veterinary Studies, Murdoch University, WA, 6150.
The metabolism of cultured mouse embryos appears to be differentfrom that of embryos developing in vivo. However, little is known
. of the effects of uterine secretions on embryo metabolism. Thepresent study was undertaken to test the effect of various molecularfractions obtained from uterine flushings on CO 2 production by mouseembryos collected on day 4 of pregnancy.
Uteri were flushed with 0.9% NaCl on days 3, 4 and 5 ofpregnancy or pseudopregnancy. The fluid from several animals waspooled and the macromolecules present in each pooled sample fractionated into four molecular weight ranges by ultrafiltration, viz.1,000-10,000 (fraction A), 10,000-100,000 (fraction B), 100,000300,000 (fraction C), 300,000+ (fraction D) daltons. Each fractionwas concentrated and the filtrate vehicle (0.9% NaCl) replaced withculture medium. The 1~C02 produced (1) over 4 hr by 10 embryosincubated in medium containing 0.28 mM [U-l~C]glucose was measuredwith and without the inclusion of uterine fractions. Data for fourreplicates are presented in Table 1.
TABLE 1. Changes in CO 2 production from glucose by mouse embryoswith the addition of different fractions of uterine flushings~
Values are the mean (± s.e.m.) per cent change from control values.
Fraction addedDonor status A B C D
Pseudopregnant Day 3 93 ± 5 106 ± 9 86 ± 13 74 ± 4Pregnant Day 3 103 ± 9 105 ± 9 102 ± 5 54 ± 3
Pseudopregnant Day 4 112 ± 8 66 ± 3 106 ± 3 58 ± 3Pregnant Day 4 98 ± 12 74 ± 6 99 ± 11 54 ± 14
Pseudopregnant Day 5 62 ± 7 75 ± 7 95 ± 17 54 ± 4Pregnant Day 5 77 ± 9 71 ± 6 59 ± 3 77 ± 15
In general, the response to flushings from pregnant and pseudopregnant animals was the same. Except for the high M.W. component(fraction D), Day 3 fluids had no effect. On day 4, fraction Bas well as fraction D was depressant whilst on day 5 all fractions(except fraction C from pseudopregnant animals) suppressed CO 2production by 25 to 50%. The suppressing effect of uterine flushingsreported in this study is in line with the observations of otherworkers (2-3) who reported marked inhibition of incorporation ofradioactive substrates in the presence of uterine secretions.
1. Brinster, R.L. (1967) Exptl. Cell Res. 47: 271-277.2. O'Neill, C. and QUinn, P. (1981) J. Reprod. Fert. 62: 257-262.3. Weitlauf, H.M. (1976) BioI. Reprod. !±: 566-571. --
107
INCORPORATION OF LABELLED AMINO ACIDS BY SHEEP EMBRYOS
R.G. Wales and C.L. Cuneo
School of Veterinary Studies, Murdoch University, WA, 6150
Between day 13 of pregnancy in the sheep and the initiationof implantation around day 18-19, considerable structural differentiation of the embryo occurs. Organogenesis is initiated withinthe embryo itself and in turn the yolk sac and allantois emergeas the trophoblast expands to occupy the uterine cavity. In thep~esent study, the incorporation of labelled amino acids was ~sedto estimate protein synthesis by each of these structures durlngthis period.
Embryos were flushed from the uteri of ewes on day 13, 15,17 and 19 of pregnancy and dissected into their component structures. The embryo and samples of the membranes present wereincubated 2.5 h in media supplemented with an equimolar mixtureof l~C-labelled amino acids. After incubation, embryonic sampleswere recovered, lyophylised, weighed and extracted with perchloricacid. On the basis of available evidence, label in the acid solublesupernatant was equated with uptake into precursor pools whilstincorporation into the acid insoluble pellet was considered asan index of protein synthesis. The results for incorporation intoprotein are presented in Table 1.
TABLE 1. Amino acid incorporation into protein by the variousembryonic structures present on days 13, IS, 17 and 19 of pregnancy.Values (ng atoms of amino acid carbon/mg dried wt/hr) are means± s.e.m. (no. of observations).
Day of Incorporation into:
Pregnancy Embryo Trophoblast Yolk sac Allantois
13 108 ± 14 (5) 6 ± 2 (6)
15 60 ± 26 (5) 7 ± 1 (7) 54 ± 21 (5)
17 30 ± 7 (5) 17 ± 6 (6) 47 ± 14 (8)
19 8 + 3 (5) 7 (1) 20 + 6 (6) 30 + 5 (7)
Incorporation of amino acids into the macromolecules of theembryo fell progressively per unit weight as developmen~ progressed.This fall in incorporation was greater than the change ln uptakeinto the precursor pool, indicating a true reduction in proteinsynthesis. The extraembryonic membranes differed 'considerablyin their profile of protein synthesis. The trophoblast had arelative low, and constant, rate of protein synthesis. The yolksac which emerges on day 15, had synthetic rates 7-8 times thoseof ~he trophoblast. By day 19, its rate of protein synthesis had
. fallen threefold. At this time an active allantois is emergingand this structure was the most active present at the time of theinitiation of implantation. The present results show that thereis both a divergence in synthetic activity between structures anda change in synthetic rate of individual structures as developmentprogresses.
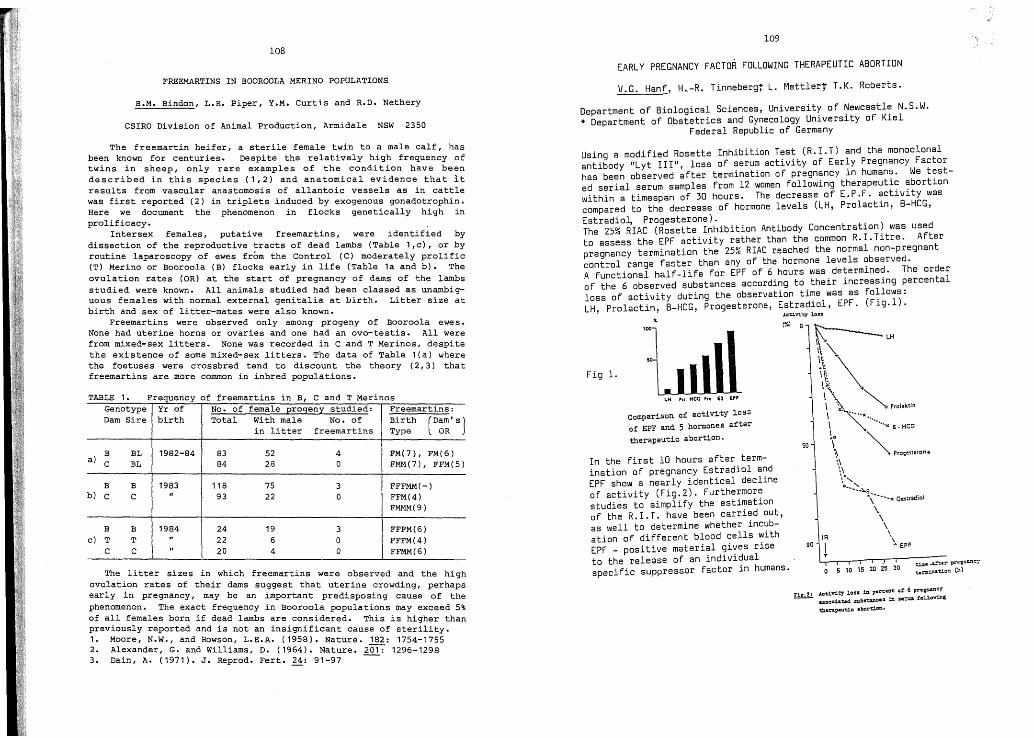
109108
EARLY PREGNANCY FACTOR FOLLOWING THERAPEUTIC ABORTION
FREEMARTINS IN BOOROOLA MERINO POPULATIONSV.G. Hanf, H.-R. Tinnebergf L. Mettlert T.K. Roberts.
B.M. Bindon, L.R. Piper, Y.M. Curtis and R.D. Nethery
CSIRO Division of Animal Production, Armidale NSW 2350
Department of Biological Sciences, University of Newcastle N.S.W.* Department of Obstetrics and Gynecology University of Kiel
Federal Republic of Germany
The. litter sizes in. which freemartins were observed and the highovulat~on rates of the~r dams suggest that uterine crowding, perhapsearly ~n pregnancy, may be an important predisposing cause of thephenomenon. The exact frequency in Booroola populations may exceed 5%of a~l females born if dead lambs are considered. This is higher thanprev~ously reported and is not an insignificant cause of sterility.1. Moore, N.W., and Rowson, L.E.A. (1958). Nature. 182: 1754-17552. Alexander, G. and Williams, D. (1964). Nature. 2017 1296-12983. Dain, A. (1971). J. Reprod. Fert. 24: 91-97 ---
'tiJD• ..a.!"":*r prtJ.gllllnc:t1:....i.n&t.!.on (11)o 5 10 15 20 2S 30
IR
IT
90
(%10~
\(\." '"\~ \\\~:'... ..~... Prolaktin
\ ·····x\ ········.x ~,HeG
SOl \ ~ Pro.esteron.
',\\\.\'
O"':'~~\""'<lOu"at!iol
\\\
\
"• EPF
'OOJ. .,1~LJllll
LH ,,1 HeG "0 £2 EPF
comparison of activity los::l
of EPF and 5 hormones after
therapeutic abortion.
In the first 10 hours after termination of pregnancy Estradiol andEPF show a nearly identical declineof activity (Fig.2). Furthermorestudies to simplify the estimationaf the R.I.T. have been carried aut,as well to determine whether incubation of different blood cells withEPF - positive material gives riseto the release of an individualspecific suppressor factar in humans.
Fig 1.
!:!!d!. .lct.ivi::r lass in p.rc.,,1: 0' 6 pr.pa.noyu:socia:ud .5U,bs'tanCc:s in se::u= !aUo'Wing
the.rapeutic abortion.
Using a modified Rosette Inhibition Test (R.I.T) and the monoclonalantibody "Lyt III", loss of serum activity of Early Pregnancy Factorhas been observed after termination of pregnancy in humans. We tested serial serum samples from 12 women following therapeutic abortionwithin a timespan of 30 hours. The decrease of E.P.F. activity wascompared to the decrease of hormone levels (LH, Prolactin, B-HCG,Estradio~ Progesterone).The 25% RIAC (Rosette Inhibition Antibody Concentration) was usedto assess the EPF activity rather than the common R.I. Titre. Afterpregnancy termination the 25% RIAC reached the normal non-pregnantcontrol range faster than any of the hormone levels observed.A functional half-life for EPF of 6 hours was determined. The orderof the 6 observed substances according to their increasing percentalloss of activity during the observation time was as follows:LH, Prolactin, B-HCG, Progesterone, Estradiol, EPF. (Fig.l).
Frequency of freemartins in B C and T MerinTABLE 1.
The freemartin heifer, a sterile female twin to a male calf, hasbe~n kn~wn for centuries. Despite the relatively high frequency oftw~ns ~n sheep, only rare examples of the condition have beendescribed in this species (1,2) and anatomical evidence that itresults from vascular anastomosis of allantoic vessels as in cattlewas first reported (2) in triplets induced by exogenous gonadotrophin.Here we document the phenomenon in flocks genetically high inprolificacy.
Intersex females, putative freemartins, were identified bydissection of the reproductive tracts of dead lambs (Table 1,c), or byroutine laparoscopy of ewes from the Control (C) moderately prolific(T) Merino or Booroola (B) flocks early in life (Table 1a and b). Theovulation rates (OR) at the start of pregnancy of dams of the lambsstudied were known. All animals studied had been classed as unambiguous females with normal external genitalia at birth. Litter size atbirth and sex of litter-mates were also known.
Freemartins were observed only among progeny of Booroola ewe s •None had uterine horns or ovaries and one had an ovo-testis. All werefrom mixed-sex litters. None was recorded in C and T Merinos, despitethe existence of some mixed-sex litters. The data of Table 1(a) wherethe foetuses were crossbred tend to discount the theory (2,3) thatfreemartins are more common in inbred populations.
, osGenotype Yr of No. of female progeny studied: Freemartins:Dam Sire birth Total With male No. of Birth (D~~' s)in litter freemartins Type
a)B BL 1982-84 83 52 4 FM(7), FM(6)C BL 84 28 0 FMM(7) , FFM(5)
B B 1983 118 75 3 FFFMM(-)b) C C " 93 22 0 FFM(4)
FMMM(9)
B B 1984 24 19 3 FFFM(6)c) T T " 22 6 0 FFFM(4)
C C " 20 4 Q FFMM(6)

110111
RESPONSE TO HYPOXEMIA AFTER ADMINISTRATION OF NERVE GROWTH FACTORANTIBODIES IN FOETAL LAMBS
J.A. Schuijers, D.W. Walker, C.A. Browne and G.D. Thorburn
Department of Physiology, Monash University,Clayton, Victoria, 3168.
IDENTIFICATION OF C19- AND C21- STEROID CONJUGATES
IN THE MALE LIZARD TILIOUA RUGOSA
P.A. Huf, A.R. Bourne and T.G. Watson
Biological Sciences, Deakin University, Vic. 3217
(1) Lupo di Prisco, C. Chieffi, G., and Delrio, G. (1967).Experientia 23: 73-74.
Steroid conjugates have rarely been investigated in lowervertebrates. The testis of reptiles, one of the least studiedvertebrate groups, seem to produce mainly testosterone. In thelizard Lacerta sicula this steroid was present only in the conjugateform (1). OUr previous studies of the male lizard Ti1igua(Trachydosaurus) rugosa have identified epitestosterone as the majorCu- steroid. Testosterone was also found but in lesseramounts. The present study is concerned with the chemicalidentification of the major endogenous CJ.9- and C21- steroidconjugates in the male T. rugosa. Previously we have shown usingboth in vivo and in vitro techniques that metabolism of 14Cprogesterone produces significant amounts of steroid conjugates (2).Analysis of steroid conjugates in male plasma was carried out bothbefore and after hydrolysis (solvolysis and glucuronidasehydrolysis). Hydrolysed steroid conjugates were analysed usingthin layer chromatography (TLC) , high performance liquidchromatography (HPLC) with U.V. detection and gas liquidchromatography (GC). Steroid conjugates were analysed directlyusing HPLC with electrochemical detection. For the latter analysisp-nitropheny1hydrazone and related derivatives were prepared.
Analysis of male lizard plasma, using TLC and GC, showed thatthe major steroids (after solvolysis) were testosterone anddehydroepiandrosterone (DHA). Several unidentified steroids werealso found. Testosterone was the major steroid found in thesolvo1ysab1e fraction following HPLC analysis with a U.V.detector. Direct analysis of steroid conjugates with HPLC showed asignificant electrochemical response corresponding to DHAsulphate. Several other, as yet unidentified, steroid su1phatesappeared to be present. Enzymatic hydrolysis indicated therelative absence of steroids in the glucuronide fraction.Epitestosterone conjugates were absent or in relatively lowconcentration.
The use of electrochemical detection with HPLC is a relativelynovel application of this technique to the direct analysis ofsteroid conjugates in biological tissues. T. rugosa appears to besimilar to other vertebrates in regard to having significant DHAsulphate. However the concentration of testosterone sulphate inthese lizards is unusually high, and with similar data from ~
sicu1a indicates that lizards may be atypical in this respect. Therelative absence of epitestosterone conjugates may be related to thehigher concentration of this steroid (compared to its epimer) inperipheral plasma.
Compo Biochem. Physio1. 70B: 661-664.(2) Bourne, A.R. (1981).
Nerve growth factor (NGF) is a protein found in a large number oftissues. in widely va~Yi~g amounts. Its actions are still relativelyunc~rtaJ.n, although J.t J.S well known that it plays an important roledurJ.n~ growth and differentiation of sensory and sympathetic neurones.In thJ.s study, the effect of anti-NGF on the development of the nervoussystem in utero was investigated in foetal sheep.
Foetal lambs were inj ected intramuscularly with ovine anti-murineNGF antibodies at 80 days gestation (full term = 145 days). Antiserahad been raised in sheep against murine NGF which was shown to behomogeneous by high performance liquid chromatography. Control foetuseswere treated with normal sheep serum. At 130 days of gestation, eachewe was made hypoxic by allowing it to breath 9% oxygen and 3% carbondioxide in nitrogen for 60 minutes. The anti-NGF treated foetuses had asignificantly lower resting partial pressure of oxygen than the controlfoetuses. A fall in partial pressure of oxygen during hypoxia of 11mmHg was observed in both groups of fetuses. A 28% increase in arterialblood pressure and a 650% increase in circulating plasma noradrenalineconce~trations was .observed in the control group of foetuses duringhypoxJ.a, however J.n the anti-NGF treated foetuses there was nosignificant increase in either plasma noradrenaline concentration orarterial blood pressure. In the control foetuses there was abradycardia during the first 15 minutes of hypoxia, whereas this did notoccur in. the anti-NGF treated foetuses, and there were no significantchanges J.n heart rate either during or after the hypoxia.
The anti-NGF treated foetuses had resting plasma noradrenalineconcentrations which were twice those seen in the control foetusesThere.were no differences in the plasma concentrations of adrenaline 0;dopamJ.ne.
These results suggest that the cardiovascular and plasmacatechol~mine changes which occur during hypoxemia in the foetal lambare attrJ.butab~e to the peripheral sympathetic nervous system, and thatthe foetal oVJ.ne sympathetic nervous system is dependent on a nervegrowth factor-like substance for its normal development.
This work was supported by an N.H.&M.R.C. Program Grant.

112
ABERRANT SPERMIOGENESIS IN A FINCH HYBRID
M.A. Swan
Department of Histology & Embryology, Sydney University,
N.S.W. Australia
While closely related avian species hybridise readily, theirhybrids are usually sterile. Since some of the structure/functionrelationships of spermiogenesis are imperfectly understoodexamination of aberrant spermiogenesis in sterile hybrids ~ay helpto explain normal processes.
. A male hybrid of the spice finch (Lonchura punctulata,L~nnaeus) X chestnut breasted finch (Lonchura castaneothorax, Gould)was anaesthetised with Saffan and the testes were-removed. Thebird was killed with Lethobarb. A small sample of testis tissuewas taken for darkfield examination1 the rest was fixed fortrans~ssion electron microscopy in Karnovsky's fixative (1), withand w~thout 2% lanthanum.
. Sperm released into buffer and examined by darkfield light~croscopy were variable in appearance; some were of normal spiralap~earance while others were split into multiple tails, one ofwh~ch was sometimes straight while the others spiralled.
. The testes contained some spermatids which were degeneratingp~~or to n~clear condensation and were in groups connected by~sshapen ~ntercellular bridges. Examination of material processedw~th lanthanum showed that Sertoli-Sertoli junctions were tight;there was no auto-immune reaction to the spermatocytes in contrastto observations in canary x goldfinch hybrids (2).
Mature spermatids lay in sperm bundles in which some sperm-tailswere of nor~l appearance, containing one axoneme; the majority ofsperm conta~ned 2 to 4 axonemes per sperm-tail. These were insertedat mult~ple sites at the base of the nucleus. The accessory fibresdevelop~ng around these axonemes were at different stages ofmaturation. These anomalies could have been caused by incompletecytokinesis or by multiple centriole replication.
(1) Karnovsky, M.J. J. Cell Biol. 27: 137A (1965).(2) Swan, M.A. Gamete Res. (in press) (1985).
113
PROGESTERONE AND OESTRADIOL IN PERIPHERAL AND UTERO-OVARIANVENOUS PLASMA DURING THE PERIPARTUM PERIOD IN THE TAMMAR
L.A. Hinds, J.D. Harder, C.A. Horn and C.H. Tyndale-Biscoe
CSIRO Division of Wildlife and Rangelands Research, Lyneham, A.C.T.
In the tammar a single ovulation occurs from alternate ovaries andso the separate contributions of the corpus luteum (CL) and Graafianfollicle to peripheral levels of progesterone (P) and oestradiol (E)can be determined by sampling the venous effluent from each side. Pand E were measured by radio-immunoassay (1, 2) in plasma from theperipheral circulation and both utero-ovarian veins during latepregnancy, parturition, post-partum oestrus and ovulation; groups of 5animals were sampled between Days 23 and 29 after reactivation oflactation-delayed pregnancy.
Concentrations of E in the peripheral plasma and in the veindraining the CL remained < 5 pg/ml on all days except the day ofparturition (12.7 pg/ml and 14.3 pg/ml respectively). Mean E levelsin the effluent from the ovary bearing the Graafian follicle weresignificantly higher than peripheral levels on Days 24-27. Thehighest average concentration (36.5 pg/ml) occurred on Day 27 coincidentwith parturition and before oestrus. Mean levels of P in peripheralplasma and in the vein draining the Graafian follicle were similar onall days. P concentrations in plasma from the vein of the CL-bearingovary were 10 fold higher (> 5.0 ng/ml) than peripheral P levels(500 pg/ml) on Days 23 to 26 but on Day 27, after parturition, levelsdecreased to basal « 250 pg/ml) for all sampling sites. These resultsprovide strong evidence that the Graafian follicle is the main sourceof E and the CL is the main source of P in the peripheral circulationat this time.
A second series of tammars (n=5) were then sampled at 6 to 12 hintervals to determine the relationships between the decrease in P (3),rise in E (2), pre-ovulatory LH surge (3), oestrus and ovulation.Parturition occurred on Day 26 or 27 after reactivation, coincident withthe precipitous decline in peripheral P levels (> 400 pg/ml to< 250 pg/ml). E rose from < 10 pg/ml to a peak of 13 to 32 pg/ml within8.3 h and oestrus occurred by 9.8 h after the drop in P. The LH surgewas dependent on the E rise and followed it by 7 h. Ovulation followedmating by about 30 h and the LH surge by 24 h. Peripartum changes inperipheral E do not appear to be involved in the initiation ofparturition but the E rise and associated change in the E:P ratio areimportant signals for post-partum oestrus and ovulation.
(1) Sernia, C., Hinds, L.A. and Tyndale-Biscoe, C.H. J. Reprod. Fert.60: 139-147 (1980)
(2) Harder, J.D., Hinds, L.A., Horn, C.A. and Tyndale-Biscoe, C.H.J. Reprod. Fert. 72: 551-558 (1984)
(3) Tyndale-Biscoe, C;H. Hinds, L.A. Horn, C.A. and Jenkin G.J. Endocr. 96: 155-161 (1983).

114115
PHOTOPERIOD INFLUENCES REPRODUCTION IN THE MALEFRUIT BAT, Pte~opus poZiocephaZus
M.A. McGuckin and A.W. Blackshaw
Department of Physiology and Pharmacology,University of Queensland, St Lucia 4067
P. poZiocephaZus is a sh t d bor - ay reeder, mating during autumn(March/April) and showing marked seasonal changes in testis weight,andtestosterone levels (1).
Three adult male P. poZiocephaZus were placed in constant longphotoperiod (16L:8D) during June (day 0), the time of testicularregr~ssion in the wild, kept at a constant temperature and fedad Z'Lb. Long photoperiod was maintained unt~l... two months before thesummer solstice (day 137) then decreased gradually (average3.~ min/day) fo~ 120 days until reaching 9L: 16D on day 257 from whichpo~nt photoper~od was held constant. Testis dimensions and bodywe~ghts were measured about every 10 days, and blood samples weretaken less f~equentlY from day 137 onward. Testis vOlume wascalculated us~ng the formula for a prolate spheroid and plasmatestosterone levels were estimated by RIA.
Table 1: Total testicular volume, body weight and testosteroneconcentration of three adult male P. poZiocephaZu8 confined to theexperimental photoperiod regimen. Mean ± s.e.m.DAY TESTES VOLUME BODY WEIGHT TESTOSTERONE
(CM3) (G) CONC. (NG/ML)a 4.29± 0.42 b* 597±11 (Day 137)
~~~ ::~: : ~:~~ ~: ~~~ : ~~ b* 1~:~~ : ~:~~ b*288 6.14 ± 0.25 a** ,b* 728 ± 57 8.74 ± 1. 23 c*308 5.70 ± 0.21 a**,b* 724 ± 48 5.47 ± 1.49 ~:*(vs day a = a, vs Qay 204 = b, vs day 137 = c; *p < 0.05, **p < 0.01)
.Long. photop.eriod cause~ a ,small but non-significant increase intest~s ~~ze wh~ch was ma~nta~ned for the duration of .thPhotoP7r~o~ but which increased significantly following :ra~::ireduct~on ~n photoperiod. The peak in testis size found on da 204(early January) occurred two months before maximum testis size yin the wild T t' , OCcurs
'. ~s. ~s s~ze then decreased, as photoperiod became shortto a level s~~n~f~cantly less than the peak though more than at da ~(full , regress~o~). Body weight showed a sharp significant rise withthe ~ncrease ~n testis size but was maintained as the t tregressed. Testosterone concentration increased with t t' . es 7sresponse t d . es ~s S~ze ~n
o ecreas~ng photoperiod and then decreased significantly toa level at day 308 that was not significantly greater than the levelat day 137.
This initial experiment shows that photoperiod can influence~eproduction in male P. poZiocephaZu8 and that this species ma be~nfluenced by both the duration and changes in the h ~Decreasi~g photoperiod prior to the summer solstice appe~r:to:oer~~~~phas7-sh~fted the seasonal reproductive changes, bringing theearl~er than observed in the wild. m on
( 1 ) McGuckin, M.A. & Blackshaw A.W. (1984) P"'o"'. A t sB • "1 ' f"... us. oc. Rap-rod.'LO". ..!2.: 23.
PMSG INDUCED OVULATION IN THE FLYING FOX 'Pte~opu8 scapuZatus
Phil Towers and Len Martin
Department of Physiology and Pharmacology,University of Queensland, St Lucia 4067
In flying foxes endometrial proliferation and progestation isrestricted to the uterine horn ipsilateral to the single preovulatoryfollicle and subsequent co~pus Zuteum (1). From a series ofP. poZiocephaZus shot in the wild, it seemed that this endometrialreaction developed late in the breeding season and in early pregnancy,in the absence of significant increases in peripheral plasmaconcentrations of oestradiol (E2) or progesterone (P4) (2).
To elucidate relationships between follicular growth, ovulation,endometrial development and peripheral E2 and P4 levels, we treated 16mature female P. scapuZatus with 15 iou. 'PMSG, 2 months before theirbreeding season. Animals were sacrificed daily over the following 6days. Blood was collected before sacrifice and peripheral E2 and P4concentrations measured by RIA. Reproductive tracts were examinedhistologically.
Plasma E2 rose from 0.2 ± 0.1* pg/ml (n=4) on days 0-1, to20 ± 7 pg/ml (n=4) on days 2-3, peaking at 254 ± 79 pg/ml (n=3) onday 4, and declining thereafter (33 ± 25 pg/ml, n=4). Plasma P4remained low until day 3 (3 ± 0.3 ng/ml, n=8) then rose over days 4, 5and 6 (respectively 6 ± 2 ng/ml, n=3; 8 ± 5 ng/ml, n=2; 34 ± 11 ng/ml,n=2).
Control ovaries had no antral follicles and adjoining endometriawere atrophic; 2 days after PMSG treatment ovarian tissues showedintense mitotic activity but adjoining endometria remained atrophic.By day 4 all ovaries contained many large (>800).J M) antral folliclesand multiple ovulations had occurred, with meiotic metaphase II ovalocated in bursa and tubes. Adjoining endometria were hypertrophiedwith intense epithelial mitotic actiVity and cytoplasmic basophiliacharacteristic of oestrogenic stimulation. By days 5-6, ovariescontained large amounts of luteal tissue and remaining antralfollicles were atretic. Adjoining endometrial epithelia showedincreasing hypertrophy and cytoplasmic eosinophilia characteristic ofprogestational stimulation; meiotic metaphase II ova were first seenin the uterine lumen on day 6.
These results indicate that in the wild, the final stages offollicular maturation and ovulation could be completed in <48 h. Evenwith the massive ovarian overstimulation achieved experimentally theperipheral E2 peak was transient. From the number of large folliclesinvolved here, we calculate that secretion from the normally singlepreovulatory follicle would produce transient increases in peripheralE2 concentrations of '\,8 pg/ml. SUch -:alues are within the rangeobserved in breeding season P. poZ'LocephaZus females lackingpreovulatory follices (3), and would be of little diagnostic value.
* mean ± SEM (n>2) or range (n=2)
( 1) Martin, L. and 'lbwers, P.A. ( 1984) P~oc. Aust. Soc. Rap~od. BioZ.
~ 8.(2) 'lbwers, P.A. and Martin, L. ( 1984) P-roc. Aust. Soc. Rep-rod. BioZ.
~ 9.Aust. MamrrnZo(3 ) 'lbwers, P.A. and Martin, L. ( 1985) (in press)

AUSTRALIAN SOCIETY FOR REPRODUCTIVE BIOLOGY
16. 2 Minutes of 15th Annual General Meeting: The minutes were receivedas a true and accurate record.
116
ACTIVE ANTI-PATERNAL IMMUNIZATION DOES NOT AFFECT THE SUCCESS OFMARSUPIAL PREGNANCY
J.C. Rodger, T.P. Fletcher & C.H. Tyndale-Biscoe
Dept. Immunology, ANU & CSIRO Div. wildlife & Rangelands Res.Canberra.
Active anti-paternal immunization does not compromise pregnancyin eutherians. However, in a preliminary study in tammar wallabies,grafting with paternal skin resulted in transient infertility (1). Inthe present study, critical monitoring of the breeding efficiency oftammar wallabies sensitized against their mate's transplantationantigens aimed to resolve the question of immunologically mediatedinfertility in marsupials. By relating the timing of mating, birthand pouch young removal to the plasma progesterone profile, it is nowpossible to determine whether an apparently infertile reproductivecycle reflects neonatal loss due to disturbance at the time ofparturition or true compromise of pregnancy.
In mid November, 16 females were randomly divided into pairs andeach pair allocated to a specific male. Within each group one femalereceived a full thickness skin graft from her prospective mate(experimental) and the other a graft of her own skin (control).Experimental females received a second graft of male skin after onemonth. The status of the grafts was assessed histologically inbiopsies taken 7 & 10 days after grafting.
Beginning in late January, the experimental group were monitoredfor 30 reproductive cycles and produced 24 pouch young whereas thecontrol animals p=oduced 28 pouch young from 33 cycles. Five of the11 apparently non-fertile cycles were judged to be normal pregnancieswhe:e the young had failed to reach the pouch (cycle length <28 days;rapld plasma progesterone decline coincident with oestrus).Therefore, true infertility was limited and although, occurringmainly in the male-skin 2 grafted group (5 cycles) this was notsignificantly different (X , p>0.5) from the controls (1 cycle) andrepresented the effect of one very poor breeder.
We conclude that allogeneic pregnancy in marsupials is notcompromised by active anti-paternal immunization. Infertilityobserved, here and in the earlier study, reflected disturbance tobreeding due to handling of the animals or the poor reproductiveefficiency of individual animals in small experimental groups.
(1) Walker, K.Z. & Tyndale-Biscoe, C.H. Aust. J. BioI. Sci.31: 173-182 (1978)
I
16. 1
16. 3
16. 4
16. 5
16. 6
16. 7
16. 8
16. 9
Minutes of 16th Annual General Meeting held at 1730
Tuesday, 28th August, 1984, Melbourne
Attendance: 80 members; an apology was received from J. Cummins
Business Arising from 16.2: No business was raised.
Treasurer's Report: A report was presented (attached) and receivedby the meeting. The Treasurer indicated that there was financiallylittle change from the position reported one year earlier.
Matters Arising from 16.4: In answer to questions the Treasurerstated that 11 students had applied for travel grants. The Chairmanproposed a vote of thanks to Bevan Miller for the excellent servicehe has rendered the Society as Treasurer.
Programme Organising Committee Report: (attached)In response to a request by the Committee concerning the success ofthe method of poster presentation a vigorous discussion ensued. Itwas generally agreed that the ~oster format was highly successful.The Chairman proposed a vote of thanks to the POC as the presentCommittee members were retiring.
Chairman's Report:The Chairman reported that the Executive Committee had met twiceduring the year (Sydney in March, Melbourne in August). As convenorof the ASRB Hormone Distribution Committee, the Chairman reportedthat materials would be available in December 1984; that theCommittee would not consider requests from commercial subscribers;that LH was in the preparative stage, and ovine material was beingprepared (report attached). He thanked the members of th: SteeringCommittee (R. Seamark, B. Bindon, L. Cahill, H. Tyndale-Blscoe)for their continued help. In response to commercial aspects of thisproject, he reported that an investigation was underway into.thepossibility of forming ASRB into a Company. Finally the Chalrmanreminded members of the changes to occur on the Executive Committee.
New Members: Eighty-one persons were admitted to membership of ASRB.
Elections: J. Cummins was elected Secretary, and G. Evans, B. Millerand C. Nancarrow were elected as Committee persons.
16.11
16.10 Constitution: The Chairman withdrew foreshadowed changes with a viewto the rewording of the changes.
General Business: T. Robinson reported that the bid to stage theInternational Symoisuum on Animal Production and ArtificialInsemination in Australia in 1988 had been unsuccessful. TheChairman thanked T. Robinson and P. Mattner for their efforts.
The Chairman commented that the Executive Committee had resolvedto follow up a request from the Secretary of the PhysiologicalSociety of Thailand for closer cooperation.
1

3
The Committee took the decision to request the Local OrganizingCommittee for space to enable posters to remain up for the entireProgramme and organized the posters into groups naming a Chairman tocarryon a poster discussion which was to take place around the poster.
REPORT OF THE A.S.R.B. PROGRAMME ORGANIZING COMMITTEE 1984
As Chairman of the Committee I would like to thank all Committeemembers (Drs Risbridger. Chamley and Staples) for their great help inthis year's activities.
The Committee wishes to emphasise the importance for members of theSociety to enclose adequate copies of the abstracts and the selfaddressed cards, as indicated in the Call for Abstracts, as thisgreatly facilitates the procedures for review and notification ofdecision regarding the abstract.
One hundred' and eight abstracts were submitted for this year'sMeeting and all papers were examined by at least two reviewers.Papers of doubtful scientific merit were then reviewed by the entireProgramme Organizing Committee and it was decided to notify theauthors that their abstracts were not acceptable in their presentform as they did not contain adequ~te scientific data to back up theconclusions reached by the investigators. Several other papers alsocontained only limited data to back up the conclusions reache4 andthe Committee would urge all authors to include sufficient data toback up their conclusions. The authors were given the opportunityto resubmit the abstracts after appropriate amendments were made asdiscussed with the Committee. Of the 108 papers one paper was withdrawn. Thirty-six papers were selected for oral presentation and theremaining seventy-two were allocated to poster presentations. Thedecision on whether a paper was included as a poster or an oralpresentation was based purely on the consideration of grouping presentations adequately to create a cohesive scientific programme. Thepapers which were viewed as good papers and bad papers by the Committeewere districuted in the oral and poster sessions.
2
Signed as a true record
Date
Page 2
A request was made for suggestions for an ASRB satellitesymposium to be held at the ESA meeting in Lorne in March,1985. A. Blackshaw requested that a set of "instructions tospeakers" be prepared.
L. Bennett spoke to the meeting about-the need for continuedsupport by ASRB for the Australian Journal of BiologicalScience.

Chairmenf
-- 1969-19731973-19771977-19811981-19831983-1985
T.J. RobinsonC.W. En1mensD.M. de KretserN.W. MooreJ.K. Findlay
ILife Members
C.W. EmmensT.J. Robinson
Junior Scientist Award
1981 P.J. Lutjen1982 R.J. Rodger & C.B. Gow1983 S.P. Flaherty1984 C. O'Neill