apostolepis longicaudata (serpentes: xenodontinae) with comments … · redescription of...
TRANSCRIPT

REDESCRIPTION OF APOSTOLEPIS LONGICAUDATA (SERPENTES:XENODONTINAE) WITH COMMENTS ON ITS HEMIPENIAL
MORPHOLOGY AND NATURAL HISTORY
FELIPE FRANCO CURCIO1,3, PEDRO M. SALES NUNES
1, MICHAEL B. HARVEY2, AND
MIGUEL T. RODRIGUES1
1Departamento de Zoologia, Instituto de Biociencias, Universidade de Sao Paulo, Caixa Postal 11.461 CEP: 05422-970,Sao Paulo, Brazil
2Department of Biological Sciences, Broward College, 3501 S.W. Davie Road, Davie, FL 33314, USA
ABSTRACT: We redescribe the rare elapomorphine snake Apostolepis longicaudata Gomes in Amaral onthe basis of a detailed examination of the holotype and comparisons with new specimens from the Cerrado ofCentral Brazil. The new data include information on coloration in life, hemipenial morphology, and naturalhistory. Two specimens identified as A. quinquelineata by previous authors are now attributed to A.nelsonjorgei, which seems to be the only species in the genus with subcaudal counts overlapping those of A.longicaudata.
RESUMO: Apresentamos a redescricao da rara especie de serpente Apostolepis longicaudata Gomes inAmaral com base no exame do holotipo e em comparacoes com tres novos exemplares do Cerrado do BrasilCentral. O trabalho inclui dados ineditos sobre a coloracao em vida, morfologia hemipeniana e historianatural. Dois especimes lineados previamente identificados como A. quinquelineata por outros autores saoredeterminados aqui como A. nelsonjorgei, que parece ser a unica especie do genero cujas contagens desubcaudais se sobrepoem as de A. longicaudata.
Key words: Apostolepis nelsonjorgei; Apostolepis nigrolineata; Apostolepis quinquelineata; Cerrado;Elapomorphini; Snakes; Taxonomy
THE SNAKE genus Apostolepis includes morethan 30 small to medium-size fossorial spe-cies widely occurring in cis-Andean SouthAmerica (Ferrarezzi et al., 2005; Harvey,1999; Lema, 2001). Along with the generaElapomorphus and Phalotris, it is allocated tothe tribe Elapomorphini and is characterizedby lack of distinct internasals (possibly fusedto their respective frontal shields) and a blackband on the tip of the tail (Boulenger, 1896;Ferrarezzi, 1994; Harvey, 1999; Zaher et al.,2009).
Because of their cryptozoic nature mostlyassociated with fossoriality, several species ofApostolepis are rare and known from fewspecimens (Harvey, 1999). Small sample sizeshinder assessment of geographic variationsand make it difficult to diagnose species in thisgenus.
Apostolepis longicaudata is an example ofsuch a rare species. Its brief description (Gomesin Amaral, 1921a) was solely based on theholotype and included data on scalation and
coloration, complemented by illustrations ofthe head, fore trunk, and ventral surface of thetail (Fig. 1). As suggested by the specific epithet,the high number of subcaudals is the mostremarkable diagnostic feature of this specieswhen compared with the lower counts of mostcongeners.
After the original description, A. longicau-data has only been mentioned in passing (e.g.,Lema, 2001; Peters and Orejas-Miranda, 1970)and, besides the holotype, no new specimenswere reported. During surveys preceding con-struction of a railroad in the Rio TocantinsDepression, municipality of Guaraı, state ofTocantins, we obtained three specimens ofA. longicaudata. On the basis of these newspecimens, as well as on direct examinationof the holotype, we provide a redescriptioncontaining data on hemipenial morphology, dis-tribution, and natural history. In the contextof this study, we also discuss some confusinginformation from the literature regarding otherstriped Apostolepis, as well as the taxonom-ic significance of two specimens of uncertainidentity that are possibly related to A. long-icaudata.3 CORRESPONDENCE: e-mail, [email protected]
Herpetologica, 67(3), 2011, 318–331
E 2011 by The Herpetologists’ League, Inc.
318

MATERIALS AND METHODS
Specimens examined and data on localitiesare listed in Appendix I. Institutional acro-nyms are IBSP (Instituto Butantan, Sao Paulo),CHUNB (Colecao Herpetologica da Universi-dade de Brasılia, Distrito Federal), MPEG(Museu Paraense Emılio Goeldi, Belem, Para),and MZUSP (Museu de Zoologia da Universi-dade de Sao Paulo, Sao Paulo). The collec-tion permit was provided by the InstitutoNacional do Meio Ambiente e Recursos NaturaisRenovaveis (193/2001-DIFAS; Processo IBAMAno 02001.003451/93-13).
To the nearest 1 mm, we measured snout–vent length (SVL) and tail length with agraduated ruler. To the nearest 0.01 mm witha digital caliper, we measured head length(from the rostral to the quadrate-mandibulararticulation), head width (at widest point),eye–nostril distance (on the right side of thehead), mid-body diameter, mid-tail diameter,and eye diameter (measured only on the rightside of head in all specimens, except that wemeasured both eyes on the holotype of A.longicaudata). Nomenclature of scalation andcolor features are those of Harvey (1999). Ven-tral scale counts were performed using the
wider-than-long system (Peters, 1964). Toothcounts were taken on the right maxillary boneonly. The diagnosis of A. longicaudata was re-constructed according to the standardized num-bered system introduced by Harvey (1999).
The hemipenis of MZUSP 12694 was pre-pared following procedures described byManzani and Abe (1988), subsequently mod-ified by Pesantes (1994) and Zaher (1999).Hemipenial calcareous structures were stainedin an alcoholic solution of alizarin red (adaptedfrom Harvey and Embert, 2008; Harvey et al.,2008; Uzzell, 1973). Nomenclature regardinghemipenial morphology follows Dowling andSavage (1960) and Zaher (1999).
To test for sexual dimorphism in numbersof subcaudals, ventrals, and relative tail lengths,we tested for normality (Shapiro–Wilk test) andhomogeneity of variance (Levene’s test). Sinceour data for the first two characters did not sat-isfy these assumptions, we used Welch’s non-parametric t-test to compare means for thesexes. For relative tail lengths, we used analysisof covariance, treating SVL as a covariate. AnF-test for equality of the regression slopes wasused to test the assumption of parallelism. Weused the PAST (Hammer et al., 2001) softwareprogram for these statistical analyses.
FIG. 1.—Original illustration of the holotype of Apostolepis longicaudata (modified from Amaral, 1921a).
September 2011] HERPETOLOGICA 319

Apostolepis longicaudata Gomes in Amaral(Figs. 2–4)
Apostolepis longicaudata: Gomes in Amaral,1921a. Santa Filomena, Piauı, Brasil.
Holotype.—IBSP 1684 (not labeled in field,destroyed by fire in 2010), an adult male fromEngenheiro Dodt, municipality o Santa Filo-mena, state of Piauı, northeastern Brazil,collected by Francisco de Assis I[n]glesias be-tween 1916 and 1918 (the ‘‘n’’ in the collector’ssurname is a typographic error; the correctspelling is ‘‘Iglesias’’). Originally described in1919, by J. Florencio Gomes, formally publishedby Amaral (1921a).
Referred specimens.—MZUSP 12692–12694(field numbers MRT 7119, 7118, and 7531 re-spectively) collected 25–28 October 2001, in the‘‘Fazenda Marupiara,’’ municipality of Guaraı,state of Tocantins, Central Brazil, by D. Pavanand F. F. Curcio.
Diagnosis.—A small species of Apostolepis,SVL up to 254 mm, differing from all con-geners by the following combination of char-acters: (1) snout rounded or slightly acu-minate, length of rostral visible from aboveslightly more than one-third its distance tofrontal; (2) six supralabials; (3) preocular–nasal contact present (i.e., nasal and preocu-lar not separated by prefrontal); (4) temporalsabsent (0+0), fifth and sixth supralabials incontact with parietals; (5) four infralabialscontacting first pair of chin shields on eachside of head; (6) ventrals 234–244 and sub-caudals 49–52 in males (females unknown);(7) dorsal head plates uniformly dark brown,except for small irregular white blotches onanteriomedial surface of prefrontals; (8) lightsupralabial blotch large, covering posteriormargin of third and almost whole area offourth supralabial; (9) white and black nuchalcollars completely absent; (10) five dorsalstripes similar in width (as wide as, or slightlywider than, one dorsal scale row); (11) dorsum
FIG. 2.—Dorasl (A), lateral (B), and ventral (C) views ofthe head of the holotype of Apostolepis longicaudata(IBSP 1684). Scale bar: 2.0 mm.
FIG. 3.—Apostolepis longicaudata from Guaraı, Tocantins (MZUSP 12693), showing general dorsal coloration anddetail of head in life (Photo: Otavio A. V. Marques).
320 HERPETOLOGICA [Vol. 67, No. 3
background light tan (orange-tan in life); (12)terminal scale directed backward, mostly white,possibly invaded by black pigment from thetail band at least on its dorsal surface; (13)chin and gular region immaculate cream.
Species with five dorsal stripes that are mostlikely to be confused with A. longicaudataare the predominantly Amazonian species A.nigrolineata and A. quinquelineata and theCerrado species A. nelsonjorgei, but over-lapping values of subcaudal counts are onlypresent in A. nelsonjorgei. Nonetheless, A.longicaudata is unambiguously distinguishablefrom these three species by a suite of char-acters: (1) condition of the white nuchal collar:A well-developed white nuchal collar is pres-ent in A. nelsonjorgei (Lema and Renner,2004a) and entirely lacking in A. longicaudata.Most specimens of A. nigrolineata have rudi-ments of a white nuchal collar, peculiarly ex-pressed as two well-defined pale blotches oneach side of the nape (Lema and Renner,1998:107,118). However, a few specimens havecomplete white collars (e.g., MPEG 16137,17671). Lema and Renner (1998) suggest thatjuveniles of A. quinquelineata may have rudi-ments of a white nuchal collar. In our com-parative material, none of the specimens of thisspecies can be considered a juvenile, but no
traces of such a structure were present even inthe smaller individual (e.g., MPEG 17817; SVL5 226 mm, tail length 5 15 mm). (2) Conditionof the black nuchal collar: A distinctive blacknuchal collar up to two dorsal scales long ispresent in A. quinquelineata (Ferrarezzi, 1993;Harvey, 1999; Lema and Renner, 1998) andabsent in A. longicaudata. (3) Temporal for-mulae: Usually 0+1, rarely 0+0 in A. nigroli-neata, (Ferrarezzi, 1993; Harvey, 1999; Lemaand Renner, 1998), contrasting with 0+0 in allknown specimens of A. longicaudata. Boulen-ger’s (1896) original description mentions notemporals in the holotype of A. quinquelineata(fifth and sixth supralabials contact parietals),but this character is apparently variable in thisspecies, because there are specimens with 0+1temporals. Thus, we do not recognize thischaracter as informative for distinguishing A.quinquelineata from A. longicaudata. (4) Blackcaudal band: In A. quinquelineata the blackcaudal band is restricted to the dorsal and dor-solateral surface of the tail (Ferrarezzi, 1993;Lema and Renner, 1998), contrasting with thecomplete black band of A. longicaudata. (5)Width of the lateral stripe: The lateral stripes ofA. quinquelineata (covering first–fourth dorsalrows) are distinctively wider than the paraver-tebral and vertebral ones (Lema and Renner,
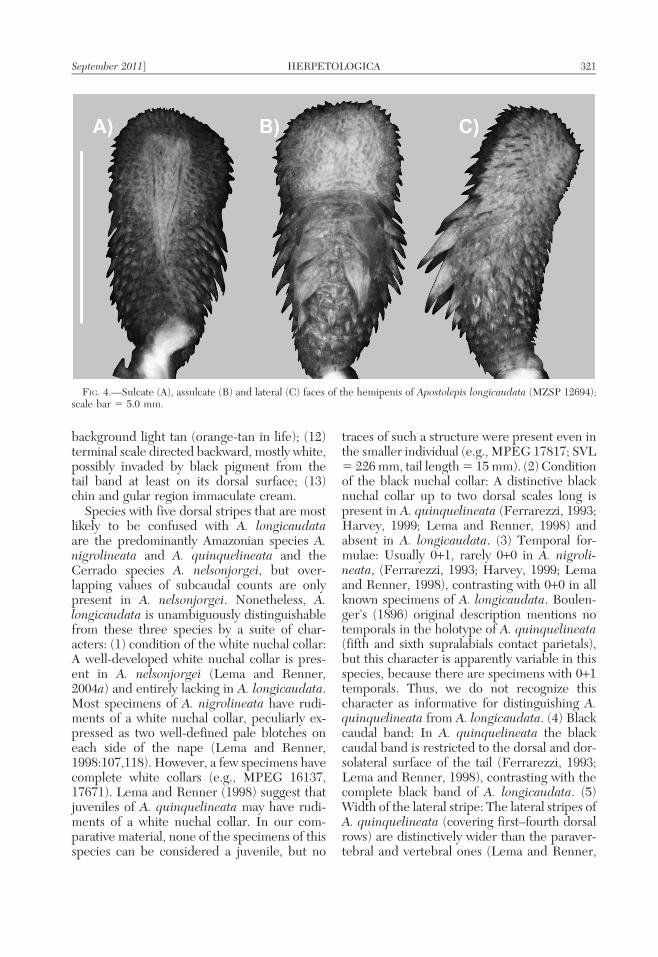
FIG. 4.—Sulcate (A), assulcate (B) and lateral (C) faces of the hemipenis of Apostolepis longicaudata (MZSP 12694);scale bar 5 5.0 mm.
September 2011] HERPETOLOGICA 321

1998), contrasting with a lateral stripe (restrict-ed to fourth dorsal row) similar in width to theparavertebral and vertebral ones in A. long-icaudata.
Five dorsal stripes are also present in Apo-stolepis arenaria, A. christineae, A. intermedia,A. lineata, A. serrana, A. striata, A. phillipsiand A. vittata (Harvey, 1999; Lema, 2002, 2004;Lema and Renner, 2004c, 2006; Rodrigues,1993), but all these taxa have considerablylower subcaudal counts and differ from A.longicaudata (characters in parentheses) in sca-lation and coloration. Apostolepis arenaria has21–31 subcaudals and distinctive white andblack nuchal collars (lacking). Furthermore,this species has 168–181 ventrals, the lowestcounts in the genus (Rodrigues, 1993). Aposto-lepis christineae has up to 29 subcaudals, anarrow black nuchal collar attaining the gularregion (lacking), a strongly projecting, flattened,and acuminate snout (rounded or slightly acu-minate), the rostral largely visible from aboveand mostly white on its dorsal surface (a smallportion visible from above and mostly blackon its dorsal surface), all stripes dark and cohe-sive (at least the paravertebral stripe diffuse), ablack caudal band restricted to the dorsum,barely reaching the lateral margins of the sub-caudals (complete caudal band), and the whiteportion of the terminal scale restricted to itsventral surface (white tip and ventral surface ofthe termimal scale; Lema, 2002). Apostolepisintermedia is only known from the holotype,which is apparently lost (Harvey, 1999; Lema,1993: 37). However, the original descriptionmentions three infralabials contacting the firstpair of chin shields (four), a pointed snout(rounded or slightly pointed), 217 ventrals(234–244), and 37 subcaudals (Koslowsky, 1898).Regarding A. lineata, the only objective dataallowing distinction from A. longicaudata areprovided by Harvey (1999: 393), who reported24 subcaudals, and Lema and Renner (2004b),who mention a distinctly projecting snout (notprojecting). Apostolepis serrana has up to 33subcaudals, besides a distinctive black nuchalcollar attaining the margins of the gular region(lacking) and an entirely white termimal scale(marked by black on the dorsal and lateralsurfaces). Apostolepis striata has considerableposterior—up to three dorsal scales—and ventralextensions of the head cap, configuring a black
nuchal collar that is lacking in A. longicaudata; thelabial blotch covers a larger area in A. striata—third–sixth supralabials—than in A. longicaudata(thirdandfourthsupralabials),andtheonlyknownspecimen has 202 ventrals (234–244) and 26subcaudals (Lema, 2004). Apostolepis philli-psi has narrow but distinctive white and blacknuchal collars (lacking), a pointed snout, and 24subcaudals (Harvey, 1999). Apostolepis vittatahas up to 28 subcaudals, a pointed and stronglyprojecting snout, five supralabials (six), and onlythree infralabials contacting the first pair of chinshields (four; Harvey, 1999).
Redescription of holotype.—A slender Apo-stolepis (mid-body diameter/SVL 5 0.012),small in size; tail relatively long, 15.9% of totallength; tail cylindrical, its diameter roughlyuniform throughout its length, slightly smallerthan mid-body diameter (mid-tail/mid-bodydiameter 5 0.79); head subcylindrical, longerthan wide (head width/head length 5 0.58),not distinct from neck; eyes small, eye diam-eter approximately one-tenth of head lengthand slightly shorter than distance from its an-terior margin to nostril (eye diameter/eye–nostril distance 5 0.75); pupil subelliptical;rostral rounded, not projecting over lowerjaw, length of its portion visible from aboveslightly more than one-third its distance toanterior border of frontal; internasals absent;prefrontals paired, longer than wide, theirlargest width two-thirds of largest length; fron-tal roughly hexagonal, longer than wide, itslargest width three-quarters of largest length,anterior border contacting posteromedial bor-ders of prefrontals, lateral borders contactingmedial margins of supraoculars and lateropos-terior borders contacting anteromedial bordersof parietals; supraoculars large, roughly trape-zoidal, slightly longer than wide, contactinglateroposterior border of prefrontal and upperposterior border of preocular anteriorly, lateralborder of frontal medially, upper border ofpostocular lateroposteriorly, and anterior bor-der of parietal posteriorly; parietals paired,longer than wide, largest width of each ap-proximately two-thirds the length of their mu-tual suture, anterior margin contacting poste-rior margin of supraocular and upper posteriormargin of postocular, anteromedial marginscontacting lateroposterior borders of frontal,lateral margins contacting upper borders of
322 HERPETOLOGICA [Vol. 67, No. 3

fifth and sixth supralabials; two enlarged occip-itals, one on each flank of nape, clearly dis-tinguishable in size and shape from dorsals;occipitals roughly rectangular, obliquely ori-ented, twice as long as wide, largely separatedby three interoccipitals (dorsal scales of verte-bral and both paravertebral rows); occipitalsanteriorly contacting lateroposterior margins ofparietals and upper posterior margin of sixthsupralabial; nasal entire, roughly triangular,posterior vertex broadly contacting preocular;nostril positioned in anterior third of nasal,partly visible from above and slightly directedanteriorly; preocular roughly pentagonal, inwide contact with nasal and forming an or-thogonal directed suture with it; postocularroughly ‘‘C’’ shaped, with feeble posterior ver-tex; postocular contacting third and fourth su-pralabials ventrally, fifth supralabial posterior-ly, parietal dorsoposteriorly, and supraoculardorsally; temporals 0+0; six supralabials, sec-ond and third contacting orbit, sixth largest,approximately as high as fifth; fifth and sixthcontacting parietals; mental subtriangular, twiceas wide as long; two pairs of chin shields, firstpair distinctively longer; seven infralabials, firstpair in contact immediately behind mental,widely separating it from chin shields; first–fourth infralabials contacting first pair of chinshields, four and five contacting second pair;fifth infralabial largest, roughly trapezoidal; 15dorsal scale rows without reductions near vent;anal plate divided; 241 ventrals; 52/52 subcau-dals; trunk broken at level of 84th ventral shield;terminal scale acuminate, directed ventrally;maxillary teeth 5+2, fourth tooth missing onright maxillary bone; head light brown; browncap covering whole dorsal and lateral surfaces ofhead, sporadically extending posteriorly overone or two dorsal scales and downward to upperhalf of second dorsal scale row, slightly belowlevel of seventh infralabial; snout mostly brown,with small irregular light spots on anteromedialsurface of prefrontals; two pale supralabialblotches; first supralabial blotch covering almostwhole area of first supralabial; second suprala-bial blotch covering posterior margin of thirdand almost whole area of fourth supralabial;white and black nuchal collars absent; chin andgular region immaculate cream, first–sixthinfralabials also immaculate; up to two-thirdsof seventh infralabial marked by brown cap;
mental immaculate; dorsal ground color lighttan, with five brown stripes similar in width (onevertebral, one paravertebral pair, and onelateral pair); all but vertebral stripe extendingto posterior margin of brown cap; vertebralstripe separated from brown cap by almostwhole length of one dorsal scale; lateral stripeslight brown, mostly cohesive, more distinctive infirst third of trunk, covering central area offourth row; paravertebral stripes light brown,diffuse, distinctively lighter than lateral stripes,covering upper margin of fourth, virtually wholearea of fifth and lower margin of sixth rows;vertebral stripe cohesive, dark brown, coveringcentral area of eighth (vertebral) row, alsomarking upper vertexes of contiguous (seventh)rows; ventrals and subcaudals immaculatecream; terminal caudal band dark brown, 10/11 subcaudals in length; terminal scale almostentirely white, with dark brown pigmentextending slightly above its dorsal surface.
Measurements of the holotype.—SVL 227 mm,tail length 43 mm, head length 4.90 mm, headwidth 2.85 mm, mid-body diameter 2.81 mm,mid-tail diameter 2.22 mm, eye-diameter 0.54/0.57 mm, eye–nostril distance 0.80 mm (onlyright side measured).
Variation and coloration in life.—Descrip-tive statistics on scale counts and measure-ments for the four known specimens of A.longicaudata are shown in Table 1. Thelargest specimen is a male (MZUSP 12692;SVL 5 254 mm, tail length 5 43); ventral andsubcaudal counts range respectively from 235to 244 and 49 to 52; no females are known.The snouts of MZUSP 12692–12694 areslightly pointed (Fig. 5A) in comparison withthe holotype (Fig. 2A). Head scalation showsno variation in the four specimens. Colorationof the holotype has considerably faded be-cause of preservation, but MZUSP 12692–12694 show minor differences between theircoloration in life (Fig. 3) and after preserva-tion in 70% ethanol. Their brown head capsare considerably darker when compared withthe tonality present in the holotype. The capsextend downward on both sides of the head,achieving the level of the second row ofdorsals (MZUSP 12693 and 12694) or further,almost to the midventral line of the neck(MZUSP 12692). The connections of the bodystripes to the head cap are also variable: the
September 2011] HERPETOLOGICA 323

TA
BL
E1.
—V
aria
tion
inso
me
sele
cted
char
acte
rsof
stri
ped
-pat
tern
edA
post
olep
isex
amin
edh
erei
n(m
ean
s6
stan
dar
dd
evia
tion
and
sam
ple
size
follo
wra
nge
sin
par
enth
eses
).
Apo
stol
epis
long
icau
data
Apo
stol
epis
aff.
long
icau
data
1A
post
olep
isaf
f.lo
ngic
auda
ta2
Apo
stol
epis
nels
onjo
rgei
Apo
stol
epis
nigr
olin
eata
Apo
stol
epis
quin
quel
inea
ta
Tai
lle
ngt
h/s
nou
t–ve
nt
len
gth
Mal
es0.
15–0
.16
(0.1
56
0.01
,n
54)
0.17
(n5
1)0.
14–0
.16
(0.1
56
0.01
,n
53)
0.13
–0.1
6(0
.14
60.
01,
n5
20)
0.09
(n5
1)
Fem
ales
—0.
13(n
51)
0.12
–0.1
4(0
.13
60.
02,
n5
2)0.
08–0
.16
(0.1
16
0.02
,n
515
)0.
07–0
.08
(0.0
86
0.00
,n
53)
Lig
htsn
outs
pot
Abs
ent,
orsm
allw
hen
pres
ent
Lar
ge,
wel
ld
efin
edL
arge
,w
ell
def
ined
Lar
ge,
wel
ld
efin
ed,
pos
sib
lyp
aire
dU
sual
lysm
all
Lar
ge,
wel
ld
efin
ed
Ven
tral
s
Mal
es23
4–24
4(2
38.5
64.
80)
229
(n5
1)—
221–
256
(238
617
.52,
n5
3)20
8–22
7(2
13.5
64.
65,
n5
21)
228
(n5
1)
Fem
ales
——
237
(n5
1)26
0–26
1(2
60.5
60.
71,
n5
2)21
4–25
9(2
30.7
610
.10,
n5
15)
231–
243
(236
.76
6.03
,n
53)
Su
bca
ud
als
Mal
es49
–52
(50.
56
1.29
)54
(n5
1)—
41–5
333
–41
(37.
56
0.48
,n
50.
48)
31(n
51)
Fem
ales
——
41(n
51)
37–4
627
–43
(30.
76
3.97
,n
53.
97)
21–2
5(2
3.3
62.
08,
n5
3)P
reoc
ula
rn
asal
con
tact
Pre
sen
tL
acki
ng
Lac
kin
gV
aria
ble
Pre
sen
tP
rese
nt
Wh
ite
nu
chal
colla
rL
acki
ng
Lac
kin
gV
esti
gial
inad
ults
Pre
sen
tP
rese
nt
(usu
ally
vest
igia
l;ra
rely
com
ple
te)
Ves
tigi
al(o
nly
adu
lts
exam
ined
)B
lack
nu
chal
colla
rL
acki
ng
Lac
kin
gL
acki
ng
Pre
sen
tL
acki
ng
Pre
sen
t1
MZ
US
P16
615.
2M
ZU
SP
2405
.
324 HERPETOLOGICA [Vol. 67, No. 3

vertebral and lateral stripes of MZUSP 12692and 12694 are not in contact with the brownhead cap, but the same stripes are connectedto its posterior border in MZUSP 12693.Paravertebral stripes contact the brown headcap in all specimens; snout spots are virtuallylacking in MZUSP 12693, restricted to sparselight brown marks on the anterolateral surfaceof the prefrontals in MZUSP 12692 andrepresented by paired well-defined small lighttan spots on the anterior area of the prefrontalsin MZUSP 12694. The first light supralabialblotch is cream and occupies most of the firstsupralabial; the second light supralabial blotchcovers almost the whole area of the fourthsupralabial and, in MZUSP 12694, it invadesmore prominently the posterior and lowerportions of the third and the anterior portionof the fifth supralabials. The dorsal ground colorof MZUSP 12692–12694 is considerably darkerthan in the holotype; likewise, the lateral brownstripes are cohesive, mostly restricted to thefourth dorsal row, flanked in some regions of thetrunk by weakly defined interrupted linesformed by brown pigment on the apexes ofscales of the third and fifth rows; paravertebraland vertebral stripes are similar to the ones ofthe holotype.
Live specimens have considerably darkercolors than preserved specimens (MZUSP12692–12694). In life, the cephalic cap, terminal
caudal band, and stripes are dark brown toalmost black; the ground color of the trunk isorange-tan; the venter is cream.
Hemipenial morphology.—Description onthebasis of the fully everted and expandedright organ of MZUSP 12964 (Fig. 4). Hemi-penis unilobed; distal end of retractor muscledivided; body corresponding to two-thirds oftotal length of organ; small spines uniformlydistributed on proximal one-third of organ’sbase, entire sulcate face, and lateral and apicalportions of capitulum; base ornamented withtwo pairs of rows of enlarged spines, onelateral and composed of about five moderate-ly-sized spines, second one parasagittal toasulcate face and composed of four distinc-tively larger spines on each row; latter pair ofspine rows delimit an almost nude medianarea on asulcate face sparsely ornamented bysmall spinules; lobe differentiated as unicaly-culated capitulum, noncapitate, covered withpapilate calyces on sulcate and lateral faces;asulcate face of lobe restricted to concavearea, surrounded by small spines, forminginverted U-shaped and completely nude area;sulcus spermaticus forked and divided ap-proximately in middle of organ; branches ofsulcus centrolineal on body, acquiring slightlycentrifugal direction on capitulum. The aliz-arin red stained the spines and spinules, butno other calcified structures were detected.
FIG. 5.—Comparisons of head shape and color patterns in dorsal perspective: (A) Apostolepis longicaudata (MZUSP12693), (B) Apostolepis aff. longicaudata (MZUSP 2405), (C) Apostolepis aff. longicaudata (MZUSP 17614) and (D) A.nelsonjorgei (CHUNB 16182). Notice the variation regarding the white nuchal collar (absent in A and B, vestigial 583 inC, present and well developed in D). Scale bar 5 5.0 mm.
September 2011] HERPETOLOGICA 325

Distribution and ecology.—Apostolepis lon-gicaudata is reported from northeastern Brazil(type locality) and the Rio Tocantins Depres-sion. The type locality is in an ecotonal areabetween the Cerrado and Caatinga biomes(Fig. 6). Nonetheless, despite the presenceof sparse xeric elements, the type locality isgenerally dominated by cerrado vegetation.No data on habitat or field notes are availa-ble for the holotype.
Specimens MZUSP 12692–12694 were col-lected in the municipality of Guaraı, Tocan-tins, about 270 km NW of the type locality(Fig. 6). Quartzitic sandy soils predominate inthe region (SEPLAN, 1999), and the land-scape is basically composed of typical opencerrado vegetation interrupted by interfluvialand gallery forests. These specimens were col-lected in pitfall traps, always checked in themorning. The pitfall trap system at each of thecollection sites included 40 buckets (height 540 cm; diameter 5 30 cm; volume 5 18 L)
assembled in 10 Y-shaped stations (Jones,1981). Each station consisted of one centraland three peripheral buckets, all connectedto the first by plastic fences 4 m long and50 cm high.
MZUSP 12693 and 12694 were collected intwo small forest fragments, both on sandy soilsand covered by thick layers of leaf litter.MZUSP 12694 was collected in a gallery forestat an elevation of about 260 m (‘‘point 1’’ ofFazenda Marupiara; see Appendix for precisecoordinates); the canopy was mostly closed,although sparse openings allowed sunlight totouch the ground in some small clearings.Other reptile species recorded in the areawere Colobosaura modesta (Gymnophthalmi-dae), Kentropyx calcarata (Teiidae), and Typ-hlops brongersmianus (Typhlopidae; collec-tion effort 480 bucket-days; sample period18–29 October 2001, 12 d). MZUSP 12693was found in an interfluvial forest about 14 kmsouthwest of the collection site of MZUSP
FIG. 6.— White triangles 5 distributional records of Apostolepis longicaudata [(1) Santa Filomena, Piauı 5 typelocality, (2) Guaraı, Tocantins 5 locality of the three new records reported herein]; black triangles 5 records of A. aff.longicaudata [(1) Linhares, Espırito Santo, (2) Cabaceiras, Paraıba].
326 HERPETOLOGICA [Vol. 67, No. 3

12694 (‘‘point 2’’ of Fazenda Marupiara; seeAppendix for precise coordinates) at a slightlyhigher elevation. Large trees were still presentin the area, though presence of lianas, smallshrubs, and several clearings were obviousevidence of disturbance. Bachia micromela,C. modesta (Gymnophthalmidae), Anolis chry-solepis (Polychrotidae), Ameiva ameiva (Teii-dae), and T. brongersmianus (Typhlopidae)were other reptile species collected at thissite (collection effort 440 bucket-days; sampleperiod 18–29 October 2001, 11 d).
In contrast to both previous individuals,MZUSP 12692 was obtained in an open cer-rado area on quartzitic sandy soils (‘‘point 3’’of Fazenda Marupiara; see Appendix I forprecise coordinates). Short and twisted treeswere sparsely distributed and small bushesof grass were frequent. Narrow tracks of baresand remained directly exposed to sun (col-lection effort: 480 bucket-days; sample period18–29 October 2001, 12 d). However, it isimportant to mention that, despite the strikingdifferences in overall structure among theforest areas of MZUSP 12693 and 12694 andthe open cerrado of MZUSP 12692, the lat-ter specimen was collected in a pitfall stationplaced contiguous to a gallery forest, quitesimilar in structure to the area of collectionof MZUSP 12694. Thus, it would not beunreasonable to consider the record ofMZUSP 12692 in an open cerrado area as apossible effect of the contact with foresthabitats to which the species might showsome degree of preference.
DISCUSSION
Until additional specimens are collected,we can only speculate about the distributionof this rare elapomorphine. Colli et al. (2002)considered A. longicaudata endemic to theCerrado biome. Since the species occurs inforested sandy soils of the Rio TocantinsDepression, it may be widespread in similarsubstrates throughout the Cerrado of north-ern and northeastern Brazil. Several squamatetaxa such as Parapostolepis polylepis (Amaral,1921b, 1930; has not yet been formallysynonymized with Apostolepis) and Amphis-baena ibijara (Rodrigues et al., 2003) appearto be largely restricted to the sandy formationsof the Parnaıba sedimentary basin, and A.
longicaudata may have a similar dis-tribution.
The specimens from the Tocantins Depres-sion have slightly acuminate snouts (Fig. 5A),contrasting with the rounded snout of theholotype (Fig 2A). This difference cannot beattributed to allometric variation, because allspecimens are adults. Alternatively, the slightlyacuminate snouts may be a local adaptation tothe particular texture of quartzitic sandy soilspresent in Guaraı.
Like other fossorial vertebrates, most Apos-tolepis species have short tails (subcaudalcounts lower than 40 scales). A relatively longtail and high subcaudal counts for the genusare the most important diagnostic featuresof A. longicaudata. The only species thatundisputedly has subcaudal counts overlap-ping with those of this species is A. nelsonjor-gei. Nonetheless, this fact is contradicted bysome recent publications where reports ofhigh subcaudal counts in other species werebased on misidentified specimens of A.nelsonjorgei.
On the basis of Lema and Renner (1998),Lema (2001:36) claimed that the high sub-caudal counts of A. longicaudata would be‘‘equal’’ to those of A. quinquelineata and A.nigrolineata. According to Boulenger (1896),the holotype of A. quinquelineata is a malespecimen with 28 subcaudals (Ferrarezzi, 1993;Hoge, 1958). Lema and Renner (1998:103) re-port 22–30 subcaudals in females and 28–53 sub-caudals in males of this species, suggesting exis-tence of an enormous discrepancy between theranges of both sexes. However, subcaudal countshigher than 40 scales occur in only three spec-imens examined by Lema and Renner (1998;MPEG 11487, 16954, and 17051). We re-examined MPEG 16954 and 17051, both with52/53 subcaudals, and identified them as A.nelsonjorgei. The third specimen (MPEG 11487,with 48 subcaudals according to Lema andRenner, 1998) was not available for study. Ex-cluding these three specimens, male A. quinque-lineata in the sample of Lema and Renner (1998)have 28–37 subcaudals, which is in better agree-ment with the variation observed in females.
Regarding A. nigrolineata, nomenclaturalconfusion explains high subcaudal counts re-ported by Lema (1997, 2001) for this spe-cies. Lema (1997) considered A. nigrolineata
September 2011] HERPETOLOGICA 327

(Peters, 1869) to be a senior synonym of A.pymi Boulenger, 1903. Nonetheless, 1 yrlater, Lema and Renner (1998) used thename A. pymi when referring to populationsof the eastern Amazon. This inconsistent useof names is apparently due to timing of thepublications involved, because Lema (1997:194, 197, 199) explicitly cites Lema andRenner (1998). Presumably, Lema and Re-nner (1998) was submitted before Lema(1997), and the features of A. pymi providedby Lema and Renner (1998) must beinterpreted as features of A. nigrolineata(see also Lema, 2001). In view of thisinformation, since Lema and Renner (1998)claim that subcaudals of A. nigrolineata rangefrom 21 to 40 in females and from 33 to 46 inmales, we believe that the report of 52subcaudals in A. nigrolineata by Lema (1997)resulted from confusion with Lema and Renner’s(1998) data for A. quinquelineata (MPEG 16954and 17051, herein identified as A. nelsonjorgei).Among specimens examined by us (Table 1),male A. nigrolineata have fewer ventrals(t21,15 5 6.17, P , 0.000), more subcaudals(t20,15 5 6.01, P , 0.000), and relatively longertails (F20,15 5 97.62, P , 0.000, Fig. 7) than
females. Moreover, the relatively shorter tails ofmale A. nigrolineata immediately distinguish thisspecies from A. longicaudata (F20,4 5 10.42, P 50.004, Fig. 7).
In summary, there are no reliable records ofmore than 46 subcaudals in A. nigrolineataand 37 subcaudals in A. quinquelineata. Onthe other hand, considering the data from theliterature (Lema and Renner, 2004a), alongwith additional specimens examined herein(Table 1), males of A. nelsonjorgei may haveup to 53 subcaudals (MPEG 16954), clearlyoverlapping the range of A. longicaudata.
During this study, we examined two striped-patterned specimens of doubtful identity (MZ-USP 2045 and MZUSP 17614) that deserveattention. Ferrarezzi (1993) considered thesespecimens related to A. longicaudata becausethey have 0+0 temporals, fifth and sixth supra-labials in contact with parietals, five dorsalstripes similar in width, a white termimal scale,and subcaudal counts high for the genus (54/54and 40/41, respectively). Nevertheless, thesame author pointed out that both specimenswere distinct from each other and from A.longicaudata with respect to features of headscalation and color pattern.
FIG. 7.—Relative tail lengths of two species of stripe-patterned Apostolepis from eastern Brazil.
328 HERPETOLOGICA [Vol. 67, No. 3

Herein, we opted for a cautious posturewith respect to these specimens, avoiding afinal decision regarding their identification;instead, we refer to them as Apostolepis aff.longicaudata. Both individuals differ from typ-ical A. longicaudata in lacking preocular-nasalcontact, as well as by presence of a large lightsnout spot (vs. preocular-nasal contact presentand snout pattern mostly black with small andirregular light blotches). In addition, MZUSP17614 has rudiments of a white nuchal collar,which is completely lacking in MZUSP 2045and in all four specimens of A. longicaudata(compare head patterns on Fig. 5). A specimenillustrated by Freitas (2003:pl. 157), fromItapebi, state of Bahia (15u579S, 39u329W,177 m; sensu www.glosk.com), which we wereunable to locate, seems to fit the characterizationof MZUSP 17614 at least with respect to colorpattern.
Since the character states related to snoutcolor and preocular-nasal contact are poly-morphic in other striped-patterned Apostole-pis (Harvey, 1999; Lema and Renner, 2004a),MZUSP 2045 and 17614 could be attributedto A. longicaudata and these features wouldbe implicitly regarded as polymorphic also forthis species. However, both specimens werecollected far from the known range of A.longicaudata. MZUSP 17614 is a female col-lected in the vicinities of the municipalityof Cabaceiras, state of Paraıba, northeasternBrazil, in the core area of the Caatinga biome(Fig. 6). The specimen was found in a mesicforest sheltered by a large block of crystallinerocks and the soil was composed of moder-ately compacted clay, densely covered withleaf litter. On the other hand, MZUSP 2045 isa male from Linhares, state of Espırito Santo,in the Atlantic Rainforest of southeasternBrazil (Fig. 6); this specimen has obviousanomalies in head scalation (Table 1).
Further study is required to determine wheth-er these specimens from eastern Brazil areconspecific with A. longicaudata. The easternlocalities appear ecologically quite different fromthe Cerrado, but resolution of this problem mustawait collection of additional specimens.
Finally, we mention that this paper has asad peculiarity. On 15 May 2010, a tragicaccident caused a fire that consumed most ofthe more than 80,000 snake specimens housed
in the collection ‘‘Alphonse Richard Hoge’’at the Instituto Butantan. This tragic lossof Neotropical snakes included the largestsamples of Brazilian taxa, assembled throughmore than a century of work. Along with sev-eral type specimens, Gomes’ holotype of A.longicaudata has apparently disappeared andis probably lost forever. Our measurementsand recent photographs of IBSP 1684 are thelast information obtained through direct ex-amination of this specimen.
Note added to proofs.— While our paperwas submitted, Lema and Albuquerque(2010) resurrected A. pymi and synonymizedA. quinquelineata with A. nigrolineata. Theauthors provided a list of characters thatdistinguish A. nigrolineata from A. pymi;however, interspecific variation in thesecharacters is greater than their brief noteindicates. For example, Lema and Albuquer-que (2010) claimed that only juvenile A.quinquelineata have five dorsal stripes. Inour sample, three adult specimens (MPEG17453, 17982–83, each more than 300 mmSVL) retained five distinct and cohesivedorsal stripes, whereas Lema and Albuquer-que (2010) claimed that at least the paraver-tebral stripes tend to fade throughout devel-opment and become indiscernible in fullygrown individuals. Herein, we continue torecognize A. nigrolineata (A. pymi included)and A. quinquelineata as defined respec-tively by Lema (1997) and Lema and Renner(1998). Nonetheless, the status of thesenames needs clarification and must beassessed elsewhere.
Acknowledgments.—We are indebted to Francisco L.Franco, Valdir J. Germano (IBSP), Ana L. Prudente,Paula C. R. de Almeida (MPEG), Hussam Zaher, andCarolina Mello (MZUSP) for allowing access to specimensunder their care. Dante Pavan coordinated fieldwork andprovided important suggestions regarding general contentof the manuscript. Hebert Ferrarezzi, Giovanna Mon-tingelli, Ana L. Prudente, and anonymous reviewersprovided constructive comments and criticisms to earlyversions of the paper. Glaucia Pontes and Thales de Lemahelped with literature. Otavio A. V. Marques providedphotographs of Apostolepis longicaudata in life; CristinaA. Rheims and Antonio D. Brescovit helped with thephotographs of the holotype of A. longicaudata. FFC issupported by a grant from the Conselho de Desenvolvi-mento Cientıfico e Tecnologico (CNPq; FFC–grant #152193/2008-1). PMSN (grant # 07/00811-8) and MTRare grateful to Fundacao de Amparo a Pesquisa do Estadode Sao Paulo (FAPESP) for support.
September 2011] HERPETOLOGICA 329

LITERATURE CITED
AMARAL, A. 1921a. Ultimos trabalhos ineditos de J. Flo-rencio Gomes: Duas novas especies de Colubrideos opi-stoglyphos brasileiros (Philodryas oligolepis e Apostole-pis longicaudata). Annaes Paulistas de Medicina e Cirurgia9:157–163.
AMARAL, A. 1921b. Contribuicao para o conhecimento dosophidios do Brasil—A. Parte 1. Quatro novas especiesde serpentes brasileiras. Anexos das Memorias doInstituto Butantan (Ofiologia) 1:1–88.
AMARAL, A. 1930. Estudos sobre ophidios dos Neotropicos.XVII. Valor systematico de varias formas de ophidiosdos Neotropicos. Memorias do Instituto Butantan4:3–68.
BOULENGER, G. A. 1896. Catalogue of the snakes in theBritish Museum (Natural History), Volume 3. BritishMuseum, London, UK.
BOULENGER, G. A. 1903. Descriptions of new snakes in thecollection of the British Museum. Annals and Magazineof Natural History, London 7:350–354.
COLLI, G. R., R. B. BASTOS, AND A. F. B. ARAUJO. 2002. Thecharacter and dynamics of the Cerrado herpetofauna.Pp. 223–241. In P. S. Oliveira and R. J. Marquis (Eds.),The Cerrados of Brazil: Ecology and Natural Historyof a Neotropical Savana. Columbia University Press,New York, New York, USA.
DOWLING, H. G., AND J. A. SAVAGE. 1960. A guide to snakehemipenis: Survey of basic structure and systematiccharacteristics. Zoologica 45:17–28.
FERRAREZZI, H. 1993. Sistematica filogenetica de Elapo-morphus, Phalotris e Apostolepis (Serpentes: Colubri-dae: Xenodontinae). 1993. Masters Thesis, Instituto deBiociencias, Universidade de Sao Paulo, Sao Paulo,Brazil.
FERRAREZZI, H. 1994. Uma sinopse dos generos eclassificacao das serpentes (Squamata) II. FamıliaColubridae. Pp. 81–91. In L. B. Nascimento, A. T.Bernardes, and G. A. Cotta (Eds.), Herpetologia noBrasil, I. PUC-MG, Fundacao Biodiversitas e FundacaoEzequiel Dias, Belo Horizonte, Brazil.
FERRAREZZI, H., F. E. BARBO, AND C. E. ALBUQUERQUE.2005. Phylogenetic relationships of a new species ofApostolepis from Brazilian Cerrado with notes on theassimilis group. Papeis Avulsos de Zoologia 45:215–229.
FREITAS, M. A. 2003. Serpentes Brasileiras. Malha-de-Sapo Publicacoes e Consultoria Ambiental, ProquigelQuımica, Bahia, Brazil.
HAMMER, Ø., D. A. T. HARPER, AND P. D. RYAN. 2001.PAST: Paleontological statistics software package foreducation and data analysis. Palaeontologia Electronica4:1–9.
HARVEY, M. B. 1999. Revision of Bolivian Apostolepis(Squamata: Colubridae). Copeia 1999:388–409.
HARVEY, M. B., AND D. EMBERT. 2008. Review of BolivianDipsas (Serpentes: Colubridae) with comments onother South American species. Herpetological Mono-graphs 22:54–105.
HARVEY, M. B., G. R. FUENMAYOR, J. R. C. PORTILLA, AND
J. V. RUEDA-ALMONACID. 2008. Systematics of theenigmatic dipsadine snake Tropidodipsas perijanensisAleman (Serpentes: Colubridae) and review of mor-phological characters of Dipsadini. HerpetologicalMonographs 22:106–132.
HOGE, A. R. 1958. Etude sur Apostolepis coronata(Sauvage 1877) et Apostolepis quinquelineata Boulen-ger 1896 (Serpentes). Memorias do Instituto Butantan28:73–76.
JONES, K. B. 1981. Effects of grazing on lizard abundanceand diversity in western Arizona. Southwestern Natu-ralist 26:107–115.
KOSLOWSKY, J. 1898. Ofıdios de Mato-Grosso (Brasil).Revista del Museo de La Plata 8:25–32.
LEMA, T. 1993. Polimorfismo em Apostolepis dimidiata(Jan, 1862) com a invalidacao de Apostolepis villaricaeLema, 1978 e Apostolepis barrioi Lema, 1978 (Ser-pentes: Xenodontinae: Elapomorphini). Acta BiologicaLeopoldensia 15:25–52.
LEMA, T. 1997. A redescription of the tropical Brazilianserpent Apostolepis nigrolineata (Peters 1869), (Colu-bridae, Elapomorphinae), synonymous with A. pymiBoulenger. Studies on Neotropical Fauna and Envi-ronment 32:193–199.
LEMA, T. 2001. Fossorial snake genus Apostolepis fromSouth America (Serpentes: Colubridae: Elapomorphi-nae). Cuadernos de Herpetologıa 15:29–43.
LEMA, T. 2002. Nova especie de Apostolepis do grupolineata do sudoeste do Brasil (Serpentes, Elapomor-phinae). Facena 18:41–52.
LEMA, T. 2004. Nova especie de Apostolepis Cope doestado de Rondonia, Brasil (Serpentes: Elapomorphi-nae). Comunicacoes do Museu de Ciencias e Tecnologia,PUCRS, Serie Zoologia 17:81–89.
LEMA, T., AND M. F. RENNER. 1998. O status deApostolepis quinquilineata Boulenger, 1896, A. pymiBoulenger, 1903 e A. rondoni Amaral, 1925 (Serpentes,Colubridae, Elapomorphini). Biociencias, Porto Alegre6:99–121.
LEMA, T., AND M. F. RENNER. 2004a. New species ofApostolepis from Alto Tocantins, Brazil, with commentson the stripe pattern species (Serpentes, Elapomorphi-nae). Biociencias, Porto Alegre 12:139–145.
LEMA, T., AND M. F. RENNER. 2004b. New specimens ofApostolepis vittata (Cope 1887) (Serpentes, Elapomor-phinae). Caderno de Pesquisa, Serie Biologia, SantaCruz do Sul 16:51–56.
LEMA, T., AND M. F. RENNER. 2004c. Description of a newspecies of Apostolepis Cope 1861 (Serpentes, Elapo-morphinae) from the Brazilian Cerrado. Acta BiologicaLeopoldensia 26:155–160.
LEMA, T., AND M. F. RENNER. 2006. A new species ofApostolepis with striped pattern, from Mato Grosso,Brazil (Serpentes, Elapomorphinae). Ciencia em Mo-vimento 15:13–18.
LEMA, T., AND N. R. ALBUQUERQUE. 2010. The identity ofApostolepis pymi and placement of A. quinquelineata inthe synonymy of A. nigrolineata (Serpentes, Xenodon-tinae). Biota Neotropica 10:343–366.
MANZANI, P. R., AND A. S. ABE. 1988. Sobre dois novosmetodos de preparo de hemipenis de serpentes.Memorias do Instituto Butantan 50:15–20.
PAYNTER, R. A., JR., AND M. A. TRAYLOR, JR. 1991.Ornithological Gazetteer of Brazil, 2nd Edition. BirdDepartment, Museum of Comparative Zoology, Har-vard College, Cambridge, Massachusetts, USA.
PESANTES, O. 1994. A method for preparing the hemipenisof preserved snakes. Journal of Herpetology 28:93–95.
330 HERPETOLOGICA [Vol. 67, No. 3

PETERS, J. A. 1964. Dictionary of Herpetology. HafnerPublishing Company, New York, New York, USA.
PETERS, J. A., AND B. R. OREJAS-MIRANDA. 1970. Catalogueof the Neotropical Squamata. Part 1. Snakes. UnitedStates National Museum Bulletin 297:1–347.
PETERS, W. 1869. Ueber neue Gattungen und neue oderweniger bekannte Arten von Amphibien (Eremias,Dicrodon, Euprepes, Lygosoma, Typhlops, Eryx,Rhynchonyx, Elapomorphus, Achalinus, Coronella,Dromicus, Xenopholis, Anoplodipsas, Spilotes, Tropi-donotus). Monatsbericht der Koniglich-PreussischenAkademie der Wissenschaften zu Berlin 1869:432–447.
RODRIGUES, M. T. 1993. Herpetofauna das dunas inter-iores do Rio Sao Francisco: Bahia: Brasil V. Duas novasespecies de Apostolepis (Ophidia, Colubridae). Mem-orias do Instituto Butantan 54:53–59.
RODRIGUES, M. T., G. V. ANDRADE, AND J. D. LIMA. 2003. Anew species of Amphisbaena (Squamata, Amphisbae-nidae) from state of Maranhao, Brazil. Phyllomedusa2:21–26.
SEPLAN (SECRETARIA DO PLANEJAMENTO E MEIO AMBIENTE,TOCANTINS). 1999, Atlas do Tocantins: subsıdios aoplanejamento da gestao territorial. Diretoria de Zonea-mento Ecologico-Economico (DEZ), Palmas, Tocantins,Brazil.
UZZEL, T. 1973. A revision of lizards of the genusPrionodactylus, with a new genus for P. leucostictusand notes on the genus Euspondylus (Sauria, Teiidae).Postilla 159:1–67.
ZAHER, H. 1999. Hemipenial morphology of the SouthAmerican Xenodontine snakes, with a proposal for amonophyletic Xenodontinae and a reappraisal ofColubroid hemipenes. Bulletin of the American Muse-um of Natural History 240:1–168.
ZAHER, H., F. G. GRAZZIOTIN, J. E. CADLE, R. W. MURPHY,J. C. MOURA-LEITE, AND S. L. BONATTO. 2009. Molecularphylogeny of advanced snakes (Serpentes, Caenophi-dia) with an emphasis on South American Xenodon-tines: A revised classification and descriptions of newtaxa. Papeis Avulsos de Zoologia 49:115–153.
.Accepted: 11 May 2011
.Associate Editor: Christopher Austin
APPENDIX I
Specimens Examined
Specimens are listed according to their respectivelocalities, organized as ‘‘COUNTRY: STATE: localityname’’; geographic coordinates are based on the WGS84datum. Coordinates of localities marked with an ‘‘*’’ wereobtained primarily from gazetteers (Paynter and Traylor,
1991; www.glosk.com) and then refined with the aid of thesoftware Google Earth 5.0.11733.9347; coordinates oflocalities marked with ‘‘**’’ were directly taken in the fieldusing a hand-held global positioning system receiver.
Apostolepis longicaudata.—BRAZIL: PIAUI: SantaFilomena* (09u079S, 45u559W, 277 m): IBSP 1684(holotype); TOCANTINS: Guaraı* (08u509S, 48u309W,259 m), ‘‘Fazenda Marupiara’’** (8u389S, 48u279W), point1** (8u369S, 48u189W, 258 m): MZUSP 12964; point 2**(8u389S, 48u259W, 296 m): MZUSP 12963; point 3**(08u159S, 48u199W, 250 m): MZUSP 12962.
Apostolepis aff. longicaudata.—BRAZIL: ESPIRITOSANTO: Linhares*: (19u239S, 40u049W, 33 m): MZUSP2405; PARAIBA: Cabaceiras**: (07u299S, 36u179W,388 m): MZUSP 17614.
Apostolepis nelsonjorgei.—BRAZIL: GOIAS: Serra daMesa dam, Point 1** (14u019S; 48u199W, 460 m): MZUSP10997; Conceicao do Araguaia* (08u159S, 49u159W,165 m) MPEG 16954; 17051; TOCANTINS: EstacaoEcologica Serra Geral do Tocantins** (11u079S, 46u469W,560 m): MZUSP 17615; Porto Nacional*, (10u429S,48u259W, 212 m): CHUNB 16182.
Apostolepis nigrolineata.—BRAZIL: PARA: Ananin-deua* (01u279S, 48u229W, 17 m): MPEG 2368, 6943,9459; Benevides, Santa Barbara* (01u229S, 48u159W,28 m): MPEG 18513; Braganca* (01u039S, 46u469W,19 m): MPEG 13009; Capitao Poco* (01u459S, 47u049W,73 m): MPEG 10461, 12170; Castanhal, Macapazinho*(01u189S, 47u569W, 41 m): MPEG 11794, 12693; ColoniaNova, BR-316 road, near Rio Gurupi* (approximatecoordinates: 01u299S, 46u229W, 50 m): MPEG 15063,15863; Conceicao do Araguaia, Rio Maria* (08u159S,49u169W, 165 m): MPEG 17671; Curupati (not located):MPEG 10887, 13260; Igarape-Acu* (01u089S, 47u379W,37 m): MPEG 925; Ilha de Mosqueiro* (01u079S,48u249W, 10 m): MPEG 12771; Inhangapi* (01u269S,47u559W, 35 m): MPEG 1571; Nova Vida, BR-316, 25 Kmdo Rio Gurupa (not located): MPEG 16227–29; Ourem*(01u339S, 47u079W, 12 m): MPEG 7016, 7019; Parua, BR-316 road (not located): MPEG 10835, 14352; Acara road,Km 16 (not located): MPEG 12803–04; Santa Maria,rodovia BR-226 (not located): MPEG 16137; Maracanaroad, km 23 (not located): MPEG 8187; Santa Rosa (notlocated): MPEG 11835–836, 12590; Santarem Novo,Trombetinha* (00u569S, 47u249W, 30 m): MPEG 7081;Santo Antonio do Taua* (01u099S, 48u089W, 51 m):MPEG 7557; Tome-Acu* (02u259S, 48u099W, 20 m):MPEG 11723; Viseu* (01u129S, 46u089W, 6 m): MPEG15126–27.
Apostolepis quinquelineata.—BRAZIL: AMAZONAS:Presidente Figueiredo, Balbina Hydroelectric PowerPlant* (02u029S, 60u019W, 122 m): MPEG 17453; RON-DONIA: Porto Velho, Samuel Hydroelectric Power Plant*(08u469S, 63u549W, 145 m): MPEG 17453, 17982–83.
September 2011] HERPETOLOGICA 331