analysis and simulation modelling of population dynamics and bioenergetics ofcryptolestes...
TRANSCRIPT

Res. Popul. Ecol. (1990)32, 235--254. ~) by the Society of Population Ecology
ANALYSIS AND S I M U L A T I O N M O D E L L I N G OF P O P U L A T I O N
DYNAMICS AND BIOENERGETICS OF CR YPTOLESTES FERR UGINEUS
(COLEOPTERA: CUCUJIDAE) IN STORED W H E A T 1
A. CAMPBELL 2 and R. N. SINHA
Agriculture Canada Research Station, 195 Dafoe Road, Winnipeg, Manitoba, R3T 2M9, Canada
INTRODUCTION
The rusty grain beetle, Cryptolestesferrugineus (Stephens) (Coleoptera: Cucujidae)
is a cosmopolitan insect pest of grain (Sinha and Watters, 1985). A common and
serious pest of stored grain on farms and elevators in the Prairie Provinces of Canada
(Sinha, 1965a; Loschiavo, 1975; Liscombe and Watters, 1962; Smith and Barker,
1987), C. ferrugineus is often found in heating grain (Sinha and Wallace, 1966) and
causes direct damage to wheat by consuming the wheat germ (Munro, 1969; Campbell
and Sinha, 1976). An extensive bibliography on this species has been compiled by
Throne (1987).
Cryptolestes ferrugineus cannot penetrate sound grain and relies on a broken or
damaged seed coat (bran layer) to reach the wheat germ on which the insect feeds
almost exclusively. Normally, one wheat germ can provide sufficient energy and
nutrients for complete larval development of one individual C. ferrugineus under op-
timal environmental conditions (Rillett, 1949; Campbell and Sinha, 1978). Although
superficially sound, most stored wheat-grain masses contain some kernels with broken
seed coats which are prone to infestation by this insect (Sinha and Watters, 1985). A
wheat-grain bulk is a short-lived and immature, human-subsidized ecosystem with a
relatively simple structure and nonregenerative food-energy supply (Sinha, 1973;
Odum, 1989). The deteriorative process of wheat in bulk storage, however, is relative-
ly complex, involving the interaction of various physical, chemical and biotic
variables. Outbreaks of C. ferrugineus have been followed in granaries containing
wheat (Sinha and Wallace, 1966; Smith, 1978, 1983). Early changes in many biotic
and abiotic variables associated with these outbreaks, however, have neither been fully
understood, nor have the pathways of energy-flow in a population of C.ferrugineus been explored.
Although C. ferrugineus is the most important insect pest of stored grain in
1 Contribution No. 1314 from Agriculture Canada Research Station, Winnipeg, Manitoba, R3T 2M9, Canada 2 Present address: Department of Fisheries and Oceans, Pacific Biological Station, Nanaimo, British Col-
umbia, V9R 5K6, Canada

236
Canada, no mathematical model simulating the population energetics of this species
has been attempted. Such models based on sound laboratory data are a prerequisite
to constructing comprehensive models of Western Canadian stored-grain ecosystems
that can be used to analyse ecosystem dynamics and advise farmers on sound manage-
ment of stored-grain pests. Recent developments of simulation modeis for beetles in-
festing wheat include the influence of various temperature and relative humidity
changes on the population dynamics of C. ferrugineus (Kawamoto et al., I989) and
Sitophilus oryzae (L.) (Hardman, 1978; Cuff and Hardman, 1980; Longstaff and Cuff,
1984).
Our objectives were to describe changes in the major biotic, bioenergetic,
chemical and physical variables in a stored-wheat ecosystem, and to develop and use a
simple computer simulation model to follow in detail the insect population energetics
during the progress of a C. ferrugineus infestation.
MATERIALS AND METHODS
Design and Sampling Procedures
Four 70-1 steel drums were each filled with 54• kg of clean Canada Western
red spring wheat No. 1, Triticum aestivum (L.) (cv. Neepawa) with 13.1-13.9%
moisture content. Each drum, placed horizontally, had four sampling points: one at
the top, one at each side and one at the bottom as described and illustrated earlier (van
Bronswijk and Sinha, 1971). The top outlet (diam 6 cm) was covered with No. 50
mesh copper-wire netting (aperture 0.3 mm) and filter paper which allowed gas ex-
change but prevented insects from escaping. Other outlets (diam 3 cm) were closed
with rubber stoppers. The drums were held in a room at 29.5-30.5~ and 30-50%
RH for 222 d (=days). Two drums of wheat without insects served as controls (con-
trol system). The other two drums each had I000 laboratory-reared (at 29.5-30.5~
and 68-72% RH on whole wheat) C. ferrugineus adults which were introduced through
the top-central opening of each drum at the beginning of the experiment (Cf system).
We chose a near optimum constant temperature level (30~ for development and
survival of C. ferrugineus populations (Rillett, 1949; Smith, 1965) to simulate
temperatures that often occur in the fall and early winter when grain is harvested on
hot days and stored in large bulks (Smith, 1983). Wheat moisture content was kept at
about 13.5% (most years Canadian grain is stored at 13.0-14.4% moisture content),
at the low end of the optimal range for C. ferrugineus population growth (Rillett, 1949;
Smith, 1965), to reduce microfloral activity (Christensen, 1982). Therefore, the only
major energy source available to the beetles would be the wheat germ, accessible
through the broken bran layers in naturally damaged wheat kernels, though C. fer- rugineus can also feed and thrive on some stored-product fungi (Sinha, 1965b;
Loschiavo and Sinha 1966).
Temperature was recorded to • I~ every 4 h at each of the four sampling loca-

237
tions (15 cm deep in the grain mass) using copper-constantan thermocouples in con-
junction with a multipoint electronic recorder.
One grain sample of 200 ml was taken from each of the four locations in each
drum at days 0, 13, 27, 45, 66, 87, 108, 137, 165, and 222; sampling dates were
somewhat irregular because of convenience in processing samples and measuring
many variables. The grain samples were collected from the bottom and two side loca-
tions by removing the rubber stoppers and allowing the wheat to flow into the sample
jar. A sample from the top-central location was taken with a brass tropedo probe
previously sterilized in alcohol and burned dry. During each sampling about 1% of
the total grain bulk was removed.
A subsample of 50 ml was taken from each sample and further subdivided to deter-
mine the values of several variables. The moisture content was determined on a wet-
mass basis using 5 g samples according to the American Association of Cereal
Chemists' method (Anon., 1962). Germination was determined and the microfloral
association was noted on 25 randomly selected kernels using the filter-paper method
(Wallace and Sinha, 1962). The weight and volume of two groups of 100 randomly
selected kernels was determined. For the Cf (Cryptolestesferrugineus) system, another
two groups of 100 randomly selected kernels were placed in plastic bags, frozen at
- -20~ and, at a later date, the kernels were thawed and dissected with a scalpel under
a binocular microscope to determine the number of each C. ferrugineus life stage that
was present. All life stages were recorded; the four larval instars were staged accor-
ding to Campbell and Sinha (1978). A total of 1400-1600 kernels from the Cf systems
(100-200 kernels per sample) were dissected for each sampling period. Only 10
kernels from each subsample of the control systems (total = 80) were dissected for each
period. Seed damage of all kernels that were subsequently dissected were noted for
the presence or absence of cracks in the bran on the wheat germ area. Also, both germ
and endosperm damage were arbitrarily classified into five categories: (1) no damage,
(2) 0.25 or a quarter, (3) 0.5 or one-half, (4) 0.75 or three-quarters, (5) 1.0 or complete-
ly eaten (Campbell and Sinha, 1976). Consumption in joules was estimated by summ-
ing the number of kernels in each germ damage category multiplied by the estimated
wheat energy lost in each damage category: (1) zero, (2) 6.4J, (3) 12.7 ] , (4) 19.1 J and
(5) 25.4J. The dissected wheat kernels were also weighed after being dried at 70~
for 48 h. Respiration of immatures was determined from another group of 100 ran-
domly chosen kernels over 12h at 3 0 + 1 C in a constant-volume Warburg
respirometer using the methods of Campbell and Sinha (1978) and Campbell et al.
(1976). The oxycaloric equivalent factor for converting from 02 to calories respired
was 0.005047 (Brody 1964) and to joules was 4.186J (= 1 cal).
In the control system, the mean dry weight of a kernel (28.5 mg+__0.08 SE; range
26.6-29.9 mg) measured from groups of 100 kernels, did not change significantly. In
the Cf system, the mean dry weight of a kernel (28.4 + 0.07 SE, range 26.6-29.7, after
dissection 28.1-----0.13 SE, range 22.5-29.7) did not change significantly during the

238
study because the variance in the weight of whole kernels was large enough to mask
any loss in weight due to the removal of the small germs.
The remaining 150 ml of wheat from each sample was sieved with 2.00 mm and
0.42 mm aperture sieves and then placed in a Berlese funnel for 24 h for extraction of
the mobile larval and adult stages (Sinha, 1964). The live and dead insects were
counted and larvae classified according to instar (Campbell and Sinha, 1978), the
volume of grain was determined and the dust oven-dried at 70--+2~ for 48h and
weighed. Fat acidity values, FAV (mg K O H required to neutralize the free fatty acids
extracted from 100 g of dry grain) were determined in duplicate on pooled samples
from each drum for each sampling date (AACC method 02-01, Anon., 1962).
Since most immature C. ferrugineus were found in the wheat germ cavities during
dissections, the mean number of larvae and pupae per 100 kernels was calculated from
the dissection of up to 1600 kernels for all the samples obtained in one sampling
period. In contrast, most of the adults were outside the kernels so the mean number of
adults per 100 kernels was obtained from the combination of the sieving and Berlese
funnel extractions. The data were divided by the grain volume of the sample and
multiplied by 4.2 ml (mean/volume of 100 kernels including intergranular spaces).
The dust weight per 100 kernels was also calculated by dividing the dust weight (mg)
by the grain volume of the sample and multiplying by 4.2. Egestion could not be ac-
curately determined from the measured dust. The insect biomass (joules) was
calculated from the product of the mean number of insects per 100 kernels, the mean
biomass (mg) and caloric value (cal/mg) of the equivalent larval, pupal or adult stage
using the energetic values reported by Campbell and Sinha (1978) multiplied by
4.186J ( = 1 cal). The mean dry weight of 100 kernels was 2.85 g. The mean energy
content of wheat germ was 25.449J/kernel, 5~0 of the energy content of a wheat
kernel.
Statistical Analysis
One way analysis of variance (ANOVA) (Nie et al., 1975) was used to compare in-
dividual variables at different locations and/or means between the control and Cf
systems for each sampling period and between periods. Distributions of the variables
were normalized by transforming the insect numbers and biomass using log10 (x+ 1),
temperature using ~-x and the other variables using arcsine ~xx prior to analysis of
variance. Where there were no significant differences (p > 0.05) between sample loca-
tions for a sample period, the data were often pooled to provide a single mean for either
the Cf and control systems for illustration purposes. The SD of the means are shown
in most figures except one where SE are shown for illustration convenience. Both SD
and SE were calculated from raw untransformed data .
Simulation Model
A simple deterministic simulation model, written in F O R T R A N 66, was

239
developed to simulate an average population of C. ferrugineus feeding on a specified
amount of energy. The universe was assumed to be 100 kernels. Published develop-
ment and survival rates (Smith, 1966), fecundity (Smith, 1962, 1963) and energy
budget (Campbell and Sinha, 1978) at 30~ and 70% RH were used as input and in-
trinsic variables in the model. Since the present study was conducted at a constant
30~ and 13.5% grain moisture content, which remained relatively constant
throughout the experiment, and the only published energy budget values for C. fer- rugineus available are at similar environmental conditions, we did not include any
feedback mechanisms in the model to accommodate the effect of temperature or
relative humidity changes on development, reproduction, survival or energetic rate
changes. The model was essentially a bookkeeping device maintaining records of
various changes in characteristics. The model followed an initial population number
of known-age individuals (i.e. 0.06 adults/100 kernels that are 45 d old) with an initial
amount of available food (i.e. 914.6J). Calculations were made for growth, respira-
tion, egestion and biomass for each life stage as the population was incremented every
half day over a specified period (i.e. 222 d). Several assumptions were made in the
model. The duration of the immature stages were egg=4.0 d; first instar larvae, La
=4.0 d; second instar larvae, L2=3.5 d; third instar larvae, L~=3.5 d; fourth instar
larvae, L4=6.0 d; L4 prepupa-----2.5 d and pupa=6.0 d (Campbell and Sinha, 1978).
Adult longevity (emergence from pupa to natural death) was 140.8 d (preoviposi-
t ion= 1.4 d, oviposition= 123.3 d, postoviposition= 16.1 d) (Smith, 1966). A beetle
was assumed to live up to 155 d (including immature and adult stages); most of the
postoviposition period was excluded because most important reproductive events hap-
pened before day 150. The maximum number of eggs produced per female per day
was 5.6 for the first 30 d and an average of 2.5 during the subsequent 93 d of adult life
(Smith, 1963, 1966). Fecundity reduced with increased adult density according to the
following formula calculated from data provided by Smith (1966):
E=4 .59F -~ (r2 =0.94, N=4)
where E = the number of eggs per female per day (EN 5.6 at low beetle densities) and
F = t h e density of female adults (assuming an equal 1 : 1 sex ratio) or the number of
adult females per 1.5 ml of grain. Since the model assumes 100 kernels was 4.2 ml,
the density of all females was prorated for 1.5 ml (i.e. divided by 2.8) prior to
estimating E; subsequently E was reduced by half for the half day iteration.
The fraction of food energy (joules) to be eaten (V) from the current available food
(A) and amount of food desired (D) by the larvae and adults was estimated from a sim-
ple negative exponential or random search model (Thompson, 1924):
V=A (1 -- e-(D/A~).
A newly hatched larva died at once if insufficient food was available to reach
adulthood; if food supply was adequate, the amount of energy required was immediate-

240
ly reserved for the entire larval feeding period. Most developing and feeding larvae
could effectively exclude other larvae and adults from entering the wheat germ cavity
by filling the entrance hole with debris (Rillett, 1949). The number of adults that fed
or starved each half day was determined from the above formula. Adults were allowed
to starve for up to 15 d before dying irrespective of adult age and density (Smith,
1966). However, only those adults that had eaten were considered to respire, egest
and produce eggs for that half day. In most insects respiration rates fall during starva-
tion (Keister and Burk, 1974). Additional mortality on the various life stages could be
produced in two ways: 1) a fixed natural mortality fraction could be applied on each
stage; the remaining uneaten reserved food of larvae that died would be returned back
to the available food supply, but the energy content of the dead larvae was lost; and 2)
a cannibalism fraction could be applied to the various immature stages; both the
energy content of the dead immatures and uneaten reserved food would be returned to
the available food energy supply. The cannibalism fraction (P) depended on the larval
density according to the following formula derived from data provided by Smith
(1966):
P=0.1180+0.2301 1oge S (r2=0.92, N=7)
where P = mortality fraction (0 < p < 0.98) and S= density or number of all larvae per
1.5 ml. The density of larvae was prorated for 1.5 from 4.2 ml (100 kernels) prior to
estimating P.
Caution is required in extrapolating the results of this study, conducted in small
grain bulks under constant laboratory conditions, to large grain masses in granaries
under natural fluctuating ambient conditions.
RESULTS AND DISCUSSION
Description of Abiotic and Biotic Changes
In general, the variables in the insect-free control system did not show any
changes over time except for the decline of the field fungus Alternaria in the first half, a
slight increase in storage-fungi AspergiUus glaucus group in the second half of the study,
and a steady increase in FAV. Consequently, only averages are shown for most
variables in the control system to compare graphically with the Cf-infested systems.
Physical changes. In the control system, the temperature remained constant, reflec-
ting the ambient room temperature of about 30~ In the Cf system, however, there
was a slight but significant (p_< 0.01) increase to a maximum average of 31.7 ~ on day
66 and subsequently a gradual decline to follow ambient room temperatures after day
108 (Fig. 1A). Because temperature or moisture content between locations in a drum
did not differ consistently, the data for each variable were pooled. Moisture content
increased only slightly but significantly (pN0.01) higher at about 13.8% for the Cf
system compared to 13.4% for the control system by day 66 and subsequently remain-
241
T e m p e r a t u r e r A i C f S y s t e m
~ . . . . . . . . ,', "" , ~ L _ c o n t r o l s y s t e m ~ ~ [ . . . . . . . . . I . . . . . . . . . [ . . . . . . . I . . . . . . . . I ' ' '
M o i s t u r e C o n t e n t B
1 4 C f S y s t e m
C o n t r o l S y s t e m 1 2 I ' ' ' ' ' . . . . I . . . . . ' ' ' ' 1 ' ' ' ' ' ' ' ' ' 1 ' ' ' ' ' . . . . i ' ' ' ' q
0 50 100 150 200 D a y s
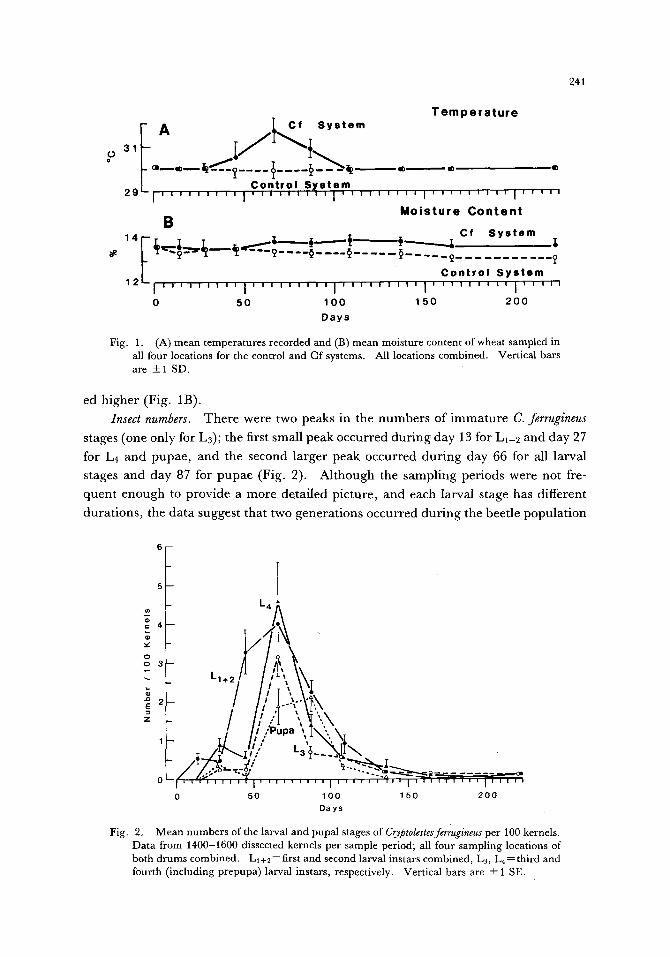
Fig. 1. (A) mean temperatures recorded and (B) mean moisture content of wheat sampled in all four locations for the control and Cf systems. All locations combined. Vertical bars are __+ 1 SD.
ed h igher (Fig. 1B).
Insect numbers. T h e r e were two peaks in the n u m b e r s of i m m a t u r e C. ferrugineus
stages (one only for L3); the first small peak occur red du r ing day 13 for L1-2 and day 27
for L4 and pupae , and the second larger peak occur red du r ing day 66 for all larval
stages and day 87 for pupae (Fig. 2). A l though the sampl ing periods were not fre-
quen t e n o u g h to provide a more detailed picture , and each larval stage has different
dura t ions , the da ta suggest that two genera t ions occur red du r ing the beetle popu la t ion
19 4
19
O o 3
19 . o E
Z
Fig. 2.
2 L1+2/~/
0 . . . . . . , , , i , , , , ~, 0 50 1 0 0 150 2 0 0
Days
Mean numbers of the larval and pupal stages of Cryptolestesferrugineus per 100 kernels. Data from 1400-1600 dissected kernels per sample period; all four sampling locations of both drums combined. Ll+2=first and second larval instars combined, L3, L4=third and fourth (including prepupa) larval instars, respectively. Vertical bars are + 1 SE.

242
growth phase; total developmental time can range from 29 to 43 d (Smith, 1966).
Decreased survival and growth rates on different life stages due to factors such as over-
crowding, quality and quanti ty of food may have influenced the t iming of the second
peak of each stage especially after the first generation or day 30.
The Cryptolestes population peaked for the total immature stages on day 66 (Fig.
3A) and for adults on day 87 (Fig. 3B). The analysis of data using A N O V A indicated
that there were no significant differences between locations except between top and bot-
tom in the number of immatures per 100 kernels (Fig. 3A). There were significantly
larger numbers of live adults (p_<0.01) in the bottom locations compared with the
20[- A Bottom-~--~T j Live I m m a t u r e s
1 +
50 - B
L i v e A d u l t s
4 0 -
3o
o t o 20
m
=,0 , /
J: 5 0 - - c . a d A d u l t s
4 0 - -
2 0 - -
1 0 - -
~..~_.~...---.-.. . . . . . . . . . . . . . . . ~ . . . . . __-~__~._--. o ..... ~ .... I ......... I ......... I ......... I " e,-~
50 100 150 2 0 0
Days
Fig. 3. Mean numbers of (A) total live immatures, (B) live adults and (C) dead adults of Cryp- tolestesferrugineus per 100 kernels sampled at the top, middle and bot tom of both d rums in the Cf system. Vert ical bars are --+ 1 SD.

243
other two locations; there were no differences (p )0 .05) in numbers of adults between
the side and top locations for all sample periods (Fig. 3B). Adults of C. ferrugineus are
positively geotactic and exhibit highly dispersive activity in wheat of low moisture con-
tent, whereas these beetles will reduce their locomotory activity and aggregate in
pockets of damp grain (Surtees, 1965; Watters, 1969). The highest number of C. fer- rugineus was also found by other similar studies at the bottom of grain bulks (White
and Sinha, 1980). After the peak beetle populations growth, a considerable number
of adults died and most were found accumulated at the bottom of the drums (Fig. 3C).
Insect bioenergetics. The estimated biomass of all live insect stages (except eggs)
peaked on day 87 and declined thereafter (Fig. 4A). The daily respiration of the im-
matures peaked on day 66 (Fig. 4B), reflecting the peak number of third and fourth in-
star larvae per 100 kernels (Fig. 2) which are known to have the highest respiration rate
120
100
80
60
40
20
0
Live Biomass of Immatures & Adults A
I Observed -
"''1... . . . . . ,
' ' ' ' ' ' ' ' ' 1 . . . . . . . . . I . . . . . . . . . I ' ' ' . . . . . . I ' ' '
12
8 o o ~ 6
4
~ 2
~ 0
1 0 0 0
8 0 0
6 0 0
4 0 0
2 0 0
0
B immature Resp i ra t ion
,J �9 -
L .:/\,
, , ,, ,, ,, ,,i , ,, , , , , , ,i ,-r-,'r-,-;-, ,, i ,, , , , , , ,, i, , ,, ,
-c . . . . . . . . . . I
~.,-,'~, .:" ~ .......... . . . . . . . . . I ' ' I '' . . . . . . I'''
50 100 150 2 0 0
Days
Fig. 4. (A) mean live biomass of immature and adult C. f e r rug ineus , (B) mean daily immature respiration and (C) mean cumulated consumption in joules "per I00 kernels. The solid lines are from observed data means (dots, vertical bars are --_+ 1 "SD) combined from all loca- tions-of both drums of the Cf system, The dashed lines are the predicted lines from the computer simulation model using combination B in Table 1.

244
of the various life stages of C. ferrugineus (Campbell and Sinha, 1978).
Only the wheat germ of the kernels was consumed. The greatest increase in
energy consumption by the beetles was during days 50-100, reaching a plateau
thereafter (Fig. 4C). The total cumulative mean consumption in joules (914.6--+42.3
SE, range 740.9-1145.3J/100 kernels) occurred on day 222 which represents the
utilization of a mean 35.9% (range 24.1-45.0~ of the total wheat germ available
(2545.1 J) or 1.8% (range 1.5-2.3%) of the total energy (50,893.4J) available in 100
kernels.
Microflora. The field fungus, Alternaria alternata, was present on nearly 50% of all
seeds examined initially but, by day 45, the fungus disappeared in both the control and
Cf systems (Fig. 5A). Thereafter, Aspergillusglaucus group infested up to a mean 21%
in the control system and 13.5% in the Cf system (Fig. 5B). The incidence of
Aspergillus was significantly (p_<0.01) lower in the Cf system than the control system
at days 108 and 137, probably because the beetles were facuhatively feeding on the
fungi (Sinha, 1965b; Loschiavo and Sinha, 1966). The incidence of Penicillium spp.
remained below a mean 17% in both systems except at the end of the study when
Penicillium reached a maximum of 22% on day 222 in the Cf system (Fig. 5C). The
low incidence of storage fungi and bacteria was presumably due to the relatively low
moisture contents of the two systems (Christensen and Kaufmann, 1969).
Deteriorative changes. Levels of damaged germ per 100 kernels increased from a
mean of 1.7 on day 27 to 31.6 on day 108 and 35.7 on day 222 (Fig. 6A). The observ-
m
6 0 -
40
2 0 - -
0
'IE
A
s , . t . m
o / n t r ~ f Alternaria
I . . . . . . . . . I ' ' . . . . . . . I . . . . . ' ' ' ' 1 ' . . . . . ' ~ ' 1 ' ' ' ' ' A s p e r g i l l u s
. . . . .
. . . . . . . . . . I . . . . . . . . . I . . . . . . . . . 1 . . . . .
C Penlctttium I
o T_ _I ~_-~_ _~. .~p.~- . . . . ~ . . . . . ~ . : . . ~ . . . . . ~ , . . . . ? . . . . . . . . . . I . . . . . . . . ~ 1 ' ' " . . . . . . I . . . . . . . . . I . . . . . . . . . I ' ' ' ~ ' 0 50 100 150 200
D a y s
Fig. 5. Mean percent kernels infected with (A) Alternaria, (B) Aspergillus, and (C) with Penicillium in the Cf and control systems. Vertical bars are + 1 SD.

245
ed m e a n n u m b e r of ke rne l s wi th cracks in the b r a n , po t en t i a l l y a l lowing C. ferrugineus access to the w h e a t g e r m , was 51.8+---4.65 S D ( r a n g e 41-67) p e r 100 kerne l s in bo th the
C f a n d con t ro l sys tems (Fig . 6A).
A l t h o u g h g e r m i n a t i o n o f i n c u b a t e d seeds f rom the con t ro l sys tem was a lmos t
100% t h r o u g h o u t the s tudy , g e r m i n a t i o n s t a r t ed dec l i n ing in the C f sys tem f rom
9 4 . 5 % on d a y 27 to 6 5 . 5 % on d a y 107 to a to ta l m e a n low of 6 0 . 5 % on d a y 222 (Fig .
6B). T h e c u m u l a t i v e loss in g e r m i n a t i o n fo l lowed the s ame p a t t e r n as d a m a g e to
w h e a t g e r m s (Fig. 6A) a n d c o n s u m p t i o n (Fig . 4C) in the C f sys tem.
= 60
.= E o o O ~ 2 0
o o .
.~ 0
_ A Kerne ls with Cracks In Bran Near Germ
, j ~ I-- "~ j ~ / ' Complete Germ Eaten
2O
E l= A w w 15
U) c
u ~ .c 0 10
~ w s
0
~ & , . . - - u
I . . . . . . . . . I . . . . . . . . . I . . . . . . . . . I ' ' ' . . . . . . I . . . . .
B C o n t r o l S y s t e m ~oo .~ .... ~_ ~ ___~ ..... { .......... .}
_~ sol- " t ~ ~ \ T I ~ o, , , . , .m
I . . . . . . . . . 1 . . . . . . . . . I . . . . . . . . . I ' ' ' . . . . . . l . . . . .
C
~ �9 . . . . ~ 2 e
-_ ._ - ,%-=, - - , -_ , :-:-,-, :-:-.--: : . . . . . ; , - _ _ z _ 7 , . �9 " " " ,oi ~ ' t ' , ~ o r " ~ = 7 , , t , '
F D cf s y s t e m
. . . . . . . . e - " - " o n t r o l S y a t e
0 50 100 150 2 0 0 D a y s
Fig. 6. (A) mean number of kernels with damaged bran on the wheat germ area in control system (dotted line) and with all germ eaten for all locations combined in the Cf system (solid line), (B) mean percent of kernels germinating in the Cf and control systems, (C) mean weight of dust per 100 kernels sampled at the top, middle and bottom of both drums in the Cf system, (D) mean fat acidity values from both the Cf and control systems. Means are for all locations combined. Vertical bars are + 1 SD.

246
Dus t we igh t was p r o n o u n c e d on ly at the b o t t o m loca t ions , re f lec t ing a d i s p r o p o r -
t i ona t e a c c u m u l a t i o n of a d u l t insects in the b o t t o m (Fig . 6C) . D u s t we igh t at the s ide
a n d top levels d i d no t exceed m o r e t h a n 2.5 mg /100 kerne ls , w he re a s dus t we igh t at the
b o t t o m i n c r e a s e d s lowly to 2.5 m g b y d a y 45, t hen rose sha rp ly to p e a k at 16 m g on d a y
108 a n d l eve led off at 9-11 rag.
T h e F A V in bo th the C f a n d con t ro l sys tems s tead i ly rose f rom 10 to 19-22 d u r i n g
the s t u d y pe r i od ; the va lue of the con t ro l sys tem was u s u a l l y 2-3 un i t s lower t h a n the
C f sys tem wi th no ove r l ap af ter d a y 40 (Fig . 6D). F u n g i were p r o b a b l y r e spons ib le in
the r ise in the F A V due to the i r h igh l ipo ly t ic c a p a c i t y ( S o r g e r - D o m e n i g g et a l . , 1955).
S i m u l a t i o n of C. ferrugineus P o p u l a t i o n Ene rge t i c s
T h e s i m u l a t i o n m o d e l p r e d i c t e d the t i m i n g a n d levels of peak n u m b e r s pe r 100
kerne l s of insec t life s tages a n d b i o e n e r g e t i c va r i ab l e s as closely as was poss ib le to the
o b s e r v e d va lues (e .g . Fig . 4) when a p p l y i n g a f ixed 7 0 % m o r t a l i t y on eggs a n d a dens i -
t y - d e p e n d e n t c a n n i b a l i s m on the second l a rva l i n s t a r ( T a b l e 1, c o m b i n a t i o n B). T h e
Table 1. Results from the Cryptolestesferrugineus population simulation model under different condi- tions of food availability, egg mortality and larval stage on which cannibalism, due to larval densi- ty, is applied. Day on which the peak of a certain category occurs is shown in brackets. Lz=seeond instar larva, L4p=fourth instar prepupa. Basic universe is 100 wheat kernels.
Condition and result Combination
A B C D E
Condition Initial energy available, joules 914.6 914.6 914.6 914.6 2545.1 Egg mortality, ~00 10 70 70 70 70 Cannibalism on larval stage Lz L 2 L~o none L 2
Result Peak mortality due to cannibalism, % 63.3 51.2 59.6 0 68.1
(day) (45) (62) (61) 0 (86) Insect biomass eaten, joules 7.5 5.0 50.2 0 23.4 Adult numbers peak 18.9 17.6 11.3 19.7 39.4
(day) (75) (88) (90) (85) (109) Fecundity (eggs/female/day) ~ 2.6 2.7 3.3 5.6 d 1.9 Biomass peak, joules b 74.5 74.5 55.3 95.9 154.9
(day) (73) (87) (78) (77) (109) Cumulated total, joules c
Consumption 922.2 919.7 964.9 914.6 2568.5 Assimilation 616.2 612.4 644.2 600.3 1740.5 Respiration 502.3 500.6 518.2 494.8 1465.3 Egestion 306.0 307.3 320.6 314.4 828.0 Production 113.9 111.8 126.0 105.5 275.0
a At peak adult density. b All life stages combined. c Includes calories recirculated by cannibalism. d No adult density-dependent effect on fecundity.

247
model could reasonably predict the timing and magnitude of the insect population
growth phase. However, the model indicated a higher consumption rate after day 66
than was observed in the Cf system (Fig. 4C). Consequently, in the model the insect
population became extinct soon after all the food was consumed and the model was
unable to maintain a low biomass of insects after the population declined, as was
observed in the drum experiments (Fig. 3B, 4A, 7, and 8). There were two genera-
tions during the insect population growth phase as indicated by the two peaks in the
predicted daily energetic variables (Fig. 4B and 7).
We examined the effect of varying mortality, cannibalism, food availability and
fecundity on the timing and size of peak numbers and biomass of the simulated Cryp-
tolestes population. Increasing the mortality of various life stages (singly or in com-
bination) reduced the predicted peak adult numbers and delayed population growth.
For example, increasing the egg mortality from 10 to 70~0 delayed peak adult numbers
by 13 d and the biomass peak by 14 d (Table 1, combinations A and B). By not apply-
ing cannibalism, adult numbers and the total biomass peak were higher and earlier
than those simulations with cannibalism applied (Table 1, combinations B, C and
( n
a
o - - )
30
20
10
o[ 0
C o n s u m p t i o n
1 R e s p i r a t i o n
/ E g e s t i o n
50 100 150 Days
Fig. 7. Daily consumption, respiration, egestion and production of C. ferrugineus infesting 100 kernels predicted by the simulatioa model. Initial energy available = 914.6 J; egg mortali- ty of 70% and density-dependent cannibalism applied to L2.
248
1000 Consumption
800 f EgesJion 6 0 0 . Assimilation I
o 400 n
:::,iiiiii . . . . procl 1
o . . . . . . . . . I . . . . . . . . . I . . . . . . . . . I . . . . . . . . . I . . . . . 0 50 100 150 2'00
Days
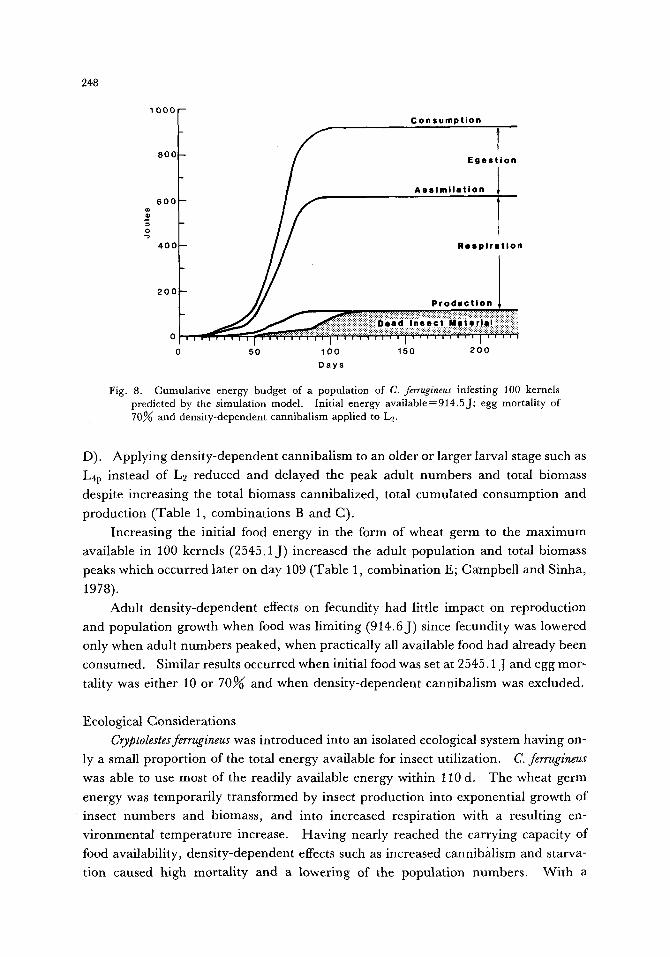
Fig. 8. Cumulative energy budget of a population of C. ferrugineus infesting 100 kernels predicted by the simulation model. Initial energy available=914.5J; egg mortality of 70% and density-dependent cannibalism applied to L2.
D). Applying density-dependent cannibalism to an older or larger larval stage such as
Lap instead of L2 reduced and delayed the peak adult numbers and total biomass
despite increasing the total biomass cannibalized, total cumulated consumption and
production (Table 1, combinations B and C).
Increasing the initial food energy in the form of wheat germ to the maximum
available in 100 kernels (2545. l j ) increased the adult population and total biomass
peaks which occurred later on day 109 (Table 1, combination E; Campbell and Sinha,
1978).
Adult density-dependent effects on fecundity had little impact on reproduction
and population growth when food was limiting (914.6J) since fecundity was lowered
only when adult numbers peaked, when practically all available food had already been
consumed. Similar results occurred when initial food was set at 2545,1 J and egg mor-
tality was either 10 or 70% and when density-dependent cannibalism was excluded.
Ecological Considerations
Cryptolestesferrugineus was introduced into an isolated ecological system having on-
ly a small proportion of the total energy available for insect utilization. C. ferrugineus was able to use most of the readily available energy within 110 d. The wheat germ
energy was temporarily transformed by insect production into exponential growth of
insec t numbers and biomass, and into increased respiration with a resulting en-
vironmental temperature increase. Having nearly reached the carrying capacity of
food availability, density-dependent effects such as increased cannibalism and starva-
tion caused high mortality and a lowering of the population numbers. With a

249
decrease of successful oviposition and low food availability, few larvae could complete
development, causing the beetle population to crash to a low level.
Quantitatively, the rate of deterioration of the whole grain bulk in the Cf system
study was only slightly faster than that found in the control system. C. ferrugineus removed about 2.6~ of the total available wheat energy, reduced the germination abili-
ty of the grain by a third, increased the temperature during the peak insect activity by
1.7~ and moisture content by 0.5~ above that of the control system. However, the
qualitative impact of C. ferrugineus infestation on the grain bulk was serious. The in-
sects removed 35~ of the total wheat germ energy, killed the seeds and made them un-
suitable for sowing purposes, and contaminated the ecosystem with feces and exu-
viae. By removing the high protein in wheat germ, C. ferrugineus renders the wheat
less marketable in a world where developed countries purchase Canadian grain for its
high level of protein.
Although the simulation model was limited in the number of variables used, it
was based on published available data and thereby provided a detailed picture of C. fer- rugineus population dynamics and use of wheat germ energy. The model also em-
phasized the lack of understanding of several of the empirical and behavioral
mechanisms involved in the interactions between some of the variables studied.
In this food-limiting environment, the classification of what constituted a crack
near the wheat germ did not indicate that all cracks would allow C. ferrugineus access to
the wheat germ since there were ca. 16~ of kernels with cracks in the bran that did not
have wheat germ eaten by the end of the study (Fig. 6). The behavioral mechanism
by which C. ferrugineus finds wheat germ is unknown. In the simulation model, we
assumed random searching and location of suitable feeding sites. Thompson's (1924)
model, originally developed for insect parasites, emulated reasonably well the feeding
relationships between G. ferrugineus and abundant levels of wheat germ, but the model
was not a good descriptor when wheat germs were comparatively rare and the sear-
ching ability of the adult insects was important. The model was obviously inadequate
for explaining the existence of a C. ferrugineus population for long periods with wheat
germ availability at low levels. Further quantitative information is required on the in-
teraction between the area of discovery and density of C. ferrugineus with wheat germ
(host) densities (Varley et al., 1973; Hassell, 1978).
Another complicating factor in this model was the lack of knowledge on the
energetic relationships between C. ferrugineus and those fungi that served as a food
source. Low population levels of C. ferrugineus would probably be able to obtain
energy indirectly from the dry wheat by feeding on fungi growing on the deteriorating
grain (Loschiavo and Sinha, 1966). An incipient low C. ferrugineus population could
maintain itself until further deterioration of the grain occurred through microfloral
activity, or invasion of another stored-product insect species able to chew through
wheat kernels, thereby making further food energy available to C. ferrugineus (White
and Sinha, 1980).

250
Density-dependent factors that influence the population process within a genera-
tion should be known so that insect population behaviour can be better understood
(Hassell, 1985). Smith (1966) showed that increased density reduced the growth, sur-
vival and fecundity of C. ferrugineus. We incorporated the effect of density on survival
or on cannabalism and fecundity using data from Smith (1966) because of their trac-
tability. However, the detailed effects of density on developmental time and energetic
conversion efficiencies of food into different individual larval stages were unknown and
could not be incorporated into the model. Increased density may have caused mutual
interference (Hassell, 1978) am~)ng searching larvae or adults, causing some in-
dividuals to stop feeding due to contact with one another and thereby, reducing energy
intake and growth. Smith (1966) showed that total developmental time from egg
hatch to adult emergence varied from 29 to 43 d; for the Cf system total developmental
times probably ranged from 29 d at low densities to about 35 d at densities of 18 lar-
vae/100 kernels (Fig. 3A). Increasing densities after the first generation ( ~ d a y 30)
may have affected the growth of individuals and consequently the timing of the second peak of Cf numbers and biomass.
The simulation trials (or experiments) indicated that cannibalism of larger com-
pared with smaller immature stages was more wasteful of developmental time and
energy, reducing the number of individuals reaching reproductive age in a C. fer-
rugineus population infesting a food-limited environment. Prepupae and pupae take
over 21 d to acquire sufficient energy to transform to adults; 25.45J are consumed,
most of which is respired or egested, to leave only 3.35J to 2.51 J in biomass, respec-
tively (Campbell and Sinha, 1978), that can be cannibalized. In contrast, L2 con-
sumes 3.98J to produce 0.33J of biomass in 11 d, but can also release the remaining
energy in the uneaten wheat germ (eg. 21.47J) when the L2 is cannibalized.
Prepupae and pupae probably provide a better defense from being cannibalized than
do smaller immatures, by plugging the entrance hole of the wheat germ cavity with
debris and silk (Rillett, 1949). However, if the entrance hole was breached the im-
mobile prepupae and pupae would be more vulnerable to cannibalism than the mobile smaller larvae.
The simulation experiments also indicated that although variation in fecundity
was linked to changes in adult density in the model, the adults did not reach a sufficient-
ly high density until most of the food was consumed; therefore, the density effect on
egg reduction came too late to have any major effect on the dynamics of C. ferrugineus population in a food-limited system. From this analysis we conclude that the publish-
ed information on density-dependent effect on fecundity (Smith, 1966) was not a major
regulatory mechanism in the C. ferrugineus population dynamics in this study.
However, reduced fecundity has the benefit of conserving energy for individual adults
during periods of stress such as starvation or high adult densities which would help to
prolong adult longevity and dispersal to more favorable areas.
The model gave a good simulation of C. ferrugineus energetics during the popula-

251
tion growth phase under the conditions of this study. Although the model was ex-
ploratory and limited to one temperature and moisture combination, the model em-
phasized how size-specific mortality can influence the rate of C. ferrugineus population
growth. The present model emphasized the need for further empirical information on
C. ferrugineus growth and bioenergetics of different development stages at various
temperature, moisture, density, starvation and food (including fungi) quality condi-
tions. The quantitative mechanisms by which C. ferrugineus can maintain itself at a
low population level is not fully understood and requires further study.
SUMMARY
The consequences of infestation of stored wheat by the rusty grain beetle, Cryp- tolestesferrugineus (Stephens) was determined for 222 d at 30~ in 70-1 drums containing
wheat at 13.5% moisture content. Temperature, grain moisture, seed damage, ger-
mination and weight, dust weight, fat acidity values (FAV), published data on growth,
reproduction, survival and cannibalism rates and energy budget were used to develop
a computer simulation model to simulate the population dynamics of C. ferrugineus at
30~ In the insect-free control system, the fungi, Alternaria alternata decreased,
Aspergillus glaucus group and PeniciUium spp. increased, probably causing a rise in FAV
of the grain. In the insect-infested system, C. ferrugineus could only eat the wheat
germ of kernels that had a broken bran layer; 35.7% of the wheat germ or 914.6 J per
100 kernels was consumed. Within two generations after initial introduction, C. fer- rugineus reached a peak in numbers and biomass polluting the ecosystem with excreta
and remains, and accelerating the deteriorative process observed in the insect-free con-
trol system by increasing respiration, temperature, FAV and reducing grain germina-
tion. After 87 d, the insect population declined to low levels. The simulation model
provided a close match between the observed and predicted numbers of insect life
stages and bioenergetic variables during the insect population growth phase. Simula-
tion trials suggested that cannibalism of larger compared with smaller immature stages
would be more wasteful of developmental time and energy, reducing the number of in-
dividuals reaching reproductive age, and that density-dependent fecundity was pro-
bably not an important regulatory mechanism of C. ferrugineus population dynamics in
this study.
ACKNOWLEDGEMENTS: We thank R.J. Bell, E. Campbell, G. G. Chee and A. M. Williamson for technical assistance; F. Cunningham for the illustrations; J. Vankirk and G. Fawkes for computer program- ming assistance; and J. M. Hardman, H. Kawamoto, N. D. G. White and C.J. Demianyk for reviewing this paper.
I~EFERENCES
Anonymous (1962) CerealLaboratory Methods. Amer. Assoc. Cereal Chem., St. Paul, Minn.

252
Brody, S. (1964) Bioenergetics and Growth. Hafner Publ. Co., Inc. New York.
Bronswijk, J. E. M. H. van- and R . N . Sinha (1971) Interrelations among physical, biological, and
chemical variates in stored-grain ecosystems; a descriptive and multivariate study. Ann. Entomol. Soc.
Amer. 69: 789-803.
Campbell, A. and R. N. Sinha (1976) Damage of wheat by feeding of some stored-product beetlesl J .
Econ. Entomol. 69: 11-13.
Campbell, A. and R. N. Sinha (1978) Bioenergetics of granivorous beetles, Cryptolestesferrugineus and
Rhyzopertha dominica (Coleoptera: Cucujidae and Bostrichidae). Can. J. Zool. 56: 624-633.
Campbell, A., N.B. Singh and R . N . Sinha (1976) Bioenergetics of the granary weevil, Sitophilus
granarius (L.) (Coleoptera: Curculionidae). Can. J. Zool. 54: 786-798.
Christensen, C. M. (1982) Storage of Cereal Grains and their Products. Amer. Assoc. Cereal Chem., St.
Paul, Minn.
Christensen, C. M. and H. H. Kaufmann (1969) Grain Storage: the Role of Fungi in Quality Loss. Univ. of
Minnesota Press, Minneapolis, Minn.
Cuff, W. R. and J. M. Hardman (1980) A development of the Leslie matrix formulation for restructur-
ing and extending an ecosystem model: the infestation of stored wheat by Sitophilis oryzae. Ecol. Modell-
ing 9: 281-305.
Hardman, J. M. (1978) A logistic model simulating environmental changes associated with the growth of
populations of rice weevils, Sitophilus oryzae, in small cells of wheat. J. appl. Ecol. 15: 65-87.
Hassell, M . P . (1978) The Dynamics of Arthropod Predator-Prey Systems. Princeton University Press,
Princeton.
Hassell, M. P. (1985) Insect natural enemies as regulating factors. J. Anim. Ecol. 34: 323-334.
Kawamoto, H., S. M. Woods, R. N. Sinha and W. E. Muir (1989) A simulation model of population
dynamics of the rusty grain beetle, Cryptolestesferrugineus in stored wheat. Ecol. Modelling 48: 137-157.
Keister, M. andJ . Burk (1974) Respiration: some exogenous and endogenous effects on rate of respira-
tion. 469-509. In M. Rockstein (ed) The Physiology of Insects, Vol. VI. Academic Press, New York.
Liscombe, E. A. R. and F. L. Watters (1962) Insect and mite infestations in empty granaries in the
prairie provinces. Can. Entomol. 94: 433-441.
Longstaff, B.C. and W. R. Cuff (1984) An ecosystem model of the infestation of stored wheat by
Sitophilus oryzae: a reappraisal. Ecol. Modelling 25: 97-119.
Loschiavo, S. R. (1975) Field tests of devices to detect insects in different kinds of grain storages. Can.
Entomol. 107: 385-389.
Loschiavo, S. R., R. N. Sinha (1966) Feeding, oviposition, and aggregation by the rusty grain beetle,
Cryptolestesferrugineus (C01eoptera: Cueujidae) on seed borne fungi. Ann. Entomol. Soc. Amer. 59: 578-
585.
Munro, H. A. U. (1969) Insect Pests in Cargo Ships. Can. Dept. Agric. Publ. 885.
Nie, N. H., C. H. Hull, J . G. Jenkins, K. Steinbrenner and D. H. Bent (1975) SPSS, 2nd ed. McGraw
Hill Book Co., New York.
Odum, E.P . (1989)
Massachusetts.
Rillett, R. O. (1949)
Sinha, R. N. (1964)
Man. 20: 19-33.
Sinha, R. N. (1965a)
Sinha, R. N. (1965b)
Ecology and our Endangered Life-support Systems. Sinauer Associates, Sunderland,
The biology of Laemophloeus ferrugineus (Steph.). Can. J. Res. D. 27" 112-148.
Mites of stored grain in western Canada-ecology and survey. Proc. Entomol. Soc.
Insects associated with stored products in Canada. Can. Insect Pest Rev. Suppl. 2.
Development of Cryptolestesferrugineus (Stephens) and Oryzaephilus mercator (Fauvel)

253
on seed-borne fungi. Entomol. Exp. Appl. 8: 309-323.
Sinha, R. N. (1973) Ecology of storage. Ann. Technol. Agric. 22: 351-369.
Sinha, R. N. and H. A. H. Wallace (1966) Ecology of insect-induced hot spots in stored grain in western
Canada. Res. Popul, Ecol. 3: 107-132.
Sinha, R. N. and F. L. Watters (1985) Insect pests of flour mills, grain elevators, and feed mills and their
control. Res. Branch Agr#. Can. Publ. 1776.
Smith, L. B. (1962) Observations on the oviposition rate of the rusty grain beetle, Cryptolestesferrugineus
(Steph.) (Coleoptera, Cucujidae). Ann. Entomol. Soc. Amer. 55: 77-82.
Smith, L. B. (1963) The effect of temperature and humidity on the oviposition of the rusty grain beetle,
Cryptolestes ferrugineus (Steph.). Proc. N. Central Br. Entomol. Soc. Amer. 18: 74-76.
Smith, L. B. (1965) The intrinsic rate of natural increase of Cryptolestesferrugineus (Stephens) (Coleoptera,
Cucujidae). J. Stored Prod. Res. 1: 35-49.
Smith, L. B. (1966) The effect of crowding on the oviposition, development and mortality of Cryptolestes
ferrugineus (Stephens)(Coleoptera: Cucujidae). J. Stored Prod. Res. 2: 91-104.
Smith, L. B. (1978) Ecology of stored grain in the Canadian Prairies. I. The distribution and size of a
low density population of Cryptolestesferrugineus (Coleoptera: Cucujidae). Can. Entomo[. 110: i281-
1292.
Smith, L, B. (1983) The relationship between wet grain, Cryptolestesferrugineus (Coleoptera: Cueujidae)
populations, and heating in wheat stored granaries. Can. Entomol. 115: 1383-1394.
Smith, L. B. and P. S. Barker (1987) Distribution of insects found in granary residues in the Canadian
Prairies. Can, Entomol. 119: 873-880.
Sorger-Domenigg, H., L. S. Cuendet and W. F. Geddes (1955) Grain storage studies. XX. Relation
between viability, fat acidity, germ damage, fluorescence value, and formazan value of commercial
wheat samples. Cereal Chem. 32: 499-506.
Surtees, G. (1965) Laboratory studies on the dispersion behaviour of adult beetles in grain. XII. The
effect of isolated pockets of damp and mouldy wheat on Cryptolestesferrugineus (Steph.) (Coleoptera,
Cucujidae). Bull. Entomol. Res. 55: 673-680.
Thompson, W. R. (1924) La th~orie math~matique de l'action des parasites entomophages et le facteur
du hasard. Ann. Fac. Sci. MarseiUe 2: 69-89.
Throne, J . E. (1987) A bibliography of the rusty grain beetle, Cryptolestes ferrugineus (Stephens) (Col-
eoptera: Cucujidae). U.S. Dept, Agr., Agr. Res. Serv. ARS-67.
Varley, G. C., G. R. Gradwell and M. P. Hassell (1973) Insect population ecology an analytical approach.
Blackwell Sci. Publ., London.
Wallace, H. A. H. and R. N. Sinha (1962) Fungi associated with hot spots in farm stored grain. Can. J.
Plant. Sci. 42: 130-141.
Watters, F. L. (1969) The locomotory activity of Cryptolestesferrugineus (Stephens) (Coleoptera: Cucu-
jidae) in wheat. Can. J. Zool. 47: 1177-1182.
�9 White, N. D, G. and.R.N. Sinha (1980) Changes in stored-wheat ecosystems infested with two combina-
tions of insect species. Can. J. Zool. 58: 1524-1534.

254
A. CAMPBELL ~ R. N. SINHA
d , ~ O ~ i ~ 5 - r ~ '~ y z , $ u ~ :~ .a -2~- Z 7 o ) J N - ~ , ~ ' T g N - 1 3 . 4 % o d , ~ o ~ o ~c 701
~ / ~ , ~ r NN, ~ : ~ � 9 �9 ~ N ~ �9 N ~ , c : ~ � 9 ~ N ~ i g f ~ (FAV)~r
�9 ~, N~, ~ * , # . t ~ t ' ~ * ~ v a e - ~ - N l ~ ' 5 1 ~ o ~ - ' - ~ ' k ~ ~''c, ~0~176
.N,::_.~[<:i~!Og"~]~,,~"~'~.~O Alternaria h;~7~" C, Aspergillus + Penicillium hit~J~ l_., ~ � 9 FAV
:~ . z , . > ' ~ / g ~ , , b / ' c ~ ' l ~ . d , ~ O ~ � 9 7g , 914. 6J/100 $ ~ 7 ~ N ~ 7 " c . ~ A ~ " C ~ 2 1~
~ L