anion exchanger 1 interacts with nephrin in...
TRANSCRIPT

Anion Exchanger 1 Interacts with Nephrin in PodocytesFiona Wu,*†‡ Moin A. Saleem,† Nicole B. Kampik,§ Timothy J. Satchwell,‡
Rosalind C. Williamson,‡ Simone M. Blattner,� Lan Ni,† Tibor Toth,† Graham White,†
Mark T. Young,‡ Mark D. Parker,‡ Seth L. Alper,¶ Carsten A. Wagner,§ andAshley M. Toye‡
*Department of Clinical Sciences, South Bristol, †Academic Renal Unit, and ‡Department of Biochemistry, School ofMedical Sciences, University of Bristol, United Kingdom; §Institute of Physiology and Zurich Center for Integrative HumanPhysiology, University of Zurich, Zurich, Switzerland; �Division of Nephrology, Department of Internal Medicine, Universityof Michigan, Ann Arbor, Michigan; and ¶Divisions of Nephrology and Molecular and Vascular Medicine, Department ofMedicine, Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, Massachusetts
ABSTRACTThe central role of the multifunctional protein nephrin within the macromolecular complex forming theglomerular slit diaphragm is well established, but the mechanisms linking the slit diaphragm to thecytoskeleton and to the signaling pathways involved in maintaining the integrity of the glomerular filterremain incompletely understood. Here, we report that nephrin interacts with the bicarbonate/chloridetransporter kidney anion exchanger 1 (kAE1), detected by yeast two-hybrid assay and confirmed byimmunoprecipitation and co-localization studies. We confirmed low-level glomerular expression of kAE1in human and mouse kidneys by immunoblotting and immunofluorescence microscopy. We observedless kAE1 in human glomeruli homozygous for the NPHS1FinMaj nephrin mutation, whereas kAE1expression remained unchanged in the collecting duct. We could not detect endogenous kAE1 expres-sion in NPHS1FinMaj podocytes in primary culture, but heterologous re-introduction of wild-type nephrininto these podocytes rescued kAE1 expression. In kidneys of Ae1�/� mice, nephrin abundance wasnormal but its distribution was altered along with the reported kAE1-binding protein integrin-linkedkinase (ILK). Ae1�/� mice had increased albuminuria with glomerular enlargement, mesangial expansion,mesangiosclerosis, and expansion of the glomerular basement membrane. Glomeruli with ILK-deficientpodocytes also demonstrated altered AE1 and nephrin expression, further supporting the functionalinterdependence of these proteins. These data suggest that the podocyte protein kAE1 interacts withnephrin and ILK to maintain the structure and function of the glomerular basement membrane.
J Am Soc Nephrol 21: 1456–1467, 2010. doi: 10.1681/ASN.2009090921
Anion exchanger 1 (AE1; SLC4A1), an SLC4 bi-carbonate transporter family member, is tran-scribed as an erythroid isoform (eAE1) and atruncated kidney isoform (kAE1) lacking aminoacids 1 through 65 in humans.1 eAE1 comprisesthe core of the multiprotein complex of integraland peripheral membrane proteins essential tothe structural integrity of the red cell membrane,and its bicarbonate/chloride activity is requiredfor gas transport (see reviews2,3). In the kidney,kAE1 is localized to the basolateral membrane ofcollecting duct type A intercalated cells. Normalterminal urinary acidification by these cells re-
quires kAE1-mediated bicarbonate reabsorptioninto the blood. Specific mutations in AE1 usually
Received September 11, 2009. Accepted April 15, 2010.
Published online ahead of print. Publication date available atwww.jasn.org.
Correspondence: Dr. Ashley M. Toye, Department of Biochemistry,School of Medical Sciences, University of Bristol, University Walk, BristolBS8 1TD, UK. Phone: 0044 117 3312111; Fax: 0044 1173312168;E-mail: [email protected]; or Dr. Fiona Wu, Department of ClinicalSciences South Bristol, Dorothy Hodgkin Building, University of Bristol,Bristol BS1 3NY, UK. Phone: 0044 117 3423553; Fax: 0044 1173423353; E-mail: [email protected]
Copyright © 2010 by the American Society of Nephrology
BASIC RESEARCH www.jasn.org
1456 ISSN : 1046-6673/2109-1456 J Am Soc Nephrol 21: 1456–1467, 2010

cause either autosomal dominant hereditary ovalo-sphero-cytosis or distal renal tubular acidosis (dRTA).4 In rare casesof homozygous recessive or compound heterozygous AE1mutations, both the erythroid and renal phenotypes canmanifest in the same individuals.5–7 In addition to theseestablished roles, low-level AE1 expression has been de-tected in the glomerulus,8,9 but its potential function orinteractions in the glomerulus are unknown.
AE1 possesses a long cytoplasmic N-terminus, a 12- to14-span transmembrane transporter domain, and a shortC-terminal cytoplasmic tail (Supplemental Figure 1). Boththe N- and C-terminal domains of kAE1 contain tyrosineresidues critical for basolateral targeting,10,11 which is likelyregulated by phosphorylation.11 The N-terminus of kAE1interacts with integrin-linked kinase (ILK),12 a protein thatbinds the cytoplasmic domains of �-integrins and cytoskel-eton-associated proteins.13,14 The kAE1/ILK interaction en-hanced kAE1 trafficking to the plasma membrane inHEK293 cells,12 but deletion of the majority of the ILK-interacting region in kAE1 did not affect its polarized traf-ficking in MDCK cells.11 Thus, the physiologic importanceof the kAE1–ILK interaction in the kidney remains unclear.We searched for proteins that interact with the C-terminusof kAE1, using a yeast two-hybrid screen of a human kidneycDNA library, and identified a novel interaction betweenkAE1 and nephrin.
Nephrin is a single-spanning transmembrane Ig super-family protein (Supplemental Figure 1) and an integralcomponent of the podocyte slit diaphragm (SD), a structurecritical to the glomerular selectivity filter.15 Mutations orgene targeting of nephrin results in congenital nephroticsyndrome.16,17 The nephrin extracellular domain contrib-utes to the structural framework of the SD via homo- andheterodimeric interactions with neighboring nephrinpolypeptides and nephrin-like homologs Neph1 andNeph2.15,18 –20 The intracellular domain of nephrin containsmultiple tyrosine phosphorylation sites and interacts withpodocin,21,22 CD2-associated protein,23,24 Nck pro-teins,25–27 the ion channel TRPC6,28,29 and adherens junc-tion proteins.30,31 These interactions anchor the SD com-plex to the underlying cytoskeleton and participate in signaltransduction. Nephrin also forms a multicomponent ter-nary complex with ILK.32 The proteinuric phenotype ofmice with podocyte-specific deletions of ILK and othercomponents of the basally situated ILK/integrin com-plex32–35 suggests that SD and basal domain signaling com-plexes of podocytes cooperate to maintain integrity of theglomerular filtration barrier.
In view of the direct associations of ILK with kAE112 andnephrin,32 we investigated the physiologic significance of thenephrin/kAE1 interaction. Our studies demonstrate the im-portance of nephrin for stable kAE1 expression in podocytesand the in vivo interdependence of levels and subcellular local-ization among kAE1, nephrin, and ILK in podocytes, suggest-ing a novel role of kAE1 in glomerular function.
RESULTS
Nephrin Interacts with kAE1 in the Yeast Two-HybridAssayA yeast two-hybrid screen of a human kidney cDNA libraryusing the C-terminus (residues 877 to 911) of human kAE1revealed an interaction with two clones identical to nephrin(GenBank accession number AF035835). This cDNA was sub-cloned, and subsequent directed two-hybrid assay confirmedthe interaction between the C-terminal domains of kAE1 andnephrin.
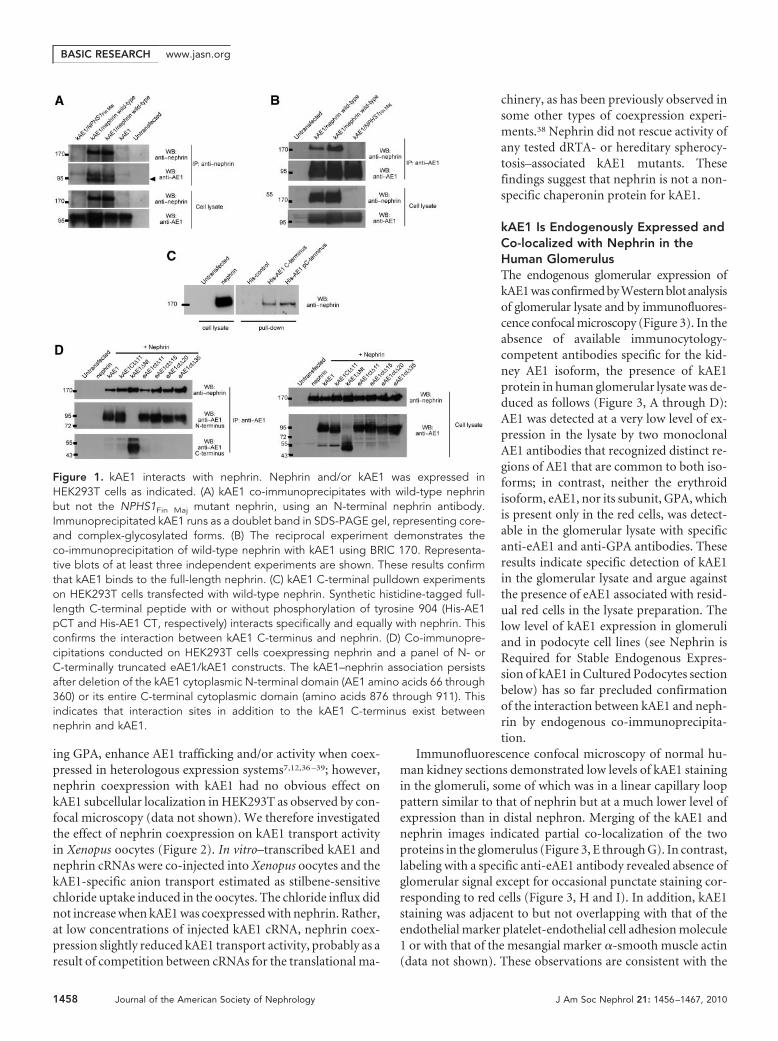
kAE1 Binds to Nephrin in HEK293T Cells, but ItsTransport Activity Is Unaffected by NephrinCoexpression in Xenopus OocytesCo-immunoprecipitation studies confirmed the interactionbetween kAE1 and nephrin (Figure 1). kAE1 was transientlyexpressed in HEK293T cells alone or in combination with ei-ther wild-type nephrin or the most prevalent nephrin mutantlinked with congenital nephrotic syndrome of the Finnishtype, a frameshift leading to a 90-residue C-terminal trunca-tion (NPHS1Fin Maj). kAE1 was co-immunoprecipitated fromthe co-transfected HEK293T cells with wild-type but notmutant nephrin using an antibody against N-terminus ofnephrin (Figure 1A). In the reciprocal experiment, wild-typenephrin was co-immunoprecipitated with kAE1 using an an-tibody against the N-terminus of kAE1 (Figure 1B). Thesefindings indicate that kAE1 and nephrin can interact to form aprotein complex.
We next used kAE1 C-terminal synthetic peptides to pulldown nephrin from the lysates of HEK293T cells overexpress-ing nephrin (Figure 1C). A synthetic peptide corresponding toeither full-length (33 amino acids; RNVELQCLDADAKAT-FDEEEGRDEYDEVAMPV) or an identical C-terminal pep-tide with phosphorylated tyrosine Y904 specifically interactedwith nephrin. Tyrosine phosphorylation of Y904 had no effecton the binding of the isolated kAE1 full-length C-terminus tonephrin.
Additional co-immunoprecipitation experiments in HEK293Tcells coexpressing nephrin and a panel of N- or C-terminallytruncated eAE1/kAE1 constructs (Figure 1D) demonstratedthat the binding of kAE1 to nephrin persisted even after dele-tion of the majority of the kAE1 cytoplasmic N-terminal do-main (amino acids 66 through 360; containing the ILK-bind-ing site) or the entire kAE1 C-terminal cytoplasmic domain(amino acids 876 to 911). Taken together, these results indicatethe existence of nephrin-binding sites in kAE1 in addition tothat in its cytosolic C-terminus. These sites may occur withinthe kAE1 transmembrane, juxtamembrane, or extracellulardomains, akin to the multiple AE1-binding sites in glycoph-orin A (GPA).4,5,36 The existence of multiple kAE1–nephrininteraction sites suggests a close association between the twomembrane proteins, but further work is needed to characterizethese additional sites.
Several proteins that interact with eAE1 in red cells, includ-
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 21: 1456–1467, 2010 kAE1 in Podocytes 1457
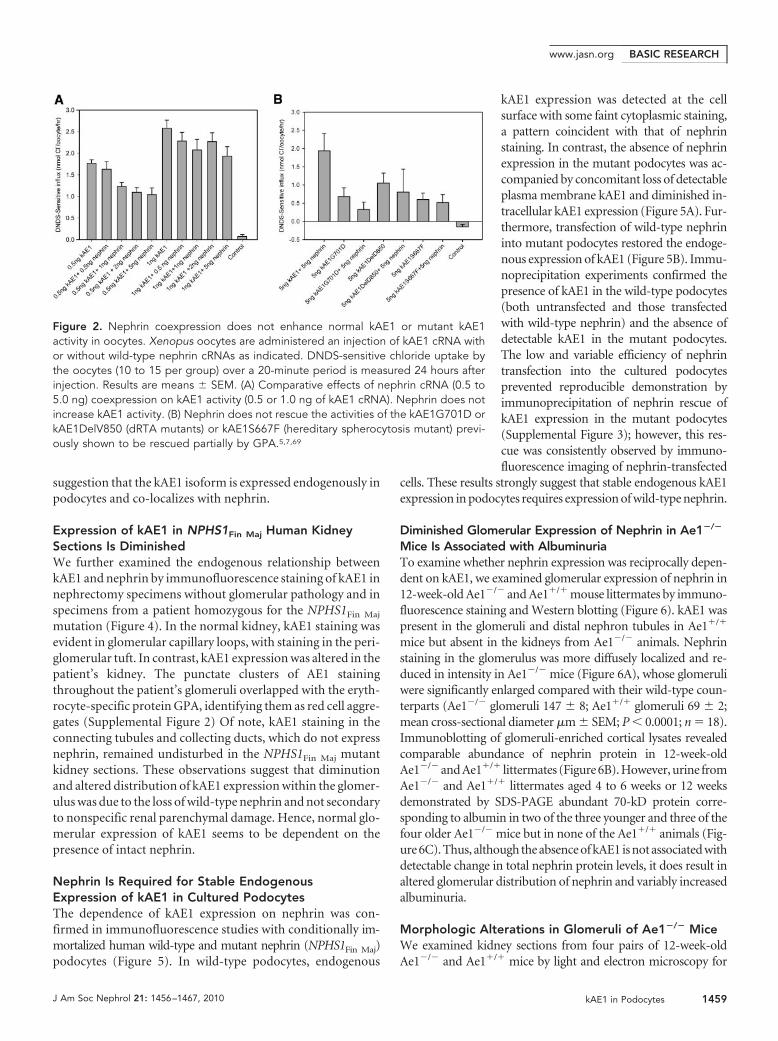
ing GPA, enhance AE1 trafficking and/or activity when coex-pressed in heterologous expression systems7,12,36 –39; however,nephrin coexpression with kAE1 had no obvious effect onkAE1 subcellular localization in HEK293T as observed by con-focal microscopy (data not shown). We therefore investigatedthe effect of nephrin coexpression on kAE1 transport activityin Xenopus oocytes (Figure 2). In vitro–transcribed kAE1 andnephrin cRNAs were co-injected into Xenopus oocytes and thekAE1-specific anion transport estimated as stilbene-sensitivechloride uptake induced in the oocytes. The chloride influx didnot increase when kAE1 was coexpressed with nephrin. Rather,at low concentrations of injected kAE1 cRNA, nephrin coex-pression slightly reduced kAE1 transport activity, probably as aresult of competition between cRNAs for the translational ma-
chinery, as has been previously observed insome other types of coexpression experi-ments.38 Nephrin did not rescue activity ofany tested dRTA- or hereditary spherocy-tosis–associated kAE1 mutants. Thesefindings suggest that nephrin is not a non-specific chaperonin protein for kAE1.
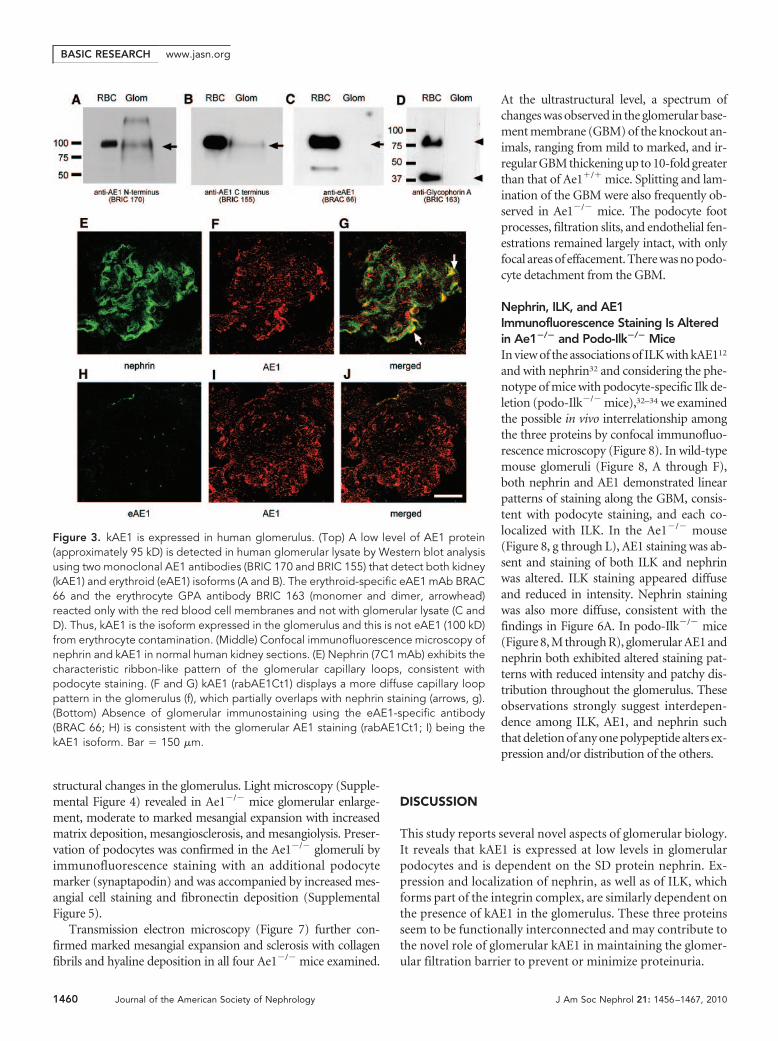
kAE1 Is Endogenously Expressed andCo-localized with Nephrin in theHuman GlomerulusThe endogenous glomerular expression ofkAE1 was confirmed by Western blot analysisof glomerular lysate and by immunofluores-cence confocal microscopy (Figure 3). In theabsence of available immunocytology-competent antibodies specific for the kid-ney AE1 isoform, the presence of kAE1protein in human glomerular lysate was de-duced as follows (Figure 3, A through D):AE1 was detected at a very low level of ex-pression in the lysate by two monoclonalAE1 antibodies that recognized distinct re-gions of AE1 that are common to both iso-forms; in contrast, neither the erythroidisoform, eAE1, nor its subunit, GPA, whichis present only in the red cells, was detect-able in the glomerular lysate with specificanti-eAE1 and anti-GPA antibodies. Theseresults indicate specific detection of kAE1in the glomerular lysate and argue againstthe presence of eAE1 associated with resid-ual red cells in the lysate preparation. Thelow level of kAE1 expression in glomeruliand in podocyte cell lines (see Nephrin isRequired for Stable Endogenous Expres-sion of kAE1 in Cultured Podocytes sectionbelow) has so far precluded confirmationof the interaction between kAE1 and neph-rin by endogenous co-immunoprecipita-tion.
Immunofluorescence confocal microscopy of normal hu-man kidney sections demonstrated low levels of kAE1 stainingin the glomeruli, some of which was in a linear capillary looppattern similar to that of nephrin but at a much lower level ofexpression than in distal nephron. Merging of the kAE1 andnephrin images indicated partial co-localization of the twoproteins in the glomerulus (Figure 3, E through G). In contrast,labeling with a specific anti-eAE1 antibody revealed absence ofglomerular signal except for occasional punctate staining cor-responding to red cells (Figure 3, H and I). In addition, kAE1staining was adjacent to but not overlapping with that of theendothelial marker platelet-endothelial cell adhesion molecule1 or with that of the mesangial marker �-smooth muscle actin(data not shown). These observations are consistent with the
Figure 1. kAE1 interacts with nephrin. Nephrin and/or kAE1 was expressed inHEK293T cells as indicated. (A) kAE1 co-immunoprecipitates with wild-type nephrinbut not the NPHS1Fin Maj mutant nephrin, using an N-terminal nephrin antibody.Immunoprecipitated kAE1 runs as a doublet band in SDS-PAGE gel, representing core-and complex-glycosylated forms. (B) The reciprocal experiment demonstrates theco-immunoprecipitation of wild-type nephrin with kAE1 using BRIC 170. Representa-tive blots of at least three independent experiments are shown. These results confirmthat kAE1 binds to the full-length nephrin. (C) kAE1 C-terminal pulldown experimentson HEK293T cells transfected with wild-type nephrin. Synthetic histidine-tagged full-length C-terminal peptide with or without phosphorylation of tyrosine 904 (His-AE1pCT and His-AE1 CT, respectively) interacts specifically and equally with nephrin. Thisconfirms the interaction between kAE1 C-terminus and nephrin. (D) Co-immunopre-cipitations conducted on HEK293T cells coexpressing nephrin and a panel of N- orC-terminally truncated eAE1/kAE1 constructs. The kAE1–nephrin association persistsafter deletion of the kAE1 cytoplasmic N-terminal domain (AE1 amino acids 66 through360) or its entire C-terminal cytoplasmic domain (amino acids 876 through 911). Thisindicates that interaction sites in addition to the kAE1 C-terminus exist betweennephrin and kAE1.
BASIC RESEARCH www.jasn.org
1458 Journal of the American Society of Nephrology J Am Soc Nephrol 21: 1456–1467, 2010
suggestion that the kAE1 isoform is expressed endogenously inpodocytes and co-localizes with nephrin.
Expression of kAE1 in NPHS1Fin Maj Human KidneySections Is DiminishedWe further examined the endogenous relationship betweenkAE1 and nephrin by immunofluorescence staining of kAE1 innephrectomy specimens without glomerular pathology and inspecimens from a patient homozygous for the NPHS1Fin Maj
mutation (Figure 4). In the normal kidney, kAE1 staining wasevident in glomerular capillary loops, with staining in the peri-glomerular tuft. In contrast, kAE1 expression was altered in thepatient’s kidney. The punctate clusters of AE1 stainingthroughout the patient’s glomeruli overlapped with the eryth-rocyte-specific protein GPA, identifying them as red cell aggre-gates (Supplemental Figure 2) Of note, kAE1 staining in theconnecting tubules and collecting ducts, which do not expressnephrin, remained undisturbed in the NPHS1Fin Maj mutantkidney sections. These observations suggest that diminutionand altered distribution of kAE1 expression within the glomer-ulus was due to the loss of wild-type nephrin and not secondaryto nonspecific renal parenchymal damage. Hence, normal glo-merular expression of kAE1 seems to be dependent on thepresence of intact nephrin.
Nephrin Is Required for Stable EndogenousExpression of kAE1 in Cultured PodocytesThe dependence of kAE1 expression on nephrin was con-firmed in immunofluorescence studies with conditionally im-mortalized human wild-type and mutant nephrin (NPHS1Fin Maj)podocytes (Figure 5). In wild-type podocytes, endogenous
kAE1 expression was detected at the cellsurface with some faint cytoplasmic staining,a pattern coincident with that of nephrinstaining. In contrast, the absence of nephrinexpression in the mutant podocytes was ac-companied by concomitant loss of detectableplasma membrane kAE1 and diminished in-tracellular kAE1 expression (Figure 5A). Fur-thermore, transfection of wild-type nephrininto mutant podocytes restored the endoge-nous expression of kAE1 (Figure 5B). Immu-noprecipitation experiments confirmed thepresence of kAE1 in the wild-type podocytes(both untransfected and those transfectedwith wild-type nephrin) and the absence ofdetectable kAE1 in the mutant podocytes.The low and variable efficiency of nephrintransfection into the cultured podocytesprevented reproducible demonstration byimmunoprecipitation of nephrin rescue ofkAE1 expression in the mutant podocytes(Supplemental Figure 3); however, this res-cue was consistently observed by immuno-fluorescence imaging of nephrin-transfected
cells. These results strongly suggest that stable endogenous kAE1expression in podocytes requires expression of wild-type nephrin.
Diminished Glomerular Expression of Nephrin in Ae1�/�
Mice Is Associated with AlbuminuriaTo examine whether nephrin expression was reciprocally depen-dent on kAE1, we examined glomerular expression of nephrin in12-week-old Ae1�/� and Ae1�/� mouse littermates by immuno-fluorescence staining and Western blotting (Figure 6). kAE1 waspresent in the glomeruli and distal nephron tubules in Ae1�/�
mice but absent in the kidneys from Ae1�/� animals. Nephrinstaining in the glomerulus was more diffusely localized and re-duced in intensity in Ae1�/� mice (Figure 6A), whose glomeruliwere significantly enlarged compared with their wild-type coun-terparts (Ae1�/� glomeruli 147 � 8; Ae1�/� glomeruli 69 � 2;mean cross-sectional diameter �m � SEM; P � 0.0001; n � 18).Immunoblotting of glomeruli-enriched cortical lysates revealedcomparable abundance of nephrin protein in 12-week-oldAe1�/� and Ae1�/� littermates (Figure 6B). However, urine fromAe1�/� and Ae1�/� littermates aged 4 to 6 weeks or 12 weeksdemonstrated by SDS-PAGE abundant 70-kD protein corre-sponding to albumin in two of the three younger and three of thefour older Ae1�/� mice but in none of the Ae1�/� animals (Fig-ure 6C). Thus, although the absence of kAE1 is not associated withdetectable change in total nephrin protein levels, it does result inaltered glomerular distribution of nephrin and variably increasedalbuminuria.
Morphologic Alterations in Glomeruli of Ae1�/� MiceWe examined kidney sections from four pairs of 12-week-oldAe1�/� and Ae1�/� mice by light and electron microscopy for
Figure 2. Nephrin coexpression does not enhance normal kAE1 or mutant kAE1activity in oocytes. Xenopus oocytes are administered an injection of kAE1 cRNA withor without wild-type nephrin cRNAs as indicated. DNDS-sensitive chloride uptake bythe oocytes (10 to 15 per group) over a 20-minute period is measured 24 hours afterinjection. Results are means � SEM. (A) Comparative effects of nephrin cRNA (0.5 to5.0 ng) coexpression on kAE1 activity (0.5 or 1.0 ng of kAE1 cRNA). Nephrin does notincrease kAE1 activity. (B) Nephrin does not rescue the activities of the kAE1G701D orkAE1DelV850 (dRTA mutants) or kAE1S667F (hereditary spherocytosis mutant) previ-ously shown to be rescued partially by GPA.5,7,69
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 21: 1456–1467, 2010 kAE1 in Podocytes 1459
structural changes in the glomerulus. Light microscopy (Supple-mental Figure 4) revealed in Ae1�/� mice glomerular enlarge-ment, moderate to marked mesangial expansion with increasedmatrix deposition, mesangiosclerosis, and mesangiolysis. Preser-vation of podocytes was confirmed in the Ae1�/� glomeruli byimmunofluorescence staining with an additional podocytemarker (synaptapodin) and was accompanied by increased mes-angial cell staining and fibronectin deposition (SupplementalFigure 5).
Transmission electron microscopy (Figure 7) further con-firmed marked mesangial expansion and sclerosis with collagenfibrils and hyaline deposition in all four Ae1�/� mice examined.
At the ultrastructural level, a spectrum ofchanges was observed in the glomerular base-ment membrane (GBM) of the knockout an-imals, ranging from mild to marked, and ir-regular GBM thickening up to 10-fold greaterthan that of Ae1�/� mice. Splitting and lam-ination of the GBM were also frequently ob-served in Ae1�/� mice. The podocyte footprocesses, filtration slits, and endothelial fen-estrations remained largely intact, with onlyfocal areas of effacement. There was no podo-cyte detachment from the GBM.
Nephrin, ILK, and AE1Immunofluorescence Staining Is Alteredin Ae1�/� and Podo-Ilk�/� MiceIn view of the associations of ILK with kAE112
and with nephrin32 and considering the phe-notype of mice with podocyte-specific Ilk de-letion (podo-Ilk�/� mice),32–34 we examinedthe possible in vivo interrelationship amongthe three proteins by confocal immunofluo-rescence microscopy (Figure 8). In wild-typemouse glomeruli (Figure 8, A through F),both nephrin and AE1 demonstrated linearpatterns of staining along the GBM, consis-tent with podocyte staining, and each co-localized with ILK. In the Ae1�/� mouse(Figure 8, g through L), AE1 staining was ab-sent and staining of both ILK and nephrinwas altered. ILK staining appeared diffuseand reduced in intensity. Nephrin stainingwas also more diffuse, consistent with thefindings in Figure 6A. In podo-Ilk�/� mice(Figure 8, M through R), glomerular AE1 andnephrin both exhibited altered staining pat-terns with reduced intensity and patchy dis-tribution throughout the glomerulus. Theseobservations strongly suggest interdepen-dence among ILK, AE1, and nephrin suchthat deletion of any one polypeptide alters ex-pression and/or distribution of the others.
DISCUSSION
This study reports several novel aspects of glomerular biology.It reveals that kAE1 is expressed at low levels in glomerularpodocytes and is dependent on the SD protein nephrin. Ex-pression and localization of nephrin, as well as of ILK, whichforms part of the integrin complex, are similarly dependent onthe presence of kAE1 in the glomerulus. These three proteinsseem to be functionally interconnected and may contribute tothe novel role of glomerular kAE1 in maintaining the glomer-ular filtration barrier to prevent or minimize proteinuria.
Figure 3. kAE1 is expressed in human glomerulus. (Top) A low level of AE1 protein(approximately 95 kD) is detected in human glomerular lysate by Western blot analysisusing two monoclonal AE1 antibodies (BRIC 170 and BRIC 155) that detect both kidney(kAE1) and erythroid (eAE1) isoforms (A and B). The erythroid-specific eAE1 mAb BRAC66 and the erythrocyte GPA antibody BRIC 163 (monomer and dimer, arrowhead)reacted only with the red blood cell membranes and not with glomerular lysate (C andD). Thus, kAE1 is the isoform expressed in the glomerulus and this is not eAE1 (100 kD)from erythrocyte contamination. (Middle) Confocal immunofluorescence microscopy ofnephrin and kAE1 in normal human kidney sections. (E) Nephrin (7C1 mAb) exhibits thecharacteristic ribbon-like pattern of the glomerular capillary loops, consistent withpodocyte staining. (F and G) kAE1 (rabAE1Ct1) displays a more diffuse capillary looppattern in the glomerulus (f), which partially overlaps with nephrin staining (arrows, g).(Bottom) Absence of glomerular immunostaining using the eAE1-specific antibody(BRAC 66; H) is consistent with the glomerular AE1 staining (rabAE1Ct1; I) being thekAE1 isoform. Bar � 150 �m.
BASIC RESEARCH www.jasn.org
1460 Journal of the American Society of Nephrology J Am Soc Nephrol 21: 1456–1467, 2010

We first tested the hypothesis that nephrin may act as achaperonin for kAE1 in the kidney. Although no enhancementof nephrin on kAE1 transport activity in Xenopus oocytes wasdetected, we did observe a dependence of kAE1 expression on
nephrin expression in podocytes, both in immortalized celllines and in intact kidney. We do not know which aspect ofkAE1 trafficking or stabilization in podocytes is dependent onnephrin. In the absence of nephrin, kAE1 may be incorrectly
targeted or once at the plasma membranemay be unstable, leading to increased deg-radation.
Strong evidence for the physiologic sig-nificance of the kAE1/nephrin interactioncomes from the abnormal histology and al-buminuria observed in Ae1�/� mice. Thesemice exhibit severe hemolytic anemia anddRTA, but glomerular abnormalities hadnot been previously reported.40 – 42 Ae1�/�
mice showed mesangiosclerosis and GBMabnormalities but no obvious podocyte ul-trastructural changes. It is possible that themesangial and GBM defects might be sec-ondary to a functional podocyte defect, butthis will need future studies. The markedalterations in nephrin expression in theAe1�/� mouse are unlikely to be simplysecondary to the hemolytic anemia per se,because our analysis of two mouse modelsof hemolysis (sickle cell disease and
Figure 4. kAE1 expression is disrupted in NPHS1Fin Maj human glomeruli. (A through F) A panel of anti-AE1 antibodies (BRIC 170 [Aand D], rabAE1Ct1 [B and E], and rabAE1Ct2 [C and F]) are used. Under wide-field microscopy, glomerular kAE1 distribution in thewild-type kidney (top) displays a diffuse capillary loop pattern, consistent with podocyte staining. In the mutant kidney (bottom), thenormal kAE1 distribution is lost and the intensely stained clusters throughout the glomeruli (white arrows) correspond to eAE1 stainingin clumps of red cells (see Supplemental Figure 2). Intense staining of kAE1 is seen in the connecting tubules surrounding the glomeruliin the top panel (deliberately oversaturated to detect the low glomerular expression of kAE1), whereas its basolateral position in thetype A intercalated cells is best appreciated in D (gray arrow). kAE1 staining in the distal tubule, where nephrin is not expressed, isundisturbed in the mutant kidney sections. (E) Red blood cells, which stain positively for eAE1, can be seen in a vessel lumen (*).Magnification, �40.
Figure 5. Stable endogenous expression of kAE1 in cultured podocytes is dependenton the presence of wild-type nephrin. Immunofluorescence staining of kAE1(rabAE1Ct1, green) and nephrin (7C1, red) in immortalized podocytes. (A) In wild-typecultured podocytes (top), low levels of endogenous kAE1 are detectable at the cellsurface along with faint cytoplasmic and nuclear staining. Nephrin staining is visiblethroughout the cells and at the plasma membrane. Both kAE1 and nephrin expressionare absent in NPHS1Fin Maj mutation podocytes (bottom). (B) Transfection of wild-typenephrin into the wild-type podocytes had little effect on the kAE1 subcellular expres-sion (top), whereas the re-introduction of wild-type nephrin into the mutant podocytesrescues the expression of kAE1 at the cell surface and in the cytoplasm (bottom). Aspreviously noted,5 rabAE1Ct1 gives a faint nuclear stain in the absence of kAE1(bottom left panel of A). Bar � 30 �m.
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 21: 1456–1467, 2010 kAE1 in Podocytes 1461

�-Thalassemia) with glomerular disease phenotypes43,44 dem-onstrated significant nephrin and AE1 expression (Supple-mental Figure 6); however, expression and distribution of bothkAE1 and nephrin were substantially altered in glomeruli ofproteinuric CD2AP�/� mice (Supplemental Figure 7). CD2APbinds nephrin in the SD23,24 and may link it to the cytoskele-ton.45 The loss of this important attachment site may destabi-lize nephrin localization and indirectly affect kAE1 expression.We as yet have not investigated the possibility of a direct inter-action of kAE1 with CD2AP in podocytes.
Importantly, we observed that localization of both ILK andnephrin was altered in Ae1�/� mice. Localization of AE1 andnephrin was similarly altered in podo-Ilk�/� mice, which de-velop GBM defects within 3 to 4 weeks of birth, coinciding withthe onset of albuminuria and preceding the development offoot process effacement and glomerulosclerosis,32–34 and inwhich altered nephrin localization has been previously report-ed.32 The �3�1-integrin–ILK complex located at the basal do-main of the podocyte foot process links the actin cytoskeletonto the extracellular matrix, mediating podocyte attachment tothe GBM, cytoskeleton modulation, and cell–matrix signal-ing.34 The functional links between the SD complex and theintegrin–ILK complex are increasingly appreciated as beingimportant for the functional integrity of the glomerular filter.
Our findings of disrupted glomerular lo-calization of nephrin, ILK, and AE1 in theAe1�/� and podo-Ilk�/� mice, together withpreviously reported interactions of ILK withAE112 and nephrin,32 strongly suggest thatthe three molecules may exist in a multipro-tein complex. Within this complex, kAE1may act as a “functional bridge” connectingthe signal complexes of the SD with those ofthe basal domain, by simultaneously bindingILK12 and nephrin, via its N- and C-terminaldomains, respectively. Although some kAE1may exist in alternative complexes, we specu-late that the multiprotein complex that doescontain kAE1, nephrin, and ILK, may includeother SD and cytoskeletal proteins (e.g.,CD2AP, Nck, Neph1 and Neph2, spectrin),extrapolating from reported interac-tions,19,20,23–26 and by analogy with the inter-actions of eAE1 with multiple erythroidcytoskeleton proteins that modulateerythrocyte mechanical and morphologicproperties.2 Of note, stable kAE1 overexpres-sion in MDCK cells alters paracellular perme-ability and may involve changes in the actincytoskeleton–linked epithelial tight junc-tions.10 It is tempting to speculate that kAE1in podocytes, rather than being required forits transport activity, may play a role in theorganization and/or function of the SD,which has features of both adherens and tight
junctions.46,47 Both kAE1 and nephrin are endocytosed from thecell surface when tyrosine phosphorylated,11,48 suggesting a po-tential mechanism of co-regulation of kAE1 and nephrin duringSD remodeling. Alternatively, the presence of kAE1 may establishpH microdomains in the SD that may potentially modulate pro-tein activity or signaling (e.g., TRPC channels). Thus, the potentialinteractions and role of kAE1 alongside other SD and adaptorproteins merit further study.
Finally, the potential effects of dRTA-associated AE1 muta-tions on glomerular function in humans should be considered.Proteinuria is not a characteristic feature of sporadic dRTA orof dRTA associated with dominant or recessive kAE1 muta-tions in the absence of (nominally) secondary chronic renalinsufficiency or nephrocalcinosis. The proposed underlyingmechanism of dominant dRTA is hetero-oligomerization ofthe mutant with the wild-type kAE1 protein, resulting in mis-localization of the wild-type protein in the type A intercalatedcells, where high levels of basolateral kAE1 are needed for bi-carbonate absorption.10,49,50 Podocytes represent a differentcellular context with distinct interacting proteins (includingnephrin) likely involved in kAE1 targeting/stability. In dRTA,sufficient levels of podocyte kAE1 may be physiologically tar-geted to allow normal function. Albuminuria may nonethelessbe present in some patients with dRTA.51
Figure 6. Nephrin expression is altered with associated albuminuria in Ae1�/� mice.(A) AE1 immunostaining is seen in the glomeruli and distal tubules of Ae1�/� mice andis absent in Ae1�/� animals. Nephrin staining appears in a capillary loop pattern in theAe1�/� glomeruli and is reduced in intensity and more diffuse in the Ae1�/� glomeruli,which are also enlarged compared with wild-type glomeruli. Bar � 20 �m. (B) Westernanalysis demonstrates comparable levels of nephrin protein in the cortical lysates fromAe1�/� and Ae1�/� animals. As expected, kAE1 protein is present in the medullarylysate from the Ae1�/� mice but absent from those of the Ae1�/� animals, in whichonly nonspecific background bands are observed. Cortical and medullary samples arechosen because these regions are enriched in nephrin and AE1 protein expression,respectively. Seventy-five micrograms of total protein was loaded per lane, and �-actinstaining confirmed equal loading. WB, Western blotting. (C) SDS-PAGE of urinesamples demonstrates the presence of a protein band corresponding to albumin(approximately 70 kD) in two of the three 4- to 6-week-old and three of four 12-week-old male Ae1�/� mice. This band is absent in wild-type littermates. Urine loading isnormalized to creatinine excretion, 5 mg/dl loaded per lane.
BASIC RESEARCH www.jasn.org
1462 Journal of the American Society of Nephrology J Am Soc Nephrol 21: 1456–1467, 2010

In conclusion, we have identified kAE1 as a physiologicallysignificant protein in the podocyte, whose newly identified in-teraction with nephrin defines a previously unknown role forkAE1 in the function of the glomerular filtration barrier. kAE1may play a role in the organization or stability of the SD andILK–integrin complexes and in facilitating cross-talk betweensubdomains of the foot process to maintain normal architec-ture under constantly changing glomerular filtration pressuresand for repair of the GBM in response to injury.
CONCISE METHODS
AntibodiesA panel of anti-human AE1 antibodies were used: Mouse monoclonal
BRIC 170 and BRIC 155 that recognized epitopes at the N- and ex-
treme C-termini of AE1, respectively10; an eAE1-specific rat mono-
clonal BRAC 66 antibody that recognized an epitope within 1 to 65
amino acids of eAE152; a mouse monoclonal BRIC 163 antibody
against GPA52; and three rabbit polyclonal antibodies raised against
residues 881 through 911 of AE1 (rabAE1Ct1; rabAE1Ct2)52 or N-
terminal AE1 (65 to 80 amino acids; rabAE1Nt1). The anti-human
nephrin antibodies used were mouse monoclonal (7C1) antibody and
a rabbit polyclonal (K2737) that recognized intracellular C-terminal
domain53 and a rabbit polyclonal antibody (H-300) raised against
amino acids 23 through 322 of the extracellular domain of nephrin
(Santa Cruz Biotechnology, Santa Cruz, CA). Two additional rabbit
polyclonal antibodies were raised against a peptide comprising the last
30 amino acids of nephrin C-terminus (N3 and N4). For the staining
of mouse tissues, polyclonal guinea pig and rabbit anti-mouse AE1,42
polyclonal rabbit anti-mouse nephrin,54 mouse monoclonal anti-
mouse ILK,33 rabbit polyclonal anti-synaptopodin (Santa Cruz Bio-
technology), mouse monoclonal anti–�-smooth muscle actin (Sig-
ma-Aldrich, St. Louis, MO), and rabbit polyclonal anti-fibronectin
(Sigma-Aldrich) antibodies were used.
Species-specific horseradish peroxidase– conjugated antibodies
(DakoCytomation, Cambridge, UK), FITC- and tetramethylrhodam-
ine-labeled antibodies (Jackson Immunoresearch, Philadelphia, PA),
or Alexa fluor 488 – and 594 – conjugated antibodies (Molecular
Probes, Invitrogen, Paisley, UK) were used in Western blotting and
immunofluorescence studies, respectively.
Plasmid ConstructsThe plasmids containing human kAE1 in the mammalian expression
vector pcDNA3.kAE155 and in the Xenopus expression vector
BSXG1.kAE137 have been described previously. pcDNA3.1.AE1mem
(lacking AE1 N-terminal residues 1 through 360) or C-terminal
p3.1eAE1 truncation constructs have been previously published.10,49 The
mammalian plasmid pcDNA3.1.nephrin has also been described.56
BSXG.nephrin was constructed by PCR amplification using Expand Taq
polymerase and pcDNA3.1.nephrin as a template and primers that incor-
porated Nhe1 sites. The resulting PCR product was cloned into pCR2.1-
TOPO vector (Invitrogen), excised, and ligated into BSXG using compat-
ible enzyme sites. NephrinFin Maj encoding the 2-bp frameshift deletion in
NPHS1 (nt121 del 2; NPHS1Fin Maj) was generated using the QuikChange
Site-Directed Mutagenesis kit using pcDNA3.1.nephrin or BSXG.neph-
rin according to the manufacturer’s protocol (Strategene, Amsterdam
Zuidoost, Netherlands). The yeast two-hybrid bait plasmid GBKT7-
AE1Ct, containing the C-terminal residues 877 through 911 of AE1, was
PCR amplified from BSXG1.kAE1 using primers that incorporated an
EcoR1 and Sal1 sites. The resulting PCR product was cloned into pCR2.1-
TOPO vector (Invitrogen), excised using restriction enzyme sites EcoR1
Figure 7. Ae1�/� mice have mesangial expansion and sclerosis and glomerular basement membrane defects. (A through H)Representative transmission electron micrographs from four pairs of 12-week-old Ae1�/� (A and B) and Ae1�/� mice (C through H) areshown. The mesangium (Mes) was abnormal in all Ae1�/� mice examined, with marked expansion, increased matrix accumulation (‡),mesangiosclerosis (black arrowhead), and deposition of hyaline and collagen fibrils (white arrow; C, E, and G). Varying degrees of GBMabnormalities are seen in three of the four mice, ranging from mild uniform (D) to marked thickening (H) up to 10-fold increase inthickness compared with the wild-type (B), with a lamellar appearance and splitting of the lamina densa clearly seen in F and H. Incontrast, the podocyte foot processes (FP) and endothelial fenestrations (EF) in Ae1�/� animals were largely intact, with only focal areasof foot process effacement, subendothelial expansion, and endothelial cell arcade formation (*, g and H). CL, capillary lumen.
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 21: 1456–1467, 2010 kAE1 in Podocytes 1463

and Sal1, and ligated into the bait vector GBKT7. Automated DNA se-
quencing was performed to validate all constructs (Geneservice, Depart-
ment of Biochemistry, Oxford, UK).
Yeast Two-Hybrid ScreenYeast two-hybrid screening was performed using a pretransformed
Matchmaker human kidney cDNA library (BD Biosciences Clontech)
and yeast strains according to the manufacturer’s protocol. The bait
plasmid was transformed into AH109 cells and mated with the pre-
transformed library strain Y187 cells containing human kidney cDNA
in the pACT2 vector. Positive colonies were selected by blue growth
on SD/-Ade/-His/-Leu/-Trp/X-�-Gal plates, which also contained 3
mM 3-amino-1,2,4-triazole. Candidate clones were recovered from
the yeast and retransformed into strain Y187 in the pGADT7 vector
(BD Biosciences Clontech), then mated back to the original AH109
[AE1] bait strains and tested again for growth on SD/-Ade/-His/-
Leu/-Trp/X-�-Gal plates. Positive colony inserts were sequenced and
identified by a BLAST search (http://ncbi.nlm.gov/blast).
Cell Culture and TransfectionHuman embryonic kidney cells (HEK293T) were cultured in DMEM
supplemented with 10% FBS, 100 U/ml penicillin, and 100 �g/ml
streptomycin under 5% CO2 at 37°C. Conditionally immortalized
human wild-type and mutant podocyte cell lines were cultured as
described previously.53,57 The mutant podocytes were derived from a
nephrectomy specimen of a patient homozygous for the NPHS1Fin Maj
frame-shift mutation (nt121 del 2) encoding a truncated 90-residue
nephrin protein. Transient transfection of podocyte cell lines with
wild-type nephrin was performed as described previously.56 Podocyte
transfection was achieved at the permissive temperature of 33°C, and
cells were allowed to differentiate for 14 days after temperature shift.
Nephrin-transfected cells were studied at passages 2 to 3.
Human Kidney TissuesHuman kidney tissue was obtained from a specimen of normal adult
donor kidney unused for transplantation (with institutional ethical
approval, University of Bristol, UK). The cortical tissue was separated
and frozen in liquid nitrogen and stored at �80°C until processed for
immunohistochemistry. Glomeruli were isolated with the sieving
method, as described previously.58
Mouse Kidney TissuesThe generation and characterization of the Ae1�/� mouse model have
been described in detail in previous publications.40,42,59 Ae1�/� mice
were generated by gene targeting, producing a null mutation with no
detectable AE1 peptides in the erythrocyte membrane and in the kidneys.
Ae1�/� animals experienced severe hemolytic anemia and growth retar-
dation at birth with high neonatal mortality. The survival rate was im-
proved to approximately 25% by reducing litter sizes after birth and using
surviving males to continue breeding.42 The kidney samples used in this
study originated from such surviving 12-week-old mice. Detailed renal
phenotyping in the surviving adult Ae1�/� mice showed spontaneous
hyperchloremic metabolic acidosis, elevated serum urea, but normal se-
rum creatinine concentrations and normal creatinine clearance, sugges-
tive of mild systemic dehydration and renal impairment.42 Cortical and
Figure 8. Nephrin, ILK, and AE1 expression is disrupted in bothAe1�/� and podoILK�/� glomeruli, suggesting an interdependencyamong the three proteins. (A through R) Confocal immunofluores-cence microscopy demonstrating the distribution of ILK, AE1, andnephrin in the glomeruli of the wild-type (6 weeks of age; A throughF), Ae1�/� (12 weeks of age; G through L), and podo-Ilk�/� mice (6weeks; M through R). In the wild-type animals, AE1 and nephrinappear in a capillary loop pattern, and the arrows in the mergedimages indicate partial co-localization of ILK with AE1 (C) and neph-rin (F), respectively, in the podocytes. In the Ae1�/� mouse (con-firmed by the absence of AE1 signal in H), staining of ILK (G and J)and nephrin (K) is diffuse and reduced in intensity, and nephrinco-localization with ILK is lost (L). (N and Q) In the podo-Ilk�/� mice,both AE1 and nephrin staining exhibit patchy distribution through-out the glomerulus. The arrowheads denote areas of ILK-deficientpodocytes in which AE1 and nephrin expression is lost. Bar � 15 �min A through C and P through R; 17.24 �m in D through F and Mthrough O; 30 �m in G through L.
BASIC RESEARCH www.jasn.org
1464 Journal of the American Society of Nephrology J Am Soc Nephrol 21: 1456–1467, 2010

medullary membrane protein extracts and kidney cryosections were pre-
pared as described previously.42 Protein quantification was performed
using a modified Lowry assay60 (Bio-Rad DC protein assay; Bio-Rad Lab-
oratories, Hercules, CA), and 75 �g of crude membrane protein was
solubilized in Laemmli sample buffer and analyzed by SDS-PAGE and
Western blotting.
Kidney specimens from 6-week-old podocyte-specific Ilk knockout
(podo-Ilk�/�) mice,33 11- to 12-week-old mouse models of �-Thalasse-
mia44 and sickle cell anemia,43 and 3-week-old CD2AP�/� mice23 were
also used for immunohistochemistry.
Co-immunonoprecipitation StudiesCo-immunoprecipitation experiments were performed as described pre-
viously.61 HEK293T cells were transfected with kAE1 (pcDNA3.1kAE1
or pcDNA3.1e AE1 truncation mutants; 5 �g) either alone or in combi-
nation with the wild-type (pcDNA3.1.nephrin; 5 or 10 �g) or mutant
(pcDNA3.1.nephrinFin Maj
; 5 �g) nephrin plasmids, using the calcium pre-
cipitation.62 After 24 hours of incubation, cells were lysed with immuno-
precipitation buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1% Non-
idet P-40, 1 mM PMSF, 10 g/ml antipain, 10 g/ml leupeptin, 100 U/ml
aprotinin, 50 mM NaF, 1 mM EDTA, and 1 mM orthovanadate) for 30
minutes on ice and clarified by centrifugation at 15,000 � g for 15 min-
utes at 4°C. Cell lysates containing equal amounts of total protein were
precleared with protein A or G-Sepharose, then incubated for 4 hours at
4°C with either protein A or G-Sepharose beads conjugated with the
H-300 or BRIC 170 antibody, respectively. The precipitated complexes
were eluted from the beads with SDS Laemmli buffer and analyzed by
SDS-PAGE and Western blotting using multiple anti-nephrin and anti-
AE1 antibodies, yielding similar results. Additional pulldown experi-
ments were conducted on HEK293T cells transfected with wild-type
nephrin, using the indicated hexahistidine-tagged synthetic C-terminal
kAE1 peptides (synthesized by University of Bristol Peptide Synthesis
facility) in conjunction with His Pur Cobolt beads (Themo Scientific).
Xenopus Oocyte Expression and Chloride TransportThe methods used for the preparation of purified cRNA and expres-
sion in oocytes and assay of 36Cl� uptake were as described previous-
ly.37
SDS-PAGE and Western Blot AnalysisProtein samples were electrophoresed on 10% SDS-polyacrylamide
gels and transferred to polyvinylidene difluoride membranes. The
blots were blocked with 5% milk powder in Tris-buffered saline/0.1%
Tween 20 and sequentially incubated with the appropriate primary
antibodies and horseradish peroxidase– conjugated secondary anti-
bodies for 1 hour at room temperature. Antibody binding was de-
tected by enhanced chemiluminescence analysis (Western Lightning;
Pierce) before exposure to x-ray film (Hyperfilm; GE Healthcare,
Amersham). Urinary protein excretion in Ae1�/� mice was deter-
mined by Coomassie blue staining of polyacrylamide gels after urine
protein electrophoresis.
Immunofluorescence MicroscopyThe immunolabeling of human kidney sections was performed as de-
scribed previously.63 Briefly, 5-�m cryosections were fixed and perme-
abilized with 2.5% paraformaldehyde and 0.3% Triton-100 or in meth-
anol/acetone (6:4 vol/vol). Sections were rinsed with PBS and blocked
with 4% BSA for 30 minutes before application of primary antibodies for
60 minutes at room temperature. Dual labeling was performed sequen-
tially, and sections were washed extensively after each incubation period.
Antigen–antibody complexes were detected with appropriate fluoro-
chrome-conjugated secondary antibodies. Sections were again washed
before mounting with Vectashield (Vector Laboratories, Burlingame,
CA). Standard wide-field images were obtained via a Leica photomicro-
scope attached to a Spot 2 slider digital camera (Diagnostic Instruments)
and were processed with Adobe Photoshop 5.0 software (Adobe, San
Jose, CA). Confocal imaging was conducted using a Leica TCS-NT con-
focal laser-scanning microscope (Leica-Microsystems, Milton Keynes,
UK) using a �63/1.32 oil immersion objective equipped with a Kr/Ar
laser.11
Cultured podocytes were fixed in methanol/acetone, blocked with
4% BSA, serially incubated with rabbit anti-Ct AE1 antibody then
with nephrin antibody 7C1, and followed by detection with suitable
fluorochrome-conjugated secondary antibodies. Cells were mounted
and imaged by confocal laser-scanning microscopy as described al-
ready.
Mouse 5-�m cryosections were fixed and labeled as for human
sections and viewed using Olympus Cell wide-field imaging system,
comprising an IX81 microscope and MT20 illumination unit, CCD
camera, and a �60 1.25 NA oil immersion lens. Confocal images were
taken on a Leica TCS-NT confocal laser-scanning microscope (as de-
scribed already). Images were assembled using Adobe Photoshop 8
software and Adobe Illustrator CS 11 software.
Ae1�/� Kidney Histology and Electron MicroscopyFor histologic analysis under light microscopy, sections were stained
with periodic acid-Schiff, Masson’s, and hematoxylin and eosin stains
using standard methods. For transmission electron microscopy, spec-
imens were fixed in 2.5% glutaraldehyde in 0.1 M phosphate buff-
er64,65 and processed through to embedding in Araldite epoxy resin.66
Ultrathin sections (100 nm) were cut, mounted on 300 mesh copper
grids, and stained with uranyl acetate67 and lead citrate.68 Ultrathin
sections were viewed on a Philips CM10, and images were captured
via SIS iTEM software.
Statistical AnalysisStatistical analysis was performed using the t test.
ACKNOWLEDGMENTS
This work was funded by National Health Service Blood and Trans-
plant Career Development Wellcome Trust Fellowship (A.M.T.),
Medical Research Council (M.A.S.), National Institutes of Health
grants DK43495 and HL000765 (S.L.A.), Swiss National Science
Foundation (3100A0-122217/1), and the 6th EU Frame work project
EUREGENE (C.A.W.).
Portions of this work were presented at the annual meeting of the
American Society of Nephrology; November 4 through 9, 2008; Phil-
adelphia, PA.
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 21: 1456–1467, 2010 kAE1 in Podocytes 1465

We thank Prof. Matthias Kretzler for providing the podo-Ilk�/�
specimens and antibodies to ILK and nephrin, Mario Schiffer for
providing the CD2AP�/� samples, and Prof. Dave Anstee for provid-
ing mAbs to AE1 and GPA. We also thank Rosey Mushens for cultur-
ing mAbs, Debbie Martin for carrying out the cryosectioning, and
Emanuelle Cordat and Prof. Reinhart Reithmeier for the
pcDNA3.1eAE1 truncation constructs.
DISCLOSURESNone.
REFERENCES
1. Kollert-Jons A, Wagner CA, Hubner S, Appelhans H, Drenckhahn D:Anion exchanger-1 in human kidney and oncocytoma differs fromerythroid AE1 in its NH2 terminus. Am J Physiol 265: F813–F821, 1993
2. Tanner MJ: Band 3 anion exchanger and its involvement in erythrocyteand kidney disorders. Curr Opin Hematol 9: 133–139, 2002
3. Alper S: Molecular physiology of SLC4 anion exchangers. Exp Physiol91: 153–161, 2006
4. Williamson RC, Toye AM: Glycophorin A: Band 3 aid. Blood Cells MolDis 41: 35–43, 2008
5. Toye A, Williamson R, Khanfar M, Bader-Meunier B, Cynober T, ThibaultM, Tchernia G, Dechaux M, Delaunay J, Bruce LJ: Band 3 Courcouronnes(Ser667Phe): A trafficking mutant differentially rescued by wildtype band3 and glycophorin A. Blood 111: 5380–5389, 2008
6. Ribeiro ML, Alloisio N, Almeida H, Gomes C, Texier P, Lemos C, MimosoG, Morle L, Bey-Cabet F, Rudigoz RC, Delaunay J, Tamagnini G: Severehereditary spherocytosis and distal renal tubular acidosis associated withthe total absence of band 3. Blood 96: 1602–1604, 2000
7. Bruce LJ, Wrong O, Toye AM, Young MT, Ogle G, Ismail Z, Sinha AK,McMaster P, Hwaihwanje I, Nash GB, Hart S, Lavu E, Palmer R,Othman A, Unwin RJ, Tanner MJ: Band 3 mutations, renal tubularacidosis and South-East Asian ovalocytosis in Malaysia and PapuaNew Guinea: Loss of up to 95% band 3 transport in red cells. BiochemJ 350: 41–51, 2000
8. Hazen-Marin D, Pasternack G, Hennigar R, Spicer S, Sens D: Immu-nocytochemistry of band 3 protein in kidney and other tissues ofcontrol and cystic fibrosis patients. Pediatr Res 21: 235–237, 1987
9. Wainwright S, Tanner M, Martin G, Yendle J, Holmes C: Monoclonalantibodies to the membrane domain of the human erythrocyte aniontransport protein. Biochem J 258: 211–220, 1989
10. Toye AM, Banting G, Tanner MJ: Regions of human kidney anionexchanger 1 (kAE1) required for basolateral targeting of kAE1 inpolarised kidney cells: Mis-targeting explains dominant renal tubularacidosis (dRTA). J Cell Sci 117: 1399–1410, 2004
11. Williamson RC, Brown AC, Mawby WJ, Toye AM: Human kidney anionexchanger 1 localisation in MDCK cells is controlled by the phosphor-ylation status of two critical tyrosines. J Cell Sci 121: 3422–3432, 2008
12. Keskanokwang T, Shandro H, Johnson D, Kittanakom S, Vilas G,Thorner P, Reithmeier R, Akkarapatumwong V, Yenchitsomanus P,Casey J: Interaction of integrin-linked kinase with the kidney chloride/bicarbonate exchanger, kAE1. J Biol Chem 282: 23205–23217, 2007
13. Wu C, Dedhar S: Integrin-linked kinase (ILK) and its interactors: A newparadigm for the coupling of extracellular matrix to actin cytoskeletonand signaling complexes. J Cell Biol 155: 505–510, 2001
14. Legate K, Montanez E, Kudlacek O, Fassler R: ILK, PINCH, and parvin:The tIPP of integrin signalling. Nat Rev 7: 20–31, 2006
15. Tryggvason K, Patrakka J, Wartiovaara J: Hereditary proteinuria syn-dromes and mechanisms of proteinuria. N Engl J Med 354: 1387–1401, 2006
16. Kestila M, Lenkkeri U, Mannikko M, Lamerdin J, McCready P, Ptaala H,Ruotsalainen V, Morita T, Nissinen M, Herva R, Kashtan C, Peltonen L,Holmberg C, Olsen A, Tryggvason K: Positionally cloned gene for anovel glomerular protein—nephrin—is mutated in congenital ne-phrotic syndrome. Mol Cell 1: 575–582, 1998
17. Putaala H, Soininen R, Kilpelainen P, Wartiovaara J, Tryggvason K: Themurine nephrin gene is specifically expressed in kidney, brain andpancreas: Inactivation of the gene leads to massive proteinuria andneonatal death. Hum Mol Genet 10: 1–8, 2001
18. Wartiovaara J, Ofverstedt L, Khoshnoodi J, Zhang J, Makela E,Sandin S, Ruotsalainen V, Cheng R, Jalanko H, Skoglund U, Tryg-gvason K: Nephrin strands contribute to a porous slit diaphragmscaffold as revealed by electron tomography. J Clin Invest 114:1475–1483, 2004
19. Gerke P, Huber T, Sellin L, Benzing T, Walz G: Homodimerization andheterodimerization of the glomerular podocyte proteins nephrin andNEPH1. J Am Soc Nephrol 14: 918–926, 2003
20. Gerke P, Sellin L, Kretz O, Petraschka D, Zentgraf H, Benzing T, WalzG: NEPH2 is located at the glomerular slit diaphragm, interacts withnephrin and is cleaved from podocytes by metalloproteinases. J AmSoc Nephrol 16: 1693–1702, 2005
21. Huber T, Kottgen M, Schilling B, Walz G, Benzing T: Interaction withpodocin facilitates nephrin signaling. J Biol Chem 276: 41543–41546,2001
22. Schwarz K, Simons M, Reiser J, Saleem M, Faul C, Kriz W, Shaw A,Holzman L, Mundel P: Podocin, a raft-associated component of theglomerular slit diaphragm, interacts with CD2AP and nephrin. J ClinInvest 108: 1621–1629, 2001
23. Shih N, Li J, Karpitskii V, Nguyen A, Dustin M, Kanagawa O, Miner JH,Shaw A: Congenital nephrotic syndrome in mice lacking CD2-associ-ated protein. Science 286: 312–315, 1999
24. Shih N, Li J, Cotran R, Mundel P, Miner J, Shaw A: CD2AP localizes tothe slit diaphragm and binds to nephrin via a novel C-terminal domain.Am J Pathol 159: 2303–2308, 2001
25. Verma R, Kovari I, Soofi A, Nihalani D, Patrie K, Holzman L: Nephrinectodomain engagement results in Src kinase activation, nephrinphosphorylation, Nck recruitment, and actin polymerization. J ClinInvest 116: 1346–1359, 2006
26. Jones N, Blasutig I, Eremina V, Ruston J, Bladt F, Li H, Huang H,Larose L, Li S, Takano T, Quaggin S, Pawson T: Nck adaptor proteinslink nephrin to the actin cytoskeleton of kidney podocytes. Nature440: 818–823, 2006
27. Blasutig IM, New LA, Thanabalasuriar A, Dayarathna TK, GoudreaultM, Quaggin SE, Li SS, Gruenheid S, Jones N, Pawson T: Phosphory-lated YDXV motifs and Nck SH2/SH3 adaptors act cooperatively toinduce actin reorganisation. Mol Biol Cell 28: 2035–2046, 2008
28. Winn M, Conlon P, Lynn K, Farrington M, Creazzo T, Hawkins A,Daskalakis N, Kwan S, Ebersviller S, Burchette J, Pericak-Vance M,Howell D, Vance J, Rosenberg P: A mutation in the TRPC6 cationchannel causes familial focal segmental glomerulosclerosis. Science308: 1801–1804, 2005
29. Reiser J, Polu K, Moller C, Kenlan P, Altintas M, Wei C, Faul C, HerbertS, Villegas I, Avila-Casado C, McGee M, Sugimoto H, Brown D, KalluriR, Mundel P, Smith P, Clapham D, Pollak M: TRPC6 is a glomerular slitdiaphragm-associated channel required for normal renal function. NatGenet 37: 739–744, 2005
30. Lehtonen E, Ryan J, Kudlicka K, Iino N, Zhou H, Farquhar M: Celljunction-associated proteins IQGAP1, MAGI-2, CASK, spectrins, andalpha-actinin are components of the nephrin multiprotein complex.Proc Natl Acad Sci U S A 102: 9814–9819, 2005
31. Lehtonen S, Lehtonen E, Kudlicka K, Holthofer H, Farquhar M: Neph-rin forms a complex with adherens junction proteins and CASK inpodocytes and in Madin-Darby canine kidney cells expressing neph-rin. Am J Pathol 165: 923–936, 2004
32. Dai C, Stolz D, Batsacky S, At-Arnauld R, Wu C, Dedhar S, Liu Y:Essential role of the integrin-linked kinase in podocyte biology: Bridg-
BASIC RESEARCH www.jasn.org
1466 Journal of the American Society of Nephrology J Am Soc Nephrol 21: 1456–1467, 2010

ing the integrin and slit diaphragm signaling. J Am Soc Nephrol 17:2164–2175, 2006
33. El-Aouni C, Herbach N, Blattner S, Henger A, Rastaldi M, Jarad G,Miner J, Moeller M, St-Arnaud R, Dedhar S, Holzman L, Wanke R,Kretzler M: Podocyte-specific deletion of integrin-liked kinase resultsin severe glomerular basement membrane alterations and progressiveglomerulosclerosis. J Am Soc Nephrol 17: 1334–1344, 2006
34. Kanasaki K, Kanda Y, Palmsten K, Tanjore H, Lee S, Leblue V, GattoneV Jr, Kalluri R: Integrin �1-mediated matrix assembly and signaling arecritical for the normal development and function of the kidney glo-merulus. Dev Biol 313: 548–593, 2008
35. Sachs N, Kreft M, van den Bergh Weerman M, Beynon A, Peters T,Weening J, Sonnenberg A: Kidney failure in mice lacking the tet-raspanin CD151. J Cell Biol 175: 33–39, 2006
36. Young MT, Tanner MJ: Distinct regions of human glycophorin Aenhance human red cell anion exchanger (band 3; AE1) transportfunction and surface trafficking. J Biol Chem 278: 32954 –32961,2003
37. Groves JD, Tanner MJ: Glycophorin A facilitates the expression ofhuman band 3-mediated anion transport in Xenopus oocytes. J BiolChem 267: 22163–22170, 1992
38. Toye AM, Ghosh S, Young MT, Jones GK, Sessions RB, Ramauge M,Leclerc P, Basu J, Delaunay J, Tanner MJ: Protein-4.2 association withband 3 (AE1, SLCA4) in Xenopus oocytes: Effects of three naturalprotein-4.2 mutations associated with hemolytic anemia. Blood 105:4088–4095, 2005
39. Sterling D, Reithmeier R, Casey J: A transport metabolon: Functionalinteraction of carbonic anhydrase II and chloride/bicarbonate ex-changers. J Biol Chem 276: 47886–47894, 2001
40. Peters L, Shivdasani R, Hanspal M, John K, Gonzlez J, Brugnara C,Gwynn B, Mohandas N, Alper S, Orkin S, Lux S: Anion exchanger1(Band 3) is required to prevent erythrocyte membrane surface lossbut not to form the membrane skeleton. Cell 86: 917–927, 1996
41. Hassoun H, Wang Y, Vassiliadis J, Lutchman M, Palek J, Aish L, Aish IS,Liu S-C, Chishti AH: Targeted inactivation of murine band 3 (AE1)gene produces a hypercoagulable state causing widespread throm-bosis in vivo. Blood 92: 1785–1792, 1998
42. Stehberger P, Shmukler B, Stuart-Tilley A, Peters L, Alper S, Wagner C:Distal renal tubular acidosis in mice lacking the AE1 (Band3) Cl�/HCO�3 exchanger (slc4a1). J Am Soc Nephrol 18: 1408–1418, 2007
43. De Paepe M, Trudel M: The transgenic SAD mouse: A model ofhuman sickle cell glomerulopathy. Kidney Int 46: 1337–1345, 1994
44. Yang B, Kirby S, Lewis J, Detloff P, Maeda N, Smithies O: A mousemodel for �0-thalassemia. Proc Natl Acad Sci U S A 92: 11608–11612,1995
45. Lehtonen S, Zhao F, Lehtonen E: CD2-associated protein directlyinteracts with the actin cytoskeleton. Am J Physiol Renal Physiol 283:F734–F743, 2002
46. Reiser J, Kriz W, Kretzler M, Mundel P: The glomerular slit diaphragmis a modified adherens junction. J Am Soc Nephrol 11: 1–8, 2000
47. Fukasawa H, Bornheimer S, Kudlicka K, Farquhar M: Slit diaphragmscontain tight junction proteins. J Am Soc Nephrol 20: 1491–1503,2009
48. Qin X-S, Tsukaguchi H, Shono A, Yamamoto A, Hidetake K, Doi T:Phosphorylation of nephrin triggers its internalization by raft-mediatedendocytosis. J Am Soc Nephrol 20: 2534–2545, 2009
49. Cordat E, Li J, Reithmeier R: Carboxyl-terminal truncations of humananion exchanger impair its trafficking to the plasma membrane. Traffic4: 642–651, 2003
50. Walsh S, Turner CM, Toye A, Wagner C, Jaeger P, Laing C, Unwin R:Immunohistochemical comparison of a case of inherited distal renaltubular acidosis (with a unique AE1 mutation) with an acquired casesecondary to autoimmune disease. Nephrol Dial Transplant 22: 807–812, 2007
51. Wrong O: Nephrocalcinosis. In: Oxford Textbook of Clinical Nephrol-ogy, 2nd Ed., edited by Davison AM, Cameron JS, Grunfeld JP, Kerr
DN, Ritz E, Winearel CG, Oxford, Oxford University Press, 1998, pp1375–1395
52. Beckmann R, Toye AM, Smythe JS, Anstee DJ, Tanner MJ: An N-terminal GFP tag does not alter the functional expression to theplasma membrane of red cell and kidney anion exchanger (AE1) inmammalian cells. Mol Membr Biol 19: 187–200, 2002
53. Saleem M, O’Hare M, Reiser J, Coward R, Inward C, Farren T, Xing C,Ni L, Mathieson P, Mundel P: A conditionally immortalised humanpodocyte cell line demonstrating nephrin and podocin expression.J Am Soc Nephrol 13: 630–638, 2002
54. Holzman L, St. John P, Kovari I, Verma R, Holthofer H, Abrahamson D:Nephrin localizes to the slit pore of the glomerular epithelial cell.Kidney Int 56: 1481–1491, 1999
55. Toye AM, Bruce LJ, Unwin RJ, Wrong O, Tanner MJ: Band 3 Walton,a C-terminal deletion associated with distal renal tubular acidosis, isexpressed in the red cell membrane but retained internally in kidneycells. Blood 99: 342–347, 2002
56. Coward RJ, Welsh GI, Koziell A, Hissain S, Lennon R, Ni L, Tavare JM,Mathieson PW, Saleem MA: Nephrin is critical for the action of insulinon human glomerular podocytes. Diabetes 56: 1127–1135, 2007
57. Coward R, Foster R, Patton D, Ni L, Lennon R, Bates D, Harper S,Mathieson P, Saleem M: Nephrotic plasma alters slit diaphragm-dependent signaling and translocates nephrin, podocin, and CD2associated protein in cultured human podocytes. J Am Soc Nephrol16: 629–637, 2005
58. Holthofer H, Reivinen J, Miettinen A: Nephron segment and cell-typespecific expression of gangliosides in the developing and adult kid-ney. Kidney Int 45: 123–130, 1994
59. Akel A, Wagner C, Kovacikova J, Kasinathan R, Kiedaisch V, Koka S,Alper S, Bernhardt I, Wieder T, Huber S, Lang F: Enhanced suicidaldeath of erythrocytes from gene-targeted mice lacking the Cl�/HCO3� exchanger AE1. Am J Physiol 292: 1759–1767, 2007
60. Lowry O, Rosebrough NJ, Farr AL, Randall RJ: Protein measurementwith the Folin phenol reagent. J Biol Chem 193: 265–275, 1951
61. Harita Y, Kurihara H, Kosako H, Tezuka T, Sekine T, Igarashi T, SeisukeHattori S: Neph1, a component of the kidney slit diaphragm, is ty-rosine-phosphorylated by the Src family tyrosine kinase and modu-lates intracellular signaling by binding to Grb2. J Biol Chem 283:9177–9186, 2008
62. Graham F, van der Eb A: A new technique for assay of infectivity ofhuman adenovirus 5 DNA. Virology 52: 456–467, 1973
63. Saleem MA, Zavadil J, Bailly M, McGee K, Witherden IR, Pavenstadt H,Hsu H, Sanday J, Satchell SC, Lennon R, Ni L, Bottinger EP, Mundel P,Mathieson PW: The molecular and functional phenotype of glomerularpodocytes reveals key features of contractile smooth muscle cells.Am J Physiol Renal Physiol 295: F959–F970, 2008
64. Sabatini DD, Bensch K, Barrnett RJ: Cytochemistry and electron mi-croscopy: The preservation of cellular ultrastructure and enzymaticactivity by aldehyde fixation. J Cell Biol 17: 19–58, 1963
65. Coetzee J, van der Merwe CF: Some characteristics of the buffervehicle in gluteraldehyde-based fixatives. J Microsc 146: 143–155,1987
66. Glauert AM, Rogers GE, Glauert RH: A new embedding medium forelectron microscopy. Nature 178: 803, 1956
67. Stempac JG, Ward RT: An improved staining method for electronmicroscopy. J Cell Biol 22: 697–701, 1964
68. Reynolds ES: The use of lead citrate at high pH as an electron-opaquestain in electron microscopy. J Cell Biol 17: 208–212, 1963
69. Tanphaichitr VS, Sumboonnanonda A, Ideguchi H, Shayakul C, Brug-nara C, Takao M, Veerakul G, Alper SL: Novel AE1 mutations inrecessive distal renal tubular acidosis: Loss-of-function is rescued byglycophorin A. J Clin Invest 102: 2173–2179, 1998
Supplemental information for this article is available online at http://www.jasn.org/.
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 21: 1456–1467, 2010 kAE1 in Podocytes 1467