antiaging glycopeptide protects human islets against...
TRANSCRIPT

Boris L. Gala-Lopez,1,2 Andrew R. Pepper,1,2 Rena L. Pawlick,1 Doug O’Gorman,3
Tatsuya Kin,3 Antonio Bruni,1,2 Nasser Abualhassan,1,2 Mariusz Bral,1
Austin Bautista,1,4 Jocelyn E. Manning Fox,1,4 Lachlan G. Young,5
Patrick E. MacDonald,1,4 and A.M. James Shapiro1,2,3
Antiaging Glycopeptide ProtectsHuman Islets Against Tacrolimus-Related Injury and FacilitatesEngraftment in MiceDiabetes 2016;65:451–462 | DOI: 10.2337/db15-0764
Clinical islet transplantation has become an establishedtreatment modality for selected patients with type 1diabetes. However, a large proportion of transplantedislets is lost through multiple factors, includingimmunosuppressant-related toxicity, often requiringmore than one donor to achieve insulin independence.On the basis of the cytoprotective capabilities of antifreezeproteins (AFPs), we hypothesized that supplementation ofislets with synthetic AFP analog antiaging glycopeptide(AAGP) would enhance posttransplant engraftment andfunction and protect against tacrolimus (Tac) toxicity. Invitro and in vivo islet Tac exposure elicited significant butreversible reduction in insulin secretion in bothmouse andhuman islets. Supplementation with AAGP resulted in im-provement of islet survival (Tac+ vs. Tac+AAGP, 31.5% vs.67.6%, P < 0.01) coupled with better insulin secretion(area under the curve: Tac+ vs. Tac+AAGP, 7.3 vs. 129.2mmol/L/60 min, P < 0.001). The addition of AAGP reducedoxidative stress, enhanced insulin exocytosis, improvedapoptosis, and improved engraftment in mice by decreas-ing expression of interleukin (IL)-1b, IL-6, keratinocytechemokine, and tumor necrosis factor-a. Finally, trans-plant efficacy was superior in the Tac+AAGP group andwas similar to islets not exposed to Tac, despite receivingcontinuous treatment for a limited time. Thus, supplemen-tation with AAGP during culture improves islet potencyand attenuates long-term Tac-induced graft dysfunction.
Islet transplantation outcomes have improved signifi-cantly in highly specialized centers, with a 5-year insulin
independence rate exceeding 50% (1). However, a pro-portion of patients require reintroduction of insulin atdelayed time points (2). Numerous factors contribute tolimited durability of both short- and long-term glycemiccontrol. Exposure to diabetogenic immunosuppressiveagents is associated with islet functional impairmentand graft loss, especially that linked to corticosteroidexposure or use of calcineurin inhibitors (CNIs), includ-ing tacrolimus (Tac) (3,4). Tac is the most potentmainstay immunosuppressant in most clinical protocols,although diabetogenicity is well documented, accountingfor an increased incidence of posttransplant diabetesmellitus (5–8). Early peak Tac levels in portal blood afterislet transplantation after oral administration may fur-ther increase islet exposure, thereby magnifying toxicity(9,10). Although undesirable, Tac has become necessaryto preventing rejection and autoimmune recurrence afterislet transplantation and may be partly responsible forlimited islet durability and a need for multiple donorsfor each recipient (11).
Antifreeze proteins (AFPs) have generated consider-able interest for their ability to protect cells under avariety of conditions. They naturally occur in Arctic andAntarctic fish as well as in other cold climate–dwellinginvertebrates and are responsible for maintaining celland tissue function at subzero temperatures (12,13).AFPs were successfully isolated in the 1950s and havedemonstrated an ability to noncolligatively lower thefreezing temperature of body fluids by binding to icecrystals (12,14). Early experiments with AFPs in the
1Alberta Diabetes Institute, University of Alberta, Edmonton, Alberta, Canada2Canadian National Transplant Research Program, University of Alberta, Edmonton,Alberta, Canada3Clinical Islet Transplant Program, University of Alberta, Edmonton, Alberta, Canada4Department of Pharmacology, University of Alberta, Edmonton, Alberta, Canada5ProtoKinetix Inc., Vancouver, Vancouver, British Columbia, Canada
Corresponding author: A.M. James Shapiro, [email protected].
Received 8 June 2015 and accepted 10 November 2015.
© 2016 by the American Diabetes Association. Readers may use this article aslong as the work is properly cited, the use is educational and not for profit, andthe work is not altered.
Diabetes Volume 65, February 2016 451
ISLETSTUDIES

field of organ and tissue transplantation showed prom-ising results, making them attractive therapeutic can-didates to protect cells against harmful conditionsassociated with procurement, preservation, and reperfu-sion (14). Moreover, benefits have been demonstratedduring cryopreservation of various cells, including isletsof Langerhans, with significant improvements in theirviability and function when supplemented with AFPduring cryostasis (15,16).
Antiaging glycopeptide (AAGP) is a 580-Da syntheticAFP analog initially developed by Géraldine Castelot-Deliencourt-Godefroy (Rouen, France) and later manu-factured by ProtoKinetix Inc. (Vancouver, BC, Canada).This new compound has improved stability, is water sol-uble, and has proven to be more potent in terms ofcytoprotective capabilities under extreme conditions(pH variations, sudden temperature changes, nutrientdeprivation, oxidative stress, ultraviolet radiation, andinflammation) (17).
In light of this evidence, significant attention is nowbeing directed toward AFPs and their potential use inreparative and regenerative medicine, particularly in thefield of transplantation. We evaluated the cytoprotectivecapacity of AAGP to protect against the diabetogeniceffect of Tac, resulting in improved islet engraftment.
RESEARCH DESIGN AND METHODS
Human Islet Isolation, Purification, and CultureHuman islet preparations were isolated from consentingdeceased multiorgan donors, as previously described (18),with intent for clinical transplantation and were onlymade available for research when the islet yield fell belowthat of the minimal mass required. Permission for thesestudies was granted by the Health Research Ethics Boardof the University of Alberta, Edmonton, AB, Canada, andafter written permission from donor families.
In Vitro Evaluation of AAGP
Islet Recovery, Viability, and Secretory FunctionExperiments with human islets in vitro included fourgroups: 1) control (islets cultured in medium alone)(Tac2), 2) islets cultured in medium containing AAGP(AAGP), 3) islets cultured in medium containing Tac(Prograf; Astellas Pharma Canada Inc., Markham, ON,Canada) (Tac+), and 4) islets cultured in medium supple-mented with AAGP (ProtoKinetix) and Tac (Tac+AAGP).Islets were cultured for 24 h with63 mg/mL AAGP beforeaddition of Tac at a clinically relevant concentration of10 ng/mL. All four groups were then cultured for anadditional 24 h.
Islets were assessed for recovery, viability, insulinrelease, oxidative stress, and cell death. Recovery ratewas calculated as the percentage of surviving islets afterculture compared with the initial count for each condi-tion. Viability was assessed by fluorescent membraneintegrity assay with counterstains using SYTO 13 greenfluorescent nucleic acid stain (Life Technologies, Burlington,
ON, Canada) and ethidium bromide (Sigma-Aldrich, ON,Canada) (19–22).
Hormonal islet secretory function was assessed byboth static glucose-stimulated insulin secretion (s-GSIS),sequentially performed at low (2.8 mmol/L) and high(16.7 mmol/L) glucose concentrations and by dynamic isletperifusion (D-IP), as described by Cabrera et al. (23).D-IP was performed at 16-min intervals by using low(2.8 mmol/L) then high 3 2 (28 mmol/L) followed by lowglucose concentrations. Glucose was infused at 100 mL/min,and results are expressed as fold change of insulin secre-tion compared with the low-glucose–stimulation baselinenormalized for 100 islet equivalents (IEQ). For s-GSIS,insulin concentrations in supernatants were measuredby ELISA (Mercodia, Uppsala, Sweden). A stimulationindex was calculated as the ratio of stimulated to basalinsulin secretion normalized by DNA. These insulinsecretion studies were always performed in vitro oncultured human islets.
Apoptosis was assayed by determining the quantity ofcleaved caspase-3 in the frozen lysates from fixed isletscultured under the aforementioned conditions by using aspectrophotometric assay (EMD Millipore, Billerica, MA).Results are expressed as fold-change increase comparedwith control.
Islet apoptosis was also examined by TUNEL staining(DeadEnd Colorimetric Apoptosis Detection System;Promega, Madison, WI) after formalin fixation, process-ing, and paraffin embedding. Costaining with insulin(1:200 concentration of anti-insulin antibody [Dako,Mississauga, ON, Canada] and DAPI [Invitrogen Molec-ular Probes, Eugene, OR]) was performed to identifygrafts and nuclei, respectively. ImageJ version 1.33 softwarewith Cell Counter plug-in (http://rsb.info.nih.gov/ij) wasused to quantify islet apoptosis by percentage of positiveTUNEL-stained area.
Reactive Oxygen Species AnalysisFrozen samples from the study groups were assayed forreactive oxygen species (ROS) released into the culturemedia by Acridan Lumigen PS-3 assay (Amersham ECLPlus Kit; Fisher Scientific Inc., Ottawa, ON, Canada) (24).Acridan Lumigen PS-3 is excited by ROS and reactivenitrogen species in the presence of hydrogen peroxide,producing chemiluminescense at 430 nm. Media sampleswere flash frozen in liquid nitrogen and stored untilanalyzed. Connaught Medical Research Laboratories cul-ture medium alone served as control, and results wereexpressed as fold-change increase compared with control.
Mixed Lymphocyte ReactionTo rule out direct drug inhibition of AAGP and Tac, a one-way mixed lymphocyte reaction (MLR) assay was per-formed to assess proliferative response of responder Tcells against antigens present on allogeneic stimulatorcells. T cells were isolated from g-irradiated splenocytes ofBALB/c mice (stimulators) and C57BL/6 mice (responders).
452 AAGP Protects Transplanted Human Islets Diabetes Volume 65, February 2016

Proliferation was measured by loss of fluorescence in-tensity by using fluorochrome 5,6-carboxyfluoresceindiacetate succinimidyl ester (Invitrogen), which spon-taneously binds to intracellular proteins shared be-tween daughter cells. Controls (Tac2, AAGP2), Tac+,and Tac+AAGP groups were studied. T-cell proliferationwas measured by flow cytometry after characterizingdifferent subpopulations by cell surface antibody stain-ing with anti-mouse T-cell receptor-b eFluor450, CD4antigen-presenting cell, and CD8a antigen-presenting celleFluor 780 (eBioscience, San Diego, CA). Acquisition wasperformed on a BD LSR II flow cytometer (BD Biosciences,Franklin Lakes, NJ), followed by analysis with FCS Expressflow cytometry software (De Novo Software, Los Angeles, CA).
Calcium ImagingMeasurements of intracellular calcium concentration forindividual human islets from the different treatmentgroups were carried out by previously described methods(25,26) at glucose concentrations of 2.5 and 25 mmol/L.Glucose-stimulated increase in intracellular calcium con-centration was expressed as area under the curve (AUC).
Capacitance StudiesMeasurement of membrane capacitance was performedon islets according to our previously established method(27,28) to determine the effect of Tac and AAGP on b-cellexocytotic responses. Cells were stimulated with a seriesof 10 depolarizations to activate voltage-dependent Ca2+
channels. Whole-cell capacitance responses were normal-ized to initial cell size and expressed as femtofarad perpicofarad (fF/pF).
In Vivo EvaluationAll mice were housed under conventional conditions, withaccess to food and water ad libitum. The care for mice wasin accordance with guidelines approved by the CanadianCouncil on Animal Care.
Transplants With Human Islets and InflammationAnalysisDiabetes was induced chemically in 8- to 12-week-old immu-nodeficient B6.129S7-Rag1tm1Mom recipient mice (TheJackson Laboratory, Bar Harbor, ME) by intraperitonealinjection of 180 mg/kg streptozotocin (Sigma-Aldrich).Mice were considered diabetic after two consecutive bloodglucose measurements $11.3 mmol/L (350 mg/dL). Re-cipients (n = 10 per group) received ;1,000 IEQ humanislets from three different isolations. Islets from each iso-lation were randomly allocated to each group (Tac2,AAGP, Tac+, Tac+AAGP) to control for potential differ-ences in each islet preparation. Islets were transplantedunder the kidney capsule as described previously (29).A minimal islet dose was used to stress the model andmaximize covert toxicity (30–32).
Three mice per group underwent acute graft explanta-tion and were killed on day 1 and 7 posttransplant todetermine proinflammatory cytokine concentrations (atboth time points), cleaved caspase-3, and TUNEL-stained
area (24 h) within the graft. For cytokine and cleavedcaspase-3 quantification, the islet grafts were excisedfrom the kidney and bisectioned, with one section flashfrozen in liquid nitrogen and stored at 280°C and theother formalin fixed and processed for TUNEL quantifica-tion. Tissue samples were subsequently lysed in acid bufferas reported previously (30). Cytokine and cleaved caspase-3determination was adjusted per gram of tissue.
Proinflammatory Cytokines and ChemokinesRelevant cytokines and chemokines (interferon-g, inter-leukin [IL]-1b, IL-6, IL-10, IL-12, keratinocyte-derivedchemokine [KC], and tumor necrosis factor-a [TNF-a])were measured with a multispot Mouse ProInflammatory7-Plex Ultra-Sensitive Kit and analyzed on a SECTOR Im-ager (Meso Scale Discovery, Gaithersburg, MD). Resultsare expressed as absolute values (pg/mL), and normal re-nal tissue lysate samples from a mouse receiving a shamoperation were used as control.
ApoptosisApoptosis was determined in the excised grafts 24 hposttransplant by quantifying cleaved caspase-3 andanalyzing percentage of death cells (TUNEL) within thegraft. Caspase-3 concentration was expressed as fold-change increase compared with normal renal tissue lysatesamples from a naive mouse.
Long-term Human Islet Graft Function AfterTransplantation in Immunodeficient MiceNonfasting blood glucose was monitored in the re-maining animals three times a week with a portableglucometer (OneTouch Ultra2; LifeScan Canada) over60 days. Normoglycemia was defined as two consecutivereadings ,11.3 mmol/L.
Intraperitoneal glucose tolerance tests (IPGTTs) wereconducted 60 days posttransplant to evaluate the capac-ity of islets to respond to a glucose bolus (3 g/kg) afterovernight fast. Blood glucose levels were monitored atbaseline time 0 and 15, 30, 60, 90, and 120 min post-injection. All results were compared with blood glucoseprofiles of naive control nondiabetic mice.
Recovery islet–bearing nephrectomies were performedon day 65 to demonstrate graft-dependent euglycemia.Both cultured islets and recovered grafts were stored at280°C and processed to measure intracellular insulin con-tent by acid-ethanol homogenization and ultrasonic lysis.The extract was neutralized and insulin measured withELISA (30).
Transplants With Mouse Islets Under ContinuousTreatment With Tac
Mouse Islet IsolationPancreatic islets were isolated from 8- to 12-week-old maleBALB/c mice (The Jackson Laboratory), as reported pre-viously (18). Islets were counted and divided into threegroups (Tac2, Tac+, and Tac+AAGP). All islets were incu-bated for 1 h under the previously described conditions,and Tac+AAGP islets were supplemented with AAGP at
diabetes.diabetesjournals.org Gala-Lopez and Associates 453

3 mg/mL during the incubation period. Recipient synge-neic BALB/c mice were also rendered diabetic with strep-tozotocin and transplanted after incubation with 500IEQ 6 10% of 90% purity under the renal capsule (18).
Subcutaneous micro-osmotic pumps (model 1002; ALZET,Cupertino, CA) were implanted in all mice at the time ofislet transplantation to provide continuous delivery of Tac.A first group (short duration) received a pump deliveringTac for 7 days at a dose of 1 mg/kg/day (Tac+ [n = 6],Tac+AAGP [n = 8]), and a second group (long duration)received pumps delivering the CNI for 28 days (Tac+ [n = 6],
Tac+AAGP [n = 6]). The Tac2 group (n = 10) receivedpumps loaded with normal saline as placebo. Steady-stateTac levels were monitored selectively from the dorsal tailvein at day 5 and ranged from 10 to 20 ng/mL (clinicallyrelevant range, tandem liquid chromatography–mass spec-trometry for continuously administered drug) (33).
Animals underwent IPGTT on day 7 (short duration)and day 14 (long duration) during the treatment courseand again on days 30 and 40 after CNI treatment cessation.Transplant islet–bearing nephrectomies were performedafter tolerance tests to prove graft-dependent function.
Figure 1—AAGP improves human islet potency in culture and protects against acute exposure to Tac. A: In vitro assessment of humanislets in culture with or without AAGP supplementation showed a significantly higher islet recovery rate after culture in the presence ofAAGP. B: No significant changes in cell viability were found after the study period. C and D: Perifusion curves comparing GSIS afterstimulation with variable glucose concentrations (low 2.8 mmol/L, high 28 mmol/L) show severely impaired islet function for the Tac2 groupand a significantly better response for groups treated with AAGP (also seen in the corresponding AUC, n = 6). Tac impairs insulin secretionwithout affecting insulin biosynthesis. s-GSIS assay and intracellular insulin content were simultaneously measured in human islets keptin culture. E: The stimulation index for the Tac+ group was significantly decreased compared with control. However, a significantimprovement was observed in insulin secretion of Tac+AAGP islets (P < 0.01), whereas no changes were seen in the intracellularcontent of insulin across the different groups (F). Data are mean 6 SEM (triplicates from four different preparations). *P < 0.05, **P < 0.01,***P < 0.001, ****P < 0.0001.
454 AAGP Protects Transplanted Human Islets Diabetes Volume 65, February 2016

Statistical AnalysisData are presented as mean 6 SEM. AUC was calculatedfor GSIS and D-IP, calcium imaging, capacitance measure-ments, and IPGTT, and differences between groups wereanalyzed with one-way ANOVA with Tukey post hoc test.P , 0.05 was considered significant, and all analyses wereperformed with GraphPad Prism software (GraphPadSoftware, La Jolla, CA).
RESULTS
AAGP Enhances Human Islet Potency in Culture andProtects Islets Against Acute Exposure to TacIsolated human islets from six different preparations werecultured in media supplemented with or without AAGPand Tac as described. After 48 h of culture, all groups were
characterized for in vitro survival, viability, function,oxidative stress, and apoptosis.
After the study period, cells were counted, resultingin a greater number of surviving islets in the AAGP-supplemented group (71.1%). Exposure to Tac clearlydecreased survival, but islets significantly recoveredwhen simultaneously supplemented with AAGP (Tac+ vs.Tac+AAGP, 31.5% vs. 67.6%, P , 0.01) (Fig. 1A). Therewas no difference in cell viability by membrane integritystain (Fig. 1B).
When comparing in vitro function by D-IP, insulinrelease was completely suppressed after Tac exposure (AUC:Tac2 vs. Tac+, 131 vs. 7.3 mmol/L/60 min, P , 0.001).However, islet function was fully restored after supplemen-tation with AAGP and comparable to the other culture
Figure 2—Islets treated with AAGP have decreased oxidative stress. A: Human islets in culture had an increased concentration of ROSwhen treated with Tac. However, supplementation with AAGP significantly decreased this effect (P< 0.05). Oxidative stress was measuredby fold increase in extracellular ROS analyzed with the Acridan Lumigen PS-3 assay (n = 5). AAGP effect is not the result of direct druginhibition with Tac. B–D: Allogeneic MLR was used to evaluate direct drug inhibition. Results show a significant decrease of T-cellproliferation in the presence of Tac, AAGP, and the combination of both, hence no direct inhibition of Tac by AAGP. Data are mean 6 SEM(n = 6). *P < 0.05, ***P < 0.001, ****P < 0.0001. TCR, T-cell receptor.
diabetes.diabetesjournals.org Gala-Lopez and Associates 455

conditions (Tac+ vs. Tac+AAGP, 7.3 vs. 129.2 mmol/L/60 min, P , 0.001) (Fig. 1C and D).
Acute Exposure to Tac Decreases Insulin Secretionbut Not BiosynthesisAliquots of 100 IEQ human islets were collected for eachgroup for s-GSIS assay and intracellular insulin content.The Tac+ group showed significant impairment of insulinsecretion, which was not observed in the Tac+AAGP group(stimulation index 1.4 vs. 10.7, P , 0.01) (Fig. 1E). How-ever, intracellular insulin content remained stable andcomparable across groups, demonstrating no changes tobiosynthesis of insulin within b-cells (Fig. 1F).
AAGP Reduces Oxidative Stress but Does Not InhibitTac FunctionOxidative stress was observed in all groups, but Tacexposure resulted in a substantial increase in ROS, whichwas ameliorated in the presence of AAGP (n = 6, P, 0.05)(Fig. 2A). To confirm that AAGP did not inhibit Tac sup-pression of T-cell proliferation, MLRs were completedwith donor splenocytes. The assay measured T-cell pro-liferative response by carboxyfluorescein diacetate succi-nimidyl ester staining. As expected, T-cell proliferationwas significantly decreased in the presence of Tac com-pared with IgG controls (n = 4, P , 0.001). Proliferationof CD8+- and CD4+-positive T cells was also significantlydecreased in the presence of Tac alone or in combinationwith AAGP (n = 4, P , 0.001 in both cases), with noimpediment to MLR suppression in the presence ofAAGP (Fig. 2B–D).
Tac Effect on Islet Intracellular Calcium Responsesand ExocytosisVarious studies were performed on human islets toelucidate a potential mechanism of action for AAGP bycharacterizing CNI-related injury and its minimization.Intracellular calcium concentrations were measured todetermine a possible influence of AAGP on glucose-stimulated calcium influx (34). No significant differencesin calcium influx were observed between groups (AUC:Tac2 209.4, Tac+ 221.6, Tac+AAGP 208.7 fF/treatment/2 s; P . 0.05), suggesting that the protective effect ofAAGP was further downstream in the secretory pathway(Fig. 3A).
Complementary membrane capacitance studies wereperformed under similar conditions as an indirect in-dicator of insulin exocytosis. There was a decreasedcumulative capacitance response in the Tac+ group com-pared with the other groups and a significantly lowerAUC compared with the Tac+AAGP group (AUC 2.9 vs.10.5 fF/pF/treatment, P , 0.001) (Fig. 3B).
AAGP Prevents Islet Apoptosis Resulting From In VitroExposure to TacExposure to Tac during culture resulted in an increasedconcentration of intracellular cleaved caspase-3 (foldchange: Tac2 vs. Tac+, 1.9 vs. 4.3, P , 0.05), which cor-responded with increased the percentage of apoptotic
islets (Tac2 vs. Tac+, 18.9% vs. 48.6%, P , 0.01). Con-versely, pretreatment with AAGP prevented Tac-inducedcell death, showing reduced levels of caspase-3 (Tac+ vs.Tac+AAGP, 4.3 vs. 2.2, P , 0.05) and a fewer number ofapoptotic cells (Tac+ vs. Tac+AAGP, 48.6% vs. 26%, P ,0.05) (Fig. 4A and B). Representative slides from TUNELhistology are shown in Fig. 4C.
AAGP Ameliorates Inflammatory ResponseImmediately PosttransplantMinimal-mass (1,000 IEQ) human islet transplants wereperformed in diabetic immunodeficient mice. Grafts fromthree animals per group (day 1 and day 7 posttransplant)were homogenized and characterized for proinflammatorycytokines and chemokines.
Acute levels of IL-1b were significantly increased inTac+ with respect to those in the sham group (18.9 vs.163.1 pg/g tissue, P , 0.001). Cytokine concentration,however, was considerably dampened in the Tac+AAGPgroup (163.1 vs. 44.9, P , 0.001), with similar excretion
Figure 3—AAGP improves insulin release by increasing islet exo-cytosis. Human islets were cultured and treated with and withoutTac/AAGP. A: Insulin secretion impairment for the Tac+ group wasalso met by a lower concentration of intracellular calcium. B: Differ-ences among groups did not reach statistical significance, butfurther studies showed that normalized membrane capacitancemeasurements were significantly worse for Tac+ islets (A, blue), in-dicating impaired exocytosis, whereas measurements were supe-rior and comparable in the Tac2 (A, red) and Tac+AAGP groups(A, black). Data are mean 6 SEM (triplicates from two isolations).**P < 0.01, ***P < 0.001.
456 AAGP Protects Transplanted Human Islets Diabetes Volume 65, February 2016

behavior on day 7 (269.5 vs. 121 pg/g tissue, n = 3, P ,0.001) (Fig. 5A and B). Similarly, IL-6 was significantlyincreased in the Tac+ group (1,414 vs. 804.7 pg/g tissue,n = 3, P, 0.001), but differences were no longer apparentat later time points (Fig. 5C and D).
Among the chemokines measured acutely posttrans-plant, KC secretion, involved in neutrophil recruitment,was significantly overexpressed in Tac+ and, again, sig-nificantly reduced in the presence of AAGP (85.4 vs. 32pg/g tissue, n = 3, P , 0.001). By day 7, the cytokine wasclearing, and differences were no longer evident (Fig. 5Eand F). TNF-a levels, on the other hand, were not sig-nificantly increased on day 1 but became notably differ-ent on day 7 (Tac2 33.9, Tac+ 76.7, Tac+AAGP 48.7 pg/gtissue; P , 0.001) (Fig. 5G and H).
Intragraft apoptosis showed an increasing trend incleaved caspase-3 concentration and TUNEL-positive cellsin the Tac+ group compared with the rest of the groups,suggesting increased in vivo cell death after CNI treat-ment and a subsequent AAGP protective effect. However,differences did not reach statistical significance (Tac+ vs.Tac+AAGP: fold change in caspase-3 2.9 vs. 1.9, not sig-nificant; TUNEL-positive cells 35.36 26.7% vs. 7.96 8%,not significant) (data not shown).
AAGP Supplementation Improves Human IsletTransplant Function Despite Tac ExposureThe remaining transplanted mice (n = 7 per group) werefollowed beyond 60 days. Delayed engraftment was observedas expected in this marginal islet mass model. Blood glu-cose improved over time in the Tac2 and Tac+AAGPgroups, with the proportion of euglycemic animals
being significantly higher than those in the Tac+ group,where all mice demonstrated poor function (P , 0.05)(Fig. 6A and B).
Thirty days posttransplant, mice underwent IPGTTto evaluate transplant function. The Tac2 andTac+AAGP groups both responded appropriately, butTac+ remained hyperglycemic at 120 min (AUC: Tac2
vs. Tac+AAGP, 92.6 vs. 91.2, P . 0.05; Tac2 vs. naive,92.6 vs. 71.4, P . 0.05; Tac+AAGP vs. naive, 91.2 vs.71.4, P . 0.05; Tac+ vs. Tac+AAGP, 149.8 vs. 91.2,P , 0.05) (Fig. 6C).
All mice reverted to their previous diabetic state afterislet-bearing nephrectomy. Insulin content was assessedas a measure of residual islet mass after 30 days. Figure6D shows significant differences between groups withreduced insulin content in grafts exposed to Tac. Again,presence of AAGP was beneficial in islet protection de-spite exposure to Tac (Tac+ vs. Tac+AAGP, 30.9 vs. 100.8ng/mL, P , 0.01).
AAGP Supplementation Improves Islet TransplantFunction Despite Continuous Recipient TreatmentWith TacIn further support of the findings, syngeneic diabetic micewere implanted with subcutaneous micro-osmotic pumpsto model continuous posttransplant Tac treatment inclinical practice. As with the in vitro findings, trans-planted islets exposed to Tac were unable to effectivelysecrete insulin or return mice to euglycemia during thetreatment course. The presence of AAGP, however, re-stored normal islet function despite Tac exposure,similar to controls (P , 0.001) (Fig. 7A and B).
Figure 4—AAGP decreases islet loss in culture due to apoptosis, even in the presence of Tac. Human islets were cultured and treated withTac/AAGP. Cell death due to apoptosis was significantly higher in Tac+ by quantification of intracellular cleaved caspase-3 (A) and byanalysis of TUNEL staining (B). Supplementation of media with AAGP restored viability and significantly decrease cell death (P < 0.05).C: Representative TUNEL slides of fixed islets from different study groups. Arrows point to TUNEL-positive nuclei. *P < 0.05, **P < 0.01.
diabetes.diabetesjournals.org Gala-Lopez and Associates 457

Similar findings were observed when using subcutane-ous pumps providing Tac treatment for a longer duration(28 days). Again, AAGP-supplemented islets functionednormally and rendered normoglycemia for all animals,despite a single 1-h AAGP treatment of islets beforetransplant (P , 0.01) (Fig. 7E and F).
Results were corroborated by IPGTTs performed inboth treatment modalities (short and long duration).Tolerance tests performed under Tac treatment showedimpaired glucose control in the Tac+ group, whereasTac+AAGP behaved similar to controls (P , 0.001)(Fig. 7C and G).
Figure 5—AAGP ameliorates inflammatory response immediately posttransplant. Proinflammatory cytokines and chemokines locallyexpressed 24 h and 7 days after transplantation. Concentrations of IL-1b (A and B), IL-6 (C and D), KC/human growth-regulated oncogene(GRO) (E and F ), and TNF-a (G and H) were significantly lower in the engrafted islets previously treated with AAGP. Cytokines weremeasured 24 h and 7 days after transplantation locally to the graft by homogenization (normalized per gram of tissue, n = 3). Data aremean 6 SEM adjusted per gram of tissue (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
458 AAGP Protects Transplanted Human Islets Diabetes Volume 65, February 2016

Tac treatment cessation resulted in normalization ofgraft function and euglycemia in all animals. RepeatIPGTTs at this stage (30 and 45 days) showed no residualdifferences between groups (Fig. 7D and H).
DISCUSSION
We demonstrate that the addition of a potent antifreezeprotein AAGP only to the islet culture media for a 48-hexposure affords considerable protection of human isletsurvival and in vitro function. This protective effect isespecially pronounced when used to prevent Tac-inducedislet toxicity.
Tac currently is regarded as a mainstay potent immu-nosuppressant given to prevent both auto- and alloim-munity after clinical islet transplantation (35). Prolongedexposure to CNI class immunosuppressants is stronglyassociated with nephrotoxicity and posttransplant diabe-tes in all organ transplants (4).
Tac is known to impair insulin secretion in the nativepancreas after pancreas and, especially, islet transplanta-tion and is characterized by impairment of early secretionand decreased biosynthesis. Several associated mechanismshave been defined, including calcineurin/nuclear factor ofactivated T-cell signaling inhibition (36), insulin gene sup-pression (37), mitochondrial arrest (38), and decreased
posttransplant vascularization (39). In the current experi-mental model, the addition of Tac resulted in a strikinginhibition of insulin secretion and cell death in vitro andimpaired islet engraftment and function in vivo. Further-more, we confirmed islet function impairment after Tacexposure in vitro and found that AAGP was able to reestab-lish insulin release despite acute exposure to high-dose Tac.
An increased loss of islets during culture was associ-ated with apoptosis observed after in vitro exposure toTac. Increase in cleaved caspase-3 and TUNEL stainingindicated significantly higher cell death in the Tac+ group,but this was diminished after AAGP supplementation.
Islet high susceptibility to hypoxia throughout allstages of cell procurement, preparation, and intraportaltransplantation relates to their intrinsic oxygen demandand size, especially that related to islet seeding density inculture (40). Islets are prone to oxidative stress due todecreased antioxidant capacity (41). These elements con-tribute to islet loss during culture and after transplan-tation. The current findings confirm an increase inoxidative stress after Tac exposure, with increased extra-cellular ROS. AAGP supplementation reduced oxidativestress in this model. Similar redox modulation findingshave also been noted when using AAGP with other celllines (17).
Figure 6—AAGP supplementation improves islet transplant function despite Tac exposure. Posttransplant graft function in immunodefi-cient mice receiving minimal human islet mass (1,000 IEQ). Islets were previously treated with or without Tac/AAGP. A and B: Pooled bloodglucose profiles and percent of mice reaching euglycemia demonstrate long-term graft function (60 days) with a nonfunctioning graft for theTac+ group. Graft-bearing nephrectomy was performed on day 60 to demonstrate graft-dependent euglycemia. A: Horizontal continuousline at 11 mmol/L indicates the normoglycemia limit. Vertical dotted line refers to graft-bearing nephrectomy on day 60, resulting in acutehyperglycemia, thus confirming the islet transplant was responsible for normoglycemia. C: IPGTT to evaluate metabolic response afterreceiving a glucose bolus. D: The Tac+ group was intolerant to high glucose, which also corresponded to less residual insulin content whengrafts were removed after 60 days posttransplant. Data are mean 6 SEM adjusted per gram of tissue (Tac2 n = 6, Tac+ n = 3, Tac+AAGPn = 7). *P < 0.05, **P < 0.01, ***P < 0.001. Tx, transplant.
diabetes.diabetesjournals.org Gala-Lopez and Associates 459
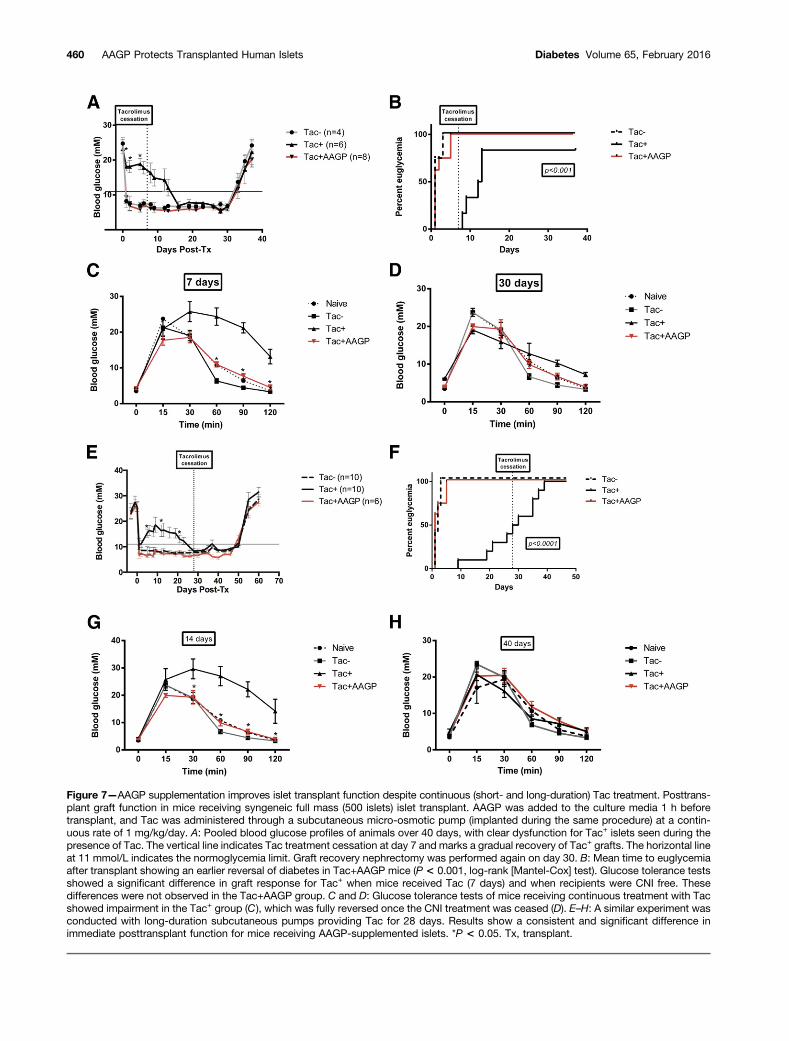
Figure 7—AAGP supplementation improves islet transplant function despite continuous (short- and long-duration) Tac treatment. Posttrans-plant graft function in mice receiving syngeneic full mass (500 islets) islet transplant. AAGP was added to the culture media 1 h beforetransplant, and Tac was administered through a subcutaneous micro-osmotic pump (implanted during the same procedure) at a contin-uous rate of 1 mg/kg/day. A: Pooled blood glucose profiles of animals over 40 days, with clear dysfunction for Tac+ islets seen during thepresence of Tac. The vertical line indicates Tac treatment cessation at day 7 and marks a gradual recovery of Tac+ grafts. The horizontal lineat 11 mmol/L indicates the normoglycemia limit. Graft recovery nephrectomy was performed again on day 30. B: Mean time to euglycemiaafter transplant showing an earlier reversal of diabetes in Tac+AAGP mice (P < 0.001, log-rank [Mantel-Cox] test). Glucose tolerance testsshowed a significant difference in graft response for Tac+ when mice received Tac (7 days) and when recipients were CNI free. Thesedifferences were not observed in the Tac+AAGP group. C and D: Glucose tolerance tests of mice receiving continuous treatment with Tacshowed impairment in the Tac+ group (C), which was fully reversed once the CNI treatment was ceased (D). E–H: A similar experiment wasconducted with long-duration subcutaneous pumps providing Tac for 28 days. Results show a consistent and significant difference inimmediate posttransplant function for mice receiving AAGP-supplemented islets. *P < 0.05. Tx, transplant.
460 AAGP Protects Transplanted Human Islets Diabetes Volume 65, February 2016

In vivo studies complemented all in vitro findings,which demonstrated that AAGP supplementation sup-pressed early inflammation and improved islet engraft-ment with long-term efficacy. AAGP-supplemented isletsshowed significantly reduced expression of IL-1b and IL-6along with decreased secretion of KC and TNF-a, despiteexposure to Tac in culture. These cytokines and chemo-kines are key participants in the posttransplant inflam-matory response and subsequent adaptive immunityactivation (42) as well as vital elements in the earlyclinical posttransplant phase (30). These findings are con-sistent with previous experiments showing reduced ex-pression of cyclooxygenase-2 expression in HeLa cellsexposed to increasing concentrations of IL-1b in the pres-ence of AAGP (17).
Tac exposure was provided by continuous subcutane-ous micro-osmotic pump for 7 and 28 days in syngeneicmice. We chose this approach in selected experimentsbecause twice-daily oral gavage of Tac would have beentoo stressful, but we wished to maintain sustained clini-cally relevant drug exposure for transplanted islets. Weobserved that marked impairment of transplanted isletsoccurred immediately following Tac exposure, which lastedthroughout Tac exposure, but this was reversible afterwithdrawal of Tac. Islets treated with AAGP, however,were protected from Tac toxicity and functioned similarto control in both the short- and the long-durationtreatment groups. Because a marked and prolongedposttransplant engraftment and functional benefit wasconsistently observed when AAGP treatment was con-fined only to the in vitro culture period, this treatmentcould potentially be readily applied in clinical studies toenhance islet engraftment and function in patientsreceiving Tac immunosuppression.
In exploring potential mechanisms of action of AAGP,we found no beneficial effect upon insulin synthesis orstorage. Furthermore, we did not find an interactiveimpact of AAGP on the immunosuppressive properties ofTac. We found that neither Tac nor AAGP affectedglucose-stimulated calcium influx in islets, which is akey element in the insulin secretion mechanism of b-cells.This information supports evidence pointing to a potentialTac mechanistic site further downstream in the secretorypathway (43). Conversely, islet capacitance measurementsin the current studies revealed significant differences be-tween Tac+ and TAC+AAGP, findings that suggest impairedinsulin exocytosis in the presence of Tac, which was re-versed by AAGP.
In conclusion, supplementation of islets with AAGPduring culture enhanced both the quality and the yield ofpostculture human islets, which translated into im-proved engraftment despite the presence of Tac. AAGPalso protected islets continuously exposed to Tac post-transplantation, with improved efficacy and decreasedinflammatory response. Clinical translation of thesefindings could potentially offer a means to protectislets both in vitro and in vivo from diabetogenic
immunosuppression after transplantation as a means toenhance single-donor islet engraftment and durablelong-term function.
Funding. Funding sources include the Diabetes Research Institute FoundationCanada, the Alberta Innovates–Health Solutions (AIHS) Collaborative Researchand Innovation Opportunities Team Award, and the Alberta Diabetes Institute.B.L.G.-L. is supported by an Izaak Walton Killam Memorial Scholarship, an AIHSClinician Fellowship, and by the Canadian National Transplant Research Pro-gram. P.E.M. is supported by a Canada Research Chair in Islet Biology.A.M.J.S. is supported through a Canada Research Chair in TransplantationSurgery and Regenerative Medicine and through AIHS as a senior scholar.Duality of Interest. L.G.Y. owns shares in ProtoKinetix Inc., the companythat owns the patent rights to the AAGP molecule. Subsequent to completion ofthis study A.M.J.S. now serves as a consultant to ProtoKinetix Inc. No otherpotential conflicts of interest relevant to this article were reported.Author Contributions. B.L.G.-L. contributed to the design and perfor-mance of experiments, data research, and writing of the manuscript. A.R.P.contributed to the performance of experiments, data research, discussion, andreview and editing of the manuscript. R.L.P. isolated mouse islets and reviewedthe manuscript. D.O. performed D-IP and reviewed the manuscript. T.K. isolatedhuman islets and reviewed the manuscript. A.Br., N.A., and M.B. researcheddata. A.Ba. performed the capacitance studies. J.E.M.F. performed calciumimaging and reviewed the manuscript. L.G.Y. contributed to the study design,provision of AAGP, and review of the manuscript. P.E.M. contributed to the dataresearch and review of the manuscript. A.M.J.S. contributed to the experimentaldesign, data research, and review and editing of the manuscript. A.M.J.S. is theguarantor of this work and, as such, had full access to all the data in the studyand takes responsibility for the integrity of the data and the accuracy of the dataanalysis.Prior Presentation. Parts of this work were presented orally at the 2015International Pancreas & Islet Transplant Association, International Xenotrans-plantation Association, and Cell Transplant Society Congress, Melbourne, VIC,Australia, 15–19 November 2015.
References1. Shapiro AM. Immune antibody monitoring predicts outcome in islet trans-plantation. Diabetes 2013;62:1377–13782. Pepper AR, Gala-Lopez B, Ziff O, Shapiro AJ. Current status of clinical islettransplantation. World J Transplant 2013;3:48–533. Nir T, Melton DA, Dor Y. Recovery from diabetes in mice by beta cell re-generation. J Clin Invest 2007;117:2553–25614. Chand DH, Southerland SM, Cunningham RJ 3rd. Tacrolimus: the good, thebad, and the ugly. Pediatr Transplant 2001;5:32–365. Yates CJ, Cohney SJ. Prediction and diagnosis of post transplant diabetes.Curr Diabetes Rev 2015;11:170–1746. Weng LC, Chiang YJ, Lin MH, et al. Association between use of FK506 andprevalence of post-transplantation diabetes mellitus in kidney transplant patients.Transplant Proc 2014;46:529–5317. Niu YJ, Shen ZY, Xu C, et al. Establishment of tacrolimus-induced diabetesin rat model and assessment of clinical treatments for post-transplant diabetesmellitus in liver transplant recipients. Clin Lab 2013;59:869–8748. Therasse A, Wallia A, Molitch ME. Management of post-transplant diabetes.Curr Diab Rep 2013;13:121–1299. Desai NM, Goss JA, Deng S, et al. Elevated portal vein drug levels ofsirolimus and tacrolimus in islet transplant recipients: local immunosuppressionor islet toxicity? Transplantation 2003;76:1623–162510. Shapiro AM, Gallant HL, Hao EG, et al. The portal immunosuppressivestorm: relevance to islet transplantation? Ther Drug Monit 2005;27:35–3711. Shapiro AM. Strategies toward single-donor islets of Langerhans trans-plantation. Curr Opin Organ Transplant 2011;16:627–631
diabetes.diabetesjournals.org Gala-Lopez and Associates 461

12. Bang JK, Lee JH, Murugan RN, et al. Antifreeze peptides and glycopep-tides, and their derivatives: potential uses in biotechnology. Mar Drugs 2013;11:2013–204113. DeVries AL. Antifreeze peptides and glycopeptides in cold-water fishes.Annu Rev Physiol 1983;45:245–26014. Amir G, Rubinsky B, Basheer SY, et al. Improved viability and reducedapoptosis in sub-zero 21-hour preservation of transplanted rat hearts using anti-freeze proteins. J Heart Lung Transplant 2005;24:1915–192915. Matsumoto S, Matsusita M, Morita T, et al. Effects of synthetic antifreezeglycoprotein analogue on islet cell survival and function during cryopreservation.Cryobiology 2006;52:90–9816. Deller RC, Vatish M, Mitchell DA, Gibson MI. Synthetic polymers enable non-vitreous cellular cryopreservation by reducing ice crystal growth during thawing.Nat Commun 2014;5:324417. ProtoKinetix Inc. AAGPs overview. http://www.protokinetix.com/aagp/overview.Accessed 16 December 201418. Kin T, Senior P, O’Gorman D, Richer B, Salam A, Shapiro AM. Riskfactors for islet loss during culture prior to transplantation. Transpl Int 2008;21:1029–103519. Kin T. Islet isolation for clinical transplantation. In The Islets of Langerhans:Advances in Experimental Medicine and Biology. Vol. 654. Islam MS, Ed. NewYork, Springer Science+Business Media, 2010, p. 683–71020. Ranuncoli A, Cautero N, Ricordi C, et al. Islet cell transplantation: in vivo andin vitro functional assessment of nonhuman primate pancreatic islets. CellTransplant 2000;9:409–41421. Ricordi C, Gray DW, Hering BJ, et al. Islet isolation assessment in man andlarge animals. Acta Diabetol Lat 1990;27:185–19522. Barnett MJ, McGhee-Wilson D, Shapiro AM, Lakey JR. Variation in humanislet viability based on different membrane integrity stains. Cell Transplant 2004;13:481–48823. Cabrera O, Jacques-Silva MC, Berman DM, et al. Automated, high-throughput assays for evaluation of human pancreatic islet function. CellTransplant 2008;16:1039–104824. Uy B, McGlashan SR, Shaikh SB. Measurement of reactive oxygen speciesin the culture media using Acridan Lumigen PS-3 assay. J Biomol Tech 2011;22:95–10725. Dezaki K, Kageyama H, Seki M, Shioda S, Yada T. Neuropeptide W in the ratpancreas: potentiation of glucose-induced insulin release and Ca2+ influxthrough L-type Ca2+ channels in beta-cells and localization in islets. Regul Pept2008;145:153–15826. Yang YH, Manning Fox JE, Zhang KL, MacDonald PE, Johnson JD. IntraisletSLIT-ROBO signaling is required for beta-cell survival and potentiates insulinsecretion. Proc Natl Acad Sci U S A 2013;110:16480–1648527. Dai XQ, Plummer G, Casimir M, et al. SUMOylation regulates insulin exo-cytosis downstream of secretory granule docking in rodents and humans.Diabetes 2011;60:838–847
28. Pigeau GM, Kolic J, Ball BJ, et al. Insulin granule recruitment and exocytosisis dependent on p110gamma in insulinoma and human beta-cells. Diabetes2009;58:2084–209229. Li D, Hao J, Yuan Y-H, et al. Pancreatic islet transplantation to the renalsubcapsule in mice. Protocol Exchange 2011:206030. McCall M, Pawlick R, Kin T, Shapiro AM. Anakinra potentiates the protectiveeffects of etanercept in transplantation of marginal mass human islets inimmunodeficient mice. Am J Transplant 2012;12:322–32931. Emamaullee JA, Merani S, Toso C, et al. Porcine marginal mass islet au-tografts resist metabolic failure over time and are enhanced by early treatmentwith liraglutide. Endocrinology 2009;150:2145–215232. Merani S, Truong W, Emamaullee JA, Toso C, Knudsen LB, Shapiro AM.Liraglutide, a long-acting human glucagon-like peptide 1 analog, improves glu-cose homeostasis in marginal mass islet transplantation in mice. Endocrinology2008;149:4322–432833. Streit F, Armstrong VW, Oellerich M. Rapid liquid chromatography-tandemmass spectrometry routine method for simultaneous determination of sirolimus,everolimus, tacrolimus, and cyclosporin A in whole blood. Clin Chem 2002;48:955–95834. Dou H, Wang C, Wu X, et al. Calcium influx activates adenylyl cyclase 8 forsustained insulin secretion in rat pancreatic beta cells. Diabetologia 2015;58:324–33335. Shapiro AM, Lakey JR, Ryan EA, et al. Islet transplantation in seven patientswith type 1 diabetes mellitus using a glucocorticoid-free immunosuppressiveregimen. N Engl J Med 2000;343:230–23836. Oetjen E, Baun D, Beimesche S, et al. Inhibition of human insulin genetranscription by the immunosuppressive drugs cyclosporin A and tacrolimus inprimary, mature islets of transgenic mice. Mol Pharmacol 2003;63:1289–129537. Hernández-Fisac I, Pizarro-Delgado J, Calle C, et al. Tacrolimus-induceddiabetes in rats courses with suppressed insulin gene expression in pancreaticislets. Am J Transplant 2007;7:2455–246238. Rostambeigi N, Lanza IR, Dzeja PP, et al. Unique cellular and mitochondrialdefects mediate FK506-induced islet b-cell dysfunction. Transplantation 2011;91:615–62339. Nishimura R, Nishioka S, Fujisawa I, et al. Tacrolimus inhibits the re-vascularization of isolated pancreatic islets. PLoS One 2013;8:e5679940. Papas KK, Avgoustiniatos ES, Tempelman LA, et al. High-density culture ofhuman islets on top of silicone rubber membranes. Transplant Proc 2005;37:3412–341441. Sklavos MM, Bertera S, Tse HM, et al. Redox modulation protects isletsfrom transplant-related injury. Diabetes 2010;59:1731–173842. Kanak MATM, Takita M, Kunnathodi F, Lawrence MC, Levy MF, Naziruddin B.Inflammatory response in islet transplantation. Int J Endocrinol 2014;2014:45103543. Uchizono Y, Iwase M, Nakamura U, Sasaki N, Goto D, Iida M. Tacrolimusimpairment of insulin secretion in isolated rat islets occurs at multiple distal sitesin stimulus-secretion coupling. Endocrinology 2004;145:2264–2272
462 AAGP Protects Transplanted Human Islets Diabetes Volume 65, February 2016