altered brain metabolism in vestibular migraine ... · brief report altered brain metabolism in...
TRANSCRIPT

Brief Report
Altered brain metabolism in vestibularmigraine: Comparison of interictaland ictal findings
Jung H Shin1, Yu K Kim2, Hyo-Jung Kim3,4 and Ji-Soo Kim1
Abstract
Background: Vestibular symptoms/signs frequently coexist with migraine, but the mechanisms of migraine-related
vestibular dysfunction remain to be elucidated. This study aimed to determine altered brain metabolism in vestibular
migraine.
Methods: Two patients with vestibular migraine underwent 18F-fluorodeoxy glucose (FDG) positron-emission tomog-
raphy (PET) during and between attacks of vestibular migraine in addition to detailed neurotological evaluation. We
analyzed the regional brain metabolism of the patients in comparison with that of age-matched healthy controls in each
patient. We also compared ictal with interictal FDG PET using a subtraction method.
Results: During the attacks, both patients showed an activation of the bilateral cerebellum and frontal cortices, and
deactivation of the bilateral posterior parietal and occipitotemporal areas. One patient also showed hypermetabolism in
the dorsal pons and midbrain, right posterior insula, and right temporal cortex while the other patient had an additional
activation of the left temporal cortex. Compared with interictal images, ictal PET showed increased metabolism in the
bilateral cerebellum, frontal cortices, temporal cortex, posterior insula, and thalami.
Conclusion: During the attacks of vestibular migraine, the increased metabolism in the temporo-parieto-insular areas and
bilateral thalami indicates activation of the vestibulo-thalamo-cortical pathway, and decreased metabolism in the occipital
cortex may represent reciprocal inhibition between the visual and vestibular systems.
Keywords
Migraine, vertigo, vestibular migraine, PET, metabolism
Date received: 9 April 2013; revised: 29 June 2013; accepted: 2 July 2013
Introduction
Vestibular symptoms/signs frequently coexist withmigraine and include dizziness/vertigo, motion sicknessand imbalance (1–5). Only recently, the Committee forClassification of Vestibular Disorders of the BaranySociety and the Migraine Classification Subcommitteeof the International Headache Society (IHS) havejointly formulated diagnostic criteria for ‘‘vestibularmigraine’’ that was also termed as ‘‘migraine-associatedvertigo’’ or ‘‘migrainous vertigo.’’ However, the mech-anisms of migraine-related vestibular dysfunctionremain to be elucidated. There have been severalhypotheses on the pathophysiology of vestibular symp-toms/signs in migraine. The vestibular nuclei receivenoradrenergic inputs from the locus ceruleus (6) andserotoninergic inputs from the dorsal raphe nucleus(7). Accordingly, activation of these nuclei duringmigraine attacks may give rise to vestibular symptoms.Since the trigeminal nucleus caudalis also has reciprocal
connections with the vestibular nuclei, and neurogenicinflammation of the trigeminal system is believed to bea mechanism of migraine (8), trigeminal activation mayprovoke vestibular symptoms during migraine attacks.
1Department of Neurology, Seoul National University College of
Medicine, Seoul National University Bundang Hospital, Republic of Korea2Department of Nuclear Medicine, Boramae Medical Center, Seoul
National University College of Medicine, Republic of Korea3Kangwon National University College of Medicine, Republic of Korea4Medical Research Institute, Seoul National University Bundang Hospital,
Republic of Korea
J.H.S. and Y.K.K. contributed equally to this paper.
Corresponding author:
Ji-Soo Kim, Department of Neurology, Seoul National University College
of Medicine, Seoul National University Bundang Hospital, 300 Gumi-
dong, Bundang-gu, Seongnam-si, Gyeonggi-do, 463-707, Republic of
Korea.
Email: [email protected]
Cephalalgia
2014, Vol 34(1) 58–67
! International Headache Society 2013
Reprints and permissions:
sagepub.co.uk/journalsPermissions.nav
DOI: 10.1177/0333102413498940
cep.sagepub.com

Otherwise, cortical spreading depression may influencethe vestibular nuclei via direct projections from the pos-terior parietal cortex (2). The direct vasodilator effect ofsubstance P, neurokinin A and calcitonin gene-relatedpeptide from the trigeminal and eighth cranial nervefibers is another hypothesis (9). Since trigeminal sen-sory innervations are prominent in the inner ear struc-tures, such as the stria vascularis and crista ampularis(10), direct activation of the inner ear structures byneuropeptides may also give rise to dizziness/vertigoand otologic findings (11) during migraine attacks.Other explanations include vasospasm-induced ische-mia of the labyrinth (12), and genetic abnormalities ofthe calcium channels, as have been found in episodicataxia type 2 and familial hemiplegic migraine that maydevelop migraine and vertigo in common (13).
Functional imaging of the brain during and betweenmigraine attacks may provide insights into the patho-physiology of migraine. Previously, positron-emissiontomography (PET) revealed hypermetabolism of thelocus cereleus and dorsal raphe nucleus during migraineattacks. Also, activation of the cingulate, bilateral insu-lae and cerebellum has been noted during the attacks.Another study also lateralized the hypermetabolic pon-tine area on the side of headache (14,15). Recently,functional magnetic resonance imaging (MRI) alsodemonstrated ictal hypermetabolism in the red nucleus,substantia nigra, pons, medial longitudinal fasciculusand periaqueductal gray (PAG) in migraineuers (16).In contrast, patients with migraine without aurashowed hypoperfusion of the postcentral, inferior tem-poral and inferior frontal gyri (17), and posterior cere-bral cortex (18,19).
According to the findings described above, activa-tion of the dorsal brainstem appears to be involved inthe generation of migraine. The dorsal raphe nucleusand PAG constitute the descending nociceptive path-ways and contribute to autonomic control via norepi-nephrinergic and serotoninergic projections (20).Hyperactivation of the dorsal brainstem nuclei leadsto pain by modulation of the central pain pathway,vasodilation of the dural arteries, plasma extravasationand release of vasoactive substances (21).
However, altered brain metabolism has not beensought during attacks of vestibular migraine. Herein,we report findings of ictal PET in two patients withvestibular migraine in comparison with interictalbrain metabolism.
Materials and methods
Subjects
Patient 1. This 30-year-old woman had suffered frommigrainous headaches for 15 years. The headache was
pulsating mostly on both sides, and was accompaniedby nausea, vomiting, phonophobia, photophobia, andvertigo. She frequently experienced flashing lights inboth eyes after the resolution of the headaches. Severeheadaches tended to develop a couple of times a monthand lasted a few hours to days. The headache and ver-tigo tended to develop simultaneously. The headachespartially responded to analgesics, and the headache fre-quency had decreased with a beta blocker (propran-olol). She denied a family history of migraine.Between the attacks of migraine headaches, sheshowed subtle left-beating nystagmus without fixationthat changed into right-beating nystagmus after hori-zontal head shaking. Horizontal smooth pursuit (SP)and saccades were normal. Other findings of neuro-logical and neurotological examinations were nonre-vealing. During the attacks of vestibular migraine, sheshowed similar findings except for prominent downbeatnystagmus after horizontal head shaking (pervertedhead-shaking nystagmus (HSN)). The patient had anictal PET three days after the onset of headache whenshe still had the symptoms, and an interictal PET twoweeks after the cessation of the headache when she wasfree of symptoms.
Patient 2. This 57-year-old woman had a history ofmigrainous headaches for about 40 years. The head-ache was throbbing usually in the right side. Theheadache began to accompany vertigo five yearsbefore. The headache and vertigo usually lasted sev-eral days with nausea and vomiting. She denied tin-nitus, ear fullness, or hearing loss. The headacheresponded to triptan. She denied a familial historyof migraine.
Between the attacks, she had no spontaneous orHSN, but showed subtle downbeat nystagmus duringstraight head hanging. Horizontal saccades and SPwere normal. The results of cervical vestibular-evokedmyogenic potential and bithermal caloric and rotatorychair tests were within normal range. During the ictuswhen the PET was taken, she had migrainous headacheand vertigo. Examination showed subtle left-beatingspontaneous nystagmus without fixation. Horizontalhead impulse tests were normal. She also showedsubtle left-beating horizontal-torsional nystagmuswith a downbeat component during head turning toeither side while supine and Dix-Hallpike maneuver ineither direction. Horizontal saccades were normal, buthorizontal SP was mildly impaired. The patient had anictal PET two days after the headache onset when shestill suffered from symptoms, and an interictal PET 10days after cessation of the headache when the symp-toms were completely resolved.
Both patients met the diagnostic criteria for vestibu-lar migraine (22).
Shin et al. 59

Oculography
In both patients, eye movements were recorded with avideo-oculography system (SMI, Teltow, Germany)between and during the attacks of vestibular migraine.Patients had recording of spontaneous nystagmus with/without fixation and gaze-evoked nystagmus in the hori-zontal (�30�) and vertical (�20�) planes. HSN wasinduced using a passive head-shaking maneuver. Aftergrasping the patient’s head firmly on both sides with thehead pitched forward by approximately 30�, the headwas shaken horizontally in a sinusoidal fashion at arate of 2.8Hz with an approximate amplitude of �10�
for 15 seconds. Detailed methods and normative datahave been described previously (23).
To induce positional nystagmus, patients lied supinefrom sitting and turned their heads to either side whilein supine. Then, patients were moved from a supine to asitting position and the head was bent down. Patientswere also subjected to right and left Dix-Hallpike man-euvers and straight head hanging test (24).
Horizontal saccades were generated by asking thepatients to follow a target moving on a light bar withranges of �5�, �10� and �15�. For each saccade,latency, accuracy and peak velocity were computedand compared with the data from 50 normal controls.The stimulus for horizontal SP was a light targetmoving in a sinusoidal pattern at peak velocities of10�/s and 20�/s. The amplitude of target motion was20�. For gain, the peak eye velocity was comparedwith the target velocity after eliminating saccades.Then, average gain from the accepted cycles was com-pared with the values from 50 age-matched controls.Impaired SP was defined when the SP gain in thepatient was less than the mean–2 standard deviations(SD) of that in the age-matched normal controls ateither peak target velocity (25).
Brain 18F-fluorodeoxy glucose (FDG)-PET scan
Patients underwent FDG-PET twice: during an attackof vestibular migraine (ictal PET) and during the head-ache-free phase (interictal PET). The subjects hadfasted for at least six hours before the PET study.The brain imaging was started 40 minutes after abolus injection of 4.8MBq/kg FDG and continuedfor 15 minutes. During the FDG equilibration periodfor 40 minutes, the patients were instructed to remainlying comfortably in a dimly lit, quiet waiting room. Allstudies were conducted using Allegro PET scanner(Philips Medical Systems, Cleveland, OH, USA) (26).Ten-minute emission scans and attenuation maps usinga Cs-137 transmission source were obtained. Imageswere reconstructed using the three-dimensional (3D)row-action maximum-likelihood algorithm with a 3Dimage filter of 128� 128� 90 matrices with a pixel
size of 2� 2� 2mm3, and attenuation correction wasperformed.
Imaging preprocessing and statistical analyses wereconducted using SPM 5 (Statistical ParametricMapping 5; Wellcome Department of CognitiveNeurology, London, UK). In each patient, the regionalabnormality of cerebral metabolic activity was deter-mined by comparing the patient’s scan with those of15 age-matched healthy women selected from our data-base. The mean age of the controls for patient 1 was29.1� 4.9 years, and for patient 2 was 56.5� 3.5 years.The significance was considered when a cluster consist-ing of at least 100 contiguous voxels exceeded a thresh-old height of p< 0.005. And the differences in theregional metabolism between the ictal and interictalimages for each patient were also assessed by subtract-ing the images from each other using Analyze 10.0workspace (AnalyzeDirect Inc, KS, USA). The voxelvalues of the ictal and interictal scans were normalizedto the mean values of the brain and subtracted. To findsignificant regional activation in the ictal scan, thethreshold was set as 2 SD from the mean values inthe subtraction image.
Results
Interictal PET
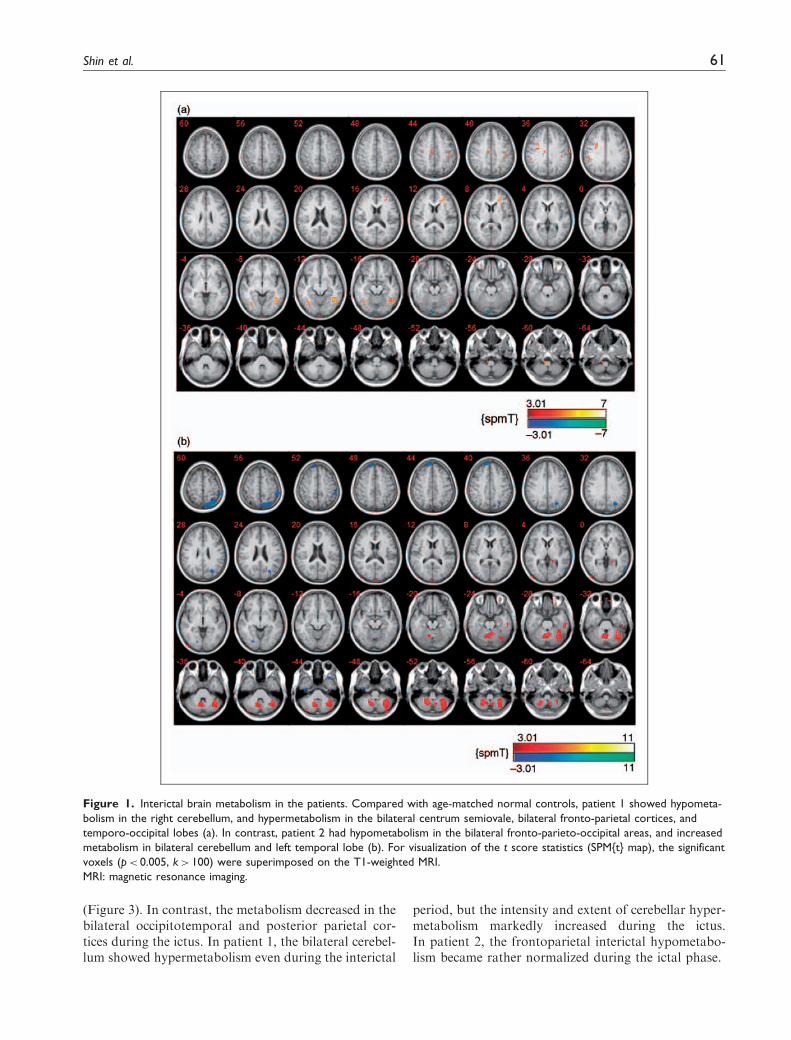
Between attacks of headache and vertigo, patient 1showed hypometabolism in the right cerebellum, andhypermetabolism in the bilateral centrum semiovale,bilateral fronto-parietal cortices and temporo-occipitallobes. In contrast, patient 2 had hypometabolism in thebilateral fronto-parieto-occipital areas, and increasedmetabolism in bilateral cerebellum and left temporallobe (Figure 1).
Ictal PET
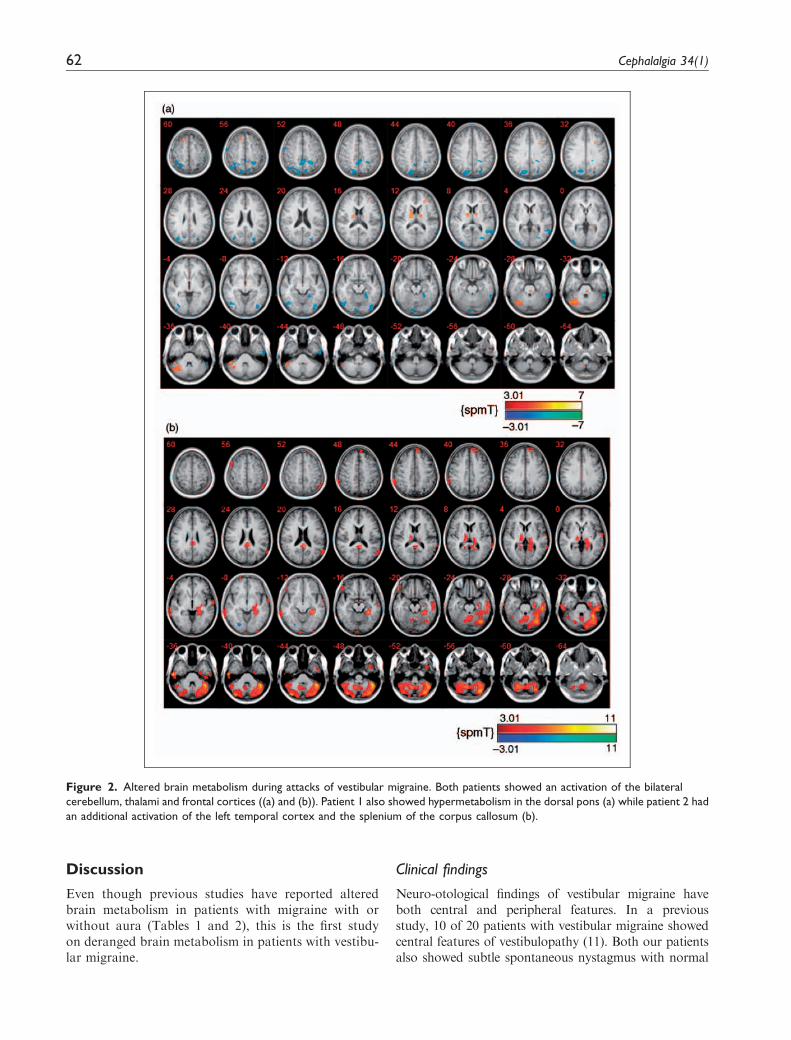
During attacks of vestibular migraine, both patientsshowed an activation of the bilateral cerebellum, thal-amus and frontal cortices. Patient 1 also showed hyper-metabolism in the dorsal pons while patient 2 had anadditional activation of left temporal cortex and thesplenium of the corpus callosum. In contrast, bothpatients exhibited hypometabolism in the posterior par-ietal and occipitotemporal areas (Figure 2).
Comparison of interictal and ictal PET
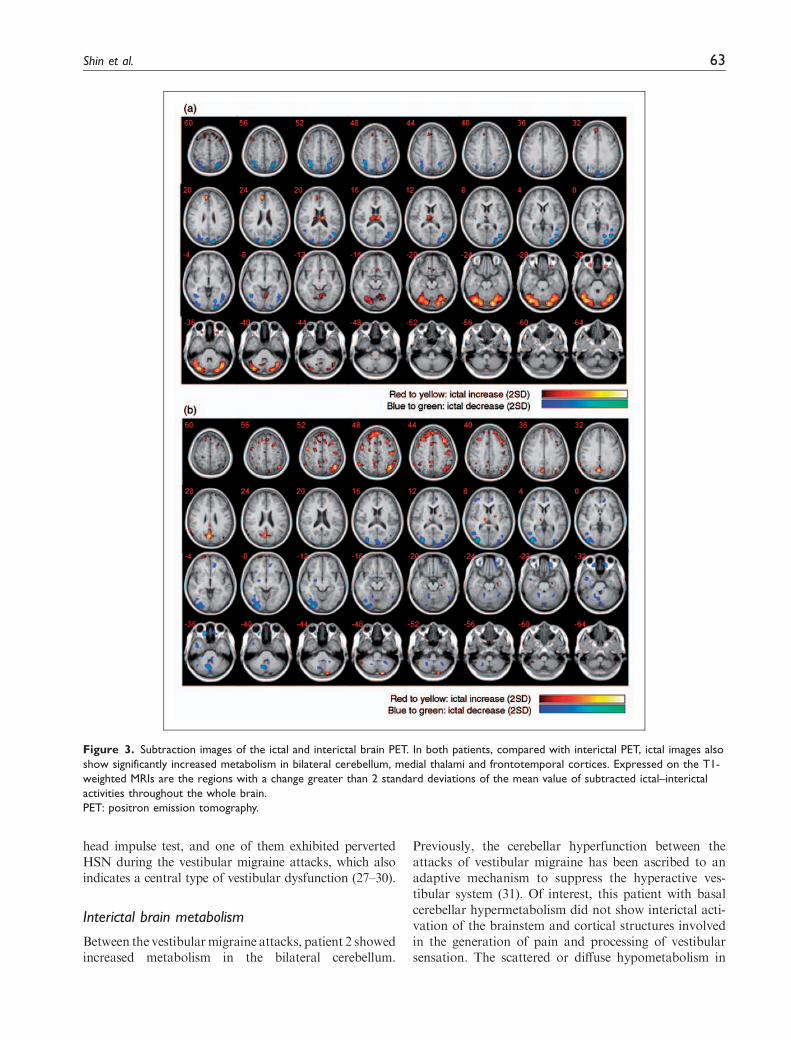
Subtraction images demonstrated the differencesbetween ictal and interictal regional metabolism.Compared with interictal images, ictal PET commonlyshowed increased metabolism of the bilateral cerebel-lum, frontal cortices and thalami in both patients
60 Cephalalgia 34(1)
(Figure 3). In contrast, the metabolism decreased in thebilateral occipitotemporal and posterior parietal cor-tices during the ictus. In patient 1, the bilateral cerebel-lum showed hypermetabolism even during the interictal
period, but the intensity and extent of cerebellar hyper-metabolism markedly increased during the ictus.In patient 2, the frontoparietal interictal hypometabo-lism became rather normalized during the ictal phase.
Figure 1. Interictal brain metabolism in the patients. Compared with age-matched normal controls, patient 1 showed hypometa-
bolism in the right cerebellum, and hypermetabolism in the bilateral centrum semiovale, bilateral fronto-parietal cortices, and
temporo-occipital lobes (a). In contrast, patient 2 had hypometabolism in the bilateral fronto-parieto-occipital areas, and increased
metabolism in bilateral cerebellum and left temporal lobe (b). For visualization of the t score statistics (SPM{t} map), the significant
voxels (p< 0.005, k> 100) were superimposed on the T1-weighted MRI.
MRI: magnetic resonance imaging.
Shin et al. 61
Discussion
Even though previous studies have reported alteredbrain metabolism in patients with migraine with orwithout aura (Tables 1 and 2), this is the first studyon deranged brain metabolism in patients with vestibu-lar migraine.
Clinical findings
Neuro-otological findings of vestibular migraine haveboth central and peripheral features. In a previousstudy, 10 of 20 patients with vestibular migraine showedcentral features of vestibulopathy (11). Both our patientsalso showed subtle spontaneous nystagmus with normal
Figure 2. Altered brain metabolism during attacks of vestibular migraine. Both patients showed an activation of the bilateral
cerebellum, thalami and frontal cortices ((a) and (b)). Patient 1 also showed hypermetabolism in the dorsal pons (a) while patient 2 had
an additional activation of the left temporal cortex and the splenium of the corpus callosum (b).
62 Cephalalgia 34(1)
head impulse test, and one of them exhibited pervertedHSN during the vestibular migraine attacks, which alsoindicates a central type of vestibular dysfunction (27–30).
Interictal brain metabolism
Between the vestibularmigraine attacks, patient 2 showedincreased metabolism in the bilateral cerebellum.
Previously, the cerebellar hyperfunction between theattacks of vestibular migraine has been ascribed to anadaptive mechanism to suppress the hyperactive ves-tibular system (31). Of interest, this patient with basalcerebellar hypermetabolism did not show interictal acti-vation of the brainstem and cortical structures involvedin the generation of pain and processing of vestibularsensation. The scattered or diffuse hypometabolism in
Figure 3. Subtraction images of the ictal and interictal brain PET. In both patients, compared with interictal PET, ictal images also
show significantly increased metabolism in bilateral cerebellum, medial thalami and frontotemporal cortices. Expressed on the T1-
weighted MRIs are the regions with a change greater than 2 standard deviations of the mean value of subtracted ictal–interictal
activities throughout the whole brain.
PET: positron emission tomography.
Shin et al. 63

bilateral fronto-parieto-occipital areas may be relatedto interictal suppression of the structures related to per-ception of pain and vestibular symptoms.
Ictal activation of the brain
Patients with migraine may suffer from cerebellarsymptoms such as dizziness, dysarthria and ataxia(32). Our patients showed normal or mildly increasedcerebellar metabolism between migraine attacks, butmarkedly enhanced metabolism in the bilateral cerebel-lum during the ictus. Previously, ocular motor signs ofthe vestibulocerebellar dysfunction, such as saccadicpursuit and gaze-evoked nystagmus, have also beenreported in vestibular migraine (11). However, in viewof the increased tilt suppression of the vestibulo-ocularreflex in a previous study on migraineurs (31), the cere-bellar hypermetabolism between or during attacks ofvestibular migraine may be better explained by anadaptive cerebellar mechanism to suppress the hyper-active vestibular system in these patients.
Bilateral medial thalami were also activated in bothpatients. Hypermetabolism in the temporal cortex andthalamus may have been caused by activation of thevestibular nucleus and the vestibulo-cortical projectionsvia the posterolateral thalamus. Indeed, patients withunilateral vestibular neuritis showed activation of bilat-eral thalami in addition to vestibular cortical areas (33).
The left temporal cortex was activated in patient 2during the vestibular migraine. In humans, vestibularprocessing involves the posterior insula, superficialpart of the temporoparietal junction, superior temporalregions, sensorimotor cortex, hippocampus and fronto-parieto-occipital areas (34). However, electricalstimulation of human brain cortex found that the tem-poro-perisylvian cortex is particularly sensitive for diz-ziness (35). Unilateral vestibular stimulation in healthyvolunteers also induces activation of both temporo-parieto-insular areas, dominantly in the stimulated side(33). So, the activation of the vestibular corticies in ourpatient is consistent with the vestibular symptomsduring the attacks. The hypermetabolism in the perisyl-vian temporal cortex, which had not been found in theprevious studies on the patients with migraine, supportsthe vestibulo-cortical activation in our patients (35).A previous study reported hypermetabolism of the pos-terior insula and thalamus in patients withmigraine eventhough there was no description of dizziness (36).
One of our patients showed an activation of the rightdorsal pons, which was also found in previous studies(14,15). Since the locus ceruleus and the dorsal raphenucleus have noradrenergic and serotoninergic projec-tions to the vestibular nuclei (7), activation of thedorsal pons in the areas of the locus ceruleus anddorsal raphe nucleus may affect the central vestibularstructures and cause dizziness.
Table 1. Brain metabolism between and during the ictus in our patients.
Patients 1/F/30 2/F/57
Interictal
Activation Rt lower medulla Bilateral cerebellum
Bilateral temporo-occipital lobes Lt temporal cortex.
Bilateral frontoparietal cortices
Bilateral corona radiata
Rt cingulatecortex
Deactivation Rt cerebellum Bilateral fronto-parieto-occipital areas
Ictal
Activation Rt cerebellum Bilateral cerebellum
Lt dorsalpons Lt inferior temporal cortex
Bilateral thalami Lt frontal cortex
Bilateral frontal cortices Bilateral medial thalami
Bilateral corona radiata Lt temporal cortex
Splenium of the corpus callosum
Deactivation Bilateral fronto-parieto-occipital areas Bilateral occipitoparietal cortices
Ictal vs interictal
Activation Bilateral cerebellum Bilateral cerebellum
Bilateral thalami Right medial thalamus
Bilateral frontal cortices Bilateral frontoparietal cortices
Lt perisylvian temporal cortex
F: female; Lt: left; Rt: right.
64 Cephalalgia 34(1)

Ictal deactivation of the brain
Our patients also showed hypometabolism of bilateraloccipitotemporal areas. Patients with visual aurashowed a perfusion defect in the unilateral occipitalarea contralateral to the visual symptoms (18,37),which is explained by spreading depression or ischemia(38). However, our patients did not report visual auraduring the attacks, and the occipitotemporal hypoper-fusion was bilateral. Previously, patients with vestibularneuritis also showed deactivations in the bilateral visualcortices (33). Accordingly, the occipitotemporal deacti-vation observed in our patients may be better explainedby reciprocal inhibition between the visual and vestibu-lar systems (39).
Limitation of the study
This study involved only two patients, which limits thegeneralization of the results to explain the pathophysi-ology of migraine, and of vestibular migraine in par-ticular, in the general population. The preliminarynature of our results should be supplemented byfuture studies with a larger number of migraineurswith or without vestibular symptoms.
Furthermore, the metabolic changes observedduring the ictus cannot be solely attributed toheadache and vertigo since the associated photopho-bia and phonophobia may have affected the corticalexcitability (40,41). Indeed, the ictal hyperactivation ofthe temporoperisylvian cortex in patient 1 mighthave been related to the phonophobia experiencedby the patient during the ictus even though occipitalhypometabolism is not consistent with the metabolicchanges observed in photophobia (15,42,43).This issue would be resolved with a comparison ofthe metabolism among migraineurs with and withoutvertigo, which again remains a subject of futurestudies.
Conclusion
During attacks of vestibular migraine, the increasedmetabolism of the temporo-parieto-insular areas andbilateral thalami indicates activation of the vestibulo-thalamo-vestibulocortical pathway, and decreasedmetabolism in the occipital cortex may represent recip-rocal inhibition between the visual and vestibularsystems.
Table 2. Altered brain metabolism during the ictus in the previous reports on migraine.
Study Subjects/symptom Activation sites Deactivation sites
Woods et al. (1994) (19) Without aura (n¼ 1) Bilateral occipitotemporal areas.
Weiller et al. (1995) (15) Without aura (n¼ 9) Dorsoraphe nucleus
Locus ceruleus
Cingulate cortex
Cutrer et al. (1998) (37) With visual aura (n¼ 4) Contralateral visual cortex.
Sanchez et al. (1999) (18) With (n¼ 14) and
without (n¼ 14) aura
Occipital cortex contralateral
to headache area
Bahra et al. (2001) (36) Without aura (n¼ 1) Rostral brain stem
Intra and extracranial vessels
Cingulate cortex
Prefrontal and posterior insular areas
Bilateral cerebellar cortices
Thalamus
Superior parietal cortex
Supplementary motor cortex
Lentiform nucleus
Cao et al. (2002) (16) With (n¼ 23) and
without (n¼ 3) aura
Red nucleus and substantia nigra
Periaqueductal gray matter
Afridi et al. (2005) (14) With (n¼ 8) and
without (n¼ 16) aura
Dorsal lateral pons
Cerebellum
Insula
Anterior cingulate
Prefrontal cortex
Putamen
Shin et al. 65

Clinical implications
. This is the first study on deranged brain metabolism in patients with vestibular migraine.
. During attacks of vestibular migraine, the increased metabolism of the temporo-parieto-insular areas andbilateral thalami indicates activation of the vestibulo-thalamo-vestibulocortical pathway, and decreasedmetabolism in the occipital cortex may represent reciprocal inhibition between the visual and vestibularsystems.
Author contributions
J.H. Shin analyzed and interpreted the data and wrote the
manuscript. Y.K. Kim and H.-J. Kim analyzed and inter-preted the data, and revised the manuscript. J.-S. Kim,MD, as the corresponding author, conducted the designand conceptualization of the study, interpretation of the
data, and drafting and revising of the manuscript.
Funding
This study was supported by a grant from the Korea Health21 R&D Project, Ministry of Health & Welfare, Republic of
Korea (A080750).
Conflicts of interest
J.-S. Kim serves as an associate editor of Frontiers in Neuro-otology and on the editorial boards of the Journal of Korean
Society of Clinical Neurophysiology, Research in VestibularScience, Journal of Clinical Neurology, Frontiers in Neuro-ophthalmology, Journal of Neuro-ophthalmology, and Case
Reports in Ophthalmological Medicine; and has receivedresearch support from SK Chemicals Co. Ltd.
J.H. Shin, Y.K.Kim andH.-J. Kimhave nothing to declare.
References
1. Kayan A and Hood JD. Neuro-otological manifestationsof migraine. Brain 1984; 107(Pt 4): 1123–1142.
2. Faugier-Grimaud S and Ventre J. Anatomic connectionsof inferior parietal cortex (area 7) with subcortical struc-tures related to vestibulo-ocular function in a monkey
(Macaca fascicularis). J Comp Neurol 1989; 280: 1–14.3. Neuhauser H, Leopold M, von Brevern M, et al. The inter-
relations of migraine, vertigo, and migrainous vertigo.
Neurology 2001; 56: 436–441.4. Evans RW, Marcus D and Furman JM. Motion sickness
and migraine. Headache 2007; 47: 607–610.5. Cha YH and Baloh RW. Migraine associated vertigo.
J Clin Neurol 2007; 3: 121–126.6. Schuerger RJ and Balaban CD. Organization of the coer-
uleo-vestibular pathway in rats, rabbits, and monkeys.
Brain Res Brain Res Rev 1999; 30: 189–217.7. Halberstadt AL and Balaban CD. Organization of projec-
tions from the raphe nuclei to the vestibular nuclei in rats.
Neuroscience 2003; 120: 573–594.8. Moskowitz MA, Nozaki K and Kraig RP. Neocortical
spreading depression provokes the expression of c-fos pro-tein-like immunoreactivity within trigeminal nucleus cau-
dalis via trigeminovascular mechanisms. J Neurosci 1993;13: 1167–1177.
9. Furman JM, Marcus DA and Balaban CD. Migrainousvertigo: Development of a pathogenetic model and struc-
tured diagnostic interview. Curr Opin Neurol 2003; 16:5–13.
10. Vass Z, Shore SE, Nuttall AL, et al. Direct evidence of
trigeminal innervation of the cochlear blood vessels.Neuroscience 1998; 84: 559–567.
11. von Brevern M, Zeise D, Neuhauser H, et al. Acutemigrainous vertigo: Clinical and oculographic findings.
Brain 2005; 128: 365–374.12. Radtke A, Lempert T, Gresty MA, et al. Migraine and
Meniere’s disease: Is there a link? Neurology 2002; 59:
1700–1704.13. Ophoff RA, Terwindt GM, Vergouwe MN, et al. Famil-
ial hemiplegic migraine and episodic ataxia type-2 are
caused by mutations in the Ca2þ channel geneCACNL1A4. Cell 1996; 87: 543–552.
14. Afridi SK, Matharu MS, Lee L, et al. A PET study
exploring the laterality of brainstem activation inmigraine using glyceryl trinitrate. Brain 2005; 128:932–939.
15. Weiller C, May A, Limmroth V, et al. Brain stem activa-
tion in spontaneous human migraine attacks. Nat Med1995; 1: 658–660.
16. Cao Y, Aurora SK, Nagesh V, et al. Functional MRI-
BOLD of brainstem structures during visually triggeredmigraine. Neurology 2002; 59: 72–78.
17. Arkink EB, Bleeker EJ, Schmitz N, et al. Cerebral perfu-
sion changes in migraineurs: A voxelwise comparison ofinterictal dynamic susceptibility contrast MRI measure-ments. Cephalalgia 2012; 32: 279–288.
18. Sanchez del Rio M, Bakker D, Wu O, et al. Perfusion
weighted imaging during migraine: Spontaneous visualaura and headache. Cephalalgia 1999; 19: 701–707.
19. Woods RP, Iacoboni M and Mazziotta JC. Brief report:
Bilateral spreading cerebral hypoperfusion during spon-taneous migraine headache. N Engl J Med 1994; 331:1689–1692.
20. Furst S. Transmitters involved in antinociception in thespinal cord. Brain Res Bull 1999; 48: 129–141.
21. Moskowitz MA. Basic mechanisms in vascular headache.
Neurologic Clin 1990; 8: 801–815.22. Lempert T, Olesen J, Furman J, et al. Vestibular
migraine: Diagnostic criteria. J Vestib Res 2012; 22:167–172.
23. Choi KD, Oh SY, Park SH, et al. Head-shaking nystag-mus in lateral medullary infarction: Patterns and possiblemechanisms. Neurology 2007; 68: 1337–1344.
24. Lee SH and Kim JS. Benign paroxysmal positional ver-tigo. J Clin Neurol 2010; 6: 51–63.
66 Cephalalgia 34(1)

25. Yang Y, Kim JS, Kim S, et al. Cerebellar hypoperfusionduring transient global amnesia: An MRI and oculo-graphic study. J Clin Neurol 2009; 5: 74–80.
26. Choi KD, Kim JS, Park SH, et al. Cerebellar hyperme-tabolism in paraneoplastic cerebellar degeneration.J Neurol Neurosurg Psychiatry 2006; 77: 525–528.
27. Minagar A, Sheremata WA and Tusa RJ. Perverted
head-shaking nystagmus: A possible mechanism.Neurology 2001; 57: 887–889.
28. Kim JS, Ahn KW, Moon SY, et al. Isolated perverted
head-shaking nystagmus in focal cerebellar infarction.Neurology 2005; 64: 575–576.
29. Huh YE and Kim JS. Patterns of spontaneous and head-
shaking nystagmus in cerebellar infarction: Imaging cor-relations. Brain 2011; 134: 3659–3668.
30. Kattah JC, Talkad AV, Wang DZ, et al. HINTS to diag-
nose stroke in the acute vestibular syndrome: Three-stepbedside oculomotor examination more sensitive thanearly MRI diffusion-weighted imaging. Stroke 2009; 40:3504–3510.
31. Jeong SH, Oh SY, Kim HJ, et al. Vestibular dysfunctionin migraine: Effects of associated vertigo and motionsickness. J Neurol 2010; 257: 905–912.
32. Lee TG, Solomon GD, Kunkel RS, et al. Reversible cere-bellar perfusion in familial hemiplegic migraine. Lancet1996; 348: 1383.
33. Dieterich M and Brandt T. Functional brain imaging ofperipheral and central vestibular disorders. Brain 2008;131: 2538–2552.
34. Lopez C and Blanke O. The thalamocortical vestibular
system in animals and humans. Brain Res Rev 2011; 67:119–146.
35. Kahane P, Hoffmann D, Minotti L, et al. Reappraisal ofthe human vestibular cortex by cortical electrical stimu-lation study. Ann Neurol 2003; 54: 615–624.
36. Bahra A, Matharu MS, Buchel C, et al. Brainstem acti-vation specific to migraine headache. Lancet 2001; 357:1016–1017.
37. Cutrer FM, Sorensen AG, Weisskoff RM, et al. Perfu-
sion-weighted imaging defects during spontaneousmigrainous aura. Ann Neurol 1998; 43: 25–31.
38. Milner PM. Note on a possible correspondence between
the scotomas of migraine and spreading depression ofLeao. Electroencephalogr Clin Neurophysiol 1958; 10: 705.
39. Brandt T, Bartenstein P, Janek A, et al. Reciprocal
inhibitory visual-vestibular interaction. Visual motionstimulation deactivates the parieto-insular vestibularcortex. Brain 1998; 121(Pt 9): 1749–1758.
40. Ambrosini A, de Noordhout AM, Sandor PS, et al. Elec-trophysiological studies in migraine: A comprehensivereview of their interest and limitations. Cephalalgia2003; 23(Suppl 1): 13–31.
41. Aurora SK and Wilkinson F. The brain is hyperexcitablein migraine. Cephalalgia 2007; 27: 1442–1453.
42. Denuelle M, Boulloche N, Payoux P, et al. A PET study
of photophobia during spontaneous migraine attacks.Neurology 2011; 76: 213–218.
43. Boulloche N, Denuelle M, Payoux P, et al. Photophobia
in migraine: An interictal PET study of cortical hyperex-citability and its modulation by pain. J Neurol NeurosurgPsychiatry 2010; 81: 978–984.
Shin et al. 67