alternatives to animal testing: research, trends ......animal use in toxicological experiments,...
TRANSCRIPT

ALTEX 20, Suppl.1/03 3
S u m m a r yC u r rent trends and issues in the development of alternatives tothe use of animals in biomedical experimentation are discussedin this position paper. Eight topics are considered and include re finement of acute toxicity assays; eye corrosion/irritation alter-natives; skin corrosion/irritation alternatives; contact sensitiza-tion alternatives; dev e l o p m e n t a l / re p roductive testing alterna-tives; genetic engineering (tra n s genic) assays; toxicoge n o m i c s ;and validation of alternative methods. The discussion of re fin e-ment of acute toxicity assays is focused primarily on dev e l o p-ments with reg a rd to reduction of the number of animals used inthe LD5 0 a s s a y. Howev e r, the substitution of humane endpointss u ch as clinical signs of toxicity for lethality in these assays is also evaluated. Alternative assays for eye corrosion/irritation aswell as those for skin corrosion/irritation are described with particular attention paid to the outcomes, both successful and un-
successful, of sev e ral validation efforts. Alternative assays forcontact sensitization and dev e l o p m e n t a l / re p roductive toxicity arep resented as examples of methods designed for the ex a m i n a t i o nof interactions between toxins and somewhat more complex p h y s i o l ogical systems. More ov e r, genetic engineering and toxi-c ogenomics are discussed with an eye toward the future of bio-l ogical experimentation in ge n e ral. The implications of gene ma-nipulation for re s e a rch animals, specific a l l y, are also ex a m i n e d .Fi n a l l y, validation methods are investigated as to their eff e c t i v e-ness, or lack there o f, and suggestions for their standard i z a t i o nand improvement, as well as implementation are rev i ewed.
1 Refinement of acute toxicity assayT h ree assays have been validated and adopted as re p l a c e m e n t sfor the conventional LD5 0 test. The assays differ primarily as to
Alternatives to Animal Testing: Research,Trends, Validation, Regulatory AcceptanceJane HugginsToxicology Consulting Services, USA-Plainsboro
Content1 Refinement of acute toxicity assay1.1 Background1.2 Refinement assays1.3 Comparison of refinement assays1.4 Humane endpoints1.5 Regulatory activities1.6 Summary, conclusions, and future work2 Alternatives to eye corrosion/irritation testing
in animals2.1 Background2.2 In vitro alternatives2.2.1 Combinatorial approaches2.2.2 Reference standards2.2.3 Mechanistic considerations2.3 Summary, conclusions, and future work3 Alternatives to skin corrosion/irritation testing
in animals3.1 Background3.2 In vitro models3.3 Validation and regulatory activities3.4 Summary, conclusions, and future work4 Alternatives to skin sensitization testing in animals4.1 Background4.2 Alternative assays4.3 Validation and regulatory activities4.4 Summary, conclusions, and future work5 Alternatives to developmental/reproductive toxicity
testing in animals5.1 Background
5.2 In vitro alternatives5.2.1 Micromass (MM) assay5.2.2 Whole embryo culture (WEC) assay5.2.3 Embryonic stem cell (EST) test5.2.4 Frog embryo teratogenesis assay (Xenopus) (FETAX)5.3 Validation and regulatory activities5.4 Summary, conclusions, and future work6 Genetic engineering methodologies6.1 Background6.2 Genetically, engineered cell lines6.3 Transgenic animals6.4 Summary, conclusions, and future work7 Gene chip technology as an alternative to animal testing7.1 Background7.2 What is a gene chip?7.3 Application of gene chips to basic research7.4 Application of gene chips to toxicological research7.5 Development of gene chips as alternatives to
animal testing7.6 Current status of gene chip technology7.7 Summary, conclusions, and future work8 Validation of alternative methodology8.1 Background8.2 What is validation and why has it failed?8.3 Improving the validation efforts8.4 Harmonization of validation process8.5 Why validate?8.6 Mechanistic understanding: what are we validating?8.7 Summary, conclusions, and future work

HU G G I N S
ALTEX 20, Suppl.1/034
the endpoint they measure; howev e r, all assays use fewer animalsthan the conventional LD5 0 test. The use of more humane end-points, such as clinical signs of toxicity, rather than lethality, isperhaps the most advanced suggestion to date reg a rding toxico-l ogical evaluation of acute ex p o s u re. Much remains to be done,h o w ev e r, with reg a rd to standardization of this appro a ch.
2 Alternatives to eye corrosion/irritation testing in animals Although much re s e a rch has been done to date on the dev e l o p-ment of viable in vitro assays of ocular corrosion and irritancy,validation of these assays has been problematic. Sev e ral re a s o n shave been postulated for the failure of validation efforts, the mostp rominent of which are the following: The in vivo test used forcomparison, the Draize test, is based on subjective scoring of tissue lesions in the ey e, providing variable estimates of eye irritancy; the non-animal method protocols were inadequate; thechoice of test substances was not well-planned; and the statisti-cal appro a ches used were not appro p r i a t e. Perhaps the mostp romising suggestion as to how to remedy these difficulties cur-rently is to use complementary alternative assays in batteries toevaluate eye corro s i o n / i r r i t a t i o n .
3 Alternatives to skin corrosion/irritation testing in animalsThe use of alternative assays has replaced, to a large extent, thetesting of corrosive or irritating substances on the skin of live animals. Examples of the assays discussed include the Corro -s i t ex® assay that uses no animal cells at all, the tra n s c u t a n e o u selectrical resistance (TER) assay that uses a small section of ra tskin, and sev e ral in vitro skin irritancy models that incorpora t ehuman skin in small quantities. From a scientific pers p e c t i v e, thereplacement of the Draize test with these assays lends greater objectivity as well as more ge n e ral re l evance to human skin cor-rosion and irritation. Validation efforts utilizing these modelshave proven satisfactory in most instances.
4 Alternatives to skin sensitization testing in animals P rog ress toward the development and re finement of alternativeassays of contact sensitization is strongly dependent upon bre a k-t h roughs in our understanding of the immune processes medi-ating the re s p o n s e. Extensive efforts directed toward validation of the local lymph node assay have borne the much-needed fruitof a “stand alone” assay that incorporates elements of both re finement and reduction. Howev e r, much more basic re s e a rchremains to be done before a fully validated replacement assay ofcontact sensitization finds regulatory support. Promising areas of re s e a rch include those in which cytokine pro files associatedwith contact sensitization are analyzed.
5 Alternatives to developmental/reproductive toxicity testing inanimalsValidation efforts are prog ressing well for in vitro assays of d ev e l o p m e n t a l / re p roductive toxicity. Results from evaluations of the MM and WEC assays as well as the EST appear to be f a v o rable; data from studies of FETAX suggest that further i m p rovements in the assay would yield greater pre d i c t i v i t y.H e n c e, our reticence to use an alternative assay to measure toxic effects on complex physiological processes such as re p ro-duction may have to yield to the results obtained from these recent evaluations.
6 Genetic engineering methodologiesThe generation and use of transgenic animals to study questionsof biomedical interest have been questioned by many in view ofthe moral and ethical dilemmas presented by these activities.That transgenic animals may contribute to the reduction of animal use in toxicological experiments, particularly studies of carcinogenicity, is not disputed. However, advocates of re-placement alternatives argue that in vitro alternatives to thistype of toxicity testing have not been given adequate attention.
7 Gene chip technology as an alternative to animal testingGene chips (DNA microarrays) represent a technology that hasalready opened many doors in basic genomic research. More-over, their value to both investigative and discovery toxicologyis becoming much more apparent as more toxicology ex-periments are conducted using them. Use of microarrays as reduction or replacement alternatives to animal testing alsoholds great pro m i s e, particularly when they are used as components of prescreening batteries, and when coupled to cell culture techniques.
8 Validation of alternative methodology Validation of alternative methods has just emerged from arather chaotic phase in which the principles behind appropriateconduct of a validation study were defined, mainly through trial and error. Much refinement has come out of this “explor-atory“ phase, including recognition that validation studiesshould be built upon a solid platform, consisting of componentssuch as good reference standards, reliable protocol transfer between laboratories, and appropriate application of biostatis-tical techniques. Efforts are now underway to apply these les-sons learned to future validation studies and to harmonize validation techniques among countries in order to maximize thepossibility that the data generated can be used worldwide.
Z u s a m m e n fassungen: A l t e r n a t iven zu Ti e r versuchen: Fo r s c h u n g ,Trends, Validierung, Akzeptanz auf behördlicher EbeneIn diesem Positionspapier werden aktuelle Trends und Ergeb-nisse in der Entwicklung von Alternativmethoden zu Tierver-suchen in der biomedizinischen Forschung erörtert. Es werdenacht Themenfelder angesprochen: Refinement akuter Toxizitäts-
tests; Alternativen zu Augenkorrosion-/reizung; Alternativen zu Hautko r ro s i o n - / reizung; Alternativen zu Ko n t a k t e m p -findlichkeit gegenüber Stoffen; Alternativen zu Entwicklungs/-R e p roduktionstests; Gentech n o l ogie (tra n s gen) Tests; To x i -c ogenomics und Validierung alternativer Methoden. Die Diskussion des Refinements akuter Toxizitätsuntersuchungen

HU G G I N S
ALTEX 20, Suppl.1/03 5
konzentriert sich in erster Linie auf Entwicklungen im Hinblickauf eine Reduzierung der Tierzahlen beim LD5 0 Test. In diesen Untersuchungen wird ausserdem die Verwendung vonm e n s ch l i chen Endpunkten wie klinische Toxizitätsmerkmale für Letalität untersucht. Bei den Alternativmethoden im Be-re i ch Au ge n ko r ro s i o n e n / - re i z u n gen sowie Hautko r ro s i o n e n /-reizungen wird das Hauptaugenmerk auf die Ergebnisse ver-s chiedener Va l i d i e r u n g s v e rs u che ge r i chtet sein. Alternative Untersuchungen im Bereich Kontaktempfindlichkeit und Ent-wicklungs-/Reproduktionstoxikologie werden als Beispiele fürMethoden dargestellt, welche zur Überprüfung von Interakti-onen zwischen Toxinen und physiolog i s ch ko m p l ex e ren Systemenkonzipiert wurden. Ausserdem werden die zukunftsträchtigenBereiche Gentechnologie und Toxicogenomics besprochen. DieAu s w i r k u n gen von Genmanipulationen insbesondere auf Versuchstiere sollen untersucht werden. Abschliessend sollenValidierungsmethoden auf ihre Effektivität hin unters u cht w e rden sowie Empfehlungen für deren Standard i s i e r u n g,Verbesserung sowie Umsetzung überprüft werden.
1 Refinement von akuten ToxizitätstestsBisher wurden drei Methoden für den Ersatz des LD50 Testsakzeptiert. Diese unterscheiden sich in erster Linie im von ihnen gemessenen Endpunkt; trotz dieses Unterschieds könnendurch diese Tests im Vergleich zum herkömmlichen LD50 TestTiere eingespart werden. Heutzutage muss der vermehrte Ein-satz von humanen Endpunkten, wie klinische Anzeichen vonToxizität, im Gegensatz zum Endpunkt Letalität als der fort-ge s chrittenste Ansatz hinsich t l i ch der toxiko l og i s ch e nBeurteilung akuter Exposition angesehen werden. Gro s s eAnstrengungen müssen jedoch in Hinblick auf die Standardi-sierung dieses Ansatzes unternommen werden.
2 Alternativen zu Tierversuchen im Bereich Augenkorrosion/-reizungObwohl bei der Entwicklung von in vitro Methoden im Bereichder Au ge n ko r ro s i o n / - reizung grosse A n s t re n g u n gen unter-nommen wurden, hat sich die Validierung der entsprechendenMethoden als pro b l e m a t i s ch hera u s gestellt. Ve rs ch i e d e n eGründe haben zu diesem Misserfolg beigetragen. Die be-kanntesten Gründe sind die folgenden: Der für den Vergleichmit dem in vitro Test herangezogene in vivo Test, der DraizeTest, beruht auf einer subjektiven Einteilung der auftretendenGewebsverletzungen am Auge, was zu unterschiedlichen Ein-schätzungen der Augenreizung führt; das Protokoll der in vitroMethode war mangelhaft; die Auswahl der Testsubstanzen wars ch l e cht geplant; die statistischen Analysemethoden ware nungeeignet. Als Lösung dieser Probleme bietet sich an, mittelsin Batterie geschalteter verschiedener komplementärer Alter-nativmethoden Au ge n ko r ro s i o ns/ - reizungs Unters u ch u n ge ndurchzuführen.
3 Alternativen zu Tierversuchen im Bereich Hautkorrosion/-reizungDer Einsatz von Alternativmethoden hat in hohem Masse Testsauf korrosive oder reizende Substanzen der Haut an lebendenTieren ersetzt. Im vorliegenden Dokument werden die folgenden
Tests beschrieben: Corrositex®, bei welchem überhaupt keineTierzellen eingesetzt werden; der transcutaneous electrical resistance (TER) Test, welcher einen kleinen Teil an Rattenhautbenötigt sowie vers chiedene in vitro Hautre i z u n g s m o d e l l e,w e l che kleine Mengen mensch l i cher Haut erfordern. Aus wissenschaftlicher Sicht verhilft der Ersatz des Draize Testsdurch die erwähnten in vitro Methoden im Bereich Haut-korrosion/-reizung zu mehr Objektivität sowie allgemeinererR e l evanz. Die mit diesen Modellen unternommenen Validierungsanstrengungen haben sich in vielen Fällen alszufriedenstellend erwiesen.
4 Alternativen zu Tierversuchen im Bereich Kontaktempfind-lichkeit Fortschritte in der Entwicklung und dem Refinement von Alter-nativmethoden im Bereich Kontaktempfindlichkeit hängen starkvon den Erfolgen bezüglich unseres Verständnisses der Immun-prozesse ab, welche die Reaktion vermitteln. Aus den grossenAnstrengungen, welche hinsichtlich der Validierung des locallymph node assay unternommen wurden, ist nun ein „standalone” Test hervorgegangen, welcher sowohl zum Refinementwie zur Reduzierung von Tierversuchen beiträgt. Nichts destot rotz muss noch vermehrt Grundlage n f o rs chung betrieben werden, bevor ein vollständig validierter Ersatztest für Kontakt-empfindlichkeit Akzeptanz auf behördlicher Ebene erlangenw i rd. Erfolgvers p re chende Fo rs ch u n g s b e re i che sch l i e s s e nAnalysen von Zytokinpro filen, welche mit Ko n t a k t e m p fin d-lichkeit assoziiert sind, ein.
5 Alternativen zu Tierversuchen im Bereich Entwicklungs-/Re-produktionstestsDie Validierungsanstrengungen für in vitro Tests im BereichE n t w i ck l u n g s - / R e p roduktionstoxizität kommen gut voran. Resultate der Evaluierung von MM und WEC Test sowie demEST Test scheinen Erfolg versprechend zu sein; Daten von FETAX-Studien zeigen, dass eine weitere Verbesserung desTests bessere Voraussagen ergeben würde. Daher sollte unsereZ u r ü ck h a l t u n g, Alternativmethoden zur Messung toxischer Effekte auf komplexe physiologische Prozesse wie Reproduktioneinzusetzen, zu den Resultaten führen, welche sich aus denfrüheren Evaluierungen ergeben haben.
6 GenmanipulationDie Erzeugung für und der Einsatz von transgenen Tieren in derbiomedizinischen Forschung hat bereits viel zu reden gegebengerade hinsichtlich moralischer und ethischer Dilemmas, diesich aus solchen Aktivitäten ergeben. Dass transgene Tierezu einer Reduzierung der Tierzahl in toxikologischen Experi-menten, im Besonderen in der Kre b s f o rs chung beitragen können wird nicht bestritten. Dennoch sind die Verfechter vonErsatzmethoden der Meinung, dass den in vitro Methoden indiesem Bereich der Toxikologieprüfung zu wenig Beachtunggeschenkt wird.
7 Gen chip Technologie als Alternative zu TierversuchenGen chips (DNA Microarrays) repräsentieren eine Technologie,welche in der Genomforschung bereits viele Türen geöffnet hat.

HU G G I N S
ALTEX 20, Suppl.1/036
1.1 BackgroundH i s t o r i c a l l y, lethality following acute exposure to a chemical has been a cor-nerstone upon which much toxicologicaldecision-making has rested. The LD50
(dose at which lethality is observed in50% of the animals tested) is often con-sidered the primary index of potentialtoxicity of a chemical and is widely usedas a tool for determining the dosages tobe used for further ex p e r i m e n t a t i o n .LD50 values are derived using multiplespecies and routes of exposure. The mostcommon species utilized is the rat, how-ever, the mouse, guinea pig, rabbit, anddog are also tested. The most commonroutes of exposure are oral (by gavage),dermal, inhalation, and intraperitonealand intravenous injection. Ideally, malesand females of equal number per dosel evel are employed and several dose levels are evaluated. The LD50 value isobtained through probit analysis of thedata obtained.
The LD50 test has come under attackfor both ethical and scientific reasons be-cause it uses a large number of animals,measures lethality as its major endpointand produces variable results. Methodsusing fewer animals have been suggestedas alternatives to the LD50 test (OECD,1992; 1996; 1998). Moreover, incorpora-tion of humane endpoints into animal
testing has been advocated for reductionof animal pain and distress (OECD,1999a).
1.2 Refinement assaysA continuum of refinement is noted inthe three assays adopted as alternatives tothe LD50 test. All three assays use feweranimals than the conventional LD 50 test,all three assays emphasize humane treat-ment of animals undergoing testing, andone assay utilizes a major endpoint otherthan lethality as its determining value.
OECD (Organization for EconomicCooperation and Development) Guide-line 423 describes the acute toxic class(ATC) method as follows: “This methodis not intended to allow the calculation ofa precise LD50, but does allow for the determination of a range of exposureswhere lethality is expected since death ofa proportion of the animals is still themajor endpoint of this test. The results ofthe test should allow for classification ac-cording to any of the commonly usedsystems. Due to the sequential nature ofthe approach, the duration of the testcould be longer than the procedure de-scribed in Test Guideline 401. The mainadvantage of this method is that it re-quires a smaller number of animals thanboth the “classical” acute oral toxicity(401) and the alternative fi xed dose
method (420). Moreover, because of thespecific provisions for dose selection andinterpretation, this method should in-crease consistency from laboratory tolaboratory.”
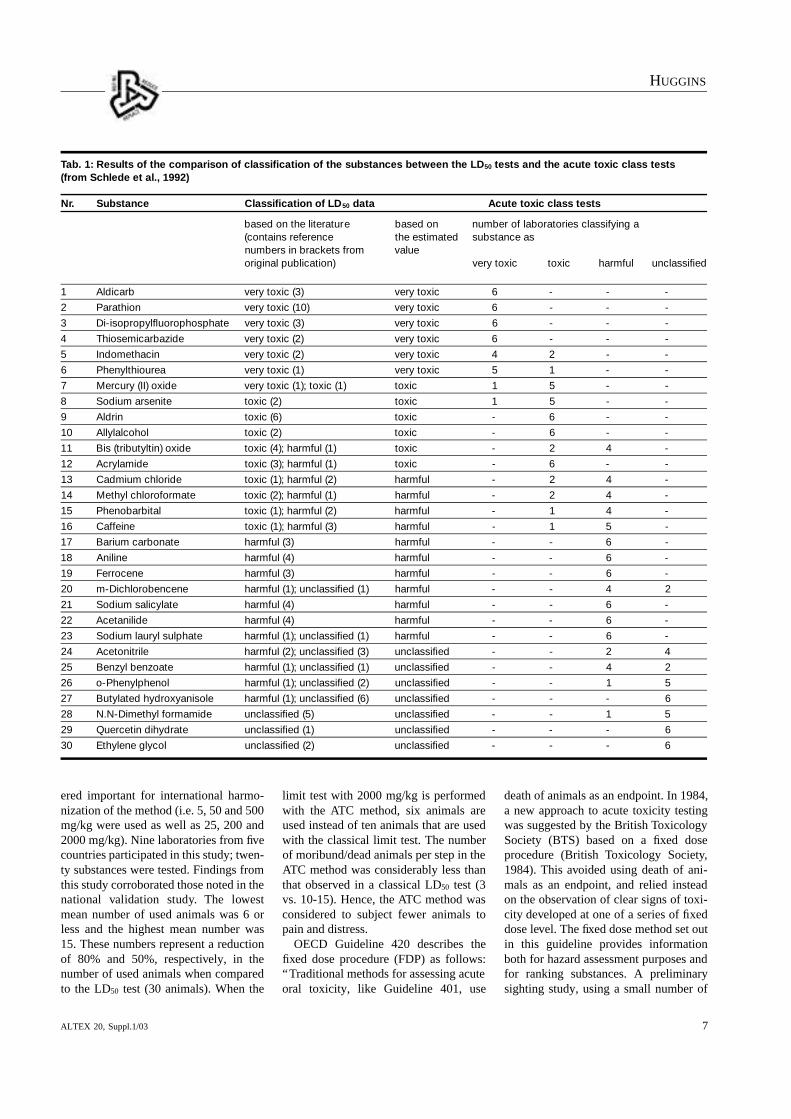
Both national and international valida-tion studies have been conducted to eval-uate the acute toxic class method as analternative to the LD50 test (Schlede etal., 1992; Schlede et. al., 1995). Resultsfrom the national validation effort indi-cated that the method produced reliableresults for the evaluation of toxicity andfor classification of chemicals accordingto the classification system of the Euro-pean Community (Tab. 1).
The ATC method also used substantial-ly fewer animals than the LD50 test, andproduced sufficient information aboutsigns of toxicity. The participants in thisstudy concluded that the ATC methodcould be applicable to routes of exposureother than oral, for example, dermal andinhalation. However, they felt that “be-cause our present knowledge of signs oftoxicity of substances with completelydifferent chemical structures is limitedand that obtaining ‘sufficient repro-ducibility’ of toxic signs is difficult, anyapproach not using death as the endpointwould be difficult to implement”.
The international validation study ofthe ATC method utilized dosages consid-
Zudem wird ihre Bedeutung für die Toxikologie umso er-sichtlicher, je mehr toxikologische Experimente unter derenEinsatz durchgeführt werden. Der Einsatz von Microarrays alsReduction oder Replacement Alternativen zu Tierversuchenmuss als vielversprechend angesehen werden, insbesonderewenn diese als Komponenten von Präscreening Batterien einge-setzt und mit Zellkulturtechniken gekoppelt werden.
8 Validierung von AlternativmethodenDie Validierung von Alternativmethoden entstand aus einer Zeit heraus, als die Richtlinien, welche einer angemessenen Durchführung einer Validierungsstudie zugrunde lagen, haupt-
sächlich über Versuch und Irrtum definiert wurden. Für das R e finement hat diese „Ve rs u chsphase” viel ge b ra cht, ein-schliesslich der Einsicht, dass Validierungsstudien einer solidenGrundlage entspringen sollten, welche aus Elementen wie guteR e f e re n z s t a n d a rds, zuverlässiger Pro t o ko l l t ransfer zwisch e nden Laboratorien und angemessener Einsatz biostatistischerMethoden bestehen. Es sind Anstrengungen im Gange, dieseErkenntnisse in zukünftige Validierungsstudien einfliessen zulassen und Validierungstechniken zwischen den verschiedenenLändern zu harmonisieren, um einen weltweiten Einsatz dergenerierten Daten zu ermöglichen.
Keywords: alternatives to animal use, 3R, validation, toxicity assays, eye corrosion, skin corrosion, contact sensitization, develop-mental/reproductive assays, genetic engineering, transgenics, toxicogenomics
1 Refinement of acute toxicity assays
HU G G I N S
ALTEX 20, Suppl.1/03 7
ered important for international harmo-nization of the method (i.e. 5, 50 and 500mg/kg were used as well as 25, 200 and2000 mg/kg). Nine laboratories from fivecountries participated in this study; twen-ty substances were tested. Findings fromthis study corroborated those noted in thenational validation study. The low e s tmean number of used animals was 6 orless and the highest mean number was15. These numbers represent a reductionof 80% and 50%, respectively, in thenumber of used animals when comparedto the LD50 test (30 animals). When the
limit test with 2000 mg/kg is performedwith the ATC method, six animals areused instead of ten animals that are usedwith the classical limit test. The numberof moribund/dead animals per step in theATC method was considerably less thanthat observed in a classical LD50 test (3vs. 10-15). Hence, the ATC method wasconsidered to subject fewer animals topain and distress.
OECD Guideline 420 describes thefixed dose procedure (FDP) as follows:“Traditional methods for assessing acuteoral toxicity, like Guideline 401, use
death of animals as an endpoint. In 1984,a new approach to acute toxicity testingwas suggested by the British ToxicologySociety (BTS) based on a fixed dose procedure (British Toxicology Society,1984). This avoided using death of ani-mals as an endpoint, and relied insteadon the observation of clear signs of toxi-city developed at one of a series of fixeddose level. The fixed dose method set outin this guideline provides informationboth for hazard assessment purposes andfor ranking substances. A preliminarysighting study, using a small number of
Tab. 1: Results of the comparison of classification of the substances between the LD50 tests and the acute toxic class tests(from Schlede et al., 1992)
Nr. Substance Classification of LD50 data Acute toxic class tests
based on the literature based on number of laboratories classifying a (contains reference the estimated substance asnumbers in brackets from value original publication) very toxic toxic harmful unclassified
1 Aldicarb very toxic (3) very toxic 6 - - -
2 Parathion very toxic (10) very toxic 6 - - -
3 Di-isopropylfluorophosphate very toxic (3) very toxic 6 - - -
4 Thiosemicarbazide very toxic (2) very toxic 6 - - -
5 Indomethacin very toxic (2) very toxic 4 2 - -
6 Phenylthiourea very toxic (1) very toxic 5 1 - -
7 Mercury (II) oxide very toxic (1); toxic (1) toxic 1 5 - -
8 Sodium arsenite toxic (2) toxic 1 5 - -
9 Aldrin toxic (6) toxic - 6 - -
10 Allylalcohol toxic (2) toxic - 6 - -
11 Bis (tributyltin) oxide toxic (4); harmful (1) toxic - 2 4 -
12 Acrylamide toxic (3); harmful (1) toxic - 6 - -
13 Cadmium chloride toxic (1); harmful (2) harmful - 2 4 -
14 Methyl chloroformate toxic (2); harmful (1) harmful - 2 4 -
15 Phenobarbital toxic (1); harmful (2) harmful - 1 4 -
16 Caffeine toxic (1); harmful (3) harmful - 1 5 -
17 Barium carbonate harmful (3) harmful - - 6 -
18 Aniline harmful (4) harmful - - 6 -
19 Ferrocene harmful (3) harmful - - 6 -
20 m-Dichlorobencene harmful (1); unclassified (1) harmful - - 4 2
21 Sodium salicylate harmful (4) harmful - - 6 -
22 Acetanilide harmful (4) harmful - - 6 -
23 Sodium lauryl sulphate harmful (1); unclassified (1) harmful - - 6 -
24 Acetonitrile harmful (2); unclassified (3) unclassified - - 2 4
25 Benzyl benzoate harmful (1); unclassified (1) unclassified - - 4 2
26 o-Phenylphenol harmful (1); unclassified (2) unclassified - - 1 5
27 Butylated hydroxyanisole harmful (1); unclassified (6) unclassified - - - 6
28 N.N-Dimethyl formamide unclassified (5) unclassified - - 1 5
29 Quercetin dihydrate unclassified (1) unclassified - - - 6
30 Ethylene glycol unclassified (2) unclassified - - - 6

HU G G I N S
ALTEX 20, Suppl.1/038
animals, is included in this guideline inorder to estimate the dose effect for toxi-city and mortality and to provide infor-mation on dose selection for the mainstudy. Results from the sighting and mainstudies enable compounds to be rankedin different classification systems, cur-rently in use.”
Van den Heuvel et al. (1990) conduct-ed an international validation study onthe fixed dose procedure as an alternativeto the classical LD50 test. 33 laboratoriesin 11 countries evaluated the toxic effectsof 20 substances using the fixed doseprocedure and compared these effects tothose obtained using the classical LD50
test (Tab. 2).This investigation produced consistent
results that were not substantially affect-ed by inter-laboratory variations and pro-vided adequate information on signs oftoxicity including their nature, time toonset, duration and outcome. Fewer ani-mals than the OECD guideline for acutetoxicity testing (401) were used and animals were subjected to less pain and
distress than the classical LD5 0 test. Utilization of this method enabled sub-stances to be ranked according to theEEC classification system. However, thisvalidation effort also highlighted thevariability between laboratories estab-lished in different countries in assess-ments of signs of toxicity on which a decision to intervene and humanely killanimals is based. As noted in the valida-tion studies of the ATC method, the in-vestigators of the FDP concluded that theprinciple of the procedure was clearlyapplicable to acute toxicity testing bydermal or inhalation routes.
OECD Guideline 425 describes the up-and-down procedure (UDP) as follows:“This test procedure is of principal valuein minimizing the number of animals re-quired to estimate the acute oral toxicityof a chemical and in estimating a medianlethal dose. The median lethal dose allows for comparison with historical data. In addition to the observation ofmortality, it allows the observation ofsigns of toxicity. The latter is useful for
classification purposes and in the plan-ning of additional toxicity tests.”
Bruce (1985) developed the UDP byfirst conducting a historical review of alarge number of conventional acute toxi-city studies. These studies were used as abasis for choosing length of time be-tween successive doses, the sex of the animals to be tested and the spacing between doses in the UDP. The secondinvestigation was a computer simulationbased on data contained in the historicaldataset. The results from the simulationwere in excellent agreement with the historical data indicating that the UDPcould be used as an alternative to theclassical LD5 0 test. Subsequently, theBruce procedure was adopted by theAmerican Society for Testing and Materials (ASTM, 1987). Bruce suggest-ed that the UDP offered substantial savings in numbers of animals althoughhe indicated that estimated LD50 valueswill be less precise than those obtainedfrom larger experiments. Moreover, hecautioned that the method may be inap-
Tab. 2: A comparison of the classification allocated to the test substances by the LD50 and fixed dose tests (from Van den Heuvel et al., 1990).*Four laboratories allocated “very toxic” on the basis of a dose ranging study only
Fixed dose tests –number of laboratories classifying compound as:
Compound LD50 test classification Very toxic Toxic Harmful Unclassified
A Nicotine Toxic - 23 3 -
B Sodium pentachlorophenate Harmful - 1 25 -
C Ferrocene Harmful/unclassified - - 3 23
D 2-Chloroethyl alcohol Toxic - 19 7 -
E Sodium arsenite Toxic - 25 1 -
F Phenyl mercury acetate Toxic 2 24 - -
G p-Dichlorobenzene Unclassified - - - 26
H Fentin hydroxide Toxic - 8 17 1
J Acetanilide Harmful - - 4 22
K Quercetin dihydrate Unclassified - - - 26
L Tetrachlorvinphos Unclassified - - 1 25
M Piperidine Harmful - 2 24 -
N Mercuric chloride Toxic - 25 1 -
P 1-Phenyl-2-thiourea Toxic harmful 12 12 2 -
R 4-Aminophenol Harmful - - 17 9
T Naphthalene Unclassified - - - 26
U Acetonitrile Harmful - - 4 22
W Aldicarb (10%) Very toxic 22* - - -
X Resorcinol Harmful - - 25 1
Y Dimethyl formamide Unclassified - - - 26

HU G G I N S
ALTEX 20, Suppl.1/03 9
propriate for chemicals typically produc-ing animal death two or more days afteradministration.
Three validation studies of the UDPprocedure have been conducted in whichthe ability of the UDP to estimate theL D5 0 was compared to that obtained using the traditional method described in Testing Guideline 401 (Bruce, 1987;Bonnyns et al. 1988; Yam et al., 1991).For all 25 chemicals evaluated, the average ratio of the LD50’s for the twomethods compared was 1.76. These dataindicate that the two methods essentiallyprovide the same point estimate of theLD50 for the chemicals tested (Fig. 1).
1.3 Comparison of refinement assays
When the up-and-down and fixed doseprocedures were compared against theclassical LD50 test by Yam et al. (1991),both methods were found to reduce thenumbers of animals used while providingadequate information for ranking the 10 materials tested according to the European Economic Commission classi-fications for acute oral toxicity. The signs
observed and the duration of signs tend-ed to vary among methods. The authorsconcluded that while different doses usedin the three methods probably accountedfor most of the observed differences in signs, they also considered that labora-tory variations in sign recording mayhave also contributed to the observed differences. In total, for the 10 test mate-rials, the classical LD50 test generated 67 signs, the up-and-down method generated 62 signs, and the fixed doseprocedure generated 49 signs. Compar-ing the fixed dose procedure and the up-and-down method with regard to autopsyfindings resulted in the finding that the fixed dose procedure produced fewerautopsy findings. This was not surpris-ing, since the fixed dose procedure gen-erally used lower doses than the up-and-d own method. Both alternatives usedfewer animals than the classical method.By using females, the up-and-dow nmethod required only 50% as many ani-mals as the fixed dose procedure, and29% as many as the classical method.The fixed dose procedure produced thefewest deaths of the three tests.
Further comparisons of the up-and-down and fixed dose procedures, andconventional LD50 test were performedby Lipnick et al. (1995). The authors’major conclusions that favor the UDP areas follows:
• The UDP generally produces an esti-mate of the LD50 that is similar to thatachieved from conventional acute toxi-city testing.
• Data on chemicals tested in the UDPlead to the same EEC acute toxicityc l a s s i fication as do those from the conventional LD50 test in 23 of 25 re-viewed cases. These results are as goodas those for the FDP vs. conventionalLD50 test where 16 out of 20 classifica-tions are coincident. For seven out of10 cases, the UDP and FDP lead to thesame classification.
• The UDP gives an estimate of the LD50
and thus data from this test method areapplicable to any acute toxicity classi-fication system. In contrast, FDP dataare directly referable to the classifica-tion system used by the EEC. How-ever, by use of the information fromthe sighting study for the FDP, classi-fication decisions can be made for other reasons.
• Testing with the UDP requires only between 6 and 10 animals of one sex,the smallest number of animals of anyprotocol. In contrast, the FDP usuallyuses 10 or 20 animals, while the con-ventional LD50 determination generallyrequires 30 animals (15 if only one sexis used). Moreover, the OECD proto-cols for the conventional test and FDPcall for a sighting study which uses upto another five animals, a sightingstudy is not needed for the UDP.
• To date, the UDP has been used toevaluate lethality as an endpoint. Giventhat the frequencies of toxic manifesta-tions are similar for the chemicals thathave been simultaneously investigatedin the UDP and the FDP (72% and64%, respectively), it seems reasonableto explore further the applicability ofthe UDP to non-lethal toxicity end-points.
• Analyses conducted here, and a reviewof the literature, indicate that the twos exes usually respond similarly inacute oral toxicity tests. When re-sponses differ, females are generally
Fig. 1: Comparison of the LD50 determined using the up-and-down method with the LD50 estimated from conventional tests for materials testes by van den Heuvel(1990) and Yam et al. (1991) (•), Bruce (1987) (◊) and Bonnyns et al. (1990) (°)(from Lipnick et al., 1995).

HU G G I N S
ALTEX 20, Suppl.1/0310
more sensitive than males. Considera-tion should be given to restricting acutetoxicity testing of chemicals to femalesunless there is information suggestingthat males are more sensitive for a given substance.
1.4 Humane endpointsRussell and Burch (1959) defined refine-ment as any development leading to a“decrease in incidence or severity of inhumane procedures applied to thoseanimals which have to be used”. Hence,incorporation of humane endpoints intoanimal testing protocols adds consider-able refinement to these studies. Animalsundergoing testing for endpoints such astumor production, infectious disease,vaccine potency, and target organ toxici-ty are treated more humanely by such ameasure and studies in which lethality isthe major endpoint measured can actual-ly be replaced by those in which othersigns of toxicity are monitored. However,as noted by the researchers below, cleardefinition of humane endpoints and de-velopment of the methods by which theycan be assessed effectively are obviouslynecessary.
Morton (2000) describes a systematicapproach for establishing humane end-points. He advocates the use of scoresheets that list the cardinal clinical signsthat are observable and measurable, andthe key clinical signs are identifi e dthrough the experience of those involvedin the research. He suggests that lists ofclinical signs be developed by ve r yclosely observing the first few animalsundergoing a new scientific procedure.The list is modified with experience untila set of signs is established that most an-imals will show during that experimentand that are relevant to the assessment ofpain and distress. These cardinal signsare set out against time in the score sheet.Use of these score sheets encouragescloser observation of animals by all in-volved at critical times in the experiment,subjective assessments are avoided to alarge extent, and consistency of scoringis increased.
Toth (2000) has advanced a data-basedapproach for predicting imminent deathand defining specific moribund condi-tions in objective terms. She indicatesthat the moribund state can be defined by
identifying the values of various vari-ables that precede imminent death thatcan serve as “signals” for preemptiveeuthanasia. She stresses that specifi cvariables should be identified andweighted in terms of their predictivevalue. However, she acknowledges thatobjective data-based criteria that predictimminent death may not always fit com-fortably into the goals of an experiment.Hypothermia, inability to rise or ambu-late, weight loss, and biochemical variables are all suggested as potentialpredictors of imminent death.
Schlede et al. (2000) discuss specifi-cally the use of humane endpoints inacute oral toxicity testing. Their evalua-tion of clinical signs was made in ratsused for validation studies of the acutetoxic class method. These data demon-strated that all forms of “convulsions” resulted in death in 94% (484/516) ofrats, and the “lateral position” resulted indeath in 79% (177/223) of rats. Clinicalsigns associated with a high mortalityrate in this study are listed in Table 3.
Wallace (2000) discusses humane end-points in cancer research and makes thefollowing suggestions:
• Tumor growth or excision should re-place “survival” endpoints.
• M a ny preparatory procedures (e.g.l ow - l evel whole body irradiation, i m m u n o s u p p r e s s ive agents, surg i c a l
ablation of endocrine glands) may re-present a greater challenge to an ani-mal that than of a developing tumor;hence, humane endpoints should con-sider the cumulative effect of all exper-imental challenges.
• Tumor development and animal condi-tion should be monitored frequentlybecause unexpected or uncontrolled tumor development can result in unnecessary animal distress or mor-tality.Olfert and Godson (2000) propose that
increases in serum levels of cytokines beused as indicators of the presence of in-fectious disease and as predictors of bothonset and outcome of infectious disease.This proposition is supported by the factthat changes in the levels of these param-eters occur early in the disease process,before severe behavioral and physiologicchanges do. Additionally, body weightchange, weight loss, and decreased activ-ity are reflective of changes in cytokinelevel. The authors suggest that changessuch as these are all measurably morehumane endpoints than is allowing pro-gression of the infectious disease withinthe animal model.
Dennis (2000) writes that “death is notusually intended in genetic engineeringstudies, but lethality or animals with severe health problems are commonlyencountered. Genetically-engineered
Tab. 3: Clinical signs associated with a high mortality rate (from Schlede, Gerner and Diener, 2000).
Clinical sign Number of rats Dead/moribund rats %
Convulsion
-Convulsion (unspecified) 43 43 100
-Clonic convulsion 218 207 95
-Tonic convulsion 96 79 82
-Tonic-Clonic convulsion 125 122 98
-Saltatory convulsion 10 10 100
Lateral position 223 177 79
Ventral position 9 9 100
Tremor 389 296 76
Gasping 143 108 76
Vocalisation 97 79 81
Extension spasm 6 6 100
Flexion spasm 8 8 100
Coma 9 9 100
Decrease of muscle tone 18 16 89
Mucoid faeces 35 27 77

HU G G I N S
ALTEX 20, Suppl.1/03 11
animals often have a decreased ability to resist disease, increased tumor pro-duction, or compromised basic bodilyfunctions such as eating or breathing”.He suggests that it is extremely impor-tant that institutions supervise and continually review ongoing studies toidentify problems as they occur and toensure that appropriate humane end-points are established.
These comments all indicate that humane endpoints can be incorporatedinto many diverse toxicological proto-cols. Perhaps the most encouraging regulatory support for doing so is foundin guidelines promulgated by the OECDwhich summarize the use of clinicalsigns as humane endpoints for experi-mental animals used in safety evaluation.Included in these guidelines is a listing oftypes of effects that should be monitoredin an adequate evaluation of an animal todetermine its condition and whetherthere might be evidence indicative ofpain and or distress:
• Changes in physical appearance (e.g.coat texture; hair soiled with urine orfaeces)
• Changes in clinical signs (e.g. respira-tion rate; posture)
• Changes in unprovoked behavior (e.g.self mutilation; compulsive behavior)
• Behavioral changes in response to ex-ternal stimuli (e.g. excitability; rightingreflex)
• Changes in body weight, and relatedchanges in food and water consump-tion
• Changes in clinical parameters (e.g.body temperature, heart and respirationrate, clinical chemistry and hema-tology).This listing concurs with many of the
suggestions given by the researchersabove. Moreover, the guiding principlesof this OECD document include thestatement that, “severe pain, suffering, ordeath are to be avoided as endpoints”.Hence, continued usage of any test wherelethality is the endpoint appears to be inconflict with these guidelines.
1.5 Regulatory activitiesThe body of information about refine-ment alternatives to the conve n t i o n a lLD50 test has reached “critical mass” inthat three alternative assays have been
extensively studied and deemed appro-priate as replacements for the conven-tional LD5 0 test by members of the scientific community and, to a certain extent, by those of the regulatory com-munity as well. Furthermore, incorpora-tion of humane endpoints (which are defined in at least one case as thosewhich do not include death) into toxicitytesting is being strongly encouraged bymembers of both communities. In De-cember 2002, the Test Guideline 401 (theconventional LD50 test) has been deletedand replaced by alternative methods ofacute toxicity testing (OECD, 1999b). Inorder to accomplish this objective, sever-al revisions in the three alternative assaysavailable need to be made. Despite thefact that the UDP is the only test that pro-vides a point estimate of the LD50, it doesnot provide estimates of the slope of the dose-response curve and confidenceinterval that are needed by regulatoryagencies in some instances. Therefore,these variables need to be included in arevised procedure. Moreover, both theFDP and ATC method need to bechanged to reflect changes in the regula-tory classification scheme brought aboutby recent global harmonization efforts.
The USEPA has agreed to revise theUDP to include a procedure that wouldprovide slope and corresponding confi-dence interval estimates. Accordingly, arevised UDP has been undergoing expertreview by the Interagency CoordinatingCommittee on Validation of AlternativeMethods (ICCVAM) (USEPA, 2000a).The revised procedure includes a modi-fied up-and-down procedure that im-proves performance, a modified limit testthat utilizes only females and provides alimit dose of 5000 mg/kg for specificregulatory purposes, and an added sup-plemental test for determining the slopeand confidence interval. The panel’s re-view of this revised procedure has beencompleted (July 25, 2000); their recom-mendations are to be posted by the end ofJanuary, 2001 (USEPA, 2000b).
1.6 Summary, conclusions, and future work
In December 2002 the Test Guideline401 (conventional LD50 test) has beendeleted. Three assays have been adopted
as replacements for the conve n t i o n a lLD50 test. The assays differ primarily asto the endpoint they measure: the ATCmethod and FDP provide ranges of values that are applicable to particularregulatory classification schemes; where-as, the UDP provides an actual point estimate of the LD50 value. However, allof these assays use fewer animals thanthe conventional LD50 test.
In addition to assays using fewer ani-mals, refinement of acute toxicity testingis being supported by such documents asthe recent set of guidelines issued by theOECD for the use of clinical signs as humane endpoints in toxicity testing.These guidelines suggest strongly thatlethality is no longer an acceptable end-point, hence, they support substitution ofan ED50 value for the LD50 value in acutetoxicity testing. Furthermore, researchersr ev i ewing the refinement assays pro-posed as alternatives to the conventionalLD50 test suggest that humane endpointscan be incorporated effectively into thoseassays.
This recent, rapid progress toward fullr egulatory acceptance of alternative assays which refine acute toxicity testingrepresents an exciting chapter in the his-tory of the alternatives to animal testingmovement. However, in view of the factthat the ultimate goal of this research isreplacement of animals in acute toxicitytesting, further work will be focused toward validation of true replacement alternatives (i.e. those that do not use animals.) Exemplary research efforts inthis area have been those by the Multi-center for the Evaluation of In VitroCytotoxicity (MEIC). The results of thiswork have just been published (Clemed-son and Ekwall, 1999; Ekwall, 1999).They were the focal point of a publicmeeting sponsored by ICCVAM that investigated alternative assays for pre-dicting acute systemic toxicity in order tolay a framework for further regulatoryacceptance (ICCVAM, 2000).
Worldwide acceptance and incorpora-tion of refinement assays such as, and including those discussed above shouldbe seen in the next five years, if not soon-er judging from recent events. Batteriesof cytotoxicity tests should also be usedmuch more frequently for prediction ofthe acute toxicity of new chemical com-

HU G G I N S
ALTEX 20, Suppl.1/0312
pounds during this time frame. The onlys i g n i ficant hurdle remaining to becleared is that of acceptance of an end-point other than lethality. Our reliance onthe LD50 as the endpoint of choice is, inmany respects, simply a product of priorconditioning as well as a lack of feasiblealternatives, neither of which should im-pede our progress now.
By the end of the next ten years, cyto-toxicity assays should have been re-searched fully enough to provide data tosupport validation of one or more ofthese methods as replacements for theuse of animals in acute toxicity testing.By the end of the next twenty years, regulatory acceptance of cytotoxicity orother similar endpoint as a reliable indicator of acute toxicity should be inevidence. Moreove r, the mechanisticevents linking cytotoxicity in cell cultureto lethality in the whole organism shouldbe well-defined.
Future research and funding effortsshould be directed toward more precisemethods of defining humane endpointsas well as validation of cytotoxicity assays as replacements for animal use inacute toxicity testing. Although muchconcerned thought and action has beendirected toward the incorporation of humane endpoints into toxicity proto-cols, further work (as noted by all the investigators cited above) is definitelyneeded in order to bring definition andresolution to observations of clinicalsigns. Systematic methods of observationand effective in-depth training of animal-handling personnel are critical to the successful implementation of humaneendpoints, particularly if the data are tobe used in a quantitative fashion. More-over, work in which patterns of toxic signdevelopment are studied in relation toboth the animal model used and thechemical(s) administered should be supported.
Many methods currently exist for themeasurement of cytotoxicity endpoints.Efforts should be made to assess whichare the most cost-effective, reliable indi -cators of acute toxicity when comparedto either animal LD50 or human acutelethality data or, perhaps, to both. Thistype of validation will prove invaluableto the development of effective predictivebatteries of these methods as well as to
their eventual acceptance by regulatorybodies. The work performed by the MEIC is a good starting point for thistype of endeavor because it providescomparative information among manydifferent types of cytotoxicity assay.
ReferencesASTM (American Society for Testing
and Materials) (1987). Standard testmethod for estimating acute oral toxi-city in rats. Designation: E 1163-87. In Annual Book of ASTM Standards,Philadelphia.
Bonnyns, E., Delcour, M. and Vral, A.(1988). Up-and-Down Method as anA l t e r n a t ive to the EC-Method forAcute Toxicity Testing. IHE ProjectNo. 2153/88/11. Institute of Hygieneand Epidemiology, Ministry of PublicHealth and the Environment, Brussels.
British Toxicology Society (1984). Special report: A new approach to the c l a s s i fication of substances and preparations on the basis of their acutetoxicity. Hum. Toxic. 3, 85-92.
Bruce, R. (1985). An up-and-down pro-cedure for acute toxicity testing. Fund.Appl. Toxicol. 5, 151-157.
Bruce, R. (1987). A confirmatory studyfor the up-and-down method for acutetoxicity testing. Fund. Appl. Toxicol. 8,97-100.
Clemedson, C. and Ekwall, B. (1999).Overview of the final MEIC results: I. The in vitro-in vivo evaluation. Toxic. In Vitro 13, 657-663.
Dennis, M. (2000). Humane endpointsfor genetically engineered animalmodels. ILAR Journal 41(2), 94-98.
Ekwall, B. (1999). Overview of the finalMEIC results: II. The in vitro-in vivoevaluation, including the selection of a practical battery of cell tests for prediction of acute lethal blood concentrations in humans. Toxic. InVitro 13, 665-673.
I C C VAM (Interagency CoordinatingCommittee for the Validation of Alter-native Methods) (2000). InternationalWorkshop on In Vitro Methods for Assessing Acute Systemic To x i c i t y.October 17-20. Arlington, VA.
Lipnick, R., Cotruvo, J., Hill, R. et al.(1995). Comparison of the up-and-down, conventional LD50, and fixed-
dose acute toxicity procedures. F d .Chem. Toxic. 33(3), 223-231.
Morton, D. B. (2000). A systematic approach for establishing humane endpoints. ILAR Journal 41(2), 80-86.
OECD (Organization for Economic Cooperation and Development) (1987).Guideline No. 401 – Acute Oral Toxicity.
OECD (1992). Guideline No. 420 –Acute Oral Toxicity: Fixed Dose Procedure.
OECD (1996). Guideline No. 423 –Acute Oral Toxicity: Acute To x i cClass.
OECD (1998). Guideline No. 425 –Acute Oral Toxicity: Up and DownProcedure.
OECD (1999a). Guidance Document onHumane Endpoints for ExperimentalAnimals Used in Safety EvaluationStudies. Paris: OECD.
OECD (1999b). OECD DocumentENV/JM (99) 19, Test Guidelines Pro-gramme, Acute Oral Toxicity Testing:Data Needs and Animal Welfare Con-siderations, 29th Joint Meeting, June8-11, Paris, France.
Offert, E. and Godson, D. (2000). Hu-mane endpoints for infectious diseaseanimal models. ILAR Journal 41(2),99-104.
Russell, W. and Burch, R. (1959). ThePrinciples of Humane ExperimentalTechnique. London: Methuen & Co.LTD. (Reissued: 1992, Unive r s i t i e sFederation for Animal Welfare, Herts,England).
Schlede, E., Mischke, U., Roll, R. andKayser, D. (1992). A national valida-tion study of the acute-toxic-classmethod – An alternative to the LD50
test. Arch. Toxicol. 66, 455-470.Schlede, E., Mischke, U., Diener, W. and
Kayser, D. (1995). The internationalvalidation study of the acute toxicclass method (oral). Arch. Toxicol. 69 ,659-670.
Schlede, E., Gerner, I. and Diener, W.(2000). The use of humane endpointsin acute oral toxicity testing. In M.Balls, A.-M. Zeller and M. E. Halder(eds.), Progress in the Reduction, Re-finement and Replacement of AnimalExperimentation (907-914). A m s t e r-dam, London, New York, Tokyo: Else-vier 11.

HU G G I N S
ALTEX 20, Suppl.1/03 13
Toth, L. (2000). Defining the moribundcondition as an experimental endpointfor animal research. ILAR Jo u r n a l41(2), 72-79.
U S E PA (United States Env i r o n m e n t a lProtection A g e n cy) (2000a). Notice(65 FR 08385): Request for Data andNomination of Expert Panel of Scientists to Participate in the Indepen-dent Peer Review Evaluation of theRevised Up-and-Down Procedure forAssessing Acute Oral To x i c i t y.
Fe d e ral Register Volume 65(34). February 18.
USEPA. (2000b). Notice (65 FR 35109)of Peer Rev i ew Meeting on the R evised Up-and-Down Procedure(UDP) as an Alternative Test Methodfor Assessing Acute Oral Toxicity; Request for Comments. Federal Regis-ter Volume 65 (106). June 1.
Van den Heuvel, M., Clar, D., Fielder, R.et al. (1990). The international valida-tion of a fixed-dose procedure as an
alternative to the classical LD50 test.Fd. Chem. Toxic. 28(7), 469-482.
Wallace, J. (2000). Humane endpointsand cancer research. ILAR Jo u r n a l41(2), 87-93.
Yam, J., Reer, P. and Bruce, R. (1991).Comparison of the up-and-dow nmethod and the fixed-dose procedurefor acute oral toxicity testing. F d .Chem. Toxic. 25, 259-263.
2 Alternatives to eye corrosion/irritation testing in animals
2.1 BackgroundCurrent testing guidelines for eye corro-sion/irritation testing promulgated byOECD (Organization for Economic Cooperation and Development) andU S E PA (United States Env i r o n m e n t a lProtection A g e n cy) use the follow i n gdefinitions for corrosion and irritation.Eye corrosion is defined as the “produc-tion of irreversible tissue damage in theeye following application of a test substance to the anterior surface of theeye”. Eye irritation is defined as the “production of reversible changes in theeye following the application of a testsubstance to the anterior surface of theeye”. (OECD, 1987; USEPA, 1998).These test guidelines also incorporate thefollowing humane considerations whichsupport the use of alternatives to eye corrosion/irritation testing: 1. “Strongly acidic or alkaline sub-
stances, for example, with a demon-strated pH of 2 or less or 11.5 orgreater, need not be tested owing totheir predictable corrosive properties.Buffer capacity should also be takeninto account.
2. Materials which have demonstrateddefinite corrosion or severe irritationin a dermal study need not be furthertested for eye irritation. It may be presumed that such substances willproduce similarly severe effects in theeyes.
3. Results from well validated and ac-cepted in vitro test systems may serveto identify corrosives or irritants such
that the test material need not be tested in vivo.”Furthermore, the number of animals
used in testing is limited by these guide-lines as follows: “A single animal shouldbe considered if marked effects are antic-ipated. If the results of this test in one animal suggest the test substance to be asevere irritant (reversible effect) or corro-sive (irreversible effect) to the eye usingthe procedure described, further testsmay not need to be performed. In casesother than a single animal test, at leastthree animals should be used. Occasion-ally, further testing in additional animalsmay be appropriate to clarify equivocalresponses.”
Moreover, a draft revised version ofOECD Guideline 405 includes the recommendation that an integrated test-ing strategy for a stepwise evaluation ofall existing information on the substanceincluding, e.g., data from human ex-perience and from in vitro tests be in-corporated (OECD, 2000). Hence, muchr egulatory support of alternatives to eye corrosion/irritation testing in animalsis in evidence. Furthermore, many alter-native models for eye corrosion/irritationtesting have been developed. Unfortu-nately, validation of these models has remained elusive. A recent report by ECVAM (European Center for the Vali-dation of Alternative Methods) discussesseveral potential reasons for this lack ofvalidation and puts forth suggested initiatives for remedy (Balls et al., 1999).These include the use of reference stan-
dards (RS), stepwise testing strategies,multivariate and other statistical tech-niques for the further analysis of datagenerated in previous validation studies,and a program of mechanistic research.
2.2 In vitro alternativesHistorically, alternatives to eye corro-sion/irritation testing in animals wereconsidered the most important assays todevelop in view of the unquestionablepain and suffering experienced by ani-mals upon which severely irritating andc o r r o s ive substances were tested. Numerous in vitro assays have been developed over the last twenty years inresponse to this need. Some of the morew e l l - k n own tests include the bov i n ecorneal opacity and permeability(BCOP) assay (Gautheron et al., 1992),the hen’s egg test – chorioallantoic mem-brane (HET-CAM) assay (Lopke, 1986),and several cytotoxicity tests (for exam-ple, 3T3-neutral red uptake (3T3-NRU))(Borenfreund and Borrero, 1984; Boren-freund and Puerner, 1985).
2.2.1 Combinatorial approachesAs mentioned above, a number of effortsdirected toward validation of a single inv i t ro assay of eye corrosion/irritationhave been conducted without consider-able success. However, when multivari-ate analysis was applied to the resultsfrom these studies, it indicated that assays used together in complementaryfashion may provide good predictive information. Multivariate analysis was

HU G G I N S
ALTEX 20, Suppl.1/0314
applied to results obtained from the European Commission/British Home Office (EC/HO) validation study (Ballset al., 1995). The analysis revealed thatcombinations of data from assays of epithelial integrity (fluorescein leakage(FL) test), ex vivo models (isolated rabbiteye (IRE), isolated chicken eye (ICE))and a cytotoxicity test (neutral red uptake(NRU)) explained more of the variabilityin the data than any single test usedalone. This finding resulted in calculation of a better prediction model(PM). Similarly, when multiva r i a t eanalysis of data obtained from a valida-tion study conducted under the auspicesof COLIPA (European Cosmetic, Toi-letry and Perfumery A s s o c i a t i o n )(Bagley et al., 1997) was performed, results comparable to those obtained inthe EC/HO study were obtained (i.e. improved PMs could be developed basedon combinations of in vitro endpoints).Furthermore, a validation study was coordinated by the Centre for Documen-tation and Evaluation of A l t e r n a t iveMethods to Animal Experiments (ZEBET) at the Bundesinstitut fürRisikobewertung (BfR), and supportedfinancially by the German Department ofResearch and Technology (BMBF)(Spielmann et al., 1993; Spielmann et al.,1996). Results from multivariate analysisof this study indicated that chemicals could be reliably classified as severe irritants (classification R41) through the combined use of the HET-CAM testand the 3T3 NRU test.
Hence, analysis (and re-analysis) of results from several validation studiessuggests that complementary pairing ofin vitro assays (usually a cytotoxicity testwith an organotypic test) can be consid-ered useful to the prediction of eye cor-rosion/irritation. Independent researchinvestigations of the predictivity of in vitro assays used in combination (batter-ies) have also yielded good results (forexample, Lewis et al., 1994; Pham andHuff, 1999; Rosenkranz and Cunning-ham; 2000).
The phrase “combinatorial approach”can be applied not only to combiningcomplementary in vitro assays into batteries of tests, but also to combiningdata from many different types of exper-iments, not just in vitro assays. When a
large combination of different data typesis analyzed, this procedure is often referred to as “tier-testing”, “hierarchicaltesting” or as a “stepwise strategy”. Results from the complementary pairingof in vitro assays can contribute much tohierarchical testing schemes such as thestepwise strategy currently suggested inthe OECD revised 405 guidelines. Thef o l l owing points from these rev i s e dguidelines outline the types of data to beconsidered prior to in vivo testing, in-cluding results from ex vivo and in vitroassays.
• Existing human or animal data: Whenthere is sufficient human data from thetest substance, it may not need to betested in animals.
• Structure activity relationships (SAR).Historical experience (including hu-man data) or testing of structurally re-lated chemicals should be evaluated. Ifthere are sufficient data to indicate theeye irritancy/corrosivity potential of achemical or mixture from analogues,the test substance can be presumed toproduce similar responses. SAR expe-riences should be interpreted cautious-ly when evaluating non-irritating/corrosive substances.
• P hysicochemical properties and chem-ical reactivity. Strongly acidic or alka-line substances which can be expectedto result in a pH in the eye of 2 or less,or 11.5 or greater, may not need to betested owing to their probable corro-s ive properties. Buffering capacity (alkaline or acidic reserve) should also be taken into consideration.
• Results from skin irritation studies.Substances that have demonstrated severe skin irritancy or corrosivity in asingle application dermal study maynot need to be tested for eye irritancyand corrosion. It can be presumed thatsuch substances will produce similarsevere effects on the eyes.
• Results from other studies. If a sub-stance is highly toxic by the dermalroute, it need not be tested in the eyesbecause it can be assumed to be highlytoxic by this route as well.
• Results from in vitro or ex vivo teststhat are generally accepted for pur-poses of hazard or risk assessment.Substances that have demonstrated thepotential in an in vitro or ex vivo
study to be corrosive or a severe irritantmay not need to be tested for irritationand corrosion in vivo. It can be pre-sumed that such substances will pro-duce similar severe effects on the eyes.
• If there is insufficient evidence withwhich to evaluate the potential eye i r r i t a t i o n / c o r r o s ivity of a substancefrom the preceding information, a skinirritation/corrosion test (see Guideline404 and its Attachment) should be performed first. If the substance isshown to produce severe skin irritationor corrosion, it can be presumed that itwould also produce similar effects inthe eyes, so that an in vivo eye test neednot be performed.ECVAM has evaluated the stepwise
s t r a t egy suggested by OECD in its r evised guidelines for acute eye irri-tation/corrosion testing and concludedthat the strategy is effective in reducingand refining the use of the Draize eye test(Worth and Fentem, 1999).
2.2.2 Reference standardsBalls et al. (1999) have suggested that areference standards approach be used invalidation studies of in vitro assays ofeye corrosion/irritation. They emphasizethat, “the term ‘reference standard’ (RS)should not be confused with ‘positivecontrol’”. Rather, they define a positivecontrol as a substance which is known togive a positive response and which isused to confirm the correct conduct ofthe assay. Alternatively, a reference stan-dard is a substance which has a knowndegree of toxicity in vivo, and which canbe used in vitro to determine the degreeof toxicity of test substances, whose effects are scaled relative to the RS. This group also hypothesizes that the reference standard approach to eye corrosion/irritation testing in vitro willinclude the following roles:
• within companies, for the developmentand cross-validation of in vitro assays
• in the validation of alternative meth-ods, as a replacement for the totallyblind approach which currently exists,so that substances can be grouped intoc a t egories defined by the referencestandards
• in regulatory toxicology, for the submission of data on selected newsubstances to authorities

HU G G I N S
ALTEX 20, Suppl.1/03 15
An evaluation of the use of referencestandards in the validation process hasbegun by the ECVAM Reference Stan-dards Working Group. Five in vitro testsof eye corrosion/irritation have beennominated as candidates for analysis.These include the ICE, BCOP, HET-CAM, NRU (neutral red uptake), Epi-Ocular™, and RBC (red blood cell) hemolysis assays. Conduct of this evalu-ation will involve testing of referencestandards from different chemicalgroups, development of a PM based onthe results from the reference standards,and application of the derived PM to asecond set of chemicals, the identities of which are unknown. This work, if successful, should lay a much neededfoundation for reliable evaluation of results from validation efforts in terms ofcomparisons to chemicals of known toxicity.
2.2.3 Mechanistic considerationsA recent article by Bruner et al. (1998)highlights the importance of understand-ing the mechanisms of eye irritation,“particularly when attempting to im-prove in vitro prediction of in vivo eye irritancy”. Efforts by ECVAM to evalu-ate the failure of validation studies of eyecorrosion/irritation have also pinpointedunderstanding mechanisms of action behind eye corrosion/irritation as criticalto any future validation/acceptance of in vitro assays of this insult (Balls et al.,1999).
Consideration of underlying mecha-nisms of action is in evidence in much ofthe research and development of in vitroassays of eye corrosion/irritation. For example, recognition of the importanceof mechanisms of action is implicit in theuse of complementary assays. Single assays of cytotoxicity or organotypic effects probably do not explain the entiremechanism of action behind develop-ment of corrosion or irritation. Further-more, measurement of different end-points within the same assay has provento be valuable in discriminating mecha-nisms of action. An excellent example of this is the BCOP assay in which Gautheron et al. (1992) realized the im-portance of measuring more than one indicator of irritation in the bov i n ecornea. Hence, the BCOP assay investi-
gates both opacity and permeability.A recent workshop held in Brussels,
Belgium in October, 1998 as a follow-upto the work discussed by Bruner abovesuggested the following areas as foci formechanistic research of eye corrosion/irritation:
• development of an appropriate set ofreference test substances for use in theresearch
• evaluation of the area and depth ofcorneal injury as markers of eye injury
• exploration of the use of early bio-markers of eye injury (for example, cytokine release)
• development of methods for evaluatingcorneal wound healing
• development of methods for assessingthe kinetics of eye injury
• development of methods for assessinginjury to nerve cells in the corneaDevelopment of an appropriate set of
reference test substances is currently un-d e r way at ECVAM by the Wo r k i n gGroup on Reference Standards. How-ever, much work remains to be done onthe remaining five areas of suggestedmechanistic research indicated above.
Evaluation of the area and depth ofcorneal injury has been addressed by several researchers. Although this workwas performed in vivo, it is thought to have important ramifications for thed evelopment of in vitro a l t e r n a t ives for ocular irritancy. Jester et al. (1996)investigated the application of in vivoconfocal microscopy (CM) to the under-standing of surfactant-induced ocular irritation. The aim of this research was to “assess the ability of in vivo confocalm i c r o s c o py to provide noninva s ively d e r ived histopathologic correlates of s u r factant-induced eye irritation fromwhich specific pathologic mechanismscould be identified”. Rats and rabbits, received anionic or cationic surfactant inone eye with the other eye serving ascontrol. Eyes were examined and scoredfor ocular irritancy subsequently using apenlight and slit-lamp. Corneas werethen evaluated by in vivo CM to evaluateepithelial layer thickness and surface epithelial cell area, corneal thickness,depth of necrosis, inflammation, fibrosis,and endothelial injury. The anionic sur-factant produced slight irritation (peak
scores of 12.4 and 8.0) and in vivo CMrevealed changes limited to the cornealepithelium.
Maurer and colleagues (1997) probedthe uses of CM microscopy further. Sur-factants of slight, mild, moderate, and severe irritancy were directly applied tothe corneas of rabbits and eyes and eye-lids were examined macroscopically andscored for irritation beginning 3 hour after dosing and periodically through day35. Concurrently, the corneas were eval-uated by in vivo CM. Three-dimensionaldata sets extending from the surface epithelium to the endothelium were assessed for surface epithelial cell size,epithelial layer thickness, total cornealthickness and depth of ke r a t i n o cy t enecrosis. Results indicated that signifi-cant differences in area and depth of in-jury occur with surfactants of differingirritancy. Furthermore, the data suggest-ed that differences at 3 hours can be usedto distinguish different levels of ocularirritation. Application of CM microscopyto evaluation of the irritancy of unknowns u r factants has also been performed(Maurer et al., 1998). Macroscopic andmicroscopic findings regarding the ocu-lar irritation of six surfactants of relative-ly unknown irritancy were compared tothose of six surfactants of known irritan-cy. The right eye of each rat tested received the surfactant directly on thecornea. Untreated left eyes served ascontrols. At 3 hours and on days 1, 3, and35, eyes and eyelids were collected formicroscopic examination. Macroscopicand microscopic findings indicated thatthree surfactants were similar to mildlyirritating surfactants and three were sim-ilar to moderately irritating surfactantspreviously studied.
The premise that cytokines can be usedas an early biomarker of corneal injury issupported by Planck (1999) who ex-plained their role in this way: “Infectionand tissue damage activate nearby cellsto produce a group of proteins, called cy-tokines, which help mediate the resultinginflammatory and repair processes. Inter-leukin-1 (IL-1), IL-6, and tumor necrosisfactor alpha (TNF1) are often calledmaster cytokines because they are pro-duced by many cell types and have mul-tiple effects on target cells including synthesis of additional cytokines. A l l

HU G G I N S
ALTEX 20, Suppl.1/0316
three cellular layers of cornea appear capable of producing and responding to these master cytokines, although theregulation of production and the reper-toire of responses are not clear. Knowl-edge of the roles of these master cy-tokines in response to corneal insultsshould enhance the development ofmethods to manipulate repair and inflam-matory processes therapeutically.”
Cytokine release has been investigatedas a measure of irritancy in dermal cells,considered by some researchers to re-present various aspects of ocular tissue.Co-cultures of human dermal fibroblastswith human epidermal keratinocytes andhuman dermal fibroblasts in three-dimensional culture have both been usedas in vitro assays of ocular irritation inwhich inflammatory response was meas-ured with cytokines (Curren et al., 1997).
Future research will undoubtedly in-clude definitive studies identifying thecytokines involved in ocular response toinjury and the kinetics of their action.This work should be greatly aided by theuse of functional human corneal equiva-lents constructed from cell lines (Griffithet al., 1999). These equivalents com-prised the three main layers of the cornea(epithelium, stroma, and endothelium).Each cellular layer was fabricated fromimmortalized human corneal cells thatwere screened for morphological, bio-chemical, and electrophysiological simi-larity to their natural counterparts.Equivalents mimicked human corneas inkey physical and physiologic functions,including morphology, biochemicalmarker expression, transparency, ion andfluid transport, and gene ex p r e s s i o n .Work performed by Sotozono et al.(1997) profiles cytokine expression f o l l owing injury to mouse corneas. Although performed in vivo, this evalua-tion and others like it, could be used on ac o m p a r a t ive basis with research per-formed in vitro, perhaps using humancorneal equivalents. Development ofmethods for evaluating corneal woundhealing is benefiting from evaluations ofocular repair mechanisms performedboth in vivo and in vitro. Although someof the experiments were not conductedwith the express purpose of developingan alternative to animal testing, themethodologies used support such a goal.
For example, Burgalassi and co-workers(2000) evaluated the effect of xyloglucan(tamarind seed polysaccharide (TSP)) onc o n j u n c t ival cell adhesion to laminin and on corneal epithelium wound heal-ing. Cultured human conjunctival cellswere labeled by addition of a tritiatedamino acid mixture. Their adhesion tolaminin-coated culture wells in the ab-sence or presence of TSP was checked byradioactivity count. TSP was also testedin vivo in animals with corneal damage.Compared to hyaluronate, TSP slightlybut significantly increased the wo u n dhealing rate in vivo. TSP (1%) also exert-ed a positive influence on cell adhesionto laminin, up to a certain laminin con-centration. These researchers concludedthat the ability of the polysaccharide to
promote corneal wound healing mightdepend on its influence on the integrin-substrate recognition system.
In another study of corneal healing,Lambiase et al. (2000) studied the effectsof nerve growth factor (NGF) on cornealrepair in human and rat corneal epithelialcells in culture and human corneal organculture. They showed that NGF is a con-stitutive molecule present and producedin normal human and rat corneas and thatin vitro human and rat corneal epithelialcells, produce, store, and release N G F, and also express high-affinity NGF receptors. In human organ culture,epithelium, ke r a t i n o cytes, and endo-thelium were shown to bind exogenousradiolabeled NGF, and epithelial cellbinding was increased after epithelium
Fig. 2: Stem cells located in limbal epithelium can be rapidly induced to enter thep roliferative population. Long term labeling with BrdU to detect slow cycling stem cells (LRCs; red stained nuclei)followed by a single pulse of 3H-TdR to detect rapidly cycling TA cells (arrows) demonstratesthat under resting conditions (A, B) all slow-cycling cells are pre f e rentially located in thelimbus, while most TA cells are located in the peripheral corneal epithelium (A, C). Anoccasional TA cell can also be observed among the limbal epithelial stem cells (arro w, B).Twenty-four hours following n-heptanol-induced central corneal wound (D, E, F), a singlepulse of 3H-TdR was administered to mice that had populations of LRCs (red stained nuclei).Many of the LRCs were now double-labeled (arrowheads, E) indicating that they hadincorporated 3H-TdR and thus were undergoing a round of DNA synthesis. In addition, therewas an increase in TA cells in the peripheral corneal epithelium (F, arrows) suggestive thatthis population also was induced to proliferate in response to wounding (from Lehrer et al.,1 9 9 8 ) .

HU G G I N S
ALTEX 20, Suppl.1/03 17
i n j u r y. The authors concluded that,“NGF plays an important role in cornealphysiopathology”.
In a study of the kinetics of cornealwound healing, Lehrer and co-workers(1998) studied the replication of cornealepithelial stem cells, and their progeny,transit amplifying (TA) cells. Using double labeling techniques, they showedthat the stem cells can be induced to enter DNA synthesis by wounding. Theyfound that TA cells of the peripheralcornea undergo at least two rounds ofD NA synthesis whereas those of the central cornea are capable of only oneround of division in response to wound-ing (Fig. 2). Moreover, cell cycle time oftransit amplifying cells can be shortenedand number of times these cells replicateis increased in response to wounding.These results could contribute to evalua-tion of endpoints derived from in vitromodels, such as the corneal equivalentdiscussed above.
Development of in vitro assays thatwill evaluate damage to the corneal nervemay benefit from recent work in assaydevelopment in the more general field ofin vitro neurotoxicity. For example, theacute neurotoxic effects of trimethyltin(TMT) have been quantified using neu-ronal networks cultured on microelec-trode arrays (Gramowski et al., 2000).Spontaneously active monolayer net-works in vitro were cultured on thin filmmicroelectrode arrays. Two diff e r e n ttypes of mouse CNS tissues exhibited“characteristic and dose-dependentchanges of their electrophysiological a c t ivity patterns after treatment withTMT”. Moreove r, rat cortical neuron cultures have been used to differentiatethe activities of structurally dive r s echemicals (e.g. 2,5-hexanedione, acryl-amide, organophosphates) (Schmuck et al., 2000). Effects on cytoskeletal ele-ments and on the energy state of the cellswere used as endpoints as well as cyto-toxicity.
Neurological endpoints have beenmeasured previously using corneal tissuecultured in vitro. Mikulec and co-wo r ke r s(1995) used in vitro rabbit cornea prepa-rations for both electrophy s i o l o g i c a lrecording and wound healing measure-ments after treatment with the cornealanalgesic, diltiazem, a calcium channel
blocker. An earlier study by Tanelian and MacIver (1990) used rabbit cornealtissue isolated and maintained in vitro tofacilitate staining, visualization, andelectrophysiologic recording of cornealnerves (Fig. 3).
This work inve s t i gated the effects produced by two methy l pyridinium fluorescent dyes on electrophysiologicresponses from corneal A-delta and Cfiber afferents. Nerve fibers were selec-tively stained by the dyes and could befollowed from their point of entry insmall nerve bundles at the cornea-scleraborder to individual free nerve endingterminals in the corneal epithelium.Hence, investigative methods, both oldand new, can perhaps be employed in thesearch for a better mechanistic under-standing of corneal nerve injury.
2.3 Summary, conclusions, and future work
Current regulatory initiatives favor andemphasize the need to incorporate alter-natives to eye corrosion/irritation testingin animals into the testing of new products for safety. Although much re-search has been done to date on the
development of viable in vitro assays ofocular corrosion and irritancy, acceptablevalidation of these assays has not beena c h i eved. Several reasons have been postulated for the failure of validation efforts, the most prominent of which arethe following. The in vivo test used forcomparison, the Draize test, is based onsubjective scoring of tissue lesions in theeye, providing variable estimates of eyeirritancy; the non-animal method proto-cols were inadequate; the choice of testsubstances was not well-planned; and thestatistical approaches used were not appropriate (Balls et al., 1999).
Several suggestions have been made as to how to remedy these difficulties.Because no single in vitro assay of ocularcorrosion/irritation can be said to encom-pass fully the in vivo response, use ofcomplementary assays in batteries mayprove to be a more suitable approach.R e - evaluation of results from several validation efforts seems to support this suggestion as do results from inde-pendent studies. Application of more discriminating methods of statisticalanalyses have also contributed to a muchbetter understanding of the relationships
Fig. 3: Diagram of the isolated cornea preparation showing placement of stimulatingprobe (STIM) on free nerve endings (FNE) terminals, micropipette used to apply dye samples (SAMP) and extracellular recording electrode (EC) placed on a nervebundle (NB). The sclera was sutured to a plexiglas ring forming a pre s s u re tight seal. Artificial aqueoushumor solution (AQH) was continuously perfused (1.0 ml/min) and temperature (TEMP; 35°C) ad pre s s u re (PRESS; 18mm Hg) monitored (from Tanelian and MacIver, 1990).

HU G G I N S
ALTEX 20, Suppl.1/0318
between/among various in vitro assayswhen used in combination. Use of com-plementary in vitro assays is also sup-ported by the current emphasis on t i e r-testing strategies that incorporatemore than one level/type of data in decision-making structures.
Finally, many voices (past and present)have recommended that we define themechanisms behind the production ofeye corrosion/irritation. Studies of thereference standards used, extent ofcorneal damage, early biomarke r s ,corneal wound healing, kinetics ofcorneal insult and repair and cornealnerve injury have all been suggested asmajor areas of research to be pursuedvigorously in the next decade.
For the present, prompt action in delin-eating which assays of ocular corro-sion/irritation to use in complementaryfashion will aid the swift incorporationof those assays into the testing of newproducts through tier-testing schemes.Re-evaluation of past validation effortscoupled with newer initiatives aimed atdefinition of a reference standards ap-proach should yield firmer ground uponwhich to plan future validation studies.These efforts will be needed, particularlyas assays of mechanisms of action be-come more prevalent and in need of validation.
Alternatives to eye corrosion/irritationtesting in animals are the oldest alterna-tives in existence and they are perhapsthe assays from which we have learnedthe most. Current re-evaluation efforts aswell as those focused on development ofnewer, mechanistically-oriented modelscan only serve to contribute more to ourunderstanding of eye corrosion/irritationand to the ultimate replacement of ani-mals in this type of toxicity testing.
ReferencesBagley, D. M. et al. (1997). A summary
report of the COLIPA internationalvalidation study on alternatives to theDraize rabbit eye irritation test. Toxi-col. In Vitro 11, 141-179.
Balls, M., Botham, P. A., Bruner, L. H.and Spielmann, H. (1995). The EC/HOinternational validation study on alter-natives to the Draize eye irritation test.Toxicol. In Vitro 9, 871-929.
Balls, M., Berg, N., Bruner, L. et al.(1999). Eye irritation testing: The wayforward. The report and recommenda-tions of ECVAM workshop 34. ATLA27, 53-77.
Borenfreund, E. and Borrero, O. (1984).In vitro cytotoxicity assays: potential al-t e r n a t ives to the Draize ocular irritancytest. Cell Biol. Toxicol. 1, 55-65.
Borenfreund, E. and Puerner, J. A .(1985). Toxicity determined in vitro bymorphological alterations and neutralred absorption. Toxicol. Lett. 24, 119-124.
Bruner, L. H., de Silva, O., Earl, L. K. etal. (1998). Report on the Colipa work-shop on mechanisms of eye irritation.ATLA 26, 811-820.
Burgalassi, S., Raimondi, L., Pirisino, R.et al. (2000). Effect of xyloglucan(tamarind seed polysaccharide) onconjunctival cell adhesion to lamininand on corneal epithelium wound healing. Eur. J. Ophthalmol. 10(1), 71-76.
Curren, R. D., Sina, J. F., Feder, P. et al.(1997). IRAG Working Group 5: Other assays. Interagency RegulatoryAlternatives Group. Food Chem. Toxi-col. 35(1), 127-158.
Gautheron, P., Dukic, M., Alix, D. andSina, J. F. (1992). Bovine cornealopacity and permeability test: an in vitro assay of ocular irritancy. Fundam. Appl. Toxicol. 18(3), 442-449.
G r a m owski, A., Schiffmann, D. andGross, G. W. (2000). Quantification ofacute neurotoxic effects of trimethyltinusing neuronal networks cultured onmicroelectrode arrays. Neurotoxicolo-gy 21(3), 331-342.
Griffith, M., Osborne, R., Munger, R. etal. (1999). Functional human cornealequivalents constructed from cell lines.Science 286 (5447), 2051-2053.
Jester, J. V., Maurer, J. K., Petroll, W. M.et al. (1996). Application of in vivoconfocal microscopy to the under-standing of surfactant-induced ocularirritation. Toxicol. Pathol. 24, 412-428.
Lambiase, A., Manni, L., Bonini, S. et al.(2000). Nerve growth factor promotescorneal healing: structural, biochemi-cal, and molecular analyses of rat andhuman corneas. Invest. Ophthalmol.Vis. Sci. 41(5), 1063-1069.
Lehrer, M. S., Sun, T. T. and Lavker, R.M. (1998). Strategies of epithelial repair: modulation of stem cell andtransit amplifying cell proliferation. J.Cell Sci. 111(Pt 19), 2867-2875.
Lewis, R. W., McCall, J. C. and Botham,P. A. (1994). Use of an in vitro test battery as a prescreen in the assess-ment of ocular irritancy. Toxicol. InVitro 8, 75-81.
Lopke, N. P. (1986). HET (Hen’s EggTest) in toxicological research. In R.Marks and G. Plewig (eds.), Skin Models. Models to Study Function andDisease of Skin (282-291). Wien, H e i d e l b e rg, New York: Springer-Verlag.
Maurer, J. K., Li, H. F., Petroll, W. M. etal. (1997). Confocal microscopic char-acterization of initial corneal changesof surfactant-induced eye irritation inthe rabbit. Toxicol. Appl. Pharmacol.143, 291-300.
Maurer, J. K., Parker, R. D. and Carr, G.J. (1998). Ocular irritation: pathologi-cal changes occurring in the rat withsurfactants of unknown irritancy. Toxi-col. Pathol. 26, 226-233.
Mikulec, A. A., Lukatch, H. S., Monroe,F. A. and MacIver, M. B. (1995). Dilti-azem spares corneal A delta mechanoand C fiber cold receptors and pre-s e r ves epithelial wound healing.Cornea 14(5), 490-496.
OECD (Organization for Economic Cooperation and Deve l o p m e n t )(1987). Guideline for Testing ofChemicals No. 405: Acute Eye Irrita-tion/Corrosion.
OECD (2000). Draft Revised Guidelinefor Testing of Chemicals No. 405:Acute Eye Irritation/Corrosion.
Pham, X. T. and Huff, J. W. (1999). Cy-totoxicity evaluation of multipurposecontact lens solutions using an in vitrotest battery. Clao J. 25, 28-35.
Planck, S. R. (1999). Cytokines incorneal injury responses. Crisp DataBase National Institutes of Health(CRISP/99/EY11921-01A1).
Rosenkranz, H. S. and Cunningham, A.R. (2000). A battery of cell toxicity as-says as predictors of eye irritation: Afeasibility study. ATLA 28(4), 603-607.
Schmuck, G., Ahr, H. J. and Schluter, G.(2000). Rat cortical neuron cultures:an in vitro model for differentiating

HU G G I N S
ALTEX 20, Suppl.1/03 19
mechanisms of chemically inducedneurotoxicity. In Vitro Mol. Toxicol.13(1), 37-50.
Sozotono, C., He, J., Matsumoto, Y. et al.(1997). Cytokine expression in the alkali-burned cornea. Curr. Eye Res.16(7), 670-676.
Spielmann, H., Kalweit, S., Liebsch, M.et al. (1993). Validation study of alter-natives to the Draize eye irritation testin Germany: cytotoxicity testing andH E T-CAM test with 136 industrial
chemicals. Toxicol. In Vitro 7, 505-510.
Spielmann, H., Liebsch, M., Kalweit, S.et al. (1996). Results of a validationstudy in Germany on two in vitro alternatives to the Draize eye irritationtest, the HET-CAM test and the 3T3NRU cytotoxicity test. ATLA 24 , 741-858.
Tanelian, D. L. and MacIve r, M. B.(1990). Simultaneous visualizationand electrophysiology of corneal A-
delta and C fiber afferents. J. Neurosci.Methods 32(3), 213-222.
U S E PA (United States Env i r o n m e n t a lProtection A g e n cy) (1998). Health E ffects Test Guidelines. OPPTS870.2400. Acute Eye Irritation. August.
Worth, A. P. and Fentem, J. H. (1999). Ageneral approach for evaluating stepwise testing strategies. ATLA 27,161-177.
3 Alternatives to skin corrosion/irritation testing in animals
3.1 BackgroundThe Draize method for evaluating skinirritation and/or corrosion has been inexistence for a long time and has beenused widely to predict the skin irrita-tion/corrosion potential of many chemi-cal compounds (Draize et al., 1944).Rabbits are the species most often usedin the assay and the number of animalsused in each study can vary from 4 to 12.In many instances, only one dose level istested. A d d i t i o n a l l y, irritancy is often assessed as a part of other studies (forexample, dermal LD5 0 studies) in whichthe same animals are used for all assess-ments. The chemical to be tested may beapplied to the abraded or intact skin ofthe animals’ backs that have been shave dor clipped. The site of application is leftunoccluded or may be occluded withvarious materials such as gauze patchesand elastic tape. Reactions are usuallyread according to a scale devised byDraize at intervals ranging from 1 hourto 7 or 14 days.
The relevance of animal irritancy data(most of which has been generated bythe Draize assay) to humans is question-able. Moreove r, the use of this assay toevaluate corrosivity or irritation hasbeen denounced in recent years for ethical reasons. Numerous alternativemethods have been developed to replacethe use of animals for evaluating skincorrosion and irritation. These includethose based on structure-activity rela-
tionships and human patch testing aswell as in vitro m e t h o d o l o g i e s .
3.2 In vitro modelsThe in vitro methodologies which serveas alternatives to skin corrosion/irritationtesting in animals range from a “test-tube” assay to excised skin to systems involving culture of skin cells on variousm e s h - l i ke frameworks. A d d i t i o n a l l y, the endpoints measured vary from simpleassays of dye retention/release to evalua-tions of more complex variables such astranscutaneous electrical resistance.
The “test tube” assay, also known asCorrositex®, is an in vitro method basedon the ability of a corrosive chemical orchemical mixture to pass through, by diffusion and/or destruction, a biobarrierand to elicit a color change in the under-lying liquid Chemical Detection System(CDS) (ICCVAM, 1999). The biobarrieris composed of a hydrated collagen matrix in a supporting filter membrane,while the CDS is composed of water andpH indicator dyes. Test chemicals andchemical mixtures, including solids andliquids, are applied directly to the biobar-rier. The time it takes for a test chemicalor chemical mixture to penetrate the bio-barrier and produce a color change in theCDS is compared to a classification chartto determine corrosivity/noncorrosivityand to identify the appropriate US De-partment of Transportation (DOT) pack-ing group. Despite criticism of the as-
s a y ’s ability to discriminate between c o r r o s ive and non-corrosive industrialchemicals (for example, Stobbe et al.,1999), the US DOT currently accepts theuse of Corrositex® to assign subcate-gories of corrosivity (packing groups) forlabeling purposes according to UnitedNations (UN) Committee of Experts onthe Transport of Dangerous Goodsguidelines. However, the US DOT limitsthe use of Corrositex® to specific chemi-cal classes, including acids, acid deriva-t ives, acyl halides, alkylamines andp o l y a l kylamines, bases, chlorosilanes,metal halides, and oxyhalides. It has alsobeen suggested that Corrositex® may beused as part of a tiered-testing strategywhere positive responses require no fur-ther testing and negative responses mustbe followed by dermal irritation testing(Scala et al., 1999).
Another method for assessing corro-s ivity is the transcutaneous electrical resistance (TER) assay. Human or ratskin is excised and changes in transcuta-neous electrical resistance across thes t ratum corneum are evaluated after exposure to a test chemical. This assayhas proven to be a good predictor of corrosivity (Botham et al., 1992; Whittleand Basketter, 1993, 1994).
Although, Corrositex® and the TER as-say are definitely valuable tools withwhich to measure the corrosive potentialof many chemical compounds, dermalcorrosion and irritancy have also been

HU G G I N S
ALTEX 20, Suppl.1/0320
studied in vitro by using various skin ir-ritancy models.
Van de Sandt et al. (1997) classified inv i t ro skin irritancy models into fourtypes: immortalized ke r a t i n o cyte celllines, conventional keratinocyte cultures,skin explant or organ cultures, and air-exposed human ke r a t i n o cyte cultures(epidermal or skin equivalents). Theycharacterized their strengths and weaknesses as follows. Immortalizedke r a t i n o cyte cell lines (most notably HaCaT cells) closely resemble normalkeratinocytes in their growth and differ-entiation characteristics and respond, invitro, to modulators of differentiation.They exhibit a remarkable stable geneticbalance over extended culture periods,making them a favorite model of normalhuman keratinocytes. They do not pos-sess the differentiation capacity to forman in vitro skin equivalent in organotypiccultures, how eve r. Normal human ke r a t i n o cyte (NHK) cultures offer distinct advantages including flexibilitywith respect to experimental time, repro-d u c i b i l i t y, and relative ease of cryo-p r e s e r vation. Their use as submerg e dmonolayer cultures is limited to testingof water-soluble test substances, how-ever, and they lack a stratum corneum resulting in increased sensitivity tochemically-induced toxicity. Skin ex-plant and organ cultures closely matchthe in vivo situation when full thicknessskin is used. The explant is placed on agrid or insert and incubated at the air-liquid interface to prevent further growth.The resulting brief survival time of theculture limits testing, however, to shortexposures. A i r- exposed reconstructedcultures represent a fourth in vitro skinirritancy model, many versions of whichare currently undergoing further develop-ment and scrutiny. Essentially, differenti-ated keratinocyte cultures are grown atthe air-liquid interface on various sub-strates, such as inert filters (Rosdy andClauss, 1990), collagen sheets (Tinois etal., 1991), de-epidermised dermis (Reg-nier et al., 1990), and fibroblast-populat-ed collagen gels (Bell et al., 1991). Themain characteristics of native skin tissue,including basal cell layer, stratum spin-osum and granulosum, as well as stratumcorneum are present in these models. Thepresence of an uninterrupted s t ra t u m
corneum in several in vitro models per-mits the application of water-insolublecompounds and final topical formula-tions. However, in some models the bar-rier function is impaired, some modelsare not available any longer, costs may be high, and results may be variable between batches.
A number of commercial models thatincorporate characteristics of the in v i t ro skin irritancy models discusseda b ove have been introduced and studiedby various researchers (for ex a m p l e ,L iving Skin Equivalent™ (Orga n o g e n e-sis), Skin™ (Advanced Tissue Sciences),EpiDerm™ (MatTek), EpiSkin™ ( L’Oreal) and PrediSkin™ (BioPredic))(Fig. 4).
Examples of research inve s t i ga t i n gskin models and corrosivity include evaluations by Lake et al. (1994), whostudied the potential corrosivity of 22compounds and 17 final product formu-lations using EpiDerm™. Ninety-sevenpercent of the compounds evaluated wereassigned the proper US DOT corrosive/non-corrosive designation. Additionally,Perkins and Osborne (1994) studied skincorrosion using several commerciallyavailable skin models. In their work, human skin equivalent cultures, ModelsZK1300™ and ZK1301™ (Adva n c e dTissue Sciences) and EpiDerm™ werecompared using MTT as the endpoint.C o r r o s ive materials were accurately distinguished from strong, moderate, andmild test materials by all three models.
M o r e ove r, numerous comparativestudies of the predictive abilities of vari-
ous models for dermal irritancy havealso been conducted (for example, Hel-man et al., 1992; Roguet et al., 1994;Wolf et al., 1995; Hayden, 1996). Mostrecently, Roguet and colleagues (1999)made an in-depth effort to evaluate several current models. They found thatEpiSkin™, EpiDerm™ a SkinEthic™model and various “in-house” (L’Oreal)models reproduced many of the charac-teristics of human epidermis. In particu-l a r, histologic examination showed acompletely stratified stratum corneum inall models. Inter-batch variations werelow for EpiDerm™ and moderate forEpiSkin™, but considerable variations(thickness of the epidermis, presence ofpycnotic cells) were noted in the SkinEthic™ model. Metabolic studiesshowed the presence of NADPH quinonereductase and glutathione-S-transferaseactivities in all commercial models. Invitro assessments of skin irritancy wereconducted using cytotoxicity (MTT), release of IL-1α, and cytoplasmic enzymes, as endpoints. After SDS treatment, inter-batch variability of MTTresults was lower for EpiDerm™, followed by EpiSkin™, the Cosmital™model and finally the SkinEthic™ model. Results of IL-1α, lactate de-hydrogenase and glutamate-oxaloacetatetransferase release showed a relativelyhigh variability intra-batch or inter-batch.
Other research efforts have investigat-ed the response of reconstructed skinmodels when used with high concentra-tions of test substance and/or with differ-
Fig. 4: EpiDerm™ skin model. (Reprinted with permission from MatTekCorporation).

HU G G I N S
ALTEX 20, Suppl.1/03 21
ent product types. Earl and colleagues(1996) questioned the relevance of in v i t ro cell cultures for measuring the cytotoxicity of high concentration testsubstances, such as surfactants or surfac-tant mixtures. They compared data gen-erated in vivo with that obtained from theagarose overlay cytotoxicity assay, andan MTT time-course assay using Epi-Derm™. The results indicated that theagarose overlay assay did not distinguishbetween any of the treatments; however,the EpiDerm™ assay broadly reflectedthe results obtained in vivo and did dis-tinguish between different surfa c t a n t sand their mixtures in vitro.
Koschier et al. (1997) compared theability of three-dimensional human skinmodels to evaluate the dermal irritancyof petroleum products. Three commer-cially supplied human skin constructs(Living Skin Equivalent™ (LSE), Epi-Derm™, and ZK1300™) were treatedwith 14 petroleum refinery streams. End-points measured were lactate dehydroge-nase, IL-1α, PGE2, and MTT conversion.Spearman rank order analysis comparingthe in vitro cytotoxicity data with the Primary Dermal Irritation Index (PDII)scores gave values of 0.54 (LSE), 0.41(ZK1300™), and 0.79 (EpiDerm™), respectively. These data indicated thatIL-1α concentrations showed reasonablecorrelations with the known in vivoirritation level, especially in the Epi-Derm™ cultures. The best prediction ofin vivo irritation in all three models appeared to come from a combination ofcytotoxicity and IL-1α m e a s u r e m e n t s(Fig. 5).
Perkins and co-workers (1999) also in-vestigated the use of human skin equiva-lent cultures with different product class-es. They directly compared in vitro to invivo human skin responses using historicor concurrent skin response data for di-verse products and ingredients including surfactants, cosmetics, anti-perspirants (AP) and deodorants (DO).EpiDerm™ was used and human clinicalprotocols were paralleled by topical dosing of neat or dilute test substances tothe stratum corneum surface of the skincultures. MTT conversion, lactate de-hydrogenase and aspartate amino-transferase release and IL-1α expressionwere monitored. For surfactants, dose-
Fig. 5: A comparison of the time course of cytotoxicity (°) estimated by relative MTTreduction, and IL-1 release (•) after direct application of material F-137 to thesurface of the three different tissue constructs: (A) LSE, (B) skin2 model ZK1300™,and (C) EpiDerm™ (from Koschier et al., 1997).

HU G G I N S
ALTEX 20, Suppl.1/0322
response curves of MTT cell viability data clearly distinguished strongly irritat-ing from milder surfactants and rank ordered irritancy potential in a mannersimilar to in vivo patch test results (Tab.4).
For AP/DO products, all the in vitroendpoints correlated well with consumerreported irritation, with IL-1α showingthe greatest capacity to distinguish irri-tancy over a broad range. IL-1α alsoshowed the best prediction of humanskin scores from 14-day cumulative irri-tancy tests of cosmetic products (Fig. 6).
The authors felt that these results con-firmed the value of cornified human skincultures as an in vitro preclinical screenfor prediction of human skin irritation responses, and identified the need to customize in vitro endpoints for irritancyprediction for different product classes.
3.3 Validation and regulatoryactivities
Several human keratinocyte and humanskin models have been evaluated by theEuropean Center for the Validation of Alternative Methods (ECVAM) for theirpredictive value (Van de Sandt et al.,1997). Furthermore, four alternativemodels were investigated as potentialcandidates for corrosion testing by ECVAM in an international validationstudy. The rat transcutaneous electricalresistance (TER) assay, Corrositex®,
S k i n2 ZK1350™ test, and EpiSkin™were all included in this effort (Fentem etal., 1998).
Two of the four tests evaluated in thevalidation study, the TER assay andEpiSkin™, met the criteria concerningacceptable underprediction and overpre-diction rates. Hence, they are consideredscientifically validated for use as replace-ments for the animal test for distinguish-ing between corrosive and non-corrosivechemicals for all of the chemical typesstudied. EpiSkin™ was the only test ableto distinguish between known R35 (UNpacking group I) and R34 (UN packinggroups II & III) chemicals, for all of thechemical types included, on an accept-able number of occasions.
The corrosive potentials of about 40%of the test chemicals could not be assessed with Corrositex®, and the assaydid not meet all of the criteria for it to be considered acceptable as a replace-ment test. How eve r, the group did decide that Corrositex® may be valid for testing specific classes of chemicals,such as organic bases and inorga n i cacids, a decision similar to that reachedby the US DOT as discussed above .Subsequent to this large validation e ffort, a prevalidation exercise has also been conducted with EpiDerm™r e - vealing excellent prediction of corro-s ivity for a wide spectrum of chemicals(Liebsch, 1999).
The Organization for Economic Coop-eration and Development (OECD) hasrecently developed a tiered-testing strate-gy for evaluating dermal corrosion andirritation that allows for the use of vali-dated and accepted in vitro m e t h o d s(OECD, 1998). Worth et al. (1998) reportan evaluation of the proposed OECDtesting strategy as it relates to the classi-fication of skin corrosives. By using a set of 60 chemicals, an assessment wasmade of the effect of applying three stepsin the strategy. The results indicate thatchemicals can be classified as corrosive(C) or non-corrosive (NC) with sufficientreliability by the sequential applicationof three alternative methods, i.e., struc-ture-activity relationships (where avail-able), pH measurements, and a single invitro method (either the rat skin transcu-taneous electrical resistance (TER) assayor the EpiSkin™ assay). They concludedthat the proposed OECD strategy for skincorrosion can be simplified without com-promising its predictivity (for example, itdoes not appear necessary to measureacid/alkali reserve (buffering capacity) inaddition to pH for the classification ofpure chemicals).
Fo l l owing this evaluation of theOECD’s tier-testing strategy for corro-sives, ECVAM recognized the need tomake substantial progress in validatingin vitro tests for skin irritation (Botham,1999). A Task Force was established
Tab. 4: Rank-order of surfactant irritancy by in vivo and in vitro tests (from Perkins et al., 1999)
Surfactant In vivo dose range Surfactant active In vivo ranka In vitro rankb
(µg/cm2) (% w/w) human skin patch skin cultures
(Irritancy Groups-A, B, Cc)
1 Sodium lauryl sulphate 22-430 0.01-0.40 1/A 1/A
2 Anionic ethoxylate (a) 22-430 0.02-0.40 2/B 3/B
3 Anionic ethoxylate (b) 22-430 0.02-0.40 3/B 4/B
4 Anionic ethoxylate (c) 22-430 0.02-0.40 4/B 5/B
5 Nonionic (a) 690-11,000 0.60-10.0 5/C 6/C
6 Amphoteric betaine 207-3300 0.20-3.00 6/C 7/C
7 Nonionic (b) 690 -11,000 0.60-10.0 7/C 2/A
Note. Lowercase (a), (b), (c) differentiate surfactants within a class.aIn vivo rank ordering is based on the cumulative irritation score, where 1 is the most irritating and 7 is the least irritating surfactant.bIn vitro ranks are based on the dose of surfactant causing 50% cytotoxicity (MTT50 values) in human skin cultures, where 1 is the most irritating or cytotoxic, and 7 is the least irritating or cytotoxic surfactant.cSurfactants within the same irritancy group (indicated by letters A, B or C) are not significantly different from each other, but aresignificantly different (p<0.05) from surfactants in other irritancy groups.

HU G G I N S
ALTEX 20, Suppl.1/03 23
which issued a challenge to develop-ers/users of relevant methods to submitdata, obtained according to a specifiedprotocol with a preliminary predictionmodel, on ten defined test chemicals.This resulted in selection of four tests –EpiSkin™, EpiDerm™, PrediSkin™,and the non-perfused pig ear model – forprevalidation testing. The main objectiveof these validation activities is to identifythose in vitro tests capable of discrimi-nating skin irritants (I) from non-irritants(NI), as defined according to EU riskphrases (R38; no label) and the harmo-
nized OECD criteria (Irritant; no label)(Fentem et al., 1999). Once these testshave completed a prevalidation assess-ment, a subsequent validation study, ifsuccessful, would then allow the com-plete OECD testing strategy for skin corrosion and irritation to be followedwithout the use of live animals.
3.4 Summary, conclusions, and future work
Alternative methodologies for the testingof dermally corrosive or irritating sub-stances include a “test-tube” assay, meas-
urements of transcutaneous electrical resistance in excised skin, and in vitroskin culture models of several types.Structure-activity-relationship modelingas well as pH determinations and acid/alkali reserve evaluations may also con-tribute to the successful determination ofdermal corrosivity or irritancy of a testcompound, particularly if used in tier-testing strategies. Of the in vitro skin irritancy systems available, skin modelkits appear to be the most popular candi-dates, perhaps due to the ease with whichthey can be used. Although the variabili-
Fig. 6: Antiperspirant/deodorant study 1. The percent of human subjects reporting either subjective or objective skin irritation for 11 products are plotted vs. each in vitro endpoint.The in vitro endpoints evaluated include cell viability (MTT assay; a), enzyme release (LDH; b) (AST; c), and extracellular IL-1α (d). Symbolsand error bars represent the mean ± SD across four replicate cultures. To compare the in vitro endpoints to the human skin irritation %response, a simple linear regression analysis was used. (r2, 0.75-0.94) (from Perkins et al., 1999).

HU G G I N S
ALTEX 20, Suppl.1/0324
ty of their response is still under investi-gation, it appears to be due in manycases to factors such as test compoundconcentration and product type.
Regulatory acceptance of in vitro mod-els of skin corrosion or irritation appearsto be no longer as substantive a hurdle asit once was as evidenced by recent USD OT and OECD initiatives. Models currently validated for use as alternativesto corrosivity testing in animals includeCorrositex® (limited to specific classes of chemicals), the rat TER assay, andEpiSkin™. EpiDerm™ has also beenprevalidated as an alternative method to corrosion testing. Validation of in vitromethodologies for skin irritation appearsto be the major “next step” in develop-ment of alternatives acceptable to regula-tors. Models of dermal irritancy current-ly undergoing this process includeEpiSkin™, EpiDerm™, Prediskin™,and the nonperfused pig ear model (Elliott et al., 1999) with results expectedby the end of 2000.
The use of alternatives to dermal c o r r o s ivity or irritancy testing in animalswill advance greatly the aims of the a l t e r n a t ives to animal testing move m e n tfor several reasons. The first, and per-haps, foremost advantage to use of thesemethods is the abolishment of testingc o r r o s ive and/or irritating substances onl ive animals. The humane implicationsof this action are straightforward and itclearly represents appropriate imple-mentation of a replacement alternative. S e c o n d l y, the number of animals usedwill be reduced by use of these assays(for example, Corrositex® uses no animal cells at all, a small section of ratskin is used in the TER assay, and mostin vitro skin irritancy models incorporatehuman skin). Finally, from a scientificp e r s p e c t ive, a test which has been questioned not only with regard to its reliance on subjective observations, bu talso as to its more general relevance tohuman skin corrosion and irritation, willh ave been replaced, in many instances,with in vitro assays using more objec-t ively evaluated endpoints. The chal-lenge remains how eve r, to relate the endpoints measured in these assays tothe development of dermal corrosionand/or irritation by more than superfic i a lex p l a n a t i o n s .
The next five years of effort in this areaof alternatives to animal testing shouldresult in widespread acceptance and integration of tier-testing strategies forcorrosivity testing. Additionally, valida-tion and regulatory acceptance of in vitroassays of dermal irritancy should also be accomplished. The next ten years of research should result in refined in vitroskin irritancy models exhibiting very little variability in response and in muchgreater use in testing laboratories. More-over, the relationship between the end-points measured in these assays and production of dermal irritancy should bebetter understood. The next 20 years ofresearch should yield models that will ef-fectively discriminate between differenttypes and levels of dermal insult perhapslending insight into the mechanisms lying beneath the subsequent pathologi-cal response. Progenitors of this type of“specialized” model include Melano-derm™ and Epi-201™ (both from Mat-Tek). Melanoderm™ is a system intowhich melanocytes have been incorpo-rated that may be useful for studyingmelanogenesis, and skin pigmentation(Klausner et al., 1999). Epi201™ is as t r a t i fied culture of proliferating and d i fferentiating human ke r a t i n o cy t e swhich, when treated with epidermalgrowth factor “develops a granular layerwhich is thicker and more heavily popu-lated with keratohyalin granules com-pared to standard EpiDerm™.” Studiesof re-epithelialisation, wound healingand normal barrier development arethought to benefit from use of modelssuch as this one (Klausner et al., 1998).
ReferencesBell, E., Parenteau, N., Gay, R. et al.
(1991). The living skin equivalent: itsmanufacture, its organotypic proper-ties and its response to irritants. Toxic.In Vitro 5, 519-596.
Botham, P., Hall, T., Dennet, R. et al.(1992). The skin corrosivity test in vitro. Results of an inter-laboratory trial. Toxic. In Vitro 6, 191-194.
Botham, P. (1999). Skin corrosion and irritation: Toward complete replace-ment. ATLA 27, 108.
Draize, J., Woodward, G. and Calvery, H.(1944). Methods for the study of irrita-
tion and toxicity of substances applied topicaly to the skin and mucous mem-branes. J. Pharm. and Exper. Therap.82, 377-390.
Earl, L., Hall-Manning, T., Holland, G. etal. (1996). Skin irritation potential ofs u r factant mixtures: using releva n tdoses in in vitro systems. ATLA 24,249. Abstract #73.
Elliott, G., Pool, C. and Bruijnzeel, P.(1999). Isolated pig ears: An in vitromodel for screening for irritant chemi-cals using stimulation of transepider-mal water loss as the test parameter.ATLA 27, 110.
Fentem, J., Archer, G., Balls, M. et al.(1998). The ECVAM international val-idation study on in vitro tests for skin corrosivity. 2. Results and evaluationby the management team. Toxic. in Vitro 12, 483-524.
Fentem, J., Botham, P., Harbell, J. et al.(1999). Prevalidation of in vitro testsfor acute skin irritation. ATLA 27, 346.
Hayden, P. (1996). Cytokine and lipidmediator release from in vitro skinmodels following detergent treatmentor physical wounding. J. Invest. Der-matol. 106(4), 936. Abstract #782.
Helman, M., Biesaga, S. and Koslo, R.(1992). Reformed human skin: Poten-tial screening tool for dermal irritationtesting. The Toxicologist 12(1), 297.Abstract #1147.
I C C VAM (Interagency CoordinatingCommittee for the Validation of Alter-native Methods) (1999). Corrositex®:An in vitro test method for assessingdermal corrosivity potential of chemi-cals. NIH Publication No. 99-4495.June.
Klausner, M., Neal, P. and Kubilus, J.(1998). Initial results with Epi-201, adeveloping in vitro model of humanepidermis. J. Invest. Dermatol. 110(4),558. Abstract. #513.
Klausner, M., Neal, P., Breyfogle, B. andKubilus, J. (1999). Melanoderm: anepidermal tissue model containingfunctional melanocytes. ATLA 27, 299.
Koschier, F., Roth, R., Wallace, K. et al.(1997). A comparison of three-dimen-sional human skin models to evaluatethe dermal irritation of selected petro-leum products. In Vitro Toxicol. 10(4),391-405.

HU G G I N S
ALTEX 20, Suppl.1/03 25
Lake, L., Swanson, J., Kubilus, J. et al.(1994). Use of an in vitro skin modelto predict chemical corrosivity. TheToxicologist 14(1), Abstract #340.
Liebsch, M., Traue, D., Barrabas, C. et a l .(1999). Prevalidation of the EpiDerm™skin corrosivity test. ATLA 27, 350.
OECD (Organization for Economic Co-operation and Development) (1998).Harmonized Integrated Hazard Classi-fication System for Human Health andE nvironmental Effects of ChemicalSubstances. Paris: OECD.
Perkins, M. and Osborne, R. (1994). Application of an in vitro method forskin corrosion testing to a variety ofcommercially available skin cultures. J. Invest. Dermatol. 102(4), 641. Ab-stract # 707.
Perkins, M. A., Osborne, R., Rana, F. R.et al. (1999). Comparison of in vitroand in vivo human skin responses toconsumer products and ingredientswith a range of irritancy potential. Toxicol. Sci. 48(2), 218-229.
Roguet, R., Regnier, M., Cohen, C. et al.(1994). The use of in vitro reconstitut-ed human skin in dermotoxicity test-ing. Toxic. In Vitro 8(4), 635-639.
Roguet, R., Beck, H., Boelsma, E. et al.(1999). Testing and improvement ofreconstructed skin kits in order to elab-orate European standards: first results.ATLA 27, 333.
Regnier, M., Asselineau, D. and Lenoir,M. (1990). Human epidermis recon-structed on dermal substrates in vitro:an alternative to animals in skin pharmacology. Skin Pharmacology 3,70-85.
Rosdy, M. and Clauss, L. (1990). Com-plete human epidermal cell differentia-tion in chemically defined medium atthe air-liquid interface on inert filtersubstrates. J. Invest. Dermatol. 95,409-414.
Scala, R., Stokes, W., Hill, R. et al.(1999). The results of an independentpeer review panel evaluation of thevalidation status of Corrositex®. ATLA27, 334.
Stobbe, J., Drake, K. and Maier, K.(1999). Comparison of dermal corro-sion values for selected industrialchemicals using Corrositex®. The Toxicologist 48(1S), 77. Abstract #359.
Tinois, E., Tiollier, J., Gaucherand, M. et al. (1991). In vitro and post-trans-
plantation differentiation of human keratinocytes grown on human type IVcollagen film of a bilayered dermal sub-stitute. E x p e r. Cell Res. 193, 310-331.
Van de Sandt, J., Roguet, R., Cohen, C.et al. (1997). The use of human ker-atinocytes and human skin models forpredicting skin irritation. The reportand recommendations of ECVA Mworkshop 38. ATLA 27, 723-743.
Whittle, E. and Basketter, D. (1993). Thein vitro skin corrosivity test. Develop-ment of method using human skin.Toxic. In Vitro 7(3), 265-268.
Whittle, E. and Basketter, D. (1994). Invitro skin corrosivity test using humanskin. Toxic. In Vitro 8(4), 861-863.
Wolf, B., Kwan, S., Moral, J. and Rheins,L. (1995). Development of in vitromethods for evaluating the epidermaland dermal effects of alpha hydroxyand amides. J. Invest. Dermatol.104(4), 639. Abstract #511.
Worth, A. P., Fentem, J., Balls, M. et al.(1998). An evaluation of the proposedOECD testing strategy for skin corro-sion. ATLA 26(5), 709-720.
4 Alternatives to skin sensitization testing in animals
4.1 BackgroundDe Silva et al. (1996) described the re-lationship between skin sensitization and in vitro alternatives as follows: “The induction of contact hypersensitivity in-volves a complex series of cellular andmolecular events. The relative contribu-tions and interdependence of these eventsare not completely understood. It re-mains to be demonstrated whether an invitro test, or several in vitro tests in com-bination which model the critical steps insensitization, can replace animal experi-ments for predicting contact allergic re-actions in humans.” Hence, the intricatecascade of events leading to developmentof skin sensitization, as elegant as it is,represents a substantial hurdle to the development of in vitro assays not unlikethe development of assays for a similarlyintricate biological system, that of repro-
duction. Alternatively, the sensitization“cascade” also provides fertile groundfor exploration of novel assay techniquesthat do not use animals.
Historically, the guinea pig has beenused extensively as the animal model ofchoice for sensitization testing. Testingtechniques vary, but the basic protocolscheme involves initial application of the sensitizer to the skin of the animal,followed by a delayed “challenge” expo-sure to the same sensitizer after a giveninterval of time. Positive reactions arescored according to degree of rednessand/or swelling at the site of application.The test can be quite lengthy taking any-where from 3 to 6 weeks to complete.
Current regulatory guidelines includes everal methods of skin sensitizationtesting. The guinea pig maximization test (GPMT) (Magnusson and Kligman,
1970) and Buehler assay (BA) (Buehler,1965) seem to be the most widely usedtechniques, endorsed by the EU, OECD,and the United States (EU, 1992; OECD,1993; USEPA, 1998). USEPA (UnitedStates Environmental Protection Agency)Guidelines suggest that, “Either sex maybe used in the Buehler test and theGPMT. If females are used, they shouldbe nulliparous and not pregnant. TheBuehler test recommends using a mini-mum of 20 animals in the treatment andat least 10 as controls. At least 10 animals in the treatment group and 5 inthe control group should be used with the GPMT, with the stipulation that if itis not possible to conclude that the testsubstance is a sensitizer after using fewer than 20 test and 10 control guineapigs, the testing of additional animals togive a total of at least 20 test and 10

HU G G I N S
ALTEX 20, Suppl.1/0326
control animals is strongly recommend-e d .” Moreove r, these guidelines statethat, “The dose level will depend on thetest method selected. In the Buehler test,the concentration of the induction doseshould be high enough to cause mild irri-tation, and the challenge dose should usethe highest non-irritating concentration.In the GPMT, the concentration of the in-duction dose should be well toleratedsystemically, and should be high enoughto cause mild-to-moderate skin irritation;the GPMT challenge dose should use thehighest non-irritating concentration.”
These requirements result in the use ofthirty animals per test, at minimum,which is a relatively large number whencompared to the number of animals usedin several of the alternative assays whichare discussed below. Furthermore, thesetechniques can employ an adjuvant thatmay enhance the severity of the responseobserved in test animals. Finally, as withthe Draize eye and skin irritation tests,the grading of responses in either of thesensitization assays discussed above depends upon the subjective, though often well-trained, assessment of a hu-man observer.
4.2 Alternative assaysEach of the alternative assays discussedbelow attempts to capture data represent-ing a particular step or steps in the “cas-cade” of events leading to contact sensi-tization. Because of the many stepsinvolved, numerous opportunities for thecapture of such data exist. How eve r,techniques for “hard-to-catch” events arestill evolving.
Two of the more well-known alternativeassays are the mouse ear-swelling test(MEST) and the local lymph node assay( L L NA). Both assays are used as screen-ing tests for skin sensitization. The LLNAhas also recently been evaluated for use asa “stand-alone” assay of skin sensitization( I C C VA M - N I C E ATM, 1999).
The MEST measures increases in ear-thickness (swelling) in the mousethat result from applying a test chemicalto the animal’s ear after these animalshave been exposed to the test material in an earlier, induction phase of the pro-tocol (Gad et al., 1986). Measurement ofe a r-thickness yields quantitative datagiving the MEST a decided advantage
over the more subjective observationsused in guinea pig tests of sensitization.The MEST has, however, been criticizedfor its lack of sensitivity (Cornacoff etal., 1988; Dunn et al., 1990). Subsequentresearch has indicated that this lack maybe remedied, at least in part, by diets rich in Vitamin A (Thorne et al., 1994;Sailstad et al., 1993).
The local lymph node assay measuresevents occurring during the inductionphase of the sensitization reaction.Specifically, this assay most often meas-ures the uptake of tritiated methyl thymi-dine by proliferating T-cells in draininglymph nodes of mice exposed to a testchemical (Kimber and Basketter, 1992).The LLNA has been studied and com-pared extensively with other assays ofskin sensitization (for example, Kimberet al., 1990, 1991, 1995; Hayes et al.,1998; Hayes and Meade, 1999; Ohta etal., 1999; Howell et al., 2000). Moreover,an evaluation of skin sensitization methods by ECVAM suggested that theassay “offers a number of advantagescompared with guinea pig methods, notleast of which are that the LLNA is relatively rapid and cost-effective. Theendpoint is quantitative and the assaydoes not require the use of an adjuvant”(De Silva et al., 1996). The recent peerreview of the LLNA conducted by the Interagency Coordinating Committee onValidation of Alternative Methods andthe National Interagency Committee forthe Evaluation of Alternative Methods(ICCVAM-NICEATM) came to similarconclusions. They stated that, “the assayperformed at least as well as currentlyaccepted guinea pig methods (GPMT/BA) for the hazard identification ofstrong to moderate chemical sensitizingagents” (ICCVA M - N I C E ATM, 1999).They also agreed with ECVAM that theassay has numerous aspects that make ita favorable alternative assay (for exam-ple, reduces animal distress, potentiallyreduces animal numbers). The LLNA hasdemonstrated several weaknesses common to alternative assays of skinsensitization (for example, failure to detect weak sensitizers, identification ofstrong irritants as sensitizers). Re-searchers in this field feel, however, thatthese irregularities may be ex p l a i n e dwith further in-depth study.
Additional, suggested alternative approaches to the study of skin sensitiza-tion have included study of the quantita-t ive structure-activity relationships ofhaptens, cytokine response induced incultured keratinocytes by haptens, con-tact allergen-induced alterations in adhesion molecules on ke r a t i n o cytes and epidermal Langerhans cells, andhapten- mediated T-cell activation byLangerhans cells (Elmets, 1996). Evalua-tion of various cell cultures systems (e.g. dendritic cells, keratinocytes, skinexplants, and co-cultures of skin cells)that support several of these approacheshas been made as well (De Silva et al.,1996).
One such approach invo l ves inter-leukin 1ß (IL-1ß) production by Langer-hans cells. This event has become linkedto development of an alternative assayfor skin sensitization because it has beenshown to be a primary mediator of the induction phase of contact sensitiv i t y(Enk and Katz, 1992; Enk et al., 1993).D evelopment of this assay has been hindered by the difficulty of obtainingLangerhans cells. How eve r, several studies seem to indicate that cultures ofdendritic cells isolated from peripheralblood can be utilized effectively to studycontact allergens (Romani et al., 1994;Lenz et al., 1993; Hoppe et al., 1995).
Reutter and colleagues (1997) studiedearly upregulation of IL-1ß in dendriticcells (DCs) following exposure to con-tact sensitizers. DCs were cultivated inserum-free medium supplemented withgranulocyte/macrophage-colony stimula-tion factor (GM-CSF) and Interleukin 4(Il-4). The DCs exhibited typical den-dritic morphology, characteristic surfacemarkers and high stimulatory capacityfor T-cells. Five-day-old DCs were incu-bated with contact sensitizers, pentade-cyl-catechol, 2,4,6-trinitrobenzene sul-fonic acid, 2,4-dinitrofluorobenzene,NiSO4, K2Cr2O7 and the irritant, sodiumd o d e cyl sulfate (SDS). IL-1ß mRNA expression was monitored by using a r everse transcriptase-polymerase chainreaction technique and non-radioactivehybridization procedures. For all contactsensitizers, expression of IL-1ß mRNAincreased, whereas treatment with the ir-ritant, SDS had no significant effect onIL-1ß expression (Fig. 7).

HU G G I N S
ALTEX 20, Suppl.1/03 27
In a subsequent study, Pichowski andco-workers (2000) evaluated further thefindings of Reutter using dendritic cellsobtained from human blood. These cellswere cultured in the presence of inter-leukin-4 (IL-4) and granulocyte/macro-phage colony stimulating factor. After 5days in culture, they exhibited a Langer-hans cell-like phenotype. These DC werethen cultured with the contact allergen,2,4-dinitrofluorobenzene (DNFB), SLSor vehicle alone and mRNA expressionfor IL-1ß mRNA (two-to threefold) incells derived from three out of eightdonors whereas IL-6 and IL-18 werelargely unaffected by allergen exposure.Sodium dodecyl sulfate treatment did notinduce IL-1ß mRNA expression in any ofthe donors investigated. Cytokine mRNAexpression was also analyzed using theprotocol described by Reutter. This protocol did not enhance the sensitivityof measurement of induced cytokine ex-pression. These researchers concludedthat, “Although selected upregulation ofIL-1ß by blood derived DC has been con-firmed, a wider range of contact allergensand irritants needs to be assessed beforethis approach could be considered forhazard identification”.
De Silva and colleagues (1996) de-scribe the migration of hapten-modified
Langerhans cells from the epidermis tothe regional lymph nodes where theystimulate hapten-specific T-cells as another measurable endpoint of contactsensitization. They cite several studies inwhich human skin explants have beenused in assays for this purpose with reasonable success (for example, recon-stituted basement membrane (Kobayashi
et al., 1994), reconstituted human epider-mis (Tammi and Maibach, 1987; Messa-di et al., 1988; Le Poole et al., 1991)).
Additionally, the activation and pro-liferation of antigen-specific T-lympho-cytes caused by primary contact of T-cells with antigen complexes ex-pressed on antigen presenting cells(APC) has been investigated as a meas-ure of contact sensitization. Moulon andassociates (1993) examined the capacityof human Langerhans’ cells (LC) to sensitize autologous T-cells to the t r i n i t r o p h e nyl hapten (TNP) in vitro.T h ey found that two-day culturedL a n g e r h a n s ’ cells, but not freshly prepared Langerhans’ cells, can induce in vitro primary proliferative reactions to the TNP hapten (Fig. 8).
The primary in vitro response wa sstrongly inhibited by monoclonal anti-bodies to major histocompatibility com-plex (MHC) class I and II, CD4 antigen,and ICAM-1 and LFA-3 adhesion mole-cules. These researchers concluded that“this in vitro priming assay, using cultured human Langerhans’ cells asAPC, might be useful to analyze the ear-ly steps of T-cell sensitization and subse-quently to develop in vitro predictivetests allowing detection of sensitizingcompounds”.
Additional work by Krasteva et al.(1996) showed that the two-day cultured
Fig. 8: Naive T-cell sensitization to TNP by cultured LC CD45RA+ T lymphocytes (105) were cultured for 5 days with graded numbers of either non-modified or TNP-treated 2-day incubated LC. T-cell proliferation was assessed by a(H3)thymidine pulse for the final 18 hrs of culture (from Moulon et al., 1993).
T cells (CD45RA+) + TNP-cLC
T cells (CD45RA+) + cLC
Fig. 7: IL-1ß mRNA expression induced by SDS, PDC, TNBS, DNFB, nickel sulfateand potassium dichromate. Signal strength was assessed by densitrometric scanning of autoradiograms andintegration by WINCAM 1.0 Software. IL-1ß data were standardized to the respective ß-actin controls. Data were expressed in arbitrary units and are representative of threeexperiments. K = control for probes solubilized in medium; DMSO = control for probessolubilized in DMSO (from Reutter et al., 1997).
600
500
400
300
200
100
0K DMSO SDS PDC TNBS DNFB NISO4 K2Cr2O7

HU G G I N S
ALTEX 20, Suppl.1/0328
human Langerhans’cell model discrimi-nated between two groups of substances:strong contact sensitizers (TNP, fluores-cein isothiocyanate (FITC), Bandrows-k i ’s base (BB) and weak sensitizers(coumarin, citronellal, and hy d r o x y -citronellal) and irritants (SDS). T h eysuggested that the assay could be used asa screening in vitro assay to eliminatestrong contact allergens before furtherpredictive animal tests have to be per-formed.
Rougier et al. (2000) have refined thiswork further by studying Langerhans-like dendritic cells (LLDC) derived fromcord blood progenitors and autologous T-lymphocytes, isolated from the sameblood sample. Treatment of day 12-14LLDC, with strong haptens trinitroben-zene sulfonic acid (TNP), flu o r e s c e i nisothiocyanate (FITC) or Bandrowski’sbase (BB), resulted in the proliferation ofT-lymphocytes, whereas weak allergensand irritants, such as sodium dodecyl sulfate (SDS) were ineffective (Tab. 5).
The use of immature (day 8) LLDCand the addition of a 48 hour stage of incubation after hapten contact, resultedin phenotypic maturation of LLDC in addition to lymphocyte activation in allthe cultures with strong haptens. The 48 hour stage of incubation, results insensitization and in some cases the induction of T-cell proliferation to cit-ronellal (1/8), coumarin (1/8) and to aprohapten p-phenylenediamine (pPDA);2/8). The phenotype of DC after 48 hoursof contact with a strong hapten, becomesthat of a mature DC (CD83+, CD86+ and
HLA-DR++). With fragrance molecules,weak haptens and prohaptens, a compa-rable phenotype is observed only when
T-lymphocytes are activated. These au-thors suggest that the unresponsivenessobserved with weak haptens, may be theconsequence of an incomplete matura-tion of LLDC.
Hence, although further work is un-doubtedly needed, three distinct events inthe “cascade” leading to contact sensiti-zation have been translated into the alter-native assays discussed above: upregula-tion of IL-1ß production by Langerhanscells, migration of hapten-modifi e dLangerhans cells to the lymph node, andthe induction of T-cell proliferation byhapten-modified Langerhans cells.
Cytokines have also been the focus ofefforts directed toward development ofa l t e r n a t ives to contact sensitization. Although early work with keratinocytecultures investigated the role of proin-flammatory key mediators as possible
Tab. 5: Forty-eight hour incubation of eight-day-old Langerhans-like dendritic cells(LLDC) with hapten is necessary to induce a proliferative response of autologous T lymphocytes (from Rougier et al., 2000).
Without incubation 48h incubation
Hapten SI Positive SI Positive experiment experiment
SDS 1.1 ± 0.04 0 (8) 1.7 ± 0.20 0 (8)
TNP 1.9 ± 0.06 0 (5) 4.3 ± 0.68 5 (5)
FITC 1.2/1.8 0 (2) 3.0/3.5 2 (2)
BB 1.5 ± 0.21 0 (8) 5.0 ± 0.56 8 (8)
pPDA 1.1 ± 0.12 0 (8) 2.1 ± 0.57 2 (8)
Coumarin 1.1 ± 0.23 0 (8) 2.1 ± 0.32 1 (8)
Citronellal 1.3 ± 0.06 0 (8) 2.4 ± 0.41 1 (8)
Stimulation index (SI) is expressed as mean index values ± 2 S.E.. Positive experimentsrefer to the number of experiments with an SI above 3; the number of cultures effectedwith different donors is indicated in brackets.
Fig. 9: Influence of increasing doses of Tween 80 (A), Triton X100 (B), BC (C) andDNCB (D) on the IL-1 (•) and IL-8 (°) intracellular level. The tissues were exposed to the different concentrations of the tested products for 20 hrsat 37°C (5% CO 2) at which time IL-1α and IL-8 were extracted and quantified by ELISA.The results of one representative experiment are given and expressed in pg ofcytokines/µg protein measured in the extract (from Coquette et al., 1999).

HU G G I N S
ALTEX 20, Suppl.1/03 29
alternatives to the Draize skin irritationassay (for example, Muller-Decker et al.,1994), recent studies have incorporatedin vitro methods that measure the kinet-ics of cytokine production in response toa l l e rgen exposure. Doubts have been expressed, however, as to the discrimina-tory power of this type of assay becausemany cutaneous chemical allergens arealso primary irritants (De Silva et al.,1996).
A recent example of the combined useof in vitro culture of skin cells and meas-urement of cytokine production aftercontact allergen exposure is that of Coquette and co-workers (1999). Theyevaluated the differential expression andrelease of cytokines by an in vitro recon-structed human epidermis following exposure to skin irritant and sensitizingchemicals. A model of reconstructed
human epidermis (RHE) was used as anin vitro model to discriminate the effectsof Tween 80, Triton X100 and benzal-konium chloride (BC) as irritants and 1-chloro-2,4-dinitrobenzene (DNCB) assensitizing agent. Epidermal irritationand sensitization were evaluated mor-phologically and on the basis of intracel-lular and extracellular levels of inter-leukin-1α and interleukin-8 (IL-8). Thecorresponding constitutive mRNA levelswere quantified and the cytotoxic re-sponse was assessed by MTT assay. Topical application of the three surfac-tants resulted in significant changes oftissue morphology and was coupled withd i fferent dose-dependent decreases incell viability corresponding to their in vivo irritant potency. The IL-1α releaseincreased concomitantly with cell viabil-ity decrease, but surprisingly, the surfac-
tants did not elicit elevated IL-8 levels. Incontrast, DNCB did not induce elevatedIL-1α release although it induced a rapiddose-dependent decrease in cell viability.DNCB did increase IL-8 release. Poly-merase chain reaction analysis demon-strated the presence of mRNA for IL-1αand IL-8; IL-1α was the most abundanttranscript found. BC, Triton X100 andDNCB upregulated IL-8 mRNA expres-sion. These researchers concluded thatthe production of IL-1α and its releaseinto the extracellular medium was as-cribed not only to direct cytotoxicity butalso to the extent of tissue stimulation.Their results demonstrated dive rgent I L - 1α and IL-8 release profiles and corresponding mRNA upregulation in response to irritants and DNCB (Fig. 9and 10, respectively).
Although additional testing is neededto clearly delineate irritant and DNCBresponse, it was suggested that it may bepossible to use a single assay that incor-porates a reconstituted skin model withmeasurements of cytokine expression todistinguish between irritant and sensitiz-ing agents as a function of induced cytokine production patterns and cell viability measurements.
Cytokines have also been proposed asadditional (non-radioactive) endpoints tobe used in conjunction with the locallymph node assay, specifically to detectmoderate sensitizers, such as mercapto-benzothiazole and hexyl cinnamic alde-hyde (Hariya et al., 1999). This assaymeasures corrected IL-2 index (CII) d e fined as including corrections forlymph node weight ratio and ratio ofCD4-positive subset. The authors con-cluded that measurement of CII signifi-cantly increased the sensitivity of the LLNA to include detection of moderatesensitizers.
Dearman and colleagues (1999) havealso investigated cytokine endpoints forthe LLNA: interferon-gamma and inter-leukin 12. Production of these cytokinesby draining lymph nodes was investigat-ed as an alternative “readout” for the LLNA. Animals were exposed to a rangeof skin sensitizers at two application concentrations. Secretion of IFN-gammaand the p40 subunit of IL-12 by draininglymph node cells was measured by cy t o k i n e - s p e c i fic enzyme-linked im-
Fig. 10: Influence of increasing doses of Tween 80 (A), Triton X100 (B), BC (C) andDNCB (D) on the IL-1 (•) and IL-8 (°) mRNA expression in the RHE. The tissues were exposed to the diff e rent concentrations of the tested products for 20 hrs at37°C (5% CO2) at which time specific mRNA were extracted and quantified by RT- P C R .Each value re p resents the mean ± SD of three independent experiments and is expressed asthe diff e rence between the ratio of cytokine mRNA/1B15 mRNA determined in tre a t e dsamples and the same ratio determined in untreated samples (from Coquette et al., 1999).

HU G G I N S
ALTEX 20, Suppl.1/0330
munosorbent assay. Parallel experimentswere run in which lymph node cell activ-ity was assessed by the usual method oftritiated thymidine incorporation in situ .All allergens produced proliferative re-sponses. Treatment with chemical aller-gen in each case caused a marked increase in IFN-gamma secretion, withparticularly high production followingexposure to oxazolone or hexyl cinnamica l d e hyde. In contrast, application ofchemical allergens was not generally associated with elevated IL-12 p40 secre-tion. However, cytokine secretion did notcorrelate closely with induction of lymphnode cell proliferation. These researchersconcluded that expression of cytokinesIFN-gamma or IL-12 by allergen-activat-ed lymph node cells does not provide areliable or sensitive endpoint for the LLNA compared to tritiated thymidineincorporation in situ.
Several investigations have been per-formed in order to define the cytokinesignals that are needed by Langerhanscells in order to migrate to reg i o n a llymph nodes after exposure to a contacta l l e rgen. Cumberbatch et al. (1997)found that both tumor necrosis factor-al-pha and interleukin-1ß are needed byLangerhans cells for such migration.They suggest that the changes induced inLC by TNF-α and IL-1ß may include thealtered expression of adhesion moleculesand acquisition of the ability to interactwith and pass through the basementmembrane. Subsequent work has sug-gested that dexamethasone, a syntheticglucocorticoid and immunosuppressant,r egulates the de nov o synthesis and/or release of TNF-α that inhibits LC migration to regional lymph nodes(Cumberbath et al., 1999).
Hence, cytokines continue to be re-searched extensively as integral compo-nents of the contact sensitization re-sponse. T h ey may perhaps be usedeffectively as measurable endpoints in alternative assays pending further devel-opment of information about their role inthe process of sensitization.
4.3 Validation and regulatory activities
Incorporation of alternative assays intoassessments of contact sensitization hasbeen addressed by several regulatory ini-
tiatives. Both the EU and OECD describes t r u c t u r e - a c t ivity relationship deve l o p-ment, the MEST and the LLNA as ac-ceptable screening procedures in assess-ments of contact sensitization (EU, 1992;OECD, 1993). If the screening test givesa positive result, the substance is regard-ed as a skin sensitizer labeled with theEU risk phrase R43 indicating “maycause sensitization by skin contact”.Negative results obtained in screeningtests have to be confirmed by using oneof the in vivo guinea pig tests describedabove.
Furthermore, US regulatory agenciesacting through the Interagency Coordi-nating Committee for Alternative Meth-ods (ICCVAM) and National InteragencyCommittee for the Evaluation of Alterna-tive Methods (NICEATM) have evaluat-ed the LLNA as a “stand-alone” assay ofcontact sensitization. The rev i ew “in-volved the evaluation of data on 209chemicals, of which both LLNA andguinea pig data were available for 126chemicals and both LLNA and human(HMT and HPTA) data were providedfor 74 chemicals. An in-depth review ofall the chemicals that have been definedin the published literature as human allergens was not conducted for this eval-uation. From the analysis generated during the review process, the accuracyof the LLNA vs. GPMT/BA was 89%(N=97), LLNA vs. all guinea pig tests(GPT) was 86% (N=126), the LLNAvs. human data was 72% (N=74),GPMT/BA vs. human was 72% (N=57),and all guinea pig tests (GPT) vs. humanwas 73% (N=62). In terms of accuracy,sensitivity, specificity, and positive andnegative predictivity, the PRP found theperformance of the LLNA to be similarto that of the GPMT/BA. Equally impor-tant, the performance of the LLNA andthe GPMT/BA was similar when eachwas compared to human data (HMT/HP-TA)” (ICCVAM-NICEATM, 1999).
M o r e ove r, the ICCVA M - N I C E AT Mpeer review panel (PRP) unanimouslyrecommended the LLNA as a stand-alone alternative for contact sensitizationhazard assessment, provided that someprotocol modifications were made (forexample, lymphocyte proliferation datashould be collected at the level of the individual animal). The PRP also con-
cluded unanimously, “that the LLNA is ad e finite improvement with respect to animal welfare (i.e., refinement and re-duction) over the currently acceptedGPMT”.
In an additional independent assess-ment of the LLNA in which the I C C VA M - N I C E ATM report was re-viewed as well as publications publishedsince the report and reports from otheragencies, the ECVAM Science AdvisoryCommittee also unanimously endorsedthe LLNA. They stated that, “the LLNAis a scientifically validated test which canbe used to assess the skin sensitizationpotential of chemicals. The LLNAshould be the preferred method, as it us-es fewer animals and causes less pain anddistress than the conventional guinea-pigmethods. In some instances, and for sci-entific reasons, the conventional methodscan be used” (ECVAM, 2000).
Hence, the current regulatory postureembraces alternative methods for evalua-tion of contact sensitization, either asscreening procedures or as “stand-alone”assays.
4.4 Summary, conclusions, and future work
Alternative assays exist for the assess-ment of contact sensitization. Several aremore well-validated than others, but alladdress measurable endpoints represen-tative of contact sensitization. The LLNAand MEST are both considered appropri-ate as screening procedures. Moreover,the LLNA has also been endorsed as a“stand-alone” procedure for assay of sensitization. Additional assays are alsounder development, all of which measurea particular event or series of events leading to sensitization. Examples in-clude interleukin 1ß (IL-1ß) productionby Langerhans cells, migration of hap-ten-modified Langerhans cells from theepidermis to regional lymph nodes, andthe activation and proliferation of anti-g e n - s p e c i fic T- l y m p h o cytes caused byprimary contact of T-cells with antigencomplexes expressed on antigen present-ing cells (APC). Cytokine profiles are also under scrutiny as possible markersof sensitization response.
Progress toward the development andrefinement of additional alternative as-says of contact sensitization is strongly

HU G G I N S
ALTEX 20, Suppl.1/03 31
dependent upon breakthroughs in our un-derstanding of the immune processesmediating the response. In fact, onemight say that they have and will contin-ue to proceed “hand-in-hand”. The ex-tensive efforts directed toward validationof the local lymph node assay have bornethe much-needed fruit of a “stand-alone”assay that incorporates elements of bothrefinement and reduction. This type ofeffort is needed to produce additional,validated assays of contact sensitization,particularly replacement assays. Thesee fforts should be encouraged by the regulatory initiatives already in place.However, much more basic research re-mains to be done before a fully validatedreplacement assay of contact sensitiza-tion finds regulatory support. One of themost pressing issues at present seems tobe the ability of any potential replace-ment assay to adequately discriminatebetween processes leading to dermal irritation versus those resulting in contactsensitization. One area of promising re-search directed toward this end is that inwhich the cytokine “profiles” resultingfrom exposure to irritants or contact sen-sitizers are being defined. That cytokine“profiles” parallel differences in type oftoxic response (dermal irritation vs. con-tact sensitization) has been demonstratedin several of the research studies dis-cussed above. Additional recent work byDearman and Kimber (1999) demon-strates that differences in cytokine ex-pression are also apparent in dermal ver-sus respiratory sensitization responses.Hence, particular cytokine “profi l e s ”may well represent measurable endpointsunique to the contact sensitization re-sponse that can be integrated effectivelyinto a replacement assay.
ReferencesBuehler, E. V. (1965). Delayed contact
hy p e r s e n s i t ivity in the guinea pig.Arch. Dermatol. 91, 171-177.
Coquette, A., Berna, N., Vandenbosch, A.et al. (1999). Differential expressionand release of cytokines by an in vitroreconstructed human epidermis fol-lowing exposure to skin irritant andsensitizing chemicals. Toxicol. In Vitro13(6), 867-877.
Cornacoff, J. B., House, R. V. and Dean,
J. H. (1988). Comparison of a ra-dioisotopic incorporation method andthe mouse ear swelling test (MEST)for contact sensitivity to weak sensitiz-ers. Fund. Appl. Toxicol. 10, 40-44.
Cumberbatch, M., Dearman, R. J. andKimber, I. (1997). Langerhans cells require signals from both tumournecrosis factor-alpha and interleukin-1beta for migration. Immunol. 92(3),388-395.
Cumberbatch, M., Dearman, R. J. andK i m b e r, I. (1999). Inhibition of d examethasone of Langerhans cell migration: influence of epidermal cytokine signals. Immunopharmacol.41(3), 235-243.
Dearman, R. J., Hilton, J., Basketter, D.A. and Kimber, I. (1999). Cytokineendpoints for the local lymph node assay: consideration of interferon-gamma and interleukin 12. J. Appl.Toxicol. 19(3), 149-155.
Dearman, R. J. and Kimber, I. (1999).Cytokine fingerprinting: characteriza-tion of chemical allergens. To x i c o l .Methods 19(1), 56-63.
De Silva, O. et al. (1996). Alternativesmethods for skin sensitisation testing.The report and recommendations ofECVAM workshop 19. ATLA 24(5),683-705.
Dunn, B. J., Rusch, G. M., Siglin, J. C.and Blaszcak, D. L. (1990). Variabilityof a mouse ear swelling test (MEST) inpredicting weak and moderate contactsensitization. Fund. Appl. Toxicol. 15 ,242-248.
ECVAM (European Center for the Vali-dation of Alternative Methods) (2000).Statement on the scientific validity oflocal lymph node assay for skin sensi-tisation testing.
Elmets, C. A. (1996). Progress in the development of an in vitro assay forcontact allergens: Results of the AvonProgram Project Experience. In VitroToxicol. 9 (2), 223-235.
Enk, A. H. and Katz, S. I. (1992). Earlymolecular events in the inductionphase of contact sensitivity. Proc. Nat.Acad. Sci. (USA) 89, 1398-1402.
Enk, A. H., Angeloni, V. L., Udey, M. C.and Katz, S. I. (1993). An essential rolefor Langerhans cell-derived IL-1ß in theintiation of primary immune responsesin skin. J. Immun. 150, 3698-2704.
EU (European Union) (1992). Annex toCommission Directive 92/69/EEC of31 July 1992 adapting to technicalprogress for the seventeenth timeCouncil Directive 67/548/EEC on theapproximation of laws, reg u l a t i o n s ,and administrative provisions relatingto the classification, packaging, and labelling of dangerous substances. Official Journal of the European Com-munities L383A, 131-136.
Gad, S. C., Dunn, B. J., Dobbs, D. W etal. (1986). Development and validationof an alternative dermal sensitizationtest: the mouse ear swelling test. Toxicol. Appl. Pharmacol. 84, 93-114.
Hariya, T., Hatao, M. and Ichikawa, H.(1999). Development of a non-radioac-tive endpoint in a modified local lymphnode assay. Food Chem. Toxicol. 37(1),87-93.
Hayes, B. B., Gerber, P. C., Griffey, S. S.and Meade, B. J. (1998). Contact hy-persensitivity to dicyclohexylcarbodi-imide and diisopropylcarbodiimide infemale B6C3F1 mice. Drug Chem.Toxicol. 21(2), 195-206.
Hayes, B. B. and Meade, B. J. (1999).Contact sensitivity to selected acrylate compounds in B6C3F1 mice: relativepotency, cross reactivity, and compari-son of test methods. Drug Chem. Toxi-col. 22 (3), 491-506.
Hoppe, U., Degwert, J. and Steckel, F.(1995). Use of CD1a - dendritic cellsand keratinocytes to characterize cellu-lar reactions involved in allergic con-tact dermatitis. J. Cell. Biochem. 21(Suppl. A), 11.
Howell, M. D., Manetz, T. Z. and Meade,B. J. (2000). Comparison of murine assays for the identification of chemi-cal sensitizers. To x i c o l ogy Methods10(1), 1-15.
I C C VAM. (Interagency CoordinatingCommittee for the Validation of Alter-n a t ive Methods) and NICEATM. (National Interagency Committee forthe Evaluation of Alternative Methods)(1999). The Murine Local Ly m p hNode Assay. A Test Method for As-sessing the Allergic Contact DermatitisPotential of Chemicals/Compounds.F e b r u a r y. NIH Publication No. 99-4494.
Kimber, I., Hilton, J. and Botham, P. A.(1990). Identification of contact aller-

HU G G I N S
ALTEX 20, Suppl.1/0332
gens using the murine local lymphnode assay. Comparisons with theBuehler occluded patch test in guineapigs. J. Appl. Toxicol. 10, 173-180.
Kimber, I., Hilton, J., Botham, P. A. et al.(1991). The murine local lymph nodeassay. Results of an inter-laboratorytrial. Toxicol. Lett. 55, 203-213.
Kimber, I. and Basketter, D. A. (1992).The murine local lymph node assay: Acommentary on collaborative studiesand new directions. Fd. Chem. Toxicol.30, 165-169.
Kimber, I, Hilton, J., Dearman, R. J. et al.(1995). An international evaluation ofthe murine local lymph node assay andcomparison of modified procedures.Toxicology 103, 63-73.
Kobayashi, Y., Staquet, M. J., Dezutter-D a m buyant, C. and Schmitt, D. (1994).Development of motility of Langer-hans cells through extracellular matrixby in vitro hapten contact. European J.of Immunol. 24, 2254-2257.
K r a s t eva, M., Peg u e t - N avarro, J.,Moulon, C. et al. (1996). In vitro primary sensitization of hapten-specif-ic T cells by cultured human epidermalLangerhans cells – a screening predic-tive assay for contact sensitizers. Clin.Exp. Allergy 26(5), 563-570.
Lenz, A., Heine, M., Schuler, G. and Romani, N. (1993). Human andmurine dendritic cells. Isolation bymeans of a novel method and pheno-typical and functional characteriza-tions. J. Clin. Invest. 95, 258-2596.
Le Poole, I. C., Das, P. K., Krieg, S. R. etal. (1991). Organotypic culture of hu-
man skin for studying wound healing.Wounds 3, 102-110.
Magnusson, B. and Kligman, A. M.(1970). Allergic Contact Dermatitis inthe Guinea Pig. Springfield, Illinois:Charles C. Thomas.
Messadi, D. V., Pober, J. S. and Murphy,G. F. (1988). Effects of recombinantgamma-interferon on HLA-DR andDQ in short term skin organ culture.Lab. Invest. 58, 61-67.
Moulon, C., Peg u e t - N avarro, J.,Courtellemont, P. et al. (1993). In vitroprimary sensitization and restimula-tion of hapten-specific T cells by freshand cultured human epidermal Langer-hans’cells. Immunol. 80(3), 373-379.
Muller-Decker, K., Furstenberger, G. andMarks, F. (1994). Keratinocyte-derivedp r o i n flammatory key mediators andcell viability as in vitro parameters ofirritancy: a possible alternative to theDraize skin irritation test. Toxicol. Appl. Pharmacol. 127(1), 99-108.
Ohta, K., Yamanaka, S. and Takaesu, Y.(1999). The evaluation of mercury sensitization by mouse ear swellingtest (MEST). Shikwa Gakuho 99 (10),821-832.
OECD (Organization for Economic Co-operation and Development) (1993).OECD Guideline for Testing of Chem-icals, No. 406. Skin Sensitisation, 9 pp. Paris, OECD.
Pichowski, J. S., Cumberbath, M., Dear-man, R. J. et al. (2000). Investigationof induced changes in interleukin 1beta mRNA expression by culturedhuman dendritic cells as an in vitro ap-
proach to skin sensitization testing.Toxicol. Vitr. 14(4), 351-360.
Reutter, K., Jaeger, D., Degwert, J. andHoppe, U. (1997). In vitro model forcontact sensitization: II. Induction ofIL-1beta mRNA in human blood-derived dendritic cells by contact sen-sitizers. Toxicol. In Vitro 11(5), 619-626.
Romani, N., Gruner, S., Brang, D. et al.(1994). Proliferating dendritic cell progenitors in human blood. J. Exp.Medicine 180, 83-93.
Rougier, N., Redziniak, G., Mougin, D.et al. (2000). In vitro evaluation of thesensitization potential of weak contact allergens using Langerhans-like den-dritic cells and autologous T cells. Tox-icol. 145(1), 73-82.
Sailstad, D. M., Tepper, J. S., Doerfler,D. L. and Selgrade, M. K. (1993).Evaluation of several variations of themouse ear swelling test (MEST) fordetection of weak and moderate con-tact sensitizers. Toxicol. Methods. 3,169-182.
Tammi, R. and Maibach, H. I. (1987).Skin organ culture: why? Int. J. Dermatol. 26, 150-160.
Thorne, P. S., Hawk, C., Kaliszewski, S.D. and Guiney, P. D. (1994). The non-invasive mouse ear swelling assay. I.Refinements for detecting weak sensi-tizers. Fund. Appl. Toxicol. 17, 790-806.
U S E PA (United States Env i r o n m e n t a lProtection Agency) (1998). Health Ef-fects Test Guidelines. OPPTS870.2600. Skin Sensitization. August.
5 Alternatives to developmental/reproductive toxicity testing in animals
5.1 BackgroundS everal years ago, Gray and Kimber(1989) reported on the value and limita-tions of several in vitro techniques for reproductive and developmental toxicity.The researchers represented in the reportemphasized the overall difficulty associ-ated with developing replacement assaysfor very complex reproductive and devel-opmental events in the whole organism.
Other researchers since that time haveechoed these sentiments (for example,Mirkles, 1996). How eve r, increased emphasis on the use of in vitro testing byregulators and industry alike, has spurredattempts to validate assays of reproduc-tive and/or developmental effects that donot use animals.
Any in vitro assay that could replacecurrent reproductive testing procedures
using animals would substantially reducethe number of animals used for biomed-ical testing. For example, current testingguidelines from the United States Envi-ronmental Protection Agency (USEPA)for reproductive/developmental toxicityscreening tests as well as prenatal devel-opmental toxicity and reproduction andfertility effects studies recommend usingten or twenty animals (at minimum) per

HU G G I N S
ALTEX 20, Suppl.1/03 33
sex per dose level. Three dose levels arerequired as well as a control group inmost cases (USEPA, 1998a, 1998b,2000a, 2000b). Hence, 80 to 160 animalsper assay are required at minimum.
Clearly, alternative assays that do notuse animals are needed. Four promisingcandidates, the micromass (MM) assay,the whole embryo culture (WEC) assay,the embryonic stem cell test (EST), andthe frog embryo teratogenesis assay –Xenopus (FETAX) assay, are discussedin-depth below.
5.2 In vitro alternatives5.2.1 Micromass (MM) assayThe MM assay may use cultures of limbbud cells or neuronal cells or both. Faust-man (1994) described these cultures asf o l l ows. “Limb or cephalic tissues (usually mesencephalic regions) are iso-lated from early to mid-organogenesisembryos and single cell suspensions are prepared using a combination of mechanical dissociation and enzymatic digestion. Cells are plated at high densi-ty and undergo differentiation into chon-
drocytes or neurons without additionalstimulation.”
In one of the first inve s t i gations of amammalian MM system, Flint (1983) de-scribed a micromass culture method forrat embryonic neural cells noting that“the formation of neurons, the binding ofneural cell projections (axons) into nerve -l i ke structures, the synthesis of neuronalcell antigen and (3H)GABA uptake stud-ies confirm that the micromass culturesystem may be a very good model of i nv i v o neuronal diff e r e n t i a t i o n ” .
Tab. 6: Concentrations of nonteratogens and teratogens inhibiting differentiation in CNS and LB cultures by 50% of the control value (IC50) and maximum concentration having no inhibitory effect (NEC) (from Flint and Orton, 1984)
Without S9 With S9
CNS LB CNS LB
Compound IC50 NEC IC50 NEC IC50 NEC IC50 NEC
Nonteratogens
Dimenhydrinate 7.0a 1.0 180.0 100.0 60.0 10.0 -b -
Glutehimide 300.0 200.0 160.0 100.0 - - - -
Teratogens
Aldrin 25.4 1.0 14.6 1.0 43.0 10.0 57.0 1.0
Aspirin 190.0 100.0 190.0 100.0 - - - -
8-Azaguanine 0.8 <0.1 0.8 <0.1 300.0 100.0 280.0 100.0
Azathioprene 162.0 100.0 9.0 <0.1 230.0 100.0 - -
Cadmium chloride 2.5 1.0 3.0 1.0 27.0 10.0 55.0 10.0
Captan 0.4 <0.1 3.8 1.0 20.0 10.0 40.0 10.0
Chloramphenicol 230.0 100.0 160.0 60.0 - - - -
Colchicine <0.1 <0.1 <0.1 <0.1 0.3 0.1 0.3 0.1
Cycloheximide <0.1 <0.1 <0.1 <0.1 0.4 0.1 0.9 0.1
Cyclophosphamide 325.0 100.0 - - <0.1 <0.1 <0.1 <0.1
Cytosine arabinoside <0.1 <0.1 <0.1 <0.1 0.8 0.1 3.0 0.1
Diazepam 150.0 100.0 150.0 100.0 - - - -
Diethylstilbestrol 2.8 1.0 1.8 0.1 - - - -
Diphenylhydantoin 64.5 25.0 78.5 50.0 250.0 100.0 - -
L-Dopa 40.0 10.0 40.0 10.0 - - 0.4 <0.1
Ethionine - - - - - - <0.1 <0.1
EDTA 2.8 1.0 - - - - - -
5-Fluorouracil <0.1 <0.1 <0.1 <0.1 0.3 <0.1 0.2 <0.1
Hydrocortisone 200.0 40.0 100.0 30.0 - - - -
Indomethacin 45.0 10.0 50.0 10.0 240.0 100.0 - -
Mebendazole <0.1 <0.1 <0.1 <0.1 4.5 1.0 1.6 0.1
Parbendazole <0.1 <0.1 0.9 <0.1 - - - -
Sulfisoxazole 160.0 100.0 210.0 10.0 - - - -
Theophylline - - 140.0 100.0 - - - -
ICI 124,206 <0.1 <0.1 <0.1 <0.1 3.5 1.0 3.5 1.0
Note: The following nonteratogens did not inhibit differentiation at any of the concentrations used: aminopyrine ascorbic acid, caffeine, I-carnosine, cyclamate, inosine, kanamycin sulfate, maltose, 6-methyluracil, nitrilotriacetic acid, quinine, saccarin, sulfanilamide,sulfisomidine, tolbutamide, ICI 89406 and ICI 123,737. Two teratogens: 2,4-Dichlorophenoxyacetic acid and thalidomide, similarly did not inhibit differentiation.aµg/ml culture mediumbNo inhibitory effect to 500 µg/ml or maximum soluble concentration (if less).

HU G G I N S
ALTEX 20, Suppl.1/0334
Shortly thereafter, Flint and Orton(1984) published results from a study inwhich 46 compounds (27 teratogens/19nonteratogens) were investigated by invitro assay using cultures of rat embryomidbrain and limb bud cells (Tab. 6).
A high specificity (89% prediction ofnonteratogens) and sensitivity (93% prediction of teratogens) were noted inthis study verifying a high predictabilityfor this MM assay.
Subsequently, Renault and colleagues(1989) investigated the micromass limbbud cell culture as a model for studyingteratogenic mechanisms. They monitoredchanges in cell differentiation and cellproliferation after treatment with terato-gens, retinoic acid and 6-aminonicoti-namide, and the non-teratogen, doxyl-amine. Retinoic acid inhibited celld i fferentiation whereas 6-aminonicoti-namide inhibited cell proliferation.Doxylamine produced a non-specific cytotoxic effect. The assay was subse-quently refined to a routine in vitro teratogenesis screening test in whichcells were cultivated in microtiter platesand chondrogenesis and cell proliferationwere simultaneously assessed after sixdays of culture. The test was validatedwith 51 reference compounds, 33 teratogens and 18 non-teratogens. Noneof the non-teratogens inhibited chondro-genesis or cell proliferation. Further ul-trastructural characterization of normaland abnormal chondrogenesis in micro-mass rat embryo limb bud cell cultureswas also performed by these researchers(Renault et al., 1995).
Characterization of neuronal micro-mass cultures has been performed byW h i t t a ker et al. (1993). T h ey concludedthat cells taken from gestational day-12embryos and cultured for 5 days in vitrocomprise at least two morphologicallydistinct cell types: fi b r o b l a s t - l i ke cellsand neurons. Antibodies to GQ ga n g l i o-side, g-aminobutyric acid (GABA), m i c r o t u bule-associated protein 2 (MAP2),MAP5, neuron-specific enolase (NSE),neural cell adhesion molecules (N-CAM), and tau yielded preferentialstaining. Hence, these wo r kers conclud-ed that the pattern of neuronal diff e r e n-tiation in CNS micromass cultures exhibits certain similarities to that ob-s e r ved in vivo. In addition, they felt that
certain markers identified in the studywere of potential utility as biomarke r sof chemically-induced deve l o p m e n t a ln e u r o t o x i c i t y.
Since the development of the MM a s s a y, it has been used widely to inve s-t i gate the developmental toxicity of numerous chemicals. Examples include,acrylamide, methylene bisacrylamideand haloperidol (Walum and Flint,1993); chlorpyrifos (Cosenza andBidanset, 1995); paraquat (Cosenza, etal., 1996); glufosinate ammonium( Watanabe and Iwase, 1996); acetylsali-cylic acid, isoniazid, penicillin G, sac-
charine, vincristine sulphate, 6-amino-nicotinamide, retinoic acid, and ama-ranth (Amacher et al., 1996);m e t hylmercury (McMillan, et al., 1997);and cadmium (Haberstroh and Kapron,1998). Recent evaluations of the MMsystem by an ECVAM expert rev i ew( B r own et al., 1995) indicated that thesecultures “represent robust test systemsfor studying potential teratogens”. T h eycited the fact that there is probably moreexperience with this system than witha ny other alternative method for repro-d u c t ive toxicology studies and that itslimitations are well understood.
Tab. 7: Toxicity of phenol, chlorophenols and selected derivates in the adult Hydra(from Mayura et al., 1991)
Chemical Adult hydra toxicity Exposure timeb
whole-log MACa (µg/ml)
Phenol 1000 20Anisole 1000 44Phenyl acetate 100 20Phenol. Na salt 1000 284-Chlorophenol 100 43-Chlorophenol 100 202-Chlorophenol 100 924-Chlorophenyl acetate 100 44-Chlorophenol, Na salt 100 42,3-Dichlorophenol 100 42,4-Dichlorophenol 10 922,5-Dichlorophenol 10 922,6-Dichlorophenol 100 203,4-Dichlorophenol 10 443,5-Dichlorophenol 10 203,5-Dichlorophenyl acetate 10 203,5-Dichlorophenol, Na salt 10 202,3,4-Trichlorophenol 1 442,3,5-Trichlorophenol 1 202,3,6-Trichlorophenol 10 682,4,5-Trichlorophenol 1 282,4,6-Trichlorophenol 10 283,4,5-Trichlorophenol 1 442,3,5-Trichlorophenyl acetate 1 282,3,5-Trichlorophenol, Na salt 1 282,3,4,5-Tetrachlorophenol 0.1 442,3,4,6-Tetrachlorophenol 1 442,3,5,6-Tetrachlorophenol 1 442,3,4,5-Tetrachloroanisole 10 922,3,5,6-Tetrachloroanisole 10 682,3,4,5-Tetrachlorophenyl acetate 0.1 442,3,4,5-Tetrachlorophenol, Na salt 0.1 44Pentachlorphenol 0.1 44Pentochloranisole 10 92Pentachlorphenyl acetate 0.1 44Pentachlorphenol, Na salt 0.1 44Tetrachloro-1,2-benzoquinone 1 92Tetrachloro-1,4-benzoquinone 10 20
a MAC (Minimum effective concentration)bTime (hr) required to reach toxic endpoint

HU G G I N S
ALTEX 20, Suppl.1/03 35
5.2.2 Whole embryo culture (WEC) assay
Mammalian embryo cultures are derivedfrom mice, rats, or rabbits as summarizedby Brown et al. (1995). Head-fold or ear-ly somite stage embryos are dissectedfree from maternal tissue, parietal yolksac and Reichert’s membrane, leavingthe visceral yolk sac and ectoplacentalcone intact. The conceptus is cultured inmedium under defined gassing condi-tions for 24-48 hours. Media containing ahigh proportion of serum is usually usedand test compounds are added to the cultures for defined periods of time.Metabolic activation systems may alsobe incorporated. A number of endpoints
can be measured such as effects on deve l-opment of the visceral yolk sac va s c u l a r i-sation and circulation; effects on haemato-poiesis, embryonic growth and diff e r e n -tiation; and dysmorphogenic eff e c t s .
Several studies have investigated theWEC assay in relationship to other as-says of developmental toxicity. Bechterand colleagues (1991) investigated theability of rat WEC to differentiate between retinoids with differing terato-genic potency. They compared the resultsobtained to those from in vivo studies ofmice and micromass cultures of rat embryonic limb bud cells and concludedthat the WEC “was an excellent tool for the preliminary testing of retinoids”.
The developmental toxicity of chlori-nated phenols was evaluated by Mayuraand co-workers (1991) utilizing Hydraattenuata and rat embryos in culture. TheWEC findings closely paralleled thosefound using Hydra embryos (Tab. 7, 8,and 9) and the authors suggested that acombination of Hydra and WEC assayswould facilitate rapid detection and ranking of hazardous chemicals associat-ed with complex mixtures of chemicalwastes.
Gregotti et al. (1992) evaluated the effects of styrene oxide in a limb bud micromass assay and a WEC system.Styrene oxide induced a dose-dependentincrease in embryolethality and embryo
Tab. 8: Comparison of phenol and chlorinated phenols at equimolar concentration (0.6mM) on growth and development of day 10 rat embryos in cultures (from Mayura et al., 1991)
Treatment Number of Yolk-sac Crown-rump Number of Morphological embryos exposed diameter* (mm) length* (mm) somites* (AV) score
Control 26 7.4 ± 0.13a 4.6 ± 0.09a 32.4 ± 0.33a 49.3 ± 0.52a
Solvent control (0.5% DMSO) 21 7.4 ± 0.13a 4.2 ± 0.12a 32.5 ± 0.36a 48.3 ± 0.49a
Phenol 10 7.0 ± 0.20a 4.6 ± 0.21a 31.5 ± 0.70a 48.9 ± 0.18a
4-Chlorophenol 12 7.3 ± 0.11a 3.9 ± 0.17b 29.5 ± 0.65b 45.6 ± 0.93b
3,5-Dichlorophenol 14 5.4 ± 0.12b 2.4 ± 0.09 20.0 ± 0.65c 17.8 ± 0.75c
2,3,5-Trichlorophenol 13 4.8 ± 0.43b 2.9 ± 0.14c 19.6 ± 0.60c 24.8 ± 1.12c
2,3,4,5-Tetrachlorophenol 7 3.7 ± 0.90c 2.3 ± 0.06d very few 5.0 ± 2.00 f
Pentachlorophenol 14 5.3 ± 0.36b 3.1 ± 0.10c 20.9 ± 0.46c 21.5 ± 0.27d
Note: Means with the same letter are not significantly different (p ≤ 0.05)*Values are means ± standard error
Tab. 9: Comparison of phenol and chlorinated phenols at equimolar concentration (0.6mM) on growth and development of day 10 rat embryos in cultures (from Mayura et al., 1991)
Treatment Number of embryos Protein∆ µg/embryo DNA∆ µg/embryo Number exposed malformed
Control 26 298.31 ± 17.2a 65.2 ± 3.2a 0
Solvent control (0.5% DMSO) 21 287.55 ± 23.9a 62.0 ± 3.5a 0
Phenol 10 294.00 ± 18.5a 67.2 ± 4.1a 0
4-Chlorophenol 12 195.00 ± 18.7b 47.6 ± 3.8b 0
3,5-Dichlorophenol 14 72.14 ± 7.0c,d 20.6 ± 1.7c,d 100%
2,3,5-Trichlorophenol 13 83.50 ± 6.9c 21.8 ± 1.5c,d *2,3,4,5-Tetrachlorophenol 7 24.50 ± 3.6d 14.7 ± 2.5d *Pentachlorophenol 14 127.50 ± 11.1c 26.5 ± 1.1c *Note: Means with the same letter are not significantly different (p ≤ 0.05)∆ Values are means ± standard error* Inhibition of growth and development

HU G G I N S
ALTEX 20, Suppl.1/0336
malformation at concentrations that werecomparable to effective concentrations in the micromass culture system. Theseresearchers felt that these findings indicated a “quantitative comparabilitybetween the two in vitro systems ana-lyzed”.
The WEC assay has also been used e ff e c t ively to differentiate the stereo-selective teratogenicity of enantiomers of the valproic acid analogue, 2-n-p r o pyl-4-pentynoic acid (4-yn-VPA ) .Andrews et al. (1995) studied the effectsof the R(+) and S(-) enantiomers of 4-yn-VPA on cultures of mouse embryo.The teratogenic effects of the S(-) enan-tiomer were clearly visible in this assay,whereas, the R(+) enantiomer producedno embryotoxic nor dysmorphogenic effects at any concentration (Tab. 10 and11).
This study represented the first illus-tration of the stereoselectivity of a devel-opmental toxicant in an in vitro test system, mirroring results obtained in anin vivo assay.
ECVAM has indicated that this in vitrosystem is a valuable tool considering itsextensive use in many laboratories and
its ability to detect dysmorphogenesis inembryos.
Moreover, they noted that it enablesnot only the detection of adverse devel-opmental effects from chemical ex -posure, but has also been shown to discriminate between the potencies ofstructurally related compounds. T h eycaution, however, that the system is lim-ited in that it is complex, covers only apart of organogenesis (a feature sharedby other in vitro assays of developmen-tal/reproductive effects), and requires afairly high level of technical sophistica-tion to perform (Brown et al., 1995).
5.2.3 Embryonic stem cell (EST) testOne of the major drawbacks of micro-mass culture systems is that they requirethe primary culture of cells isolated fromembryos. Although avian embryos can beused in place of mammalian embryos,the isolation of fresh cells is still a limiting factor in the development ofthese systems. Researchers have noted,however, that embryonic stem (ES) celllines can be established from mammalianblastocysts. Essentially, embryonic stemcells can be maintained in an undifferen-
tiated state in the presence of feeder lay-ers and/or purified leukemia inhibitoryfactor (LIF). Upon removal of the LIF,ES cells differentiate into a variety of celltypes (Brown et al., 1995). Optimizationof culture conditions for ES cells hasbeen studied by several laboratories(Laschinski et al., 1989; Mummery et al.,1993) and attempts at validation of theassay as predictive of teratogenicity havebeen undertaken.
In one such study, Newall and Beedles (1994) examined the effects of25 compounds on mouse ES cells andcompared them to results obtained withrat primary embryonic cells in a micro-mass test protocol. The stem-cell testpredicted the teratogenicity of thesecompounds with a similar sensitiv i t yand specificity to the micromass test.The authors noted that an added adva n-tage of the stem-cell test was that “thetest used a propagated cell line; therewas no use of animals”. This particularmethod utilized colony formation as ameasurable endpoint of adverse eff e c t .S u b s e q u e n t l y, as discussed below, otherendpoints have been shown to be moreuseful as predictive tools.
Tab. 11: The effects of R-4-yn VPA on embryonic development in whole-embryo culturea (from Andrews et al., 1995)
C o n c e n t r a t i o n Yolk-sac Crown- Head length Somite Developmental Dead Abnormal Ending pH(mmol/liter) diameter rump length (mm) number score (%) (%)
(mm) (mm)
0 3.3 ± 0.08 2.7 ± 0.07 1.4 ± 0.05 21 ± 0.5 41 ± 1.1 6 19 7.05 ± 0.05
0.075 3.3 ± 0.09 2.7 ± 0.07 1.3 ± 0.04 20 ± 0.5 40 ± 1.3 14 21 7.05 ± 0.05
0.15 3.3 ± 0.07 2.7 ± 0.07 1.3 ± 0.04 19 ± 0.5 39 ± 1.1 0 44 7.07 ± 0.05
0.3 3.3 ± 0.07 2.7 ± 0.06 1.3 ± 0.04 20 ± 0.5 41 ± 1.1 0 25 7.07 ± 0.05
0.6 3.3 ± 0.08 2.6 ± 0.07 1.4 ± 0.04 20 ± 0.5 40 ± 1.1 6 31 7.12 ± 0.05
1.2 3.2 ± 0.07 2.7 ± 0.07 1.2 ± 0.04 19 ± 0.5 39 ± 1.1 0 25 7.07 ± 0.05
a Values are means ± SE for 14-16 embryos.* p ≤ 0.05 compared to controls.
Tab. 10: The effects of S-4-yn-VPA on embryonic development in whole-embryo culturea (from Andrews et al., 1995)
C o n c e n t r a t i o n Yolk-sac Crown- Head length Somite Developmental Dead Abnormal Ending pH(mmol/liter) diameter rump length (mm) number score (%) (%)
(mm) (mm)
0 3.4 ± 0.08 2.8 ± 0.07 1.35 ± 0.05 21 ± 0.5 41 ± 1.3 7 15 7.09 ± 0.04
0.075 3.4 ± 0.08 2.8 ± 0.06 1.34 ± 0.04 20 ± 0.5 41 ± 1.1 0 38 7.13 ± 0.05
0.15 3.3 ± 0.07 2.7 ± 0.07 1.31 ± 0.04 21 ± 0.5 38 ± 1.1 0 50* 7.13 ± 0.05
0.3 3.2 ± 0.08 2.6 ± 0.06 1.24 ± 0.04 19 ± 0.5 38 ± 1.1 0 56* 7.13 ± 0.05
0.6 3.2 ± 0.08 2.4 ± 0.06* 1.06 ± 0.04* 18 ± 0.5* 31 ± 1.1* 0 94* 7.19 ± 0.05
1.2 3.1 ± 0.13 2.3 ± 0.15* 0.93 ± 0.08* 16 ± 1.3* 25 ± 2.0* 69* 100* 7.28 ± 0.05
a Values are means ± SE for 13 - 16 embryos.* p ≤ 0.05 compared to controls.

HU G G I N S
ALTEX 20, Suppl.1/03 37
Utilizing a somewhat different proto-col, Spielmann and colleagues (1997)evaluated an embryonic stem cell testcomposed of two permanent mouse celllines, 3T3 fibroblasts and embryonicstem cells (D3). This evaluation investi-gated the effectiveness of 12 differentendpoints and endpoint ratios in predict-ing the embryotoxic potential of 16 testcompounds. Three endpoints (inhibitionof differentiation of ES cells into cardiacmyoblasts (identified by contraction) andcytotoxicity of ES cells and 3T3 cells(MTT-test)) were found to show a bettercorrelation to the embryotoxic propertiesof the test chemicals than the other end-points. Utilizing these endpoints, 50%inhibition concentrations for differentia-tion (ID50) and cytotoxicity (IC50D3 andIC503T3) were calculated from concen-tration-response curves. When coupledwith linear discriminant analysis, these
data formed the basis of a classificationscheme in which test chemicals were assigned to three classes: embryotoxic,moderately embryotoxic and stronglyembryotoxic. Using this model, all sixteen test chemicals were correctly assigned to their in vivo classes of em-bryotoxicity.
Subsequently, an interlaboratory com-parison of results obtained using the stemcell test described above was performed(Spielmann et al., 1998; Scholz et al.,1999a). Three test chemicals represent-ing three classes of embryotoxicity (non-, weak/moderate, and strong) wereevaluated by three different laboratoriesas part of the protocol transfer phase (II)of prevalidation experiments. In the finalphase (III) of prevalidation (protocol performance), nine test chemicals and apositive control of known embryotoxicpotential in vivo and similar potential in
animals and in humans were selected andtested by three laboratories. Because theoriginal prediction model calculated forthe EST was unsatisfactory, an improvediPM was utilized to analyze the resultsobtained from Phases I and II. The application of the iPM to phase III datafrom two of the participating laboratoriesprovided correct predictivities of 93 and94%, respective l y. Application of theiPM to all data from the three laborato-ries involved in Phase II of the study p r ovided a 100% correct prediction. Although there was some overlap in thechemicals tested in the different phasesof the study, the iPM proved to be moreappropriate than the original PM for datasets from different laboratories. Scholz etal. (1999b) discusses in-depth the im-proved prediction model (iPM). Unlikethe original PM, the improved PM incorporated as one variable the relative
Tab. 12: Results of the interlaboratory study for Hydroxyurea (from Bantle et al., 1994)
L a b o r a t o r y R e p l i c a t e L C5 0 Mean LC5 0 L C5 0 E C5 0 Mean EC5 0 E C5 0 M e a n T I M C I Gc M e a n M C I G( n ) (mg ml- 1) (mg ml- 1) CVa (mg ml- 1) E C5 0 C V ( % ) T Ib C V ( % ) (mg ml- 1) M C I G C V ( % )
( % ) (mg ml- 1) (mg ml- 1)
1 1 0.45 0.16 0.25
2 0.49 0.45 7.3 0.15 0.16 7.8 2.8 14.5 0.15 0.18 25.7
3 0.41 0.18 0.15
2 1 0.71 0.17 0.10
2 0.32 0.51 31.2 0.10 0.15 23.7 3.4 17.3 0.15 0.15 27.2
3 0.50 0.18 0.20
3 1 0.45 0.21 0.40
2 2.19 1.11 69.5 0.41 0.23 58.4 4.8 54.1 0.50 0.53 23.4
3 0.68 0.08 0.70
4 1 0.80 0.14 0.10
2 d 1.08 25.9 0.22 0.19 18.7 5.7 7.0 0.15 0.17 37.4
3 1.36 0.21 0.25
5 1 0.23 0.07 0.20
2 0.12 0.23 40.8 0.12 0.11 26.8 2.1 45.6 0.30 0.30 27.2
3 0.35 0.14 0.40
6 1 1.82 0.28 0.20
2 1.24 1.56 15.5 0.33 0.26 24.0 6.0 36.1 0.50 0.28 54.6
3 1.63 0.18 0.15
7 1 0.55 0.15 0.15
2 0.40 0.44 18.6 0.12 0.13 10.9 3.4 8.5 0.10 0.12 20.2
3 0.36 0.12 0.10
a Coefficient of variationb Mean teratogenic index (TI) calculated by dividing the mean LC50 by the mean EC50 for each laboratoryc Minimum concentration to inhibit growthd An LC50 could not be calculated because there was insufficient mortality in the concentration range tested

HU G G I N S
ALTEX 20, Suppl.1/0338
distance between IC503T3 and ID5 0, instead of the ratio ID50/IC50D3 that wasused previously.
Brown et al. (1995) noted that “EScells offer several new approaches withrespect to screening for embryotoxicityin vitro, enabling the use of differentiat-ing permanent embryonic cells”. Theyalso suggested that continued develop-ment of this assay should be supported.The endpoints used in the EST to date,those of cytotoxicity and effects on dif-ferentiation, have correlated reasonablywell with results obtained with in vivoteratogens. Many researchers feel, how-ever, that there are relevant endpoints yetto be discovered with this assay perhapsin conjunction with the use of transgenicanimals, novel immunologic stainingmethods and/or studies of changes ingene expression in ES cells.
5.2.4 Frog embryo teratogenesis assay (Xenopus) (FETAX)
Unlike the three assays discussed above,FETAX evaluates the effect of toxicantson the development of embryos from anon-mammalian species, the SouthAfrican clawed frog (Xenopus laevis).Originally developed by Dumont (1983), the assay has undergone a num-ber of interlaboratory validation studies(Bantle et al., 1994a, 1994b, 1996, 1999)(Tab. 12). Moreover, in 1991, the Ameri-can Society for Testing and Materials(ASTM) developed a test guideline for FETAX, which was subsequently r evised and republished in 1998 (ASTM, 1991, 1998). Recent evaluationsof FETAX data have focused on the utility of the assay as a screening protocol for deve l o p m e n t a l / r e p r o -d u c t ive toxicity testing in mammals as well as for ecotoxicological studies of potential reproductive hazard(NICEATM, 2000).
The rationale behind using FETAX as a monitor for teratogenic events inmammals is that embryonic developmentduring the first 96 hrs in Xenopus paral-lels many of the major processes of human organogenesis. FETAX measuresm o r t a l i t y, malformations and growth inhibition in a 96-hour in vitro whole-embryo test. Functional deficits resultingfrom exposure to a teratogenic substanceare not monitored in FETAX.
However, metabolic activation systemscan be incorporated into FETAX allow-ing for evaluation of substances that mayrequire metabolism in order to producet e r a t o g e n i c i t y. The concentration estimated to induce lethality in 50% ofthe exposed embryos (LC5 0) and the concentration estimated to induce mal-formations in 50% of the exposed embryos (EC50) are calculated based onmortality and malformation fi n d i n g s .The teratogenic index (TI) (LC50 dividedby the EC50) is also calculated. Growth isestimated by measuring the head to taillength of the embryos. The minimumconcentration to inhibit growth (MCIG)is determined by comparing the mean96-hour head to tail length of the treatedembryos at each treatment concentrationto that of the control embryos. Any oneof these three criterion (TI, growth inhibition, or severity of induced mal-formations) is used to identify a test material as teratogenic.
5.3 Validation and regulatory activities
The Organization for Economic Cooper-ation and Development (OECD) guide-lines for developmental toxicity do notexplicitly restrict developmental toxicitytesting to mammals (OECD, 1981,1983a, 1983b, 1995, 1996). However,most current regulatory guidelines ford eve l o p m e n t a l / r e p r o d u c t ive toxicity testing include tests that use animals,particularly mammals, when consideringpotentially adverse effects on humans.As noted above, the complexity of thedevelopmental and reproductive systemsin humans and other animals has hindered the search for an eff e c t ive alternative assay or battery of assays.This hindrance, not surprisingly, is reflected in the current lack of strong regulatory initiatives supporting replace-ment alternatives to deve l o p m e n t a l /reproductive toxicity testing in animals.However, use of alternative assays asscreening tests for reproductive toxicantsseems to be gaining a foothold as valida-tion efforts provide additional evidencethat several of these assays are indeedsuitable predictors of in vivo reproduc-tive toxicity.
E C VAM (European Center for the Validation of A l t e r n a t ive Methods) is
continuing work with a full validation of the MM and WEC assays as well as the EST (Prieto, 2000). Moreover,Genschow et al. (2000) cite encouragingresults regarding development and use ofprediction models for these assays. ThePMs generated for the MM and WEC assays, provided 81% (MM) and 72%(WEC) correct classifications. The im-proved PM (iPM) for the EST resulted in79% correct classifications when com-pared to embryotoxic potential definedby in vivo testing. Although, the resultsgiven for the MM and WEC assays maybe modified downward when the entirecontingent of 20 chemicals is tested, thepercentages obtained thus far indicate areasonable degree of correlation betweenthese three in vitro assays and their in vivo counterparts.
An important, although often ove r-looked, aspect of validation experiments,is that they often serve to refine the protocols undergoing validation. Exam-ples of this are found in results fromwork with both the MM and WEC assaysas cited by Genschow. Analysis of MMdata indicated that the midbrain cells inthe MM assay did not provide re-producible data. Hence, prevalidation ofthis assay only includes data obtainedfrom limb bud cells. Furthermore, be-cause the concentration response curvesof the differentiation of limb bud cellsmonitored by AB staining and cytotoxic-ity assessed by neutral red uptake werealmost identical, only differentiation oflimb bud cells was considered because itshowed a higher predictive power thancytotoxicity. Moreover, although a widespectrum of results is available for evaluation in the WEC assay (i.e. 16 parameters for general morphologicaldevelopment and 30 types of malforma-tions), only morphological development(total morphological score) and the per-centage of malformed embryos provideds u fficient information for predicting embryotoxic potential in a PM. This typeof refinement (toward more relevant endpoints for prediction modeling) tendsto simplify assay procedures. Ultimately,this simplification should enhance the effectiveness of in vitro assays, whethert h ey monitor developmental and/or reproductive toxicity or other types oftoxicity.

HU G G I N S
ALTEX 20, Suppl.1/03 39
The USEPA has also recently request-ed that the Interagency CoordinatingCommittee on the Validation of Alterna-tive Methods (ICCVAM) evaluate thevalidation status of FETAX. In responseto this request, ICCVAM in conjunctionwith NICEATM (National To x i c o l o g yProgram Interagency Center for the Eval-uation of Alternative Testing Methods)has finished a critical evaluation of F E TAX (NICEATM, 2000). In this evaluation, NICEATM assessed the status of FETAX as a screening assay fordetection of potential human teratogens.FETAX has been proposed as a screeningtest that, if positive, would indicate a potential human hazard, and, in the ab-sence of other data, would be considereda presumptive teratogen/developmentaltoxicant. A negative FETAX responsewould not necessarily indicate the absence of a hazard, and negative re-sponses would be followed by definitivein vivo mammalian testing. NICEATMcompiled information from 276 studiesinvolving 137 substances as it pertains tohuman development hazard assessment.
F E TAX performance characteristics(i.e. accuracy, sensitiv i t y, specifi c i t y,positive predictivity, negative predictivi-ty, false positive rate, and false negativerate) against rat, mice, and/or rabbit teratogenicity test results or human teratogenicity study results were deter-mined. The decision criteria used in determining the performance characteris-tics of FETAX included single decisioncriteria (TI > 1.5; TI > 3.0; MCIG/LC50
< 0.30) and multiple decision criteria (TI > 1.5 plus MCIG/LC50 < 0.30; TI > 3.0 plus MCIG/LC50 < 0.30). When amultiple decision criterion was used, testsubstances were classified as positivewhen both the TI values were greaterthan the decision point (1.5 or 3.0) andthe MCIG/LC50 ratio was less than 0.3;equivocal when one, but not both, criteri-on were positive; and nega t ive when neither criterion was positive.
The values obtained from this analysissuggested that the use of FETAX as ascreen, based on current decision criteria,may pose some problems, although theassay is considered robust in many respects. Three approaches were ad-vanced for modifying decision criteria to increase the ability of FETAX to cor-
rectly identify developmental toxicants.The first approach would be to evaluatethe EC50 based on characteristic mal-formations (i.e., those increasing in incidence and severity with increasingtest substance concentration) only.(Note: An atlas addressing X e n o p u smalformations for use with FETAX hasrecently been published (Bantle et al.,1998)). The second approach is to calculate a point estimate for the dosethat inhibits growth by 50% rather thanusing an MCIG. The third approachwould be to use 95% confidence intervalsfor statistically identifying TI values (and other point estimates) that are significantly different from the decisioncriteria value. This report also found excessive variability in the LC50, EC50,TI, and especially the MCIG within andacross laboratories participating in thevalidation studies conducted with FETAX thus far. No immediate reasonfor this variability is apparent, althoughthe lack of classification of malforma-tions as to type and severity seems to bea contributing factor. On a more positivenote, NICEATM’s evaluation indicatedthat the projected cost (<$25,000) andstudy duration (less than two months),for a Good Laboratory Practice (GLP)compliant FETAX study, with and with-out metabolic activation were reasonablein comparison to the cost of a completerat Prenatal Developmental To x i c i t yStudy (approximately $120,000).
Overall, validation of several in vitroassays of deve l o p m e n t a l / r e p r o d u c t ivetoxicity seems to be reaching a criticalculmination.
5.4 Summary, conclusions, and future work
In summary, validation efforts are pro-gressing well for four in vitro assays of developmental/reproductive toxicity.Results from evaluations of the MM andWEC assays as well as the EST appear to be favorable; data from studies of FETAX suggest that further improve-ments in the assay would yield greaterp r e d i c t iv i t y. The fact that the OECD does not restrict its reproductive testingguidelines to mammals, coupled with recent initiatives on the part of theUSEPA seem to suggest that regulatorysupport of alternatives assays of develop-
ment and reproductive toxicity is present,although, perhaps not as vigorous as itcould be. This reticence may reflect boththe complexity of the reproductive cycleand the need for validated in vitro assays,which are, as suggested by the discussionabove, “on the horizon”.
The next five years of research in thisarea should yield assays that are accept-able to both scientific and regulatorycommunities as screening tools for developmental and reproductive toxicity.This acceptance should promote muchmore widespread incorporation ofscreening batteries utilizing these assaysinto toxicity testing regimens for newcompounds. Schwetz (1993) has pointedout, how eve r, that an in vivo s c r e e nshould perhaps be performed first with asmall number of animals over a range ofdose levels simply to see if the com-pound does indeed cause reproductive ordevelopmental effects.
Over the next ten years, it may be possible to develop in vitro assays thatrepresent much more selectively the various components of the reproductivecycle (i.e. the complexity of the systemwill be much more well-represented).This should help tremendously in identi-fying the exact locus of toxic action andin acceptance of in vitro assays as true replacement alternatives. Moreove r,Brown et al. (1995) recommended that,“Methods using human semen for testingthe effects of chemicals on mature spermshould be refined and validated”.
Development of alternative assays ofdevelopment and reproduction over thenext twenty years may well incorporate“pattern of response” approaches such as those described in discussions of proteomics or toxicogenomics. Brown(1998) has already stated, “It is importantthat these new strategies (embryonic approaches) are not bedevilled by naiveexpectations, particularly in the earlystages of their use. The V word (valida-tion) should be locked away, in favor of “profiling”, in which we ask ‘Can this chemical affect this particular pathway?’” Moreover, he predicts that,“Once answers are available for manychemicals and pathways, patterns of response will be assembled, and thesemay allow the prediction of some typesof developmental toxicity.” T h e s e

HU G G I N S
ALTEX 20, Suppl.1/0340
thoughts are mirrored in the NICEATMreport on FETAX (2000) that suggeststhat cDNA microarray technology would “greatly increase the utility of FETAX for identifying and prioritisingdevelopmental hazards”. Hence, “profil-ing” of chemical pathways after repro-ductive toxicant exposure with cDNAmicroarray technology may well be thenext definitive step in development of a l t e r n a t ives to the use of animals in d eve l o p m e n t a l / r e p r o d u c t ive toxicity testing.
ReferencesAmacher, D., Stadler, J., Schomaker, S.
and Verseil, C. (1996). The compara-tive testing of eight coded chemicals inthe rat limb bud micromass and rat embryo culture systems. AlternativesLab. Animal 24 (6), 945-954.
Andrews, J. E., Ebron-McCoy, M., Bojic,U. et al. (1995). Validation of an in vitro teratology system using chiralsubstances: stereoselective terato-genicity of 4-yn-valproic acid in cultured mouse embryos. Toxicol. Appl. Pharmacol. 132 (2), 310-316.
ASTM (American Society for Testingand Materials) (1991). Standard Guidefor Conducting the Frog Embryo Ter-atogenesis Assay – Xenopus (FETAX).ASTM E1439–91. In Annual Book ofASTM Standards, Philadelphia, PA.
ASTM (1998). Standard Guide for Conducting the Frog Embryo Terato-genesis Assay – Xenopus (FETAX).ASTM E1439-98. In Annual Book ofASTM Standards, Philadelphia, PA.
Bantle, J. A., Burton, D. T., Dawson, D.A. et al. (1994a). Initial interlaboratoryvalidation study of FETAX: Phase Itesting. J. Appl. Toxicol. 14, 213-223.
Bantle, J. A., Burton, D. T., Dawson, D.A. et al. (1994b). FETAX interlabora-tory validation study: Phase II testing.E nv i ron. Toxicol. Chem. 13, 1629-1637.
Bantle, J. A., Finch, R. A., Burton, D. T.et al. (1996). FETAX interlaboratoryvalidation study. Phase III, part 1 test-ing. J. Appl. Toxicol. 16, 517-528.
Bantle, J. A., Dumont, J. N., Finch, R. A.et al. (1998). Atlas of Abnormalities: AGuide for the Performance of FETAX,2 nd ed. Stillwater, OK: Oklahoma StateUniversity Press.
Bantle, J. A., Finch, R. A., Fort, D. J. etal. (1999). Phase III interlaboratorystudy of FETAX, part 3 – FETAX validation using 12 compounds withand without an exogenous metabolica c t ivation system. J. Appl. To x i c o l .19(6), 447-472.
Bechter, R., Terlouw, G. D., Tsuchiya,M. et al. (1991). Teratogenicity ofarotinoids (retinoids): comparison ofthe whole embryo culture system withthe in vivo mouse model and the limbbud cell culture. Te ra t o l ogy 44(3), 29A.
Brown, N., Spielmann, H., Bechter, R. etal. (1995). Screening chemicals for re-productive toxicity: the current alter-natives. The report and recommenda-tions of ECVAM workshop 12. ATLA23, 868-882.
B r own, N. A. (1998). Deve l o p m e n t a ltoxicology in vitro: current status andimminent advances. Hum. Exp. Toxi-col. 17(9), 483.
Cosenza, M. and Bidanset, J. (1995). E ffects of chlorpyrifos on neuronal development in rat embryo midbrainmicromass cultures. Vet. Hum. Toxicol.37(2), 118-121.
Cosenza, M., Bidanset, J. and Johnson,D. E. (1996). In vitro/in vivo correlation of paraquat induced devel-opmental neurotoxicity. To x i c o l og i s t30 (I Part 2), 244.
Dumont, J. N., Schultz, T. W., Buchanan,M. W. and Kao, G. L. (1983). III. FrogEmbryo Teratogenesis Assay: Xeno-pus (FETAX) – A short-term assay applicable to complex environmentalmixtures. In M. Waters, S. Sandhu, J.Lewtas, L. Claxton, N. Chernoff, andS. Nesnows (eds.), Short-term Bio-assays in the Analysis of Complex E nv i ronmental Mixtures ( 3 9 3 - 4 0 5 ).New York: Plenum.
Faustmann, E. M. (1994). Mammalianmicromass embryo cell cultures: appli-cations in developmental toxicity eval-uation and mechanistic investigations.In Vitro Toxicol. 7(2), 158.
Flint, O. P. (1983). A micromass culturemethod for rat embryonic neural cells.J. Cell Sci. 61, 247-262.
Flint, O. P. and Orton, T. C. (1984). An invitro assay for teratogens with culturesof rat embryo midbrain and limb bud cells. Toxicol. Appl. Pharmacol.76, 383-395.
Genschow, E., Scholz, G., Brown, N. etal. (2000). Development of predictionmodels for three in vitro embryotoxic-ity tests in an ECVAM va l i d a t i o nstudy. In Vitro & Molecular Toxicology13(1), 51-65.
Gray, T. and Kimber, I. (1989). Repro-ductive and developmental toxicology:the value and limitations of in vitrotechniques. Toxic. In Vitro 3(3), 241-242.
Gregotti, C., Manzo, L., Costa, L. G. andFaustman, E. M. (1992). Developmen-tal toxicity of styrene oxide in the post-implantation embryo culture. Toxico-logist 12(1), 333.
Haberstroh, K. M. and Kapron, C. M.(1998). Cellular mechanisms of cad-mium embryonic toxicity. Teratology57(4/5), 228.
Laschinski, G., Vogel, R. and Spielmann,H. (1989). Establishment of embryon-ic stem cells as a test system for embryotoxicity. Teratology 40(3), 289.
Mayura, K., Smith, E. E., Clement, B. A.and Phillips, T. D. (1991). Evaluationof the developmental toxicity of chlorinated phenols utilizing Hydra attenuata and postimplantation rat embryos in culture. Toxicol. A p p l .Pharmacol. 108(2), 253-266.
McMillan, M. J., Ou, Y. C., Ponce, R. A.et al. (1997). Inve s t i gation of the temporal correlation betweenm e t hylmercury induced cell cycle arrest and indution of cell cycle specific genes. Teratology 55(1), 60.
Mirkles, P. (1996). Prospects for the d evelopment of validated screeningtests that measure developmental toxicity potential: view of one skeptic.Teratology 53(6), 334-338.
Mummery, C. L., Slager, H. G., vanInzen, W. et al. (1993). Regulation ofg r owth and differentiation in early d evelopment: of mice and models. Reprod. Toxicol. 7(1), 145-154.
N I C E ATM (National Toxicology Pro-gram Interagency Center for the Evaluation of Alternatives Toxicologi-cal Methods) (2000). Background R ev i ew Document. Frog Embryo Teratogenesis Assay – Xenopus (FETAX). March 10.
Newall, D. R. and Beedles, K. E. (1994).The stem-cell test: A novel in vitro assay for teratogenic potential. Toxic.

HU G G I N S
ALTEX 20, Suppl.1/03 41
In Vitro 8(4), 697-701. OECD (Organization for Economic
Cooperation and Deve l o p m e n t ) .(1981). OECD guideline for testing of chemicals 414: Te r a t o g e n i c i t y.OECD, Paris.
OECD (1983a). OECD guideline fortesting of chemicals 415: One-genera-tion reproduction toxicity study.OECD, Paris.
OECD (1983b). OECD guideline fortesting of chemicals 416: Two-genera-tion reproduction toxicity study.OECD, Paris.
OECD (1995). OECD guideline for testing of chemicals 421: Reproduc-tion/developmental toxicity screeningtest. OECD, Paris.
OECD (1996). OECD guideline for test-ing of chemicals 422: Combined re-peated dose toxicity study with the re-p r o d u c t i o n / d evelopmental toxicityscreening test. OECD, Paris.
Prieto, P. (2000). ECVA M ’s in-houseprevalidation/validation studies in theareas of haematotoxicity, reproductivetoxicity, metabolism-mediated toxicityand epithelial barrier function. Sci. Total. Environ. 247, 349-354.
Renault, J., Caillaud, J., Melcion, C. andCordier, A. (1989). Micromass limbbud cell culture: a model for studying teratogenic mechanisms. Te ra t o l og y40(3), 295.
Renault, J., Caillaud, J. and Chevalier, J.(1995). Ultrastructural characteriza-tion of normal and abnormal chondro-genesis in micromass rat embryo limbbud cell cultures. Toxicol. Appl. Phar-macol. 130(2), 177-187.
Scholz, G., Pohl, I., Genschow, E. et al.(1999a). Embryotoxicity screening using embryonic stem cells in vitro:Correlation to in vivo teratogenicity.Cells Tissues Organs 165(3-4), 203-211.
Scholz, G., Genschow, E., Pohl, I. et al.(1999b). Prevalidation of the embryon-ic stem cell test (EST). A new in vitro embryotoxicity test. Toxic. In Vi t ro13(4-5), 675-681.
Schwetz, B. A. (1993). Utility of in vitroassays: summary of internationalworkshop on in vitro methods in reproductive toxicity. Reprod. Toxicol.7, 171-173.
Spielmann, H., Pohl, I., Dsoring, B. et al.(1997). The embryonic stem cell test,an in vitro embryotoxicity test usingtwo permanent mouse cell lines: 3T3fibroblasts and embryonic stem cells.In Vitro Toxicol. 10(1), 119-127.
Spielmann, H., Scholz, G., Bremer, S. andH o l z h u e t t e r, H. G. (1998). In vitro test-ing of embryotoxicity using the embry-onic stem cell test (EST): an interlaboratory comparison. In Vi t roCell Dev. Biol. - Animal. 34 (3 Pt 2), 11A.
U S E PA (United States Env i r o n m e n t a lProtection A g e n cy) (1998a). HealthE ffects Test Guidelines. OPPTS870.3770. Prenatal Deve l o p m e n t a lToxicity Study.
U S E PA (1998b). Health Effects Te s tGuidelines. OPPTS 870.3800. Repro-duction and Fertility Effects.
U S E PA (2000a). Health Effects Te s tGuidelines. OPPTS 870.3550. Repro-duction and Developmental ToxicityScreening Test.
U S E PA (2000b). Health Effects Te s tGuidelines. OPPTS 870.3650. Com-bined Repeated Dose Toxicity Studywith Reproduction and Deve l o p-mental Toxicity Screening Test.
Walum, E. and Flint, O. P. (1993). Selec-tive effects of acrylamide, methylenebisacrylamide, and haloperidol on neu-ronal d evelopment in rat embryomidbrain micromass cultures. In VitroToxicol. 6(2), 125-134.
Watanabe, T. and Iwase, T. (1996). Developmental and dysmorphogeneiceffects of glufosinate ammonium onmouse embryos in culture. Teratog.Carcinog. Mutagen. 16(6), 287-299.
Whittaker, S., Wroble, J., Silbernagel, S.and Faustman, E. (1993). Characteri-zation of cy t o s keletal and neuronalmarkers in micromass cultures of ratembryonic midbrain cells. Cell Biol.Toxicol. 9(4), 359-375.
6 Genetic engineering methodologies
6.1 BackgroundTo say that genetic engineering is a controversial issue is to make an under-statement. The controversy stems fromthe moral debate over whether thetremendous contribution “engineered”genes can make to medical science is justified in view of the possibility of inappropriate, misguided use of this relatively new technology. Genetic engi-neering can indeed be used to enhancethe welfare of both humans and animals.However, questions remain as to whetherhumane considerations can be incorpo-rated into the medical research needed todevelop applications of genetic engineer-
ing. Moreover, there are concerns overwhether the application itself is, in someinstances, humane.
Examples of recent events in the realm of genetic engineering include thepatenting of a process for engineeringhuman cells that can be used to grow avariety of human tissues in the laborato-ry (Anonymous, 1998). Researchersclaim that the techniques could be usedto grow an endless supply of human o rgans for transplant. Furthermore, human genes have been successfully in-serted into a pair of lambs supposedlyendowing them with the DNA to aid burnvictims (Arent, 1999). Moreove r, the
American Medical Association recentlylistened to proposed ethical guidelinesfor animal-to-human organ or cell trans-plants. The committee is recommending,among other things, that human subjectsin clinical trials of animal-to-humantransplantation – or xenotransplantation– be required to waive their traditionalright to drop out of a study. This sugges-tion has met with considerable resistancefrom groups such as the Campaign for Responsible Transplantation (Philip-koski, 2000). Hence, genetic engineeringcontinues to find numerous applicationsin the field of biomedical research and to provo ke heated debate over the

HU G G I N S
ALTEX 20, Suppl.1/0342
ethical implications of those applica-tions.
What specific contributions can genet-ic engineering make to the animal welfare movement? How are animals ad-versely affected by genetic engineering?Quite possibly, answers will be found asgenetic engineering evolves. However,preliminary consideration of the issueswill contribute much to our preparedness.
Such consideration has been the focusof several meetings convened under theauspices of the European Center for the Validation of Alternative Methods( E C VAM). One workshop on genetic engineering was organized in “recogni-tion that genetically engineered celllines, particularly those expressing xeno-biotic metabolizing enzymes, are beingused increasingly in toxicological andpharmacological studies” (Weibel et al.,1997). An additional ECVAM gatheringon the use of transgenic animals in theEuropean Union was held to “formulate
a set of guidelines to assist regulatory authorities in the EU in deciding whetherto permit and/or how to regulate re-search involving transgenic animals”.(Metpham et al., 1998). Much work isstill needed, however, to ensure that thebenefits of genetic engineering can be realized without sacrificing animal welfare.
6.2 Genetically engineered cell linesGenetically engineered cells can be usedfor determining the activity of specificenzymes in the metabolic profiles ofxenobiotics, to screen chemicals for t o x i c i t y, and to evaluate molecular mechanisms of action. Heretofore, although representing a useful alternativetest system for cytotoxicity or genotoxic-i t y, established cells lines lacked sufficient endogenous xenobiotic metab-olizing enzyme activity to make them agood predictor of human or animal response to toxicants. However, Weibel
et al. (1997) indicate that xenobiotic metabolizing enzymes have now beenexpressed in bacteria, yeast, insect cells,and mammalian cells. T h ey cite 20 examples of cell lines, mostly derivedfrom human and hamster tissues, whichh ave been genetically engineered for expression of xenobiotic metabolizingenzymes in the last decade. They alsosuggest that human cells expressing human xenobiotic metabolizing enzymesare more likely to be predictive of humansusceptibility to toxicants than non-human cells.
Crespi has written extensively aboutthe use of genetically engineered humancells. He suggests that, “Identification ofthe human enzymes invo l ved in the metabolism of a series of investigationalcompounds (e.g. candidates for drug development) can help direct the choice,among the alternatives, of the best in-ve s t i gational compound to take into d evelopment, can help focus in vivo
Fig. 11: Mutagenic activation of IQ in the cell lines expressingCYP1A2 and NAT.ANM-13 and ANP-25 are cell lines expressing CYP1A2 plus NAT orN AT2, re s p e c t i v e l y. Top, relative survival rates after a 24-h tre a t m e n twith IQ. Bottom, mutant frequency expressed as the number of 6-t h i o - g u a n i n e - resistant cells/106 viable cells (from Yanagawa et al.,1994). o: A2R-5; ◊: ANM-13, •: ANP-25
Fig. 12: Mutagenic activation of MeIQx in the cell linesexpressing CYP1A2 and NAT (from Yanagawa et al., 1994). o: A2R-5; ◊: ANM-13, •: ANP-25

HU G G I N S
ALTEX 20, Suppl.1/03 43
studies to specific areas of concern andcan help elucidate the mechanisms forinterindividual variability in response ortoxicity” (Crespi, 1995). He also com-ments that much of the progress that hasbeen made in the development of cellculture systems that contain xenobiotic-metabolising capacities is due to recom-binant DNA techniques. ComplementaryD NA (cDNA) encoding nearly every major human P450 form has been isolat-ed and sequenced.
Crespi and Penman (1997) have studied the use of single P450 enzymesystems, relative to multienzyme systemsusing cDNA. T h ey suggest that this approach has major advantages and disadvantages. Direct incubation of a testcompound with microsomes preparedfrom cells expressing a single cy-tochrome P450 form coupled with ananalysis of metabolite formation or lossof parent compound can yield significantinsight into cytochrome P450 “form-se-lective” metabolism of a test compound.Use of single P450 enzyme systems canalso be used to study the inhibition of human, drug-metabolizing cytochromesP450. In a recent study, microtiter plate-based fluorometric assays for theactivities of five principal drug-metabo-lizing enzymes, CYP1A2, CYP2C9,CYP2C19, CYP2D6 and CYP3A4 wereevaluated. (Crespi et al., 1997). The authors found that their observed IC50
values were in good agreement withthose reported for the same enzyme/in-hibitor pairs reported by others.
Single-enzyme experiments may notpredict, how eve r, the activity of the balance of enzymes present in humanl iver microsomes. In their rev i ew of genetic engineering, Weibel et al. (1997)recommended that, “For studies onmechanisms of metabolic activation, enzymes acting in concert, such as cytochrome P4501A2 and N-acetyltrans-ferases or sulphotransferases, need to bec o - expressed in recombinant systems.This also applies to enzymes involved incompeting metabolic pathways”. Crespi(1995) concurs by suggesting that, “co-expression of many enzymes within asingle cell will be an important techniquefor the analysis of the balance of Phase Iand Phase II enzymes on overall metabo-lism and/or toxicity of xenobiotics”. He
and others demonstrated the feasibility ofthis approach with the development ofMCL-5 cells that coexpress five cDNAs(Crespi et al., 1991). More recently,S awada and Kamataki (1998) have r ev i ewed the stable expression of cytochrome P450 in mammalian celllines and its application to toxicity testing. In their discussion the successfulreconstruction of an entire metabolic activation system for certain heterocyclicamines is described. The system consistsof mammalian cell lines that simultane-ously express a form of human P450 anda phase II enzyme, N-acetyltransferase(Fig. 11 and 12) (Yanagawa et al., 1994).
Kranendonk et al. (1999) have also recently reported on the genetic engineering of four new Escherichia colitester bacteria, coexpressing humanCYP1A1, CYP2A6, CYP3A4 or CYP3A5with human NADPH cytochrome P450reductase (RED) by a biplasmid co-expression system, recently developed toexpress human CYP1A2 in the testerstrain MTC. They found these four newstrains to “contain stable CYP- andRED-expression, significant CYP-activi-ties and demonstrated significant bio-activation activities with several diagnos-tic carcinogens” (Tab. 13).
However, Crespi and Penman (1998)have cautioned that considerable workremains to be done to develop appropri-
ate approaches for use of these data topredict the consequences of human expo-sure to xenobiotics. Both appropriate validation of systems containing human,c D NA - expressed xenobiotic meta-bolizing enzymes and integration of experimental results from host cells expressing different amounts of enzymeactivity than those found in vivo need tobe addressed.
Other areas of research benefiting fromhuman cells genetically engineered tocontain cytochrome P450 metabolic enzymes include cancer research andt h e r a py. Recent examples include thework of Hanna et al. (2000) who re-searched the association of cytochromeP450 1B1 (CYP1B1) polymorphismswith functional differences in estrogenhydroxylation activity. They evaluatedwhether inherited variants of CYP1B1differ from wild-type CYP1B1 in estro-gen hydroxylase activity by developingrecombinant wild-type and five polymor-phic variants of CYP1B1. The CYP1B1variants formed 4-OH-E2 as the mainproduct (as did the wild type), but dis-played 2.4- to 3.4-fold higher catalyticefficiencies. The variant enzymes alsoexceeded wild-type CYP1B1 with re-spect to 2- and 16alpha-hydroxylationa c t iv i t y. The authors concluded that inherited alterations in CYP1B1 estrogenhydroxylation activity may be associated
Tab. 13: Mutagenic activities of chemical carcinogens in strain MTC/CYP expressingfive different human CYP isoforms (from Kranendonk et al., 1999)
Carcinogen Strain Mutagenic activity(revertants per nanomole± standard deviation)
2AA MTC1A2 3433 ± 447
MTC2+S9 108 ± 4a
AFB1 MTC1A1 77 ± 12
MTC1A2 5437 ± 555
MTC2A6 421 ± 10
MTC3A4 2349 ± 330
MTC3A5 978 ± 59
MTC2+S9 13247 ± 732a
B[a]P MTC1A1 798 ± 60
MTC2+S9 84 ± 8a
DMBA MTC1A1 682 ± 77
MTC2+S9 5 ± 1b
a Determined with non-induced strain MTC1A2b Determined with strain MX100

HU G G I N S
ALTEX 20, Suppl.1/0344
with significant changes in estrogen metabolism and, “thereby, may possiblyexplain interindividual differences inbreast cancer risk associated with estro-gen-mediated carcinogenicity”. More-ove r, Waxman et al. (1999) have reviewed recent advances in cancer genetherapy in which an intratumoral prodrugactivation strategy based on the combina-tion of a cytochrome P450 gene with thegene encoding NADPH-P450 reductaseis utilized. Transduction of tumors with aprodrug-activating P450 gene, followedby prodrug treatment, greatly increasesintratumoral formation of activated drugmetabolites. This leads to “more efficientkilling of the transduced tumor cellswithout a significant increase in host toxicity“.
Among the conclusions and recom-mendations of the ECVAM report(Weibel et al, 1997) regarding further de-velopment of recombinant systems werethe following:
- There is a need to develop systemswhich express xenobioticmetabolizing enzymes, other thanthe cytochromes P450.
Recent work by Hu et al. (1999) studied the transport and metabolic char-acterization of Caco-2 cells expressingCYP3A4 and CYP3A4 plus oxidoreduc-tase. They found that the monolayers of newly transfected cells (CYP3A4 +oxidoreductase) have significantly in-creased levels of CYP3A4 activ i t i e scompared to untransfected cells. Thesecell monolayers also have desirable mor-phological and transport characteristicsthat are similar to untransfected cells(Tab. 14).
- For risk assessment purposes,xenobiotic metabolizing enzymes of commonly used species oflaboratory animals need to beexpressed in recombinant cellsystems. Priority should be given to rat enzymes.
Recent work by Grove et al. (2000) in-vestigated the influence of rat UGT1A7on B[a]P-induced cytotoxicity. Humanlymphoblastoid L3 cells were transfectedwith pMF6 (control expression vector),p167Dtk2 (microsomal epoxide hydro-lase expression vector), or P167Dtk2-1A7 (epoxide hydrolase/UGT1A7 coex-pression vector), and the cell populations
were compared for sensitivity to B[a]P-induced effects. The data obtained sug-gested that UGT1A7 may be preferen-tially active toward B[a]P-quinones andthat UGT1A7 may represent the PAH-inducible UGT activity previously impli-cated in protection against toxic redoxcycling by B[a]P-3,6-quinone.
- It is essential to establishmammalian cell lines which exhibithigh levels of enzyme activities (for example, of the magnitudeachievable in bacterial recombinantsystems). Preferably, the host cells should be of human origin.
Crespi (1995) echoed these sentimentssuggesting that “higher expression levelsare necessary for routine use of such asystem for metabolite analysis”.
Recent work by Scott et al. (1999)studied the expression and partial puri-fication of a recombinant secretory formof human liver carboxylesterase (CaE)from human cells in which the car-boxylesterase was overexpressed. Car-boxylesterases play an important role inthe detoxification of xenobiotic chemi-cals that contain organophosphate compounds; high activity is found in theliver. In Scott’s work, a single amino acidchange was made in the C-terminal retrieval signal of a human liver CaE. Arecombinant plasmid, pRc/CMV- m h-
CaE, containing the amino acid change,was isolated and stably integrated intohuman 293T cells. Expression of the altered cDNA resulted in secretion of anactive CaE up to levels of 500 enzymeunits per liter of growth medium. Thissecretory CaE displayed isoelectric focusing patterns similar to those of then a t ive enzyme with no observa b l echanges in activity. The secreted enzymewas partially purified and CaE retained ahigh level of enzymatic activity.
All of these research findings indicatethat progress is steadily being made inrecombinant genetic engineering as it relates to toxicology and drug metabo-lism as well as to areas such as cancer research.
6.3 Transgenic animalsTransgenic animals are a product of genetic engineering and they are a goodexample of the dichotomy inherent tothis area of research as mentioned earlierin this article. There are definitely usesfor transgenic animals that contribute agreat deal to toxicological research.However, their negative effects on animalw e l fare may outweigh their contribu-tions. Metpham et al. (1998) state that“while transgenic animals might allowreduction and refinement in animal
Tab. 14: Metabolic properties of model compounds in Caco-2 cell variants (from Hu et al., 1999)
Metabolic rates (pmol/min/mg protein)
Parameters/drugs TC7 clone CYP3A4#4 CYP3A4+OR#8
Drugs
testosterone (Tes) BDLa 7-32 5.20 ± 0.48
nifedipine (Nif) 1 2-10 1.36 ± 0.08
linopirdine (Lin) BDL 12.3 ± 1.4 ND
midazolam ND 1.09 ± 0.12 ND
warfarin BDL BDL ND
Drug + Modulator (% Control)
Tes + ketoconazole BDL 7.0 ± 0.0 ND
Nif + ketoconazole 12 ± .16 5 ± 0.2 ND
Nif + TPA NDb 145 ± 5 ND
Lin + ketoconazole ND 57.6 ± 3.8 ND
Lin + TPA ND ND 4.71 ± 0.07c
Tes + TPA ND 183 ± 21 7.57 ± 0.44c
a “BDL” stands for “below detection limit” of 0.25 pmol/min/mg protein.b “ND” stands for “not determined”.c Actual rates of metabolism with a unit of pmol/min/mg protein.

HU G G I N S
ALTEX 20, Suppl.1/03 45
use via more precise gene targeting inbreeding programs, these objectives arethreatened by transgenic procedureswhich could promote greater animal use, agreater variety of applications, and an in-creased likelihood of animal suff e r i n g ”.
Currently transgenic animals are usedin numerous basic research applications,as well as in the development of desir-able traits in agricultural products andl ivestock. T h ey are considered “bio-reactors” for the mass production of bio-logical products. They are also used totest vaccines. Moreover, several types oftransgenic animals are either used or “ e nvisaged” for specific toxicologicaltesting procedures.
One of the most widely discussed usesof transgenesis in toxicology is that oftransgenic mice as predictive models foridentifying carcinogens. Advantages tousing this model or others like it includeuse of fewer animals, performance of theassay in a short time period, and muchlower cost than the average two-year rodent bioassay. Yamamoto et al. (1998)states that, “In vitro testings, i.e. muta-genicity tests, transformation assays, etc.may be helpful to predict carcinogenici-ty; but such in vitro testing is still subsidiary. Carcinogenicity testing usingexperimental animals is the only way forthe prospective identification of possiblehuman carcinogens”. They indicate thatanimals susceptible to carcinogens (i.etransgenic (Tg) animals) are indispensa-ble and describe the validation of trans-genic mice carrying the human prototypec-Ha-ras gene as a bioassay model forrapid carcinogenicity (Tab. 15).
Others have also evaluated transgenicmice as predictive models for carcino-genesis. Eastin (1998) describes a preliminary assessment by the U.S. National Toxicology Program (NTP) ofthe utility of the p53def (hemizygous forthe tumor-suppressor gene) and theTg.AC (carrier of an activated H-rasoncogene) transgenic mice strains asmodels for identifying chemical carcino-gens and assessing risk. Moreove r, Contrera and DeGeorge (1998) as well asMonro and MacDonald (1998) discussthe evaluation of the carcinogenic poten-tial of pharmaceuticals using transgenicmice in response to recommendations bythe International Conference on Harmon-
isation (ICH) that allow the substitutionof one species in the standard 2-year cancer bioassay with a short- or medium-term alternative model (ICH, 1996).
Furthermore, Thompson et al. (2000)have recently investigated a battery ofprescreening methods based on compar-isons with the Tg.AC mouse model. Theydeveloped a battery based on correct responses to 24 compounds tested pre-viously in Tg.AC mice. Included in thebattery were assays chosen to reflect molecular pathways possibly involved inthe Tg.AC papilloma formation: zeta-globin promoter-luciferase construct sta-bly expressed in K562 cells (Zeta-Luc)and three of the stress-response element-chloramphenicol acetyltransferase (CAT )fusion constructs stably expressed inHepG2 cells that are a part of the CAT-Tox (L)iver assay. Stress response elements chosen were the c-fos promot-e r, the gadd153 promoter, and p53 response element repeats. The gadd153-CAT assay showed the strongest correla-tion with activity in the Tg-AC assay.This correlation was improved further byadding the Zeta-Luc assay as a second-stage screen. Hence, efforts to augmentthe Tg.AC mouse model with other as-says appear promising.
Use of the transgenic mouse bioassayfor carcinogenicity has been criticized byadvocates of in vitro bioassays, however.For example, Van Zeller and Combes(1999) evaluated an interlaboratory c o l l a b o r a t ive study organized by the International Life Sciences Institute (ILSI) that included four transgenic carcinogenicity assays in mice (the c-Ha-ras, Tg.AC, P53+/- and XPA systems).They concluded that, “to date, the datasuggest that none of these assays is appropriate for inclusion in a carcino-genicity testing strategy”. They suggest-ed that, “more emphasis should be placedon developing replacement alternativeassays which are capable of identifyingand characterizing carcinogens of humanrelevance”. The Syrian hamster embryocell transformation assay was consideredto represent perhaps a more appropriateassay for evaluation of human carcino-genesis by these researchers.
Additional criticism has been directedat transgenic models for prion disease(Jenkins and Combes, 1999). Prion
diseases are fatal neurological disorderscaused by proteinaceous agents knownas prions. An example of this type of disease is that of Creutzfeldt Macombdisease in humans. These researchersfeel that the use of transgenic mice tostudy prions should be augmented by inv i t ro a l t e r n a t ives. Examples of these alternatives include cell-free conversionassays, cell culture systems, immunoas-says for the pathogenic form of the prionprotein, and perhaps yeast-based systemsbecause prion-like proteins have beenidentified in yeast.
These criticisms of transgenic animalmodels reflect a growing concern aboutgreater use of transgenic animal modelswithout appropriate consideration of invitro alternatives which may, in fact, beas predictive, as the transgenic models.In response to these and other animalw e l fare issues about transgenesis,Delpire et al. (1999) have proposed anew ethical scheme addressing the use oflaboratory animals for biomedical purposes. T h ey suggest that current European Union legislation relating tothe use of laboratory animals was im-plemented before transgenesis and its ap-plications were appropriately researched.They recommend that “a specific clusterof questions dealing with transgenic animals” could be incorporated at theEuropean level when reviews of projectproposals involving animal experimenta-tion occur.
What specifically can go wrong withtransgenic animals? Van Reenen andBlockhuis (1993, 1997) present threegeneral factors that can result in negativeeffects in transgenic animals: reproduc-tive and other biotechnological interven-tions, mutations, and expression of thetransgene. Metpham et al. (1998) discussthese factors in-depth. They suggest thatin vitro procedures employed both beforeand after microinjection can lead to in-creased gestation length, body weight,incidence of dystocia, and perinatal lossand anomalies as compared to in vivoprocedures. Furthermore, foreign DNAoften becomes integrated within or adja-cent to an endogenous gene creating anew mutation that has deleterious effectson the host. Finally, transgenic animalsmay express characteristics that areharmful to them. The degree of harm ex-

HU G G I N S
ALTEX 20, Suppl.1/0346
Tab. 15: Results of rapid carcinogenicity testing using rasH2 mice (from Yamamoto et al., 1998)
Chemicals tested Dose Route of Rapid tumor Tumor Malignant administration responsea incidenceb tumors
in Tg miceTg N o n - T g
Mutagenic (Salmonella) carionogensp-Cresidine 0.25 or 0.5% for 26 wk Feed +* Tg ≅ Non-Tg + +
Cyclophosphamidec 30mg/kg x 2/wk for 25 wk Gavage +/- Tg ≅ Non-Tg + -
DENc 90mg/kg x 1 Intraperitoneal +* Tg > Non-Tg* + -
1,2-Dimethylhydrazineg 20mg/kg x 1/wk for 20 wk Subcutaneous +* Tg > Non-Tg* + -
4HAQOe 10 or 20mg/kg x 1 Intravenous +* Tg > Non-Tg* ++ +
MAMc 20mg/kg x 1/wk for 6 wk Subcutaneous + Tg > Non-Tg + -
Melphalan 0.3 or 1.5mg/kg x 1/wk for 26 wk Intraperitoneal + Tg > Non-Tg + -
MNNGc 2.5mg x 1 Gavage +* Tg > Non-Tg* + -
MNUc 75mg/kg x 1 or 15mg/kg x 5 Intraperitoneal +* Tg > Non-Tg* + -
NNKe 3 or 6mg x 1/wk for 2 wk Intraperitoneal +* Tg < Non-Tg* + -
4NQOc 15mg/kg x 1 Subcutaneous +* Tg > Non-Tg* + -
Phenacetine 0.7 or 1.4% for 24 wk Feed +* Tg > Non-Tg* + -
Procarbazinee 6 or 12mg/kg x 3/wk for 24 wk Intraperitoneal + Tg > Non-Tg + -
4,4’-Thiodianilinee 2000 or 4000 ppm for 24 wk Feed +* Tg > Non-Tg* + -
Thiotepae 1 or 2mg/kg x 3/wk for 24 wk Intraperitoneal +* Tg > Non-Tg* + -
Vinyl carbamatef 60mg/kg x 1 Intraperitoneal +* Tg > Non-Tg* ++ +
4-Vinyl-1-cyclohexene diepoxidee 5 or 10mg x 5/wk for 24 wk Dermal +* Tg > Non-Tg* ++ +
Nonmutagenic (Salmonella) carcinogensBenzenee 50 or 100 mg/kg x 5/wk for 24 wk Gavage +* Tg > Non-Tg* + -
Cyclosporine 10 or 25 mg/kg x 5/wk for 24 wk Gavage + Tg > Non-Tg + -
1,4-Dioxanee 0.5 or 1% for 24 wk Water + Tg > Non-Tg* + -
Ethyl acrylatee 100 or 200 mg/kg x 5/wk for 24 wk Gavage +* Tg > Non-Tg* + -
Ethylene thiouread 0.3% for 28 wk Feed +* Tg ≅ Non-Tg + +
1,1,2-Trichloroethanee 100 or 200mg/kg Gavage -
Mutagenic (Salmonella) noncarcinogensp-Anisidine 0.225 or 0.45% for 26 wk Feed -
8-Hydroxyquinolinee 1500 or 3000 ppm for 24 wk Feed -
4-Nitro-o-phenylenediaminee 3750 or 7500 ppm for 24 wk Feed +/- Tg ≅ Non-Tg - -
Nonmutagenic (Salmonella) noncarcinogensResorcinol 225mg/kg x 5/wk for 26 wk Gavage -
Rotenonee 600 or 1200 ppm for 24 wk Feed -
Xylene (mixed) 500 or 1000 mg/kg x 5 /wk for 24 wkGavage
a Rapid tumor response means that tumors actually developed within 26 weeks, (28 weeks for ethylen thiourea) in rasH2 (Tg) mice inresponse to tested chemicals.
(+*) “Positive” indicates that the incidence of at least one type of tumor developed in chemical-treated Tg mice was significantly (p<0.05;Fisher´s exact test) higher than that in control vehicle-treated Tg mice.
(+) indicates that the incidence of at least one type of tumor developed in chemical-treated Tg mice was ≥25% but not statisticallysignificant versus control.
(+/-) “Positive-negative” indicates that the incidence of at least one type of tumor developed in chemical treated Tg mice was more than13% but less than 25%.
(-) “Negative” indicates that the incidence of at least one type of tumor developed in chemical treated Tg mice was ≤13% or that tumorincidence was similar between the vehicle-treated control and dosed groups.
b Tumor incidence: Incidence of at least one type of tumor (in cases of p-cresidine and ethylene thiourea, incidences of diffuse hyperplasiaof urinary bladder and thyroid follicular cell hyperplasia, respectively) developed in chemical-treated Tg mice was statistically differentfrom that of corresponding non-Tg mice (p<0.05; Fisher´s exact test)
References of the original publication c Data from Yamamoto et al (5). d Data from Yamamoto et al. (6). e Data from Urano et al. (7). f Data from Mitsumori et al (8). g Data from Itoh et al. (unpublished data).

HU G G I N S
ALTEX 20, Suppl.1/03 47
pressed was noted to be dependent on thebiological properties of the resulting pro-tein, the tissue in which transgenes areexpressed, the route of secretion of thegene product, and the level of transgeneexpression.
In direct contrast to concerns that theharmful effects of transgenic expressionbe avoided, are transgenic models of disease. These animals are used as toolsin which the harmful effects of trans-genic expression are actively sought. Assuch they represent an area of transgenicresearch in which the moral dilemma between animal welfare and biomedicaladvance can perhaps be most clearly observed.
That research involving transgenicmodels of disease is proliferating is certainly true (for example, Ware et al.,2000 (Bernard-Soulier syndrome); Shiraiet al., 2000 (prostate carcinogenesis);Bardeesy et al., 2000 (melanoma); G u y e r, 2000 (neurodeg e n e r a t ive dis-eases)). Unfortunately, the current focusof much of this research seems to be generation of a transgenic model withoutmuch consideration for the welfare of theanimals invo l ved. Metpham suggests,however, that “…it should be possible togenerate animals that have a number ofsalient features of the disease but do notdevelop the full disease condition”. Thissuggestion should be achievable in viewof the enhanced capability with whichg e n e t i c i s t ’s can accurately identifycausative genes in the disease process.Hardouin and Nagy (2000) in their re-view of mouse models for human diseasecite the fact that “In the last two decadesby the advent of novel and very efficientgenetic tools, the creation of diseasemodels is significantly advancing. Weh ave reached the stage where mouse geneticists feel there is no limitation increating phenocopies (or genocopies) ofa ny mutations or chromosomal aberra-tions identified in disease”. Hopefully,this capability will result in productionof more humane transgenic diseasem o d e l s .
More significant perhaps than the ef-fects of transgene expression of any kind,are the more general, but perhaps less obvious, effects of introduction of trans-genes into a host organism. Van der Meerand colleagues (1999), in an important
attempt to differentiate between the effects of the technique of transgenesisand the effects caused by the expressionof the transgene, studied behavioral responses and morphological develop-ment of newborn mice. Newborn animalswere subjected to various behav i o r a ltests and their growth and morphologicalcharacteristics were measured from birthup to the age of 3 weeks. The results indicated that the microinjected DNA-construct influenced the survival rateduring the first 2-3 days after birth. Theaverage loss of pups was about 10%, incontrast to the groups without the DNAconstruct, in which none of the pupsdied. Body weight gain was significantlylower for the DNA-construct pups, butonly during the first 11 days. No signifi-cant differences in morphological char-acteristics or behavioral deve l o p m e n twere noted. The authors suggested that,“Before general conclusions about theextent to which the technique of transge-nesis affects animal welfare can bedrawn, more transgenic lines should bestudied in this way.” Evaluations such asthis one should indeed be performed ifwe are to draw viable conclusions from da-ta generated from any transgenic animal.
6.4 Summary, conclusions, and future work
Genetic engineering offers tremendousopportunities for the advancement ofbiomedical research in many areas, including toxicology. The expression ofmetabolic enzymes in human cell culturesystems represents a major toxicologicaltool with which to study drug and/or toxicant metabolism and elicit a clearerunderstanding of many mechanisms ofaction. Current research trends are to-ward the isolation of the activity of singleenzymes in a metabolic profile followedby study of the activities of multiple enzymes acting in sequence as is the casein vivo. Considerable progress has beenmade thus far. However, caveats havebeen issued with regard to the extent towhich these systems can be reasonablycompared to in vivo situations. Appropri-ate validation of much of this work remains to be accomplished.
The generation and use of transgenicanimals to study questions of biomedicalinterest have been questioned by many in
view of the moral and ethical dilemmaspresented by these activities. That trans-genic animals may contribute to the reduction of animal use in toxicologicalexperiments, particularly studies of car-cinogenicity, is not disputed. However,a d vocates of replacement alternatives argue that in vitro alternatives to this typeof toxicity testing have not been givenadequate attention. Moreover, althoughtransgenic models of disease offer a potent method with which to study dis-ease conditions, animal welfare is oftennot a foremost consideration in many ofthese research endeavors.
Despite the controversy surroundinguse of transgenic animals, recent recom-mendations by the ICH have indicatedthat a transgenic model can be consid-ered as a replacement for one species in chronic carcinogenic evaluations ofpharmaceutical products. Several suchmodels are undergoing evaluation andbattery-type approaches incorporatingmore than one model have been suggest-ed. Many individuals however, remaincircumspect of the use of transgenic animals in general for humane reasons.Actions such as those in which a “specif-ic cluster of questions dealing with transgenic animals” is incorporated intoa ny rev i ew of research proposals inwhich transgenic models are used shouldbe encouraged.
M o r e ove r, the blatant disregard for animal welfare that is often inherent inany experimental rush to define the bio-chemical lesion resulting in a particulardisease should be discouraged. As suggested by Metpham above, animalmodels of disease should be designed sothat the salient features of the disease areemphasized and any unnecessary un-toward effects on the animal minimized.G iven the very sophisticated genetictools we have on hand these days, thisshould not be an insurmountable, or evendifficult, task.
ReferencesAnonymous (1998). Human cell growth
patented. http://www.wired.com/news/technology/0,1282,16730l,00.html
Arent, L. (1999). Lambs get humangenes. http://www.wired.com/technol-ogy/0,1282,20874,00.html

HU G G I N S
ALTEX 20, Suppl.1/0348
Bardeesy, N., Wong, K., DePinho, R. andChin, L. (2000). Animal models ofmelanoma: recent advances and future prospects. A d v. Cancer Res. 79, 123-156.
Contrera, J. and DeGeorge, J. (1998). Invivo transgenic bioassays and assess-ment of the carcinogenic potential ofpharmaceuticals. Environ. Hlth. Per-spect. 106(1), 71-80.
Crespi, C., Gonzalez, F., Steimel, D. etal. (1991). A metabolically competenthuman cell line expressing five com-plementary DNAs encoding procar-cinogen-activating enzymes: Applica-tion to mutagenicity testing. Chem.Res. Toxicol. 4, 566-572.
Crespi, C. (1995). Xenobiotic-metaboliz-ing human cells as tools for pharmaco-logical and toxicological research.Adv. Drug Res. 26, 180-235.
Crespi, C. and Penman, B. (1997). Use ofc D NA - expressed human cy t o c h r o m eP450 enzymes to study potential drug-drug interactions. Adv. Pharmacol. 43,171-188.
Crespi, C., Miller, V. and Penman, B.(1997). Microtiter plate assays for in-hibition of human, drug-metabolisingcytochromes P450. Anal. Bioch e m .248, 188-190.
Crespi, C. and Penman, B. (1998). Use ofhuman engineered cells in toxicology.Hum. Exp. Toxicol. 17(9), 485.
Delpire, V., Mepham, T. and Balls, M.(1999). A proposal for a new ethicalscheme addressing the use of laborato-ry animals for biomedical purposes.ATLA 27, 869-881.
Eastin, W. (1998). The U.S. NationalToxicology Program evaluation oftransgenic mice as predictive modelsfor identifying carcinogens. Environ.Hlth. Perspect. 106 (Suppl. 1), 81-84.
Grove, A., Llewellyn, G., Kessler, F. etal. (2000). Differential protection byrat UDP-glucuronosyltransferase 1A7against Benzo[a]pyrene-3,6- quinoneversus Benzo[a]pyrene-induced cyto-toxic effects in human lymphoblastoiccells. Toxicol. Appl. Pharmacol.162(1), 34-43.
Guyer, C. (2000). Recent developmentsin animal models of human neurode-generative diseases. Toxicol. Pathol.28(2), 363-366.
Hanna, I. H., Dawling, S., Roodi, N. et al.(2000). Cytochrome P450 1B1 (CYP1B1)
pharmacogenetics: association of poly-morphisms with functional diff e r e n c e sin estrogen hydroxylation activ i t y. C a n-cer Res. 60(13), 3440-3444.
Hardouin, S. and Nagy, N. (2000).Mouse models for human disease.Clin. Genet. 57, 237-244.
Hu, M., Li, Y., Davitt, C. et al. (1999).Transport and metabolic characteriza-tion of Caco-2 cells ex p r e s s i n gCYP3A4 and CYP3A4 plus oxidore-ductase. Pharm. Res. 16(9), 1352-1359.
ICH (International Conference on Har-monisation) Expert Safety Wo r k i n gGroup (1996). Testing for carcino-genicity of pharmaceuticals. FederalRegister 61, 43298-43300.
Jenkins, E. and Combes, R. (1999).Transgenic models for prion disease:Have they outlived their useful pur-pose? ATLA 27, 827-838.
Kranendonk, M., Carreira, F., Theisen, P.et al.(1999). Escherichia coli MTC, ahuman NADPH P450 reductase com-petent mutagenicity tester strain for theexpression of human cytochrome P450isoforms 1A1, 1A2, 2A6, 3A4, or 3A5:catalytic activities and mutagenicitystudies. Mutat. Res. 441(1), 73-83.
Metpham, T., Coombes, R., Balls, M. etal. (1998). The use of transgenic animalsin the European Union. The Report andRecommendations of ECVAM Wo r k-shop 28. ATLA 26, 21-43.
Monro, A. and MacDonald, J. (1998).Evaluation of the carcinogenic poten-tial of pharmaceuticals. Drug Safety18(5), 309-319.
Philipkoski, K. (2000). The future: partman, part beast. http://www. w i r e d . c o m/n ew s / t e c h n o l o g y / 0 , 1 2 8 2 , 4 0 5 1 2 , 0 0 . h t m l
Sawada, M. and Kamataki, T. (1998).Genetically engineered cells stably ex-pressing cytochrome P450 and theirapplication to mutagen assays. Mutat.Res. 411(1), 19-43.
Scott, D., Chacko, T., Maxwell, D. et al.(1999). Expression and partial purification of a recombinant secretoryform of human liver carboxylesterase. Protein Expr. Purif. 17(1), 16-25.
Shirai, T., Takahashi, S., Cui, L. et al.(2000). Experimental prostate carcino-genesis – rodent models. Mutat. Res.462(2-3), 219-226.
Thompson, K., Rosenzweig, B., Tsong,Y. and Sistare, F. (2000). Evaluation of
in vitro reporter gene induction assaysfor use in a rapid prescreen for com-pound selection to test specificity in theT g - AC mouse short-term carcinogenic-ity assay. Toxicol. Sci. 57(1), 43-53.
Van der Meer, M., Costa, P., Baumans,V., Olivier, B. and Van Zutphen, B.(1999). Welfare assessment of trans-genic animals: behavioural responsesand morphological development ofnewborn mice. ATLA 27, 857-868.
Van Reenen, C. and Blockhuis, H.(1993). Investigating welfare of dairycalves involved in genetic mode: prob-lems and perspectives. Livestock Pro-duction Science 36, 81-90.
Van Reenen, C. and Blockbuis, H.(1997). Evaluation of welfare of trans-genic farm animals: lessons from acase study in cattle. Journal of theRoyal Swedish Academy of Agricultureand Forestry. 136, 99-105.
Van Zeller, A. and Combes, R. (1999).Transgenic mouse bioassays for car-cinogenicity testing: A step in the rightdirection? ATLA 27, 839-846.
Ware, J., Russell, S. and Ruggeri, Z.(2000). Generation and rescue of amurine model of platelet dysfunction:the Bernard-Soulier syndrome. P ro c .Natl. Acad. Sci. USA 97(6), 2803-2808.
Waxman, D., Chen, L., Hecht, J. andJounaidi, Y. (1999). Cytochrome P450-based cancer gene therapy: recent ad-vances and future prospects. D r u gMetab. Rev. 31(2), 503-522.
Weibel, F., Andersson, T., Casciano, D. etal. (1997). Genetically engineered celllines: Characterisation and applica-tions in toxicity testing. The Reportand Recommendations of ECVA MWorkshop 26. ATLA 25, 625-639.
Yamamoto, S., Urano, K., Koizumi, H. etal. (1998). Validation of transgenicmice carrying the human prototype c-Ha-ras gene as a bioassay model forrapid carcinogenicity testing. Environ.Hlth. Perspect. 106 (Suppl. 1), 57-69.
Yanagawa, Y., Sawada, M., Deguchi, T.,Gonzalez, F. and Kamataki, T. (1994).Stable expression of human CYP1A2and N-acetyltransferases in Chinesehamster CHL cells: mutagenic activation of 2-amino-3-methylimida-zo[4,5-f]quinoline and 2-amino-3,8-d i m e t hylimidazo[ 4,5-f]quinoxaline.Cancer Res. 54, 3422-3427.

HU G G I N S
ALTEX 20, Suppl.1/03 49
7.1 BackgroundDNA microarrays, commonly referred toas “gene chips”, form the basis of a newfield of endeavor within toxicology,known as “toxicogenomics”. The majorissue to be addressed in toxicogenomicsis the relevance of the endpoint(s) measured to the toxicological insult(s)studied. Questions concerning the re-lationship between the genetic changesobserved on a microarray after exposureto a toxic chemical and the toxic insultproduced at the biochemical, cellular, ortissue level remain to be answered. Thistype of question however, haunts most, ifnot all, techniques which are purported tobe alternatives to the use of animals intoxicity testing. The answer is oftenfound through analysis of biochemicalmechanisms of toxic insult and oftensupported through exhaustive validationefforts that analyze the degree of correla-tion existing between in vitro alternativesand animal or human assays. In the caseof gene chips, this analysis will have tobe extended to analysis of genetic mech-anisms of toxic insult. However, if re-liable links between genetic changes andensuing toxic responses can be made,and these genetic changes can be faith-fully identified with gene chip techno-logy, then gene chips would indeed re-present an alternative method whichwould greatly reduce the number of, ifnot replace entirely, the animals used inthese experiments.
7.2 What is a gene chip?Gene chips have been defined with re-gard to both construction techniques andfunction. Ermolaeva et al. (1998) regard-ed gene chips as one of “several newtechnologies for studying the simultane-ous expression of large numbers ofgenes”. They divided these technologiesinto serial and parallel methods, and de-fined parallel approaches (such as micro-arrays) as those based on hybridizationof mRNA or DNA to cDNA immo-bilized on glass or to synthetic oligonu-cleotides immobilized on silica wafers or “chips”. Bowtell (1999) provides anexpanded definition. His microarray is a“complete system” composed of three
parts involving sample preparation (‘thefront end’), array generation and sampleanalysis (‘middleware’) and data han-dling and interpretation (the ‘back end’).The ‘front end’ requires preparation ofRNA from various sources, the ‘middle-ware’ is developed through target prepa-ration, labeling and construction of thearray, and the ‘back end’is characterizedby techniques for moving and handlingthe large quantities of data generated bythese array experiments.
B r own and Botstein (1999) provide adetailed description of the development ofc D NA microarrays. Arrays of discreteD NA sequences are printed on glass mi-croscope slides using a robotic “arrayer”and the relative abundance of each genesequence in two DNA or RNA samplesisolated from two different cell popula-
tions is compared. The two samples are labeled with different fluorescent dyes andthen mixed and hybridized with the ar-rayed DNA spots. Subsequently, flu o r e s-cence measurements are made with a mi-croscope that illuminates each DNA spotand measures fluorescence for each dyes e p a r a t e l y. These measurements are thenused to determine the ratio and, hence, ther e l a t ive abundance, of the sequence ofeach specific gene in the two mRNA orD NA samples. These researchers indicatethat although cDNA and oligonucleotidemicroarrays differ in detail, they share thesame “essential simplicity of ex p e r i m e n-tal design” which they attribute to thes p e c i ficity and affinity of complementarybase-pairing. Figure 13 contains a simpli-fied diagram of sample preparation andhybridization to cDNA microarrays.
7 Gene chip technology as an alternative to animal testing
Fig. 13: Simplified overview of the method for sample preparation and hybridizationto cDNA microarrays. For illustrative purposes, samples derived from cell culture are depicted, although othersample types are amenable to this analysis (from Nuwaysir et al., 1999).

HU G G I N S
ALTEX 20, Suppl.1/0350
Lockhart et al. (1996), while acknowl-edging the advantages of cDNA spottingonto chips, pointed out that in order tomonitor many genes, a large number ofcDNAs or PCR products must be pre-pared, purified, quantitated, catalogued,and spotted onto a solid support. Theydescribe development of an approachbased on hybridization to small, high-density arrays containing thousands ofsynthetic oligonucleotides. These arraysare designed based on sequence informa-tion and are synthesized in situ using acombination of photolithography andoligonucleotide chemistry. The advan-tages to this approach are that oligonu-cleotide array construction is “direct,combinatorial synthesis of appropriateoligonucleotides based on sequence information alone eliminat[ing] the necessity of preparing and handlingclones, PCR products, and cDNA s .” Furthermore, they suggest that becauseoligonucleotide probes for each gene arespecifically chosen and synthesized inknown locations on the arrays, hybridiza-tion patterns and intensities can be inter-preted with no additional sequencing orcharacterization.
Whether cDNA or oligonucleotide arrays, DNA-chip products are definitelybenefiting from improvements in fabrica-tion techniques that were developed initially for computer chip manufacture.As a result, organic structures (such assegments of DNA or mRNA) can now beapplied to a substrate of inorganic materials. Inorganic substrates for DNA-chips are usually glass or plastic wafers.H ow eve r, with the development of microassays that can be done on a chip,miniaturized glass tubes and reservoirsare also being utilized. Bonding of genetic sequences on a microchip usingphotolithographic processes as well asdeposition of sequences onto the chip using a droplet sprayer or a robot are currently the principal methods used tojoin organic structures with inorganicsubstrates in chip manufacture (Hencke,1998a).
Gene chips have also been classifiedby function (Hencke, 1998b). Three basic classes have been delineated. Thefirst type is a sequencing chip that servesas a template for analyzing unknownD NA sequences. The second type is
categorized as an expression chip. Theseare designed to determine the degree ofexpression of a certain genetic sequenceby measuring the rate or amount of mes-senger ribonucleic acid being producedby the target gene. The third type of chipis devoted to comparing the degree of genomic hybridization on the chip produced by different samples of DNA.
7.3 Application of gene chips to basic research
Perhaps the most extensive effort usingcDNA microarrays was the first pub-lished report of gene expression of an entire genome, that of the yeast, Saccha-romyces cerevisiae.
DeRisi et al. (1997) studied changes ingene expression during the shift fromanaerobic to aerobic metabolism occur-ring in this organism. Briefly, yeast openreading frames, amplified by polymerasechain reactions, were analyzed withDNA microarrays containing about 6400distinct DNA sequences printed ontoglass slides by a robotic printing device.Exponentially growing cells were harvested at 9 hours and at 2 hour inter-vals thereafter and mRNA was isolated ateach interval. Fluorescently-labeled cDNA was prepared by reverse transcrip-tion in the presence of green- and red-labeled deoxyuridine triphosphate(dUTP) and then hybridized to the microarrays. Subsequently, data from theseries of seven samples consisting ofmore than 43,000 expression-ratio measurements were organized into adatabase for analysis. These investigatorsfound that gene expression in glucose-rich medium, is remarkably stable. Veryfew changes in gene expression werenoted. How eve r, when glucose was depleted from the growth medium,marked changes in expression were not-ed (e.g. mRNA levels for approximately710 genes were induced by a factor of atleast 2, and the mRNA levels for approx-imately 1030 genes declined by a factorof approximately 2). The authors notedthat about half of these differentially expressed genes had no currently recog-nized function and were not, as of yet,named. T h ey concluded that DNA microarrays provided a simple and eco-nomic way to explore gene expressionpatterns on a genomic scale, indicating
that even simple experiments such as thisone, can yield vast amounts of data. Inaddition, they warned that perhaps thegreatest challenge lay not in the use ofmicroarrays, but in the effective organi-zation and analysis of the large amountsof data generated.
Oligonucleotide chips have also beenused to evaluate gene-expression patternsin nearly all open reading frames of S. cerevisiae during anaerobic and aero-bic metabolism (Wodicka et al., 1997).Cellular poly (A)+ mRNA was extractedfrom yeast cells grown in rich and mini-mal media, and hybridized to a set offour arrays that contained more than260,000 specifically chosen oligonu-cleotides synthesized using light-directedcombinatorial chemistry. More than 87%of all yeast mRNAs were detected inyeast cells grown in rich medium. Approximately 19% of all RNAs werepresent at less than 0.1 copies per cell,50% between 0.1 and 1 copy per cell,25% between 1 and 10 copies per celland 5% at more than 10 copies per cell.The overall distribution of mRNA levelswas very similar for cells grown in mini-mal medium. As in the study using cDNA arrays discussed above, a smallnumber of genes were identified withdramatically different expression levelsin rich and minimal media. The authorsconcluded that the combination of highlyparallel detection and monitoring meth-ods (such as oligonucleotide arrays), mutant strains, and use of genetic screensprovide a very powerful basis for investi-gation of yeast biochemistry. They envi-sioned research findings in yeast utilizingthese methods as contributing to agreater understanding of genetic mecha-nisms in more complex organisms.
N u waysir et al. (1999) have listed numerous additional examples of recentresearch utilizing either cDNA oroligonucleotide microarrays. cDNA microarray investigations include thoseof A rabidopsis thaliana R NA, yeastRNA, yeast genomic DNA, tumorigenicversus non-tumorigenic human tumorcell lines, human T-cells, and human inflammatory disease-related genes(Schena et al, 1995, 1996; DeRisi et al.,1996; Shalon et al., 1996; Lashkari et al.,1997; Heller et al., 1997). Alternatively,oligonucleotide microarrays have been
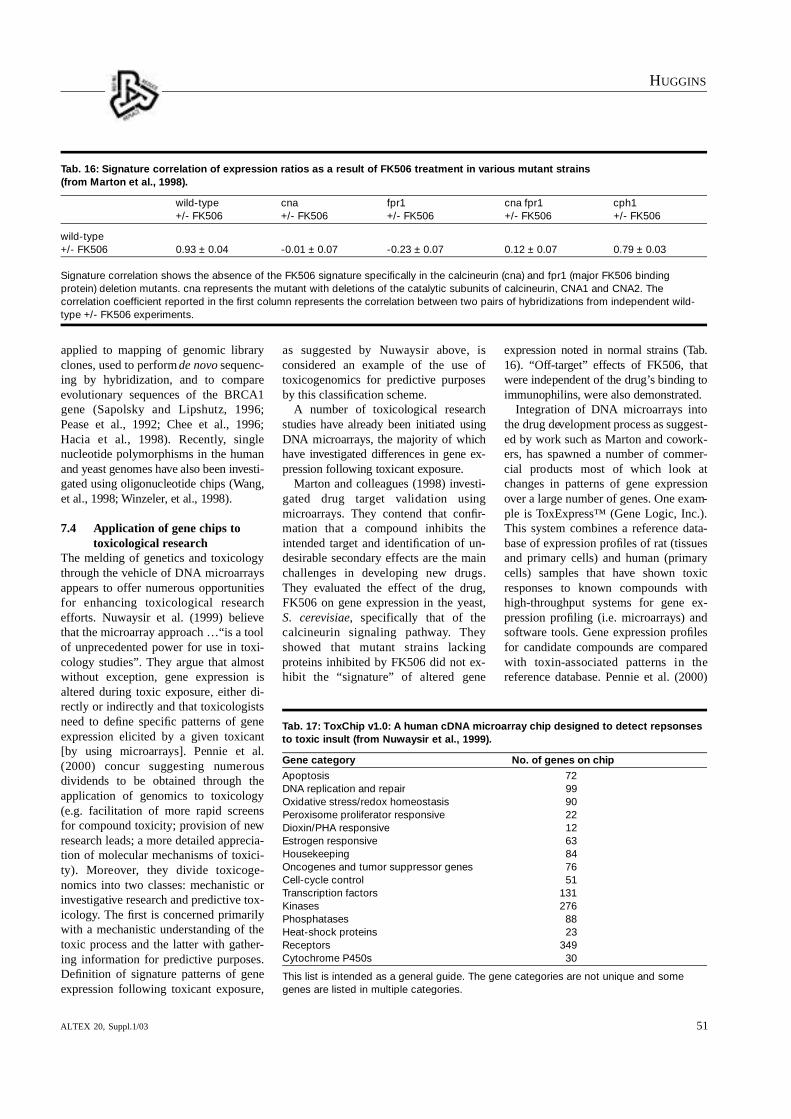
HU G G I N S
ALTEX 20, Suppl.1/03 51
applied to mapping of genomic libraryclones, used to perform de novo sequenc-ing by hybridization, and to compareevolutionary sequences of the BRCA1gene (Sapolsky and Lipshutz, 1996;Pease et al., 1992; Chee et al., 1996; Hacia et al., 1998). Recently, single nucleotide polymorphisms in the humanand yeast genomes have also been inve s t i-gated using oligonucleotide chips (Wa n g ,et al., 1998; Wi n z e l e r, et al., 1998).
7.4 Application of gene chips to toxicological research
The melding of genetics and toxicologythrough the vehicle of DNA microarraysappears to offer numerous opportunitiesfor enhancing toxicological research efforts. Nuwaysir et al. (1999) believethat the microarray approach …“is a toolof unprecedented power for use in toxi-cology studies”. They argue that almostwithout exception, gene expression is altered during toxic exposure, either di-rectly or indirectly and that toxicologistsneed to define specific patterns of geneexpression elicited by a given toxicant[by using microarrays]. Pennie et al.(2000) concur suggesting numerous dividends to be obtained through the application of genomics to toxicology(e.g. facilitation of more rapid screensfor compound toxicity; provision of newresearch leads; a more detailed apprecia-tion of molecular mechanisms of toxici-ty). Moreove r, they divide toxicoge-nomics into two classes: mechanistic orinvestigative research and predictive tox-icology. The first is concerned primarilywith a mechanistic understanding of thetoxic process and the latter with gather-ing information for predictive purposes.Definition of signature patterns of geneexpression following toxicant exposure,
as suggested by Nuwaysir above, is considered an example of the use of toxicogenomics for predictive purposesby this classification scheme.
A number of toxicological researchstudies have already been initiated usingD NA microarrays, the majority of whichh ave inve s t i gated differences in gene ex-pression following toxicant ex p o s u r e .
Marton and colleagues (1998) investi-gated drug target validation using microarrays. They contend that confir-mation that a compound inhibits the intended target and identification of un-desirable secondary effects are the mainchallenges in developing new drugs.They evaluated the effect of the drug,FK506 on gene expression in the yeast,S. cerev i s i a e, specifically that of the calcineurin signaling pathwa y. T h eys h owed that mutant strains lacking proteins inhibited by FK506 did not ex-hibit the “signature” of altered gene
expression noted in normal strains (Ta b.16). “Off - t a rget” effects of FK506, thatwere independent of the drug’s binding toimmunophilins, were also demonstrated.
Integration of DNA microarrays intothe drug development process as suggest-ed by work such as Marton and cowork-ers, has spawned a number of commer-cial products most of which look atchanges in patterns of gene expressionover a large number of genes. One exam-ple is ToxExpress™ (Gene Logic, Inc.).This system combines a reference data-base of expression profiles of rat (tissuesand primary cells) and human (primarycells) samples that have shown toxic responses to known compounds withhigh-throughput systems for gene ex-pression profiling (i.e. microarrays) andsoftware tools. Gene expression profilesfor candidate compounds are comparedwith toxin-associated patterns in the reference database. Pennie et al. (2000)
Tab. 16: Signature correlation of expression ratios as a result of FK506 treatment in various mutant strains (from Marton et al., 1998).
wild-type cna fpr1 cna fpr1 cph1+/- FK506 +/- FK506 +/- FK506 +/- FK506 +/- FK506
wild-type+/- FK506 0.93 ± 0.04 -0.01 ± 0.07 -0.23 ± 0.07 0.12 ± 0.07 0.79 ± 0.03
Signature correlation shows the absence of the FK506 signature specifically in the calcineurin (cna) and fpr1 (major FK506 bindingprotein) deletion mutants. cna represents the mutant with deletions of the catalytic subunits of calcineurin, CNA1 and CNA2. Thecorrelation coefficient reported in the first column represents the correlation between two pairs of hybridizations from independent wild-type +/- FK506 experiments.
Tab. 17: ToxChip v1.0: A human cDNA microarray chip designed to detect repsonsesto toxic insult (from Nuwaysir et al., 1999).
Gene category No. of genes on chip
Apoptosis 72DNA replication and repair 99Oxidative stress/redox homeostasis 90Peroxisome proliferator responsive 22Dioxin/PHA responsive 12Estrogen responsive 63Housekeeping 84Oncogenes and tumor suppressor genes 76Cell-cycle control 51Transcription factors 131Kinases 276Phosphatases 88Heat-shock proteins 23Receptors 349Cytochrome P450s 30
This list is intended as a general guide. The gene categories are not unique and somegenes are listed in multiple categories.

HU G G I N S
ALTEX 20, Suppl.1/0352
h ave referred to this comparison ofchanges in gene expression with refer-ence profiles as the “pattern recognition”approach to toxicology.
Comparison to a reference set of “toxicant signature” gene patterns is also a major feature of ToxChip v1.0(Nuwaysir et al., 1999). 2900 humangenes compose ToxChip v1.0 includingD NA replication and repair genes, apoptosis genes, and genes responsiveto PAHs and dioxin-like compounds,peroxisome proliferators, estrogeniccompounds, and oxidant stress (Tab. 17).
Transcription factors, oncogenes, tumor suppressor genes, cyclins, kinases,phosphatases, cell adhesion and motilitygenes, and homeobox genes are alsopresent. 84 “housekeeping” genes thatare used to normalize signal are on thechip as well. A hypothetical toxicity pro-tocol using ToxChip v1.0 is as follows.Dose- and time-course parameters wouldbe established for a series of toxicantswithin a given prototypic class (e.g. poly-cyclic aromatic hydrocarbons (PA H s ) .Cells would then be treated with theseagents at a fixed toxicity level (as meas-ured by cell survival), RNA would beh a r vested, and toxicant-induced gene
expression changes would be evaluatedby hybridization to ToxChip v1.0. Whenthe “toxicant signature” is determined,the genes within this signature areflagged and subsequent experiments withchemicals of unknown action rely oncomparison to these patterns establishedfor each prototypic class of chemicals.
Another approach to assessing toxicantexposure through changes in gene ex-pression utilizes smaller microarrays, upon which selected gene sequenceshave been placed (e.g. ToxBlot arrays)(Pennie et al., 2000). ToxBlot arrays ofboth human and mouse genes have beenconstructed which contain, in each case,about 2400 cDNA sequences, represent-ing about 600 genes. Broad gene classesrepresenting cancer, immunology, en-docrinology and neurobiology, as well assequences thought to be useful to inves-t i ga t ive toxicology, predevelopment toxicology, and safety assessment are integrated into these arrays (Tab. 18).
A database on gene function, distribu-tion, and allelic variations for each generepresented has also been established.Research inve s t i gations of toxicant action on several target organ systemsh ave been performed using To x B l o t
arrays. Treatment of cultured cells withnatural, synthetic, and phytoestrogens atseveral doses and time points resulted ina pattern of consistent gene expressionchanges. Similarly, hepatic toxicity following exposure of HepG2 cells toethanol, paracetamol, hydrogen peroxideand carbon tetrachloride was character-ized by transcript profiling at selecteddoses for all four compounds. Consistentchanges in gene expression were ob-served here also.
7.5 Development of gene chips as alternatives to animal testing
The relationship of gene chips to alterna-t ives research can perhaps be best described as one of tantalising potential.As indicated by several of the researcherswhose work was described above, DNAmicroarrays can be a very powerful tool,particularly when used to link changes in gene expression with changes in bio-chemical processes. That this tool can beused effectively in toxicological investi-gations is evident from work monitoringchanges in gene expression following exposure to toxicants. However, how cangene chips contribute to the reduction or replacement of animals in medical ex-
Tab. 18: Broad gene classes included on the ToxBlot Microarrays (from Pennie et al., 2000)
Cancer Immunology Endocrinoloy and Investigative Predevelopment Safety neurobiology toxicology toxicology assessment
Apolipoprotein genes Basic transcription Basic transcription Acetyl CoA pathway Bcl/Bax family Bcl/Bax familyfactors factors
Basic transcription Cell adhesion CYP Bcl/Bax family CYP CYPfactors molecules
Bcl/Bax familiy Cell surface receptors Drug metabolism Drug metabolism Acetyl CoA pathway Drug metabolism
CYP genes Chemokines Extracellular matrix Immediate early genes Ion channels GST
Caspases Extracellular matrix GABA receptors/ GST GST Heat shock transport proteins
cdc/cdk s Heat shock proteins Heat shock proteins CYP Histones Liver acute-phase markers
Cyclins Interleukins Ion channels Basic transcription Heat shock proteins Markers for factors GI tract physiology
GST Metalloproteinases Neurotransmitter- Steroid hormone Steroid regulated Oxidative-stress metabolising enzymes receptors genes markers
Heat shock proteins Neurotrophic factors/ Steroid regulated receptors genes
Immediate early genes Peptide hormones Thyroid hyperplasiamarkers
Interleukins Steroidogenesis/aromatase
Matrix metalloproteins Steroid hormone receptors
Steroid hormone receptors

HU G G I N S
ALTEX 20, Suppl.1/03 53
perimentation? The answer(s) lies in theintegration of gene chips effectively intoprescreening protocols for unknow ncompounds, in the coupling of genechips with cell and tissue culture models,and, in a better understanding of the genetic effects of toxicant insult.
Nuwaysir et al. (1999) emphasizes thatalthough the field of toxicology usesmany in vivo model systems, includingrats, mice and rabbits, to evaluate potential toxicity, a variety of in vitrotechniques already exist to measure toxi-city, many of which measure toxicant-induced DNA damage (e.g. Ames test,Syrian hamster embryo cell transforma-tion assay, micronucleus assays, un-scheduled DNA synthesis assays). Allthese methods are predicated upon alter-ations in gene expression. Hence, genechips are yet another addition to, if not areplacement for, this collection of in vitro assays. Assays that measure DNAdamage are already an integral part ofmost toxicology prescreening programs.The gene chip pattern recognition ap-proach could also most definitely be incorporated into many prescreening batteries for new compound development(e.g. new drug development).
Pennie et al. (2000) suggest that thethroughput requirements of patternrecognition approaches using gene chipswill…“almost certainly necessitate em-ploying in vitro culture systems.” Theirwork (discussed above) did just that, em-ploying both estrogenic and hepatic cellsin culture. However, they caution, that although practical advantages to cell cultures, exist, there are major disadvan-tages. Compound-induced changes intranscription many not necessarily reflectaccurately the response of the correspon-ding organ in vivo and availability of celllines may be limited (particularly ifmechanistic endpoints are sought). Alsoof note is the fact that in vitro modelingis unlikely to be an effective replacementfor animals when species, strain, sex, orroute of administration are important factors in development of the toxic re-sponse.
Despite the limitations listed above,much recent research has combined cellcultures with DNA microarrays. Holdenand cowo r kers (1999) have analyzed altered hepatocyte gene expression fol-
lowing exposure to carbon tetrachlorideby using human hepatoma cells (HepG2cell line). A total of 580 genes were ana-lyzed; 40 were found to exhibit modulat-ed expression after exposure to CCl4
(Tab. 19).Rodi et al. (1999) evaluated the effect
of phenobarbital in rodent hepatocytes.More than 300 genes were found to ex-hibit modulated expression after this exposure. Mouse liver cells have beenused to evaluate changes in expression ofcytochrome P4501A1 and 1A2 genes af-ter beta-naphthoflavone exposure (Barto-siewicz et al., 2000). Upregulation ofCYP1A2 was noted at doses lower thanthose detected using Northern blottingtechniques. A human bladder epithelialcell line has been used to assess the ef-fects of arsenite on cell proliferation(Simeonova et al., 2000). Expression ofgenes associated with cell growth (e.g. c-fos, c-jun, and EGR-1) and cell arrest(e. g. GADD153 and GADD45) wasmodulated. Furthermore, the National Institutes of Environmental Health Sci-ences (NIEHS) is currently solicitinggrant proposals for development of alter-natives to animals for toxicity testingspecifically those which combine cellculture and microarray technology(NIEHS, 2000). Proposals are to includemeans by which “it is possible to assessgene expression in cells in culture andcells in vivo and to then devise methodsto make the cell cultures mimic the in vivo situation at least with regard to geneexpression.” Hence, the combination ofcell culture and DNA microarray tech-nologies is being considered seriously as
a method by which replacement alterna-tives to animal testing can be derived.
Although utilizing microarrays as pre-screens of toxicant action can easily beconsidered a useful reduction alternative,establishing the relationship betweenchanges in gene expression and resultingtoxic response may be key to acceptanceof assays utilizing microarrays as re-placement alternatives. Pennie et al.(2000), however, warn that establishingthis linkage can be fraught with difficul-ties and may require assistance from proteomic technology in order to exam-ine parallels between gene and proteinexpression modulation after toxic expo-sure. They suggest that microarray ex-periments pose a question of “certainty”(i.e. are changes in gene expression ‘real’, and if so, does altered gene ex-pression translate into protein produc-tion?). They also argue that changes ingene expression that do not result in al-tered protein production are of relevanceand should not be dismissed. Indeed,they challenge us to adopt a broader viewof biological relevance, one that acceptsgreater intellectual discomfort, in orderto embrace a more holistic perspective ofbiological change. In essence, eve n t stranspiring on a gene chip may not a lways be reduced/refined to discretebiochemical pathways. T h ey should nevertheless, be considered valuable.
7.6 Current status of gene chip technology
H e n c ke (1999) summarizes the rapidprogress of chip technology as occurringin four areas: sensitivity, microfabrica-
Tab. 19: Selected genes found to be differentially expressed in HepG2 cells followingtreatment with CCl4 for 8 hours. Results given as fold induction relative to the DMFcontrol from a single experiment (from Holden et al., 1999).
Gene Description Induction (fold)
IL-8 Chemotactic factor/neutrophil activation +7
IL-1ß Immunoregulatory cytokine +3.8
c-fms/csf-1 Tyrosine-protein kinase -12.8
p55CDC Cell division control +4
STAT6 IL-4 signalling -5.5
STK2 Serine/threonine-protein kinase +7
RAB-6 Ras-related protein +8
Apopain/Caspase-3 Executes apoptosis +4
Bcl-X/Bax Accelerates apoptosis +6.6

HU G G I N S
ALTEX 20, Suppl.1/0354
tion, density, and integration. Increasedsensitivity is being sought through betterlabeling technologies such as time-r e s o l ved fluorescence, dendrimers, iodine radioisotopes, and chemical locks.A balance will need be struck, however,between the complexity of the testingprocedures and the sensitivity of thetests. Microfabrication of chips is alreadyin existence as indicated by the presenceof pico- and microliter-sized compart-ments; nanoscale devices are now alsounder construction. Increasing the densi-ty of the arrays packed onto DNA chipsis an objective that is rapidly beingreached. Single-chip arrays may nowcontain 20.000 wells of 100 nl volume;n ewer technology promises to makeavailable single-molecular layer DNAbound to thin films of gold. An integrat-ed chip, also known as a “lab-on-a-chip”seems to be where much work in chiptechnology remains to be done. Thesechips integrate sample preparation andanalysis, reducing the “complete system”as designated by Botwell above, to twoor fewer components. Improved mi-crofluidic capabilities to capitalize on themultiple uses of capillary electrophoresisas well as on-chip chemiluminescence,and electrochemiluminescence assaysare being honed for integration into chipmanufacture.
Many of the research possibilities forgene chips have yet to be explored andnovel ideas for their use continue tospring forth. How eve r, major hurdlescurrently face manufacturers and users ofthe chips, the primary example of whichis quality.
Government regulators and users willhave to be guaranteed that the chips areof the highest quality possible. Becausethe effective manufacture of DNA mi-croarrays is still undergoing deve l o p-ment/refinement, quality control issuescontinue to require re-definition. Severallarge companies have launched a groupcalled the Genetic Analysis TechnologyConsortium (GATC) to evaluate these issues. This group intends to standardizearray-based genetic analysis, “paving theway for more-affordable and productivedevelopment of diagnostic and therapeu-tic products” (Hencke, 1999).
In addition to issues regarding genechip quality, concerns surrounding the
economics of gene chip use need to beaddressed. A new technology may p r ovide unique solutions to current problems, but if it’s not affordable, noone will use it. Marketing gene chips as aless expensive option to current diagnos-tic procedures is probably the only wayin which they will be accepted in the cur-rent marketplace. Unfortunately, makinggene chip manufacture less costly willrequire the development of cost-effectivetechniques for both information manage-ment and sample preparation compo-nents considering the large quantities ofdata requiring analysis produced fromthese experiments. This will not be aneasy task.
7.7 Summary, conclusions, and future work
In summary, gene chips (DNA micro-arrays) represent a technology that hasalready opened many doors in basic genomic research. Moreover, their valueto both investigative and discovery toxi-cology is becoming much more apparentas more toxicology experiments are con-ducted using them. Use of microarrays asreduction or replacement alternatives toanimal testing also holds great promise,particularly when they are used as com-ponents of prescreening batteries, andwhen coupled to cell culture techniques.
In the next five years research effortsdirected toward use of “toxicant signa-tures” and pattern recognition approach-es to develop reproducible screeningtechniques for structurally similar groupsof chemicals will probably be the mostfruitful. This work may also be extendedto include groups of genes involved inparticular target organ toxicity. Eitheroption will likely produce a method foruse as a reduction alternative that can bei n t egrated into prescreening batteries.General changes in gene expression willbe monitored and reproducible patternswill be sought. In vitro assays in which a mechanistic relationship betweenchanges in gene expression and toxicantinsult is well defined should be validatedin the next ten years. Regulatory accept-ance of this type of assay should occurduring that time as well. In the nexttwenty years, many of the genes affectedby toxicant action should be identifiedsupporting efforts focused on under-
standing mechanisms of action. Whenaccurate parallels can be drawn betweenchanges in gene expression and exposureto toxic chemicals, gene chips will, without a doubt, function admirably asreplacements for animals in toxicity testing.
ReferencesBartosiewicz, M., Trounstine, M., Bark-
er, D. et al. (2000). Development of atoxicological gene array and quantita-t ive assessment of this technology.Arch. Biochem. Biophys. 376(1), 66-73.
Bowtell, D. (1999). Options available:From start to finish – for obtaining ex-pression data by microarray. NatureGenetics (Supplement) 21, 25-32.
Brown, P. and Botstein, D. (1999). Ex-ploring the new world of the genomewith DNA microarrays. Nature Genet-ics (Supplement) 21, 33-37.
Chee, M., Yang, R., Hubbell, E. et al.(1996). Accessing genetic informationwith high-density DNA arrays. Science274, 610-614.
DeRisi, J., Penland, L., Brown, P. et al.(1996). Use of a cDNA microarray toanalyse gene expression patterns in hu-man cancer. Nature Genetics 14, 457-460.
DeRisi, J., Iyer, V. and Brown, P. (1997).Exploring the metabolic and geneticcontrol of gene expression on a ge-nomic scale. Science 278, 680-686.
Ermolaeva, O., Rastogi, M., Pruitt, E. etal. (1998). Data management andanalysis for gene expression arrays.Nature Genetics 20, 19-23.
Gene Logic, Inc. (2000). http://www. g e n e -logic.com/products/white-tox.pdf
Hacia, J., Makalowski, W., Edgemon, K.et al. (1998). Evolutionary sequencecomparisons using high-density oli-ognucleotide arrays. Nature Genetics18(2), 155-158.
Heller, R., Schena, M., Chai. A. et al.(1997). Discovery and analysis of inflammatory disease-related genes us-ing cDNA microarrays. P roc. Natl.Acad. Sci. USA 94 , 2150-2155.
Hencke, C. (1998a). DNA-chip technolo-gies. Part 1: Research fundamentalsand industry catalysts. IVD Technolo-gy.

HU G G I N S
ALTEX 20, Suppl.1/03 55
H e n c ke, C. (1998b). DNA-chip tech-nologies. Part 2: State-of-the art andcompeting technologies. IVD Technol-ogy.
Hencke, C. (1999). DNA-chip technolo-gies. Part 3: What does the futurehold? IVD Technology.
Holden, P., James, N., Brooks, A. et al.(1999). Analysis of altered hepatocytegene expression induced by carbontetrachloride (CCl4) using microarraytechnology. Hum. Exp. Toxicol. 18(8) ,522.
Lashkari, D., DeRisi, J., McCusker, J. etal. (1997). Yeast microarrays forgenome wide parallel genetic and geneexpression analysis. Proc. Natl. Acad.Sci. USA 94, 13057-13062.
Lockhart, D., Dong, H., Byrne, M. et al.(1996). Expression monitoring by hy-bridization to high-density oligonu-cleotide arrays. Nature Biotechnology14, 1675-1680.
Marton, M., DeRisi, J., Bennett, H. et al.(1998). Drug target validation andidentification of secondary drug targete ffects using DANN microarrays. Nature Medicine 4(11), 1293-1301.
NIEHS (National Institute for Environ-mental Health Sciences) (2000). Solic-itation of the Public Health Service forSmall Business Innovation ResearchContract Proposals. PHS 2001-1.
National Institute for EnvironmentalHealth Sciences (NIEHS). Section X.Research Topics: 079 Development ofAlternatives to Animals for ToxicityTesting. p. 37.
Nuwaysir, E. F., Bittner, M., Trent, J. etal. (1999). Microarrays and toxicolo-gy: The advent of toxicogenomics.Molecular Carc i n ogenesis 24, 153-159.
Pease, A., Solas, D. and Fodor, S. (1992).Parallel synthesis of spatially address-able oliognucleotide probe matrices.Abstract. American Chemical Society.203, 34.
Pennie, W. D., Tugwood, J., Oliver, G.and Kimber, I. (2000). The principlesand practice of toxicogenomics: appli-cations and opportunities. Toxicologi-cal Sciences 54, 277-283.
Rodi, C., Bunch, R., Curtiss, S. et al.(1999). Revolution through genomicsin investigative and discovery toxicol-ogy. Toxicol. Pathol. 27, 107-110.
S a p o l s ky, R. and Lipshutz, R.(1996).Mapping genomic library clones usingoligonucleotide arrays. Genomics 33,445-456.
Schena, M., Shalon, D., Davis, R. andBrown, P. (1995). Quantitative moni-toring of gene-expression patterns witha complementary DNA microarray.Science 270, 467-470.
Schena, M., Shalon, D., Heller, R. et al.(1996). Parallel human genome analy-sis: Microarray-based expression monitoring of 1000 genes. Proc. Natl.Acad. Sci. USA 93, 10614-10619.
Shalon, D., Smith, S. and Brown, P.(1996). A DNA microarray system foranalyzing complex DNA samples using two-color fluorescent probe hybridization. Genome Research 6,639-645.
Simeonova, P., Wang, S., Toriuma, W. etal. (2000). Arsenic mediates cell pro-liferation and gene expression in thebladder epithelium: association witha c t ivating protein-1 transactiva t i o n .Cancer Res. 60(13), 3445-3453.
Wang, D., Fan, J., Siao, C. et al. (1998).L a rge-scale identification, mapping,and genotyping of single-nucleotidepolymorphisms in the human genome.Science 280, 1077-1082.
Winzeler, E., Richards, D., Conway, A. etal. (1998). Direct allelic va r i a t i o nscanning of the yeast genome. Science281, 1194-1197.
Wodicka, L., Dong, H., Mittmann, M. etal. (1997). Genome-wide expressionmonitoring in Saccharomyces cerevisi-ae. Nature Biotechnology 15, 1359-1367.
8 Validation of alternative methodology
8.1 BackgroundIn an early article about cell culture alternatives to animal testing, Goldberg(1986) wrote that, “The need is stressedfor adequate standardization and valida-tion of CCS (cell culture systems), andmeticulous conduct of tests using suchsystems.“ Mehlman et al. (1989) expand-ed these sentiments in a later report onmethods to reduce, refine and replace animal testing in industrial laboratories.They stated that, “There is a pressingneed for the systematic and scientificallysound validation of non-animal alterna-tives techniques to reduce the use of animals in toxicology testing while satis-fying requirements for the protection of
public safety”. Both statements reflectthe fact that validation of test methodshas been recognized early on as essentialto the development and subsequent acceptance of any alternative method bymany researchers in this area.
Gad (1990) subsequently wrote that as c i e n t i fic consensus had been formed on the requirements and process for vali-dation (of alternative methods). Althoughthis consensus might be considered debatable at times, serious attempts atstandardizing the requirements of valida-tion and the process by which it can beachieved have been performed (Balls etal., 1990; Balls et al., 1995a; Balls et al.,1995b). Moreove r, a research fa c i l i t y
specifically designed to investigate thevalidation of alternative methods, the European Center for the Validation of A l t e r n a t ive Methods (ECVAM), hasbeen established. Figure 14 contains a diagram of a proposed va l i d a t i o nprocess.
Furthermore, Goldberg et al. (1993)suggested that, “A framework capable offostering the validation of new methodsis essential for the effective transfer ofn ew technology from the research laboratory to practical use”. An exampleof part of this framework in place in theUnited States is the recent establishmentof the Interagency Coordinating Com-mittee on the Validation of Alternative

HU G G I N S
ALTEX 20, Suppl.1/0356
Methods (ICCVAM) working in concertwith the National Toxicology ProgramInteragency Center for the Evaluation ofA l t e r n a t ive Toxicological Methods(NICEATM). This type of linkage is alsoa strong indication that regulatory bodieshave recognized the validation process asintegral to effective development of a viable alternative method.
8.2 What is validation and whyhas it failed?
Definitions of validation vary. However,the following excerpts represent funda-mental descriptions of validation as theyapply to alternatives to animal testing.Balls et al. (1990) state that, “Validationis the process by which the reliability andrelevance of a procedure are establishedfor a specific purpose”. Moreover, a laterperspective indicates that, irrespective ofpurpose, or the type of validation study,the key factor is that it is of a scientific,rather than a political, nature. (Balls etal., 1995a). This latter view emphasizesan important distinction that is necessary,although often overlooked, for the main-tenance of objectivity in a regulatory are-na that often exhibits strongly politicalovertones.
Despite the realization that validationis an important part of the developmentand ultimate regulatory acceptance of analternative method, numerous problemshave arisen with the appropriate conductof validation studies. Balls and cowork-ers (1995a) compiled the following listof notable general deficits:
• The goals of the validation study werenot sufficiently well defined, and therewere differences of opinion as to whatthe objectives should be, and how bestto achieve them.
• Many of the studies were poorly de-signed and planned.
• The responsibilities of the participantshave not been clearly defined and/orfulfilled.
• The validation study has been inade-quately managed due to, for example,funding and/or communication prob-lems.
• Standard protocols defining all aspectsof the test procedure have been lack-ing, or have not been strictly adhered toby some of the participating laborato-ries.
Confirm Alternative Method isDeveloped Sufficiently to Allowan Assessment of Reliabilityand Relevance • Evidence of Relevance
• Adequate Protocols and SOPs• Evidence of Reliability
• Adequate Prediction Model
Assay Inception:Initial Design andDevelopment (Empiricalor Mechanistic Basis)
Optimise Assay and/orDevelop New PredictionModel
Value in optimising
assay?
• Design Validation Study• Assemble Reference Set of
Test Substances
• Assess Quality of In VivoData
• Conduct the Validation Study• Collect Data
• Analyse Data
Assess Relevance of AlternativeMethod• Best theoretical performance
• Performance relative to in vivo
• Mechanistic Basis• Other factors
Assess Reproducibility of TestResults
Assess Reproducibility ofPredictions from Alternative
Method
Is AlternativeMethod
Reliable?
Is AlternativeMethod
Relevant?
Abandon Assay
Use Method Seek Acceptance
Validation Study
No
No
No
Yes
Yes
Yes
Fig. 14: Validation process (from Bruner et al., 1996).

HU G G I N S
ALTEX 20, Suppl.1/03 57
• Relevant test chemicals, for which highquality and unambiguous in vivo (ani-mal and/or human) toxicity data wereavailable, could not be identified or ob-tained.
• Participating laboratories have failed tomeet the deadlines set for completionof the testing, submission of data, andother critical stages of the study.
• Participants in the validation studyhave been unable to agree on how toevaluate the data or on the interpreta-tion of the test results.
• Laboratories have committed them-selves to participating in a validationstudy, but have subsequently droppedout once the study was under way, forfinancial or logistical reasons.Furthermore, reasons for the failure of
s everal specific validation studies of alternatives to eye irritation testing haverecently been investigated (Balls et al.,1999). The following conclusions weredrawn:• The in vivo test to which most of in
vitro alternatives were compared pro-vided highly variable results due to a subjective scoring system
• Many non-animal protocols were inad-equate
• The choice of test substances was notrigorously defined
• The choice of statistical methods fordata analysis was not as robust as itcould have beenAlthough several of the conclusions
drawn from this investigation were simi-lar to the more general ones listed above,it should be noted that most in vivo mod-els of toxicity testing do not exhibit asmuch variability as that observed withthe Draize eye irritation test. Therefore,this particular validation problem mayapply only to attempts to validate eye ir-ritation alternatives.
As judged from these investigations,the in vitro assay undergoing validationmay fail due to causes that have less todo with the actual value of the assay thanwith the conduct of the validation study.
In an additional critique of the valida-tion process, Koeter (1995) wrote thatvalidation is a highly charged conceptunderstood as necessary by many, butunable to produce more widespread acceptance of alternative methods. Hecited two reasons for this failure: “First,
the results of the validation studies mayhave been unsatisfactory, which couldmean that either the method subjected tovalidation failed to show the desired relevance and reliability, or the validationstudy as such yielded inconclusive results. Secondly, despite clear-cut (sup-porting) results from the validation exer-cise, toxicologists/regulators appear re-luctant actually to use the data providedfor hazard and risk assessment proce-dures because of a lack of confidencewith the (types of) endpoints of the newtest.” He considered the latter reason tobe the “major hurdle” to the acceptanceof alternative methods. However, his firstreason, in synchrony with the views ofother researchers, emphasizes the needfor adequate conduct of validation stud-ies.
8.3 Improving the validation process
General remedies have been advancedfor improving the validation process(Balls et al., 1995a). Suggestions in-clude:
• A clear and unequivocal statement ofwhat the validation study is designed toaccomplish
• A well-defined plan for the study• A sufficiently large set of test sub-
stances covering the relevant chemicalclasses, the different categories of cos-metic ingredients, and the range of tox-ic endpoints to be evaluated
• In vivo data of high quality on all thetest substances to be used
• Evidence that the methods to be evalu-ated are scientifically-sound, relevant,reproducible, and that they have thepotential to replace the animal test inquestion
• An optimized protocol for each test,with any necessary standard operatingprocedures
• A clear description of how each alter-native method can be used to predict anin vivo endpoint
• Agreed statistical procedures for test-ing whether the methods can predictthe in vivo endpoints defined by theirdevelopers
• Agreed criteria to be met in order tos h ow that an alternative method could successfully and safely replacean animal test
• Results which meet these criteriaM o r e ove r, specific suggestions for
enhancing the validation of eye irritationalternatives have been made (Balls et al.,1999). These included the following:
• A new approach to the validation of in vitro tests for eye irritancy, based onthe use of reference standards
• The use of stepwise testing strategieswhich reduce and refine the use of ani-mals in eye irritation testing
• The use of multivariate and other sta-tistical techniques for the further analy-sis of data generated in previous vali-dation studies
• A program of research aimed at under-standing the underlying mechanisms ofeye irritationHence, ways to improve the validation
process in general as well as specificsuggestions aimed at a particular type ofvalidation study, that of the eye irritationalternative, have been put forth.
In addition, perhaps one of the mostimportant concepts suggested as a meansof improving validation efforts has beenthat of “prevalidation”. Prevalidation wasproposed to: (1) standardize test proce-dures into a formal protocol; (2) establishmultiple laboratories competent in per-forming the test method in line withGood Laboratory Practice (GLP) princi-ples; (3) develop the test’s predictionmodel (the algorithm which converts invitro data to a predicted in vivo hazard)(Fig. 15); and (4) supply independentevaluation of the performance of the test protocol and the prediction model(Curren and Southee, 2000).
Curren et al. (1995) defined prevalida-tion as consisting of three phases. PhaseI was defined as Protocol Developmentin which a GLP-compliant protocol andStandard Operating Procedures (SOPs)would be created, and determination ofintralaboratory reproducibility would beaccomplished. If successful, Phase I a c t ivities would then be followed byPhase II, Protocol Tr a n s f e r. ProtocolTransfer would consist of transfer of themethod to a second laboratory using thedefined protocol and SOPs, determina-tion of interlaboratory reproducibility,further refinement of the protocol ifneeded, submission of the optimized protocol to a documentation source, e.g.INVITTOX, and evaluation of suitability

HU G G I N S
ALTEX 20, Suppl.1/0358
for progression to Phase III. Phase III,Protocol Performance, would requiretransfer of the method to a third labora-tory, testing of coded materials in at leastthree laboratories, preparation of a reporton the performance of the method, andc o n firmation or redefinition of the prediction model.
The concept of prevalidation has beenevaluated by applying it “in practice” tos everal in vitro assays including the embryonic stem cell (EST) assay, thebovine corneal opacity and permeability(BCOP) assay, the fluorescein leakagetest, the EpiDerm™ Skin Corrosiv i t ytest, and the EpiDerm™ phototoxicitytest. Recent applications have also included the transcutaneous electrical resistance (TER) and Episkin™ assaysas well as the 3T3 NRU assay. Applica-tion of prevalidation procedures to evaluation of these assays was success-ful; several of them have been approvedas scientifically valid by the ECVAMScientific Advisory Committee (ESAC)(Balls and Corcelle, 1998a; Balls andCorcelle, 1998b; Balls and Corcelle,1998c). Curren and Southee advise, how-eve r, that eff e c t ive communication iscritical to successful prevalidation effortsas is the “spirit of cooperation”. Theynote that one of the most persistent
problems in these prevalidation efforts,as with full-blown validation attempts,was obtaining an adequate number of testchemicals with acceptable in vivo datawith which to conduct a prevalidation exercise.
A specific area of both prevalidationand validation studies which has alsocome under close scrutiny is that of thebiostatistical techniques used to drawin vitro/in vivo correlations. These tech-niques represent one of the most heavilyweighted factors used in determiningwhether sufficient correlation exists between in vitro and in vivo methods tojustify saying that the in vitro method isp r e d i c t ive of in vivo t o x i c i t y. In an attempt to provide some basis for stan-dardization of these techniques, Bruneret al. (2000) address this issue when describing objective assessment of testmethod performance in validation studies. They state that, “In order tom a ke further progress in developing validation methodology, two topics thatneed further exploration are how to ade-quately measure the predictive capacityof toxicity tests evaluated in validationstudies, and what values obtained fromthese measurements of predictive capac-ity are indicative of acceptable testmethod performance.” They suggest that
this issue deserves special considerationin view of the numerous data types derived from the toxicity test methodsunder evaluation and the plethora of statistical techniques which have beenused to assess performance of the testmethod. They analyzed four simulatedvalidation study data sets using three different approaches: calculation of cor-relation coefficients, determination of the 95% prediction interval (PI) and cal-culation of sensitiv i t y, specificity and accuracy as defined by Cooper et al.(1979). They found that calculation ofthe 95% PI provides the most informa-tion regarding performance of the testmethod under study. They also found thatdividing data sets into positive and nega-tive toxicity classifications followed bythe calculation of sensitivity, specificityand accuracy, leads to significant infor-mation loss.
8.4 Harmonization of validation efforts
As indicated above, improvement andstandardization of validation efforts hasreceived vigorous attention in the lastfive years. This attention was felt to bejustified in view of the fact that alterna-tives to eye irritation testing in animals,which have been in existence the longest
Inputs Conversion Algorithms
Output
Quantitative Data Quantitative Data Algorithm
Integration of Outputs From
Other Algorithms
In Vivo ToxicityPrediction
Censored Data Censored Data Algorithm
Qualitative Data Qualitative DataAlgorithm
Non-Qualified Data Non-Qualified Data Algorithm
Other Data Other Data Algorithm
Fig. 15: Converting alternative method data into an in vivo toxicity prediction (from Bruner et al., 1996).

HU G G I N S
ALTEX 20, Suppl.1/03 59
and about which the majority of data hasbeen generated, could not be validated tothe extent necessary to assure regulatoryacceptance in Europe or the UnitedStates. Although much work remains tobe accomplished regarding the validationof eye irritation alternatives, improve-ments made to the validation process resulting from this inquiry, particularlythose involving a “prevalidation” of thealternative test method, have been ap-plied successfully to areas of researchother than those of eye irritation alterna-tives (for example, skin corrosivity).
Fentem (2000) in noting the progressmade toward eff e c t ive validation of alternative methods observed that a retro-s p e c t ive approach using data alreadygenerated by industrial laboratories or inthe scientific literature had been used byICCVAM to validate the local lymphnode assay. Subsequent validation of theCorrositex® assay by this same body,however, relied upon data from formalvalidation studies. She concluded fromthis and other validation efforts by E C VAM that validation was now actually “in practice” as a reality as opposed to a theory. She suggested thatthe procedure for conducting formal val-idation studies adopted by ECVA Mcould form the starting point for discus-sions on international acceptance of validation procedures and their outcomesand that perhaps the OECD (Organiza-tion for Economic Cooperation and D evelopment) could oversee thisprocess.
ECVAM’s validation procedure (Ballset al., 1995a) consists of the followingmain points:• Test development
• Prevalidation (involving three phasesas discussed above)
• Validation (involving a formal interlab-oratory study)
• Independent assessment
• Progression toward regulatory accept-anceICCVAM has established its own set
of validation steps (NIEHS, 1997) as follows:
• Identify needs for new and/or im-proved test methods
• Understand toxic mechanisms
• Incorporate new science and technolo-gy into test methods
• Optimize transferable protocol
• Determine reliability and relevance• Independent peer rev i ew evaluation
of validation status• Determine acceptability for regulatory
risk assessmentAs can be seen, much overlap exists
between the two methods of validation.The first three steps of ICCVA M ’s approach are probably encompassed bythe first step in ECVA M ’s strateg y. Optimization of a transferable protocol(ICCVAM) is the objective of the preval-idation procedure suggested by ECVAM.Determination of reliability and rele-vance (ICCVAM) is the outcome of asuccessful validation study that includesinterlaboratory comparisons (ECVAM).Finally, independent assessments by ex-perts are a part of both schemes as ismovement toward regulatory acceptanceupon successful completion of the previous steps in either strategy. Hence,major differences between the two s t r a t egies are not apparent. How eve r,much work probably remains to be donewith regard to achieving a consensusabout issues of a more technical nature,such as biostatistical methods of evalua-tion as discussed above. Therefore, efforts directed toward harmonization ofvalidation methods are warranted andsuggestions by those such as Fentema b ove, indicating that ECVA M ’s approach could perhaps serve as a start-ing point for discussion, are pertinent.
8.5 Why validate?Brown (1992) noted that, “It appears thatthe past 10 years have produced relative-ly few advances in this use of alternativetests. I will suggest that this is becausetoo much effort has been concentrated onso-called ‘validation’ studies.” It wouldappear from the discussion above and thecomments below, that some of this efforthas finally borne fruit, particularly inview of the effective validation of sever-al alternative methodologies through useof improved validation methods.
Fentem (2000) states that, “Principally,validation studies are conducted to provide objective information on newtests, to show that they are robust andtransferable between laboratories, andthat the data generated can be relied upon for decision-making purposes (for
example, for the identification and label-ing of a potential skin corrosive, severeeye irritant, teratogen, etc.).”
Others have confirmed the value ofvalidation efforts as follows:
• Curren et al. (1998), in their report ofthe 13th meeting of the Scientific Groupon Methodologies for the Safety Eval-uation of Chemicals (SGOMSEC),write that “S c i e n t i fic principles de-mand that before newly developed al-ternative methods for safety testing arefully embraced by the industrial or reg-ulatory community, they reliably andre p roducibly predict the designatedtoxic end point. The process used to de-termine reliability and reproducibilityis termed validation, and it generallyculminates with a highly controlled,blinded study using multiple chemicalsand laboratories. It is imperative thatthe validation study is designed to con-firm the previously established repro-ducibility and predictive power of theassay.”
• Tennant (1998), in his discussion ofevaluation and validation issues in thedevelopment of transgenic mouse car-cinogenicity bioassays, states that, “Al-though results in transgenic modelsmay not be completely concordant withlong-term bioassays, the data can beused effectively in chemical and drugsafety assessments. Further, it is pro-posed that validation of the models isreadily achievable via ongoing stud-ies.”
• Artiges (1999) in his description of therole of the European Pharmacopoeia indeveloping alternative methods writesthat, “All these changes [deletion or re-placement of animal tests, reduction inthe number of animals used and possi-bilities of refining tests in ways thatcause less suffering] require extensivevalidation work and collaborative tri-als in a number of control laboratoriesto ensure that the quality standards aremaintained with the alternative meth-ods.”Hence, validation seems to have
“come of age” after a lengthy process ofd e finition and refinement, despite negative comments to the contrary in theearly stages of its development.

HU G G I N S
ALTEX 20, Suppl.1/0360
8.6 Mechanistic understandings: what are we validating?
One of the primary challenges of devel-oping alternative models of toxicity test-ing has been the question of relevance ofendpoint measured in vitro to the toxicresponse noted in the whole animal. In terms of validation, many partiesquestion the wisdom of validating anendpoint that may or may not have muchto do with the basic mechanism of actionof a toxic chemical. However, others a rgue that if a good correlation isachieved between the endpoint measuredin an in vitro assay and an in vivo end-point, then the in vitro assay is indeedpredictive of the in vivo response. Theyalso point out that determining the mech-anism of action of a toxicant is, for mosttoxicants, a very lengthy procedure.
As noted in an earlier portion of thisposition paper, a good example of thisquandary regarding relevance is recentwork with gene chips. Pennie et al.(2000) have argued that changes in geneexpression that do not result in alteredprotein production are of relevance andshould not be dismissed. They suggestthat we adopt a broader view of biologi-cal relevance and realize that events tran-spiring on a gene chip may not always bereduced/refined to discrete biochemicalpathways. They indicate that these eventsshould nevertheless, be considered valuable. This perspective is undoubted-ly true for other areas of alternatives toanimal testing and should definitely beincorporated it into our thinking aboutvalidation efforts.
8.7 Summary, conclusions, and future work
Experiments designed to evaluate howclosely results from an in vitro assay mirror those obtained from an in vivoassay have been performed practicallysince the beginning of the alternatives to animal testing movement (and much earlier if we look at early experimentsperformed “in vitro” in many disci-plines). However, only recently has thistype of exercise been subjected to attempts at formalization and standardi-zation. These relatively recent eff o r t sh ave been prompted by the need to evaluate objectively those in vitro assaysthat could be used to reduce, refine, or
replace animals in medical experimenta-tion in order to promote regulatory acceptance of them.
Validation of alternative methods hasjust emerged from a rather chaotic phasein which the principles behind appropri-ate conduct of a validation study weredefined, mainly through trial and error.Much refinement has come out of this“exploratory” phase, including recogni-tion that validation studies should bebuild upon a solid platform, consisting ofcomponents such as good reference standards, reliable protocol transfer between laboratories, and appropriateapplication of biostatistical techniques.Efforts are now underway to apply theselessons learned to future validation studies and to harmonize validation techniques among countries in order tomaximize the possibility that the datagenerated can be used worldwide.
So, despite the fact that some in vitroassays may not have been validated inthe past due to faults inherent in the validation study and not in the assay,future attempts to validate in vitro assayswill benefit from a better understandingof the requirements of validation andstandardization of the conduct of valida-tion studies. Furthermore, validation ofendpoints that are not directly correlatedwith a defined mechanism of action isslowly becoming more acceptable. Thisis due, in part, to our increasing realiza-tion of the fact that our knowledge aboutmechanisms of action at any given timemay be limited. It is hoped that this perspective will be taken into considera-tion by regulatory bodies charged withthe task of evaluating and ultimately,accepting alternatives to animal testingas viable tests of toxicity. If so, Koeter’ssecond reason for the lack of regulatoryacceptance of alternatives mentioneda b ove (i.e. “toxicologists/regulators appear reluctant actually to use the dataprovided for hazard and risk assessmentprocedures because of a lack of confi-dence with the (types of) endpoints of thenew test.”) may be set aside as no longerviable.
Validation, while not an actual alterna-tive to animal testing, certainly can besaid to be the “gateway” through whicheach proposed alternative must pass inorder to find acceptance as a viable test
of toxicity. Therefore, the methods bywhich we conduct validation studies areas important, if not more so, than thoseby which we conduct an alternative assay. Our previous attempts at valida-tion have been well meant, but expen-sive, and, in some cases, not so wellplanned or executed. We have, however,through reassessment, honed the tech-nique and applied it successfully to produce validated alternatives to animaltesting. All that truly remains to be doneis to see that the technique is used consistently to evaluate alternative meth-ods, thereby contributing to a muchneeded “atmosphere of trust” among scientists, the industrial sector, and regu-lators.
ReferencesArtiges, A. (1999). Alternatives to ani-
mals in the development and control ofbiological products for human and vet-erinary use. The role of the EuropeanPharmacopoeia. Dev. Biol. Stand. 101 ,29-35.
Balls, M., Blaauboer, B., Brusick, D. etal. (1990). Report and recommenda-tions of the CAAT/ERGATT workshopon the validation of toxicity test proce-dures. ATLA 18, 313-337.
Balls, M., Blaauboer, B., Fentem, J. et al.(1995a). Practical aspects of the vali-dation of toxicity test procedures. Report and recommendations of ECVAM workshop 5. ATLA 23, 129-147.
Balls, M., De Klerck, W., Baker, F. et al.(1995b). Development and validationof non-animal tests and testing strate-gies: the identification of a coordinatedresponse to the challenge and the opportunity presented by the sixthamendment to the cosmetics directive.Report and recommendations of ECVAM workshop 7. ATLA 23, 398-409.
Balls, M. and Corcelle, G. (1998a).Statement on the scientific validity ofthe rat skin electrical resistance (TER)test (an in vitro test for skin corrosivi-ty). ATLA 26, 275-277.
Balls, M. and Corcelle, G. (1998b).Statement on the scientific validity ofEPISKIN™ test (an in vitro test forskin corrosivity). ATLA 26, 278-280.

HU G G I N S
ALTEX 20, Suppl.1/03 61
Balls, M. and Corcelle, G. (1998c).Statement on the scientific validity ofthe 3T3 NRU PT test (an in vitro testfor phototoxic potential). ATLA 26,281-283.
Balls, M., Berg, N., Bruner, L. et al.(1999). Eye irritation testing: The wayforward. The report and recommenda-tions of ECVAM workshop 34. ATLA27, 53-77.
Brown, N. (1992). Hazard identificationin developing toxicology – the role ofnonwhole mammal tests. Reprod. Tox-icol. 6(2), 178.
B r u n e r, L., Carr, G. and Curren, R.(2000). Is it good enough? Objectiveassessment of toxicity test method performance in validation studies. InM. Balls, A.-M. van Zeller and M. E.Halder (eds.), Progress in the Reduc-tion, Refinement, and Replacement ofAnimal Experimentation, (375-384).Amsterdam, London, New Yo r k ,Tokyo: Elsevier.
C o o p e r, J., Saracci, R. and Cole, P.(1979). Describing the validity of carcinogen screening tests. Br. J. Can-cer 39, 87-89.
Curren, R., Southee, J., Spielmann, H. etal. (1995). The role of prevalidation inthe development, validation and acceptance of alternative methods. ATLA 23, 211-217.
Curren, R., Bruner, L., Goldberg, A. andWalum, E. (1998). 13th Meeting of theScientific Group on Methodologies forthe Safety Evaluation of Chemicals(SGOMSEC): validation and acutetoxicity testing. E nv i ron. Health Perspect. 106(2), 419-425.
Curren, R. and Southee, J. (2000).P r evalidation: successes justify theconcept. In M. Balls, A.-M. van Zellerand M. E. Halder (eds.), Progress inthe Reduction, Refinement, and Re-placement of Animal Experimentation,(359-363). Amsterdam, London, NewYork, Tokyo: Elsevier.
Fentem, J. (2000). Validation in practice– from theory to reality. In M. Balls,A.-M. van Zeller and M. E. Halder(eds.), P rog ress in the Reduction, Refinement, and Replacement of Ani-mal Experimentation, (365-373). Amsterdam, London, New Yo r k ,Tokyo: Elsevier.
Gad, S. (1990). Recent developments inreplacing, reducing, and refining ani-mal use in toxicologic research andtesting. Fundam. Appl. Toxicol. 15(1) ,8-16.
Goldberg, A., Frazier, J., Brusick, D. etal. (1993). Framework for the valida-tion and implementation of in vitrotoxicity tests: Report of the validationand technology transfer committee of
the Johns Hopkins Center for Alterna-tives to Animal Testing. J. Am. Coll.Toxicol. 12(1), 23-30.
Goldberg, L. (1986). Charting a coursefor cell culture alternatives to animaltesting. Fundam. Appl. Toxicol. 6(4),607-617.
Koeter, H. (1995). Validation: A highlycharged concept. Toxicol. In Vitro 9(6),851-856.
Mehlman, M., Pfitzer, E. and Scala, R.(1989). A report on methods to reduce,refine and replace animal testing in in-dustrial toxicology laboratories. CellBiol. Toxicol. 5(3), 349-358.
NIEHS (National Institutes of Environ-mental Health Science) (1997). Valida-tion and regulatory acceptance of toxicological test methods: A report ofthe ad hoc interagency coordinatingcommittee on the validation of alter-n a t ive methods. NIH publication no. 97-3981. Research Triangle Park, NC, USA.
Pennie, W. D., Tugwood, J., Oliver, G.and Kimber, I. (2000). The principlesand practice of toxicogenomics: appli-cations and opportunities. Toxicologi-cal Sciences 54, 277-283.
Tennant, R. (1998). Evaluation and validation issues in the development oftransgenic mouse carcinogenicitybioassays. Environ. Health Perspect.106(2), 473-476.