attempts to restore abduction of the paralyzed equine arytenoid
TRANSCRIPT

Attempts to Restore Abduction of the Paralyzed EquineArytenoid Cartilage I. Nerve-Muscle Pedicle Transplants
N.G. Ducharme, F.D. Horney, G.D. Partlow and T.J. Hulland
ABSTRACT
The purpose of this project was toadapt a surgical technique fromhumans and dogs to horses in which aportion of an accessory muscle ofrespiration and its nerve supply istransplanted to a denervated dorsalcricoarytenoid muscle. Anatomicaldissections in seven horses revealedtwo possible donor nerve-pediclegrafts: the omohyoid and the sterno-thyrohyoid, both innervated by abranch of the first and second cervicalnerves. Histochemical evaluations intwo ponies of the dorsal cricoaryte-noid, omohyoid and sternothyrohyoidmuscles revealed similar proportionsof fiber types 1 and 2 in all threemuscles. Electromyographic studies inthese two ponies revealed that theomohyoid and sternothyrohyoid mus-cles contract synchronously withrespirations during forced inspirationunder general anesthesia. Based onsurgical ease of access, a 1 cm2 portionof the omohyoid muscle at the point ofpenetration of the second cervicalnerve was used as a nerve-musclepedicle graft in an attempt to reinner-vate the left dorsal cricoarytenoidmuscle in four ponies. These fourponies (as well as three others whichserved as controls) had previouslyundergone left recurrent laryngealnerve transection. All seven poniesendoscopically showed signs of com-plete left laryngeal hemiplegia imme-diately postoperatively. Animals weremonitored endoscopically for 30 weeksafter surgery. The three control poniesshowed no abduction of the arytenoid
cartilage. In addition, in these threeponies, histological and histochemicalevidence of denervation with theexpected changes of muscle fiberatrophy and fibrosis were present inthe dorsal cricoarytenoid muscle.Three of the four experimental ponieshad histochemical evidence of acomplex muscle fiber hypertrophy andabnormal fiber pattern in the dorsalcricoarytenoid muscle as well as type 1and 2 muscle fiber groupings whichseemed to be an indication ofsubtermi-nal axonal sprouting in reinnervation.Endoscopic examination, however,revealed partial restoration of aryte-noid abduction in only one of the fourponies where nerve-pedicle transplan-tation was performed.
RESUME
Cette experience visait a adapter unetechnique chirurgicale, utilisee chez leshumains et les chiens, a des chevauxauxquels on transplante une partied'un muscle accessoire de la respirationet son apport nerveux, dans un musclecrico-arytenoidien dorsal denerve'. Desdissections anatomiques, chez septchevaux, revelerent la possibilited'obtenir deux muscles donneurs degreffes innervees, a savoir: l'omo-hyoldien et le sterno-thyro-hyoidien,tous deux innerves par une branche dupremier et du second nerfs cervicaux.L'evaluation histochimique des mus-cles crico-arytenoidien, omo-hyoidienet sterno-thyro-hyoidien, chez deuxponeys, y revela une proportionsemblable de fibres des types #1 et #2.
L'electromyographie effectuee chez cesdeux poneys revela que les musclesomo-hyoidien et sterno-thyro-hyoldien se contractent de faconsynchrone avec les respirations, durantl'inspiration forcee, lors d'anesthesie.En se basant sur l'acces chirurgicalfacile du muscle omo-hyoidien, lesauteurs en preleverent un morceau de 1cm2, au point de penetration dudeuxieme nerf cervical, et l'utiliserentcomme greffe musculaire innervee,dans une tentative de restaurerl'innervation du muscle crico-arytenoidien dorsal gauche, chezquatre poneys. Ceux-ci, tout commeles trois temoins, avaient precedem-ment subi une section du nerf laryngerecurrent gauche. Lendoscopie permitde constater que les sept poneysaffichaient une hemiplegie laryngeegauche complete, immediatementapres l'intervention precitee. On lessoumit ensuite au monitorage endo-scopique, pendant 30 jours. Les troistemoins ne presentierent aucune abduc-tion du cartilage arytenoide; de plus,leur muscle crico-arytenoidien dorsalafficha l'evidence histologique ethistochimique dune denervation quis'accompagnait d'une atrophie et d'unefibrose prevues de ses fibres muscu-laires. Trois des quatre poneys experi-mentaux presentaient l'evidence histo-chimique dun complexe: hyper-trophie-arrangement anormal desfibres du muscle crico-arytenoidiendorsal, ainsi que des regroupements defibres des types #1 et #2, qui semblerentcorrespondre a un bourgeonnementaxonal subterminal de reinnervation.L'endoscopie revela toutefois une
Department of Clinical Studies (Ducharme, Horney), Department of Biomedical Sciences (Partlow) and Department of Pathology (Hulland), OntarioVeterinary College, University of Guelph, Guelph, Ontario N IG 2W 1. Present address of Dr. Ducharme: Large Animal Clinic, New York State College ofVeterinary Medicine, Cornell University, Ithaca, New York 14853.
This project was supported by the Guelph Centre for Equine Research.
Submitted March 18, 1988.
Can J Vet Res 1989; 53: 202-209202

restauration partielle de l'abduction del'arytenoide chez un seul des quatreponeys qui avaient subi une transplan-tation de tissu musculaire innerve.
INTRODUCTION
Laryngeal hemiplegia is a commoncause of respiratory disorder in theperformance horse (1). The pathophy-siological basis of this disease in horsesis a peripheral neuropathy resulting ina decrease or absence of motor activityof the dorsal cricoarytenoid (DCA)muscle, the primary arytenoid cartil-age abductor (2,3). Different causes ofrecurrent laryngeal nerve damage havebeen suggested: periphlebitis, traumaand congenital neurogenic defects (3-5). The dorsal cricoarytenoid muscle aswell as most other intrinsic muscles ofthe larynx are innervated by therecurrent laryngeal nerve (3-6). Basedon experimentally created left laryn-geal hemiplegia, the loss of abductorfunction results in obstruction toinspiratory flow (1). This explains thenoisy respiration and the decrease inperformance level (4).
In horses, to restore sufficientfunction for racing or running, theinsertion of a laryngeal prosthesis(laryngoplasty) is presently the surgicaltechnique of choice (4,7,8). The aim ofthis technique is to fix the affectedarytenoid cartilage into an abductedposition (4). The success rate for racingsoundness ranged from 38-90% (7-1 1).Complications associated with thesurgery include chronic infection of theprosthesis, formation of suture sinus,ossification of cartilage, dysphagia,esophageal obstruction, pneumonia,intralaryngeal granulomatous polyps,right sided laryngospasm duringexercise, laryngeal edema, chondritisand coughing (7,8, 10,11). In somehorses, the arytenoid cartilage seemedto return to an abducted position aftervarious periods of time and, therefore,the animal's performance level againdecreased. For these reasons, someinvestigators do not recognize thistechnique of fixation as satisfactory forracing soundness (7,13), or indicatethat "it is time for substantial progressto be made" (14).
Laryngeal reinnervation was firstreported in humans by Horsley in 1909(14). Since then, many surgical
techniques have been described, mostof which resulted in limited success inachieving reinnervation (16-19). Atechnique of nerve-muscle pedicletransplantation described by Tucker in1970 offered promising results (20-24).With this technique a nerve and aportion of its attached muscle, whichcontracts normally during inspiration,is implanted into the posterior cricoa-rytenoid muscle (22,23). In humansand dogs, function is restored in two tosix weeks (20,23). Proponents of thissurgical technique have suggested thatthe rapid restoration of functionoccurs only because of the requirementfor sufficient healing to propagateimpulses from the transplanted muscleto the recipient muscle (24). Histologi-cal studies have demonstrated that themajority (80%) of the nerve fibers inthe pedicle do not degenerate (21).
It has been shown that reinnervationas late as one to three years followingdenervation is possible (25,26). Thedorsal cricoarytenoid muscle has beensuccessfully reinnervated in dogs aftera period of denervation of six months(27). In humans, successful reinnerva-tion has been reported in cases rangingfrom 7 months to 22 years of denerva-tion (23). Therefore, it seems reasona-ble to assume that in performanceanimals in which endoscopic examina-tion is done frequently, laryngealhemiplegia would be discovered earlyenough to allow reinnervation beforeirreversible changes of muscle fibrosisand atrophy intervened.The objective of this study was to
determine if an accessory muscle ofrespiration can serve as a nerve-musclepedicle donor in the equine patient andreinnervate a recently paralyzed leftdorsal cricoarytenoid muscle.
MATERIALS AND METHODS
All procedures complied with theGuide to the Care and Use ofExperimental Animals (CanadianCouncil on Animal Care).
DETERMINATION OF DONOR MUSCLE
Two mature ponies with unknownhistories were obtained from a nearbystockyard and determined, after acomplete physical examination, to bein good condition. Endoscopic exami-nation performed using a flexible fiber
optic endoscope (Model C 100,Olympus Corporation, Lake Success,New York) and without sedationrevealed symmetrical arytenoid cartil-ages motion.The ponies were premedicated
intravenously with chloral hydrate andanesthesia was induced with anintravenous bolus of thiamylal sodium(5 mg/kg body weight). Immediatelyfollowing induction, tracheal intuba-tion was performed using a cuffedendotracheal tube and anesthesia wasmaintained with halothane and oxygendelivered through a semiclosed system.With the animal in right lateralrecumbency with the head extendedand following aseptic preparation ofthe left proximal cervical area, a 15 cmcurvilinear incision was made ventralto the linguofacial vein. Using delicateblunt dissection, the omohyoid musclewas separated from the jugular andlinguofacial veins. Further dissectionallowed exposure of the left dorsalcricoarytenoid (DCA) muscle.
Simultaneous electromyographicrecordings were obtained using twobipolar 30 micron electrodesimplanted directly in the left dorsalcricoarytenoid muscle which served ascontrol. The other bipolar electrodeswere implanted in the omohyoid andsternothyrohyoid muscles. All record-ings were obtained at a sensitivity of10-20 millivolts/division using anelectromyograph (Teca Electromyo-graph Model M, Pleasantville, NewYork). The electromyographic record-ings indicated that the omohyoid andsternothyrohyoid muscles contractsynchronously with the dorsal cricoa-rytenoid muscle during forcedinspiration.
Following the recording, throughthe same surgical approach, a cube ofmuscle about 0.5 cm in each dimensionwas removed from the dorsal cricoa-rytenoid, omohyoid and sternothyro-hyoid muscles. The samples werefrozen immediately in solid CO2 cooledisopentane for adenosine triphospha-tase (ATPase) histochemical stainingto identify fiber types. All threemuscles had a similar proportion offiber type with both types making up atleast 40% of the total fiber population(Fig. 1). Ease of surgical access led toselection of a branch of the secondcervical nerve entering the omohyoidmuscle as the origin of a nerve-musclepedicle.
203

Fig. 1. Normal dorsal cricoarytenoid muscle. Note the checkerboard pattern due to the arrangementof the different type of muscle fiber. ATPase preparation, alkaline preincubation, X65.
NERVE-MUSCLE PEDICLETRANSPLANTATION
A. Animals Seven ponies wereobtained at a nearby animal stockyardand determined to be in good condi-tion by physical examination anddetermination of hematocrit, totalplasma protein levels and the acid/base and electrolyte status. Endos-copic examination without sedationrevealed symmetrical movement of the
arytenoid cartilages. Of the sevenponies, four were randomly allocatedto the experimental group and three tothe control group. Disproportionateallocation was chosen because ofexpected consistency in the controlgroup where any spontaneous reinner-vation was felt to be unlikely.
B. Surgical Procedures - Undergeneral anesthesia (previously des-
Fig. 2. Schematic representation of left cranial cervical area and the relation of the surgicalincision. Wing of atlas - A, linguofacial vein - Lf, maxillary vein - Mv, jugular vein - Jv,jugular groove - Jg.
cribed), the four experimental ponieswere subjected to the followingprocedures. A 20 cm linear skinincision was made adjacent to theventral border of the left brachioce-phalic muscle (Fig. 2). The incisionwas extended between the maxillaryvein ventrally and the ventral borderof the brachiocephalic muscle dor-sally. A ventral branch of the secondcervical nerve was identified andfollowed to its entry on the dorsome-dial border of the omohyoid muscle(Fig. 3). A 1 cm2 section of muscleincluding the site of entry of the nervewas resected, creating a nerve-musclepedicle. The DCA muscle wasapproached by blunt dissectionventral to the common carotid artery.The nerve-muscle pedicle was thensutured into a slit made perpendicularto the long axis of the fibers in theDCA muscle with two to fournonabsorbable 5-0 polypropylenesutures (simple interrupted pattern)(Fig. 3). The maxillary vein and thebrachiocephalic muscle were reap-posed with 2-0 polyglactin 910 in asimple continuous pattern. At thecaudal aspect of the incision the leftrecurrent laryngeal nerve was thentransected and a 2 cm sectionremoved. The skin and the subcutae-nous tissues were reapposed similarly.The ponies in the control group weresubjected to an identical surgicalprocedure except that the nervemuscle-pedicle was placed back in theomohyoid muscle.
Endoscopic evaluation was per-formed without sedation through theventral meatus of the right nostrilusing a flexible fiber optic endoscope.This was done preoperatively, the dayafter surgery and every two weeksuntil the 12th postoperative week.At that time, under general anesthe-
sia and the same surgical approach, abiopsy of the left dorsal cricoarytenoidmuscle of an experimental pony wasobtained. This muscle biopsy wasfrozen immediately and ATPasestaining techniques (alkaline preincu-bation), hemotoxylin and eosin andsilver stains were used to evaluate thestate of the muscle. At that timehistopathological evidence of reinner-vation was observed and it wasdecided to prolong the observationperiod for another 18 weeks for the sixremaining animals (three in the
204
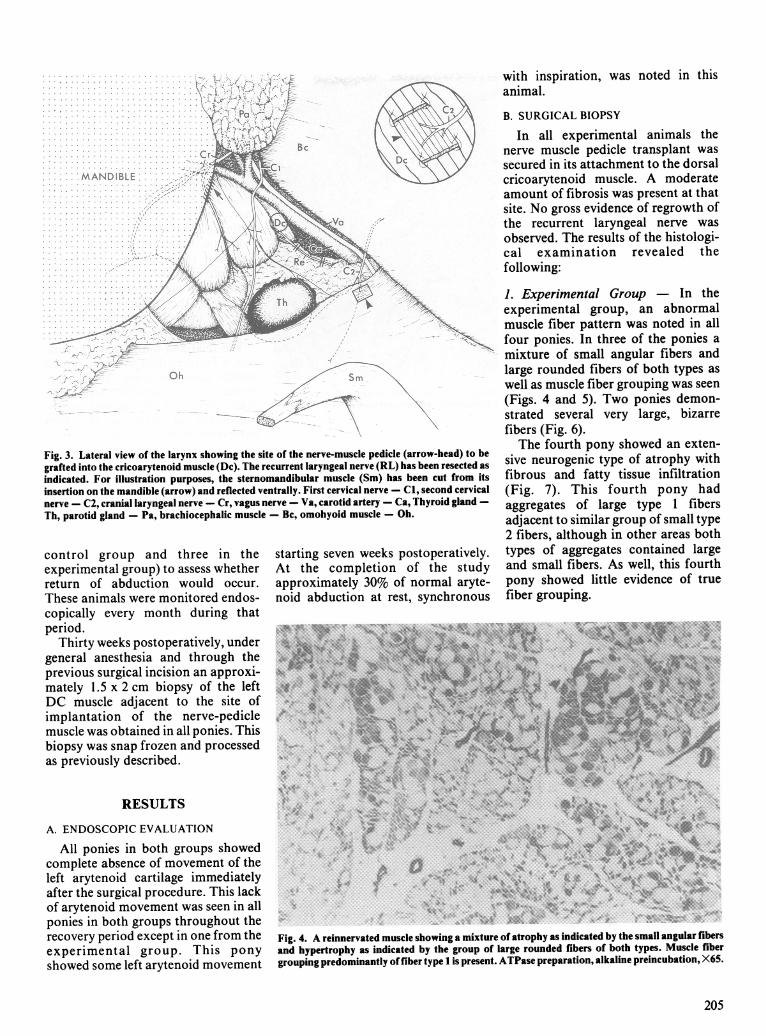
Fig. 3. Lateral view of the larynx showing the site of the nerve-muscle pedicle (arrow-head) to begrafted into the cricoarytenoid muscle (Dc). The recurrent laryngeal nerve (RL) has been resected asindicated. For illustration purposes, the sternomandibular muscle (Sm) has been cut from itsinsertion on the mandible (arrow) and reflected ventrally. First cervical nerve - Cl, second cervicalnerve - C2, cranial laryngeal nerve - Cr, vagus nerve - Va, carotid artery - Ca, Thyroid gland -Th, parotid gland - Pa, brachiocephalic muscle - Bc, omohyoid muscle - Oh.
control group and three in theexperimental group) to assess whetherreturn of abduction would occur.These animals were monitored endos-copically every month during thatperiod.
Thirty weeks postoperatively, undergeneral anesthesia and through theprevious surgical incision an approxi-mately 1.5 x 2 cm biopsy of the leftDC muscle adjacent to the site ofimplantation of the nerve-pediclemuscle was obtained in all ponies. Thisbiopsy was snap frozen and processedas previously described.
RESULTS
A. ENDOSCOPIC EVALUATION
All ponies in both groups showedcomplete absence of movement of theleft arytenoid cartilage immediatelyafter the surgical procedure. This lackof arytenoid movement was seen in allponies in both groups throughout therecovery period except in one from theexperimental group. This ponyshowed some left arytenoid movement
starting seven weeks postoperatively.At the completion of the studyapproximately 30% of normal aryte-noid abduction at rest, synchronous
with inspiration, was noted in thisanimal.
B. SURGICAL BIOPSY
In all experimental animals thenerve muscle pedicle transplant wassecured in its attachment to the dorsalcricoarytenoid muscle. A moderateamount of fibrosis was present at thatsite. No gross evidence of regrowth ofthe recurrent laryngeal nerve wasobserved. The results of the histologi-cal examination revealed thefollowing:
1. Experimental Group - In theexperimental group, an abnormalmuscle fiber pattern was noted in allfour ponies. In three of the ponies amixture of small angular fibers andlarge rounded fibers of both types aswell as muscle fiber grouping was seen(Figs. 4 and 5). Two ponies demon-strated several very large, bizarrefibers (Fig. 6).The fourth pony showed an exten-
sive neurogenic type of atrophy withfibrous and fatty tissue infiltration(Fig. 7). This fourth pony hadaggregates of large type 1 fibersadjacent to similar group of small type2 fibers, although in other areas bothtypes of aggregates contained largeand small fibers. As well, this fourthpony showed little evidence of truefiber grouping.
-:
Fig. 4. A reinnervated muscle showing a mixture of atrophy as indicated by the small angular fibersand hypertrophy as indicated by the group of large rounded fibers of both types. Muscle fibergrouping predominantly of fiber type 1 is present. ATPase preparation, alkaline preincubation, X65.
205
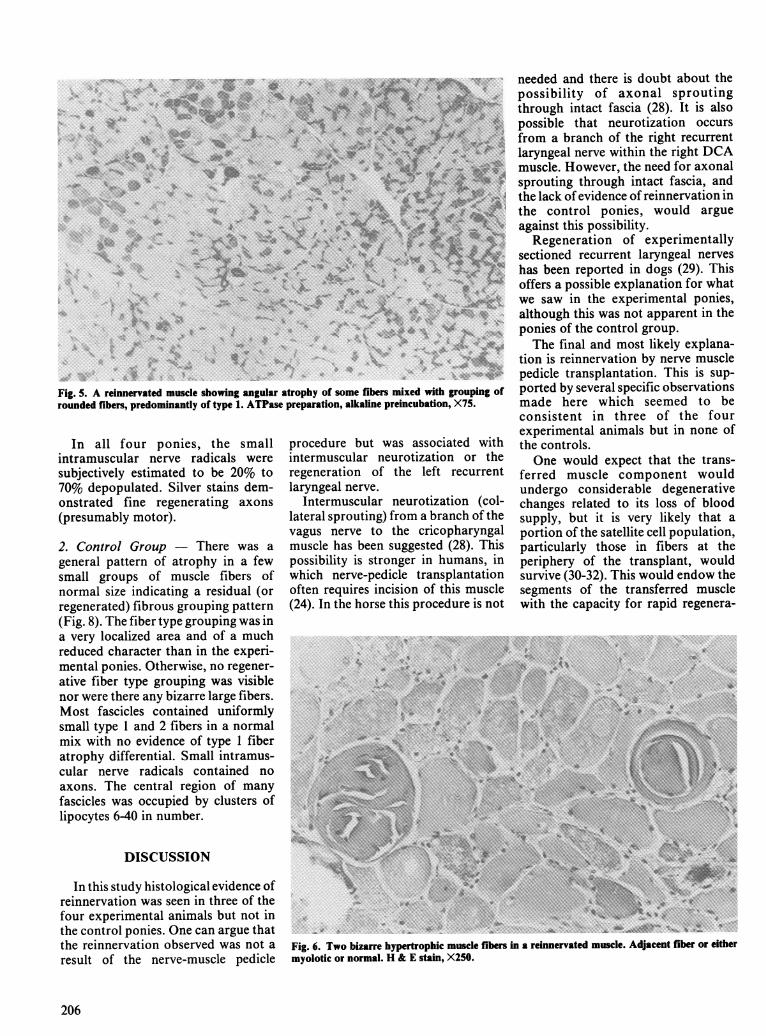
Fig. 5. A reinnervated muscle showing angular atrophy of some fibers mixed with grouping ofrounded fibers, predominantly of type 1. ATPase preparation, alkaline preincubation, X75.
In all four ponies, the smallintramuscular nerve radicals weresubjectively estimated to be 20% to70% depopulated. Silver stains dem-onstrated fine regenerating axons(presumably motor).
2. Control Group - There was ageneral pattern of atrophy in a fewsmall groups of muscle fibers ofnormal size indicating a residual (orregenerated) fibrous grouping pattern(Fig. 8). The fiber type grouping was ina very localized area and of a muchreduced character than in the experi-mental ponies. Otherwise, no regener-ative fiber type grouping was visiblenor were there any bizarre large fibers.Most fascicles contained uniformlysmall type 1 and 2 fibers in a normalmix with no evidence of type 1 fiberatrophy differential. Small intramus-cular nerve radicals contained noaxons. The central region of manyfascicles was occupied by clusters oflipocytes 6-40 in number.
DISCUSSION
In this study histological evidence ofreinnervation was seen in three of thefour experimental animals but not inthe control ponies. One can argue thatthe reinnervation observed was not aresult of the nerve-muscle pedicle
procedure but was associated withintermuscular neurotization or theregeneration of the left recurrentlaryngeal nerve.
Intermuscular neurotization (col-lateral sprouting) from a branch of thevagus nerve to the cricopharyngalmuscle has been suggested (28). Thispossibility is stronger in humans, inwhich nerve-pedicle transplantationoften requires incision of this muscle(24). In the horse this procedure is not
needed and there is doubt about thepossibility of axonal sproutingthrough intact fascia (28). It is alsopossible that neurotization occursfrom a branch of the right recurrentlaryngeal nerve within the right DCAmuscle. However, the need for axonalsprouting through intact fascia, andthe lack of evidence of reinnervation inthe control ponies, would argueagainst this possibility.
Regeneration of experimentallysectioned recurrent laryngeal nerveshas been reported in dogs (29). Thisoffers a possible explanation for whatwe saw in the experimental ponies,although this was not apparent in theponies of the control group.The final and most likely explana-
tion is reinnervation by nerve musclepedicle transplantation. This is sup-ported by several specific observationsmade here which seemed to beconsistent in three of the fourexperimental animals but in none ofthe controls.One would expect that the trans-
ferred muscle component wouldundergo considerable degenerativechanges related to its loss of bloodsupply, but it is very likely that aportion of the satellite cell population,particularly those in fibers at theperiphery of the transplant, wouldsurvive (30-32). This would endow thesegments of the transferred musclewith the capacity for rapid regenera-
t~~~~~~~~~~~~~~~~~X.tFig. 6. Two bizarre hypertrophic muscle fibers in a reinnervated muscle. Adjacent fiber or eithermyolotic or normal. H & E stain, X250.
206
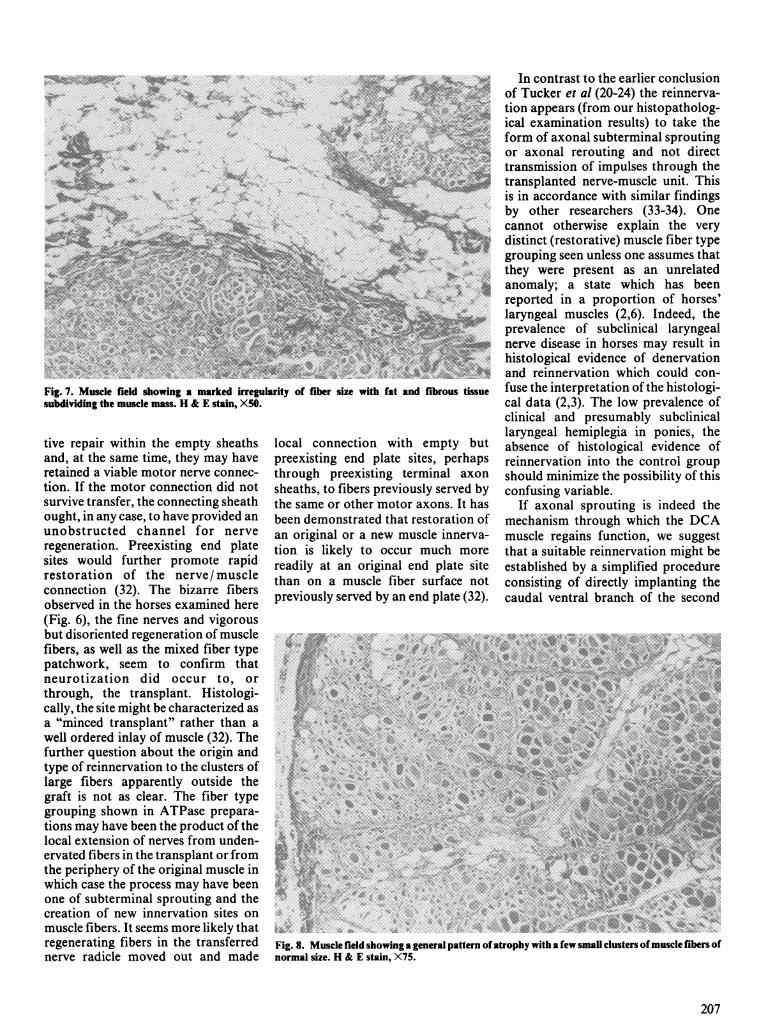
Fig. 7. Muscle field showing a marked irregularity of fiber size with fat and fibrous tissuesubdividing the muscle mass. H & E stain, X50.
tive repair within the empty sheathsand, at the same time, they may haveretained a viable motor nerve connec-tion. If the motor connection did notsurvive transfer, the connecting sheathought, in any case, to have provided anunobstructed channel for nerveregeneration. Preexisting end platesites would further promote rapidrestoration of the nerve/muscleconnection (32). The bizarre fibersobserved in the horses examined here(Fig. 6), the fine nerves and vigorousbut disoriented regeneration of musclefibers, as well as the mixed fiber typepatchwork, seem to confirm thatneurotization did occur to, orthrough, the transplant. Histologi-cally, the site might be characterized asa "minced transplant" rather than awell ordered inlay of muscle (32). Thefurther question about the origin andtype of reinnervation to the clusters oflarge fibers apparently outside thegraft is not as clear. The fiber typegrouping shown in ATPase prepara-tions may have been the product of thelocal extension of nerves from unden-ervated fibers in the transplant or fromthe periphery of the original muscle inwhich case the process may have beenone of subterminal sprouting and thecreation of new innervation sites onmuscle fibers. It seems more likely thatregenerating fibers in the transferrednerve radicle moved out and made
local connection with empty butpreexisting end plate sites, perhapsthrough preexisting terminal axonsheaths, to fibers previously served bythe same or other motor axons. It hasbeen demonstrated that restoration ofan original or a new muscle innerva-tion is likely to occur much morereadily at an original end plate sitethan on a muscle fiber surface notpreviously served by an end plate (32).
In contrast to the earlier conclusionof Tucker et al (20-24) the reinnerva-tion appears (from our histopatholog-ical examination results) to take theform of axonal subterminal sproutingor axonal rerouting and not directtransmission of impulses through thetransplanted nerve-muscle unit. Thisis in accordance with similar findingsby other researchers (33-34). Onecannot otherwise explain the verydistinct (restorative) muscle fiber typegrouping seen unless one assumes thatthey were present as an unrelatedanomaly; a state which has beenreported in a proportion of horses'laryngeal muscles (2,6). Indeed, theprevalence of subclinical laryngealnerve disease in horses may result inhistological evidence of denervationand reinnervation which could con-fuse the interpretation of the histologi-cal data (2,3). The low prevalence ofclinical and presumably subclinicallaryngeal hemiplegia in ponies, theabsence of histological evidence ofreinnervation into the control groupshould minimize the possibility of thisconfusing variable.
If axonal sprouting is indeed themechanism through which the DCAmuscle regains function, we suggestthat a suitable reinnervation might beestablished by a simplified procedureconsisting of directly implanting thecaudal ventral branch of the second
Fig. 8. Muscle field showing a general pattern ofatrophy with a few small clusters ofmuscle fibers ofnormal size. H & E stain, X75.
207

cervical nerve only into the dorsalcricoarytenoid muscle. This mightminimize fibrous tissue reactionassociated with the transplant of avascular muscle unit and this, in turn,may enhance axonal sprouting.
Selective type 1 muscle fiberatrophy was not observed to occurwith regularity, unlike the previouslyreported situation in cats (35). We donot have an explanation for thisdifference at this time.The positive endoscopic evidence of
mild abduction in the fourth experi-mental pony was not supported by thehistopathological findings. It ispossible that the biopsy was notrepresentative of the entire muscle.One should also consider whether theendoscopic evidence of abduction isdue to a passive abduction associatedwith descent of the larynx or to activeabduction due to reinnervation of theDCA muscle by the nerve-pedicletransplant.Although not documented in
horses, in humans and dogs inspira-tory descent of the larynx produceslateral movement of the arytenoidcartilage partially through the tensionof contraction of the cricothyroid,omohyoid, and sternothyroid muscles(36-39). These muscles lead to simul-taneous descent and distraction of thelarynx from the hyoid bone stretchingthe supporting ligaments of thelarynx. The longitudinal stress is thenreflected to the quadrangular mem-brane which attaches on the arytenoidcartilages. The tension on the aryte-noid cartilages causes them to slidelaterally and then rotate dorsally onthe oblique cricoarytenoid articula-tion leading to arytenoid abduction.Enhancement of this mechanismduring recurrent laryngeal paralysishas been suggested by Crumley (28).He suggested that postoperativemuscular scarring of the DCA resultsin caudolateral displacement of thearytenoid cartilage resulting in itsabduction by increasing longitudinaltension on the supporting ligaments ofthe larynx. This would also decreasethe craniomedial gliding movementsof the arytenoid cartilage duringcricothyroid inspiratory contractionsand give the cricothyroid muscle andthe extrinsic laryngeal dilators (omo-hyoid, sternothyroid muscles) a firmerand more caudal fulcrum enhancing
208
this usually subtle abduction. Wecannot rule out this mechanism;moderate muscular fibrosis wasobserved during the biopsy procedure.Left arytenoid cartilage movement,however, was not observed in thecontrol ponies postoperatively.
Histopathological evidence of rein-nervation was not associated withabduction in the other three experi-mental ponies. Possibly the reinnerva-tion obtained was too focal to allowmeaningful contraction of the dorsalcricoarytenoid muscle. It is possiblethat the nerve/ muscle resection usedwas unacceptable (i.e. does not fireconsistently during inspiration). Theelectromyogram (EMG) analysisindicated contraction of the omohyoidmuscle during inspiration. This wasobtained with the animal anesthetizedand with increased dead space rebrea-thing, presumably leading to hyper-capnia. Therefore, it is conceivablethat the omohyoid muscle does notcontract during inspiration at rest, butdoes contract during exercise. Perhapsendoscopic evaluation should beperformed during or after strenuousexercise.
In conclusion, from this pilot studyit appears that reinnervation of thedorsal cricoarytenoid muscle is possi-ble. However, reinnervation of theDCA with a nerve-muscle pedicleusing a branch of the second cervicalnerve and a section of the omohyoidmuscle did not result in normalabduction of the left arytenoidcartilage. In addition, further work isnecessary to verify the mechanismsand structures involved in abductionof the equine arytenoid cartilages.
ACKNOWLEDGMENTS
The authors would like to thankMs. Wynn Halina for the schematicillustrations.
REFERENCES
1. DERSKEN JF, STICK JA, SCOTT EA.Effects of laryngeal hemiplegia andlaryngoplasty on airway flow mechanics inexercising horses. Am J Vet Res 1986; 47:16-26.
2. DUNCAN ID, GRIFFITHS IR,McQUEEN A. The pathology of equinelaryngeal hemiplegia. Acta Neuropathol(Berl) 1974; 27: 337-348.
3. COLE CR. Changes in the equine larynxassociated with laryngeal hemiplegia. Am JVet Res 1946; 7: 69-77.
4. MARKS D, MacKAY-SMITH MP,CUSHING LS, LESLIE JA. Observationson laryngeal hemiplegia in the horse andtreatment by abductor muscle prosthesis.Equine Vet J 1969; 2: 159-167.
5. GUNN HM. Further observations onlaryngeal skeletal muscle fibers in "normal"horses. Equine Vet J 1973; 5: 77-80.
6. GUNN HM. Histochemical observationson laryngeal skeletal muscle fibers in"normal" horses. Equine Vet J 1972; 4: 144-148.
7. GOULDEN BE, ANDERSON LJ. Equinelaryngeal hemiplegia: Part III. Treatmentby laryngoplasty. NZ Vet J 1982; 30: 1-5.
8. BAKER GJ. Laryngeal hemiplegia in thehorse. Compend Contin Educ 1983; 5: 561-568.
9. HUSKAMP B, BOCKEMHOFF G.Ergebnise der Kehikopfpfeif-eroperationnach Marks U.A. Der Pratische Tieraerztl1978; 4: 302-306.
10. GREET TRC, BAKER GJ, LEE R. Theeffect of laryngoplasty on pharyngealfunction in the horse. Equine Vet J 1979; 11:153-158.
11. SPEIR VC. A retrospective surgery of theracing performance of Australian horsesafter surgery for carpal injuries andlaryngeal hemiplegia. Proc Am AssocEquine Pract 1976: 335-345.
12. HUSKAMP B. Komplikationen NachKehlkopf Operationem. Der PratischeTieraertztl 1980; 61: 848-858.
13. COOKS WE. Biomechanics of the equinelarynx in health and disease. Proc 8th AnnuVet Surg Forum, Chicago, 1980.
14. SPEIRS VC. Laryngeal surgery - 150years on. Equine Vet J 1987; 19: 377-383.
15. HORSLEY JA. Suture of the recurrentlaryngeal nerve with report of a case. TransSouth Surg Gynecol Assoc 1909; 22: 161-170.
16. FRAZIER CH, MOSSER WB. Treatmentof recurrent laryngeal nerve paralysis bynerve anastomosis. Surg Gynecol Obstet1926; 43: 134-199.
17. COLLEDGE L. On the possibility ofrestoring movements in a paralyzed vocalcord by nerve anastomosis. Br Med J 1925;2: 547-548.
18. BALLANCE C. Some experiments onnerve anastomosis. Proc Mayo Clin 1928; 3:317-318.
19. GORDON JH, McCABE BF. The effect ofaccurate neurorrhaphy on reinnervationand return of laryngeal function. Laryngo-scope 1968; 78: 236-250.
20. TUCKER HM, HARVEY JE, OGURAJH. Vocal core remobilization in the caninelarynx. Arch Otolaryngol 1970; 92: 530-533.
21. TUCKER HM, OGURA JH. Vocal cordremobilization in the canine larynx: anhistological evaluation. Laryngoscope1971; 81: 1602-1606.
22. HENGERER AS, TUCKER HM. Resto-ration of abduction in the paralyzed vocalcord. Arch Otolaryngol 1973; 97: 247-250.
23. TUCKER HM. Human laryngeal reinner-vation. Laryngoscope 1976; 86: 769-779.
24. TUCKER HM. Nerve-muscle pediclereinnervation of the larynx: avoidingpitfalls and complications. Ann OtolRhinol Laryngol 1982; 91: 440-444.
25. GUTAMN E, YOUNG JZ. Reinnervationof the muscle after various periods ofatrophy. J Anat 1946; 78: 15-52.

26. BOWDEN RCM, GUTMANN E. Dener-vation and reinnervation of human volun-tary muscle. Brain 1944; 67: 20-72.
27. LYONS RM, TUCKER HM. Delayedrestoration of abduction in the paralyzedcanine larynx. Arch Otolaryngol 1972; 100:176-179.
28. CRUMLEY RL. Experiments in laryngealreinnervation. Laryngoscope 1982; 92(suppl 30): 1-27.
29. CRUMLEY RL. Regeneration of therecurrent laryngeal nerve. OtolaryngolHead Neck Surg 1982; 90: 442-447.
30. FAULKNER JA, MAXWELL LC,WHITE TP. Characteristics of autograftedmammalian skeletal muscle. In: Mauro A,Bischoff R, Carlson BM, Shafig SA,Konigsbery I, Lipton B, eds. Muscle
Regeneration. New York: Raven Press,1979: 485-492.
31. CARLSON BM, HANSEN-SMITH FM,MAGON DK. The life history of a freemuscle graft. In: Mauro A, Bischoff R,Carlson BM, Shafig SA, Konigsbery I,Lipton B, eds. Muscle Regeneration. NewYork: Raven Press, 1979: 493-507.
32. CARLSON BM, FAULKNER JA. Theregeneration of skeletal muscle fibersfollowing injury: A review. Med Sci SportsExerc 1983; 15: 187-198.
33. CHANG SY. Studies of early laryngealreinnervation. Laryngoscope 1986: 95: 455-457.
34. MEIKLE D, TRACH RE, CUMMINGSCW. Reinnervation of skeletal muscle; acomparison of nerve implantation with
neuromuscular pedicle transfer in an animalmodel. Ann Otol Laryngol 1987: 96: 152-157.
35. DREYFUS, J, SCHAPIRA G. Biochemicde l'atrophie musculaire. Som Hosp (Paris)1948; 24: 1999.
36. FINK B. Folding mechanism of the humanlarynx. Acta Otolaryngol 1974; 78: 124-128.
37. STORE RE, NUTHALL AL. Relativemovements of the thyroid and cricoidcartilage assessed by neural stimulation indogs. Acta Otolaryngol 1974; 78: 134-140.
38. KOTHY MN, HAUGEN LK. The mechan-ics of laryngeal function. Acta Otolaryngol1970; 70: 203-211.
39. FINK RB, BASEK M, EPAUGHIN V. Themechanism of opening of human larynx.Laryngoscope 1956; 66: 410-425.
209