an introduction to mathematical modeling - math user …othmer/papers/sem_intro.pdf · an...
TRANSCRIPT

An introduction to mathematical modeling
of signal transduction and gene control networks
Hans G. Othmer
Department of Mathematics
University of Minnesota
Minneapolis, MN

Overview
First Lecture: An introduction to mathematical modeling of signal transduction andgene control networks
• Examples of signal transduction, metabolic and gene control networks
• What is it we want to understand?
• The mathematical description of chemical reactions
• Analytical and computational techniques
Second lecture: Analysis of a model of signal transduction and motor control
• Signal transduction in bacteria
– The basic input-output behavior
– Response to steps and slow ramps
– The gain in the signal transduction pathway
– The bacterial motor
– Monte Carlo results

Third lecture: Analysis of a model for Drosophila melanogastersegment polarity genes
• Background on Drosophila development
• The basic facts about segment polarity genes
• Description of the Boolean representation
• Analysis of wild-type and heat-shock behavior
• Expression patterns in mutants
• A two-step model

Biochemical networks
• Signal transduction networks: The pathways and the molecular components, such as
kinases, G-proteins, sencond messengers, ..., involved in transducing a signal from one
location to another. Frequently used in the context of transduction of extra- into
intracellular signals.
• Metabolic networks: The pathways and the molecular components (metabolites,
enzymes, control factors) involved in the biosynthesis of new components, the conversion
of molecular ‘foodstuffs’ into energy, etc. One of the most important examples is the
glycolytic pathway, which converts sugars into energy-storing molecules such as ATP.
• Gene expression networks:The pathways and components, such as genes, polymerases,
transcription factors, etc., that are involved in gene expression, mRNA translation, etc.

An early view of signal transduction
To understand, next, how external objects that strike
the sense organs can incite [the machine] to move its
members in a thousand different ways: think that
(a) the filaments (I have already often told you that
these come from the innermost part of the brain and
compose the marrow of the nerves) are so arranged
that they can very easily be moved by the objects of
that sense and that
(b) when they are moved, with however little force,
they simultaneously pull the parts of the brain from
which they come, and by this means open the en-
trances to certain pores in the internal surface of the
brain ..
Thus if fire A is near foot B, the particles of this fire
have force enough to displace the area of skin they
touch; and thus pulling the little thread (cc) which
you see attached there, they simultaneously open the
entrance to the pore (de) where this thread termi-
nates; just as, pulling on one end of a cord, one si-
multaneously rings a bell which hangs on the oppo-
site end.
R. Descartes -De Homine

A global view of signal transduction
Lipid-soluble molecules can
pass through the cell mem-
brane, but most signals are
proteins or peptides and these
require more machinery ...
Y
Nucleus
Receptor
Cell
factorSoluble
Ligand

Receptors can transduce signals in a variety of ways

Receptor phosphorylation and G-protein activation: two majortransduction pathways

The G-protein motif

Some of the functions of G proteins

One mode involving phosphorylation: the RTK pathway

Downstream activation via RTKs

A Ras-mediated pathway

The signal transduction pathway in E. coli
+CH3R
ATP ADPP~
flagellarmotor
Z
Y
PY
~
PiB
B~P
Pi
CW-CH3
ATP
WA
MCPs
WA
+ATT
-ATT
MCPs

Metabolic networks
• Metabolism : The cellular process by which organic molecules are synthesized or
degraded, usually via enzyme-catalyzed reactions
• The interconnected components and reactions form a network, called themetabolicnetwork
E B
C
D
AE
P2
P
P
: A B
D
C
3
A
: A
P
P
P
C
4
5
6
E1 :
:
: C
D
: B1
2
3
E2
E3
E4
E5 E1 E6
E6E5
E4
Rather than viewing reactions in isolation, as on the left, we should begin thinking about the
underlying structure of the network, as well as the individual reactions, as on the right!

The glycolytic reactions
Glycolysis: Thelysis or split-
ting of glucose
Stage 2
Stage 1
molecules
Glycolysis
Stage 3Production of NADH
and ATP
Breakdown of complex

Gene control networks, development, and the French flag problem

The map from egg to adult in Drosophila melanogaster

The general structure of the gene network in Drosophila

The details of the segment polarity gene network

The general structure of signal transduction cascades
1. General scheme
Outside
Inside
Signal Transduction
Internal Response
Signal Propagation
External Signal External Signal
Signal Detection
55555555555555555555555555555555555555555555555555555555
55555555555555555555555555555555555555555555555555555555

Amplification of signals

Adaptation to constant signals
A single step change in the signal produces a single response and a return to the basal level of
activity, while a sequence of steps produces a sequence of responses.
Thus the system both adapts AND maintains sensitivity to further changes in the stimulus.

The role of adaptation in the visual system

What is it we want to understand about these networks?
• How do we describe their dynamical behavior? Do we use
1. a continuous state space and deterministic descriptions via ODE’s,
2. a Boolean representation of ON/OFF states and logical functions that determine the
dynamics, or
3. a stochastic description, in which we follow individual molecules.
How do we decide which is the appropriate choice?
• What are the ‘attractors’ in the dynamics? Are they steady states or fixed points, periodic
attractors, or are they more complicated?
• How do we define and then compute measures of amplification, sensitivity and gain?
• How sensitive is the behavior to variations in the parameters? In the structure of the
network itself ? From a dynamical systems point of view this might be viewed as a
question of ‘structural stability’; another way of phrasing this is to ask whether certain
features of the dynamics are robust.

What are the big questions about these networks?
• Why have the networks evolved to their present form? To what extent do the physics and
chemistry influence the structure of the networks? Said otherwise, if one wants to
synthesize Z from A, how many distinct (nontrivial) paths are there? The answer bears
on the inverse problem of inferring the network from expression patterns. Are there
design principles and evolutionary approaches that can help us understand the observed
structure?
• Many signal transduction systems adapt to constant signals but retain the ability to
respond to further changes. What structure in a dynamical system guarantees this
characteristic?
• Can we give a useful precise definition of robustness? How do we determine when
‘parametric robustness’ suffices and when ‘redundancy’ is necessary? Should we expect
robustness at the individual level, or can population-level feedbacks correct
individual-level variances in response to signals?

Two views on complex networks .......
”When the number of factors coming into play in a phenomenological complex is too large,
scientific method in most cases fails. One need only think of the weather, in which case the
prediction even for a few days ahead is impossible. Nevertheless, none doubts that we are
confronted with a causal connection whose causal components are in the main known to us.
Occurrences in this domain are beyond the reach of exact prediction because of the variety of
factors in operation, not because of any lack of order in nature.”
Albert Einstein
”Though signalling pathways are often drawn as simple linear chains of events, they are
rarely that simple. Frequently there is feedback, cross-talk between pathways and enough
branching to make one want to change to a less complicated field of biology.
In order to fully understand these pathways, we need a convenient and powerful model to
complement the experimental research. Though there have been relatively few attempts to
model signalling pathways using computers, it seems likely that this will very soon become a
major area of study.”
J. Michael Bishop, Science 267:1617

The mathematical description of chemical reaction networks
• The stochiometryof a chemical reaction denotes the molar ratios in which molecules
are converted into products in that reaction.
A + 2 B C
1 2 1
BA
11
k k
Rate = kA Rate = kAB2MAK:
Here, and unless specified otherwise later, we consider only mass action kinetics
(MAK ), which means that the rates are monomials in the concentrations, raised to the
power given by the stoichiometry, of the components that react together.
• Thestochiometric matrixν = (νij) provides a mathematical encoding of the
stoichiometry and topology of a reaction network
21P
A
P B
D
C
reactions
1 0 00 1 0 00 0 1 10 0 1 0
−1 −1−1
−1
00
00
P4
P5
P6
P3 −1 = ν reactants

The mathematical description of chemical reaction networks ...
Define the concentration and rate vectors
c =
c1
c2...
cn
P (c) =
P1(c)
P2(c)...
Pn(c)
Then the concentration evolves in time according the differential equation
dc
dt= νP (c)
(1)c(0) = c0
c
c
c
2
1
3

Reaction invariants
Suppose that there are vectorsξk ∈ Rn, k = 1, . . . ,m such that
νT ξk = 0 for k = 1, . . . ,m.
What does this mean for the dynamics? Clearly
d
dt< ξk, c >=< ξk, νP (c) >=< νT ξk, P (c) >= 0,
i.e., the quantity< ξk, c > is constant in time. We call this a kinematic invariant, and itaffects the evolution of the composition as follows.< ξk, c >= constant defines a linearmanifold, and the intersection of this manifold with the positive ‘orthant’ of compositionspace defines the reaction simplex.
c
c
c3
2
1
Reaction simplex
c(0)
Remark: Note thatνT has a nontrivial
null space if the number of species ex-
ceeds the number of reactions.

Enzyme-catalyzed reactions
LetE denote an enzyme, letS denote the substrate, letES denote the complex formed
whenS binds toE, and letP denote the product
E + S ES E + Pk k1 2
k−1
Herec = (E,S,ES, P )T and
ν =
−1 1 1
−1 1 0
1 −1 −1
0 0 1
The rank ofν is 2 and therefore dimN (νT ) = 2, so there are two kinematic invariants. We
can choose these asξ1 = (1, 0, 1, 0)T andξ2 = (0, 1, 1, 1)T . Thus the reaction simplex is
two-dimensional. How could we have predicted thisa priori?
Exercise: Determine how the reaction invariants reflect the conservation of atomic species in
the reaction
H2 +O−→←− H2O.

A Cartoon Model of Excitation and Adaptation
Excitation
Change
Adaptation
ResponseSignal
dy1
dτ=
S(τ)− (y1 + y2)τE
dy2
dτ=
S(τ)− y2
τA
For example,S(t) could be proportional to the fraction of receptors occupied.
y1 =S0τAτA + τE
(e−τ/τA − e−τ/τE )
y2 = S0(1− e−τ/τA)
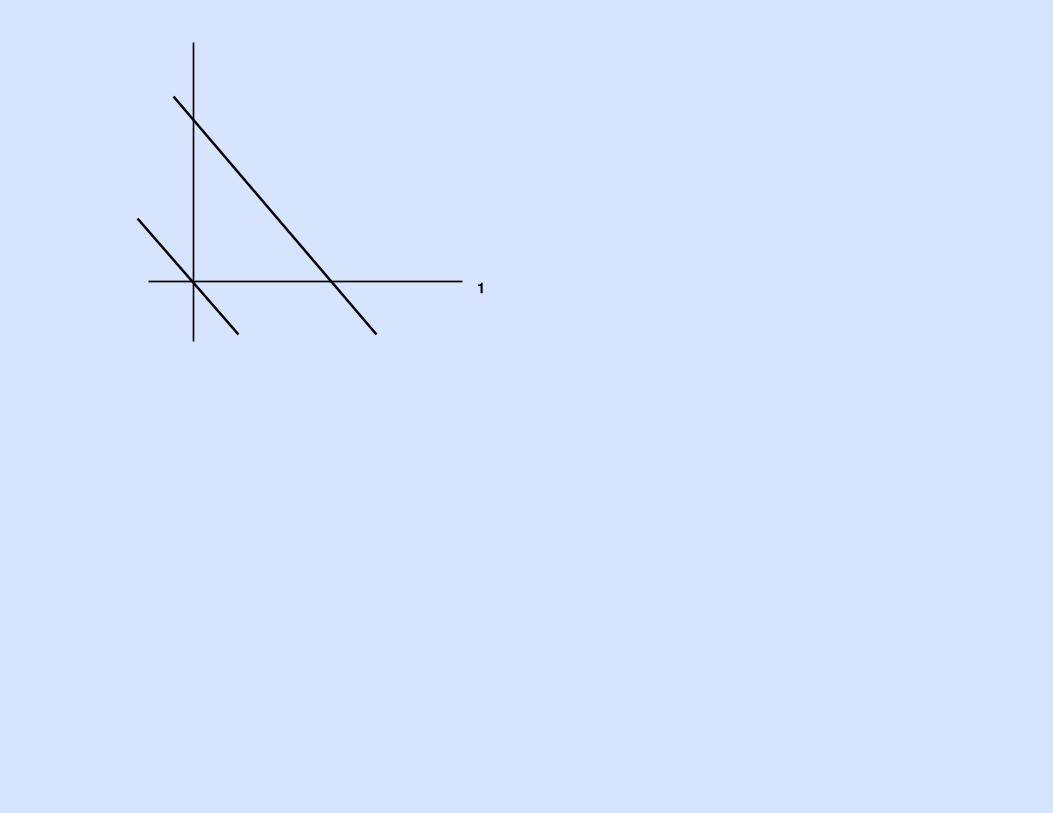
1y
y2(a)
1S(τ) = 0 S(τ) = S
1y
y2(b)
S(τ) = 0 S(τ) = S1 S(τ) = S2
If τE << τA, then forτ >> τE, y1 relaxes to
y1 ∼ S0e−τ/τA ≡ S0 − y2(τ) = τAy2.
u ≡ y2 satisfies
du
dt+
1τAu =
1τA
dS
dt

Adaptation, Sensitivity and Gain
dx
dt= f(x, S(t))
Response
R = G(x(t)).
At steady state supposex = X(S); then adaptation requires that
dRdS
=∑ ∂G
∂xi
∂xi∂S
= 〈∇xG(X(S)), (Dxf)−1(X(S), S)DSf(X(S), S)〉 = 0
Definitions of gain and sensitivity
g0 ≡dRdS
=∑ ∂G
∂xi
∂xi∂S
gLS ≡dRd lnS
= Sg0
gLR ≡d lnRdS
=1Rg0 gLL ≡
d lnRd lnS
=S
Rg0
Thus the steady-state gain vanishes in a system that adapts perfectly!

Sensitivity of transient solutions
∂x
∂S= Φ(t, 0)
∂x
∂S(0) +
∫ t
0
Φ(t, τ)DSf(x(τ), S(τ))dτ
g = max[0,∞)
g(t)
Reported bacterial gains range from g =∼ 6 to g =∼ 55

Example
Cartoon model
dy1
dt=
θ(t)− (y1 + y2)τE
dy2
dt=
θ(t)− y2
τA
Step toθ1 at t = 0
y1 =θ1τA
τA + τE(e−t/τA − e−t/τe)
∂y1
∂θ1=
τAτA + τE
(e−t/τA − e−t/τE )
Thus
g0 =y1
θ1gLS = θ1g0 = y1
gLL =θ1
y1g0 ≡ 1 gLR =
g0
Rg0 ≡
1θ1
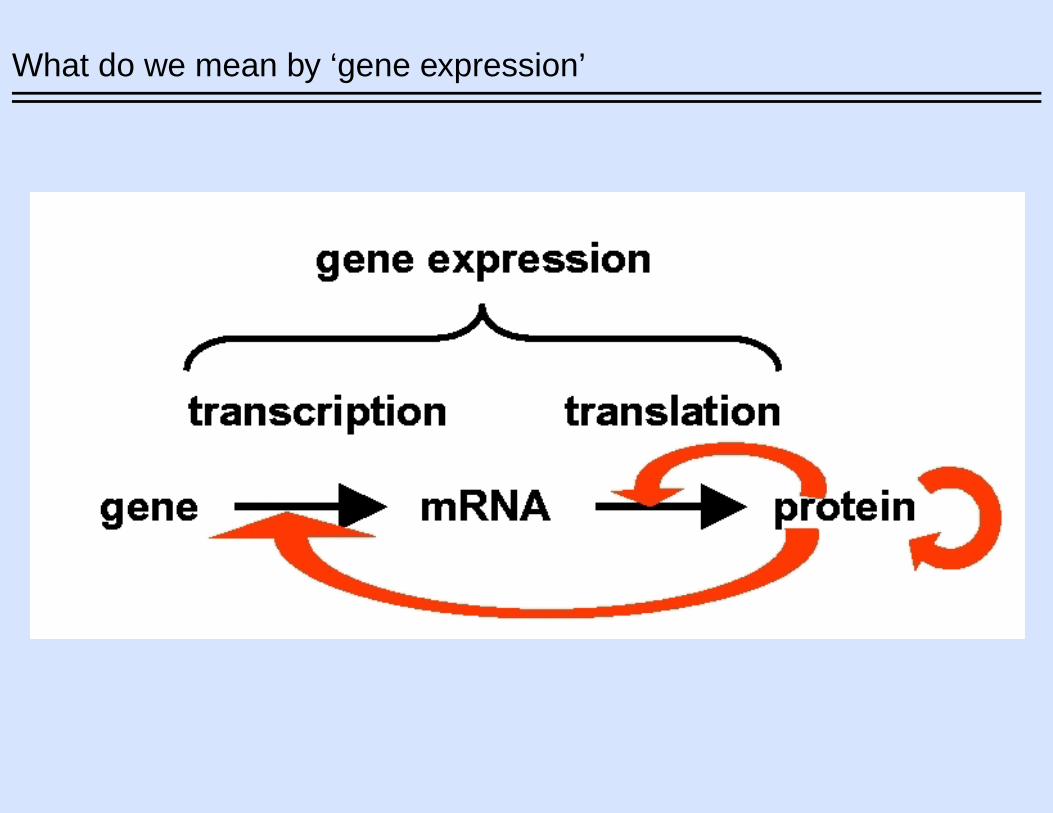
What do we mean by ‘gene expression’
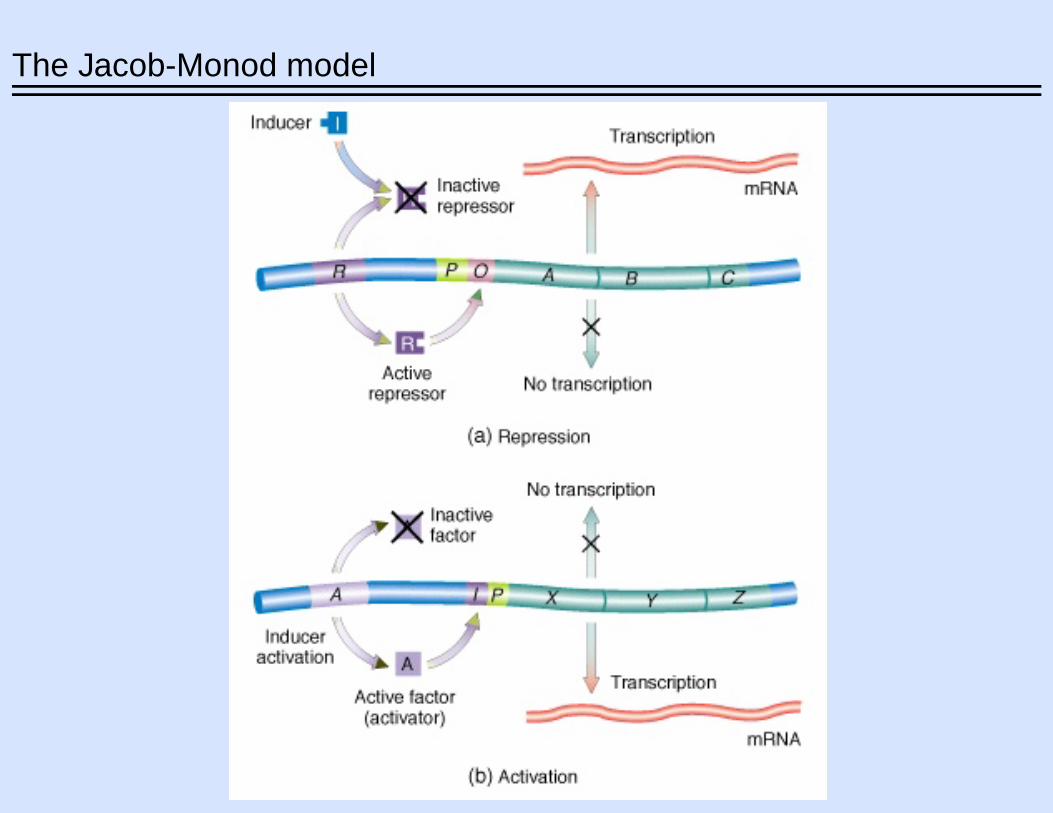
The Jacob-Monod model

The four basic modes of transcription control
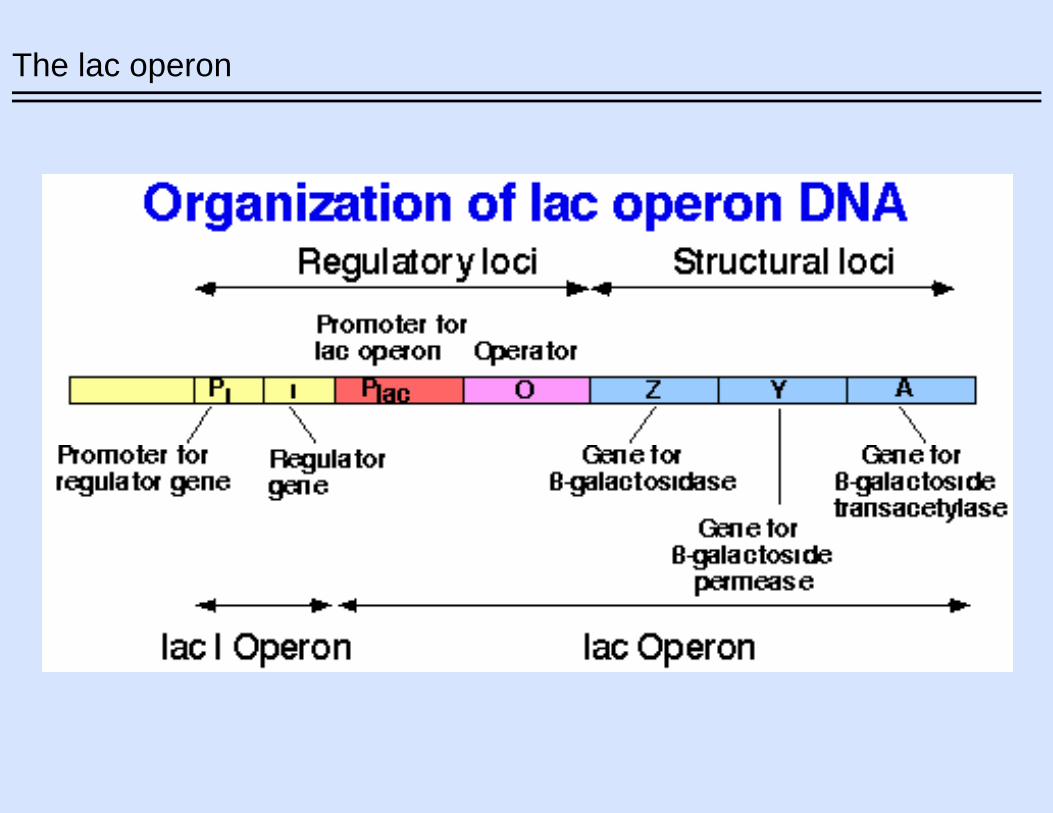
The lac operon

The effect of lactose

Eucaryotic transcription control

The loci of control and modification

Four types of models
• Boolean models
.... in which everything is either ON or OFF and the state space is a finite set
• Mixed models
.... in which some things are either ON or OFF, while others vary continuously
• Deterministic continuous models
.... in which everything varies continuously, the state space is a subset ofRn,
and the dynamics are deterministic
• Stochastic models
.... in which we recognize that ‘reactions ’ occur one molecule at a time

The Boolean approach

One realization of the ‘not if’ or ‘C and (not D)’

A general description of a gene control pathway based on feedback

The governing equations for a ‘continuous’ description
dS1
dt= R(Sn+1)− AN
VNT1(S1, S2)− k1S1
dS2
dt=ANVC
T1(S1, S2)− k2S2
dS3
dt= k2S2 − k3S3
dS4
dt= k3S3 − k4S4
...dSjdt
= kj−1Sj−1 − kjSj , j = 5, . . . , n− 1...dSndt
= kn−1Sn−1 − knSn −ANVC
Tn(Sn, Sn+1)
dSn+1
dt=ANVN
Tn(Sn, Sn+1)− kn+1Sn+1
wherekj = kj + kj for 4 ≤ j ≤ n− 1.

The control functions for inducible and repressible systems
R+ pS RSp, K1 = RSp/R · Sp
R+O OR, K2 = OR/R ·O
R = repressor,O = operator, andS = effector.
Rt = R+RSp = R(1 +K1Sp)
Ot = O +OR = O(1 +K2R)
Fraction of operator regions free of repressor:
f(S) =O
Ot=
1 +KtSp
K +K1Sp(2)
K = 1 +K2Rt > 1

For a repressible system
R+ pSK1� RSp
RSp +OK2� ORSp
In this case
f(S) =1 +K1S
p
1 +K1KSp(3)

Quantitative characterization of some gene control systems
Enzyme Effector p K1 K2Rt
Inducible
β-Galactosidase Isopropylthio- 1.91 2.5× 1010M−2 2.5× 103
galactoside
Histidine-NH3-lyase Imadizole 2.04 1.7× 1010M−2 26
propionate
Urocanase Histidine 2.3 4.3× 1012M−2 102
Mannitiol Ribitol 3.13 — —
dehydrogenase
Repressible
IMP dehydrogenase Guanine 0.91 —
XMP aminase Guanine 0.68 —
Alkaline PO3−4 0.93 2× 103M−1 5× 103
aFrom Yagil and Yagil (1971).