antibody glomerular filtration in
TRANSCRIPT

The Acute Effects of Antiglomerular Basement Membrane
Antibody upon Glomerular Filtration in the Rat
THE INFLUENCE OF DOSEANDCOMPLEMENTDEPLETION
ROLANDC. BLANTZ, Department of Medicine, University of California, San DiegoSchool of Medicine, and the Veterans Administration Hospital, San Diego,California 92161
BRYANJ. TUCKER, Department of Medicine, University of California, San DiegoSchool of Medicine, San Diego, California 92093
CURTIS B. WILSON, Department of Immunopathology, Scripps Clinic and ResearchFoundation, La Jolla, California 92037
A B S T RA C T Recent studies from this laboratoryhave revealed that single nephron filtration rate (sngfr)decreases significantly within 1 h of the administra-tion of large doses of complement-fixing antiglomeru-lar basement membrane antibody (AGBM Ab) inplasma-expanded Munich-Wistar rats. This reductionin sngfr was due to decreases in nephron plasmaflow (rpf) and the glomerular permeability coefficient(LpA) utilizing direct evaluation of all pertinent pres-sures, flows, and permeabilities. With identical micro-puncture techniques, we have determined (a) therespective influences of rpf and LpA upon sngfr byexamining the effects of differing doses of AGBMAb,and (b) the specific effect of complement fixationupon the reduction in sngfr. In normal rats, low dose(1.4 lug/g body wt) AGBMAb decreased sngfr from57.9+3.4 to 50.8+±3.9 nl/min per g kidney wt (kw)(P < 0.001), and this was due to a 10% reductionin rpf and a decrease in LpA from 0.069+0.014 incontrol to 0.041±0.007 nl/s per g kw per mmHg (P< 0.02). At the high dose (2.3 gg/g body wt),sngfr fell dramatically from 58.4±4.0 to 7.6±3.8 nl/minper g kw (P < 0.001), and this effect upon filtration was
This is publication 1371 from the Department of Im-munology, Scripps Clinic and Research Foundation, La Jolla,Calif. 92037.
Dr. Blantz is a Clinical Investigator of the VeteransAdministration. B. J. Tucker is a graduate student in AppliedMechanics and Engineering Sciences, Univ. of Calif., SanDiego.
Received for publication 15 August 1977 and in revisedform 10 November 1977.
the result of an 86% reduction in rpf and a decreasein LpA from 0.092+0.020 to 0.007+0.004 nl/s per g kwmmHg (P < 0.001). Therefore, at lower doses sngfrfell primarily as a result of a 40% reduction in LpA anda 10% decrease in rpf; however, at the high dosemassive reductions in both rpf and LPA led to thelarge decrease in sngfr.
In complement-depleted rats, receiving identicaldoses, low-dose AGBMAb no longer reduced thesngfr, but a reduction in LpA persisted (other factorscompensating to maintain sngfr). At the high dose,complement depletion ameliorated the reduction insngfr (55.1+2.4 to 37.2+3.4 nl/min per g kw mmHg)by nearly eliminating the vasoconstriction but onlypartially diminished the reduction in LpA (0.097+0.020to 0.032+0.004 nl/s per g kw mmHg, P < 0.05).
Complement depletion prevented the migration ofpolymorphonuclear leukocytes (present in largernumbers after the high dose of AGBMAb) into thecapillary and eliminated vasoconstriction. Comple-ment depletion resulted in a lesser effect of high-dose AGBMAb upon LpA than in normal rats, andthis is likely due to lesser polymorphonuclearleukocyte effects upon capillary surface area. Thepersistent reduction in LpA observed in complement-depleted rats correlated with separation of the endo-thelial cell from the glomerular basement membraneafter AGBMAb. AGBMAb diminishes glomerularultrafiltration by decreasing LpA and altering theendothelial surface of the glomerular membrane, andthis effect is not totally dependent upon the fixation ofcomplement.
J. Clin. Invest. © The American Society for Clinical Investigation, Inc., 0021-9738/78/0401-0910 $1.00910

INTRODUCTION
Recently (1), we demonstrated that nephron filtrationrate (sngfr)' in rats decreased within 60 min after theadministration of a large dose of complement-fixing anti-glomerular basement membrane antibody (AGBMAb).The various determinants of nephron filtration weremeasured utilizing micropuncture techniques thatallow all pertinent pressures, flows, and permeabilitiesto be determined directly in the control conditionand after antibody administration. The study revealedthat sngfr decreased because of reductions in boththe glomerular permeability coefficient (LpA) andnephron plasma flow (rpf). This experimental approachto the assessment of glomerular immune injury pro-vides certain significant advantages. With micropunc-ture techniques not only can changes in sngfr thatfollow immune injury be evaluated accurately, but alsofactors that affect the filtration rate can be assessedindividually both before and after the immune insult.
Two questions relevant to the specific mechanismsleading to reduced nephron filtration have been posedin this study. Firstly can we separate the respectiveinfluences of the factors that lead to reduction insngfr: decreased rpf and reduced LpA? Secondly, whatrole does activation of the complement system play inreducing the filtration rate after glomerular immuneinjury? Wehave examined these two issues by studyingthe changes that occur in the factors influencingglomerular filtration when (a) high and low doses ofAGBMAb are administered and (b) when complementactivity is depleted with cobra venom factor (2).
METHODSExperiment animals. The current studies were performed
on Munich-Wistar rats (180-240 g body wt), bred and main-tained in a colony housed at the Animal Research Facilityat the Veterans Administration Hospital, San Diego, Calif.
Preparation of AGBMAb. AGBMAb was produced byimmunizing rabbits repeatedly with 10-20 mgof rat glomeru-lar basement membrane (GBM) in complete Freund'sadjuvant. Rat GBMwas prepared by a modification of themethod of Krakower and Greenspon (3). The attainment ofnephrotoxic levels of AGBMAb in rabbits was recognizedwhen intravenous injection of rabbit serum induced acute
1Abbreviations used in this paper: AGBM,Ab, antiglomeru-lar basement membrane antibody; AR, afferent arteriolarresistance; C, protein concentration; CVF, cobra venom factor;AP, hydrostatic pressure gradient across glomerular mem-brane; EFP, effective filtration pressure; EFP, mean effectivefiltration pressure; ER, efferent arteriolar resistance; GBM,glomerular basement membrane; kw, kidney weight; L,A,glomerular permeability coefficient; ITA, systemic oncoticpressure; irE, efferent arteriolar oncotic pressure; PG, glomer-ular capillary hydrostatic pressure; PMN, polymorphonuclearleukocytes; Pt, Bowman's space hydrostatic pressure; rpf,nephron plasma flow; sngfr, single nephron filtration rate;x*, normalized unit glomerular capillary length.
proteinuria in rats. Rabbit serum was then collected, pooled,absorbed with rat plasma and peripheral blood cells, and thegammaglobulin fraction separated and concentrated by pre-cipitation at a final concentration of 50% saturated ammoniumsulfate. The gamma globulin fractions obtained by thisprocedure and normal rabbit gammaglobulin fractions werepair labeled with I131 and I125 radioactive iodine, and theamount of kidney-fixing antibody was quantitated using thepaired label isotope technique (4, 5). The pool of AGBMAbutilized in this study was obtained from a different group ofrabbits than that used in the previous study (1).
Methods utilized for decomplementation. Cobra venomfactor (CVF; kindly prepared by Dr. Richard J. Ulevitchof Scripps Clinic and Research Foundation, La Jolla, Calif.)(6) was injected intraperitoneally into one group of rats. Eachrat received 85 U/kg body wt of CVF at 48, 44, 40, 28, and24 h before micropuncture for a total dose of 425 U/kg bodywt (7). Blood was drawn from the tail vein approximately16 h before micropuncture surgery, and the serum assayedfor C3 levels by double immunodiffusion in gel. Ratspretreated with CVF had <5% of the C3 immunoreactantnormally present in the Munich-Wistar rat. The effectivenessof complement depletion in all rats studied was also assessedby immunofluorescence of the glomeruli for C3.
In separate studies, CVF at the doses used had noquantitative effect upon the binding of AGBMAb to renaltissue as shown by the quantitative paired label radioisotopetechnique (4, 5).
Micropuncture studies evaluating glomerular ultrafiltra-tion before and after AGBMAb. The micropuncture proto-col utilized in this study was nearly identical to thatdescribed in our previous study on glomerular immuneinjury (1). Surgical preparation was as previously described,and all studies were paired with iso-oncotic plasma expansion(2.5% body wt administered over 60 min) which waschosen as the control condition (1). A separate infusion of[14C]inulin dissolved in isotonic NaCl-NaHCO3 (0.5% bodywt/h) was begun at the time of plasma expansion and wasdelivered at approximately 40 ,uCi/h.
After equilibration of radioactive inulin, control measure-ments of glomerular capillary and Bowman's space hydro-static pressure (utilizing a servo-nulling device with 1- to2-,um tip pipettes; 1, 8) and of sngfr (n = 5) were obtainedand at least three samples of efferent peritubular capillaryblood from "star" vessels were obtained (8).
After completion of the control measurements, either lowor high dose (1.4 or 2.3 ,ug/g kidney wt [kw]) AGBMAbwas administered intravenously over 5 min in a volume of400 ,ul isotonic NaCl-NaHCO3. Urine flow decreased acutelywith infusion of high doses of AGBMAb but was changedlittle by the lower dose. 15 min after initiation of AGBMAbinfusion, all pressure, filtration rate, and efferent proteinconcentration measurements were repeated and completedwithin 45 min. In a previous study on the mechanism ofglomerular immune injury with AGBMAb, we have deter-mined that inulin remains a valid marker of glomerularultrafiltration even after antibody is administered (1).
Analytic methods. Protein concentration in systemic andefferent peritubular blood samples was measured by a micro-adaptation of the Lowry protein method (8-10). Sngfr,glomerular filtration rate, renal plasma flow and renal bloodflow, urine and plasma sodium and potassium concentrationswere determined as described in the previous study (1).
Morphological studies. Tissue for histologic, immuno-fluorescent, and electron microscope studies was obtainedfrom both kidneys at the termination of the study andprocessed as previously described (1, 11, 12). The range of
Antiglomerular Basement Membrane Antibody: Dose and Complement Effect 911

TABLE IThe Effect of AGBMAb upon Various Clearances Indices at Both Doses in
Normal and Complement-Depleted Rats
UNaV* UkVt UV§
C E C E C E
lteqImin zeq/min ;LI/min
Normal ratsLow-dose group (n = 6)" 2.6 1.1¶ 1.2 0.6¶ 24 8.0¶
±0.6 +0.4 ±0.2 ±0.2 +7 ±2.7
High-dose group (n = 6)** 5.1 0.10¶ 1.4 0.20¶ 40 1.01±+1.7 ±0.03 +0.3 +0.07 +6 ±0.2
Complement-depleted ratsLow-dose group (n = 9)" 1.8 1.4 1.0 0.8 17.1 15.0
±0.4 +0.3 +0.3 ±0.1 ±5.4 +4.0
High-dose group (n = 6)** 2.3 0.5¶ 1.7 0.3¶ 21 2.6¶±0.5 +0.2 +0.3 ±0.1 ±3 ±1.0
* Sodium excretion.t Potassium excretion.§ Urine volume."1.4 ,ug/g body wt of AGBMAb.¶ Experiment (E) significantly different from control (C), P < 0.05 or less.** 2.3 ,ug/g body wt of AGBMAb.
polymorphonuclear leukocytes (PMN) per glomerulus wasrecorded by light microscopy. Morphologic grading wasbased on a 0-4+ scale. On light microscope examination,the glomerular capillary lumens appeared irregular with vari-
able degrees of encroachment, which is presumably relatedto the endothelial abnormalities seen by an electron micro-scope as well as to PMN infiltration. This change wasgraded according to the approximate number of capillary
TABLE IIEffect of Dose and Complement Depletion upon Pressures, Flows, Permeabilities,
and sngfr before and after the Administration of AGBMAb
MAP* PG P, AP sngfr 7WA
C E C E C E C E C E C E
mmHg mmHg mmHg mmHg nil/minig kw mmHg
Normal ratsLow dose 115 119 61 61 19 13 42 48 58 51 19 19
(1.4 ug/g body wt) +6 +411 ±3 +2 +2 ±2 ±2 +2 +3 ±4 ±1 +1P >0.1 >0.8 <0.01 <0.02 <0.001 >0.9
High dose 108 116 60 55 19 10 41 45 58 8 22 20(2.3 ,ug/g body wt) ±8 +5 ±2 ±2 +1 +1 +1 ±2 ±4 ±4 ±1 ±1
P >0.1 >0.1 <0.001 <0.05 <0.001 >0.5
Complement-depleted ratsLow dose 117 118 57 54 18 12 39 42 50 47 20 18
(1.4 ,ug/g body wt) ±4 ±4 ±2 ±2 ±1 +2 +2 +2 ±4 ±5 ±1 +1P >0.5 >0.3 <0.01 >0.3 >0.3 <0.05
High dose 109 114 58 57 18 8 40 48 55 37 22 23
(2.3 jug/g body wt) +6 +6 +3 ±2 +2 ±2 +2 ±1 ±2 +3 ±1 +2
P >0.1 >0.6 <0.01 <0.01 <0.005 >0.1
* Mean arterial pressure.4 IE, efferent arteriolar oncotic pressure.§ Superficial nephron filtration fraction.
II SEM.
912 R. C. Blantz, B. J. Tucker, and C. B. Wilson
loops involved with 1 and 2+ changes denoting 25 and50%, respectively. By electron microscopy, the degree ofendothelial separation was graded 1 + when scattered areas orsegments of separation were noted, 2+ when separationinvolved the entire circumference of capillary loops of somecapillaries, and 3+ when roughly half of the endotheliumwas partially or completely separated. Foot process fusionwas graded largely by the area involved with 1 and 2+ cor-responding roughly to 25 and 50%, respectively. PMNap-proximated to the GBMwere scored simply according toease of identification, i.e., rare, occasional, or many.
Calculations. Superficial nephron filtration fraction, rpf,afferent arteriolar resistance (AR), and efferent arteriolarresistance (ER) and oncotic pressure (X) from protein con-centration (C) were calculated as described in our previouspublications (1, 8, 9, 13-15).
The four factors that define the sngfr-hydrostatic pres-sure gradient acting across the glomerular membrane (AP),systemic oncotic pressure (ITA), glomerular permeability co-efficient (LpA), and rate of rpf (6, 7) interrelate in thefollowing manner.
Because AP = PG - Pt, where PG = directly measured glo-merular capillary hydrostatic pressure and P, = Bowman'sspace hydrostatic pressure, the effective filtration pressure(EFP) can be defined as follows: EFP = AP - r, where ir= oncotic pressure.
As a consequence of glomerular ultrafiltration, 7T risesalong the length of the glomerular capillary (x*) as a resultof the increase in protein concentration (C). The meanEFP (EFP) is defined as follows:
EFP =| (AP - ir)dx*,
where x* = normalized unit glomerular capillary length.
Changes in rpf modify the EFP profile along x* by affectingthe rate at which protein is concentrated and the rate ofrise in Xr along x*.
The sngfr can therefore be defined as follows: sngfr= LpA x EFP, where L1A is the glomerular permeabilitycoefficient and which in turn is a product of thehydraulic conductivity (L,) of the glomerular membrane and(A) the total filtering surface area of the glomerular capil-lary.
Statistical analysis. The significance of data betweencontrol and experimental states among all four groups incontrol and experimental states was determined by two- andthree-way analysis of variance and by Student's t test whereappropriate (16, 17).
RESULTS
Measurements were made both before and after AGBMAb infusion in four separate groups-low- and high-dose normal rats and low- and high-dose complement-depleted rats.
Studies utilizing either low-dose (1.4 ,g/g body wt)or high-dose (2.3 ,ug/g body wt) AGBMAb wereperformed in alternating sequence with normal andcomplement-depleted rats. The changes in urinevolume, sodium excretion, and potassium excretionafter AGBMAb infusion at both doses are summarizedin Table I. All groups except the low dose of AGBMAb in complement-depleted rats showed uniform re-ductions in excretion. Administration of AGBMAb didnot alter mean arterial blood pressure in any of thefour respective experimental groups (P > 0.1).
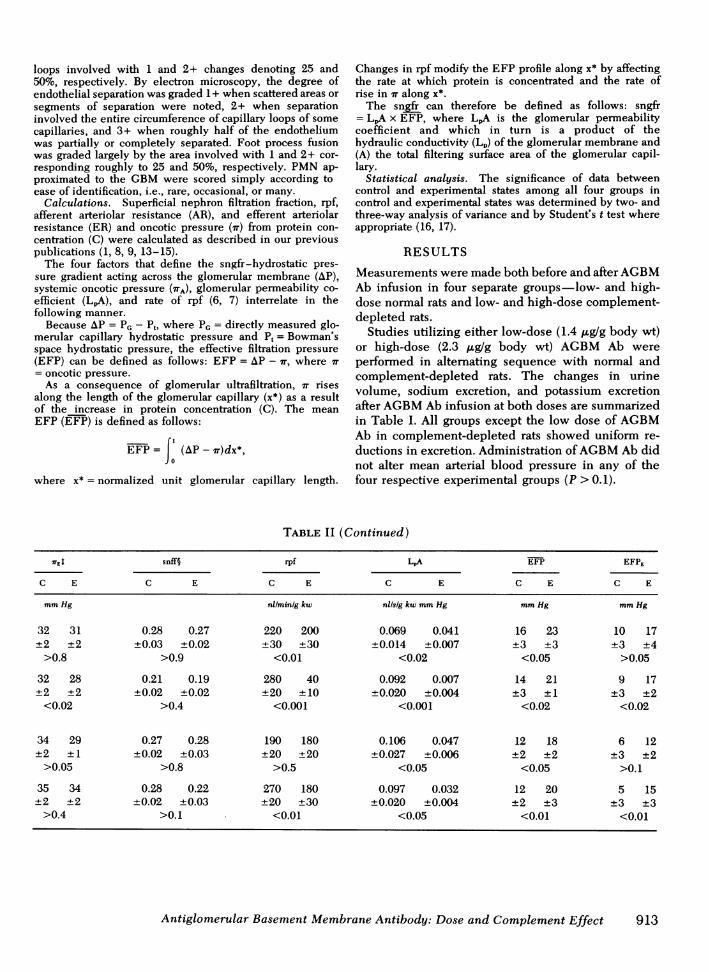
TABLE II (Continued)
7TE snff§ rpf LA EFP EFPE
C E C E C E C E C E C E
mmHg nl/min/g kw nilsig kw mmHg mmHg mmHg
32 31 0.28 0.27 220 200 0.069 0.041 16 23 10 17+2 ±2 ±0.03 ±0.02 ±30 ±30 ±0.014 ±0.007 ±3 ±3 ±3 ±4
>0.8 >0.9 <0.01 <0.02 <0.05 >0.05
32 28 0.21 0.19 280 40 0.092 0.007 14 21 9 17±2 ±2 ±0.02 ±0.02 ±20 ±10 ±0.020 ±0.004 ±3 ±1 ±3 ±2
<0.02 >0.4 <0.001 <0.001 <0.02 <0.02
34 29 0.27 0.28 190 180 0.106 0.047 12 18 6 12±2 ±1 ±0.02 ±0.03 ±20 ±20 ±0.027 ±0.006 ±2 ±2 ±3 ±2
>0.05 >0.8 >0.5 <0.05 <0.05 >0.1
35 34 0.28 0.22 270 180 0.097 0.032 12 20 5 15±2 ±2 ±0.02 ±0.03 ±20 ±30 ±0.020 ±0.004 ±2 ±3 ±3 ±3
>0.4 >0.1 <0.01 <0.05 <0.01 <0.01
Antiglomerular Basement Membrane Antibody: Dose and Complement Effect 913

Effect of dose of AGBMAb. No changes wereobserved on the kidney surface after low dose AGBMAb infusion. With the higher dose, the kidney becameless firm, and some decrease in color intensity of thesurface was noted, possibly reflecting the reducednephron blood flow. As shown in Table II, the meansngfr fell somewhat (58±3 vs. 51±4 nl/min per g kw)after a low dose of AGBMAb. This reduction wassignificantly greater at the higher dose 58±4 ascompared to 8±4 nl/min perg kw). The mean rpfdecreased minimally but significantly after the lowerdose of AGBMAb and to a significantly greaterextent after the higher dose. The mean superficialnephron filtration fraction remained unchanged at bothlow and high doses of AGBMAb.
Bowman's space hydrostatic pressure (P,) fell from19±2 to 13±2 mmHg after low-dose AGBMAbinfusion (P < 0.01; Table II). At the higher dose Ptfell from 19±1 to 10±1 mmHg (P < 0.001). The PG,however, was not changed by AGBMAb at eitherlow dose (61±3 vs. 61±2 mmHg; P < 0.8) or highdose (60±2 vs. 55±2; P < 0.1). The net hydrostaticpressure gradient (AP) increased with both low-doseAGBMAb (42±2 vs. 48±2 mmHg; P < 0.02) and high-dose infusion (41±1 vs. 45±2 mmHg; P < 0.05), sothat the net effect on sngfr was positive rather thannegative.
The afferent (AR) and efferent (ER) arteriolarresistances were unchanged (11±1 vs. 14±3 x 109dyn s/cm5; P > 0.3) and 10±1 vs. 12±2 x 109 dyns/cm5; P > 0.4), respectively, after low-dose AGBMAb infusion. However, at the higher dose AR in-creased from 8±1 to 161±70 x 109 dyn-s/cm5 (P< 0.001) and ER from 7.0±0.5 to 148±70 x 109 dyn-s/cm5 (P < 0.001).
Plasma C was identical at both the low-dose (5.8±0.2vs. 5.8±0.1 g/100 ml) and high-dose AGBMAb(6.3±0.2 vs. 5.9±0.3 g/100 ml; P > 0.05). The meansystemic oncotic pressure (7TA) was not altered by lowand high doses of AGBMAb (Table II). The hematocritswere also unchanged at either dose of AGBMAb(P > 0.1).
The EFP at the afferent end of the glomerularcapillary (EFPA) increased after administration of low-dose AGBMAb from 23±2 to 29 ± 2 mmHg (P< 0.01) and after high-dose infusion 19±2 vs. 25±1 mmHg (P < 0.05). The EFP at the efferent end of thecapillary (EFPE) increased from 10±3 to 17±4 mmHg (P < 0.05) after low-dose infusion and from 9±3to 17±2 mmHg (P < 0.02) after the higher dose.The EFP rose from 16±3 to 23±3 mm Hg (P<0.05) after low-dose AGBMAb administration andfrom 14±3 to 21±1 mmHg (P < 0.02) after the higherdose. Finding that sngfr decreased at both doses ofAGBMAb in spite of increased EFP was also ob-served in our previous study on glomerular immune
injury. This suggests that reductions in LpA werecritical to the decrease in sngfr observed at bothdoses of AGBMAb (Table II).
The mean LpA fell after low-dose AGBMAb infusionand decreased to a significantly greater extent (P< 0.01), as did sngfr, with the higher dose of AGBMAb (Table II; 0.092+0.020 vs. 0.007±0.004 nl/s per g kwmmHg) (P < 0.001). Thus, the influence of rpf canbe separated from that of LpA by varying the dose ofAGBMAb.
Effect of prior complement depletion upon low-and high-dose AGBMAb. The appearance of thekidney surface was basically unaltered in complement-depleted rats by both low and high doses of AGBMAb which suggests that high doses of AGBMAb havea lesser influence after complement depletion.
After complement depletion, low-dose AGBMAb nolonger significantly reduced sngfr: 50±4 vs. 47±5 nl/min per g kw; P > 0.3; n = 9 (Table II). At the higher dose,prior complement depletion provided an even more im-pressive protection effect: sngfr decreased from 55±2 to37±3 nl/min per g kw; P < 0.005; n = 6 (Table II), amuch lesser reduction than observed in rats with anintact complement system (P < 0.005).
The beneficial effects of complement depletion weresimilarly observed when rpf was evaluated. The rpfwas 189±17 in control and 183±21 nl/min per g kw(P > 0.5) after low-dose AGBMAb (Table II), sig-nifying that complement depletion totally preventedthe vasoconstriction observed in rats with intactcomplement given low-dose AGBMAb. Also, withhigh-dose AGBMAb, a reduction in rpf persisted,but to a much lesser extent (P < 0.005), at 209±23and 184±30 nl/min per g kw (P < 0.01), respectively.
However, P, also decreased in the low-dose, comple-ment-depleted group (P < 0.01; Table II) and in thehigh-dose complement-depleted group. PG was un-changed by low-dose AGBMAb infusion in thecomplement-depleted group as well. Similarly, high-dose AGBMAb did not alter PG. As a result, AP wasnow unchanged by the lower dose but increasedafter the high-dose Ab infusion.
AR was unchanged in the low-dose group (17±2 vs.23±4 x 109 dyn s/cm5; P > 0.3) and at the higherdose (12+2 vs. 16±2 x 109 dyn s/cm5; P > 0.1). ERwas also unchanged at the lower dose (15±3 vs. 18±5x 109 dyn s/cm5) and higher dose (11±2 vs. 14±2x 109 dyn s/cm5; P > 0.1).
Plasma C (CA) was reduced slightly at the lowerdose (6.0±0.2 vs. 5.5±0.3 g/100 ml; P < 0.05) but wasunchanged at the higher dose (6.2±0.3 vs. 6.5±0.3 g/100 ml; P > 0.2). Similarly, the IrA was reducedslightly in the low-dose group (Table II) but wasunchanged at the higher dose.
The initial driving force for ultrafiltration was in-creased slightly at the lower dose (19±2 vs. 23±1 mm
914 R. C. Blantz, B. J. Tucker, and C. B. Wilson

Hg; P < 0.05) and to a somewhat greater extent at thehigher dose (19±2 vs. 25+3 mmHg; P <0.01). TheEFP at the efferent end of the capillary was 6±3 mmHg in control and 12±2 mmHg (P > 0.1) after low-dose infusion; it increased from 5±3 to 15±3 mmHg(P < 0.01) after the higher dose of antibody. Althoughsngfr remained constant, the EFP increased at thelower dose in the complement-depleted group (12±2vs. 18±2 mmHg; P < 0.05). At the higher dose, theEFP, again increased from 12±2 to 20±3 mmHg(P < 0.01).
Although sngfr remained constant, the reduction inLpA persisted with lower dose AGBMAb infusion(0.106+0.027 vs. 0.047±0.006 nl/s per g kw mmHg;P < 0.05) (Table II). The persistent decrease in LpAwith no change in sngfr suggests that the beneficialeffects that complement depletion exerted upon thelow-dose group was mediated by preventing thevasoconstriction observed with AGBMAb in the groupwith an intact complement system. The increase inEFP which permitted constant sngfr resulted fromreduced ITA, decreased LpA, and a numerical in-crease in AP. At the higher dose, complementdepletion significantly diminished the extent to whichLpA was reduced (0.097±0.020 vs. 0.032±0.004 nl/sper g kw * mmHg; P < 0.05). In fact, the L,A after high-dose AGBMAb infusion in the complement-depletedgroup was not different from the experimental LpA inboth low-dose groups (P > 0.05) (Table II).
In summary, complement depletion prevented thereduction in sngfr in the low-dose AGBMAb groupprimarily by preventing a decrease in rpf, but didnot affect the antibody-mediated reduction in LpA. At
the higher dose, the decrease in sngfr persisted inspite of complement depletion, but to a much lesserdegree. This beneficial effect of complement deple-tion at the higher dose resulted from a lesser degree ofvasoconstriction and a decrease in the magnitude ofreduction in LpA. However, at both doses in normaland complement-depleted rats, there remains a per-sistent reduction in LpA after AGBMAb administra-tion that appears independent of the complementsystem.
Immunofluorescence and morphologic findings.Immunofluorescence study revealed that all rats hadtypical heavy, linear deposits of rabbit IgG alongtheir GBM (Fig. 1). The deposits of rabbit IgGwere perhaps more prominent in the high-dosegroup, but the differences could not be clearlydistinguished among dosage groups by immuno-fluorescence. Rat C3 accompanied the rabbit IgG inthe normal complementemic animals (Fig. 1). Glo-merular C3 deposits were virtually absent in thecomplement-depleted rats (Fig. 1).
Morphologic changes that accompanied AGBMAbinfusion were similar but quantitatively different forthe four groups of rats, and there was only minorintragroup variation (Table III). The high-dose nor-mal complementemic group was the most severelyaffected. With light microscopy, PMNs could bedetected easily within the glomerular capillarylumina, where as many as 15 were present perglomerulus (Fig. 2). The endothelial cytoplasm ap-peared irregular which, combined with the PMNs,appeared to compromise the glomerular capillarylumina; however, the extent of compromise was dif-
FIGURE 1 The typical linear- deposits of rabbit AGBMAb IgG are shown along the GBMof a rat from a low-dose CVF-treated group in A. In B, the irregular linear deposits ofC3 that accompanied the IgG in normal complementemic rats are seen. The virtualabsence of C3 found in the complement-depleted CVF-treated rats is visualized in C.(Fluorescein isothiocyanate conjugated anti-rabbit IgG in A, and anti-rat C3 in B and C. x312.)
Antiglomerular Basement Membrane Antibody: Dose and Complement Effect 915

TABLE IIIIndividual Correlations of Physiologic and Morphologic Changes with AGBMAb Infusion at Low
and High Doses in Normal and Complement-Depleted Rats
Morphologic changes
Light microscopy Electron microscopyPhysiologic changes
Capillary Endo- Focalsngfr rpf PMN irregularity thelial foot
LpA glomer- and sepa- PMN processRat control change control change change ulus narrowing* rationt on GBM fusion§
num- nl/minig ±% nl/min/g ±% ±%ber kw kw
Normal ratsLow-dose group 1 47 0 143 +10 -40 0-3 tr-+"1 1-2+ Occasional tr-+
2 65 -23 251 -20 -44 1-5 tr- + 1-2+ Moderate tr- +6 68 -15 275 -4 -41 0-4 tr- + 2+ Occasional tr- +9 62 +7 158 +91 -62 2-5 tr- + 1-2+ Many +
11 55 -26 305 -61 -26 2-8 tr 2+ Many +15 50 -12 194 -21 0 1-6 tr- + 1-2+ Moderate +
High-dose group 3 49 -23 262 -61 -86 ND¶ ND 1-2+ Many tr-+4 54 -49 220 -86 -94 3-8 1-2+ 1-2+ Many +5 75 -70 277 -91 -95 3-10 2+ 1-2+ Many +7 51 -46 337 -85 -89 3-10 2+ 1-2+ Many tr-+
10 63 -60 299 -96 -96 5-15 2+ 2-3+ Many 1-2+13 58 -57 261 -97 -99 5-15 2+ 2-3+ Many 1-2+
Complement-depletedrats
Low-dose group 1 49 -30 182 +11 -83 0-1 tr 1-2+ 0 tr4 34 -38 140 -69 -21 0-4 0 1-2+ Occasional tr- +6 43 +17 135 +30 +13 0-2 0 tr-+ 0 tr8 39 +26 205 -4 +8 0-2 0 tr- + Rare tr9 76 -19 304 -16 -72 0-2 0 1-2+ 0 tr- +
12 54 +19 163 +22 +20 0-2 0 tr 0 tr14 42 -11 201 -30 -38 0-2 0 1+ 0 tr15 57 -6 204 -10 -42 0-2 0 1-2+ Rare tr- +16 54 0 164 +57 -76 0-3 0 1-2+ Rare +
High-dose group 3 59 -27 236 -16 -49 0-3 tr 1-2+ 0 tr5 64 -48 227 +33 -77 0-4 tr 1-2+ Occasional tr- +7 55 -54 211 -37 -75 0-3 tr ND ND ND
10 55 -38 161 -28 -77 0-3 tr 2+ Rare tr- +11 53 -7 288 -21 -26 0-4 tr 1-2+ 0 tr- +13 46 -16 132 -2 -43 0-3 tr 1-2+ Occasional tr- +
* Graded according to the approximate number of capillary loops involved with 1 and 2+ changes denoting 25 and 50%,respectively.I Graded 1+ when scattered areas or segmental separation were noted, 2+ when separation involved the entire circumferenceof some capillary loops, and 3+ when partial or complete involvement was present in roughly half of the capillary lumens.§ Graded according to extent with 1 and 2+ corresponding roughly to 25 and 50%, respectively.'tr, trace.¶ ND, not done.
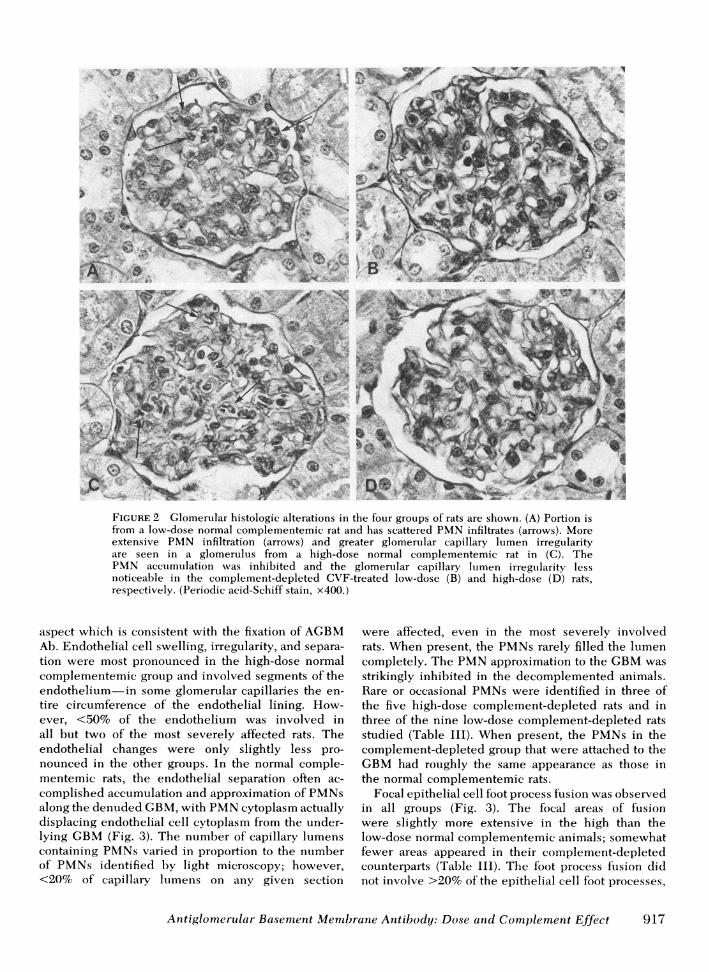
ficult to estimate. Similar but less extensive changes No extraglomerular or vascular histologic alterationswere observed in the low-dose normal complemente- were noted.mic group (Fig. 2). The complement-depleted rats Endothelial abnormalities, approximation of PMNwere virtually free of PMNaccumulation in compari- along the GBM, and focal foot process fusion wereson with their complementemic counterparts, and had observed in various combinations by electron micros-less noticeable endothelial irregularity and little copy (Table III). The GBM itself appeared some-compromise in their capillary lumens (Fig. 2 B and D). what irregular, particularly on its subendothelial
916 R. C. Blantz, B. J. Tucker, and C. B. Wilson
a @swNo N tttt Ws-*
<e
.s. a>... .. *t..
T..s d.. , *
.. ... }\E
v bs XC:.,,,4 / ..
...^ ; -p-\ g; ....* ;\t t
2 .'\.ssV{q
\k--.yS. j,,/ 't/r
FIGuRE 2 Glomerular histologic alterations in the four groups of rats are shown. (A) Portion isfrom a low-dose normal complementemic rat and has scattered PMNinfiltrates (arrows). Moreextensive PMN infiltration (arrows) and greater glomerular capillary lumen irregularityare seen in a glomerulus from a high-dose normal complementemic rat in (C). ThePMN accumulation was inhibited and the glomerular capillary lumen irregularity lessnoticeable in the complement-depleted CVF-treated low-dose (B) and high-dose (D) rats,respectively. (Periodic acid-Schiff stain, x400.)
aspect which is consistent with the fixation of AGBMAb. Endothelial cell swelling, irregularity, and separa-tion were most pronounced in the high-dose normalcomplementemic group and involved segments of theendothelium-in some glomerular capillaries the en-tire circumference of the endothelial lining. How-ever, <50% of the endothelium was involved inall but two of the most severely affected rats. Theendothelial changes were only slightly less pro-nounced in the other groups. In the normal comple-mentemic rats, the endothelial separation often ac-complished accumulation and approximation of PMNsalong the denuded GBM,with PMNcytoplasm actuallydisplacing endothelial cell cytoplasm from the under-lying GBM(Fig. 3). The number of capillary lumenscontaining PMNsvaried in proportion to the numberof PMNs identified by light microscopy; however,<20% of capillary lumens on any given section
were affected, even in the most severely involvedrats. When present, the PMNs rarely filled the lumencompletely. The PMNapproximation to the GBMwasstrikingly inhibited in the decomplemented animals.Rare or occasional PMNs were identified in three ofthe five high-dose complement-depleted rats and inthree of the nine low-dose complement-depleted ratsstudied (Table III). When present, the PMNs in thecomplement-depleted group that were attached to theGBMhad roughly the same appearance as those inthe normal complementemic rats.
Focal epithelial cell foot process fusion was observedin all groups (Fig. 3). The focal areas of fusionwere slightly more extensive in the high than thelow-dose normal complementemic animals; somewhatfewer areas appeared in their complement-depletedcounterparts (Table III). The foot process fuision didnot involve >20% of the epithelial cell foot processes,
Antiglomerular Basement Membrane Antibody: Dose and Complement Effect 917

EP
e., 4f N
jt-.*' * .e J*. ; f_f i.4~~~~ z ~-w;
3/
i
-z :~~~~~~h
S. %i //E# / - Xs
xi f, t
r F
t si, tS +
; , .JS +P_ .-;
.; _\:, . /. e rr.,, ,,Y..,,,,$_ ft
¢;
. C1 4.g s*-< gs-
df ;; s; s-,- t ,>/ j
{'. {wo... M
" b('_zla;,'1_, i. , . ' .
_ I bVi J ta'
.J i. Iutt^f 1g B I 12m
FIGuRE 3 Electron microscope studies of a high-dose normal complementemic rat (A) and acomplement-depleted CVF-treated rat (B) are shown. The subendothelial aspect of the GBM(arrows) appeared irregular, presumably representing the fixation of anti-GBM antibody.A PMN was found approximated along the GBM displacing the endothelium (EN) inthe normal complementemic rat in IL. Endothelial separation occurred in the complement-depleted rat seen in B; however, PMN infiltration was almost completely lacking. Focalareas of epithelial cell (EP) foot process fusion were found and were particularly evidentin A. (x 17,750.)
except in the two most severely affected high-doserats. This finding was not clearly related to thepresence of PMNs.
Electron microscope studies revealed minor individ-ual variations within animals of each group. Eventhough portions of four glomeruli were studied ineach rat, the possibility of sampling error was feltto preclude meaningful, detailed animal to animalcorrelation. It is interesting, however, that the threerats in which LpA did not decrease had the leaststriking endothelial change in the low-dose comple-ment-depleted group.
DISCUSSION
Micropuncture techniques permit direct evaluation ofglomerular fluxes, flows, pressures, and permeabilitycoefficients and can be usefully applied as quantita-tive assay tools to examine the series of factors andevents that lead to the altered glomerular filtrationthat follows immune injury (1). Previous studiesutilizing these tools have demonstrated that within1 h after immune insult, the glomerular filtration ratefalls because of both alterations in the LpA andincreases in renal vascular resistance (1). To gain
918 R. C. Blantz, B. J. Tucker, and C. B. W'ilson
:' i;4 :-A!. I
Ic-I
.. I ...I .-I !.;,.J

further insights into the exact mechanisms involvedin this early phase of immune injury, we have deter-mined whether these two events leading to reducedsngfr are intimately linked or whether vasoconstrictionand permeability effects are separable mechanisms.Utilizing differing doses of AGBMAb, we have beenable to separate quantitatively these two mechanismsand their respective influence upon filtration. At higherdoses of AGBMAb, massive reductions in both LpA andrpf occurred, and both factors contributed to the largedecrease in sngfr. However, at the lower dose, al-though a minimal vasoconstrictive effect occurred, asignificant reduction in LpA was largely responsiblefor the decrease in filtration rate.
Although the qualitative conclusions that decreasesin both rpf and LpA contribute to reduced filtra-tion rate in the early phase of immune injury arecommon to this and our previous study on thisissue (1), there were significant quantitative dif-ferences observed between the antibody preparationsutilized. Doses of 225 and 450 ,ug of AGBMAb inour previous study produced similar quantitative ef-fects on rpf, LpA, and sngfr. However, the cur-rent microtechniques have permitted us to determinethat these same low and high doses resulted inquantitative differences after the antibody utilized inthis study. The specific reasons for the quantitativedifferences between antibody preparations generatedto the same antigen is not known, but may relateto secondary effects such as the respective capacitiesof antibody preparations to fix complement. Also,similar high doses of antibody preparations led todifferent degrees of PMNaccumulation, also possiblyrelated to the complement activity generated by therespective preparations at identical doses adminis-tered.
The fixation of complement is critical in the processof immune injury. The studies of Cochrane et al. (2),and reviews of Muller-Eberhard (18), and Cochraneand Janoff (19) have stressed the role of complementin the pathogenesis of experimental glomerulonephri-tis. Activation of the complement cascade has multipleeffects, including the attraction of PMN, the enhance-ment of cellular adherence, and the activation andrelease of a variety of biologically active substances.Judgments as to the role of complement fixationhave been based primarily upon evaluating the renalmorphology and the proteinuria that follow immunechallenge (2, 18). No studies have evaluated the pos-sible beneficial effects upon glomerular ultrafiltrationand the respective determinants of filtration that mightresult from preventing complement fixation.
The present studies in the complement-depletedrat and ultrastructural analysis of all experimentalgroups have provided information on the mechanisms
leading to vasoconstriction and reduced LpA and thespecific role of the complement system in the processof immune injury. CVF, which appears to be cobraC3B (20), interacts with factors B and D of the alterna-tive complement pathway to form a C3 cleavingenzyme (7), thereby causing C3 depletion. Comple-ment depletion largely prevented the acute vaso-constriction that occurred at both doses of AGBMAb.Inasmuch as the vasoconstriction was probably func-tional, results from studies on complement depletionsuggest that vasoconstriction was not due to acutefixation of antibody alone. It is possible that activa-tion of the complement system causes the local releaseof intrarenal substances capable of mediating constric-tion of resistance vessels. It is also possible thatcertain components of the complement cascade mayhave vasoconstrictor activity (18). Complement de-pletion also prevented migration of PMN into theglomerular capillary, but this complement-mediatedevent is unlikely to be the direct cause of increasedvascular resistance, since the cells accumulated onlywithin the capillary, normally a region of verylow resistance in the renal vasculature (15). Ourprevious study on glomerular immune injury alsoprovided evidence that the decrease in rpf andmigration of PMNs were separate effects of comple-ment activation. With a different AGBMAb, sig-nificant vasoconstriction occurred but fewer PMNswere noted within the capillary 1 h after the in-fusion of the complement-fixing AGBMAb thanwas noted in the high dose group in this study (1).Although vasoconstriction and migration of PMNsaregreatly dependent upon complement fixation, the twoevents are presumably the unrelated effects of comple-ment activation. Where individual rats were studied,the degree of infiltration of the capillaries with PMNscorrelated strongly with the degree of rpf reduction(both presumably effects of complement fixation; TableIII). We cannot exclude the possibility that PMNssomehow contribute directly to the increase in vascularresistance.
Prevention of complement activation also had a majoreffect upon LpA. With the higher dose of AGBMAb,complement depletion greatly decreased the effect ofantibody upon LpA. In the normal rat, LpA wasreduced 92% after high-dose AGBMAb; this effectwas diminished to 67% in the complement-depleted,high-dose group. This beneficial effect of complementdepletion correlated with the prevention of PMNsmigrating into the capillary. Although the effectsof complement activation upon PMNs and vaso-constriction appear to be unrelated events, we believethat there is reasonable evidence that PMNs maymediate in part the larger LpA reduction observedwhen normal rats were injected with high doses of
Antiglomerular Basement Membrane Antibody: Dose and Complement Effect 919

AGBMAb. At high doses, PMNs reduced capillarysurface to a small extent in normal complementemicrats by adherence, but in addition may have had agreater effect by partially obstructing certain capillaryconduits within the glomerulus, diverting glomerularblood flow through fewer channels. This latter effectis impossible to evaluate quantitatively but shouldreduce effective filtering surface area without con-tributing measurably to increased vascular resistancebecause the capillary normally contributes little tototal renal vascular resistance. At lower doses of anti-body, there was no beneficial effect of complementdepletion upon the reduction in LpA, but therewere also fewer PMNs within the capillary at thisdose. In addition, at the higher dose, LpA remainedlow in spite of complement depletion. We thereforeconclude that complement depletion partially amelio-rates the effect of a high dose of antibody upon LpAby preventing any loss of capillary surface arearesulting from migration of PMNsinto the capillary.
AGBMAb fixation exerts a significant and persistenteffect upon LpA that appears independent of the com-plement system. The quantitative reductions in LpAafter antibody infusion in low-dose, in low-dosecomplement-depleted, and in high-dose complement-depleted groups were not significantly different (TableII). This persistent reduction in LpA did not resultfrom loss of surface area secondary to the adherenceof PMNs but was associated with structural altera-tions upon the endothelial surface of the glomerularmembrane and, to a lesser extent, with focal fusion ofepithelial cell foot processes. The detachment of endo-thelial cells, an effect noted in all groups (and in aprevious study; 1), appears to be the logical cause ofthis persistent decrease in LpA within 1 h of AGBMAbinfusion. The causes of reduction in LpA are additive.The first is complement dependent and related to lossof capillary surface area (A) from PMN accumula-tion; the second is independent of the complement sys-tem and due to structural alterations upon the endo-thelial surface of the capillary membrane which leads toreductions in local capillary permeability (Lp).
In a previous study on glomerular immune injury, wehave postulated that fixation of antibody globulin tothe GBMmay have produced separation of the endo-thelial cell from the underlying basement membraneby interfering with the potentially charge-dependentattachment of these structures (1). This endothelialseparation may lead to a relatively less "well-stirred" compartment in which serum proteins maybecome concentrated from the lack of "stirring"effect of capillary blood flow. Although we can becertain that much of the alteration in capillary permea-bility (Lu) is independent of complement activation,the data do not permit us to conclude that endothelialchanges result from antibody fixation alone. It remains
possible that other mediators of immune injury andthe steric effects of AGBM Ab fixation are in-dependent of the complement cascade and may have apart in producing the endothelial injury.
There may be other mechanisms which furtherdecrease LpA and affect glomerular filtration at longerintervals after immune injury. Ultrastructural andproliferative cellular changes (21) could contribute tofurther reductions in LPA at a later time, and therole of the complement system in this process is lesswell defined.
It is difficult to extrapolate the results of the cur-rent study on the effects of differing doses of AGBMAb to the clinical state associated with AGBMAb-induced nephritis. Although large amounts of AGBMAb can be demonstrated within the glomeruli of pa-tients with this form of nephritis (22, 23), it is notlikely that such a large quantity of AGBMAb aswas utilized in this study (with respect to weightof the animal) could be generated into the circulationin such a short time period in the clinical condition.It is, therefore, not likely that vasoconstriction is thedominant mode of the initial glomerular filtration ratereduction in the analogous clinical condition but ratherthe reduction in LpA. Also the present studies havenot defined that specific substances that mediate vaso-constriction. Histologic and immunofluorescent ex-amination of renal tissue revealed neither structuralalterations nor immunoglobulin deposition withinresistance vessels, suggesting that vasoconstriction re-sulted from functional causes rather than structuralalterations.
The application of micropuncture techniques to theevaluation of glomerular immune injury provides aquantitative assessment of substances that mediatedecreased sngfr and the extent of their influence. Thecurrent study demonstrates that the vasoconstrictionobserved is a phenomenon associated with large dosesof antibody and, in the acute condition, requires anintact complement system. Mechanisms leading to re-duced filtration rate secondary to immune injury canbe further separated into complement-dependent ef-fects and into noncomplement-dependent mechanismsrelated to altered capillary hydraulic permeabilityand to changes in the ultrastructure of the glomeru-lar membrane.
ACKNOWLEDGMENTS
We extend our appreciation to Ms. Ann Chavez for herexcellent secretarial assistance and to Leslie Gushwa forher excellent technical assistance. Wealso thank Dr. CharlesCochrane of Scripps Clinic and Research Foundation for hisadvice.
These studies were supported through grants and contractsfrom the National Institutes of Health (HL 14914, Al 07007,and AM20043) and from the Veterans Administration MedicalResearch Service.
920 R. C. Blantz, B. J. Tucker, and C. B. Wilson

REFERENCES
1. Blantz, R. C., and C. B. Wilson. 1976. Acute effectsof antiglomerular basement membrane antibody on theprocess of glomerular filtration in the rat. J. Clin.Invest. 58: 899-911.
2. Cochrane, C. G., E. R. Unanue, and F. J. Dixon. 1965. Arole of polymorphonuclear leukocytes and complement innephrotoxic nephritis. J. Exp. Med. 122: 99-116.
3. Krakower, C. A., and S. A. Greenspon. 1951. Localizationof the nephrotoxic antigen within the isolated renalglomerulus. Arch. Pathol. 51: 629-639.
4. Unanue, E. R., and F. J. Dixon. 1965. Experimentalglomerulonephritis. V. Studies on the interaction ofnephrotoxic antibodies with tissues of the rat. J.Exp. Med. 121: 697-714.
5. Pressman, D., E. D. Day, and M. Blau. 1957. The use ofpaired labelling in the determination of tumor-localizingantibodies. Cancer Res. 17: 845-850.
6. Ballow, M., and C. G. Cochrane. 1969. Two anti-complementary factors in cobra venom: hemolysis ofguinea pig erythrocytes by one of them. J. Immunol.103: 944-952.
7. Gotze, O., and H. J. Muller-Eberhard. 1977. The alterna-tive pathway of complement activation. Adv. Immunol.24: 1-35.
8. Blantz, R. C. 1974. Effect of mannitol upon glomerularultrafiltration in the hydropenic rat. J. Clin. Invest.54: 1135-1143.
9. Blantz, R. C., F. C. Rector, Jr., and D. W. Seldin. 1974.Effect of hyperoncotic albumin expansion upon glomeru-lar ultrafiltration in the rat. Kidney Int. 6: 209-221.
10. Lowry, 0. H., N. J. Rosebrough, A. I. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193: 265-275.
11. Allison, M. E. M., C. B. Wilson, and C. W. Gottschalk.1974. Pathophysiology of experimental glomerulonephri-tis in rats. J. Clin. Invest. 53: 1402-1423.
12. Wilson, C. B. 1976. Immunohistopathology of the kidney.
In Manual of Clinical Immunology. H. R. Rose and H.Friedman, editors. American Society of Microbiology,Washington, D. C. 692-700.
13. Blantz, R. C. 1975. The mechanism of acute renalfailure after uranyl nitrate. J. Clin. Invest. 55: 621-635.
14. Blantz, R. C., K. S. Konnen, and B. J. Tucker. 1975.Glomerular filtration response to elevated ureteralpressure in both the hydropenic and plasma-expandedrat. Circ. Res. 37: 819-829.
15. Blantz, R. C., K. S. Konnen, and B. J. Tucker. 1976.Angiotensin II effects upon the glomerular microcircula-tion and ultrafiltration coefficient of the rat. J. Clin.Invest. 57: 419-434.
16. Bliss, C. I. 1967. Statistics in Biology. Vols. I and II.McGraw-Hill Book Company, New York.
17. Dunn, 0. J., and V. A. Clark. 1974. Applied Statistics:Analysis of Variance and Regression. John Wiley & Sons,Inc., New York. 221-304.
18. Muller-Eberhard, H. J. 1974. The complement systemand nephritis. In Adv. Nephrol. 4: 3-15.
19. Cochrane, C. G., and Janoff. 1974. The Arthus reaction.A model of neutrophil and complement mediated injury.In The Inflammatory Process. Vol. III. B. W. Zweifach,L. Grant, and R. T. McCluskey, editors. Academic Press,Inc., New York. 2nd edition. 85-162.
20. Alper, C. A., and D. Balavitch. 1976. Cobra venom factor:evidence for its being altered cobra C3 (the 3rd com-ponent of complement). Science (Wash. D. C.). 191:1275-1276.
21. Maddox, D. A., J. L. Troy, C. R. Robertson, and B. M.Brenner. 1973. Dymanics of glomerular ultrafiltration inthe rat. IV. Determination of the ultrafiltration coefficient.J. Clin. Invest. 52: 1500-1508.
22. Wilson, C. B., and F. J. Dixon. 1973. Anti-glomerularbasement membrane antibody induced glomerulonephri-tis. Kidney Int. 3: 74-89.
23. McPhaul, J. J., Jr., and J. D. Mullins. 1976. Glomerulo-nephritis mediated by antibody to glomerular basementmembrane: immunological, clinical, and histopatho-logical characteristics. J. Clin. Invest. 57: 351-361.
Antiglomerular Basement Membrane Antibody: Dose and Complement Effect 921