analysis ofthedefectsof temperature-sensitive mutantsof ... · inthis paper wereport ona...
TRANSCRIPT

JOURNAL OF VIROLOGY, Mar. 1977, p. 1140-1148Copyright X) 1977 American Society for Microbiology
Vol. 21, No. 3Printed in U.S.A.
Analysis of the Defects of Temperature-Sensitive Mutants ofVesicular Stomatitis Virus: Intracellular Degradation of
Specific Viral ProteinsDAVID KNIPE,' HARVEY F. LODISH, AND DAVID BALTIMORE*
Department of Biology, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139
Received for publication 10 September 1976
The metabolism of viral RNA and proteins has been studied in cells infectedwith temperature-sensitive mutant strains of vesicular stomatitis virus. Certainviral proteins encoded by the mutant strains, usually the putative mutantprotein for the assigned complementation group, were shown to be degradedmore rapidly at the nonpermissive temperature than were the wild-type pro-teins. Group III mutants (tsG33, tsM301) encode M proteins which are degradedthree- to fourfold faster than the wild-type protein. This defect cannot be fullyrescued by coinfection with wild-type virus, and thus the defect appears to be inthe M protein itself. Mutants tsM601(VI) and tsG41(IV) encode N proteins whichare degraded much faster than the wild-type protein and also share the propertyof being defective in replication of viral RNA, suggesting a correlation betweenthese phenotypic properties. Furthermore, the L proteins of tsGll(I) andtsG13(I) are more labile than the wild-type protein at the nonpermissive temper-ature. The G protein of tsM501(V) did not undergo the change in electrophoreticmobility previously shown to be the result of sialylation, suggesting that it isdefective in maturation or glycosylation at the nonpermissive temperature.Three of the mutants previously isolated in this laboratory, tsM502(V),tsM601(VI), and tsM602(VI), were shown to be defective in viral RNA synthesisat the nonpermissive temperature. Mutant tsM601(VI) was defective mainly inviral RNA replication, whereas tsM502(V) appeared to be totally defective forviral RNA transcription and replication at the nonpermissive temperature.
Temperature-sensitive mutants of vesicularstomatitis virus (VSV) have been isolated inseveral laboratories and classified into six com-plementation groups (2, 3, 6, 18, 19, 21). Group Icontains more than one-half of the mutantsisolated, and some of these are defective fortranscription of mRNA species in vivo at thenonpermissive temperature (15, 16, 20, 23). Inaddition, reconstitution studies of the tran-scriptase enzyme showed that the L protein isdefective in these mutants (7, 8). Similar recon-stitution studies have led to the suggestion thatgroup IV mutations are defective in the N pro-tein (14). Mutants in group III are believed tobe defective in the M protein because, in in-fected cultures shifted from the nonpermissiveto permissive temperatures, M protein previ-ously labeled at the nonpermissive temperaturecannot be incorporated into virions (11). GroupV mutations can be complemented by RNAtumor viruses (24), have been shown to be de-fective in maturation of the glycoprotein (11),
I Present address: Committee on Virology, University ofChicago, Chicago, IL 60637.
and thus appear to reside in the gene for Gprotein. No consistent phenotype has been ob-served for the group II mutants.
In this paper we report on a new property ofcertain classes of mutants, lability of the puta-tive mutant viral protein at the nonpermissivetemperature. In addition, we have observedelectrophoretic differences between certain vi-ral proteins of the Glasgow and Massachusettsvirus strains. Analysis of the fate of specificmutant gene products in cultures coinfectedwith two virus strains allowed us to furthercharacterize the nature of the mutations.
MATERIALS AND METHODSGrowth of viruses. Wild-type VSV was prepared
as described previously (9). Mutants of the Glasgowseries and tsO45(V) were obtained from Craig Prin-gle, Glasgow University. Purified B virions fromplaque-purified stocks of these viruses were used inall experiments. Plaque-purified stocks of the Mas-sachusetts mutants were kindly supplied by CarlRettenmier. Stocks were grown by infecting L-cellsat 0.1 PFU/cell and allowing the infection to proceedfor 20 h at 31°C (9). To conserve the plaque-purifiedpreparations, these unpurified stocks were used for
1140

DEGRADATION OF MUTANT VSV PROTEINS 1141
the experiments in this paper. All but two of themutants [tsM502(V) and tsM603(VI)] demonstratedwild-type levels ofRNA synthesis at the permissivetemperature. Labeled viral particles prepared fromcultures infected with these two mutants sedi-mented only as B virions. Thus, there was no evi-dence of T particle contamination of the prepara-
tions.Measurement of virus-specific RNA accumula-
tion. Viral RNA accumulation was studied as de-scribed by Stampfer et al. (22), except that all cul-tures were buffered by the addition of 10 mM TES[N - tris(hydroxymethyl)methyl - 2 -aminoethane-sulfonic acid]-25 mM HEPES (N-2-hydroxyethylpi-perazine-N'-2-ethanesulfonic acid), pH 7.4. Actino-mycin D was added to 5 ug/ml, and [3H]uridine(New England Nuclear Corp.; 5,6-3H, 20 to 40 Ci/mmol) was added to 1 ,Ci/ml at 30 min postinfectionto follow virus-specific RNA accumulation. At 1.5-hintervals, portions of the cultures were transferredinto 5% trichloroacetic acid, and the precipitateswere collected on membrane filters (MilliporeCorp.). The RNA was hydrolyzed from the filterswith 0.5 ml of NH40H, and the radioactivity was
determined by scintillation spectrometry in a diox-ane-based scintillation cocktail.
Sucrose gradient analysis of cytoplasmic virus-specific RNA. Infected cells labeled with [3H]uridinewere lysed in RSB (10 mM Tris-hydrochloride [pH7.41-10 mM NaCl-1.5 mM MgCl2) with 1% NonidetP-40, and nuclei were removed by centrifugation for5 min at 1,000 x g. The cytoplasmic extract was
made 1% with sodium dodecyl sulfate (SDS), layeredon a 15 to 30% sucrose gradient in SDS buffer (0.1 MNaCl-10 mM Tris-hydrochloride [pH 7.51-1 mM
EDTA-0.5% SDS), and centrifuged for 11 h at 27,000rpm at 25°C in a Beckman SW27 rotor. Fractionswere collected by pumping from the bottom of thetube through a flow cell in a Gilford spectrophotom-eter to monitor the absorbance at 260 nm. An equalvolume of 25% trichloroacetic acid and 0.1 mg ofyeast RNA was added to the samples, and the radio-activity in the precipitate was determined as above.
Pulse-chase labeling of viral proteins. Cells were
infected by following the protocol previously de-scribed (9), except that infections were buffered bythe addition of TES-HEPES (pH 7.4) as describedabove. At 4 to 5 h postinfection, the cells were har-vested, washed with Earle saline solution, and re-
suspended at 2 x 106 cells/ml in Earle saline supple-mented with TES-HEPES. The culture was labeledwith 6 to 12 ,Ci of [35S]methionine (New EnglandNuclear Corp.; 2 to 400 Ci/mmol) per ml for 5 min.Unlabeled methionine was added and the incuba-tion was continued for periods up to 60 min. Atspecific times, portions of the infected culture were
removed and kept at 4°C. The cells were lysed di-rectly in this solution by the addition of 0.1 volumeof 10% Nonidet P-40-5% deoxycholate. Nuclei were
removed by centrifugation at 1,000 x g for 5 min.and the proteins were concentrated from the super-
natant by the addition of 9 volumes of acetone. Theprecipitate was dissolved in gel sample buffer, andthe proteins were analyzed by SDS-polyacrylamidegel electrophoresis as described previously (9).
RESULTS
Degradation of specific viral proteins en-coded by temperature-sensitive mutants.While the synthesis of viral proteins in cellsinfected with temperature-sensitive mutant vi-rus strains was being analyzed, it became ap-parent that certain viral polypeptides accumu-lated to much lower levels than in cells infectedwith wild-type virus. To determine whetherthis deficiency was due to decreased rates ofsynthesis or increased rates of degradation, weperformed a series of pulse-chase experiments(Fig. 1 and 2). Cells infected with wild type ortemperature-sensitive strains of virus werepulse-labeled with [35S]methionine for 5 min ateither 31'C or after a temperature shift from 31to 390C. At this time one-half of the infectedculture was removed, and the remainder wasincubated with a large excess of unlabeled me-thionine for 60 min. The viral proteins from thecells and medium were recovered as describedabove and subjected to polyacrylamide gel elec-trophoresis. The wild-type viral proteins werestable during the chase period, except that theM protein was partially degraded at 390C (Fig.1). At 31'C the wild-type M protein was consid-erably more stable. The decrease in electropho-retic mobility of the wild-type G protein duringthe chase period should also be noted. The basisfor this change has been shown to involve theaddition of sialic acid (9).The mutant virus tsM301(III) demonstrated a
lower level of accumulation of the M protein inlong labeling periods at 390C, but when infectedcultures were pulse-labeled for 5 min, the Mprotein was present in approximately normalamounts relative to cells infected with wild-type virus (Fig. 1). After the chase period, theM protein was virtually absent from the cul-ture, indicating a very rapid degradation rate;at 310C the tsM301(III) M protein was muchmore stable.
Quantitation of the percentage of the originalamount of M protein remaining at varioustimes during a 1-h chase period revealed thatthe tsM301(III) M protein had a half-life of 20min at 390C, whereas the M protein of wild typeand other mutant strains had half-lives of 60min (Fig. 3). At 31'C the M proteins of all virusstrains showed less than 20% turnover duringthe 60-min chase period.The M and G proteins of the tsM301(III) virus
also showed a reduced electrophoretic mobilityrelative to the wild-type virus proteins [Fig. 1,wild-type and tsM301(III) coinfection]. The Mand G proteins encoded by tsM302(III) andtsM303(III) showed the same electrophoreticmobilities as the tsM301(II) proteins (D.
VOL. 21, 1977

1142 KNIPE, LODISH, AND BALTIMORE
FIG. 1. Pulse-chase labeling of cultures infected with wild type or temperature-sensitive mutants of VSV.Cultures were infected at a multiplicity of 10 at 310C. At 4 to 5 h postinfection the cells were collected bycentrifugation and resuspended in Earle saline supplemented with TES-HEPES. One-half of each samplewas incubated at 390C and the other halfat 31 0C. After a 10-min warming period the cultures were labeled for5 min with [35S]methionine. At that time one-half of each culture was placed at 0 to 40C, and the remainderwas incubated with excess unlabeled methionine for 60 min. The cells were lysed directly in this solution bythe addition of 0.1 volume of10% Nonidet P-40-5% deoxycholate. Nuclei were removed by centrifugation at1,000 x g for 5 min, and the proteins were concentrated from the supernatant by the addition of9 volumes ofacetone. The precipitate was dissolved in gel sample buffer, and the proteins were analyzed by gel electropho-resis. Exposure times: wt, 24 h; wt + tsM301(III), 36 h; tsG33(III), 390C, 36 h; 310C, 24 h; tsG41(IV),48 h; tsM601(VI), 24 h.
Knipe, Ph.D. thesis, Massachusetts Institute ofTechnology Cambridge, 1976), and thus it be-came necessary to explain how alterations inthe mobility of two proteins could be caused bya temperature-sensitive mutation. An identicaldifference in the electrophoretic mobilities ofthe proteins of our wild-type virus andtsG33(III) has been found (Knipe, Ph.D. the-sis), and we now believe that tsM301(III),tsM302(III), and tsM303(III) all represent acci-dental reisolates of tsG33(III). The tsG33(III),like tsM301(III), encoded an M protein with anincreased degradation rate at 39°C, yet it wasmore stable at 31°C (Fig. 1). The increaseddegradation rate of the M protein was not ageneral property of the Glasgow virus strainsbecause other temperature-sensitive mutantsavailable to us did not show this feature [e.g.,tsGll(I), tsG13(I), and tG41(IV); Fig. 1 and 2].To determine whether the degradation of the
M protein in cells infected with tsM301(III) wasdue to the lack of some stabilizing factor or to adefect intrinsic in the protein which made it
more susceptible to intracellular proteases,cells were coinfected with wild-type virus andtsM301(III) [we will continue to use the termtsM301(III), even though we suspect that thevirus may actually be tsG33(III)]. ThetsM301(III) M protein, the upper band of thedoublet in the M region, was degraded at afaster rate than the wild-type M protein at39°C, yet at 31°C the two proteins were presentat equal levels after the chase period (Fig. 1).Due to the proximity of the two bands (see Fig.5), it was difficult to determine whether therate of degradation of the tsM301(III) M proteinwas exactly the same as in singly infected cul-tures. However, it was obvious that the degra-dation rate of the tsM301(III) M protein wasmuch faster than the M protein of the wild-typevirus and that the wild-type virus could notrescue the M protein encoded by the mutantvirus. Therefore, we conclude that the M pro-tein of tsM301(III), presumably due to a defectin the molecule, is more sensitive to intracellu-lar proteases at the nonpermissive tempera-
J. VIROL.

DEGRADATION OF MUTANT VSV PROTEINS 1143
ts G 13(I)390 310
ts M 501 (V)390 31°
ts M502(V)39" 310
_ _ m
0 60 0 60 0 60 0 60 0 60 0 60 0 60 0 60minutes of chase
FIG. 2. Pulse-chase labeling of cultures infected with temperature-sensitive mutants. These experimentswere performed as described in the legend to Fig. 1. Exposure times: tsG11 (I), 24 h; tsG13(I), 24 h; tsM501 (V),390C, 72 h; 310C, 24 h; tsM502(V), 36 h.
lo0t
80
60
40
-
's 200
I OC tsM60180-t
- 0sM301 tsM5O160 IVA
0 20 40minutes of chose
FIG. 3. Rate of degradation of the M proteins en-
coded by various virus strains. A pulse-chase experi-ment similar to the one described in the legend to Fig.1 was performed, except that portions of the culturewere removed at 0, 15, 30, and 60 min of chase. Thecytoplasmic and extracellular proteins were concen-
trated and subjected to polyacrylamide gel electro-phoresis. The amount of M protein in each samplewas determined and expressed as a percentage of theamount present at the beginning of the chase.
ture. This conclusion is consistent with the pre-vious suggestion that group III mutations arein the M protein (11).Mutants were also found that coded for N
proteins which were degraded rapidly at thenonpermissive temperature. In cells infectedwith tsG41(IV), the N protein was present inthe cells at normal levels after the 5-min pulse-label, but decreased greatly during the chaseperiod at 390C, indicating a rapid rate of degra-dation (Fig. 1). However, at 310C the proteinwas quite stable. Similarly, cells infected withtsM601(VI) showed a nearly complete loss of theN protein during the 60-min chase period at390C, but the protein was very stable at 3100.Determination of the amount of N protein re-maining at various periods of chase in cellsinfected with these mutants revealed that at390C the N proteins had half-lives from theinitial slope of degradation of approximately 10min. Cultures infected with wild-type VSV andother mutants showed degradation of less than20% of the N protein during the 60-min chaseperiod at 390C. In addition, all of the proteinsshowed less than 15% degradation at 310C (Fig.4). The other mutant that showed this pheno-type of a rapidly degrading N protein wastsG12(I) (data not shown)-a group I mutant
ts Gll (I)390 310
sM601
NN ~~~~~~~~~wt- sG41- tsM50
x~~~~~~~~l
VOL. 21, 1977
)_

1144 KNIPE, LODISH, AND BALTIMORE
' 201000
-tsG4I
tsM66
0 20 40 60minutes of chase
FIG. 4. Rate of degradation of the N proteins en-coded by various virus strains. The amount of Nprotein in each of the samples from the experimentdescribed in the legend to Fig. 3 was also quantitatedand expressed as a percentage of the amount presentat the beginning of the chase.
that nevertheless is defective for replication of40S viral RNA (16). Therefore, these three mu-tants shared the property of a thermolabile Nprotein and a defect in replication of viral RNA,suggesting a link between these two phenotypicproperties.
Figure 2 shows examples of another subclassof group I mutants. Pulse-chase experimentswith tsGll(I) and tsG13(I) revealed that the Lprotein was lost at 39°C, whereas at 31°C theprotein was quite stable. This is consistent withthe previous assignment of the defect in group Imutants to the L protein (7, 8). The data for thegroup II mutants are not shown, but we ob-served no special protein degradation for eithertsG21(II) or tsG22(II). For the group V mutant,tsM501, the M protein was degraded at a rateslightly faster than wild type but slower thantsM301(III) (Fig. 2 and 3). In addition, a more
striking defect was evident: at 39°C the G pro-tein did not undergo the change in mobilityascribed previously to the addition of sialicacid. This will be described in more detail in theaccompanying paper (9), but it suggests a defectin the G protein. Mutant ts 045(V) showed simi-lar behavior of the G protein (data not shown).An additional property of tsM501(V) as well asother group V mutants was that incorporationof [35S]methionine into viral proteins was five-to tenfold higher in cells labeled at 31°C than incells labeled immediately after a shift from 31to 39°C (Fig. 2). We are unsure whether thisreflects an alteration in protein synthesis or inuptake of the amino acid label.For the mutant tsM502(V), the M protein was
degraded at a rate nearly that of tsM301(III),and there was no obvious defect in maturationof the G protein. However, analysis of this mu-tant is further complicated by the fact that ithas a defect in RNA synthesis and nucleocapsidassembly (see Table 1 and reference 9).
In each of the experiments described in thissection, the cells were lysed directly in theEarle saline used as the labeling medium so asto recover all of the proteins that may havebeen incorporated into extracellular viral parti-cles. In some sets of samples, presumably dueto acetone precipitation of the crude cytoplasm,a small amount of radioactively labeled mate-rial remained at the top of the gel. This did not,however, affect the quantitation of the relativeamounts of the viral proteins because similarresults were obtained from duplicate samplesthat contained less labeled material at the ori-gin. We do not believe the degradation of spe-cific viral polypeptides described here was dueto any artifact of labeling or lysing the cells inEarle saline because similar results on the defi-ciencies of specific viral proteins were observedin cells labeled in complete medium lackingmethionine and washed carefully prior to dis-solving the total cells in gel sample buffer (9).
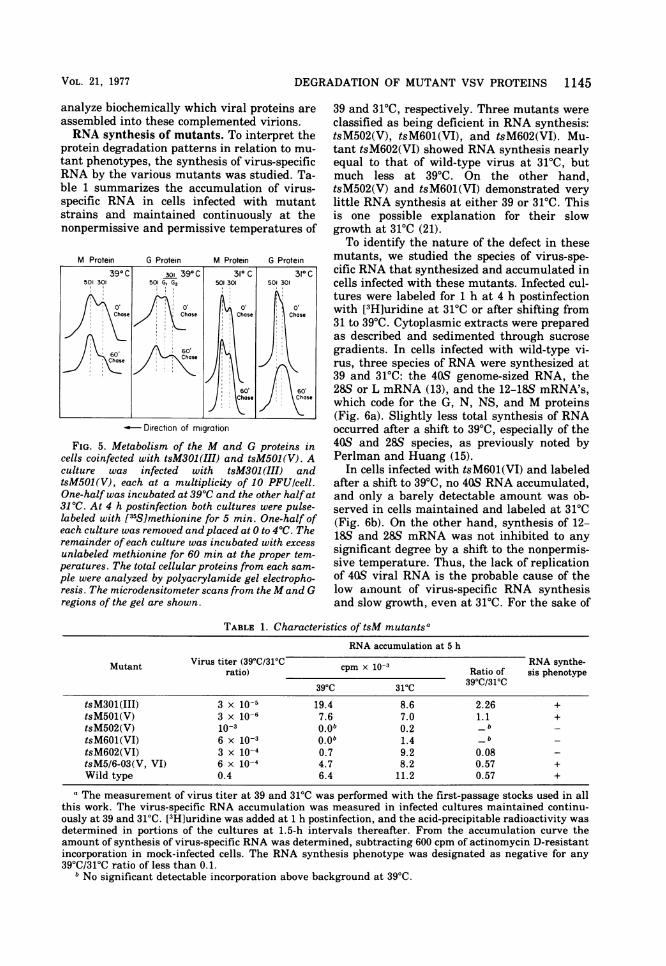
Biochemical analysis of the M and G pro-teins in cells coinfected with tsM301(III) andtsM501(V). Because of the mobility differencebetween the M and G proteins of tsM301(III)and tsM501(V), we were capable of resolvingthe gene products of these two mutants by gelelectrophoresis even in doubly infected cells. Ithad been shown previously that the two mu-tants are capable of complementation, so weexamined the fate of the specific gene productsin cells coinfected with the two virus strains. At390C the M protein of tsM301(III) was degradedat a faster rate than the tsM501(V) M protein(Fig. 5), whereas at 31'C the two proteinsturned over at the same rate. Also, at 390C theG protein of the tsM501(V) virus did not un-dergo the change of mobility associated withthe addition of sialic acid, whereas the G pro-tein of tsM301(III) did change in mobility dur-ing the 60-min chase period. At 30'C both pro-teins shifted in mobility. Therefore, it appearedthat the defects in tsM501(V) and tsM301(III)were in the G and M proteins, respectively, andnot in trans-active secondary proteins. We wereable to demonstrate genetic complementationin these cells coinfected with tsM301(III) andtsM501(V), but we have been unable to detectany increase in viral proteins in total extracel-lular viral particles (data not shown), presum-ably due to uninfectious particles produced bythe cells. Therefore, we have been unable to
J. VIROL.
DEGRADATION OF MUTANT VSV PROTEINS 1145
analyze biochemically which viral proteins are
assembled into these complemented virions.RNA synthesis of mutants. To interpret the
protein degradation patterns in relation to mu-tant phenotypes, the synthesis of virus-specificRNA by the various mutants was studied. Ta-ble 1 summarizes the accumulation of virus-specific RNA in cells infected with mutantstrains and maintained continuously at thenonpermissive and permissive temperatures of
-Direction of migration
FIG. 5. Metabolism of the M and G proteins incells coinfected with tsM301(III) and tsM501(V). Aculture was infected with tsM301 (III) andtsM501(V), each at a multiplicity of 10 PFU/cell.One-half was incubated at 39°C and the other halfat31°C. At 4 h postinfection both cultures were pulse-labeled with [35S]methionine for 5 min. One-half ofeach culture was removed and placed at 0 to 4°C. Theremainder of each culture was incubated with excess
unlabeled methionine for 60 min at the proper tem-peratures. The total cellular proteins from each sam-ple were analyzed by polyacrylamide gel electropho-resis. The microdensitometer scans from theM and Gregions of the gel are shown.
39 and 31°C, respectively. Three mutants wereclassified as being deficient in RNA synthesis:tsM502(V), tsM601(VI), and tsM602(VI). Mu-tant tsM602(VI) showed RNA synthesis nearlyequal to that of wild-type virus at 31°C, butmuch less at 39°C. On the other hand,tsM502(V) and tsM601(VI) demonstrated verylittle RNA synthesis at either 39 or 31°C. Thisis one possible explanation for their slowgrowth at 31°C (21).To identify the nature of the defect in these
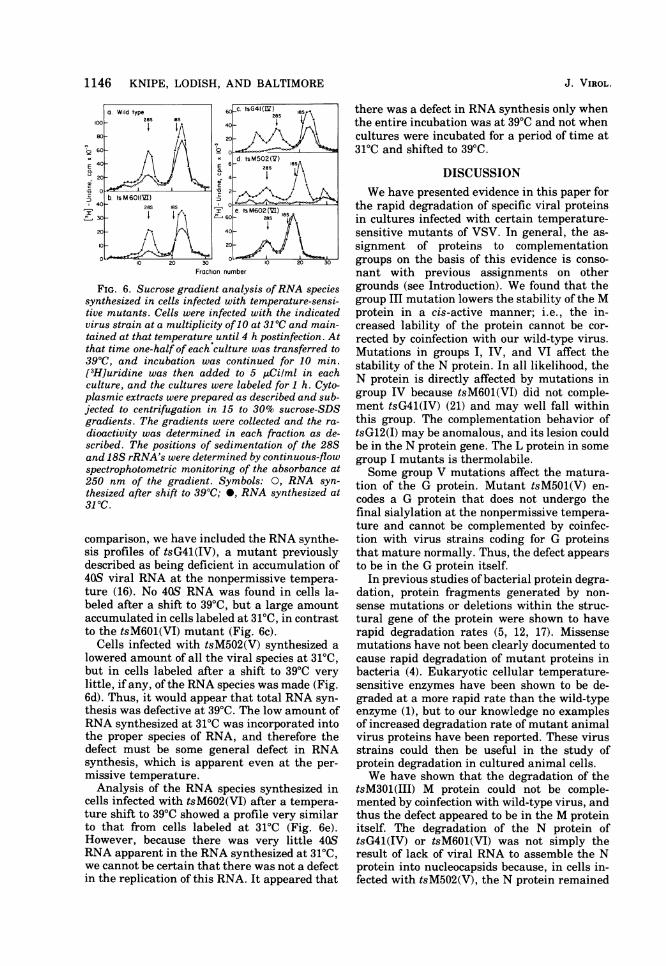
mutants, we studied the species of virus-spe-cific RNA that synthesized and accumulated incells infected with these mutants. Infected cul-tures were labeled for 1 h at 4 h postinfectionwith [3H]uridine at 31°C or after shifting from31 to 39°C. Cytoplasmic extracts were preparedas described and sedimented through sucrosegradients. In cells infected with wild-type vi-rus, three species of RNA were synthesized at39 and 31°C: the 40S genome-sized RNA, the28S or L mRNA (13), and the 12-18S mRNA's,which code for the G, N, NS, and M proteins(Fig. 6a). Slightly less total synthesis of RNAoccurred after a shift to 39°C, especially of the40S and 288 species, as previously noted byPerlman and Huang (15).
In cells infected with tsM601(VI) and labeledafter a shift to 39°C, no 40S RNA accumulated,and only a barely detectable amount was ob-served in cells maintained and labeled at 31°C(Fig. 6b). On the other hand, synthesis of 12-18S and 28S mRNA was not inhibited to anysignificant degree by a shift to the nonpermis-sive temperature. Thus, the lack of replicationof 40S viral RNA is the probable cause of thelow amount of virus-specific RNA synthesisand slow growth, even at 31°C. For the sake of
TABLE 1. Characteristics of tsM mutantsa
RNA accumulation at 5 h
Mutant Virus titer (39C/31C cpm x 10-3 a RNA synthe-ratio) Ratio of sis phenotype39°C 310C 39°C/31°C
tsM301(III) 3 x 10-5 19.4 8.6 2.26 +tsM501(V) 3 x 10-6 7.6 7.0 1.1 +tsM502(V) 10-3 O.Ob 0.2 _ btsM601(VI) 6 x 10 3 O.Ob 1.4 b _tsM602(VI) 3 x 10-4 0.7 9.2 0.08 -tsM5/6-03(V, VI) 6 x 10-4 4.7 8.2 0.57 +Wild type 0.4 6.4 11.2 0.57 +
a The measurement of virus titer at 39 and 310C was performed with the first-passage stocks used in allthis work. The virus-specific RNA accumulation was measured in infected cultures maintained continu-ously at 39 and 310C. [3H]uridine was added at 1 h postinfection, and the acid-precipitable radioactivity wasdetermined in portions of the cultures at 1.5-h intervals thereafter. From the accumulation curve theamount of synthesis of virus-specific RNA was determined, subtracting 600 cpm of actinomycin D-resistantincorporation in mock-infected cells. The RNA synthesis phenotype was designated as negative for any390C/313C ratio of less than 0.1.
b No significant detectable incorporation above background at 390C.
VOL. 21, 1977
1146 KNIPE, LODISH, AND BALTIMORE
100
a. Wild type28S 8ss
l I I
b ts M 601Cz)28S 20
10 20 30
I
E
-I
,0-c tsG4 AV Iss
2Sa
10-0 3
Fraction number
FIG. 6. Sucrose gradient analysis ofRNA speciessynthesized in cells infected with temperature-sensi-tive mutants. Cells were infected with the indicatedvirus strain at a multiplicity of10 at 31 0C and main-tained at that temperature until 4 h postinfection. Atthat time one-half of each culture was transferred to390C, and incubation was continued for 10 min.[3H]uridine was then added to 5 ,tCilml in eachculture, and the cultures were labeled for 1 h. Cyto-plasmic extracts were prepared as described and sub-jected to centrifugation in 15 to 30% sucrose-SDSgradients. The gradients were collected and the ra-
dioactivity was determined in each fraction as de-scribed. The positions of sedimentation of the 28Sand 18S rRNA's were determined by continuous-flowspectrophotometric monitoring of the absorbance at250 nm of the gradient. Symbols: 0, RNA syn-thesized after shift to 390C; 0, RNA synthesized at31 0C.
comparison, we have included the RNA synthe-sis profiles of tsG41(IV), a mutant previouslydescribed as being deficient in accumulation of40S viral RNA at the nonpermissive tempera-ture (16). No 40S RNA was found in cells la-beled after a shift to 390C, but a large amountaccumulated in cells labeled at 31TC, in contrastto the tsM601(VI) mutant (Fig. 6c).
Cells infected with tsM502(V) synthesized alowered amount of all the viral species at 31'C,but in cells labeled after a shift to 390C very
little, if any, of the RNA species was made (Fig.6d). Thus, it would appear that total RNA syn-thesis was defective at 390C. The low amount ofRNA synthesized at 310C was incorporated intothe proper species of RNA, and therefore thedefect must be some general defect in RNAsynthesis, which is apparent even at the per-
missive temperature.Analysis of the RNA species synthesized in
cells infected with tsM602(VI) after a tempera-ture shift to 390C showed a profile very similarto that from cells labeled at 31'C (Fig. 6e).However, because there was very little 40SRNA apparent in the RNA synthesized at 310C,we cannot be certain that there was not a defectin the replication of this RNA. It appeared that
there was a defect in RNA synthesis only whenthe entire incubation was at 390C and not whencultures were incubated for a period of time at31'C and shifted to 390C.
DISCUSSIONWe have presented evidence in this paper for
the rapid degradation of specific viral proteinsin cultures infected with certain temperature-sensitive mutants of VSV. In general, the as-signment of proteins to complementationgroups on the basis of this evidence is conso-nant with previous assignments on othergrounds (see Introduction). We found that thegroup III mutation lowers the stability of the Mprotein in a cis-active manner; i.e., the in-creased lability of the protein cannot be cor-rected by coinfection with our wild-type virus.Mutations in groups I, IV, and VI affect thestability of the N protein. In all likelihood, theN protein is directly affected by mutations ingroup IV because tsM601(VI) did not comple-ment tsG41(IV) (21) and may well fall withinthis group. The complementation behavior oftsG12(I) may be anomalous, and its lesion couldbe in the N protein gene. The L protein in somegroup I mutants is thermolabile.Some group V mutations affect the matura-
tion of the G protein. Mutant tsM501(V) en-codes a G protein that does not undergo thefinal sialylation at the nonpermissive tempera-ture and cannot be complemented by coinfec-tion with virus strains coding for G proteinsthat mature normally. Thus, the defect appearsto be in the G protein itself.
In previous studies of bacterial protein degra-dation, protein fragments generated by non-sense mutations or deletions within the struc-tural gene of the protein were shown to haverapid degradation rates (5, 12, 17). Missensemutations have not been clearly documented tocause rapid degradation of mutant proteins inbacteria (4). Eukaryotic cellular temperature-sensitive enzymes have been shown to be de-graded at a more rapid rate than the wild-typeenzyme (1), but to our knowledge no examplesof increased degradation rate of mutant animalvirus proteins have been reported. These virusstrains could then be useful in the study ofprotein degradation in cultured animal cells.We have shown that the degradation of the
tsM301(III) M protein could not be comple-mented by coinfection with wild-type virus, andthus the defect appeared to be in the M proteinitself. The degradation of the N protein oftsG41(IV) or tsM601(VI) was not simply theresult of lack of viral RNA to assemble the Nprotein into nucleocapsids because, in cells in-fected with tsM502(V), the N protein remained
80
b 60
E40CL
20
-60
40I
30
20
10
0
J. VIROL.

DEGRADATION OF MUTANT VSV PROTEINS 1147
soluble and did not assemble into nucleocapsidsat 39TC (9), yet it was not rapidly degraded(Fig. 2). In addition, at 31'C in cells infectedwith tsM601(VI) the N protein is quite stable,yet replication of viral RNA is very low (Fig. 6).This suggests that the degradation of the Nprotein is not a result of the absence of stable40S RNA. The converse relationship -that theabsence of stable 40S RNA is due to defective Nprotein -seems probable and suggests two pos-sible explanations for the lack of 40SRNA inthese cells. The defect in the N protein couldprevent the synthesis of 40SRNA at the non-permissive temperature, or the lack of accumu-lation of 40SRNA could be due to the inabilityof the N protein to encapsidate the 40S RNAproperly, leaving the RNA susceptible to rapiddegradation by intracellular nucleases. Wehave been unable to find any 40S RNA aftershort pulse-labels in these cells (data notshown), but it is not possible by these experi-ments to completely exclude rapid degradation.In vitro systems having the capacity for replica-tion as well as transcription will be required toanswer this question.Our observation that tsM601(VI) resembles
tsG41(IV) in several phenotypic propertiesraises the question of the present status of com-plementation group VI. Mutant tsM602(VI) ap-peared to be defective for RNA synthesis wheninfected cultures were maintained continuouslyat 390C, but when cultures were initially in-fected at 31'C and shifted to 390C prior to label-ing, no defect was apparent in the RNA speciessynthesized. This conclusion is subject to thereservation that we have been able to detectvery little 40S RNA in cells infected withtsM602(VI), even at the permissive tempera-ture. The proteins of tsM602(VI) show no in-creased lability, a further difference fromtsM601(VI). The double mutant tsM5/6-03(V,VI) shows no defect in RNA synthesis, but theG protein does not undergo the change in mo-bility associated with the addition of sialic acid(data not shown). This latter property is proba-bly the result of the group V mutation. Thus, inspite of the apparent similarity betweentsM601(VI) and tsG41(IV), the heterogeneousphenotypes associated with the group VI muta-tions suggest that group VI may truly be sepa-rate from the other complementation groups.
ACKNOWLEDGMENTSWe gratefully acknowledge the technical assistance of
Martin Brock.D.K. was supported by a National Science Foundation
predoctoral fellowship during part of this work and a PublicHealth Service traineeship during the remainder. H.F.L.was the recipient of Public Health Service research careerdevelopment award GM-50175 from the National Institute
of General Medical Sciences. D.B. is an American CancerSociety research professor. This work was supported byPublic Health Service grants AI-08814 and AI-08388 fromthe National Institute of Allergy and Infectious Diseases,American Cancer Society grant E559, and Public HealthService grant CA-12174 from the National Cancer Institute.
LITERATURE CITED
1. Capecchi, M., N. Capecchi, S. Hughes, and G. Wahl.1974. Selective degradation of abnormal proteins inmammalian tissue culture cells. Proc. Natl. Acad.Sci. U.S.A. 71:4732-4736.
2. Flamand, A. 1969. Etude des mutants thermosensiblesdu virus de la stomatite vesiculaire mise en pointd'un test de complementation. C.R. Acad. Sci. Paris268:2305-2308.
3. Flamand, A. 1970. Etude genetiques du virus de lastomatite vesiculaire: clasement de mutants thermo-sensibles spontanes en groupes de complementation.J. Gen. Virol. 8:187-195.
4. Goldberg, A. L., and J. F. Dice. 1974. Intracellularprotein degradation in mammalian and bacterialcells. Annu. Rev. Biochem. 43:835-869.
5. Goldschmidt, R. 1970. In vivo degradation of nonsensefragments in E. coli. Nature (London) 228:1151-1156.
6. Holloway, A. F., P. K. Y. Wang, and D. V. Cormack.1970. Isolation and characterization of temperature-sensitive mutants of vesicular stomatitis virus. Virol-ogy 42:917-926.
7. Hunt, D. M., S. Emerson, and R. R. Wagner. 1976.RNA-negative temperature-sensitive mutants of ve-sicular stomatitis virus: L protein thermosensitivityaccounts for transcriptase restriction of group I mu-tants. J. Virol. 18:596-603.
8. Hunt, D. M., and R. R. Wagner. 1974. Location of thetranscription defect in group I temperature-sensitivemutants of vesicular stomatitis virus. J. Virol. 13:28-35.
9. Knipe, D., D. Baltimore, and H. F. Lodish. 1977. Matu-ration of viral proteins in cells infected with tempera-ture-sensitive mutants of vesicular stomatitis virus.J. Virol. 21:1149-1158.
10. Knipe, D., H. F. Lodish, and D. Baltimore. 1977. Local-ization of two cellular forms of the vesicular stomati-tis viral glycoprotein. J. Virol. 21:1121-1127.
11. Lafay, F. 1974. Envelope proteins of vesicular stomati-tis virus: effect of temperature-sensitive mutations incomplementation groups III and V. J. Virol. 14:1220-1228.
12. Lin, S., and I. Zabin. 1972. ,B-Galactosidases rates ofsynthesis and degradation of incomplete chains. J.Biol. Chem. 247:2205-2211.
13. Morrison, T., M. Stampfer, D. Baltimore, and H. F.Lodish. 1974. Translation of vesicular stomatitis vi-rus mRNA by extracts from mammalian and plantcells. J. Virol. 13:62-72.
14. Ngan, J. S. C., A. F. Holloway, and D. V. Cormack.1974. Temperature-sensitive mutants of vesicular sto-matitis virus: comparison of the in vitro polymerasedefects of group I and group IV mutants. J. Virol.14:765-772.
15. Perlman, S., and A. S. Huang. 1973. RNA synthesis ofvesicular stomatitis virus. V. Interactions betweentranscription and replication. J. Virol. 12:1395-1400.
16. Perlman, S. M., and A. S. Huang. 1974. Virus-specificRNA specified by the group I and IV temperature-sensitive mutants of vesicular stomatitis virus. Inter-virology 2:312-325.
17. Platt, T., J. Miller, and K. Weber. 1970. In vivo degra-dation of mutant lac repressor. Nature (London)228:1154-1156.
18. Pringle, C. R. 1970. Genetic characteristics of condi-tional lethal mutants of vesicular stomatitis virus
VOL. 21, 1977

1148 KNIPE, LODISH, AND BALTIMORE
induced by 5-fluorouracil, 5-azacytidine, and ethylmethane sulfonate. J. Virol. 5:559-567.
19. Pringle, C. R. 1970. The induction and genetic charac-teristics of conditional lethal mutants of vesicularstomatitis virus, p. 567-582. In R. D. Barry and B. W.J. Mahy (ed.), The biology of large RNA viruses.Academic Press Inc., London.
20. Printz-Ane, C., A. Conbard, and C. Martinet. 1972.Study ofthe transcription and the replication of vesic-ular stomatitis virus by using temperature-sensitivemutants. J. Virol. 10:889-895.
21. Rettenmier, C., R. Dumont, and D. Baltimore. 1975.Screening procedure for complementation-dependent
mutants of vesicular stomatitis virus. J. Virol. 15:41-49.
22. Stampfer, M., A. Huang, and D. Baltimore. 1969. Ribo-nucleic acid synthesis of vesicular stomatitis virus. I.
Species of ribonucleic acid found in Chinese hamsterovary cells infected with plaque-forming and defec-tive particles. J. Virol. 4:154-161.
23. Unger, J. T., and M. E. Reichmann. 1973. RNA synthe-sis in temperature-sensitive mutants of vesicular sto-matitis virus. J. Virol. 12:570-578.
24. Zavada, J. 1972. Pseudotypes of vesicular stomatitisvirus with the coat of murine leukemia and of avianmyeloblastosis viruses. J. Gen. Virol. 15:183-191.
J. VIROL.