atp synthase: convert a proton gradient into chemical synthesis through connected motors feb 2002...
TRANSCRIPT

ATP Synthase:Convert a Proton Gradient
into Chemical Synthesis Through Connected Motors
Feb 2002Physics 498TBP, UIUC

Photosynthetic Apparatus of Purple Bacteria
LH-II LH-I
cytochrome c2
ATPase
cytoplasm
periplasm
ADP ATP
hbc1
H+
H+
e-
Q/QH2/Q
RC
RC - Photosynthetic Reaction Center
LH – Light Harvesting Complex
ATP-synthase

All enzymes are beautiful, but “ATP synthase” is one of the most:
- Beautiful because of its “3D structure”,
- Unusual because of its “structural complexity and reaction mechanism”,
- Important because “everyday an active graduate student makes more than his/her body weight of ATP”.

ATP + H2O ADP + H+ + PO42- + H+
-12.2 kBT = -30.6 kJ/mol = -7.3 kcal/mol
1. Introduction
hydrolysis
synthesis
(ADP = Adenosine diphosphate)
ATP, universal energy carrier of living systems
(ATP = Adenosine triphosphate)
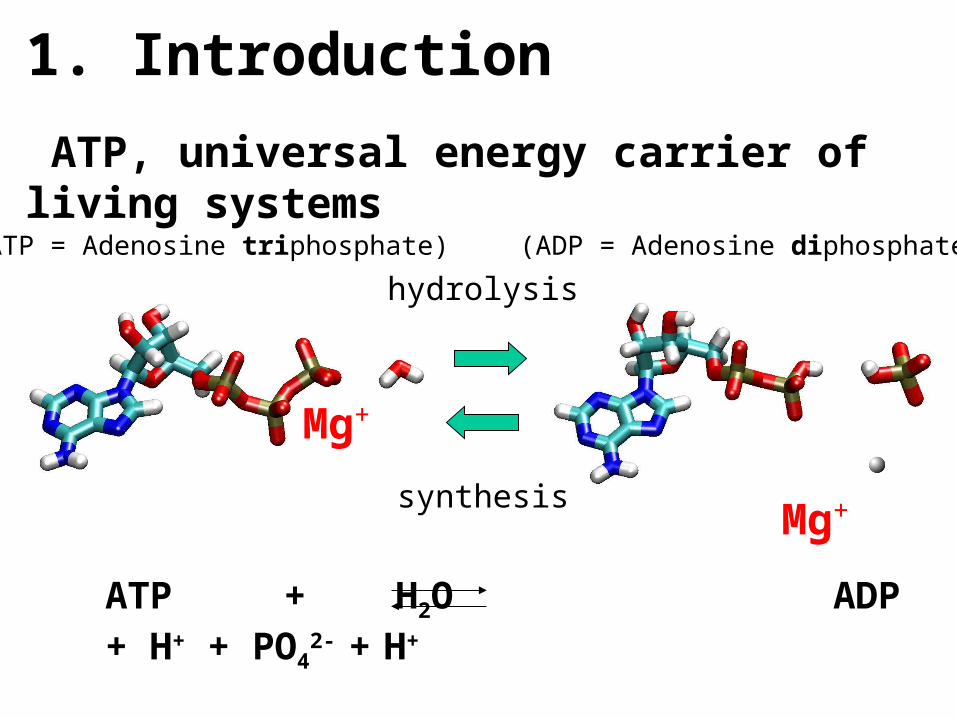
ATP + H2O ADP + H+ + PO42- + H+
-12.2 kBT = -30.6 kJ/mol = -7.3 kcal/mol
1. Introduction
hydrolysis
synthesis
(ADP = Adenosine diphosphate)
ATP, universal energy carrier of living systems
(ATP = Adenosine triphosphate)
Mg+
Mg+

Why do we consume so much ATP?
Many reactions of cell function and growth are thermodynamically unfavorable.
A thermodyamically unfavorable reaction can be driven by a favorable one.
Suppose a cell needs to perform the unfavorable conversion of A into B: A B, G° ' = +4 kcal/mol
K eq ' = [B] eq / [A] eq = 10 (- G° ' / 1.36) = 10 (- (+4) / 1.36) =1.15 10-3
No spontaneous formation of B ([B] / [A] = 1.15 10-3), so most of A remains unconverted.
We can make much more of B if we couple A B with a favorable reaction,
ATP + H2O ADP + Pi + H+ , G° ' ' = -7.3 kcal/mol
Couple hydrolysis of one ATP molecule to every conversion of A into B…

ATP hydrolysis shifts the equilibria of coupled reactions
Coupled reaction:A B G° '= +4 kcal/molATP + H2O ADP + Pi + H+ G° ' ' = -7.3 kcal/mol------------------------------------------------- ---------------------------A + ATP + H2O B+ ADP + Pi + H+ G° tot= - 3.3kcal/mol
[B]eq [ADP] eq [Pi] eq
K 'eq = ------------ • -------------------- = 10 ( - G° ' / 1.36) = 10 (- (-3.3) / 1.36 ) = 2.67 102
[A]eq [ATP] eq
[B]eq [ATP] eq ----- = K 'eq ---------------[A]eq = [ADP] eq [Pi] eq
If the cell keeps it’s [ATP] / ([ADP] [Pi ]) ratio at about 500, [B] eq/ [A] eq = 2.67 102 500 = 1.34 105, most of A has been converted!
Without ATP hydrolysis, this was 1.15 10-3, so
A B conversion has been increased by a factor of 108.
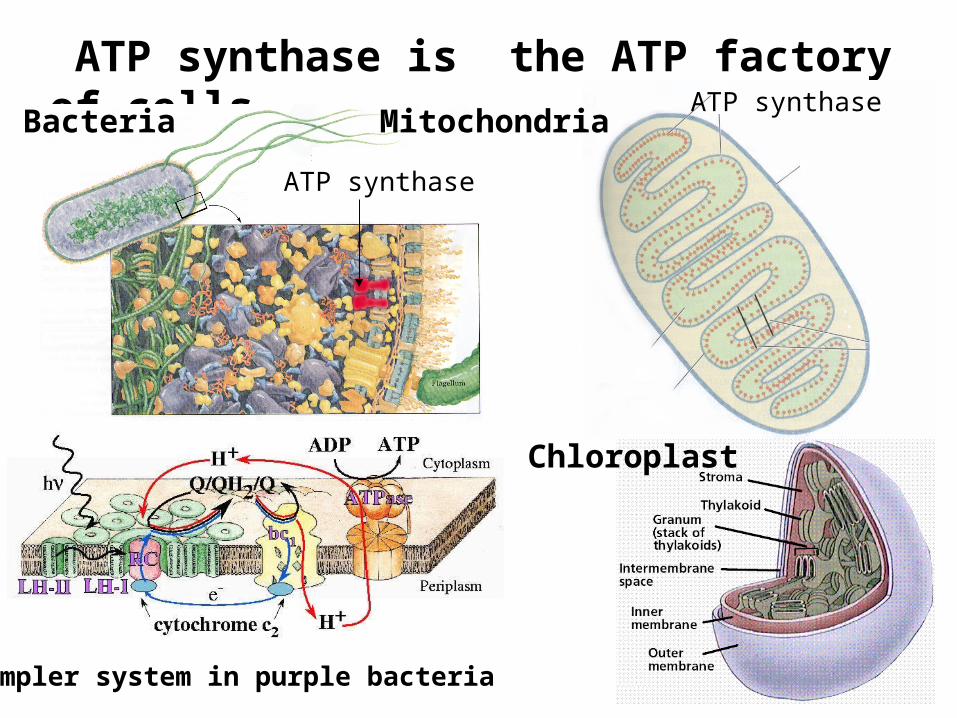
ATP synthase is the ATP factory of cellsATP synthase
Bacteria Mitochondria
Chloroplast
ATP synthase
simpler system in purple bacteria

ATPase is a
molecular motor

Structure of ATP synthase
Soluble part, F1-ATPase- consists of 5 subunits (3 , 3 , 1 , 1 and 1 ) - catalyzes/hydrolyses ATP
Membrane-bound part, F0 Complex- consists of 3 subunits (1 a, 2 b and 9-12 c) - involved in proton translocation.
~ 80 Å
~ 200 Å
~ 60 Å
~ 60 Å
~ 100Å

ATPase Structure Data
Stock et al., Science (1999)
John E. WalkerNobel Prize in Chemistry 1997

Top view
ATP synthase X-ray structure (Walker’94)
C10
Abrahams et al., Nature 370, 621 (1994); Stock et al., Science 286, 1700 (1999)
E
E
DP
DP
TP
TP

One shaft, two motors
Soluble part, F1-ATPase-Synthesizes ATP when torque is applied to it (main function of this unit)-Produces torque when it hydrolyzes ATP (not main function)
Membrane-bound part, F0 Complex- Produces torque when positive proton gradient across membrane(main function of this unit)- Pumps protons when torque is applied (not main function)
~ 80 Å
~ 200 Å
~ 60 Å
~ 60 Å
~ 100Å
Torque is transmitted between the motors via the central stalk.

Hydrogen gradient causes Fo to spin
Junge et al. FEBS 25153 (2001)
It takes about ~20kBT to produce one ATP molecule.
The Fo motor transforms a protonmotive force with PMF =(2.3RT/F)pH + into a rotary torque (R and F are the gas and Faraday constants).
Typical contributions to PMF are: pH difference: 80mV=3.2kBT membrane potential: 140mV=5.6kBT
Thus, at least three protons should pass across the membrane to allow synthesis of one ATP molecule.

Steered Molecular Dynamics (SMD) of F0 unit:• ~100k atoms (17k protein, 39k membrane, 42k solvent)• System size ~100x100x100A. PBC, const T, const p.• Applying constant torque to the central stalk ( subunit).
Focus on:• The central stalk connection with the c subunit assembly;• Rotation of the c subunit assembly in the membrane;• Dynamics of protein residues involved in proton transfer.
Ilya Balabin
a
b1
b2
c
F0 ATP Synthase Converts Proton Gradient into Rotation
How can a local (3 A) proton transfer reaction turn the large (20 A) c-subunit ?

How local H+ motion generates a rotary torque
Brownian forces induce random rotations of the rotor in both directions
The binding sites near the stator can lose or gain protons depending on the rotor position and proton concentration outside of the channel.
Only 2 out of 12 binding sites can be simultaneously in the vicinity of the stator
Unprotonated binding sites cannot leave the rotor/stator interface because of the energy barrier of ~45kBT caused by the difference in the dielectric constants (mem=3,stator=10)

G. Oster and H Wang (1998)
Due to asymmetry of the proton concentrations, the state is more likely to occur then the and states. Therefore, the rotor moves on average to the left.
The charged group at the middle of the stator prevents the rotor in the state from moving to the right. The proton will dissociate back to the channel when approaching a positively charged group. This increases efficiency of energy utilization by the motor.Summarizing: the difference in proton concentrations and asymmetry of the stator geometry facilitate transformation of a random motion into a unidirectional rotation.

The Simulation

ATP F1 Synthase
t (ps)
RM
SD, C
a (A
)
stalk only
full protein
Simulation conditions: Explicit solvent MD, Full Coulomb int., Particle Mesh Ewald, system prepared in NpT, final equilibration dynamics in NVE4.6 days/ns on 256 processors (Origin 2000), 327,000 atoms total, 51,000 atoms -- protein and nucletoide, 276,000 atoms -- water and 100 mM [salt]

N
C
-subunit
Bottom view
ATP synthesis mechanism
Boyer, P. D. Biochim. biophys. Acta 1140, 215 (1993)
3.1 Binding Change Mechanism (Boyer’93)
The applied torque causes rotation of the -subunit which causes cyclic transformation of three catalytic sites.

Synthesis: Binding Change Mechanism
DP (Loose)- Loosely binding ligands.- Catalytically “inactive”.
TP (Tight)- Tightly binding ligands.- Catalytically “active”.
E (Open)- Low affinity for ligands- Catalytically “inactive”
Energy of proton gradient is used to- BIND ADP and Phosphate- RELEASE tightly bound ATP
G. Oster and H Wang (1998)

Hydrolysis is Synthesis in Reverse
Boyer, P. D. Biochim. biophys. Acta 1140, 215 (1993)
ATP hydrolysis can be thought of as a rotary engine.

Direct Observation of ATPase Rotary motion
Noji et al., Nature 386, 299 (1997); Pänke et al. FEBS Lett. 472, 34 (2000)
Yoshida’97

Summary
1. ATP synthase produces ATP via a rotary mechanism. The electrochemical energy from the cross-membrane proton gradient is changed into a torque, and the torque changed into chemical bond energy.
2. ATP synthase can operate in reverse under the proper conditions, changing chemical bond energy to a torque, then the torque into a proton gradient.

The Brownian Ratchet