aging implicit sequence learning and white matter integrity
TRANSCRIPT

AGING, IMPLICIT SEQUENCE LEARNING, AND WHITE MATTER INTEGRITY
A Dissertation submitted to the Faculty of the
Graduate School of Arts and Sciences of Georgetown University
in partial fulfillment of the requirements for the degree of
Doctor of Philosophy in Psychology
By
Ilana Jacqueline Bennett, M.A.
Washington, DC August 31, 2009

ii
AGING, IMPLICIT SEQUENCE LEARNING, AND WHITE MATTER INTEGRITY
Ilana Jacqueline Bennett, M.A.
Thesis Advisor: Darlene V. Howard, Ph.D.
ABSTRACT
Cognitive neuroscience of aging research investigates neural mechanisms
associated with cognitive stability and decline in older versus younger adults. This
field has been dominated by neuroimaging techniques that emphasize brain function
over structure, gray over white matter, and isolated brain regions over neural networks.
Furthermore, the scope of the field has been limited by a focus on explicit cognitive
processes (e.g., working memory, executive functions). To broaden our current
understanding, this dissertation examined relationships among healthy aging, implicit
sequence learning, and white matter integrity. In the first study, diffusion tensor
imaging (DTI), which measures diffusion of molecular water, was used to characterize
age-related differences in multiple measures of white matter integrity. Results revealed
age-related declines in fractional anisotropy (FA), a measure of diffusion coherence,
with the magnitude of these differences being largest in frontal white matter.
Additional measures of diffusion parallel (axial diffusivity, AD) and perpendicular
(radial diffusivity, RD) to the primary diffusion direction revealed region-specific
patterns of age group differences that may reflect differential aging of microstructural
(e.g., degree of myelination) and macrostructural (e.g., coherence of fiber orientation)
properties of white matter. In the second study, white matter integrity correlates of

iii
implicit sequence learning were examined in younger and healthy older adults. Implicit
sequence learning was assessed with the alternating serial reaction time task (ASRT).
Significant learning was seen in both age groups (i.e., faster and more accurate
responses to frequent, pattern-related events compared to intervening, random events),
with an age-related decline in the late learning stage. In line with functional imaging
studies that identified subcortical (e.g., caudate, hippocampus) and cortical (e.g.,
dorsolateral prefrontal cortex, DLPFC) correlates of implicit sequence learning,
integrity of caudate-DLPFC and hippocampus-DLPFC tracts was related to sequence
learning in the ASRT, and these relationships did not differ with age group.
Additionally, age-related differences in caudate-DLPFC tract integrity mediated age-
related differences in sequence learning. Taken together, results of these studies
support the theory of white matter disconnection, which proposes that age-related
declines in white matter integrity may explain age-related cognitive declines, as
communication between distributed cortical regions involved in the task is disrupted.

iv
ACKNOWLEDGEMENTS
This dissertation would not have been possible without the efforts of many
individuals. First and foremost, I would like to thank the healthy older adults who
participate in our research. These successful agers inspire my work and motivate me to
uncover the neurobiological underpinnings of their cognitive advantage.
Of my many mentors, I am deeply indebted to my academic parents, Drs.
Darlene and Jim Howard. Together, they have provided guidance and support for both
my professional and personal development during the past five years. I am also
thankful to the imaging experts on my committee, Drs. Dave Madden and Chandan
Vaidya, who enabled me to embark on this diffusion tensor imaging project and were
central in helping me obtain an NIH grant. I would like to give additional thanks to Dr.
Madden for his inherent mentoring demeanor that makes every email communication
another invaluable lesson for me. Finally, I would like to acknowledge my early
mentors, Drs. Arnie Starr, Elizabeth Parker, and especially Edward Golob who taught
me to write like a scientist, which made this dissertation a slightly less arduous task.
My friends and family, with their endless love and support, are responsible for
making this journey so enjoyable. Fellow graduate students Alexis Lauricella, Amy
Lowenstein, Jennifer Romano, Jessica Simon, Kelly Barnes, Melanie Stollstorff, and
Samantha Harvell were always on hand for a laugh or a cry. I am grateful to call this
incredible group of budding researchers my friends. I would like to give special thanks
to my lab mate, Jessica Simon, whose inquisitive nature and dedication to her work

v
constantly inspire me to be a better researcher; and to my class mate, Amy Lowenstein,
who persevered through every milestone by my side and always had a mature
perspective that I strived to see. Words cannot express how thankful I am to my
parents, Ivan Bennett and Kathy and Dennis Stajic, for I would not be who I am today
without their influence. Finally, I am grateful for the unwavering support of my sisters,
Amy Bennett and Sasha Stajic, and for my long-time friend, Jamie Brown, who
believes I am a better scientist than I could even aspire to be.

vi
TABLE OF CONTENTS
CHAPTER I: INTRODUCTION............................................................................................. 1
CHAPTER II: AGE-RELATED DIFFERENCES IN MULTIPLE MEASURES OF WHITE MATTER:
A DIFFUSION TENSOR IMAGING STUDY IN HEALTHY AGING........................................... 7
Materials and Methods.................................................................................................. 11
Results…....................................................................................................................... 16
Discussion..................................................................................................................... 25
CHAPTER III: WHITE MATTER INTEGRITY CORRELATES OF IMPLICIT SEQUENCE
LEARNING IN HEALTHY AGING...................................................................................... 38
Method…...................................................................................................................... 42
Results and Discussion................................................................................................. 51
General Discussion....................................................................................................... 61
CHAPTER IV: GENERAL DISCUSSION............................................................................. 66
Future Directions.......................................................................................................... 66
Limitations.................................................................................................................... 69
APPENDIX...................................................................................................................... 72
REFERENCES.................................................................................................................. 75

vii
LIST OF FIGURES
Figure 1. Statistical maps showing white matter clusters with significant age group
differences in fractional anisotropy (FA)...................................................................... 18
Figure 2. Scatterplot showing age group differences in FA as a function of the cluster’s
anterior-posterior location............................................................................................. 25
Figure 3. Population maps showing the three tracts of interest separately for younger
and older adults............................................................................................................. 50
Figure 4. Line graphs showing overall learning performance for mean of median
reaction time and proportion correct in both groups, and bar graphs showing
differences scores across the three stages of learning................................................... 52
Figure 5. Scatterplots showing significant relationships between white matter tract
integrity (FA) and sequence learning............................................................................ 56

viii
LIST OF TABLES
Table 1. Demographic and neuropsychological test results.......................................... 13
Table 2. Descriptions of white matter clusters.............................................................. 19
Table 3. Age group differences in white matter integrity............................................. 20
Table 4. Statistics for age group differences in diffusivity measures........................... 21
Table 5. Descriptions of tractography seed and target regions..................................... 49

1
CHAPTER I: INTRODUCTION
The aging population will increase to ~71 million people over the age of 65
years by 2030, accounting for almost 20% of the total population in the United States
(Goulding, Rogers, & Smith, 2003). Because of this abundance of older adults, there
will be an increased need to promote successful aging and minimize cognitive decline.
Researchers in the cognitive neuroscience of aging are contributing to this goal by
studying how the brain changes with aging and whether these neural changes affect
cognitive functioning. A number of recent books and special issue journal articles have
increased awareness of both cognitive neuroscience of aging (Cabeza, Nyberg, & Park,
2005; Grady, 2008) and the broader field of lifespan cognitive neuroscience (Bialystok
& Craik, 2006; Craik, 2006; Crone & Huizinga, 2006; Muller & Mayes, 2001), which
marks an important step toward advancing our understanding of the neural substrates
underlying cognition and their relation to healthy and pathological development and
aging.
Despite these advances, the scope of cognitive neuroscience of aging research
has been limited in at least two important ways. First, our current understanding has
been restricted by imaging techniques that focus on brain function over brain structure,
gray matter more than white matter, and localized brain regions versus brain systems.
That is, research in this field has been dominated by studies using functional imaging
techniques, including positron emission tomography (PET) and functional magnetic
resonance imaging (fMRI), that assess age-related differences in regional gray matter

2
activity during performance of various tasks (Cabeza, 2001; Langley & Madden,
2000). Studies using other techniques, such as structural magnetic resonance imaging
(sMRI), also examine age group differences in isolated brain regions, relating the
volume of these gray and white matter structures to cognitive performance (e.g., Raz,
Gunning-Dixon, Head, Dupuis, & Acker, 1998). Second, cognitive neuroscience of
aging research has primarily examined a small set of cognitive processes including
working memory, episodic memory, visual perception, and attention (Cabeza, 2001;
Grady, 2008). As a result of this limited focus, certain cognitive functions, such as
implicit forms of learning and memory, have received little attention in reviews of the
cognitive neuroscience of aging (e.g., Grady, 2008; Hedden & Gabrieli, 2004).
The first of these limitations can be avoided by employing a relatively new
structural imaging technique, diffusion tensor imaging (DTI)(Basser, Mattiello, &
LeBihan, 1994; Pierpaoli & Basser, 1996). DTI measures the diffusion, or movement,
of water, which can be used as a proxy for the integrity of the underlying white matter
microstructure. White matter integrity in discrete brain regions can be assessed via
voxel-based comparisons such as tract-based spatial statistics (TBSS)(Smith et al.,
2006; Smith et al., 2007). However, integrity from white matter tracts that connect
distributed brain regions can be examined using fiber tracking, or tractography
methods (Beaulieu, 2002; Jones, 2008); thus, taking neural networks into account. In
healthy aging, the integrity of these tracts is affected by a variety of age-related white

3
matter changes, including atrophy, neuron loss, and demyelination (Gunning-Dixon,
Brickman, Cheng, & Alexopoulos, 2009; Raz, 2005).
The second limitation is readily overcome by employing implicit learning
paradigms. In general, implicit learning describes sensitivity to regularities in the
environment (e.g., repeated sequences of events or spatial patterns) that occurs without
intent to learn or awareness of what has been learned (Seger, 1994). Implicit
acquisition of subtle sequential regularities plays a role in learning a variety of every
day activities (e.g., playing sports, typing) and high-level cognitive skills (e.g.,
language, social intuition). For example, language acquisition initially involves
implicitly learning to extract phonemes from speech streams through repeated
exposure to these word chunks (Kuhl, 2004). The ability to extract predictable
relationships from the environment is also important for adapting to change (e.g., new
people, places, and technologies)(Reber, 1993). In healthy aging, this allows older
adults to respond effectively to their environment in the face of declines in explicit
forms of learning (Craik & Jennings, 1992).
Capitalizing on the limitations of previous cognitive neuroscience of aging
research, the primary aim of this dissertation was to examine relationships among
healthy aging, implicit sequence learning, and white matter integrity. More
specifically, two studies will use DTI to first characterize age-related differences in
multiple measures of white matter integrity (Chapter II), and then assess white matter
integrity correlates of implicit sequence learning in younger and healthy older adults in

4
light of age-related integrity differences (Chapter III). The studies presented here were
conceptualized within the framework of the theory of white matter disconnection (e.g.,
Andrews-Hanna et al., 2007; Charlton et al., 2008; O'Sullivan et al., 2001). According
to this theory, age-related declines in cognitive functioning are due to disrupted
communication between distributed cortical regions involved in a task as a result of
age-related damage to the underlying white matter.
Age group differences in white matter integrity will be examined in discrete
clusters using TBSS analyses (Chapter II) and in white matter tracts using tractography
(Chapter III). In line with previous findings, it is expected that older adults will have
reduced fractional anisotropy (FA), a measure of the uniformity of diffusion
orientation, compared to younger adults, with the magnitude of these age group
differences being largest in anterior white matter (Gunning-Dixon et al., 2009;
Sullivan, Adalsteinsson, & Pfefferbaum, 2006). Additional measures of axial (AD) and
radial (RD) diffusivity, which measure diffusion parallel and perpendicular to the
primary diffusion direction respectively, will also be examined. Research suggests that
these measures are selectively sensitive to changes in specific neural substrates, with
AD reflecting axonal damage or loss and RD reflecting differences in the degree of
myelination (Budde et al., 2007; Nair et al., 2005; Song et al., 2003; Song et al., 2002;
Song et al., 2005). Based on this evidence, previous DTI aging studies have found that
decreased FA in older adults is often accompanied by an age-related increase in RD,
suggesting that compromised white matter integrity in healthy aging may be attributed

5
to loss of myelin in older versus younger adults (Bhagat & Beaulieu, 2004; Madden et
al., 2009; Zhang et al., 2008). However, more recent work suggests that these
interpretations may be misleading, especially in regions with multiple white matter
tracts, and thus should be accepted with caution (Wheeler-Kingshott & Cercignani,
2009).
Implicit sequence learning will be assessed in the second study with the
alternating serial reaction time (ASRT)(Howard & Howard, 1997), a modified version
of the original serial reaction time task (SRT)(Nissen & Bullemer, 1987), in which
participants view a series of stimuli and respond to the location of each event.
Unbeknownst to them, stimuli that follow a repeating sequence of locations alternate
with stimuli that occur in randomly determined locations. Implicit learning is seen as
faster reaction times and/or higher accuracy to pattern-related events compared to
random events, without explicit knowledge of the repeating regularity. Age group
differences in learning are expected based on previous research showing that older
adults learn significantly less than younger adults when subtle regularities are used,
such as those in the ASRT where the alternating structure forces the smallest repeating
units of the pattern to span three trials (Bennett, Howard, & Howard, 2007; Curran,
1997; Howard et al., 2004).
This dissertation will be the first to examine white matter integrity correlates of
implicit sequence learning using DTI. Previous functional imaging and patient research
indicates that implicit sequence learning is mediated by fronto-striatal (Curran, 1995)

6
and medial temporal lobe (Rose, Haider, Weiller, & Buchel, 2002; Schendan, Searl,
Melrose, & Stern, 2003) networks. Therefore, it is expected that the integrity of
separate tracts connecting the caudate and hippocampus to dorsolateral prefrontal
cortex (DLPFC) will be related to sequence learning in the ASRT. These integrity-
sequence learning relationships are predicted to be present in younger and healthy
older adults, consistent with two functional imaging studies that found both age groups
activated similar gray matter regions during implicit sequence learning (Daselaar,
Rombouts, Veltman, Raaijmakers, & Jonker, 2003; Aizenstein et al., 2006). Finally, in
a direct test of the white matter disconnection theory, analyses will assess whether
decreased integrity in older versus younger adults in white matter tracts involved in
implicit sequence learning (i.e., caudate-DLPFC and hippocampus-DLPFC) mediate
age-related deficits in sequence learning.
Taken together, results of the two studies presented in this dissertation will
advance the field of cognitive neuroscience of aging by further characterizing patterns
of healthy age-related decline in white matter integrity, assessing relationships between
white matter integrity and implicit sequence learning in younger and older adults, and
determining whether age group differences in white matter integrity underlie age-
related deficits in implicit sequence learning. These data will be the first to
demonstrate that age-related differences in implicit learning can be explained by white
matter disconnection theories as complex coordination within the underlying neural
networks is compromised by age-related declines in white matter integrity.

7
CHAPTER II: AGE-RELATED DIFFERENCES IN MULTIPLE MEASURES OF WHITE
MATTER INTEGRITY: A DIFFUSION TENSOR IMAGING STUDY
This Chapter has been published as: Bennett, Madden, Vaidya, Howard, and
J.H. Howard, Jr. (2009). Age-related differences in multiple measures of white matter
integrity: A diffusion tensor imaging study of healthy aging. Human Brain Mapping.
Introduction
Brain aging research has been dominated by examinations of age-related
differences in the structure and function of gray matter (Cabeza et al., 2005), with the
other half of the brain—white matter—having been largely ignored. The lack of
attention to white matter aging in the past likely resulted from limitations in imaging
technology, because the relatively recent advent of diffusion tensor imaging (DTI) has
led to widespread in interest in age-related changes in white matter.
DTI is a magnetic resonance imaging (MRI) technique that measures the
diffusion of molecular water (Basser et al., 1994; Pierpaoli & Basser, 1996). Water
diffuses 3-7 times more rapidly along the length of axons aligned in white matter tracts
compared to movement perpendicular to the axons (Le Bihan, 2003; Pierpaoli, Jezzard,
Basser, Barnett, & Di Chiro, 1996) because the latter is restricted by axonal cell
membranes, myelin sheaths, and neurofilaments (Beaulieu, 2002). Various properties
of water diffusion can be calculated from DTI-based eigenvalue measures (λ1, λ2, and
λ3; which indicate the rate of diffusion along the three principal axes of the diffusion
ellipsoid), providing information about the integrity of these white matter structures.

8
Mean diffusivity (MD) and fractional anisotropy (FA) are the two most
commonly used measures of white matter integrity. MD is the average amount of water
diffusion, calculated as the average diffusivity across all three eigenvalues, with higher
values denoting increased rate of diffusion. On the other hand, FA refers to the
coherence of the orientation of water diffusion, independent of rate. It is calculated as
the fraction of total diffusion that can be attributed to anisotropic diffusion, which is
derived from the normalized variances of the three eigenvalues (Basser & Pierpaoli,
1996), with higher values corresponding to a more consistent diffusion orientation.
Taken together, a breakdown in white matter integrity would be seen as higher MD
and/or lower FA.
Accordingly, it is not surprising that the most pervasive findings from DTI
aging research are age-related increases in MD and decreases in FA (Abe et al., 2002;
Grieve, Williams, Paul, Clark, & Gordon, 2007; Head et al., 2004; Hsu et al., 2008;
Hugenschmidt et al., 2008; Pfefferbaum, Adalsteinsson, & Sullivan, 2005;
Pfefferbaum et al., 2000; Salat et al., 2005; Sullivan & Pfefferbaum, 2006; Yoon,
Shim, Lee, Shon, & Yang, 2008). These age differences in integrity are seen
throughout the brain, with most studies reporting the magnitude of the difference to be
larger in anterior white matter compared to posterior regions (Abe et al., 2002;
Ardekani, Kumar, Bartzokis, & Sinha, 2007; Bucur et al., 2008; Grieve et al., 2007;
Gunning-Dixon et al., 2009; Head et al., 2004; Hedden & Gabrieli, 2005; Madden et
al., 2009; Pfefferbaum et al., 2005; Pfefferbaum & Sullivan, 2003; Salat et al., 2004;

9
Salat et al., 2005; Sullivan, Rohlfing, & Pfefferbaum, 2008; Zahr, Rohlfing,
Pfefferbaum, & Sullivan, 2009). This effect, referred to as the anterior-posterior
gradient, has also been seen in the time course of age-related declines in white matter
integrity, with age differences in FA occurring earlier in frontal white matter (Yoon et
al., 2008).
What remains relatively unknown is what these age differences in white matter
integrity mean in terms of underlying neural substrates. With the aim of shedding light
on this issue, DTI aging studies have begun to examine the more frequently used
integrity measures (FA, MD) in relation to axial (AD) and radial (RD) diffusivity,
which correspond to diffusion in the primary (λ1) and perpendicular ((λ2 +λ3)/2)
directions, respectively. Though DTI cannot resolve neural changes within a voxel, this
shift was motivated by animal research indicating that these measures may be
selectively sensitive to specific neural changes, with AD reflecting axonal differences
(e.g., axonal damage or loss)(Budde et al., 2007; Song et al., 2003) and RD reflecting
differences in the degree of myelination (Budde et al., 2007; Nair et al., 2005; Song et
al., 2003; Song et al., 2002; Song et al., 2005).
Previous DTI aging studies that included AD and RD measures have revealed
two prominent patterns of age differences in diffusivity. In the first pattern (Radial
Increase Only), age-related decreases in FA are primarily associated with a significant
increase in RD, but not AD (Bhagat & Beaulieu, 2004; Davis et al., 2009; Madden et
al., 2009; Zhang et al., 2008). In the second pattern (Radial/Axial Increase), age-related

10
decreases in FA are associated with significant increases in both RD and AD (Sullivan
et al., 2006; Sullivan et al., 2008; Zahr et al., 2009). Both patterns are characterized by
an increase in RD with aging. Given the animal literature cited above showing that
increased RD is associated with decreased myelin, and evidence that healthy aging is
accompanied by myelin damage and loss (Peters, 2002), it is not surprising that these
patterns are often interpreted to reflect age-related demyelination. However, because
the focus has been on age differences in RD, potentially important age differences in
AD, and the corresponding neural underpinnings, have been largely ignored.
Shifting attention to age differences in AD may provide insight into a number
of neural substrates underlying age differences in white matter integrity. Diffusion
anisotropy is known to be influenced by both microstructural, cellular variables (e.g.,
axonal packing density, degree of myelination, axonal diameter, and inflammation) and
macrostructural, voxel-level variables (e.g., axonal organization such as aligned,
crossing, and “kissing” fibers)(Giorgio et al., 2008; Mori & Zhang, 2006; Pierpaoli et
al., 1996). Thus, if AD does reflect axonal differences as suggested by the animal
literature cited above, then inferences can be made about the contribution of axon-
based microstructural variables to white matter aging. In addition, we may identify
macrostructural level changes with aging by examining the regional variation of age
differences in AD, because the organization of underlying white matter tracts would
need to be considered.

11
Therefore, the present study assessed region-specific patterns of age differences
in multiple measures of white matter integrity. In order to furthering our understanding
of the neural substrates underlying the widely-reported age differences in FA, younger
and healthy older adults were compared on diffusivity measures (AD and RD; MD was
not included because it is not independent of these measures) in white matter regions
that showed significant age differences in FA, with particularly attention to age
differences in AD. Regional variation of these patterns may reflect differential aging of
the underlying white matter, comparable to studies showing different patterns of aging
across gray matter regions (e.g., Head, Snyder, Girton, Morris, & Buckner, 2005; Raz
et al., 2005). Furthermore, because white matter integrity is compromised (i.e.,
decreased FA) in a variety of populations, including individuals diagnosed with
schizophrenia, multiple sclerosis, and Alzheimer’s disease (Sullivan & Pfefferbaum,
2003; Sundgren et al., 2004), our results may help identify region-specific patterns of
diffusivity that are unique to healthy aging.
Materials and Methods
Participants
Fourteen Georgetown University undergraduate students (18-20 years old) and
14 older adults (63-72 years old) who responded to advertisements in the Washington
Post Health Section were recruited. Demographic and neuropsychological
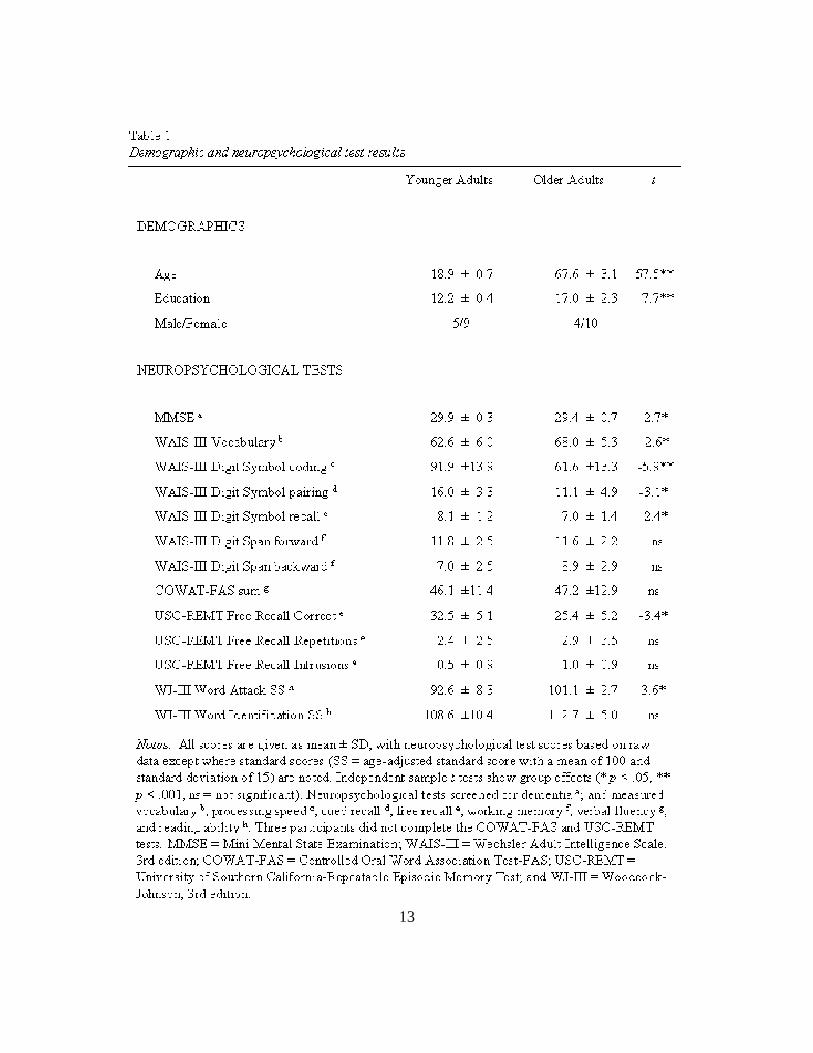
characterizations for each group are presented in Table 1. All participants gave
informed consent, and received either payment or course credit for their participation.

12
The Georgetown University Institutional Review Board approved the experimental
procedures.
Prior to participation, individuals were screened for conditions that would
prevent them from being able to enter the MRI scanner. These conditions included
being pregnant, having ferrous metal implants, having difficulty lying in the supine
position for 30 minutes, and being claustrophobic. In addition, individuals were
excluded prior to scanning if they reported having a health condition or neurological
disease or disorder that is known to influence cognitive functioning and/or contribute
to white matter pathology (e.g., stroke, dementia, diabetes, and uncontrolled depression
or hypertension).
General Procedure
Participants completed three days of testing. On the first day, they completed
screening procedures (informed consent, biographical and health screen questionnaires,
and MRI safety form) and the MRI scanning protocol. On the second and third days of
testing, participants completed the comprehensive neuropsychological test battery (see
Table 1) and a computer-based learning task (data not presented here).
MRI scanning protocol. Participants were scanned using the 3.0 Tesla MRI
system (Siemens Magnetom Trio, Erlangen, Germany) at Georgetown University’s
Center for Functional and Molecular Imaging. An imaging technician positioned
participants in the scanner, laying them in the supine position with a circularly
polarized head coil. A mirror mounted on the head coil allowed them to watch cable
13

14
television programming during scanning. Fitted padding was used to minimize head
movements.
A high resolution T1-weighted structural scan (MPRAGE) was acquired first
with the following parameters: scan time = 7:23 minutes, TR = 2300 ms, TE = 2.94
ms, TI = 900 ms, 9o flip angle, 1 slab, 160 sagittal slices with a 1.0 mm slice thickness,
and FOV = 256 x 256 mm with a 256 x 256 matrix resulting in an effective resolution
of 1.0 mm3 isotropic voxels. A Neurologist reviewed these images, and no participant
exhibited a clinically significant structural abnormality that was atypical for their age
(e.g., lesions, excessive atrophy).
Two 35-direction diffusion weighted echo planar imaging sequences were then
acquired using gradient values of b = 0 and b = 1000 s/mm2 applied in 35 orthogonal
directions. Each acquisition had the following parameters: scan time = 4:39 minutes,
TR = 7700 ms, TE = 100 ms, 55 axial interleaved slices with a 2.5 mm slices thickness
with no gap, and FOV = 240 x 240 mm with a 64 x 64 matrix resulting in an effective
resolution of 2.5 mm3 isotropic voxels. The entire scanning session took approximately
30 minutes.
Data analysis
Diffusion data processing. Diffusion-weighted data were separately processed
for each participant using the University of Oxford’s Center for Functional Magnetic
Resonance Imaging of the Brain (FMRIB) Software Library (FSL) release 4.0
(http://www.fmrib.ox.ac.uk/fsl). First, the two diffusion acquisitions were concatenated

15
in time to increase the signal-to-noise ratio. The first volume within this merged data
file that did not have gradient applied (i.e., the first b = 0 image) was used to generate a
binary brain mask with the Brain Extraction Tool. These data were then corrected for
head movement and eddy current distortions using Eddycorrect, which aligns all the
volumes. Finally, DTIfit was used to independently fit diffusion tensors to each voxel,
with the brain mask limiting the fitting of tensors to brain space. The output of DTIfit
yielded voxelwise maps of FA, AD (λ1), and RD (average of λ2 and λ3) for each
participant.
Between-group t-tests. A between-group skeleton-wise t-test analysis was
performed for the FA measure using Tract-Based Spatial Statistics (Smith et al., 2006).
In this analysis, individual FA maps were non-linearly aligned to the FMRIB58_FA
template, affine transformed into MNI152 1 mm3 standard space, and then averaged
across all participants to form a mean FA image. The mean FA image was used to
generate a white matter skeleton that identifies the center of white matter tracts shared
by all participants. This mean FA skeleton was thresholded at 2000 (corresponding to
FA > 0.2) to exclude voxels containing gray matter or cerebrospinal fluid. Pre-aligned
FA images from each participant were registered to the mean FA skeleton by searching
for maximum FA values perpendicular to the skeleton. This second registration
corrected for any misalignment from the initial nonlinear registration and affine
transformation. The resulting skeletonized FA data were then subjected to a skeleton-
wise (i.e., voxel-wise comparison limited to voxels within the mean skeleton)

16
comparison of FA between younger and older adults. This between-group t-test was
conducted using the Randomise tool, which tests the t value at each voxel against a
null distribution generated from 5000 random permutations of group membership. The
output contained statistical maps corrected for multiple comparisons at the cluster level
(cluster-forming threshold t > 3.0, p < .05).
Significant clusters from the FA analysis were separately masked and labeled
with reference to the JHU ICBM-DTI-81 White Matter Labels, part of the FSL atlas
tools. To obtain mean FA values for each participant from the significant clusters, each
mask was projected back into the native space of each individual’s FA map using the
FSL Deproject tool. Back projected masks were then binarized and multiplied by the
individual’s FA map, leaving just the FA values for voxels within that cluster, which
were then averaged. The same back projection and averaging was conducted for each
of the non-FA maps (λ1, λ2, and λ3) to obtain mean values for each participant for the
significant clusters from the FA analysis. Values for the λ2 and λ3 maps were averaged
to get a single measure of mean RD for each individual.
Finally, between-group t-test analyses were conducted separately for AD and
RD at each of the 28 significant clusters from the FA analysis. Results for each
diffusivity measure were corrected for multiple comparisons using Bonferroni
corrections for 28 comparisons.
Results
Neuropsychological Data

17
Neuropsychological test results, seen in Table 1, revealed the typical pattern of
age effects, with all participants performing within the age-expected range. That is,
older adults performed significantly worse than younger adults on measures of
processing speed, cued recall, and free recall, but not vocabulary or reading ability. All
participants had Mini-Mental State Examination (Folstein, Folstein, & McHugh, 1975)
scores that were ≥ 28.
Age Group Differences in FA
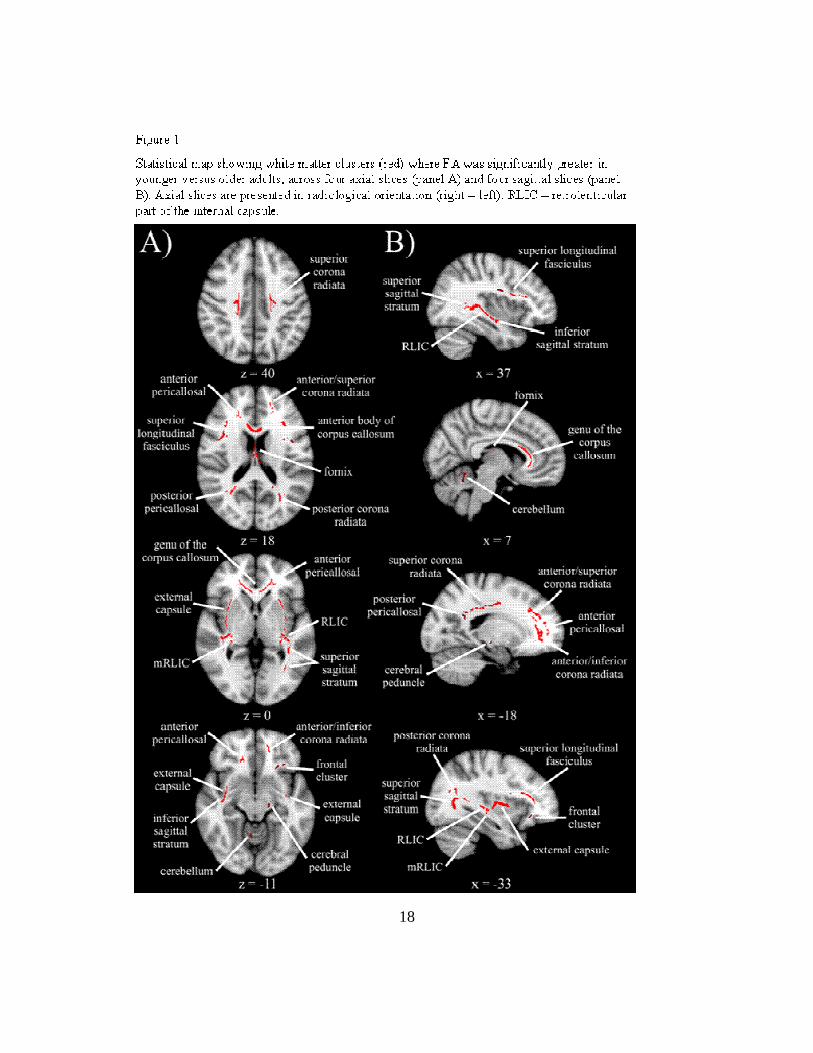
Results of the skeleton-wise between group t-test for FA are presented in
Figure 1. Significantly lower FA was seen in older adults compared to younger adults
in 28 white matter clusters (see descriptions in Table 2), which included two midline,
11 bilateral, and four unilateral regions. Three of the unilateral clusters were in the left
hemisphere, suggesting a slight hemispheric asymmetry, especially in frontal regions
(left anterior/inferior corona radiata, left frontal cluster). Consistent with the notion that
the integrity of white matter connections degrade with age, there were no regions with
significantly higher FA in older versus younger adults.
Age Group Differences in Diffusivity Measures
Between group t-test results for the non-FA, diffusivity measures are
summarized in Table 3, and the corresponding statistics are presented in Table 4. These
data revealed that age-related decreases in FA are characterized by three distinct
patterns of age differences in diffusivity measures. In the first pattern, Radial Increase
Only, clusters with significant age-related decreases in FA also had significant age-
18

19
related increases in RD, but no age group effects in AD. For the second pattern,
Radial/Axial Increase, clusters in which FA decreased with age also had significantly
higher RD and AD values for older adults relative to younger adults. In the third

20
pattern, Radial Increase/Axial Decrease, clusters with significant age-related decreases
in FA also had significantly higher RD and lower AD in older versus younger adults.

21
Age Group Differences in RD across Diffusivity Patterns

22
Significant age-related increases in RD were seen in all clusters that had
significant age differences in FA. To determine whether the magnitude of the age
difference in RD differed across the three diffusivity patterns, an Age Group (younger,
older) x Pattern (Radial Increase Only; Radial/Axial Increase; Radial Increase/Axial
Decrease) ANOVA was conducted for the RD measure. As expected, results revealed a
significant main effect of Age Group, F (1,26) = 65.7, p < .001, indicating that RD was
higher in older (8.52 ± 2.48 x 10-4 mm2/sec) compared to younger (5.39 ± 1.07 x 10-4
mm2/sec) adults. There was also a significant main effect of Pattern, F (2,52) = 224.1,
p < .001, with higher RD in the Radial/Axial Increase pattern (8.52 ± 2.30 x 10-4
mm2/sec) compared to the Radial Increase Only (3.23 ± 0.65 x 10-4 mm2/sec) and
Radial Increase/Axial Decrease (5.16 ± 0.54 x 10-4 mm2/sec) patterns. Importantly, the
Age Group x Pattern interaction, F (2,52) = 43.3, p < .001, revealed that the age-related
difference in RD (older minus younger adults) was significantly larger for the
Radial/Axial Increase pattern (3.78 x 10-4 mm2/sec) compared to the Radial Increase
Only (0.90 x 10-4 mm2/sec) and Radial Increase/Axial Decrease (0.80 x 10-4 mm2/sec)
patterns. A follow-up 2 Age Group x 2 Pattern ANOVA confirmed that the magnitude
of the age difference in RD was comparable in the Radial Increase Only and Radial
Increase/Axial Decrease patterns (p > .20).
Age Group Differences in AD across Diffusivity Patterns
As can be seen in Table 3, the three diffusivity patterns differ with respect to
age differences in AD. That is, clusters either had no significant age differences in AD

23
(Radial Increase Only), significantly higher AD in older versus younger adults
(Radial/Axial Increase), or significantly lower AD in older versus younger adults
(Radial Increase/Axial Decrease). To determine whether the magnitude of the age
difference in AD was significantly different across the three diffusivity patterns, an
Age Group x Pattern ANOVA was conducted for the AD measure. Results revealed
significant main effects of Age Group, F (1,26) = 14.3, p < .001 and Pattern, F (2,52) =
211.1, p < .001, such that AD was higher in older (1.40 ± 0.28 x 10-3 mm2/sec) versus
younger (1.34 ± 0.10 x 10-3 mm2/sec) adults, and higher in the Radial/Axial Increase
pattern (1.61 ± 0.20 x 10-3 mm2/sec) compared to the Radial Increase Only (1.24 ± 0.05
x 10-3 mm2/sec) and Radial Increase/Axial Decrease (1.26 ± 0.05 x 10-3 mm2/sec)
patterns. There was also a significant Age Group x Pattern interaction, F (2,52) = 58.3,
p < .001, which indicated that the age-related difference in AD (older minus younger
adults) was significantly larger for the Radial/Axial Increase pattern (3.15 x 10-3
mm2/sec) compared to the Radial Increase Only (-0.05 x 10-3 mm2/sec) and Radial
Increase/Axial Decrease (-0.08 x 10-3 mm2/sec) patterns. Importantly, a follow-up 2
Age Group x 2 Pattern ANOVA revealed a significant interaction, F (1,26) = 6.8, p <
.02, indicating that the magnitude of the age difference in AD was significantly
different for the Radial Increase Only and Radial Increase/Axial Decrease patterns.
Anterior-Posterior Gradient
There was evidence of an anterior-posterior gradient in the magnitude of the
age-related difference in FA (see Figure 2). A correlation between the percent age

24
group difference in FA (calculated as FA in older adults minus FA in younger adults,
divided by the average FA value) and the location of each cluster (measured as the
average y coordinate) revealed a significant positive relationship, r = .40, p < .04, such
that age-related decreases in FA were larger in more anterior clusters. When analyzed
separately for each hemisphere, the relationship remained positive (left hemisphere, r =
.43; right hemisphere, r = .39), but no longer attained significance (p’s > .13). The
strength of the correlation was magnified when only superior clusters with z
coordinates above or traversing zero were assessed, r = .60, p < .01, whereas it did not
approach significance for inferior clusters, p > .85. This latter result is comparable to
previous research in which significant anterior-posterior gradients of age differences in
FA are reported for superior white matter (e.g., Davis et al., 2009; Pfefferbaum et al.,
2005).
Before concluding that anterior white matter is more susceptible to aging than
posterior regions, it is important to recognize that the age-related decline is regionally
complex. For example, variability of the magnitude of age differences was greater in
posterior clusters, which included regions with both the largest (left inferior sagittal
stratum, right cerebellum) and smallest (bilateral retrolenticular part of the internal
capsule and posterior pericallosal white matter) age group differences in FA. Note that
the fornix was removed from this analysis and from Figure 2 because the percentage of
age-related FA decrease was more than four standard deviations above the average
difference of the remaining white matter clusters.

25
Discussion
Summary of findings

26
The present study examined age group differences in white matter
microstructure by comparing younger and healthy older adults on multiple measures of
white matter integrity. The main finding was that age-related decreases in FA were
associated with three region-specific patterns of age differences in diffusivity measures
(AD and RD).
Radial Increase Only. In most clusters, decreased FA in older adults was
associated with an age-related increase in RD but no age difference in AD. This has
been the predominant pattern reported in several previous DTI aging studies (Bhagat &
Beaulieu, 2004; Davis et al., 2009; Fjell et al., 2008; Madden et al., 2009; Zhang et al.,
2008). The present findings replicated earlier studies that also showed this pattern in
frontal (Zhang et al., 2008), posterior pericallosal (Vernooij et al., 2008; Zhang et al.,
2008), superior longitudinal fasciculus (Madden et al., 2009; Sullivan et al., 2008), and
sagittal stratum (Burzynska et al., 2009) white matter.
Radial/Axial Increase. In other clusters, decreased FA in older adults was
associated with an age-related increase in both RD and AD, which has also been the
primary pattern reported in several other DTI aging studies (Sullivan et al., 2006;
Sullivan et al., 2008; Zahr et al., 2009). The current findings replicated some earlier
reports of this pattern in the genu of the corpus callosum (Burzynska et al., 2009;
Sullivan et al., 2006; Sullivan et al., 2008; Zahr et al., 2009), external capsule
(Burzynska et al., 2009; Sullivan et al., 2008), and fornix (Burzynska et al., 2009;
Sullivan et al., 2008; Vernooij et al., 2008; Zahr et al., 2009). A few studies found that

27
the genu of the corpus callosum (Davis et al., 2009; Madden et al., 2009; Vernooij et
al., 2008) or external capsule (Bhagat & Beaulieu, 2004) fit into the Radial Increase
Only pattern, which may be linked to the possibility that the Radial Increase Only and
Radial/Axial Increase patterns reflect different degrees of severity of the same
underlying neural changes that affect RD.
Radial Increase/Axial Decrease. Remaining clusters with decreased FA in
older adults had an age-related increase in RD and decrease in AD. This pattern was
only recently indentified in another DTI aging study (Burzynska et al., 2009), with two
earlier studies having each observed this pattern in a single white matter region. The
present finding of the Radial Increase/Axial Decrease pattern in frontal white matter
(anterior pericallosal, anterior/superior corona radiata) overlaps with earlier
observations in the superior frontal gyrus (Bhagat & Beaulieu, 2004), superior corona
radiata (Burzynska et al., 2009), and frontal forceps (Burzynska et al., 2009; Sullivan
et al., 2008). This pattern was also previously seen in the retrolenticular part of the
internal capsule (Burzynska et al., 2009). In contrast to the other patterns, possible
neural substrates underlying the Radial Increase/Axial Decrease pattern in healthy
aging have only recently been proposed (e.g., lesion-induced axonal loss and gliosis;
Burzynska et al., 2009).
Age-related increases in RD
In the present study, age-related increases in RD were characteristic of all
clusters in each of the three patterns, suggesting that the neural changes underlying RD

28
increases with healthy aging affect white matter microstructure throughout the brain.
However, the magnitude of the age difference in RD was significantly larger in the
Radial/Axial Increase pattern, indicating that the severity of these neural changes may
be exacerbated in these clusters.
Animal literature has shown RD increases in mice that have been genetically
modified to be myelin deficient (Nair et al., 2005; Song et al., 2002) or treated with
cuprizone to induce myelin loss (Song et al., 2005). Combined with evidence that
myelin damage and loss is prevalent in healthy aging (Peters, 2002), these findings
suggest that age-related demyelination contributes to RD increases in older compared
to younger adults. However, myelin is not the only factor contributing to restricted
diffusion measured by FA because anisotropic diffusion can occur in the absence of
myelin (Beaulieu, 2002; Berman et al., 2005; Le Bihan, 2003; Pierpaoli et al., 1996).
Thus, it may be an oversimplification to conclude that age-related increases in RD are
due solely to demyelination in healthy aging.
Alternatively, some researchers have proposed that the primary determinant of
anisotropy is the packing density of axons within a voxel (Beaulieu, 2002; Shenkin et
al., 2003; c.f., Pierpaoli et al., 1996). Axonal packing density encompasses a variety of
microstructural level variables (e.g., degree of myelination, axonal diameter,
inflammation, etc.) that can influence the amount of extracellular water between axons,
and thus the amount of diffusion in the non-primary direction. One benefit of this
explanation is that it applies to multiple white matter changes known to occur with

29
aging (Gunning-Dixon et al., 2009; Peters, 2002), but that cannot be separately
assessed with the current resolution of most DTI sequences (Sullivan, et al. 2008).
Region-specific differences in the degree of age-related change in axonal
packing density may explain why the age difference in RD was significantly larger in
the Radial/Axial Increase pattern. For example, age differences in microstructural level
variables contributing to decreased axonal packing density may be mild enough in
Radial Increase Only and Radial Increase/Axial Decrease clusters to only increase RD.
However, more severe decreases in axonal packing density from, for example, a
greater loss of myelin or axons in aging, would lead to a global increase in
extracellular water, resulting in larger RD increases and subsequent AD increases in
Radial/Axial Increase clusters. This hypothesis is supported by evidence that the genu
of the corpus callosum, a Radial/Axial Increase cluster, contains a many small
diameter unmyelinated and thinly myelinated axons (Aboitiz, Scheibel, Fisher, &
Zaidel, 1992), the latter of which are highly susceptible to degeneration and
demyelination in aging (Marner, Nyengaard, Tang, & Pakkenberg, 2003; Tang,
Nyengaard, Pakkenberg, & Gundersen, 1997).
Age-related decreases in AD
Previous DTI aging articles that have examined AD and RD tend to focus on
age-related increases in the latter, which, as mentioned above, are often interpreted to
reflect age-related demyelination (e.g., Davis et al., 2009). As a result, the DTI aging

30
literature has largely ignored age differences in AD and its potential underlying neural
substrates.
In the present study, the three observed patterns differed with respect to age
effects for the AD measure, with either no significant age differences in AD (Radial
Increase Only), significantly higher AD in older adults (Radial/Axial Increase), or
significantly higher AD in younger adults (Radial Increase/Axial Decrease). As can be
seen from the statistics presented in Table 4, 80% of the Radial Increase Only clusters
have non-significant age-related decreases in AD. Importantly, however, follow-up
comparisons revealed that the magnitude of the age-related decrease in AD was
significantly larger in the Radial Increase/Axial Decrease pattern compared to the
Radial Increase Only pattern, indicating that these patterns are quantitatively different.
One interpretation of age-related decreases in AD can be inferred from previous
research on the effect of ischemic stroke on DTI-based measures of white matter
integrity in humans (Pierpaoli et al., 2001; Thomalla et al., 2004) and mice (Song et al.,
2003). These studies suggest that axonal degeneration and subsequent gliosis that
follow an ischemic incident or lesion formation lead to a disruption of diffusion
coherence, which decreases diffusion parallel to the primary diffusion direction.
Burzynska, et al. (2009) recently interpreted a Radial Increase/Axial Decrease pattern
in this manner. However, it may not completely explain the present results given that
examination of high resolution structural scans by a Neurologist did not reveal any
clinically significant structural abnormalities. Furthermore, this pattern of diffusivity

31
change is commonly seen following insults in regions with consistent fiber orientation
(Pierpaoli et al., 2001), whereas Radial Increase/Axial Decrease clusters in the present
study often contained projection tracts in regions with many crossing fibers, especially
inferior-to-superior tracts that pass through long range anterior-to-posterior tracts (e.g.,
thalamic radiations in the RLIC, anterior/superior and superior corona radiata, and
anterior pericallosal clusters; and corticospinal tract in superior corona radiata and
cerebral peduncle clusters).
We propose an alternative interpretation: age-related AD decreases may result
from disrupted macrostructural organization. Radial Increase/Axial Decrease clusters
with multiple white matter tracts likely have less coherent diffusion in any given
orientation, and thus decreased AD. On its own, this effect should not differ across age
groups (Head et al., 2004). However, macrostructural organization of these regions
may be disrupted in older adults because of existing age differences in microstructural
variables that affect axonal packing density (e.g., demyelination and axonal
shrinkage)(Stadlbauer, Salomonowitz, Strunk, Hammen, & Ganslandt, 2008; Zhou,
Goto, Otsuka, Moriyama, & Nakamura, 1997), which may magnify the AD decrease in
these regions with aging. In other words, AD in a cluster with crossing fibers would
reflect the average primary diffusion direction of two highly aligned tracts in younger
adults, but the average of two loosely aligned tracts (due to age-related decreased
packing density) in older adults, leading to lower AD in the latter group. Future
research will be necessary to determine whether the Radial Increase/Axial Decrease

32
pattern results from age-related axonal degeneration versus age differences in
macrostructural organization.
Anterior-posterior gradient
Results of the present study support an anterior-posterior gradient, with the
magnitude of the age difference in FA being significantly larger in anterior white
matter clusters. The present findings are in line with histological studies showing that
anterior white matter is more susceptible to age-related alterations on variables
affecting measures of integrity (e.g., myelin loss or damage)(e.g., Peters, 2002), and
studies showing that other changes in the aging brain (e.g., gray matter shrinkage,
neurotransmitter functioning) predominantly affect frontal regions (Raz, 2000; Salat et
al., 2004; West, 1996). Nonetheless, it is important to note that the anterior-posterior
gradient of age differences may be misleading because age differences in white matter
integrity often occur throughout the brain, as seen in the present data (see Figure 1) and
in previous studies (Head et al., 2004; Madden et al., 2004; Salat et al., 2005), with
substantial variability in the magnitude of age differences across regions (see Figure 2).
Limitations
The primary limitation of this study is that DTI, as implemented here and in
most previous studies, cannot assess individual neural substrates that contribute to
anisotropy and diffusivity smaller than the 1-3 mm voxel size, such as degree of
myelination, fiber orientation, and axonal loss. Thus, future studies using either higher
resolution DTI or other techniques that can assess white matter microstructure at the

33
cellular level (e.g., magnetic resonance spectroscopy) will be necessary to validate
interpretations of patterns of age differences in diffusivity, such as those proposed
here.
Nonetheless, certain DTI-based studies may be useful in furthering our
understanding of these patterns. For example, future studies that can incorporate
measures of intervoxel coherence (e.g., lattice index) may be able to assess whether
clusters with decreased FA and AD are in regions with crossing fibers (i.e., in regions
with low lattice index), which may provide additional support for the theory that the
Radial Increase/Axial Decrease pattern reflects age differences in macrostructural
organization in these regions. Similarly, future studies could examine the classification
of various white matter regions longitudinally to provide additional support for the
notion that the patterns reflect varying degrees of severity of underlying neural
changes. For example, such studies may show that at younger ages where white matter
aging is minimal, most regions are classified as the Radial Increase Only pattern,
whereas with increasing age and severity of underlying microstructural changes, more
regions may be classified by the Radial/Axial Increase pattern. The presence of non-
significant trends for age-related increases and decreases in AD in most regions
classified by the Radial Increase Only pattern (see Table 4) indicate that clusters have
the potential to be reclassified over time depending on the progression of underlying
microstructural and/or macrostructral level white matter changes.

34
Another potential limitation of the present study stems from the use of Tract-
Based Spatial Statistics (TBSS). This technique was chosen because it involves a high
quality across-subject registration procedure that relies on a “skeleton” created from
the center of white matter tracts common to all participants (Smith et al., 2006; Smith
et al., 2007), which minimizes alignment issues commonly seen when comparing
groups, such as healthy older versus younger adults, with gross-level brain differences
(e.g., atrophy, ventricular enlargement, sulcal expansion). This approach restricts
analyses to major white matter tracts, thus reducing the area of white matter that could
be identified as having significant age differences in FA. However, it has the benefit of
reducing false identification of group differences because it excludes white matter
regions subject to partial voluming effects due to, for example, gray matter atrophy
(Hugenschmidt et al., 2008) or cerebrospinal fluid (CSF) contamination (Bhagat &
Beaulieu, 2004; Hirsch, Bock, Essig, & Schad, 1999; Papadakis et al., 2002) that can
decrease anisotropy and increase diffusivity measures. Thus, the skeletonizing
technique used here minimized the likelihood that partial voluming effects can explain
the present results, even in Radial/Axial Increase pattern clusters that are surrounded
by gray matter and CSF.
The present analyses restricted age group comparisons of RD and AD to
regions with significant age differences in FA because the primary aim was to make
inferences about the possible neural substrates underlying this commonly observed age
difference in white matter integrity. However, certain patterns of age differences

35
cannot be detected using this approach (e.g., regions with increased RD and/or AD, but
not FA), and potential variations in the pattern of effects across voxels within each
cluster was not assessed. Importantly, the region specificity of patterns identified here
showed substantial overlap with findings from earlier DTI aging studies that used
unrestricted analyses (i.e., voxel-wise comparisons for each diffusivity
measure)(Burzynska et al., 2009; Vernooij et al., 2008; Zhang et al., 2008). Similarly,
when skeleton-wise comparisons were conducted separately for each diffusivity
measure (AD, RD) using the present data (not shown), every Radial/Axial Increase
region continued to show the same pattern, as did the majority of Radial Increase Only
(9/15) and Radial Increase/Axial Decrease (4/9) regions. Together, these results
suggest that limiting analyses to regions with significant age differences in FA had
minimal impact on the generality of the present results.
Conclusions and implications
The present study used the well-established approach of examining AD and RD
in conjunction with FA to assess patterns of age differences in diffusivity and to make
inferences about the underlying neural substrates. Of the nine previous DTI aging
studies that have also assessed age differences in these diffusivity measures, only one
recent report has taken steps to categorize these patterns and discuss the relevance of
age-related AD decreases (Burzynska et al., 2009), complementing the present study.
However, this study is one of the first to propose that a macrostructural level variable
may underlie age differences in white matter integrity.

36
Taken together, results revealed three patterns of age differences in diffusivity
in white matter clusters that had significant age-related decreases in FA. In line with
evidence from animal research, we propose that these three patterns reflect various
combinations of at least two qualitatively different changes in underlying white matter
that differentially affect RD and AD. First, age-related increases in RD, seen in all
white matter clusters in the present study, may reflect an age-related decrease in axonal
packing density from microstructural level variables including demyelination, axonal
loss or damage, and inflammation. These underlying neural changes may be mild in
certain brain regions (Radial Increase Only pattern), whereas other regions that are
more susceptible to aging, such as the genu of the corpus callosum, may have moderate
to severe changes leading to increased RD and AD (Radial/Axial Increase pattern).
Second, age-related decreases in AD, seen in the Radial Increase/Axial Decrease
pattern, may reflect age-related axonal degeneration. However, given that this pattern
primarily occurred in white matter regions with crossing fibers, the age-related
decrease in the coherence of diffusion may instead be due to age-related differences in
macrostructural organization. Results also revealed that age differences in FA were
larger in anterior white matter clusters, consistent with the anterior-posterior gradient
of aging, though differences were seen throughout the brain.
This study takes an important step toward understanding the neural substrates
underlying differences in white matter integrity in healthy aging, though future
research replicating these results and interpretations is necessary. Because many

37
populations are characterized by decreased FA (e.g., schizophrenia, multiple sclerosis,
mild cognitive impairment, and Alzheimer’s disease), regional specificity of the
various patterns of diffusivity may help dissociate changes in white matter integrity
that are unique to healthy aging. Previous research suggests that age differences in FA
are largest and occur earliest in frontal brain regions (i.e., the anterior-posterior
gradient), but it may be possible to identify specific patterns of diffusivity occurring in
specific locations that are characteristic of healthy brain aging, such as increases in all
diffusivity measures occurring earliest in the genu of the corpus callosum.

38
CHAPTER III: WHITE MATTER INTEGRITY CORRELATES OF IMPLICIT
SEQUENCE LEARNING IN HEALTHY AGING
Implicit learning refers to sensitivity to regularities in the environment that
occurs without awareness (Seger, 1994). This ubiquitous process plays a role in skills
ranging from language acquisition (Kuhl, 2004) to social intuition (Lieberman, 2000),
which involve extracting predictable words from speech streams and non-verbal cues
from social interactions, respectively. Implicit learning is essential for adapting to new
people, places, and technologies; particularly in aging as explicit forms of learning
decline (Craik & Jennings, 1992).
The present study focuses on implicit probabilistic sequence learning, a form of
implicit learning that involves extracting regularities from a series of events. This type
of learning can be assessed with the alternating serial reaction time (ASRT) task
(Howard & Howard, 1997), in which participants view four open circles on the screen
and respond to the location of the circle that fills in black. Unbeknownst to
participants, every second stimulus follows a repeating pattern and intervening stimuli
are randomly determined. Sequence-specific learning is seen as faster reaction time
and/or higher accuracy to the predictable, pattern-related events that occur with greater
probability than random events.
Behaviorally, implicit sequence learning is sometimes spared in aging. For
example, older adults learn as well as younger adults in studies using sequences with
simple deterministic structure, where single items (0th order structure) or pairs of items

39
(1st order structure) occur more frequently than others (Curran, 1997; Frensch &
Miner, 1994; Howard & Howard, 1989). However, age-related deficits often appear
when more complex sequences are used, such as those in which the lowest level of
regularity to be learned spans three (2nd order structure) or four (3rd order structure)
consecutive trials (Bennett et al., 2007; Curran, 1997; Howard et al., 2004; Howard &
Howard, 1997). When present, the magnitudes of these age-related differences are
greater in later stages of learning as younger adults continue to learn after older adults
reach a plateau.
Neurobiologically, implicit sequence learning has repeatedly been shown to
involve fronto-striatal networks (Curran, 1995; Doyon et al., 2009; Seger, 2006). This
includes projections between the striatum (caudate, putamen) and frontal regions such
as the dorsolateral prefrontal cortex (DLPFC), and primary, supplementary (SMA), and
premotor areas. There is also evidence that medial temporal lobe (MTL) regions such
as the hippocampus and parahippocampal gyrus (entorhinal, parahippocampal, and
perirhinal cortices) are involved in implicit learning of complex sequences (Rose et al.,
2002; Schendan et al., 2003). Whereas the putamen and frontal motor regions are
thought to be involved in motor demands of the task (e.g., response preparation and
execution), both the caudate and MTL have been implicated in the formation of
associations necessary for learning (e.g., stimulus-stimulus and stimulus-response
associations)(Curran, 1995). Research suggests that these two structures may be
recruited at different stages of implicit sequence learning (Schendan et al., 2003), with

40
the MTL being essential for rapid association formation early in learning and the
caudate being involved in forming associations into later stages of learning (Poldrack
& Packard, 2003).
Previous research has examined neural correlates of implicit sequence learning
in healthy aging. This literature consists of two functional imaging studies that each
observed task-related activity in expected regions for both younger and older adults
(e.g., striatum, frontal regions, hippocampus). However, one study found that the
magnitude of these effects did not differ with age group (Daselaar et al., 2003),
whereas the second study reported an age-related decrease in activity in the prefrontal
cortex and putamen (Aizenstein et al., 2006). Thus, the effect of aging on these brain-
behavior relationships remains unresolved. In addition to these mixed findings, age
group differences in activation patterns for early versus late learning were not assessed.
Together, these earlier examinations of age group differences in the neural
correlates of implicit sequence learning were restricted to isolated cortical and
subcortical gray matter regions (except see Aizenstein et al., 2002). One way to avoid
this limitation is with diffusion tensor imaging (DTI) tractography, which assesses the
integrity of white matter tracts that connect gray matter regions within distributed
neural networks. DTI measures the diffusion, or movement, of molecular water, which
is summarized for each voxel as an ellipsoid where the eigenvalues λ1, λ2, and λ3
indicate the rate of diffusion along its three principal axes (Basser et al., 1994;
Pierpaoli & Basser, 1996). Tractography analyses can then reconstruct white matter

41
tracts, which are assumed to follow the trajectory of the primary diffusion directions
(λ1) of adjacent voxels (see Jones, 2008 for an overview of tractography methods), and
measures of white matter integrity can be calculated from these tracts using the DTI-
based eigenvalues.
Fractional anisotropy (FA) is a commonly used measure of integrity that refers
to the coherence of the orientation of water diffusion, independent of rate. It is
calculated as the fraction of total diffusion that can be attributed to anisotropic
diffusion, which is derived from the normalized variances of the three eigenvalues
(Basser & Pierpaoli, 1996). A single white matter tract with high integrity would have
more consistent diffusion in one orientation, and thus a higher FA value. In healthy
aging, significant decreases in FA are often reported, with the age group differences
being largest in frontal tracts (e.g., Davis et al., 2009; Sullivan et al., 2008). When
accompanied by an age-related decrease in cognitive performance, studies often find
that age group differences in FA from underlying white matter tracts mediate age
group differences in cognition (Madden et al., 2009; Perry et al., 2009; Zahr et al.,
2009). However, previous DTI tractography studies have not examined whether
relationships between tract-based measures of FA and cognitive performance differ
significantly across age groups (i.e., whether these relationships are moderated by age
group).
To address gaps in earlier research, the present study will pursue two aims
regarding relationships between ASRT performance and DTI tractography-based

42
measures of integrity in younger and healthy older adults. The primary aim will be to
examine white matter integrity correlates of implicit sequence learning using
correlations between FA from three bilateral subcortical-cortical tracts and
performance on the ASRT. Given the functions of the gray matter regions these tracts
connect (i.e., association formation versus response preparation and execution), it is
expected that sequence learning will relate to FA in the caudate-DLPFC and
hippocampus-DLPFC tracts, but not the putamen-SMA tract. Step-wise regression
analyses will also be used to determine whether these FA-sequence learning
relationships are moderated by age group. Based on the mixed findings from earlier
functional imaging studies, it is unknown whether the magnitude of these relationships
will vary with age. The secondary aim will separate learning into three stages to assess
age group differences in the time course of learning, and whether white matter integrity
correlates of sequence learning vary with practice. It is expected that larger age group
differences in sequence learning will be seen in the late versus early learning stage, and
that hippocampus-DLPFC tract FA will relate to early learning whereas caudate-
DLPFC tract FA will relate to learning at all stages.
Method
Participants
Fourteen Georgetown University undergraduate students (18.9 ± 0.7 years old,
9 female) and 14 older adults (67.6 ± 3.1 years old, 10 female) who responded to
advertisements in the Washington Post Health Section were recruited.

43
Neuropsychological characterizations for each group are presented in Table 1. All
participants gave informed consent, and received either payment or course credit for
their participation. The Georgetown University Institutional Review Board approved
the experimental procedures.
Prior to participation, individuals were screened for conditions that would
affect their ability to complete the computer-based task (e.g., uncorrected vision,
arthritis, and back problems that would make it difficult to see items presented on the
computer screen or comfortably push the response buttons), prevent them from being
able to enter the MRI scanner (e.g., being pregnant, having ferrous metal implants,
having difficulty lying in the supine position for 30 minutes, and being
claustrophobic), or influence their cognitive functioning and/or contribute to white
matter pathology (e.g., stroke, dementia, diabetes, and uncontrolled depression or
hypertension).
General Procedure
Participants completed three days of testing. On the first day, they completed
screening procedures (informed consent, biographical and health screen questionnaires,
and MRI safety form) and the MRI scanning protocol. On the second and third days of
testing, participants completed the comprehensive neuropsychological test battery (see
Table 1) and a computer-based implicit sequence learning task, the ASRT.
MRI scanning protocol. Participants were scanned using the 3.0 Tesla MRI
system (Siemens Magnetom Trio, Erlangen, Germany) at Georgetown University’s

44
Center for Functional and Molecular Imaging. An imaging technician positioned
participants in the scanner, laying them in the supine position with a circularly
polarized head coil. A mirror mounted on the head coil allowed them to watch cable
television programming during scanning. Fitted padding was used to minimize head
movements.
A high resolution T1-weighted structural scan (MPRAGE) was acquired first
with the following parameters: scan time = 7:23 minutes, TR = 2300 ms, TE = 2.94
ms, TI = 900 ms, 9o flip angle, 1 slab, 160 sagittal slices with a 1.0 mm slice thickness,
and FOV = 256 x 256 mm with a 256 x 256 matrix resulting in an effective resolution
of 1.0 mm3 isotropic voxels. A neurologist reviewed these images, and no participant
exhibited a clinically significant structural abnormality that was atypical for their age
(e.g., lesions, excessive atrophy).
Two 35-direction diffusion weighted echo planar imaging sequences were then
acquired using gradient values of b = 0 and b = 1000 s/mm2 applied in 35 orthogonal
directions. Each acquisition had the following parameters: scan time = 4:39 minutes,
TR = 7700 ms, TE = 100 ms, 55 axial interleaved slices with a 2.5 mm slices thickness
with no gap, and FOV = 240 x 240 mm with a 64 x 64 matrix resulting in an effective
resolution of 2.5 mm3 isotropic voxels. The entire scanning session, which included
three functional runs with a different task (data not reported here), took approximately
45 minutes.

45
Alternating serial reaction time task (ASRT). Participants viewed four black
outlined open circles presented in a row on a 17 in. PC computer monitor. On each
trial, one circle filled in black and participants were instructed to press one of four
buttons that corresponded to the target locations using their dominant hand. If the first
response was incorrect, the target remained on the screen until the correct response was
made. A 120 ms interval separated a correct response on one trial and presentation of
the target on the following trial. Participants completed 45 88-trial blocks, with each
block containing 8 practice trials followed by 10 repetitions of the 8-element sequence.
Short breaks were offered after each block.
Unbeknownst to participants, this ASRT task contained 2nd order sequential
structure, meaning that the location of the target on every other trial was determined by
a repeating sequence, and intervening trials were randomly determined. The six
possible repeating sequences can be presented as 1r2r3r4r, 1r2r4r3r, 1r3r2r4r, 1r3r4r2r,
1r4r2r3r, and 1r4r3r2r; where the number refers to the location of the target from left to
right, and ‘r’ refers to a random trial where the target could occur at any of the four
locations. Each participant was randomly assigned one of these repeating sequences.
Participants were instructed to respond to each target as quickly as possible. As
older adults tend to make fewer errors than younger adults, which complicates age
comparisons, differential feedback previously shown to match the groups on overall
accuracy (Bennett et al., 2007; Negash, 2003) was used to maintain performance at
92% accuracy. The feedback display included mean reaction time and accuracy scores

46
for a given block for younger adults, but only mean reaction time scores for older
adults. Depending on their mean accuracy for a given block, a statement also prompted
participants to “focus more on speed” (> 93% for younger and 90% for older adults) or
“focus more on accuracy” (< 91% for younger and 80% for older adults). Feedback
stated “speed and accuracy are about right” when mean accuracy scores were between
91-93% for younger adults and 80-90% for older adults.
Explicit awareness. Two tests of explicit awareness were used. First, a brief
post-experiment interview assessed explicit knowledge of the repeating sequence with
questions ranging from general (“Do you have anything to report about the task or
stimuli?”) to specific (“Did you notice a pattern, and if so, can you describe it to me?”).
Participants then completed a card sort task in which they sorted cards depicting all
possible combinations of three consecutive trials (three rows of four circles, with one
filled in on each row representing the target for that trial) into piles indicating whether
that triplet occurred more often or less often.
ASRT data analysis
Sequence learning is thought to result from individuals learning the relative
frequency of the smallest repeating units of a sequence rather than the alternating
pattern-random structure (Howard et al., 2004; Perruchet, Gallego, & Savy, 1990).
Thus, for ASRT sequences with second-order structure, participants learn that certain
three-element sequence chunks, or runs of three consecutive trials referred to as
triplets, occur more frequently that others. High frequency triplets are those in which

47
the first and third elements are part of the repeating pattern (e.g., 112, 122, 132, 142 for
the sequence 1r2r3r4r), whereas low frequency triplets occur with lower probability
because they do not fall on pattern trials (e.g., 113 and 124). Sequence learning will
therefore be assessed with triplet type effects, where performance on high and low
frequency triplets is compared for reaction time and accuracy measures. As in previous
research, certain low frequency triplets, repetitions (e.g., 111 and 222) and trills (e.g.,
121 and 232), will be omitted because responses to them are influenced by preexisting
response biases, rather than learning per se (Howard et al., 2004).
Diffusion data analysis
Pre-processing. Diffusion-weighted data were separately processed for each
participant using the University of Oxford’s Center for Functional Magnetic
Resonance Imaging of the Brain (FMRIB) Software Library (FSL) release 4.0
(http://www.fmrib.ox.ac.uk/fsl). First, the two diffusion acquisitions were concatenated
in time to increase the signal-to-noise ratio. The first volume within this merged data
file that did not have gradient applied (i.e., the first b = 0 image) was used to generate a
binary brain mask with the Brain Extraction Tool. These data were then corrected for
head movement and eddy current distortions using Eddycorrect, which aligns all the
volumes. Finally, DTIfit was used to independently fit diffusion tensors to each voxel,
with the brain mask limiting the fitting of tensors to brain space. The output of DTIfit
yielded voxelwise maps of FA for each participant.

48
Tractography. Probabilistic fiber tracking was conducted separately in each
participant using FMRIB’s Diffusion Toolbox (FDT)(Behrens et al., 2003). First,
diffusion parameters were estimated at each voxel using BEDPOSTX, in which
Markov Chain Monte Carlo sampling generates a probability distribution function of
the principle diffusion direction. ProbtrackX was then used to estimate the distribution
of connections between seed and target regions (see Table 5). To generate this
connectivity distribution, 10,000 streamline samples were initiated from all voxels
within the seed regions, traveling along the probability distribution functions of local
voxels (steplength = 0.5 mm, curvature threshold = 0.2), until they terminated in voxels
within the target regions. The output contains a connectivity value for each voxel that
represents the number of streamline samples that passed through that voxel.
Three bilateral tracts of interest were generated using the seed and target
regions described in Table 5. These included caudate-DLPFC (dorsolateral prefrontal
cortex), hippocampus-DLPFC, and putamen-SMA (supplementary motor area). Seed
and target regions were masked in standard space (MNI152 2 mm3 standard brain),
which has been shown to be a highly reproducible way of generating tracts (Heiervang,
Behrens, Mackay, Robson, & Johansen-Berg, 2006), and a midline exclusion mask (at
x = 0) was used to restrict tracking to the left or right hemisphere. In order to conduct
tractography in diffusion space, Flirt was used to register the masks from standard
space to each individual’s diffusion space using a transformation with 6 degrees of
freedom. Resulting tracts were thresholded at 15% of the maximum connectivity value,

49
leaving only those voxels with a high likelihood of being connected to the seed
regions. To illustrate these tracts (see Figure 3), population maps were created by using
the reverse transformation to convert thresholded tracts from diffusion space into
standard space, and then overlaying tracts from each individual onto the standard
MNI152 2 mm3 brain.
Tract-based measures of integrity. Mean FA values were calculated for the
three bilateral tracts of interest for each participant. Each thresholded tract was
binarized and multiplied by the individual’s FA map, leaving just the FA values for

50
voxels within that tract, which were then averaged. All tracts were visually checked for
quality. Due to significant distortions (e.g., pixilated, non-continuous tracts; branching
to non-target regions) some individual’s tracts were excluded from analyses of the left
caudate-DLPFC (two younger, one older), right caudate-DLPFC (two older), left and

51
right hippocampus-DLPFC (one younger), and right putamen-SMA (one younger, two
older) tracts.
Results and Discussion
Neuropsychological profiles
The neuropsychological test results in Table 1 revealed the typical pattern of
age effects, with all participants performing within the age-expected range. That is,
older adults performed significantly worse than younger adults on measures of
processing speed, cued recall, and free recall, but not vocabulary or reading ability. All
participants had Mini-Mental State Examination scores (Folstein et al., 1975) that were
≥ 28.
ASRT performance
Sequence learning. For each participant, median reaction times for correct trials
and percentage of correct responses were calculated separately for each triplet type for
each block, and then averaged into nine, five-block epochs. Mean of median reaction
times and mean accuracy scores were analyzed separately in Age Group (Young, Old)
x Triplet Type (High Frequency, Low Frequency) x Epoch (1-9) repeated measures
ANOVAs, with Age Group as a between-subject variable, and Triplet Type and Epoch
as within-subject variables. These data are presented in Figure 4 (Overall
Performance).
For reaction time, skill learning was seen as a significant main effect of Epoch,
F (8, 208) = 33.8, p < .001, in which responses became faster with practice. Sequence
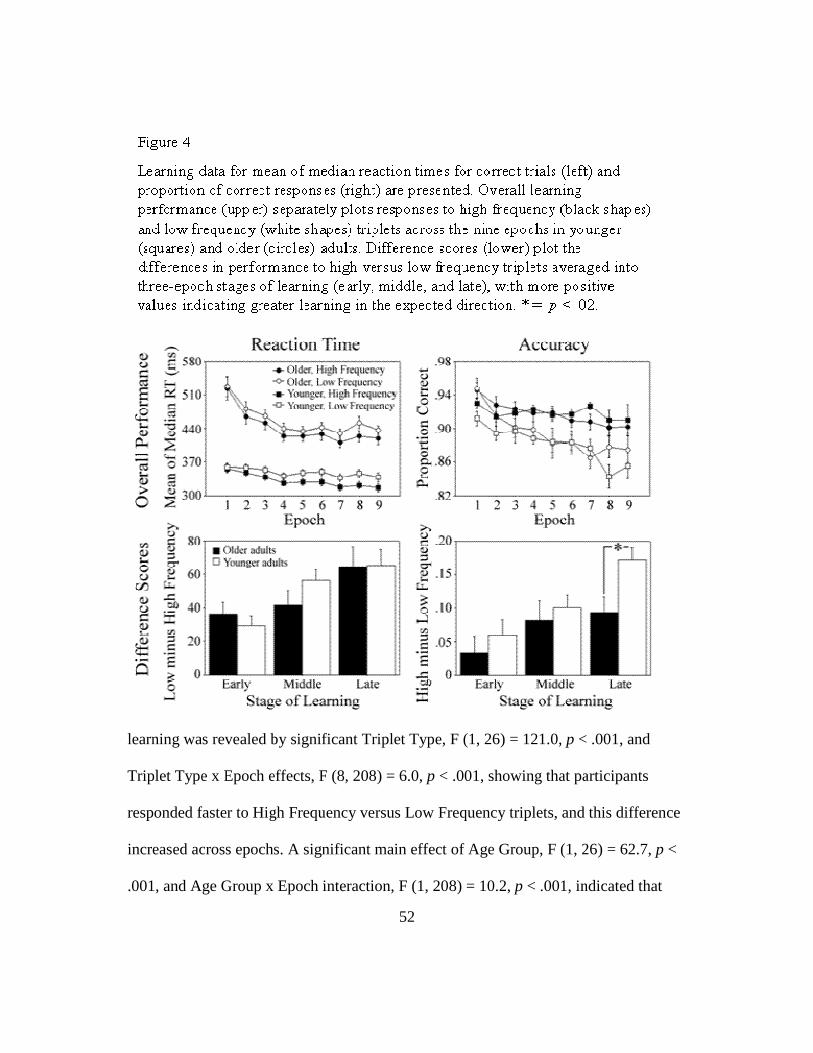
52
learning was revealed by significant Triplet Type, F (1, 26) = 121.0, p < .001, and
Triplet Type x Epoch effects, F (8, 208) = 6.0, p < .001, showing that participants
responded faster to High Frequency versus Low Frequency triplets, and this difference
increased across epochs. A significant main effect of Age Group, F (1, 26) = 62.7, p <
.001, and Age Group x Epoch interaction, F (1, 208) = 10.2, p < .001, indicated that

53
responses of older adults were slower and increased more with practice compared to
younger adults. However, in contrast to our predictions, there were no group
differences in sequence learning as seen by non-significant Age Group x Triplet Type
and Age Group x Triplet Type x Epoch interactions, p’s > .45.
Similar results were seen for accuracy. A significant main effect of Epoch, F (8,
208) = 9.4, p < .001, revealed skill learning as a decrease in accuracy with practice,
which is typical in sequence learning tasks as participants make increasingly more
errors on pattern-inconsistent trials as they learn the regularity. Sequence learning was
again observed as significant Triplet Type, F (1, 26) = 48.8, p < .001, and Triplet Type
x Epoch effects, F (8, 208) = 3.5, p < .001, with participants responding more
accurately to High Frequency versus Low Frequency triplets, and this difference
increasing across epochs. There were no significant effects with Age Group, p’s > .12,
indicating that there were no age group differences in sequence learning, and that the
differential feedback was successful in equating both age groups on overall accuracy.
Implicitness. Self-reports from the post-experiment interview did not reveal any
explicit awareness. Five younger adults and five older adults reported noticing a
regularity in the location of target presentations, but no individual was able to
accurately describe the nature of the regularity. Similarly, participants did not
recognize that High Frequency triplets occurred more often than Low Frequency
triplets. Mean recognition ratings were calculated for each participant as the proportion
of times each triplet type was reported as having occurred “frequently” during the

54
recognition task. An Age Group (Young, Old) x Triplet Type (High Frequency, Low
Frequency) mixed design ANOVA showed a significant main effect of Triplet Type, F
(1, 26) = 347.2, p < .001, with Low Frequency triplets (0.67 ± 0.05) being rated as
occurring more often than High Frequency triplets (0.33 ± 0.05) across age groups.
Taken together, these behavioral data revealed implicit sequence learning in
younger and older adults as faster and more accurate responses to high frequency
versus low frequency triplets, with no explicit awareness of this regularity in either age
group. In contrast to our predictions based on earlier work (Curran, 1997; Howard et
al., 2004; Howard & Howard, 1997), this analysis did not reveal significant age group
differences in sequence learning. One advantage of this unexpected finding is that any
age-related differences in white matter integrity correlates of implicit sequence
learning cannot be attributed to group differences in the magnitude of learning.
However, this also results in one limitation of the present study in that certain results,
such as finding that age group does not moderate FA-sequence learning relationships
(see below), may be limited to groups with equivalent learning.
Age group differences in white matter integrity
Separate between-group t-tests compared FA in younger and older adults for
each tract of interest. Significant age group differences were seen in the left, t (23) = -
2.82, p < .01, and right, t (24) = -3.71, p < .01, caudate-DLPFC tracts, with higher FA
in younger adults compared to older adults (left young, 0.31 ± 0.05; left old, 0.25 ±

55
0.06; right young, 0.32 ± 0.04; right old, 0.25 ± 0.04). FA did not differ across groups
in bilateral hippocampus-DLPFC or putamen-SMA tracts, p’s > .13.
The age-related decrease in caudate-DLPFC tract integrity is consistent with
previous DTI tractrogaphy studies that report age group difference in FA in frontal
white matter tracts (e.g., Davis et al., 2009; Sullivan et al., 2008). It also fits with other
known changes in the aging brain that primarily affect frontal and striatal regions. For
example, structural imaging studies show robust age-related shrinkage of prefrontal
and striatal gray matter, functional imaging studies find hyper- and hypo-activation of
frontal regions in aging, and neurochemical studies report significant age-related
disruption of neurotransmitter systems associated with the frontal-striatal network (see
Hedden & Gabrieli, 2005 for review). For the hippocampus-DLPFC tract, the lack of
an age group difference is in line with findings that MTL structures are relatively
unaffected in healthy aging (Head et al., 2005; Hedden & Gabrieli, 2005).
White matter integrity correlates of sequence learning
Correlations were performed between FA from the tracts of interest and triplet
type learning scores for reaction time and accuracy, calculated as the difference in
performance on high versus low frequency triplets (low minus high for reaction time,
high minus low for accuracy) summed across all epochs. To maximize power for
detecting these effects, all participants were included regardless of age group.
Significant effects are presented in Figure 5.

56
As expected, left caudate-DLPFC tract FA was positively correlated with triplet
type learning scores for reaction time, r = +0.49, p < .02, and accuracy, r = +0.44, p <
.03; and right hippocampus-DLPFC tract FA was significantly related to triplet type
learning scores for reaction time r = +0.41, p < .04. There was also a trend between the
accuracy learning score and left putamen-SMA tract FA, p < .09. Remaining effects
did not attain significance, p’s > .23.

57
Age group does not moderate FA-sequence learning relationships. Stepwise
regression analyses assess whether relationships between FA and ASRT sequence
learning were different in younger versus older adults. Separate analyses were
conducted using triplet type learning scores for reaction time and accuracy as outcome
variables. The predictor variable age group was entered into the model first, followed
by FA from the tracts of interest (left caudate-DLPFC and right hippocampus-DLPFC
for reaction time, left caudate-DLPFC for accuracy), and the interactions between age
group and FA.
Results revealed that only left caudate-DLPFC tract FA remained a significant
predictor of sequence learning for reaction time, F (1, 24) = 7.30, r = +0.49, p < .02,
and accuracy, F (1, 24) = 5.47, r = +0.49, p < .03. The interaction term was not a
significant predictor of sequence learning for either outcome measure, indicating that
age group did not moderate the FA-sequence learning relationships. That is, the
magnitude of these relationships was similar in younger and older adults.
Controlling for overall reaction time. The impact of response speed on FA-
sequence learning relationships was assessed using a measure of overall reaction time
in the ASRT (mean reaction time on all trials). Correlation analyses revealed that
overall reaction time was significantly related to sequence learning for the accuracy
measure, r = -.44, p < .02, and FA in the left, r = -.53, p < .01, and right, r = -.42, p <
.04, caudate-DLPFC tract. Relationships for reaction time and remaining tracts were
not significant, p’s > .11.

58
Follow-up multiple regression analyses showed that, when controlling for
overall reaction time, left caudate-DLPFC FA was no longer a significant predictor of
sequence learning for the accuracy measure, p > .30, though the overall model
remained significant, F (2, 24) = 5.14, p < .02, with overall reaction time as a
marginally significant predictor, r = -.42, p < .06. Importantly, FA in the left caudate-
DLPFC and right hippocampus-DLPFC tracts remained marginally significant
predictors of sequence learning for the reaction time measure after controlling for
overall reaction time, p’s < .06, indicating that these learning effects are independent of
overall speed.
Stages of sequence learning and relation to white matter integrity
Age group differences in later learning. Following from earlier behavioral
research indicating that age group differences in sequence learning are largest at later
stages (e.g., Howard et al., 2004), and the observation that younger adults appeared to
show more learning in the last few epochs on the accuracy measure compared to older
adults, the present study assess age group differences in the time course of learning by
separating sequence learning into three stages (early, middle, and late). Between-group
t-tests compared triplet type effects for younger and older adults at each learning stage.
For each participant, the difference in performance on high versus low frequency
triplets was calculated separately for each epoch, and then summed into three, three-
epoch time periods. These data are presented in Figure 4 (Difference Scores).

59
There were no significant age-related differences in sequence learning at any
stage for reaction time, p’s > .18, and the early and middle learning stages for
accuracy, p’s > .45. However, as expected, there was a significant age-related decrease
in late learning for accuracy, t (26) = -2.65, p < .02, indicating that younger adults
(0.17 ± 0.07) learned more than older adults (0.09 ± 0.09) later in practice. This age
group difference cannot be attributed to older adults being more cautious (i.e., making
fewer errors) with practice, because as Figure 4 reveals, younger and older adults have
nearly identical accuracy on the high frequency triples.
White matter integrity correlates of sequence learning stages. Previous
functional imaging research suggested that the hippocampus is engaged early in
learning whereas the caudate is engaged throughout learning (Poldrack & Packard,
2003; Schendan et al., 2003). To assess whether white matter integrity correlates of
sequence learning also vary with stages of learning, correlation analyses were
conducted between FA and triplet type learning effects for reaction time and accuracy
at each learning stage.
Results revealed that, for reaction time, learning in the middle stage was
positively correlated with FA in the left caudate-DLPFC, r = +0.44, p < .01, and right
hippocampus-DLPFC, r = +0.44, p < .03, tracts. For accuracy, the late learning stage
was significantly related to FA in the left caudate-DLPFC tract, r = +0.62, p < .01, and
marginally significant in the right caudate-DLPFC tract, r = +0.38, p < .06. These
findings are generally consistent with predictions that integrity of the hippocampus-

60
DLPFC tract would relate to early learning, integrity of the caudate-DLPFC tract
would relate to early and later stages of learning, and putamen-SMA tract FA would
not be related to sequence learning. Note that correlations with sequence learning in
the early stage may not have been detected because the magnitude of the learning
scores were significantly lower in the early versus middle and late stages for both
reaction time (32.8 versus 49.0 and 65.0) and accuracy (0.05 versus 0.09 and 0.13).
Age group differences in later learning are mediated by age group differences
in FA. The previously reported age-related decrease in the late learning stage for
accuracy, in combination with the age-related decrease in FA in the left and right
caudate-DLPFC tracts, suggest that age group differences in FA may mediate age
group differences in ASRT sequence learning. To test this, hierarchical regression
analyses were conducted separately for these two tracts in accordance with the
procedures laid out by Baron and Kenny (Baron & Kenny, 1986).
When age group was the sole predictor of the late learning score for accuracy,
21.3% of the variance in learning was age-related. When FA in the left caudate-
DLPFC tract was entered before age group, FA remained a significant predictor,
accounting for 41.2% of the variance, but age group no longer accounted for a
significant portion of the variance. To calculate the degree to which FA attenuated the
amount of variance in ASRT sequence learning that can be explained by age, the
amount of variance uniquely associated with age group (after partialling out the effect
of FA in the left caudate-DLPFC tract) was subtracted from the amount of variance

61
associated with age group as the sole predictor, and then divided by the amount of
variance associated with age group as the sole predictor (see Salthouse, 1991). This
procedure revealed that FA attenuated age-related variance in sequence learning by
93.0%.
The same analysis for FA in the right caudate-DLPFC tract revealed that
neither FA nor age group remained significant predictors of the later learning score for
accuracy when they were both entered into the model. Thus, FA in this tract was not a
significant mediator of the age group-sequence learning relationship.
General Discussion
The present study was the first to use DTI tractography to examine tract-based
white matter integrity correlates of implicit sequence learning, and to assess whether
these relationships were moderated by healthy aging. The main result from our primary
aim was that FA in the caudate-DLPFC and hippocampus-DLPFC tracts, but not the
putamen-SMA tract, was related to sequence learning in the ASRT. Importantly, the
relationships between FA and sequence learning were not moderated by age group in
that they did not differ for younger and older adults. For our secondary aim that
involved separating learning into three stages, results revealed significant age-related
decreases in late learning (for accuracy). In addition, FA in the hippocampus-DLPFC
tract was related to earlier learning (middle learning stage), and FA in the caudate-
DLPFC tract was related to earlier and later learning (middle and late learning stages).

62
Further analyses revealed that the age-related decrease in sequence learning was
mediated by the age-related decreases in caudate-DLPFC tract integrity.
Of central importance here was whether sequence learning in the ASRT was
related to white matter integrity from tracts connecting discrete gray matter regions
that were previously identified as being involved in the task. Using a different measure
of white matter integrity, one earlier study had found that one measure of implicit
learning (skill learning) was related to the degree of white matter hyperintensity (i.e.,
white matter lesion) burden in older adults (Aizenstein et al., 2002). The present study
expanded these findings by examining relationships between sequence-specific
learning and white matter integrity in younger and older adults, and using tract-based
measures of integrity that are more sensitive to specific white matter regions
(compared to periventricular and deep white matter).
Results revealed that sequence learning was significantly related to integrity of
the caudate-DLPFC and hippocampus-DLPFC tracts, with greater sequence learning
relating to higher integrity. These findings may be due to the role of the caudate and
hippocampus in forming learning-related associations (Curran, 1995), such as
associations among the locations of stimuli in frequently occurring triplets, or between
a stimulus location and the corresponding motor response. In support of this view,
hippocampus-DLPFC tract integrity was related to early learning, whereas caudate-
DLPFC integrity was related to learning at early and late stages, which is consistent
with evidence that the hippocampus is engaged in forming rapid associations early in

63
learning, whereas the caudate is involved in forming associations throughout learning
(Poldrack & Packard, 2003; Schendan et al., 2003). The marginally significant
relationship between sequence learning and FA in the putamen-SMA tract may reflect
this network being involved in response selection based on sequence-specific implicit
knowledge (Curran, 1995).
Importantly, the white matter integrity correlates of sequence learning did not
differ for younger and older adults. Finding that age group did not moderate these FA-
sequence learning relationships is consistent with one functional imaging study that
found similar gray matter regions activated during implicit sequence learning
performance in younger and older adults (Daselaar et al., 2003), though a comparable
study found different degrees of activation across age groups (Aizenstein et al., 2006).
Together, these data suggest that younger and older adults are calling on the same
neural networks during sequence learning, with this study providing the first evidence
that sequence learning depends on the integrity of fronto-striatal (caudate) and MTL
networks.
One of the most interesting findings from the present study was that age-related
decreases in FA in the left caudate-DLPFC tract mediated age-related decreases in the
ASRT late learning score for accuracy. In fact, integrity of this caudate-DLPFC tract
attenuated the amount of variance in sequence learning that could be attributed to age
group by 93%. This finding is consistent with the notion that age-related declines in
implicit sequence learning may be due to healthy brain aging of the underlying frontal

64
and striatal regions (see Hedden & Gabrieli, 2005), with the present study providing
the first evidence for the role of fronto-striatal white matter integrity mediating age-
related differences in implicit sequence learning. One explanation for this mediating
relationship is that age-related changes in white matter (e.g., axonal loss or degree of
myelination) may alter communication between distributed gray matter structures
within a neural network, for example by disrupting the timing of neural signals, which
in turn affects higher-level cognitive processes that depend on these communications
(e.g., Andrews-Hanna et al., 2007; Nordahl et al., 2006; Sullivan & Pfefferbaum,
2006).
Conclusions and future directions
In summary, the present results complement functional imaging and patient
studies of gray matter substrates involved in implicit sequence learning, finding that
the integrity of white matter tracts connecting these distributed regions (caudate-
DLPFC, hippocampus-DLPFC) is significantly related to sequence learning in the
ASRT. Furthermore, the present results provide evidence that neural substrates of
implicit sequence learning do not vary with healthy aging. Future research combining
DTI and functional imaging will be central to advancing our understanding of the
interaction between white matter integrity and gray matter activity within the neural
networks underlying implicit sequence learning. Such research would also improve
interpretations of the mediating role of white matter integrity, identifying whether age-
related declines in white matter integrity are associated with age-related differences in

65
functional activity, which in turn lead to decreased performance in older versus
younger adults.

66
CHAPTER IV: GENERAL DISCUSSION
Two studies were presented that characterized age-related differences in
multiple measures of white matter integrity (Chapters II and III), identified white
matter integrity correlates of implicit sequence learning in younger and healthy older
adults (Chapter III), and determined that age-related integrity differences mediated
age-related differences in implicit sequence learning (Chapter III). Together, these
findings can be interpreted in light of the white matter disconnection theory (e.g.,
Andrews-Hanna et al., 2007; Charlton et al., 2008; O'Sullivan et al., 2001). That is,
white matter integrity decreases were seen in healthy older adults compared to younger
adults. In turn, this compromised white matter may have disrupted the quality or timing
of communication between distributed cortical regions within the neural networks
involved in the ASRT (e.g., fronto-striatal network). As a result, performance was
affected as seen by significantly less sequence learning in the ASRT in later stages of
learning for older versus younger adults.
Future Directions
In contrast to the vast majority of cognitive neuroscience of aging research that
has focused on age-related differences in functional activity in discrete gray matter
regions during performance of explicit cognitive tasks, the present studies emphasized
age group differences in white matter tracts and their relation to implicit learning
performance. In addition to broadening the scope of cognitive neuroscience of aging

67
research, these data will stimulate future research that addresses key issues in the field,
and research aimed at fostering successful aging.
One prominent issue in cognitive neuroscience of aging research is qualifying
the relationship between brain structure and brain function, especially in the context of
age-related cognitive stability and decline. Functional imaging studies have revealed
both greater and less activity in older adults compared to younger adults across a
variety of tasks (Grady, 2008; Reuter-Lorenz, 2002). These under-activations are
thought to reflect inadequate recruitment of relevant brain regions, which are often
associated with age-related performance declines, whereas over-activations may reflect
compensatory activity (Cabeza, 2002; Reuter-Lorenz & Cappell, 2008) or non-
selective recruitment/inefficient processing (Logan, Sanders, Snyder, Morris, &
Buckner, 2002; Rypma et al., 2006), especially in the absence of age-related
differences in performance. The various functional activity-behavior relationships are
further thought to be mediated by age differences in underlying brain structure. For
example, one review proposed that certain age-related white matter changes (e.g.,
demyelination) may lead to mild disruptions of communication between brain regions
that may be compensated for by increased functional activity and thus improved
performance, whereas other changes (e.g., white matter hyperintensities, WMH) may
lead to more severe disruptions that would decrease activity and performance (Grady,
2008).

68
Future cognitive neuroscience of aging research should aim to integrate brain
function, structure, and cognitive performance to further our understanding of these
complex relationships. In the present study, such an undertaking would have been
invaluable for providing information about relationships between brain structure and
function in implicit learning. Follow-up projects may be able to determine whether the
age-related decrease in the integrity of the caudate-DLPFC tract is associated with an
age-related decrease (under-recruitment) or increase (non-selective recruitment) in
frontal activity. This may determine whether the sequence learning deficit in older
versus younger adults is due to inadequate recruitment of the same networks engaged
by younger adults (e.g., fronto-striatal and MTL networks examined here), or whether
older adults are also calling on additional brain regions (networks not examined here).
Of further interest is whether compensatory structure-function relationships would be
limited to frontal brain regions, which showed the largest age group differences in
integrity, or whether integrity differences that were seen throughout the brain (see
Chapter II) affect global patterns of brain activity.
Research aimed at promoting successful aging may also benefit from the
present studies. As mentioned earlier, implicit learning supports independent living by
enabling people to adapt to change, including new settings, people, and technologies.
Although implicit sequence learning declines are seen in healthy older adults in later
stages of learning, these implicit learning systems remain somewhat intact in healthy
and pathological aging. Thus, implicit learning tasks and their underlying neural

69
networks may be targeted by training programs aimed at maximizing successful aging
and recovery after trauma or stroke, which are frequent in old age (e.g., Camp, Bird, &
Cherry, 2000).
Limitations
Two main limitations of the present data were the relatively small sample size,
and the inclusion of high functioning older adults. Small sample sizes reduce the
ability to detect significant effects. This may explain why certain well-defined results,
such as age-related decreases in high-order sequence learning, were not readily
detected in the second study presented in Chapter III. It may also underlie the non-
significant interaction from the moderation analysis in Chapter III. That it, rather than
the null result reflecting that age group did not moderate the FA-learning relationships,
it may be that we did not have enough power to detect this effect. This limited power
issue may also be due to the high functioning older adults examined here, who showed
substantial overlap in their sequence learning scores with the younger group. These
older individuals may affect the generalizability of the present results because their
performance and underlying white matter may not be characteristic of “normal” or low
functioning older individuals.
Additional limitations stem from the fact that there is currently no “gold
standard” for interpreting diffusion data (Moseley, Bammer, & Illes, 2002) as the
implementation of DTI is still in its infancy, especially in cognitive neuroscience
research. Thus, responsibility falls on the researchers to carefully consider their data in

70
light of a few notable limitations. For instance, measures of white matter integrity are
considerably less accurate in certain brain regions, including areas with crossing fibers,
fiber degeneration, gray matter, and voxels with more than one tissue type (i.e., those
affected by partial volume)(Catani, 2006; Moseley et al., 2002). These inaccuracies
become especially important when making inferences about neural substrates
underlying differences in white matter integrity, as is commonly done with
examinations of axial and radial diffusivities (Wheeler-Kingshott & Cercignani, 2009).
In Chapter II, the location and organization of underlying fiber structure were taken
into account when patterns of age group differences were characterized based on these
diffusivity measures, which should have minimized the impact of this limitation to the
present results. Nonetheless, it will be necessary to confirm the proposed
interpretations with alternative techniques that are capable of identifying, accounting
for, or avoiding these error-prone regions (e.g., high-resolution DTI, multiple-fiber
DTI tractography).
Similarly, tractography analyses are subject to inaccuracies. For instance,
although highly reproducible seed and target masks were used here, the size and
placement of these masks may have resulted in erroneous tract reproductions, possibly
by over- or under-estimating a given tract of interest (Catani, 2006). There is also
uncertainty about the nature (anterograde versus retrograde) and functionality of
reconstructed tracts (Johansen-Berg & Behrens, 2006; Mori & Zhang, 2006; Powell et
al., 2004). These issues are especially relevant when the tracts are not major, well-

71
defined fiber pathways, as was the case for the tracts reconstructed in Chapter III.
Significant correlations between tract integrity and ASRT learning measures suggest
that the tracts examined here were at least partially accurate, and that they are
functionally relevant to implicit sequence learning. However, future research
replicating these results is necessary.
A broader limitation that should be addressed in future DTI aging research is
the impact of white matter hyperintensities (WMH) in relationships between white
matter integrity and cognition in healthy aging. As mentioned in the Discussion section
of Chapter II, WMH were not taken into account in this dataset because there were no
clinically significant structural abnormalities identified on the high resolution structural
scans. Nonetheless, WHM are present in healthy older adults, especially in frontal and
periventricular white matter (Nebes et al., 2006; Raz & Rodrigue, 2006), and their
prevalence increases with cerebrovascular risk factors commonly seen in healthy aging
(e.g., hypertension, silent strokes, and high blood pressure)(Gunning-Dixon & Raz,
2000; Longstrength et al., 1996). These regions, which are characterized by significant
white matter damage (e.g., myelin rarefaction, vessel endothelium breakdown,
microvascular disease)(Gunning-Dixon et al., 2009), may have affected the present
results, and should be acknowledged in future studies.

72
APPENDIX
Supplemental material from Chapter III
Following from the Chapter II findings regarding diffusivity measures of white
matter integrity, additional analyses assessed age-related differences in axial (AD) and
radial (RD) diffusivity in the tracts of interest from Chapter III, and related these
diffusivity measures to ASRT sequence learning scores. As in the earlier chapter, AD
and RD were used to make inferences about neural substrates underlying significant
differences in white matter integrity.
Age group differences in diffusivity measures. Separate between-group t-tests
compared AD and RD in younger and older adults for each tract of interest. For the
caudate-DLPFC tracts, significant age-related decreases in FA were accompanied by
age-related decreases in AD that were marginally significant for the left hemisphere, t
(23) = -2.01, p < .06, and significant for the right hemisphere, t (24) = -3.62, p < .01,
but no age group differences in RD. Based on earlier research (Budde et al., 2007;
Song et al., 2003), one explanation may be that age-related axonal differences (e.g.,
axonal damage or loss) contribute to this decline in white matter integrity. However,
given that the caudate-DLPFC tracts traverse long range anterior-posterior fibers that
pass through the internal and external capsules (e.g., anterior thalamic radiations,
superior longitudinal fasciculus), age group differences in macrostructural organization
may also be a contributing factor.

73
For the putamen-SMA tracts, younger and older adults did not differ on FA.
However, there were significant age-related increase in RD in the left hemisphere tract,
t (26) = 2.44, p < .03, and increases in both AD, t (23) = 2.28, p < .04, and RD, t (23) =
2.45, p < .03, in the right hemisphere tract. As proposed in Chapter II, this pattern
likely reflects age-related decreases in axonal packing density from demyelination,
axonal loss or damage, or inflammation in older versus younger adults, that are mild
enough to only increase RD in the left hemisphere, but more severe in the right
hemisphere, increasing both RD and AD (Bennett, Madden, Vaidya, Howard, &
Howard, 2009). Previous research has found evidence both for (Ardekani et al., 2007)
and against (Takao et al., 2009) asymmetry of age-related differences in white matter
integrity, though the neural underpinnings of these asymmetric effects warrants further
investigation.
Like analyses with FA, there were no significant age group differences in
diffusivity measures for the hippocampus-DLPFC tracts, p’s > .18. The lack of an age
group difference in the hippocampus-DLPFC tract suggests that not all tracts are
compromised in healthy aging.
Diffusivity correlates of sequence learning. Correlations were performed
between AD and RD from the tracts of interest and triplet type learning scores for
reaction time and accuracy across all epochs (overall scores) and for each of the three
stages of learning. The overall learning score for accuracy was significantly related to
AD in the left putamen-SMA tract, r = -0.38, p < .05; with the late learning score for

74
accuracy being related to AD in the left caudate-DLPFC tract, r = +0.40, p < .05, RD in
the right hippocampus-DLPFC, r = -0.44, p < .03, and RD in left putamen-SMA, r = -
0.52, p < .01.
The relationship between the late learning for accuracy and left caudate-
DLPFC tract AD was also seen with FA, suggesting that the correlation for FA may
have reflected individual differences in axonal integrity (e.g., homogeneous
orientation, axonal number or density). The remaining relationships did not overlap
with significant effects for FA, though axonal or myelin integrity may also underlie
these effects. It is worth noting that the pattern of findings seen here is not consistent
with earlier research. That is, half of the diffusivity-sequence learning relationships
were significant for RD, which does not fully support the notion that integrity-
cognition relationships are primarily due to individual differences in the degree of
myelination (Davis et al., 2009; Madden et al., 2009).
Taken together, these analyses revealed various patterns of age-related
differences in diffusivity measures across the three tracts of interest, suggesting that
different underlying neural substrates are responsible to for age-related declines in
white matter integrity. Furthermore, individual differences in a variety of neural
changes (e.g., both axonal and myelin-related) likely underlie individual differences in
cognitive functioning.

75
REFERENCES
Abe, O., Aoki, S., Hayashi, N., Yamada, H., Kunimatsu, A., Mori, H., Yoshikawa, T.,
Okubo, T., & Ohtomo, K. (2002). Normal aging in the central nervous system:
Quantitative MR diffusion-tensor analysis. Neurobiology of Aging, 23, 433-
441.
Aboitiz, F., Scheibel, A. B., Fisher, R. S., & Zaidel, E. (1992). Fiber composition of
the human corpus callosum. Brain Research, 598, 143-153.
Aizenstein, H. J., Butters, M. A., Clark, K. A., Figurski, J. L., Andrew Stenger, V.,
Nebes, R. D., Reynolds, C. F., 3rd, & Carter, C. S. (2006). Prefrontal and
striatal activation in elderly subjects during concurrent implicit and explicit
sequence learning. Neurobiology of Aging, 27, 741-751.
Aizenstein, H. J., Nebes, R. D., Meltzer, C. C., Fukui, M. B., Williams, R. L., Saxton,
J., Houck, P. R., Carter, C. S., Reynolds, C. F., 3rd, & DeKosky, S. T. (2002).
The relation of white matter hyperintensities to implicit learning in healthy
older adults. International Journal of Geriatric Psychiatry, 17, 664-669.
Andrews-Hanna, J. R., Snyder, A. Z., Vincent, J. L., Lustig, C., Head, D., Raichle, M.
E., & Buckner, R. L. (2007). Disruption of large-scale brain systems in
advanced aging. Neuron, 56, 924-935.
Ardekani, S., Kumar, A., Bartzokis, G., & Sinha, U. (2007). Exploratory voxel-based
analysis of diffusion indices and hemispheric asymmetry in normal aging.
Magnetic Resonance Imaging, 25, 154-167.

76
Baron, R. M., & Kenny, D. A. (1986). The moderator-mediator variable distinction in
social psychological research: Conceptual, strategic, and statistical
considerations. Journal of Personality and Social Psychology, 51, 1173-1182.
Basser, P. J., Mattiello, J., & LeBihan, D. (1994). MR diffusion tensor spectroscopy
and imaging. Biophysical Journal, 66, 259-267.
Basser, P. J., & Pierpaoli, C. (1996). Microstructural and physiological features of
tissues elucidated by quantitative-diffusion-tensor MRI. Journal of Magnetic
Resonance, B 111, 209-219.
Beaulieu, C. (2002). The basis of anisotropic water diffusion in the nervous system - a
technical review. NMR in Biomedicine, 15, 435-455.
Behrens, T. E. J., Woolrich, M. W., Jenkinson, M., Johansen-Berg, H., Hunes, S.,
Clare, S., Matthews, P. M., Brady, J. M., & Smith, S. M. (2003).
Characterization and propagation of uncertainty in diffusion-weighted MR
imaging. Magnetic Resonance in Medicine, 50, 1077-1088.
Bennett, I. J., Madden, D. J., Vaidya, C. J., Howard, D. V., & Howard, J. H. Jr. (2009).
Age-related differences in multiple measures of white matter integrity: A
diffusion tensor imaging study of healthy aging. Human Brain Mapping.
Bennett, I. L., Howard, J. H. Jr., & Howard, D. V. (2007). Older adults reveal implicit
learning of subtle third-order sequential structure. The Journals of Gerontology.
Series B, Psychological Sciences and Social Sciences, 62, 98-103.

77
Berman, J. I., Mukherjee, P., Partridge, S. C., Miller, S. P., Ferriero, D. M., Barkovich,
A. J., Vigneron, D. B., & Henry, R. G. (2005). Quantitative diffusion tensor
MRI fiber tractography of sensorimotor white matter development in premature
infants. Neuroimage, 27, 862-871.
Bhagat, Y. A., & Beaulieu, C. (2004). Diffusion anisotropy in subcortical white matter
and cortical gray matter: Changes with aging and the role of CSF-suppression.
Journal of Magnetic Resonance Imaging, 20, 216-227.
Bialystok, E., & Craik, F. I. M. (Eds.). (2006). Lifespan Cognition: Mechanisms of
Change. New York, NY: Oxford University Press.
Bucur, B., Madden, D. J., Spaniol, J., Provenzale, J. M., Cabeza, R., White, L. E., &
Huettel, S. A. (2008). Age-related slowing of memory retrieval: Contributions
of perceptual speed and cerebral white matter integrity. Neurobiology of Aging,
29, 1070-1079.
Budde, M. D., Kim, J. H., Liang, H. F., Schmidt, R. E., Russell, J. H., Cross, A. H., &
Song, S. K. (2007). Toward accurate diagnosis of white matter pathology using
diffusion tensor imaging. Magnetic Resonance in Medicine, 57, 688-695.
Burzynska, A. Z., Preuschhof, C., Backman, L., Nyberg, L., Li, S. C., Lindenberger,
U., & Heekeren, H. R. (2009). Age-related differences in white-matter
microstructure: Region-specific patterns of diffusivity. Manuscript submitted
for publication.

78
Cabeza, R. (2001). Cognitive neuroscience of aging: Contributions of functional
neuroimaging. Scandinavian Journal of Psychology, 42, 277-286.
Cabeza, R. (2002). Hemispheric asymmetry reduction in older adults: The HAROLD
model. Psychology and Aging, 17, 85-100.
Cabeza, R., Nyberg, L., & Park, D. (Eds.). (2005). Cognitive neuroscience of aging:
Linking cognitive and cerebral aging. New York, NY: Oxford University Press.
Camp, C. J., Bird, M. J., & Cherry, K. E. (2000). Retrieval strategies as a rehabilitation
aid for cognitive loss in pathological aging. In R. D. Hill, L. Backman & A. S.
Neely (Eds.), Cognitive Rehabilitation in Old Age (pp. 224-248). New York:
Oxford University Press.
Catani, M. (2006). Diffusion tensor magnetic resonance imaging tractography in
cognitive disorders. Current Opinion in Neurology, 19, 599-606.
Charlton, R. A., Landau, S., Schiavone, F., Barrick, T. R., Clark, C. A., Markus, H. S.,
& Morris, R. G. (2008). A structural equation modeling investigation of age-
related variance in executive function and DTI measured white matter damage.
Neurobiology of Aging, 29, 1547-1555.
Craik, F. I. (2006). Brain-behavior relations across the lifespan: A commentary.
Neuroscience and Biobehavioral Reviews, 30, 885-892.
Craik, F. I., & Jennings, J. M. (1992). Human memory. In F. I. M. Craik & T. A.
Salthouse (Eds.), The Handbook of Aging and Cognition (1st ed., pp. 51-110).
Hillsdale, NJ: Lawrence Erlbaum Assoc.

79
Crone, E., & Huizinga, M. (2006). Preface special issue 'Advances in Developmental
Cognitive Neuroscience'. Neuropsychologia, 44, 2015-2016.
Curran, T. (1995). On the neural mechanisms of sequence learning. PSYCHE, 2.
Curran, T. (1997). Effects of aging on implicit sequence learning: Accounting for
sequence structure and explicit knowledge. Psychological Research, 60, 24-41.
Daselaar, S. M., Rombouts, S. A., Veltman, D. J., Raaijmakers, J. G., & Jonker, C.
(2003). Similar network activated by young and old adults during the
acquisition of a motor sequence. Neurobiology of Aging, 24, 1013-1019.
Davis, S. W., Dennis, N. A., Buchler, N. G., White, L. E., Madden, D. J., & Cabeza, R.
(2009). Assessing the effects of aging on long white matter tracts using
diffusion tensor tractography. Neuroimage, 46, 530-541.
Doyon, J., Bellec, P., Amsel, R., Penhune, V., Monchi, O., Carrier, J., Lehericy, S., &
Benali, H. (2009). Contributions of the basal ganglia and functionally related
brain structures to motor learning. Behavioral Brain Research, 199, 61-75.
Fjell, A. M., Westlye, L. T., Greve, D. N., Fischl, B., Benner, T., van der Kouwe, A. J.,
Salat, D., Bjornerud, A., Due-Tonnessen, P., & Walhovd, K. B. (2008). The
relationship between diffusion tensor imaging and volumetry as measures of
white matter properties. Neuroimage, 42, 1654-1668.
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). "Mini-mental state". A
practical method for grading the cognitive state of patients for the clinician.
Journal of Psychiatric Research, 12, 189-198.

80
Frensch, P. A., & Miner, C. S. (1994). Effects of presentation rate and individual
differences in short-term memory capacity on an indirect measure of serial
learning. Memory and Cognition, 22, 95-110.
Giorgio, A., Watkins, K. E., Douaud, G., James, A. C., James, S., De Stefano, N.,
Matthews, P. M., Smith, S. M., & Johansen-Berg, H. (2008). Changes in white
matter microstructure during adolescence. Neuroimage, 39, 52-61.
Goulding, M. R., Rogers, M. E., & Smith, S. M. (2003). Trends in aging--United States
and worldwide. Morbidity and Mortality Weekly Report, 52, 101-106.
Grady, C. L. (2008). Cognitive neuroscience of aging. Annals of the New York
Academy of Sciences, 1124, 127-144.
Grieve, S. M., Williams, L. M., Paul, R. H., Clark, C. R., & Gordon, E. (2007).
Cognitive aging, executive function, and fractional anisotropy: A diffusion
tenor MR imaging study. American Journal of Neuroradiology, 28, 226-235.
Gunning-Dixon, F. M., Brickman, A. M., Cheng, J. C., & Alexopoulos, G. S. (2009).
Aging of cerebral white matter: A review of MRI findings. International
Journal of Geriatric Psychiatry, 24, 109-117.
Gunning-Dixon, F. M., & Raz, N. (2000). The cognitive correlates of white matter
abnormalities in normal aging: A quantitative review. Neuropsychology, 14,
224-232.
Head, D., Buckner, R. L., Shimony, J. S., Williams, L. E., Akbudak, E., Conturo, T. E.,
McAvoy, M., Morris, J. C., & Snyder, A. Z. (2004). Differential vulnerability

81
of anterior white matter in nondemented aging with minimal acceleration in
dementia of the Alzheimer type: Evidence from diffusion tensor imaging.
Cerebral Cortex, 14, 410-423.
Head, D., Snyder, A. Z., Girton, L. E., Morris, J. C., & Buckner, R. L. (2005). Frontal-
hippocampal double dissociation between normal aging and Alzheimer's
disease. Cerebral Cortex, 15, 732-739.
Hedden, T., & Gabrieli, J. D. (2005). Healthy and pathological processes in adult
development: New evidence from neuroimaging of the aging brain. Current
Opinion in Neurology, 18, 740-747.
Hedden, T., & Gabrieli, J. D. E. (2004). Insights into the ageing mind: A view from
cognitive neuroscience. Nature Reviews Neuroscience, 5, 87-96.
Heiervang, E., Behrens, T. E., Mackay, C. E., Robson, M. D., & Johansen-Berg, H.
(2006). Between session reproducibility and between subject variability of
diffusion MR and tractography measures. Neuroimage, 33, 867-877.
Hirsch, J. G., Bock, M., Essig, M., & Schad, L. R. (1999). Comparison of diffusion
anisotropy measurements in combination with the flair-technique. Magnetic
Resonance in Medicine, 17, 705-716.
Howard, D. V., & Howard, J. H., Jr. (1989). Age differences in learning serial patterns:
Direct versus indirect measures. Psychology and Aging, 4, 357-364.

82
Howard, D. V., Howard, J. H., Jr., Japikse, K., DiYanni, C., Thompson, A., &
Somberg, R. (2004). Implicit sequence learning: Effects of level of structure,
adult age, and extended practice. Psychology and Aging, 19, 79-92.
Howard, J. H., Jr., & Howard, D. V. (1997). Age differences in implicit learning of
higher order dependencies in serial patterns. Psychology and Aging, 12, 634-
656.
Hsu, J. L., Leemans, A., Bai, C. H., Lee, C. H., Tsai, Y. F., Chiu, H. C., & Chen, W. H.
(2008). Gender differences and age-related white matter changes of the human
brain: a diffusion tensor imaging study. Neuroimage, 39, 566-577.
Hugenschmidt, C. E., Peiffer, A. M., Kraft, R. A., Casanova, R., Deibler, A. R.,
Burdette, J. H., Maldjian, J. A., & Laurienti, P. J. (2008). Relating imaging
indices of white matter integrity and volume in healthy older adults. Cerebral
Cortex, 18, 433-442.
Johansen-Berg, H., & Behrens, T. E. (2006). Just pretty pictures? What diffusion
tractography can add in clinical neuroscience. Current Opinion in Neurology,
19, 379-385.
Jones, D. K. (2008). Studying connections in the living human brain with diffusion
MRI. Cortex, 44, 936-952.
Kuhl, P. K. (2004). Early language acquisition: Cracking the speech code. Nature
Reviews Neuroscience, 5, 831-843.

83
Langley, L. K., & Madden, D. J. (2000). Functional neuroimaging of memory:
implications for cognitive aging. Microscopy Research and Technique, 51, 75-
84.
Le Bihan, D. (2003). Looking in the functional architecture of the brain with diffusion
MRI. Nature Reviews Neuroscience, 4, 469-480.
Lieberman, M. D. (2000). Intuition: a social cognitive neuroscience approach.
Psychological Bulletin, 126, 109-137.
Logan, J. M., Sanders, A. L., Snyder, A. Z., Morris, J. C., & Buckner, R. L. (2002).
Under-recruitment and nonselective recruitment: Dissociable neural
mechanisms associated with aging. Neuron, 33, 827-840.
Longstrength, W. T., Jr., Manolio, T. A., Arnold, A., Burke, G. L., Bryan, N., Jungreis,
C. A., Enright, P. L., O'Leary, D., & Fried, L. (1996). Clinical correlates of
white matter findings on cranial magnetic resonance imaging of 3301 elderly
people. Stroke, 27, 1274-1282.
Madden, D. J., Spaniol, J., Costello, M. C., Bucur, B., White, L. E., Cabeza, R., Davis,
S. W., Dennis, N. A., Provenzale, J. M., & Huettel, S. A. (2009). Cerebral
white matter integrity mediates adult age differences in cognitive performance.
Journal of Cognitive Neuroscience, 21, 289-302.
Madden, D. J., Whiting, W. L., Huettel, S. A., White, L. E., MacFall, J. R., &
Provenzale, J. M. (2004). Diffusion tensor imaging of adult age differences in
cerebral white matter: Relation to response time. Neuroimage, 21, 1174-1181.

84
Marner, L., Nyengaard, J. R., Tang, Y., & Pakkenberg, B. (2003). Marked loss of
myelinated nerve fibers in the human brain with age. Journal of Comparative
Neurology, 462, 144-152.
Mori, S., & Zhang, J. (2006). Principles of diffusion tensor imaging and its
applications to basic neuroscience research. Neuron, 51, 527-539.
Moseley, M., Bammer, R., & Illes, J. (2002). Diffusion-tensor imaging of cognitive
performance. Brain and Cognition, 50, 396-413.
Muller, M. M., & Mayes, A. R. (2001). Cognitive neuroscience: Perception, attention,
and memory. Neuroscience and Biobehavioral Reviews, 25, 463-464.
Nair, G., Tanahashi, Y., Low, H. P., Billings-Gagliardi, S., Schwartz, W. J., & Duong,
T. Q. (2005). Myelination and long diffusion times alter diffusion-tensor-
imaging contrast in myelin-deficient shiverer mice. Neuroimage, 28, 165-174.
Nebes, R. D., Meltzer, C. C., Whyte, E. M., Scanlon, J. M., Halligan, E. M., Saxon, J.
A., Houck, P. R., Boada, F. E., & DeKosky, S. T. (2006). The relation of white
matter hyperintensities to cognitive performance in the normal old: Education
matters. Aging, Neuropsychology, and Cognition, 13, 326-340.
Negash, S. (2003). Adult age differences in implicit learning of short and higher-order
sequential patterns. Washington, DC: Georgetown University.
Nissen, M. J., & Bullemer, P. (1987). Attentional requirements of learning: Evidence
from performance measures. Cognitive Psychology, 19, 1-32.

85
Nordahl, C. W., Ranganath, C., Yonelinas, A. P., Decarli, C., Fletcher, E., & Jagust,
W. J. (2006). White matter changes compromise prefrontal cortex function in
healthy elderly individuals. Journal of Cognitive Neuroscience, 18, 418-429.
O'Sullivan, M., Jones, D. K., Summers, P. E., Morris, R. G., Williams, S. C., &
Markus, H. S. (2001). Evidence for cortical ''disconnection'' as a mechanism of
age-related cognitive decline. Neurology, 57, 632-638.
Papadakis, N. G., Martin, K. M., Mustafa, M. H., Wilkinson, I. D., Griffiths, P. D.,
Huang, C. L., & Woodruff, P. W. (2002). Study of the effect of CSF
suppression on white matter diffusion anisotropy mapping of healthy human
brain. Magnetic Resonance in Medicine, 48, 394-398.
Perruchet, P., Gallego, J., & Savy, I. (1990). A critical reappraisal of the evidence for
unconscious abstraction of deterministic rules in complex experiment
situations. Cognitive Psychology, 22, 493-516.
Perry, M. E., McDonald, C. R., Hagler, D. J., Jr., Gharapetian, L., Kuperman, J. M.,
Koyama, A. K., Dale, A. M., & McEvoy, L. K. (2009). White matter tracts
associated with set-shifting in healthy aging. Neuropsychologia.
Peters, A. (2002). The effects of normal aging on myelin and nerve fibers: A review.
Journal of Neurocytology, 31, 581-593.
Pfefferbaum, A., Adalsteinsson, E., & Sullivan, E. V. (2005). Frontal circuitry
degradation marks healthy adult aging: Evidence from diffusion tensor
imaging. Neuroimage, 16, 891-899.

86
Pfefferbaum, A., & Sullivan, E. V. (2003). Increased brain white matter diffusivity in
normal adult aging: Relationship to anisotropy and partial voluming. Magnetic
Resonance in Medicine, 49, 953-961.
Pfefferbaum, A., Sullivan, E. V., Hedehus, M., Lim, K. O., Adalsteinsson, E., &
Moseley, M. (2000). Age-related decline in brain white matter anisotropy
measured with spatially corrected echo-planar diffusion tensor imaging.
Magnetic Resonance in Medicine, 44, 259-268.
Pierpaoli, C., Barnett, A., Pajevic, S., Chen, R., Penix, L. R., Virta, A., & Basser, P.
(2001). Water diffusion changes in Wallerian degeneration and their
dependence on white matter architecture. Neuroimage, 13, 1174-1185.
Pierpaoli, C., & Basser, P. J. (1996). Toward a quantitative assessment of diffusion
anisotropy. Magnetic Resonance in Medicine, 36, 893-906.
Pierpaoli, C., Jezzard, P., Basser, P. J., Barnett, A., & Di Chiro, G. (1996). Diffusion
tensor MR imaging of the human brain. Radiology, 201, 637-648.
Poldrack, R. A., & Packard, M. G. (2003). Competition among multiple memory
systems: Converging evidence from animal and human brain studies.
Neuropsychologia, 41, 245-251.
Powell, H. W., Guye, M., Parker, G. J., Symms, M. R., Boulby, P., Koepp, M. J.,
Barker, G. J., & Duncan, J. S. (2004). Noninvasive in vivo demonstration of the
connections of the human parahippocampal gyrus. Neuroimage, 22, 740-747.

87
Raz, N. (2000). Aging of the brain and its impact on cognitive performance:
Integration of structural and functional findings. In F. I. M. Craik & T. A.
Salthouse (Eds.), The Handbook of Aging and Cognition (2nd ed., pp. 1-90).
Mahwah, NJ: Lawrence Erlbaum Assoc.
Raz, N. (2005). The aging brain observed in vivo: Differential changes and their
modifiers. In R. Cabeza, L. Nyberg & D. Park (Eds.), Cognitive Neuroscience
of Aging: Linking Cognitive and Cerebral Aging (pp. 19-57). New York, NY:
Oxford University Press.
Raz, N., Gunning-Dixon, F. M., Head, D., Dupuis, J. H., & Acker, J. D. (1998).
Neuroanatomical correlates of cognitive aging: Evidence from structural
magnetic resonance imaging. Neuropsychology, 12, 95-114.
Raz, N., Lindenberger, U., Rodrigue, K. M., Kennedy, K. M., Head, D., Williamson,
A., Dahle, C., Gerstorf, D., & Sacker, J. D. (2005). Regional brain changes in
aging healthy adults: General trends, individual differences, and modifiers.
Cerebral Cortex, 15, 1676-1689.
Raz, N., & Rodrigue, K. M. (2006). Differential aging of the brain: Patterns, cognitive
correlates and modifiers. Neuroscience and Behavioral Reviews, 30, 730-748.
Reber, A. S. (1993). Implicit learning and tacit knowledge: An essay on the cognitive
unconscious. New York, NY: Oxford University Press.
Reuter-Lorenz, P. A. (2002). New visions of the aging mind and brain. TRENDS in
Cognitive Sciences, 6, 394-400.

88
Reuter-Lorenz, P. A., & Cappell, K. A. (2008). Neurocognitive aging and the
compensation hypothesis. Current Directions in Psychological Science, 17,
177-182.
Rose, M., Haider, H., Weiller, C., & Buchel, C. (2002). The role of medial temporal
lobe structures in implicit learning: An event-related FMRI study. Neuron, 36,
1221-1231.
Rypma, B., Berger, J. S., Prabhakaran, V., Bly, B. M., Kimberg, D. Y., Biswal, B. B.,
& D'Esposito, M. (2006). Neural correlates of cognitive efficiency.
Neuroimage, 33, 969-979.
Salat, D. H., Buckner, R. L., Snyder, A. Z., Greve, D. N., Desikan, R. S., Busa, E.,
Morris, J. C., Dale, A. M., & Fischl, B. (2004). Thinning of the cerebral cortex
in aging. Cerebral Cortex, 14, 721-730.
Salat, D. H., Tuch, D. S., Greve, D. N., van der Kouwe, A. J., Hevelone, N. D., Zaleta,
A. K., Rosen, B. R., Fischl, B., Corkin, S., Rosas, H. D., & Dale, A. M. (2005).
Age-related alterations in white matter microstructure measured by diffusion
tensor imaging. Neurobiology of Aging, 26, 1215-1227.
Salthouse, T. A. (1991). Mediation of adult age differences in cognition by reductions
in working memory and speed of processing. Psychological Science, 2, 179-
183.

89
Schendan, H. E., Searl, M. M., Melrose, R. J., & Stern, C. E. (2003). An fMRI study of
the role of the medial temporal lobe in implicit and explicit sequence learning.
Neuron, 37, 1013-1025.
Seger, C. A. (1994). Implicit learning. Psychological Bulletin, 115, 163-196.
Seger, C. A. (2006). The basal ganglia in human learning. Neuroscientist, 12, 285-290.
Shenkin, S. D., Bastin, M. E., MacGillivray, T. J., Deary, I. J., Starr, J. M., &
Wardlaw, J. M. (2003). Childhood and current cognitive function in healthy 80-
year-olds: A DT-MRI study. Neuroreport, 14, 345-349.
Smith, S. M., Jenkinson, M., Johansen-Berg, H., Rueckert, D., Nichols, T. E., Mackay,
C. E., Watkins, K. E., Ciccarelli, O., Cader, M. Z., Matthews, P. M., &
Behrens, T. E. J. (2006). Tract-based spatial statistics: Voxelwise analysis of
multi-subject diffusion data. Neuroimage, 31, 1487-1505.
Smith, S. M., Johansen-Berg, H., Jenkinson, M., Rueckert, D., Nichols, T. E., Miller,
K. L., Robson, M. D., Jones, D. K., Klein, J. C., Bartsch, A. J., & Behrens, T.
E. (2007). Acquisition and voxelwise analysis of multi-subject diffusion data
with tract-based spatial statistics. Nature Protocols, 2, 499-503.
Song, S. K., Sun, S. W., Ju, W. K., Lin, S. J., Cross, A. H., & Neufeld, A. H. (2003).
Diffusion tensor imaging detects and differentiates axon and myelin
degeneration in mouse optic nerve after retinal ischemia. Neuroimage, 20,
1714-1722.

90
Song, S. K., Sun, S. W., Ramsbottom, M. J., Chang, C., Russell, J., & Cross, A. H.
(2002). Dysmyelination revealed through MRI as increased radial (but
unchanged axial) diffusion of water. Neuroimage, 17, 1429-1436.
Song, S. K., Yoshino, J., Le, T. Q., Lin, S. J., Sun, S. W., Cross, A. H., & Armstrong,
R. C. (2005). Demyelination increases radial diffusivity in corpus callosum of
mouse brain. Neuroimage, 26, 132-140.
Stadlbauer, A., Salomonowitz, E., Strunk, G., Hammen, T., & Ganslandt, O. (2008).
Age-related degradation in the central nervous system: Assessment with
diffusion-tensor imaging and quantitative fiber tracking. Radiology, 247, 179-
188.
Sullivan, E. V., Adalsteinsson, E., & Pfefferbaum, A. (2006). Selective age-related
degradation of anterior callosal fiber bundles quantified in vivo with fiber
tracking. Cerebral Cortex, 16, 1030-1039.
Sullivan, E. V., & Pfefferbaum, A. (2003). Diffusion tensor imaging in normal aging
and neuropsychiatric disorders. European Journal of Radiology, 45, 244-255.
Sullivan, E. V., & Pfefferbaum, A. (2006). Diffusion tensor imaging and aging.
Neuroscience and Biobehavioral Reviews, 30, 749-761.
Sullivan, E. V., Rohlfing, T., & Pfefferbaum, A. (2008). Quantitative fiber tracking of
lateral and interhemispheric white matter systems in normal aging: Relations to
timed performance. Neurobiology of Aging.

91
Sundgren, P. C., Dong, Q., Gomez-Hassan, D., Mukherji, S. K., Maly, P., & Welsh, R.
(2004). Diffusion tensor imaging of the brain: Review of clinical applications.
Neuroradiology, 46, 339-350.
Takao, H., Abe, O., Yamasue, H., Aoki, S., Kasai, K., Sasaki, H., & Ohtomo, K.
(2009). Aging effects on cerebral asymmetry: A vovel-based morphometry and
diffusion tensor imaging study. Magnetic Resonance Imaging.
Tang, Y., Nyengaard, J. R., Pakkenberg, B., & Gundersen, H. J. (1997). Age-induced
white matter changes in the human brain: A stereological investigation.
Neurobiology of Aging, 18, 609-615.
Thomalla, G., Glauche, V., Koch, M. A., Beaulieu, C., Weiller, C., & Rother, J.
(2004). Diffusion tensor imaging detects early Wallerian degeneration of the
pyramidal tract after ischemic stroke. Neuroimage, 22, 1767-1774.
Vernooij, M. W., de Groot, M., van der Lugt, A., Ikram, M. A., Krestin, G. P.,
Hofman, A., Niessen, W. J., & Breteler, M. M. (2008). White matter atrophy
and lesion formation explain the loss of structural integrity of white matter in
aging. Neuroimage, 43, 470-477.
West, R. L. (1996). An application of prefrontal cortex function theory to cognitive
aging. Psychological Bulletin, 120, 272-292.
Wheeler-Kingshott, C. A. M., & Cercignani, M. (2009). About "axial" and "radial"
diffusivities. Magnetic Resonance in Medicine, 61, 1255-1260.

92
Yoon, B., Shim, Y. S., Lee, K. S., Shon, Y. M., & Yang, D. W. (2008). Region-
specific changes of cerebral white matter during normal aging: A diffusion-
tensor analysis. Archives of Gerontology and Geriatrics, 47, 129-138.
Zahr, N. M., Rohlfing, T., Pfefferbaum, A., & Sullivan, E. V. (2009). Problem solving,
working memory, and motor correlates of association and commissural fiber
bundles in normal aging: A quantitative fiber tracking study. Neuroimage, 44,
1050-1062.
Zhang, Y., Du, A. T., Hayasaka, S., Jahng, G. H., Hlavin, J., Zhan, W., Weiner, M. W.,
& Schuff, N. (2008). Patterns of age-related water diffusion changes in human
brain by concordance and discordance analysis. Neurobiology of Aging.
Zhou, M., Goto, N., Otsuka, N., Moriyama, H., & Nakamura, Y. (1997). Morphometric
analyses of axons in the lateral corticospinal tract with ageing process.
Okajimas Folia Anatomica Japonica, 74, 133-138.