are the platyhelminthes a monophyletic primitive group? an ...molevol.cmima.csic.es/carranza/pdf/are...
TRANSCRIPT

Are the Platyhelminthes a Monophyletic Primitive Group? An Assessment Using 18s rDNA Sequences
Salvador Carranza, Jaume Bagu&i, and Marta Riutort Departament de Genetica, Facultat de Biologia. Universitat de Barcelona, Spain
In most zoological textbooks, Platyhelminthes are depicted as an early-emerging clade forming the likely sister group of all the other Bilateria. Other phylogenetic proposals see them either as the sister group of most of the Protostomia or as a group derived from protostome coelomate ancestors by progenesis. The main difficulty in their correct phylogenetic placing is the lack of convincing synapomorphies for all Platyheh-ninthes, which may indicate that they are polyphyletic. Moreover, their internal phylogenetic relationships are still uncertain. To test these hypotheses, new complete 18s rDNA sequences from 13 species of “Turbellaria” have been obtained and compared to published sequences of 2 other “Turbellaria,” 3 species of parasitic Platyhelminthes, and several diploblastic and deuterostome and protostome triploblastics. Maximum-parsimony, maximum-likelihood, and neighbor-joining meth- ods were used to infer their phylogeny. The results show the order Catenulida to form an independent early- branching clade and emerge as a potential sister group of the rest of the Bilateria, while the rest of Platyhelminthes (Rhabditophora), which includes the parasites, form a clear monophyletic group closely related to the protostomes. The order Acoela, morphologically considered as candidates to be ancestral, are shown to be fast-clock organisms for the 18s rDNA gene. Hence, long-branching of acoels and insufficient sampling of catenulids and acoels leave their position still unresolved and call for further studies. Within the Rhabditophora, our analyses suggest (1) a close relationship between orders Macrostomida and Polycladida, forming a clear sister group to the rest of orders; (2) that parasitic platyhelminthes appeared early in the evolution of the group and form a sister group to a still- unresolved clade made by Nemertodermatida, Lecithoepitheliata, Prolecithophora, Proseriata, Tiicladida, and Rhab- docoela; and (3) that Seriata is paraphyletic.
Introduction
The origin of Bilateria (organisms which display bilateral symmetry and clear anteroposterior polarity) can be considered the most important unsolved problem in systematic biology. These animals also share the char- acteristic of possessing three clearly distinct cell layers (i.e., they have a true mesoderm); hence, they are col- lectively called triploblastics. Historically, the presence/ absence of a true coelom (i.e., a system of cavities with- in the mesoderm), and, hence, whether the most primi- tive body form was acoelomate, pseudocoelomate, or coelomate, has been the main issue in discussion of the origin of the Bilateria. The most classical view, adopted in the majority of zoological textbooks, sees the acoe- lomate Platyhelminthes as an early-emerging clade forming the likely sister group of all the other Bilateria, which themselves would be divided into two coelomate supergroups, protostomes and deuterostomes (Hyman 1951; Salvini-Plawen 1978; fig. 1A). Since egg cleavage in Platyhelminthes is spiral, as in most protostomes, while it is radial in most deuterostomes, this proposal implies several modifications of early embryogenesis in the deuterostome line with respect to the protostome one. To skip that problem, another phylogenetic scheme sees two Bilateria supergroups: the “Spiralia” on the one hand, including the Platyhelminthes, and the “Ra- dialia” on the other (fig. 1B). Under that scheme, the coelom must therefore have originated twice, once in
Key words: 18s rDNA, Platyhelminthes, phylogeny, metazoan, monophyletic primitive group.
Address for correspondence and reprints: Dr. Marta Riutort, Depar- tament de Genktica, Facultat de Biologia, Universitat de Barcelona, Di- agonal 645, 0807 1 Barcelona, Spain. E-mail: [email protected].
Mol. Biol. Evol. 14(5):485497. 1997 0 1997 by the Society for Molecular Biology and Evolution. ISSN: 0737-4038
the Spiralia (after the split of one branch from Platy- helminthes) and once in the Radialia (Ax 1987; Brusca and Brusca 1990). A third view is based on the idea that the gastral pouches of coelenterates are homologous with the gastral pouches (enterocoels) that give rise to the coeloms in deuterostomes. Therefore, features of deuterostome development are assumed to be primitive among Bilateria. In this scheme, protostome develop- mental features are derived, coelom is of early origin, Platyhelminthes are considered derived from a coelo- mate ancestor by progenesis (Rieger 1985) or by reduc- tion of coelomic cavities in the adult (Remane, Starch, and Welsch 1980), and the hypothetical ancestor of the Bilateria would be an “archicoelomate” (Siewing 1980) (fig. 10
Knowledge of the actual phylogenetic position of Platyhelminthes is paramount to decide among these al- ternatives. Platyhelminthes, or flatworms, display a va- riety of body forms and are succesful inhabitants of a wide range of environments. The majority of their 20,000 extant species are parasitic (classes Trematoda, Monogenea and Cestoda). The free-living forms (class Turbellaria) are primarily epifaunal or infaunal inhabi- tants of the marine and freshwater benthos, but marine and freshwater pelagic and terrestrial forms also occur. The free-living forms range from less than 1 mm to about 50 cm long. Some parasites (tapeworms) may at- tain lengths of several meters. Major diagnostic features of the phylum (synapomorphies) are disputed. Ehlers (1985, 1986; see also Ax 1987) has proposed as auta- pomorphies some features of protonephridia, multicilia- tion in epidermal cells, and absence of mitosis in epi- dermal and other somatic cells, but such views have been contested (Smith, Tyler, and Rieger 1986; Rohde 1990). This casts doubt on the monophyly of the Pla-
485
_ ~_. ---

486 Carranza et al.
* - Cnidaria
Platyhelminthes
Protostomia
Deuterostomia
Cnidaria
Platyhelminthes
Protostomia
Deuterostomia ‘Radialia’
Platyhelminthes
Protostomia
Deuterostomia
FIG. 1 .-Conflicting traditional phylogenies 01 nl the origin of Pla-
Bilateria
‘Spiralia’
Bilateria
tyhelmmthes and the acoelomate condition. A, evolutionary tree based on the assumption that the acoelomate condition is primitive within the triploblasts. Under this view, Platyhelminthes form the sister group of all the other Bilateria, which themselves would be divided into two coelomate supergroups, protostomes and deuterostomes (after Hyman, 1951). B, An evolutionary tree based on a very early splitting of Bi- lateria into two supergroups, the “Spiralia,” including the Platyhel- minthes, and the “Radialia.” In this scheme, the acoelomate condition is primitive within the triploblastic spiralians, making Platyhelminthes the first descendant group of this lineage but not the sister group of the Bilateria (after Ax 1987; Brusca and Brusca 1990). C, An evolu- tionary tree based on the proposal that the acoelomate condition arose through neoteny from developmental stages of protostomes prior to the embryonic appearance of coelomic cavities. In this scheme, coelomic cavities (enterocoels) in deuterostomes are homologous to gastral pouches of the diblastic coelenterates, protostome coelomic cavities (schizocoels) are derived, and the hypothetical ancestor of the bilateria would be an “archicoelomate” (after Sewing 1980; Rieger 1985).
tyhelminthes. Indeed, the common denominator of all recently proposed phylogenetic schemes for the platy- helminths is recognition of three clearly monophyletic groups: one containing the Acoela and Nemertoderma-
tida (“Acoelomorpha” of Ehlers 1985, 1986), one con- taining the Catenulida, and the third containing all other turbellarian orders together with the parasitic classes (“Rhabditophora” of Ehlers 1985, 1986) (fig. 2A). As there are no convincing synapomorphies for all Platy- heminthes, this may indicate that Platyhelminthes are polyphyletic and that Catenulida, Acoelomorpha, and Rhabditophora may be unrelated taxa. As first suggested by Ehlers (1985, 1986) and Ax (1987), this means that the free-living class, the Turbellaria, are paraphyletic. Indeed, features defining it (free-living life style and the body covered by a ciliated epidermis) are plesiomor- phies (Ehlers 1985, 1986; Ax 1987). Finally, affinities between the different “turbellarian” orders, as well as whether a parasitic group is monophyletic or polyphy- letic and, if the former, which is its closest “turbellari- an” taxon, are uncertain and a matter of debate. Al- though phylogenetic schemes, based mainly on morpho- logical characters and electron microscope features, have been proposed (Karling 1974, fig. 2b; Ehlers 1985; Rohde 1990), presently available information cannot de- cide among them.
Morphological and embryological comparisons be- tween Platyhelminthes and presumed close phyla (e.g., gnathostomulids, pseudocoelomates, gastrotrichs, acan- thocephalans, rotifera) have so far been unable to answer the main question on the phylogenetic position of Pla- tyhelminthes as related to the rest of the Bilateria and its mono- or polyphyletic status. This is because these phyla share few informative homologous anatomical or embryological features and because it is very difficult to distinguish homologous similarity (homology) from convergent or parallel similarity (analogy). Moreover, most Platyhelminthes are small and soft-bodied organ- isms which lack a fossil record. Sequence data obtained from ribosomal RNA or DNA offer an important new source of informative characters for inferring high-level phylogenetic relationships for many taxa and provide an independent test of hypotheses based on morphological characters (Woese 1987; Field et al. 1988; Turbeville, Field, and Raff 1992; Adoutte and Philippe 1993; Riu- tort et al. 1993; Smothers et al. 1994; Winnepenninckx, Backeljau and De Wachter 1995; Friedrich and Tautz 1995; Bridge et al. 1995). The main reasons for using 18s rDNA or rRNA have been repeatedly reviewed (Woese 1987; Sogin 1991).
18s rDNA sequences, either partial or complete, from some species of Platyhelminthes have been used either in general molecular analysis of animal evolution (Field et al. 1988; Riutort et al. 1993; Adoutte and Phi- lippe 1993; Philippe, Chenuil, and Adoutte 1994; Wada and Satoh 1994) or in specific studies of particular phyla or groups of phyla (e.g., protostome worms: Winnepen- ninckx, Backeljau, and De Wachter 1995; Myxozoa: Smothers et al. 1994; Dicyemid Mesozoa: Katayama et al. 1995; Nemertini: Turbeville, Field, and Raff 1992; Arthropoda: Turbeville et al. 1991). In most studies, Pla- tyhelminthes appear as the sister group of Bilateria. In others, they cluster with nematodes, acanthocephalans, and some protostomes in ill-defined paraphyletic groups. Ingroup studies of Platyhelminthes phylogeny have

The Phylogenetic Status of Platyhelminthes 487
Catenulida I Catenulida Acoela
Nemertodermatida I Acoelomorpha
Macrostomida
Polycladida
Lecithoepitheliata
Prolecithophora
Proseriata Rhabditophora
Tricladida Rhabdocoela
l’gphloplanoitla
Rhabdocoela Dalgellioida
Neodermata
B
Catenulida
Acoela
Nemertodermatida Macrostomida
Polycladida
Lecithoepitheliata
Rhabdocoela
Neodermata
Prolecithophora
Proseriata
Tricladida
FIG. 2.-A, Phylogeny of Platyhelminthes according to Ehlers (1985). B, Evolutionary tree depicting the relationships among the orders of Turbellaria and the parasitic platyhelminths (after Karling 1974).
mainly concerned parasitic groups (Baverstock et al. 1991; Blair 1993; Rohde et al. 1993, 1995) and a few turbellarian orders (Riutort et al. 1992, 1993; Katayama et al. 1993; Rohde et al. 1993, 1995). The main conclu- sions are that all the major parasitic groups constitute a monophylum, that class Trematoda is monophyletic whereas Monogenea may be paraphyletic, that there are contradictory views as regards the closest free-living turbellarian to parasitic groups, and that Platyhelminthes may be polyphyletic (with acoels forming a separate clade; Katayama et al. 1995). Most of these data, how- ever, should be taken very cautiously because most stud- ies used only partial 18s sequences and because “low- er” platyhelminth orders (Catenulida, Nemertodermati- da, Lecithoepitheliata, and Macrostomida) either were not included or were represented by a single species.
We report here the complete 18s rDNA sequences of 13 species of free-living “Turbellaria” belonging to 10 orders. Published complete sequences of two other free-living turbellaria and three parasitic species were also included in some of the analyses. Sequence data were analyzed with maximum-parsimony, distance-ma- trix, maximum-likelihood, and pattern of resolved nodes (PRN) methods. Our main focus has been to test: (1) the monophyletic or polyphyletic status of the Platy- helminthes, (2) whether Platyhelminthes can be consid- ered the sister group of the rest of bilaterians, (3) the internal relationships among the extant “turbellarian” orders, and (4) which is the closest free-living represen- tative of the parasitic groups.
Materials and Methods Organisms
Organisms were chosen to provide representatives of the 11 orders of Turbellaria. In addition, published Platyhelminthes sequences were used. Species repre- sented are shown in table 1, and the 13 species for which new sequence data were collected are indicated. Because there is no consensus as to the sister group of Platy- helminthes (see Introduction), we used as outgroup the
choanoflagellate Sphaeroeca volvox. Since diploblastics have been clearly shown to be the sister group of all Triblastics, the cnidarian Anemonia sulcatu, the cteno- phore Beroe cucumis, and the placozoan Trichoplux ad- haerens were also included in the analysis. Represen- tatives of Deuterostomia were the starfish Asterias amu- rensis, the cephalochordate Brunchiostoma jloridue, and the amphibian Xenopus laevis, while Aphonopelma sp. (Arthropoda, Aracnida), Odiellus troguloides (Arthro- poda, Arachnida), Eisenia fetidu (Annelida, Oligochae- ta), Lunice conchilega (Annelida, Polychaeta), Liolo- phuru juponica (Mollusca, Polyplacophora), Crussos- trea virginica (Mollusca, Bivalvia), and Prostoma eil- hardi (Nemertea) were used as representatives of Protostomia. Because recent analyses indicate that pseu- docoelomates, namely the phyla Nematoda and Nema- tomorpha, may form the sister group of the rest of Bi- lateria, and since Gastrotricha and Acanthocephala ap- pear as a likely sister groups of Platyhelminthes (Win- nepenninckx et al. 1995; Doolittle et al. 1996), sequences from the nematomorph Gordius aquaticus, the gastrotrich Lepidodermella squammata, and the acanthocephalan Moliniformis moliniformis were includ- ed in some of the analyses. All nonplatyhelminth species are listed in table 2.
DNA Extraction, Amplification, and Sequencing
High-molecular-weight DNA was purified accord- ing to a modification (Garcia-Fernandez, Bagufia, and Sal6 1993) of the guanidine isothiocyanate method ini- tially described for RNA (Chirgwin et al. 1979). The entire length of the 18s rDNA was amplified in two fragments of approximately equal length by the poly- merase chain reaction (Saiki et al. 1985) using the prim- ers: 1E TACCTGGTTG ATCCTGCCAG TAG; 5R, CTTGGCAAAT GCTTTCGC; 5F, GCGAAAGCAT TTGCCAAGAA; and 9R, GATCCTTCCG CAGGTT- CACC TAC. The resulting fragments were either blunt end cloned and sequenced as described elsewhere (Car- ranza et al. 1996) or directly sequenced (Nemertinoides,

488 Carranza et al.
Table 1 Table 2 List of Platyhelminth Species Used in this Study and GenBank Accession Numbers
List of Nonplatyhelminth Species Included in this Study and GenBank Accession Numbers
Accession Number
Name in Figures
Class Turbellaria
Order Tricladida
Crenobia alpina* . . . . . . . . . . . M58345 Dendrocoelum lacteum* . . . . . M58346
Order Proseriata
Monocelis lineata* . . . . . . . . . . U45961 Archiloa rivularis” . . . . . . . . . . U70077
Order Lecithoepitheliata
Geocentrophora sp.* . . . . . . . . U70080
Order Rhabdocoela
Mesocastrada sp.* . . . . . . . . . . U70082
Order Nemertodermatida
Nemertinoides elongatus* . . . . U70084
Order Prolecithophora
Urastoma sp.*. . . . . . . . . . . . . . U70086
Order Macrostomida
Macrostomum tuba*. . . . . . . . . U7008 1 Microstomum lineare* . . . . . . . U70083
Order Polycladida
Discocelis tigrina* . . . . . . . . . . U70079 Planocera multitentaculata . . . D17562
Triclad- 1 Triclad-2
Proseriate- 1 Proseriate-2
Lecithoepitheliate
Rhabdocoel
Nemertodermate
Prolecithophoran
Macrostomid- 1 Macrostomid-2
Polyclad- 1 Polyclad-2
Order Catenulida
Stenostomum leucops” . . . . . . . U70085 Catenulid
Order Acoela
Convoluta pulchra” . . . . . . . . . U70078 Acoel- 1 Convoluta naikaiensis . . . . . . . D17558 Acoel-2
Class Cestoda Order Cyclophyllida
Echinococcus granulosus. . . . . U27015 Cestode
Class Trematoda
Subclass Aspidogastrea
Order Aspidobothria
Lobatostoma manteri . . . . . . . . L 169 11
Subclass Digenea
Trematode- 1
Order Schistosomida Schistosoma mansoni. . . . . . . . M62652 Trematode-2
NOTE.--New sequences reported in this paper are marked with an asterisk.
Mesocastrudu, and Archiloa). Both chains were com- pletely sequenced.
Sequence Alignment and Phylogenetic Analyses
Sequence data were aligned by hand with the help of a computer editor. Alignment gaps were inserted to account for putative length differences between the se- quences. A secondary-structure model (Gutell et al. 1985) was used in order to optimize alignment of ho- mologous nucleotide positions, resulting in a total of 1,322 positions that could be used in the phylogenetic analyses (675 being variable and 428 being parsimony- informative when the 33 species are compared).
Distance analyses were calculated using the PHY- LIP program package v. 3.52 (Felsenstein 1993) and MUST v. 1.0 (Philippe 1993). A distance matrix of the
Phylum Chordata
Accession Number
Name in Figures
Subphylum Cephalochordata
Branchiostoma j7oridae . . . . . . . M9757 1 Cephalochordate Subphylum Vertebrata
Xenopus laevis . . . . . . . . . . . . . . X04025 Vertebrate
Phylum Echinodermata Asterias amurensis . . . . . . . . . . . D 14358 Echinoderm
Phylum Mollusca
Class Polyplacophora Liolophura japonica. . . . . . . . . . X702 10 Mollusc- 1
Class Bivalvia Crassostrea virginica. . . . . . . . . X603 15 Mollusc-2
Phylum Arthropoda Class Arachnida
Order Opiliones Odiellus troguloides . . . . . . . . . . X8 144 1 Arthropod- 1
Order Araneae Aphonopelma sp. . . . . . . . . . . . . X 13457 Arthropod-2
Phylum Annelida Class Clitellata
Eisenia fetida . . . . . . . . . . . . . . . X79872 Annelid- 1 Class Polychaeta
Lanice conchilega . . . . . . . . . . . X79873 Annelid-
Phylum Nemertini
Prostoma eilhardi . . . . . . . . . . . U29494 Nemertine
Phylum Gastrotricha Lepidodennella squammata. . . . U29198 Gastrotrich
Phylum Nematomorpha
Gordius aquaticus . . . . . . . . . . . X87985 Nematomorph
Phylum Acanthocephala
Moliniformis moliniformis . . . . . 219562 Acanthocephalan
Phylum Placozoa Trichoplax adhaerens . . . . . . . . L 10828 Placozoan
Phylum Cnidaria
Anemonia sulcata. . . . . . . . . . . . X53498 Cnidarian
Phylum Ctenophora Beroe cucumis . . . . . . . . . . . . . . D 15068 Ctenophore
Phylum Sarcomastigophora
Sphaeroeca volvox. . . . . . . . . . . 234900 Choanoflagellate
aligned sequences was generated using the program DNADIST and corrected with the two-parameter meth- od of Kimura (1980). The distances were then converted to phylogenetic trees using FITCH (Fitch and Margo- liash 1967) and the neighbor-joining (NJ) method of Sai- tou and Nei (1987) provided by the NEIGHBOR pro- gram. Bootstrap resampling (Felsenstein 1985) was ac- complished with the use of the programs SEQBOOT (1,000 replicas) and CONSENSE. FASTDNAML v. 1.1. la (with global rearrangements and reordering of species) was used for maximum-likelihood analyses (Felsenstein 1981; Olsen et al. 1994), and a bootstrap analysis (n = 100) was performed. Maximum-parsi- mony (MP) analyses (Camin and Sokal 1965) were cal- culated with the PAUP computer program v. 3.1.1

The Phylogenetic Status of Platyhelminthes 489
100 T&lad-2 -&lad-l
I Rhabdocoel
r, Lecithoepitheliate
Proleci thonhoran
Macrostokrid-1 Macrostomid-2
Catenulid Gastrotrich
I I ’ Acanthocephalan
I
Choanoflagellate
Acoel-2 Acoel-1 I
I I
0.05
DEUTEROSTOMIA
PROTOSTOMIA + ASCHELMINTHES”
‘PLATYHELMINTHES’
ASCHELMINTHES
‘PLATYHELMINTHES
DIPLOBLASTS
PROTOZOA
FIG. 3.-Neighbor-joining tree including two acoel sequences. All branch lengths are drawn to scale. Note long branches leading to and separating the two acoel species, which indicates they may be fast-clock organisms. Lack of resolution within the Bilateria clade results from the loss of information due to the difficulty in aligning the acoel sequences. Note that Platyhelminthes cannot be considered monophyletic because some Aschelminthes branch between different Platyhelminthes clades, here represented as “Platyhelminthes.” Numbers at nodes are percentages of 1,000 bootstrap replicates that support the branch. Values are shown only if over 50%. This is an unrooted tree, rooted defining the choanoflagellate as the outgroup. For species names, see tables 1 and 2.
(Swofford 1993) using a heuristic search procedure and a branch-swapping algorithm. A bootstrap analysis (n = 100) was also performed. The trees were rooted using the SSU rRNA sequence of the “choanoflagellate” Sphaeroeca volvox. In all analyses, gaps were consid- ered as missing data.
PRN (Lecointre et al. 1994) was performed using the program MUST v. 1.0, to prepare the data, and PRN v. 1 to test the robustness of the nodes obtained. Instead of simply examining the bootstrap proportions (BP) at important nodes as a criterion of robustness of the cor- responding nodes, PRN introduces a procedure of BP analysis which involves following the values of BP as a function of increasing number of nucleotides. PRN was used for the same taxa used in the distance analysis, but only the parsimony-informative positions were in- cluded, under the following conditions. The alignments of the N species were each submitted to random sam- pling of a given number of sites through the use of the PRN program running on UNIX platforms. Ten different sequence lengths were chosen (10, 15, 25, 50, 75, 100, 150, 200, 250, 300), and, for each, 200 samples were drawn. Thus a total of 2,000 subsets of sequence align- ments were obtained, each including all N species. Each of these subsets was used to construct a neighbor-joining tree, which was submitted to 1,000 bootstrap replicates.
Selection of the nodes was then carried out to keep only those with an ascending tendency and that appeared more frequently. At a given node, one could therefore graphically display the evolution of BP as a function of the number of nucleotides used to generate the tree.
The sequence alignment and the weighting mask which determined the nucleotide positions taken for the analyses are available from the ftp site: por- thos.bio.ub.es/users/ftp/pub/teu/l8Sphylogeny.
Results
A distance tree including all the sequences is rep- resented in figure 3. Some features are consistent with what is known of animal evolution. As expected, the diploblast branched before all triploblast groups. How- ever, the most interesting feature of this tree is the long branches leading to and separating the two acoel rep- resentatives. In fact, acoel sequences diverge so much from the rest of the organisms studied (not only from Platyhelminthes) that alignment of sequences turned out to be very difficult. Nonetheless, at the cost of losing many informative sites, the two acoels and all the rep- resentatives from the other groups (diploblasts, protos- tomes, “aschelrninthes,” deuterostomes, and platyhel- minths) were included. The analysis considers only

490 Carranza et al.
Cephalochordate
J 1001 Arthropod-2
Arthropod-l
Triclad-1 T&lad-2
Rhabdocoel Proleci thonhoran
Proseriate-1 a Proseriate-2
__I A Lecithoenitheliate 111
r-l, 100 Tremkode-1 1001 Trematode-2
i 99 q Nemertodern%fode
r Polvclad-1
Catenulid Placozoan
Cnidarian C tenophore
Choanoflagellate PROTOZOA
DEUTEROSTOMIA
PROTOSTOMIA
ASCHELMINTHES
PLATYJTELMINTHES
DIPLOBLASTS
FIG. 4.-Distance tree. Fitch-Margoliash (F-M) and neighbor-joining (N-J) methods gave the same topology, only the second being rep- resented here. Note that Platyhelminthes cannot be considered monophyletic because the Catenulida form an early-branching sister group to the rest of Bilateria, whereas the rest of Platyhelminthes (=Rhabditophora + Nemertodermatida) constitute a clear monophyletic group which appears as the sister group of a protostomate + deuterostomate clade. Numbers at nodes are percentages of 1,000 bootstrap replicates that support the branch, only values over 50% being represented, with the exception of the bilaterian sister group to Catenulida (45%). All branch lengths are drawn to scale. For species names, see tables 1 and 2.
those positions that were unambigously aligned for the two acoels. This reduced the available positions from 1,322 to 1,13 1 and the variables from 675 to 5 12. In this tree (rooted with the choanoflagellate Sphaeroeca volvox), the acoels constitute an early sister branch to the rest of bilaterians (67% bootstrap). However, the loss of informative sites leaves the internal relationships of the bilaterian group unresolved. The very long branches of the two acoels suggest that we are in front of fast- clock organisms; hence, their position in the tree could be artifactual. To avoid their disturbing influence, they were not included in subsequent analyses.
The distance analyses, Fitch-Margoliash and neigh- bor-joining methods, not including the acoels, resulted in identical topologies (fig. 4). Two general features of the tree were unexpected. First, Platyhelminthes appear to be a paraphyletic group, as the order Catenulida do not cluster with the rest of Platyhelminthes. Although weakly supported (45% bootstrap), Catenulida appears in this tree as the most primitive bilaterian, as it is the first branching after the diblastics. PRN analysis shows the node for Catenulida + Diplobasts + choanoflagel- late versus the rest of organisms to be a promising node (results not shown), although almost 2,000 parsimony- informative sites will be needed to obtain 100% boot-
strap. Second, the rest of Platyelminthes forms a mono- phyletic clade (99% bootstrap) which appears, although only weakly supported (less than 50% bootstrap), as the sister group of the rest of the Bilateria. However, pro- tostomes do not form a monophyletic group but a para- phyletic one, as nematomorphs, acantocephalans, and gastrotrichs branch inside them. Moreover, and even more significant, the degree of divergence inside the Pla- tyhelminthes ranks with that among protostomes and deuterostomes (compare the long branches separating any two platyhelminths to the branches separating pro- tostomes or deuterostomes).
The 50% majority-rule consensus tree of 10 max- imum-parsimony (MP) trees of equal length is shown in figure 5. It has a length of 1,943 steps, with a consis- tency index (CI) of 0.495 and a retention index (RI) of 0.504. Platyhelminthes and Deuterostomia are well-sup- ported monophyletic groups (91% and 8 l%, respective- ly), while the Protostomia show a sequential branching pattern not well supported by bootstrap (only 27%), sim- ilar to that obtained in the distance tree (fig. 4). Al- though most of the more parsimonious trees (9 of 10) group platyhelminths with deuterostomes, this grouping is not supported by bootstrap (only 24%). This analysis and the bootstrap analyses suggest that the available se-

1. - -. -~ _._ ___~___ -________~ -
The Phylogenetic Status of Platyhelminthes 491
c3;gate I d
241 lOO- Proleci thophoran Rhabdocoel Leci thoepi theli ate Proseriate-1 Proseriate-2 Nemertodermate Trematode-1 Trematode-2 Cestode Polyclad-1 Polyclad-2 Macrostomid-2 Macrostomid-1 Annelid-l Annelid- Mollusc-1 Nemerti ne Mollusc-2 Arthropod-l Arthropod-2 Catenuli d Placozoan Cnidarian Ctenophore Choanoflagellate
60 34 66 801
24 100 10 90
86 100
54 80
71 91 100
100 100 1001
47 10 100
100 27 90
100
I DEUTEROSTOMIA
‘PLATYJTELMINTHES
PROTOSTOMIA
1 ‘PLATYHELMINTHES
I DIPLOBLASTS
1 PROTOZOA
FIG. 5.-Fifty percent majority-rule consensus of 10 most parsimonious trees (percentiles indicated below the nodes). Bootstrap values are indicated above the nodes, only those over 50% being shown. Total length of the tree is 1,943 steps; the overall consistency index is 0.495. Branch lengths are unrelated to evolutionary distance. The topology of this tree only differs from the distance tree of figure 4 in that the monophyletic ‘Platyhelminthes’ (=Rhabditophora + Nemertodermatida; Ehlers 1985) form a clade with the deuterostomes. However, the low bootstrap values for this clade and for the protostome lineage suggests instead a polytomy (see text). For species names, see tables 1 and 2.
quence data are insufficient to unambiguously infer the branching order among the three groups (protostomes, deuterostomes, and platyhelminthes). The order Caten- ulida is, again, the sister group of the rest of bilaterians, with a bootstrap value of 71%.
Maximum likelihood (ML; fig. 6) yielded a similar topology to the distance tree (fig. 4) as regards the par- aphyly of protostomates. Again, pseudocoelomates (here represented by the nematomorph Gordius) fall within the protostomate clade branching with the two arthro- pods and forming a sister group to a group formed by the monophyletic Platyhelminthes (excluding the Caten- ulida) and the other pseudocoelomates, the acantho- cephalan Moliniformis, and the gastrotrich Lepidoder- mellu. In this analysis, Platyhelminthes as well as pseu- docoelomates appear buried within a general protosto- mate clade. Again, the exception is the Catenulida, which is the first taxon branching after the diploblastics, although the bootstrap value for the Protostome + Deu- terostome + Platyhelminthes clade is again low (45%).
The resulting internal phylogeny of Platyhelmin- thes is basically the same with all the methods used (figs. 3-6). As stated above, Catenulida do not cluster inside the Platyhelminthes, and it is not possible to es- tablish the position of Acoela. The main monophyletic group corresponds to Rhabditophora (sensu Ehlers 1985, 1986) plus Nemertodermatida and includes the parasitic species (Neodermata). This group divides, in turn, into two monophyletic clades: one including the orders Ma- crostomida and Polycladida; the other, the rest of the
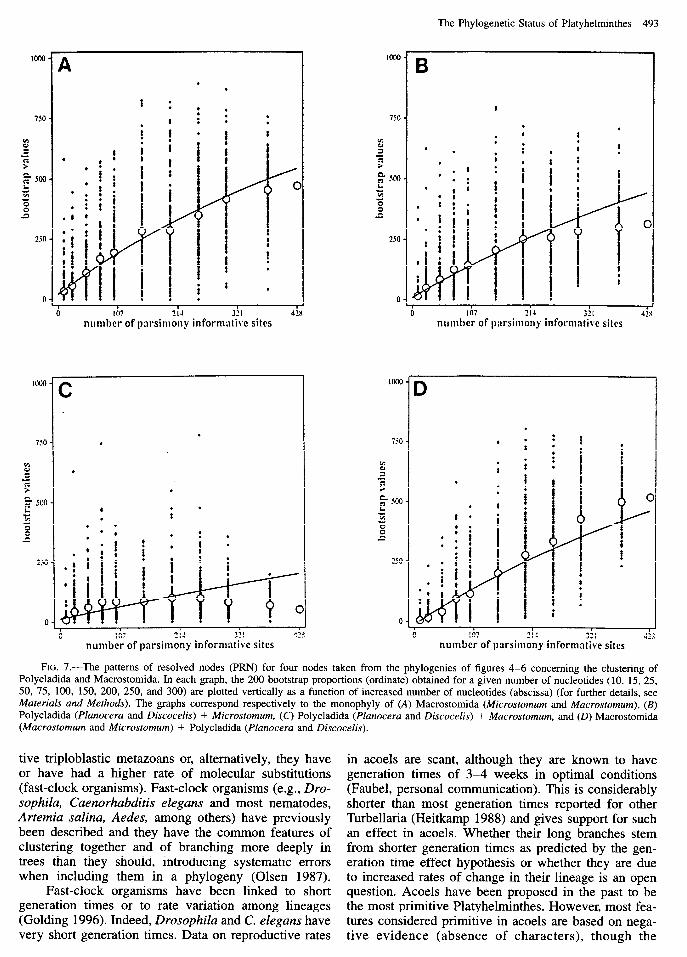
orders including the parasitic classes. Maximum-parsi- mony trees, however, show an early branching of Mu- crostomum (family Macrostomidae) as compared to Mi- crostomum (family Microstomidae) plus Polycladida (fig. 5). Moreover, in the ML tree (fig. 6) we find the two macrostomids branching independently within this clade. PRN analysis shows this to be due to the fact that Macrostomum is more divergent from the Polycladida than Microstomum is (fig. 7). The node for Microsto- mum + Macrostomum (fig. 7A) is better supported than the ones joining Polycladida to either Microstomum (fig. 7B) or Mucrostomum (fig. 7C), and nearly as well as the one joining Polycladida and Macrostomida as a whole (fig. 70). However, the uncertain clustering of Macrostomum with the Polycladida, due to a greater di- vergence, should cause the existence of a greater number of homoplasies of this species to other species from out- er groups, and this makes it branch independently in the MP and ML analysis.
Leaving aside the clade formed by Macrostomida and Polycladida, the internal relationships of the rest of Rhabditophora + Nemertodermatida are not well re- solved, with the exception of three clear monophyletic lineages: Tricladida (Crenobia + Dendrocoelum), Pro- seriata (Monocelis + Archiloa), and the parasites (Lob- atostoma, Echinococcus, and Schistosoma), all with maximum bootstraps (100%).
Discussion This paper explores one of the most important un-
solved problems in systematic biology, the origin of Bi-

492 Carranza et al.
L-r 56 Mollusc-1 - I Annelid-l
I Nematomorph Gastrotrich
AcanthoceDhalan I Triilad-1 c Triclad-2
Rhabdocoel Leci thoepi theliate
Proleci thophoran
_jJ_ Cestode
_gj Nemertodermate ,- Macrostomid-1
- Macrostomid-2 Catenulid
DEUTEROSTOMIA
PROTOSTOMIA
ASCHELMINTHES
PLATYHELMINTHES
DIPLOBLASTS
PROTOZOA
0.05
FIG. 6.-Maximum-likelihood tree including representatives from Nematomorpha, Gastrotricha, and Acantocephala. As in the distance and MP trees, the Catenulida is the most primitive bilaterian and protostomes cannot be considered monophyletic. The monophyletic Platyhelminthes (=Rhabditophora + Nemertodermatida; Ehlers 1985) are sister to a clade including Gastrotricha and Nematomorpha and both are buried within the protostomian phyla. Numbers at nodes are percentages of 100 bootstrap replicates that support the branch, only those over 50% being represented, with the exception of the sister group to Catenulida (45%). For species names, see tables 1 and 2.
lateria. Using complete 18s rDNA sequences and dif- ferent methods of phylogenetic analysis, we have stud- ied the monophyletism, phylogenetic position, and in- ternal phylogeny of one of the most likely candidates for sister group of all the other Bilateria, the Platyhel- minthes. The analyses show the Catenulida branching after the diploblasts, whereas the Rhabditophora, which form a monophyletic clade, branch early on in the evo- lution of the protostomates. In addition, all analyses give a similar internal phylogeny for the Rhabditophora, with the parasites as a monophyletic group branching unex- pectedly early within the evolution of the group.
The Acoela: Primitive or Derived Fast-Clock Organisms?
Our 18s analyses show Acoela as the first Bilateria to branch after the diploblasts, although with only mod- erate support (67% bootstrap) (fig. 3). An early diver- gence of acoel flatworms in triploblast evolution had previously been reported by Katayama et al. (1993) based on partial 18s rDNA sequences. Later, Katayama et al. (1995) used the acoel Convoluta naikaiensis with two dicyemids, one mixozoan, the nematode Caenor- habditis elegans, and other diploblast and triploblast or- ganisms to position the mesozoan dicyemids within the Metazoa. A clade made by the acoel, dicyemids, mix-
ozoans, and C. elegans was found to be the sister group to the rest of Bilateria. All the members of this clade, however, had very long branches, which may explain why species so diverse grouped together.
The NJ tree supporting the early branching of acoels (fig. 3) leads, however, to several inconsistencies. First, protostomes and deuterostomes cluster together but with low bootstrap (less than 50%), and protostomes appear paraphyletic, with arthropods and the single ne- matomorph included forming a sister group to the deu- terostomes. When acoels are not introduced, deuteros- tomes appear highly supported (94%, 81%, and 84% in NJ, MP and ML trees, respectively; figs. 4-6), whereas protostomes remain paraphyletic because aschelminthes (NJ trees; fig. 4) or Platyhehninthes (ML trees; fig. 6) appear buried within them. Second, the bulk of Platyh- elminthes appears only weakly supported (5 1% in fig. 3); instead, when acoels are not included, Platyhelmin- thes are very highly supported (figs. 4-6; see below). Finally, and most importantly, the lines leading to and separating the two acoels are extremely long, which may lead to artifactual grouping. Indeed, another long-branch organism, the acanthocephalan Moliniformis, is attracted close to acoels. Early branching of acoels may be ex- plained in two ways. Either they really are very primi-
The Phylogenetic Status of Platyhelminthes 493
. .
IO7 21-1 .I!1 nrlmbcr Of pilr.SinlOny illfOrllliltiYC sites
.
IO7 ?I-! 321 number of parsimony informative sites
(
B
t
: . . . . : . l
: : . : . . : * f . . ; I 6 : i
t ’ i . f t
. I : C kil.‘/i( *it ’
a l
4 I I . m-l
numhr of 321
informntivc 41x
sitcs
D
1
1 I07 21-1 321 number of parsimony informntive sites
FIG. 7.-The patterns of resolved nodes (PRN) for four nodes taken from the phylogenies of figures 4-6 concerning the clustering of Polycladida and Macrostomida. In each graph, the 200 bootstrap proportions (ordinate) obtained for a given number of nucleotides (10, 15, 25, 50, 75, 100, 150, 200, 250, and 300) are plotted vertically as a function of increased number of nucleotides (abscissa) (for further details, see Materials and Methods). The graphs correspond respectively to the monophyly of (A) Macrostomida (Microstomum and Mucrostomum), (B) Polycladida (Plunoceru and Discocelis) + Microsfomum, (C) Polycladida (Plunoceru and Discocelis) + Mucrostomum, and (D) Macrostomida (Mucrostomum and Microstomum) + Polycladida (Plunoceru and Discocelis).
tive triploblastic metazoans or, alternatively, they have or have had a higher rate of molecular substitutions (fast-clock organisms). Fast-clock organisms (e.g., Dro- sophila, Caenorhabditis elegans and most nematodes, Artemia salina, Aedes, among others) have previously been described and they have the common features of clustering together and of branching more deeply in trees than they should, introducing systematic errors when including them in a phylogeny (Olsen 1987).
Fast-clock organisms have been linked to short generation times or to rate variation among lineages (Golding 1996). Indeed, Drosophila and C. elegans have very short generation times. Data on reproductive rates
in acoels are scant, although they are known to have generation times of 3-4 weeks in optimal conditions (Faubel, personal communication). This is considerably shorter than most generation times reported for other Turbellaria (Heitkamp 1988) and gives support for such an effect in acoels. Whether their long branches stem from shorter generation times as predicted by the gen- eration time effect hypothesis or whether they are due to increased rates of change in their lineage is an open question. Acoels have been proposed in the past to be the most primitive Platyhelminthes. However, most fea- tures considered primitive in acoels are based on nega- tive evidence (absence of characters), though the

494 Carranza et al.
“brain” and the longitudinal nerve cords closely resem- ble the cnidarian and the ctenophore condition (Haz- sprunar 1996). Recent reappraisals, however, place acoels as a rather derived group (Ehlers 1985; Smith, Tyler, and Rieger 1986; Willmer 1990).
Are the Catenulida an Early Bilaterian Group?
In the past, Catenulida have been considered as an aberrant member of Platyhelminthes (Reisinger 1924) or to represent one of the first offshots in their evolution (Ax 1963; Karling 1974). Some (Sterrer and Rieger 1974) have raised the question of whether this group should even be classified within the Platyhelminthes. This is because the postulated synapomorphies between Catenulida and Rhabditophora: protonephridia, ciliary rootlet system, and mode of epidermal replacement (Eh- lers 1985) have not been proved to be homologous (Smith, Tyler, and Rieger 1986), and synapomorphies linking Catenulida to Acoelomorpha are not known. Synapomorphies of the Catenulida are the unpaired ex- cretory system, the special organization of the cyrtocyte, the dorsorostral position of the male copulatory organ, and aciliary spermatozoa (Karling 1974; Ehlers 1985). Other features are plesiomorphies, common to most Pla- tyhelminthes, and some (e.g., sparsely ciliated epi- dermis, monociliated epidermal sensory receptors, lack of frontal glands, and lack of rhabdites) are common to lower Eumetazoa.
With the exception of trees incorporating acoels (fig. 3), Catenulida is the first group branching after the diploblasts, although with low or moderate bootstrap support (45%, 7 1 %, and 45% for NJ, MP, and ML trees, respectively). Moreover, the promising PRN node found for Catenulida + Diploblasts + choanoflagellates (Pro- tozoa) is a good indicator of the basal position of this group. However, their basal position in all trees could be a consequence of a greater rate of evolutionary change for this group (similar to what happens with Acoela). In trees constructed using algorithms reflecting rate inequalities along branches such as those used here (fig. 4), one essentially carries out a relative-rate test on several species simultaneously, a rate difference being manifested by a difference in branch lengths (Philippe et al. 1994). Clearly at variance to what happened with acoels, the catenulid branch is similar in length to the rest of metazoans (only some platyhelminths have some- what longer branches). A relative-rate test (Wilson, Carl- son, and White 1977) was carried out, resulting in very similar distances for deuterostomes (2 1,8%), proto- stomes (21,4%), “Platyhelminthes” (23,5%), and Caten- ulida (Stenostomum) (20,7%) when calculated using the choanoflagellates as an outgroup. Similar results are ob- tained when the diploblasts are used as outgroup (18% for deuterostomes, 17.9% for protostomes, 19.2% for “Platyhelminthes” and 17.6% for the Catenulida with respect to the ctenophore). In all cases, only the “Pla- tyhehninthes” seem to have slightly higher rates.
When “lower” groups, reportedly considered basal to the triploblastic Bilateria (Winnepenninckx et al. 1995), such as phylum Gastrotricha, phylum Acantho- cephala, and phylum Nematomorpha (classically clas-
sified as phylum Aschelminthes) were included in the analysis, they fell within the protostomates (NJ tree, fig. 4) or formed the sister group of Rhabditophora (ML tree, fig. 6, for Gastrotricha and Acanthocephalan). In both cases, Catenulida branched earlier than any other Bilateria, including these presumptive lower groups. However, when acoels are introduced (NJ trees, fig. 3), acantocephalans and gastrotricha branch sequentially af- ter them, although this probably results from attraction among long-branch groups. Gastrotricha and Acantho- cephala were reported by Winnepenninckx et al. (1995) to form a weakly supported clade with the Platyhelmin- thes. Our analyses, using more species, support the re- lationship of rhabditophoran platyhelminthes with these aschelminth groups, calling for further studies.
Altogether, and despite the fact that only a single representative has been sequenced so far, our data sup- port the hypothesis that Catenulida constitute an inde- pendent clade that branched off early in the evolution of Bilateria and, hence, that Platyhelminthes probably are paraphyletic.
The Bulk of “Platyhelminthes”: the Monophyletic Rhabditophora
The bulk of “Platyhelminthes,” the so-called Rhabditophora sensu Ehlers (1985), appear in all anal- yses as a clear monophyletic group. Rhabditophora, which includes the parasitic classes (Neodermata), has a main autapomorphy, the presence of lamellated rhab- dites, and several synapomorphies such as the presence of duo-gland adhesive system and duo-cell weir and multiciliated terminal cell in the protonephridia. There is ample consensus in considering the Rhabditophora as monophyletic (Karling 1974; Ehlers 1985; Smith, Tyler, and Rieger 1986; Rohde 1990). This is supported here by very high or moderate bootstraps (99% and 91% for NJ and MP trees, respectively, and 65% for ML trees) and agrees with the morphological character-based phy- logenetic schemes of Karling and Ehlers (see fig. 2). 18s sequence data, however, show a major difference with them: the clustering of Nemertodermatida within Rhab- ditophora and not with Acoela forming the Acoelomor- pha sensu Ehlers (1985). Because only one species (Nemertinoides elongatus) has been analyzed here, the position of nemertodermatids should be left open.
Internally, NJ and ML trees reproduce two main monophyletic clades with high or moderate bootstrap values: Macrostomida + Polycladida and the rest of Rhabditophora plus Nemertodermatida. Macrostomida and Polycladida appear as close groups in Karling’s ( 1974) and Ehlers’ ( 1985) phylogenetic proposals (fig. 2) but do not cluster together. Smith, Tyler, and Rieger (1986) see Polycladida within a clade with the bulk of Rhabditophora, whereas Macrostomida form a sister group to the enigmatic Haplopharyngida. Finally, ac- cording to Rohde (1990), Macrostomida form a clade with Prolecithophora this being the sister group of Pro- seriata and Neodermata. This state of flux as regards Macrostomida may reflect the uncertainties of linking Macrostomidae and Microstomidae in a single order (Rieger, personal communication), indicating the need

to include more species in future analyses. Despite these difficulties, the high bootstraps found in NJ trees, the promising PRN nodes for Microstomidae + Macrostom- idae (fig. 7A) and Polycladida + Macrostomidae (fig. 7D), and the presence of homocellular gonads and en- dolecithal eggs in both groups are good indicators of close affinities between Polycladida and Macrostomida.
The sister group to Polycladida + Macrostomida reproduces the Neoophora (Westblad 1948; Ehlers 1985) with high or moderate bootstraps (78%, 86%, and 57% for NJ, MP and ML trees, respectively). The main syn- apomorphies of this group are the presence of hetero- cellular female gonads and the ectolecithal eggs. The internal phylogeny of Neoophora, however, is not well resolved. Rohde (1990) has noted that apomorphies for the Lecithoepitheliata, Prolecithophora, Proseriata, and Tricladida are insufficient to determine their position within the Platyhelminthes. The low bootstraps found here for most neoophoran branchings indicate that at the present level of knowledge, 18s rDNA sequences are unable to settle the issue. Even so, all trees reproduce some interesting regularities. First, leaving Nemertoder- matida aside, parasites appear as a clear monophyletic group basal to the rest of neoophorans (see below). Sec- ond, Tricladida often clusters with Rhabdocoela with low or moderate bootstraps (although PRN nodes are promising; data not shown), forming a highly derived group. And finally, Proseriata always show a rather basal position close to the parasites (Neodermata) and far from their presumed sister group, the Tiicladida. These results indicate that Seriata are a paraphyletic group and that Neodermata are a monophyletic basal group.
Parasitic platyhelminthes (Neodermata; Ehlers 1985) are most often considered either the sister group of the Dalyellioida, rhabdocoels with doliiform pharynx (Ehlers 1985; Brooks 1989), or an early branch of rhab- docoels which retained the less specialized rosulate pharynx (Kotikova and Joffe 1988). Based on the sim- ilarity between ultrastructure of the flame bulbs of Neo- dermata and those of most “primitive” proseriates, Roh- de (1990) considers them not related to rhabdocoels but as the sister group of the Proseriata. The NJ, MP and ML trees built here support the monophyly of Neoder- mata and its basal position within the evolution of neoo- phorans. Cestodes begin their life cycle as parasites of arthropods, the digenetic trematodes as parasites of mol- lusts. It is widely accepted that most invertebrate phyla were established as distinct and disparate groups by the Cambrian, as shown by the Ediacaran and Burgess Shale fauna (Conway Morris 1993; Valentine, Erwin, and Ja- blonski 1996), and there are sound reasons to believe that parasitism is as old as predation, symbiosis, and mimicry. Therefore, it is also sound to think that certain primitive-looking free-living “turbellarians” became parasites of invertebrates, probably molluscs (Rohde 1995), during Cambrian times. Later, when vertebrates appeared, they added a fish host and a final piscivorous host, giving the typical life cycle with two larval stages and three host species (Stunkard 1975). This plausible scenario perfectly fits the early branching of parasitic groups from primitive “turbellarians” found here and
The Phylogenetic Status of Platyhelminthes 495
argues against proposals on parasitism as a late devel- opment from advanced turbellarian rhabdocoels (Ehlers 1985; Brooks 1989).
Taxonomic and Phylogenetic Considerations
The general topology of the trees here obtained agrees to a large extent with previous proposed phylog- enies of the animal kingdom (Field et al. 1988; see the reanalysis of their data by Patterson 1989 and Lake 1990), followed by those of Christen et al. (1991), Adoutte and Philippe (1993), Chenuil(1993), and Wain- right et al. (1993). All show two main features: first, a clear split between diploblasts and triploblasts; second, a poor resolution for the major coelomate phyla, es- pecially for protostomes often giving big multifurcations that include annelids, molluscs, arthropods, nemertines, and other minor phyla. In addition, the positions of pseudocoelomates and acoelomates remain uncertain. Philippe, Chenuil, and Adoutte (1994) have provided convincing evidence that unresolved bushy multifurca- tions arise because the resolving power of the presently available complete 18s rDNA database is only about 40 Myr. There is now amply documented evidence, namely from fossil records, that the period of early and rapid origin and diversification of the major invertebrate phy- la, the so-called “Cambrian explosion” occurred in less than 40 Myr. In other words, multifurcations derived from molecular data have provided molecular corrobo- ration for the Cambrian “explosion.” In this context, the uncertain position of the Rhabditophora as regards pro- tostomates and deuterostomates (see figs. 3-6) may in- dicate that the bulk of “Platyhelminthes” appeared and evolved during this fast Cambrian radiation. In contrast, the early branching of Catenulida supports an acoelo- mate grade of organization for the first bilaterian, al- though the small number of species studied leaves the issue unresolved and calls for a deeper study.
As regards the hypothesis put forward on the origin of Bilateria (fig. lA--C) the phylogenetic derivations of our results could also be of interest. First, Platyhelmin- thes as a whole cannot be considered the sister group of the rest of Bilateria as they are paraphyletic. Second, the bulk of “Platyhelminthes,” represented by the mono- phyletic Rhabditophora, are more related, as could be expected, to prostostomes than to deuterostomes. This suggests that Rhabditophora are either protostomates or, most likely, an early branch on the lineage leading to the bulk of protostomates (fig. 1B). This rules out the hypothesis that sees “Platyhelminthes” (Rhabditophora) as the most primitive Bilateria (fig. IA) or as a derived group from coelomate ancestors (fig. lc). Third, all analyses and trees, with the exception of those incor- porating acoels, show the catenulid Stenostomum branching after the diploblasts and earlier than any tri- ploblast here studied, including potential primitive groups such as gastrotrichs, acanthocephalans, and ne- matomorphs (Winnepenninckx et al. 1995).
To summarize, we have shown that Platyhelminthes as a whole do not seem to be primitive. Although our results do not contradict the idea of Platyhelminthes be- ing monophyletic, the persistence of the node, albeit

496 Carranza et al.
with a low bootstrap, locating the Catenulida as the sis- ter group to the rest of the Bilateria, casts some doubts on its monophyly. Long-branching of acoels and the paucity of sampling in both acoels and catenulids calls for a large sampling of these primitive groups which hold promise to be the key to the evolution of the Bi- lateria.
Acknowledgments
We thank the following persons for help in locating and providing turbellarian material: M. Curini-Galleti, A. Falleni, J. Gamo, B. I. Joffe, D. T J. Littlewood, M. Litvaitis, C. Norena-Janssen, C. Novell, R. Ponce de Leon, M. Reuter, R. Rieger, E. Schockaert, J. Smith III, and S. Tyler. We also thank Gonzalo Giribet, Miguel Angel Arnedo, and Carles Ribera for helpful comments and discussions. We are grateful to one reviewer, whose constructive criticism and comments helped to refine this manuscript. This study was supported by a DGI- CYT grant PB90-0477 and a CIRIT grant GRQ93-1044 to J.B.
LITERATURE CITED
ADOUTTE, A., and H. PHILIPPE. 1993. The major lines of meta- zoan evolution: summary of traditional evidence and les- sons from ribosomal RNA sequence analysis. Pp. l-30 in Y. PICHON, ed. Comparative molecular neurobiology. Birk- hauser Verlag, Basel.
Ax, l? 1963. Relationships and phylogeny of the Turbellaria. Pp. 1-68 in E. C. DOUGHERTY, ed. The lower metazoa. University of California Press, Berkeley-Los Angeles, Calif.
-. 1987. The phylogenetic system. John Wiley & Sons, Chichester.
BAVERSTOCK, l? R., R. FIELKE, A. M. JOHNSON, R. A. BRAY, and I. BEVERIDGE. 1991. Conflicting phylogenetic hypoth- esis for the parasitic Platyhelminthes tested by partial se- quencing of 18s ribosomal RNA. Int. J. Parasitol. 21:329- 339.
BLAIR, D. 1993. The phylogenetic position of the Aspidoboth- ria within the parasite flatworms inferred from ribosomal RNA sequence data. Int. J. Parasitol. 23:169-178.
BRIDGE, D., C. W. CUNNINGHAM, R. DESALLE, and L. W. Buss. 1995. Class-level relationships in the phylum Cnidaria: mo- lecular and morphological evidence. Mol. Biol. Evol. 12: 679-685.
BROOKS, D. R. 1989. A summary of the database pertaining to the phylogeny of the major groups of parasitic platyhelmin- thes, with a revised classification. Can. J. Zool. 67:714-720.
BRUSCA, R. C., and G. J. BRUSCA. 1990. Invertebrates. Sinauer, Sunderland, Mass.
CAMIN, J. H., and R. R. SOKAL. 1965. A method for deducing branching sequences in phylogeny. Evolution 19:3 11-326.
CARRANZA, S., G. GIRIBET, C. RIBERA, J. BAGUIQA, and M. RIUTORT. 1996. Evidence that two types of 18s rDNA co- exist in the genome of Dugesia (Schmidtea) mediterrunea (Patyhelminthes, Turbellaria, Tricladida). Mol. Biol. Evol. 13:824-832.
CHENUIL, A. 1993. Etude des relations de parent6 entre les principaux groupes d’invertebres protostomiens par ampli- fication, sequencage et comparaison de portions du gene de I’ARN 28s. Ph.D. thesis, Universite Sciences et Techniques du Languedoc., Montpellier, France.
CHIRGWIN, J. M., A. E. PRZBYLA, R. J. MACDONALD, and W. RUTTER. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294-5299.
CHRISTEN, R., A. RATTO, A. BAROIN, R. PERASSO, K. G. GRELL, and A. ADOU-ITE. 1991. An analysis of the origin of metazoans, using comparison of partial sequences of the 28s rRNA, reveals an early emergence of triploblasts. EMBO J. 10:499-503.
CONWAY MORRIS, S. 1993. The fossil record and the early evolution of the Metazoa. Nature 361:219-225.
DOOLITTLE, R. E, D. E FENG, S. TSANG, G. CHO, and E. LIT- TLE. 1996. Determining divergence times of the major king- doms of living organisms with a protein clock. Science 271: 470-477.
EHLERS, U. 1985. Das Phylogenetische System der Plathehnin- thes. G. Fischer, Stuttgart, Germany.
~ 1986. Comments on a phylogenetic system of the pla- tyhehninthes. Hydrobiologia 132: 1-12.
FIELD, K. G., G. J. OLSEN, D. J. LANE, S. J. GIOVANNONI, M. T. GHISELIN, E. C. RAFF, N. R. PACE, and R. R. RAFF. 1988. Molecular phylogeny of the animal kingdom. Science 239: 748-753.
FELSENSTEIN, J. 1981. Evolutionary trees from DNA sequenc- es: a maximum likelihood approach. J. Mol. Evol. 17:368- 376.
- 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783-79 1.
-. 1993. PHYLIP (phylogeny inference package). Ver- sion 3.5~. University of Washington, Seattle.
FITCH, W. M., and E. MARGOLIASH. 1967. Construction of phy- logenetic trees. Science 155:279-284.
FRIEDRICH, M., and D. TAUTZ. 1995. Ribosomal DNA phylog- eny of the major extant Arthropod classes and the evolution of myriapods. Nature 376: 165-167.
GARCIA-FERNANDEZ, J., J. BAGuRIA, and E. SALK). 1993. Ge- nome organization and expression of the planarian hom- eobox genes Dth-1 and Dth-2. Development 118:241-253.
GOLDING, B. 1996. When was life’s first branch point? Cur-r. Biol. 6(6):679-682.
GUTELL, R. R., B. WEIBSER, C. R. WOESE, and H. E NOLLER. 1985. Comparative anatomy of 16S-like ribosomal RNA. Prog. Nucleic Acid Res. Mol. Biol. 32: 155-216.
HASZPRUNAR, G. 1996. The Mollusca: coelomate turbellarians or mesenchymate annelids? Pp. l-28 in J. TAYLOR, ed. Or- igin and evolutionary radiations of the Mollusca. Oxford University Press, Oxford.
HEITKAMP, U. 1988. Life-cycles of microturbellarians of pools and their strategies of adaptation to their habitats. Fortschr. Zool. 36:449-456.
HYMAN, L. H. 1951. The invertebrates. Vol. 2. Platyhelminthes and Rhynchocoela: the Acoelomate Bilateria. McGraw Hill, New York, N.Y.
KARLING, T. G. 1974. On the anatomy and affinities of the turbellarian orders. Pp. 1-16 in N. W. RISER and M. I? MORSE, eds. Biology of the Turbellaria. McGraw Hill, New York, N.Y.
KATAYAMA, T., H. WADA, H. FURUYA, N. SATOH, and M. YA- MAMOTO. 1995. Phylogenetic position of the dicyemid Me- sozoa inferred from 18s rDNA sequences. Biol. Bull. 189: 8 l-90.
KATAYAMA, T, M. YAMAMOTO, H. WADA, and N. SATOH. 1993. Phylogenetic position of acoel turbellarians inferred from partial 18s rDNA sequences. Zool. Sci. 10:529-536.
KIMURA, M. 1980. A simple method for estimating evolution- ary rates of base substitution through comparative studies of nucleotide sequences. J. Mol. Evol. 16: 11 l-120.

The Phylogenetic Status of Platyhelminthes 497
KOTIKOVA, E. A., and B. I. JOFFE. 1988. On the nervous sys- tem of the dalyellioid turbellarians. Forts&r. Zool. 36: 191- 194.
LAKE, J. A. 1990. Origin of the metazoa. Proc. Natl. Acad. Sci. USA 87:763-766.
LECOINTRE, G., H. PHILIPPE, H. L. V LB, and H. LE GUYADER. 1994. How many nucleotides are required to resolve a phy- logenetic problem? The use of a new statistical method ap- plicable to variable sequences. Mol. Phylogenet. Evol. 3: 292-309.
OLSEN, G. J. 1987. Earliest phylogenetic branchings: compar- ing rRNA-based evolutionary trees inferred with various techniques. Cold Spring Harb. Symp. Quant. Biol. LII:825- 837.
OLSEN, G. J., H. MATSUDA, R. HANGSTROM, and R. OVERBEEK. 1994. FastDNAml: a tool for construction of phylogenetic trees of DNA sequences using maximum likelihood. Com- put. Appl. Biosci. 10:41-48.
PATTERSON, C. 1989. Phylogenetic relations of major groups: conclusions and prospects. Pp. 471-478 in B. FERNHOLM, K. BREMER, and H. J~RNVALL, eds. Hierarchy of life. Mol- ecules and morphology in phylogenetic analysis. Excerpta Medica, Amsterdam.
PHILIPPE, H. 1993. MUST, a computer package of management utilities for sequences and trees. Nucleic Acids Res. 21: 5264-5272.
PHILIPPE, H., A. CHENUIL, and A. ADOUT~E. 1994. Can the Cambrian explosion be inferred through molecular phylog- eny? Development (Suppl.): 15-25.
PHILIPPE, H., U. S~RHANNUS, A. BAROIN, R. PERASSO, E GAS- SE, and A. ADOUTTE. 1994. Comparison of molecular and paleontological data in diatoms suggest a major gap in the fossil record. J. Evol. Biol. 7:247-265.
REISINGER, E. 1924. Die Gattung Rhynchoscolex. Z. Morphol. Gkol. Tiere l:l-37.
I&MANE, A., V. STORCH, and U. WELSCH. 1980. Systematische Zoologie. G. Fischer, Stuttgart, Germany.
RIEGER, R. 1985. The phylogenetic status of the acoelomate organisation within the Bilateria: a histological perspective. Pp. 101-122 in S. CONWAY MORRIS, J. D. GEORGE, R. GIB- SON, and H. M. PLATT, eds. The origins and relationships of lower invertebrates. Oxford University Press, Oxford.
RIUTORT, M., K. G. FIELD, R. R. RAFF, and J. BAGU&& 1993. 18s rRNA sequences and phylogeny of Platyhelminthes. Biochem. Syst. Ecol. 21:7 l-77.
RIUTORT, M., K. G. FIELD, J. M. TURBEVILLE, R. R. RAFF, and J. BAGU&& 1992. Enzyme electrophoresis, 18s t-RNA se- quences, and levels of phylogenetic resolution among sev- eral species of freshwater planarians (Platyhehninthes, Tri- cladida, Paludicola). Can. J. Zool. 70: 1425-1429.
ROHDE, K. 1990. Phylogeny of Platyhehninthes, with special reference to parasitic groups. Int. J. Parasitol. 20:979-1007.
- 1995. The origins of parasitism in the Platyhelminthes. Int. J. Parasitol. 24: 1099-l 115.
ROHDE, K., C. HEFFORD, J. T. ELLIS, I? R. BAVERSTOCK, A. M. JOHNSON, N. A. WATSON, and S. DITTMANN. 1993. Con- tributions to the phylogeny of Platyhehninthes based on partial sequencing of 18s ribosomal DNA. Int. J. Parasitol. 23:705-724.
ROHDE, K., A. M. JOHNSON, F? R. BAVERSTOCK, and N. A. WATSON. 1995. Aspects of the phylogeny of Platyhelmin- thes based on 18s ribosomal DNA and protonephridial ul- trastructure. Hydrobiologia 305:27-35.
SAIKI, R. K., S. J. SCHARF, E FALOONA, K. B. MULLIS, G. T. HORN, H. A. ERLICH, and N. ARNHEIM. 1985. Enzymatic
amplification of P-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anaemia. Science 233: 1076-1078.
SAITOU, N., and M. NEI. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406-425.
SALVINI-FLAWEN, L. VON. 1978. On the origin and evolution of the lower Metazoa. Z. Zool. Syst. Evolutionsforsch. 16: 40-88.
&EWING, R. 1980. Das Archicoelomatenkonzept. Zool. Jahrb. Syst. 103:439-482.
SMITH, J. I? S., S. TYLER, and R. R. RIEGER. 1986. Is the Turbellaria polyphyletic? Hydrobiologia 132: 13-2 1.
SMOTHERS, J. E, C. D. VON DOHLEN, L. H. SMITH JR., and R. D. SPALL. 1994. Molecular evidence that the myxozoan pro- tists are Metazoans. Science 265: 17 19-1721.
SOGIN, M. L. 1991. Early evolution and the origin of eukary- otes. Curr. Opin. Genet. Dev. 1:451-456.
STERRER, W., and R. RIEGER. 1974. Retronectidae-a new cos- mopolitan marine family of Catenulida (Turbellaria). Pp. 63-92 in N. W. RISER and M. I? MORSE, eds. Biology of the Turbellaria. McGraw Hill, New York, N.Y.
STUNKARD, H. W. 1975. Life-histories and systematics of par- asitic flatworms. Syst. Zool. 24:378-385.
SWOFFORD, D. L. 1993. PAUP: phylogenetic analysis using parsimony. Version 3.1.1. Illinois Natural History Survey, Champaign.
TURBEVILLE, J. M., K. G. FIELD, and R. R. RAFF. 1992. Phy- logenetic position of the phylum Nemertini, inferred from 18s rRNA sequences: molecular data as a test of morpho- logical character homology. Mol. Biol. Evol. 9:235-249.
TURBEVILLE, J. M., D. M. PFEIFER, K. G. FIELD, and R. R. RAFF. 1991. The phylogenetic status of Arthropods, as in- ferred from 18s rRNA sequences. Mol. Biol. Evol. 8:669- 686.
VALENTINE, J. W., D. H. ERWIN, and D. JABLONSKI. 1996. De- velopmental evolution of metazoan bodyplans: the fossil ev- idence. Dev. Biol. 173:373-38 1.
WADA, H., and N. SATOH. 1994. Details of the evolutionary history from invertebrates to vertebrates, as deduced from the sequences of 18s rRNA. Proc. Natl. Acad. Sci. USA 91:1801-1804.
WAINRIGHT, I? O., G. HINKLE, M. L. SOGIN, and S. K. STICKEL. 1993. Monophyletic origins of the metazoa: an evolutionary link with fungi. Science 260:340-342.
WESTBLAD, E. 1948. Studien iiber Skandinavische Turbellaria Acoela V. Ark. Zool. 41A:l-82.
WILLMER, I? 1990. Invertebrate relationships. Cambridge Uni- versity Press, Cambridge.
WILSON, A. C., S. S. CARLSON, and T J. WHITE. 1977. Bio- chemical evolution. Annu. Rev. Biochem. 46:573-639.
WINNEPENNINCKX, B., T. BACKELJAU, and R. DE WACHTER. 1995. Phylogeny of protostome worms derived from 18s rRNA sequences. Mol. Biol. Evol. 12641-649.
WINNEPENNINCKX, B., T BACKELJAU, L. Y. MACKEY, J. M. BROOKS, R. DE WACHTER, S. KUMAR, and J. R. GAREY. 1995. 18s rRNA data indicate that Aschelminthes are poly- phyletic in origin and consists of at least three distinct clades. Mol. Biol. Evol. 12: 1132-l 137.
WOESE, C. R. 1987. Bacterial evolution. Microbial. Rev. 51: 221-271.
MANOLO GOUY, reviewing editor
Accepted January 24, 1997