aggregation of sponge cells - the journal of biological chemistry
TRANSCRIPT

Aggregation of Sponge Cells A NOVEL MECHANISM OF CONTROLLED INTERCELLULAR ADHESION, BASING ON THE INTERRELATION BETWEEN GLYCOSYLTRANSFERASES AND GLYCOSIDASES*
(Received for publication, May 30, 1978)
Werner E. G. Miiller,$ 8 Rudolf K. Zahn,$ 0 B ranko Kurelec,$ Isabel Miiller,$ 9 Gerd Uhlenbruck,l and Peter Vaithl
From the $Physiologisch-Chemisches Institut, Universitat, Duesbergweg, 6500 Mainz, West Germany, the aCenter for Marine Research, Institute Ruder BoskoviC, 52210 Rovinj, Yugoslavia, and the BMedizinische Universittitsklinik, Abteilung Experimentelle Innere Medizin, Kerpener Str. 15, 5000 Koln, West Germany
The biochemical and functional properties of p-glu- curonidase and j?-galactosidase, isolated and partially purified from the sponge Geodia cydonium, were stud- ied. The two glycosidases are not only localized in the cytoplasm but are also associated at a high activity with the cell membrane.
The aggregation receptor, a low molecular weight cell surface-bound glycoprotein, is deglucuronylated by the action of the Geodia /3-glucuronidase both in the state at which the receptor is bound to the cell surface and in its isolated form. The deglucuronylated aggre- gation receptor can be reglucuronylated enzymatically by the extracellularly occurring homologous glucuron- osyltransferase.
Untreated cells lose their aggregation potency if they are incubated under conditions optimal for P-glucuron- idase activity. Cells, depleted of membrane-bound p- glucuronidase, do not show any reduction of their ag- gregation potency under identical conditions. Cells that carry on their cell surface deglucuronylated molecules and are biologically characterized by only a low aggre- gation potency can be reglucuronylated in the presence of glucuronosyltransferase and UDP-glucuronic acid. These restored cells show again the original high ag- gregation potency.
From the results presented it is assumed that cell aggregation can occur after glucuronylation of the ag- gregation receptor by the glucuronosyltransferase via a linkage of the aggregation factor with the aggrega- tion receptor. Some evidence is presented, indicating that cell separation is the consequence of an activation of the cell membrane bound P-glucuronidase, which results in a deglucuronylation of the aggregation recep- tor.
Among the different biological models studied to under- stand cell-cell aggregation processes both on the cellular and biochemical level, the retina system (for a survey, see Refs. 1
and 2) and the sponge cell system (for a survey, see Ref. 3) are the most widely used. It is owing to Moscona (4), who suc- ceeded to isolate an “extracellularly functioning material” from the sponge Microciona prolifera which promotes reag-
* This work has partially been supported by the Academy of Science and Letters, Maim, Germany and by the Bundesministerium fiir Forschung und Technologie. This manuscript is No. XIX in a series of publications emanating from our laboratory and dealing with aggregation of sponge cells. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
gregation of single cells. This “material” was later termed aggregation factor; in 1973 the factor was purified and char- acterized from the sponge Geodia cydonium (5, 6).
The aggregation factor of Geodia is associated with high molecular weight particles with a sedimentation coefficient (s&J of 90 (6a); in the presence of minute amounts of Ca”, the particles form complexes which have a sedimentation coefficient of 3,000 (6). The Geodia particles occur in the intercellular space and are found to act multifunctionally. The first subunit identified was the aggregation factor (5, 7); later a series of glycosyltransferases was identified as subunits of the Geodia particles: sialyltransferase (8), glucuronosyltrans- ferase (9), and galactosyltransferase (9). Hence, the Geodia particles represent a multiglycosyltransferase system associ- ated with an aggregation promoting activity.
The Geodia particles, in particular the aggregation factor associated with them, bind to specific cell surface molecules, the so-called aggregation receptors, which are of low molecular weight (Mr = approximately 20,000; s$J,~ = 2.6) and consist chemically of glycoproteins (10). The termini of the aggrega- tion receptor which are involved in the binding with the
aggregation factor were determined to consist of glucuronic acid (11). A further cell membrane component, the antiaggre- gation receptor (Mr = 180,000), a glycoprotein with D-galactose termini (12), modulates the activity of the aggregation factor; the antiaggregation receptor is most likely involved in a con- trolled segregation and subsequent reaggregation of the cells in randomly aggregated cell clumps.
Stimulated by the study of Aoyagi et al. (13), demonstrating the presence of a variety of glycosidases on the surface of mammalian cells, we searched for the cell surface enzymes neuraminidase, ,&glucuronidase, and ,&galactosidase. Only the latter two enzymes could be detected, partially purified, and characterized. In addition, attempts were made to deter- mine the role of both the ,&galactosidase and the P-glucuron- idase in the control of the reaggregation process, with the main emphasis on the latter enzyme. From our studies, we hypothesized that the ,&glucuronidase and the glucuronosyl- transferase might control the initial events of cell aggregation, mediated by the aggregation factor and the aggregation recep- tor, while the ,&galactosidase and the galactosyltransferase might play a role in the secondary event the “sorting out.”
MATERIALS AND METHODS
Chemicals
p-Nitrophenyl-ol-D-galactopyranoside, p-nitrophenyl-P-D-galacto- pyranoside, p-nitrophenyl-a-D-glucoside, p-nitrophenyl-P-r-gluco- side,p-nitrophenyl-P-D-glucuronide,p-nitrophenyl-a-D-mannoside,p- nitrophenyl, UDP-glucuronic acid, ethylene glycol bis(P-aminoethyl
1280
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

Controlled Intercellular Adhesion 1281
ether)N,N’-tetraacetic acid and 5,5’-dithiobis(2-nitrobenzoic acid) were from Sigma; Sepharose 4B, DEAE-Sephadex A-25, Sephadex G- 100, and dextran blue were from Deutsche Pharmacia, Freiburg (West Germany); Nonidet NP-40 was from Deutsche Shell, Hamburg (West Germany); [8-3H]GMP (specific activity, 4.6 Ci/mmol), UDP-D-[ U- i4C]galactose (specific activity, 337 mCi/mmol), UDP-D-[ U-‘4C]glu- curonic acid (specific activity, 321 mCi/mmol), and D-[ U-‘*C]glucu- ronic acid (specific activity, 76 mCi/mmol) were from the Radiochem- ical Centre, Amersham (England); and UDP-galactose was from Calbiochem.
Animals
The siliceous sponge Geodia cydonium Jam. (Tetractinellida) was collected in the vicinity of Rovinj (Yugoslavia).
Buffers and Solutions
Calcium- and magnesium-free artificial sea water and calcium- and magnesium-containing artificial sea water were composed as de- scribed (8).
Isolation of Cells
The dissociation of sponge tissue into separate cells was performed in CMF’ as described (5). The preparation of dissociated cells con- sisted of three main size classes (8). Counting and characterization of cells were performed as described (5, 14).
Cell Membranes
The cell membranes were isolated from cells, dissociated in CMF, according to the “Tris method” described by Warren et al. (15). The resulting membrane fraction was dialyzed (24 h, 2°C) against 3 ml of CMF medium containing 0.1% Nonidet NP-40. Chemical analysis of the membrane fraction revealed: 44.1% protein, 39.6% lipid, and 8.7% neutral carbohydrates. The membrane fraction contained a 5’.nucle- otidase activity of 3.6 nkat of [3H]GMP hydrolyzed/mg of protein. The membranes were dissolved in saline (0.9% NaCl) at a concentra- tion of 1.2 mg of protein/ml.
The procedures for isolation of the aggregation factor (Step V with a protein content of 1.5 mg/ml and a specific aggregation-promoting activity of 6.1 x 10” aggregation units/mg of protein has been used) and isolation of the aggregation receptor (Step IV has been used) have all been described in detail (5, 7, 8, 10). The aggregation activity is expressed in aggregation units (AU) as described (5).
Aggregation Assay
In the standard assay, a suspension with 25 + 5 x 10” cells/ml of ASW was placed in glass tubes and rollered at 35 rpm at 20°C (5). The assays contained the aggregation factor in a concentration of 1 x lo4 AU/ml. The diameter of the aggregates formed was determined microscopically according to the equation D = m in which D is the “diameter,” a is the longer axis, and b is the shorter axis of the aggregate.
Acceptor Preparation
In order to convert the aggregation receptor into a form lacking n-glucuronic acid, 0.5 ml of the highly purified aggregation receptor (= 2.3 mg of neutral carbohydrate), dissolved in saline, was added to 0.5 ml of a solution containing 20 mM sodium phosphate (pH 5.0), 400 mM NaCl, 5 IIIM 2-mercaptoethanol, and 10 nkat of Geodia p-glucu- ronidase (Step IV). The mixture was incubated for 2 h at 22°C and subsequently heated for 5 min at 100°C to inactivate the enzyme. After centrifugation (150,000 x g, 2O”C, 5 min), the supernatant was collected and served as acceptor for the experiments described under “Reglucuronylation of Aggregation Receptor.”
The acceptors (deglucuronylated aggregation receptor and dega- lactosylated aggregation receptor) used for the studies described under “Changes of /&Glucuronosyltransferase and ,L-Galactosyltrans- ferase Activity during Reaggregation” were prepared as previously described (9).
Glycosyltransferases
The high molecular weight particle-bound preparation (Step IV; Ref. 7), which carries, besides the aggregation factor (7), the three
’ The abbreviations used are: CMF, calcium- and magnesium-free artificial sea water; ASW, calcium- and magnesium-containing artifi- cial sea water; EGTA, ethylene glycol bis(,&aminoethyl ether)N,N’- tetraacetic acid; DTNB, 5,5’-dithiobis(2-nitrobenzoic acid).
glycosyltransferases (sialyltransferase, Ref. 8; glucuronosyltransfer- ase, Ref. 9; galactosyltransferase, Ref. 9) as subunits, was used for the glycosyltransferase reactions. The specific activities of the enzymes were as follows: sialyltransferase 9.1 pkat/mg of protein, glucurono- syltransferase 18.3 pkat/mg of protein, and galactosyltransferase 27.7 pkat/mg of protein,
Glycosyltransferase Assays
The ,L-glucuronosyltransferase was measured in a reaction mixture containing 10 mivf MgCL, 300 mM NaCl, 50 mM Tris-HCl (pH 8.2), 0.1 IIIM UDP-[ U-‘?]glucuronic acid (4 X 10’ dpm/pmol), 20 ~1 of deglu- curonylated aggregation receptor (2.8 pg of protein/assay), and 30 ~1 of the enzyme sample in a final volume of 80 pl. The incubation was carried out for 60 min at 22’C. The acid-insoluble radioactivity was determined as described (9).
The ,&galactosyltransferase assay was composed and performed as described for the glucuronyltransferase assay with the exception that 0.1 mM UDP-[U-‘4C]galactose (5 x 10’ dpm/pmol) instead of UDP- glucuronic acid was used and degalactosylated aggregation receptor served as the acceptor.
Isolation and Purification of ,l-Glucuronidase and p- Galactosidase
All procedures were carried out at 0-4°C. A typical preparation (starting with 50 g of material) is summarized in Table I.
Step I: Initial Extract-The sponge was cut into 2-mm cubes and suspended 1:3 (w/v) in 100 mM Tris-HCl buffer (pH 7.5) containing 5 mM 2-mercaptoethanol and 0.1% Nonidet NP-40 and homogenized with 10 strokes of a Dounce homogenizer. The homogenate was centrifuged at 20,000 x g for 30 min at 2°C. The supernatant (120 ml) was collected and contained 8.3 mg of protein/ml; it was concentrated in a dialysis tubing with polyethyleneglycol to 25 ml (Fraction 1).
Step 2: Sepharose 4B-Fraction 1 was passed through a Sepharose 4B column (4.5 x 20 cm) to remove the high molecular weight aggregation factor (5). The column was previously equilibrated with 100 mrvr Tris-HCl buffer (pH 7.5; 100 mM NaCl and 5 IIIM P-mercap- toethanol) and, after application of the sample, it was eluted with the same buffer. Three-milliliter fractions were collected. Both the p- glucuronidase- and the ,f3-galactosidase activity appeared within the Z’& (16) range 0.23 to 0.30 (Fraction 33-40). The pooled fractions (24 ml; Fraction 2) were directly subjected to a DEAE-Sephadex column.
Step 3: DEAE-Sephadex-Fraction 2 was applied to a DEAE- Sephadex column as described in Fig. 1. The two glycosidases (,f- glucuronidase and P-galactosidase) were obtained by stepwise gra- dient; they appeared after elution with 0.25 M NaCl. The enzyme- containing fractions (Fraction 20-24) were pooled; they contained 6.1 nkat of ,&glucuronidase and 2.0 nkat of P-galactosidase in a total volume of 25 ml (0.88 mg of protein/ml). The enzyme fractions were concentrated in a dialysis tubing with polyethylene glycol to 2 ml (Fraction 3).
Step 4: Sephadex G-100-Fraction 4 was passed over a column of Sephadex G-106 (Fig. 2). The ,&glucuronidase activity appeared as an asymmetric peak near the void volume ( V,/VO value: 1.3). The p- galactosidase activity was completely separated from the ,&glucuron- idase one; maximal activity was determined around a V,/ VO value of 1.9. The ,&glucuronidase Fractions 14 to 19 were pooled (Fraction 4); this preparation contained the enzyme with a specific activity of 0.857 nkat/mg of protein (0.53 mg of protein/ml). Fractions 22 to 26
TABLE I
Purification of P-glucuronidase and ,t-galactosidase
Step and fraction
1. Initial extract
2. Sepharose 4B
/&Glucuronidase P-Galactosidase
Specific Total en- zyme ac- Yield Specific Total en-
activity zyme ac- Yield tivity activity tivity
nkatlmg nkat % nkat/mg nkat 5% 0.012 11.9 100 0.003 3.0 100
0.059 9.4 79 0.002 2.5 84
3. DEAE-Sepha- 0.277 6.1 51 0.091 2.0 67 dex
4. Sephadex 0.857 5.4 45 0.652 1.5 49 G-100
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

Controlled Intercellular Adhesion
Fraction number
FIG. 1. DEAE-Sephadex chromatography of Fraction 2. Fraction 2, containing in a volume of 120 ml 550 mg of protein, was applied to a DEAE-Sephadex (A-25) column (3 x 20 cm), previously equilibrated with 100 mM Tris-HCl buffer (pH 7.5; 100 mM NaCl and 5 mM 2- mercaptoethanol). For elution, the Tris buffer was stepped up with 0.25 M and 0.5 M NaCl (----). Five-milliliter fractions were collected and assayed for enzyme activity. X. . . . . X, A~80 nm; O----O, p-glucu- ronidase activity; 0- - -0, P-galactosidase activity.
Fraction number
FIG. 2. Purification of Fraction 2 on Sephadex G-100. A column (2 x 20 cm) of Sephadex G-100, previously equilibrated with 10 mM Tris-HCl buffer (pH 7.5; 200 mM NaCl and 5 mM 2-mercaptoethanol), was loaded with 2 ml of Fraction 3 and eluted with the equilibration buffer; 2-n-J fractions were collected. The arrow marks the position of V0 (dextran blue marker). Abscissa (upper scale): V,/Vo values (16). x..... X, Azso ,,,,,; +---O, j?-glucuronidase activity; 0- - -0, p- galactosidase activity.
containing the ,&galactosidase were pooled (Fraction 4); the specific activity of the enzyme was determined to be 0.652 nkat/mg of protein (0.23 mg of protein/ml).
The purification scheme is summarized in Table I. A 71-fold increase in the specific activity of the P-n-glucuronidase was achieved, in the case of ,8-n-galactosidase, a 217-fold enrichment was obtained. Under the standard incubation conditions used, the two enzyme preparations were free of the following glycosidases: oc-n-galactosid- ase, a-n-glucosidase, P-n-glucosidase, and a-n-mannosidase.
Assay of Glycosidases
The amount of a given glycosidase activity was determined using the p-nitrophenyl derivatives (17). The standard assay (1 ml) con- tained the following constituents: 10 mM sodium phosphate (pH 5.0), 200 mM NaCl, 5 mM 2-mercaptoethanol, 1 mM concentration of the p- nitrophenyl derivative and routinely 50 ~1 of the enzyme preparation. The incubation was usually carried out for 60 min at 22°C. The reaction was terminated with 1 ml of 0.4 M glycine/NaOH buffer (pH 10.5) and the absorbance of the released p-nitrophenol measured at 420 nm. The substrates used were thep-nitrophenyl derivatives from a-n-galactoside, ,8-n-galactoside, a-n-glucoside, P-n-glucoside, p-D- glucuronide, and a-n-mannoside.p-Nitrophenol was used as standard. The specific activity is expressed as katals per mg of protein.
Labeling of Aggregation Receptor with Glucuronic Acid
Labeling of aggregation receptor was performed by addition of D-
[‘%]glucuronic acid (3 pCi/ml) to a 50-ml culture of 25 + 5 X lo6
cells/ml of ASW. After 24 h of cultivation (2O”C), the cells were harvested and washed twice with ASW by centrifugation. Subse- quently, the aggregation receptor was solubilized from the cell mem- brane and partially purified according to a procedure described earlier (10). Briefly, the cells were suspended in tricbloroacetic acid and homogenized. After centrifugation, the supernatant was dialyzed until neutral pH and subsequently poured into acetone. The precipitate formed was collected, dissolved, and then purified by DEAE-Sepha- dex ion exchange chromatography and finally by gel filtration through Sephadex G-50. The resulting partially purified aggregation receptor preparation had a specific radioactivity of 8.300 dpm/lO pg of neutral carbohydrate.
Analytical Methods
For protein determination the method of Lowry et al. (18) was used. Neutral carbohydrates were determined according to the method of Dubois et al. (19); lipids, according to Hinsberg et al. (20); and 5’-nucleotidase activity, according to Miiller et al. (21). The molecular weight, basing on the V,/VO value, was determined accord- ing to Determann (16).
Polyacrylamide gel electrophoresis in dodecyl sulfate was per- formed according to Laemmli (22) with an 8% polyacrylamide gel. The glycoprotein was located by the periodic acid-Schiff reaction (23) and densitometric scanning was performed using an ISCO model UA4 absorbance monitor.
RESULTS
Properties of ,8-D-Glucuronidase-The partially purified sponge /3-glucuronidase has a broad pH optimum between 6.5 and 4.0; at pH above 7.5 and beloti 3.0 its activity is lower than 20%. The divalent cations, Mg2+ as well as Ca2+ (at a molarity of 5 mM), have no effect on the enzyme activity; addition of 20 mM EDTA to the assays was also without influence on the enzyme reaction. For full enzyme activity, 200 mM NaCl must be added to the assays; in the absence of NaCl and in the presence of 500 mM NaCl, the enzyme activity is inhibited by more than 70%.
The temperature optimum was determined to be around 2O’C. The temperature coefficient Qlo (24) at 22”C:12”C is 2.6 and that at 22”C:32”C is 2.2. The molecular weight of the enzyme estimated from the elution behavior through Sepha- dex G-100 is approximately 80,000.
Properties of P-D -GaZactosidase-The pH dependence of the partially purified P-galactosidase is almost identical with the one observed in the case of /?-n-glucuronidase; the opti- mum is at 5.0. Addition of 5 mM Ca2+, 5 mM Mg”+, and 20 mM EDTA to the galactosidase assay is without influence on the enzyme activity. Estimations gave a salt optimum at 200 mM NaCl; at a low ionic strength (absence of NaCI), the activity decreased by 90% and, in the presence of higher salt concen- trations (500 mM NaCl), the enzyme reaction is reduced by 35%.
The temperature optimum lies around 22°C. The Q10 value of 22”C:12”C is 2.9 and that of 22”C:32”C is 1.8. The apparent molecular weight of the ,&galactosidase is about 25,000, as found by gel filtration with Sephadex G-100.
Localization of /3- Glucuronidase-The ,&glucuronidase ac- tivity was determined in the standard assay using either intact cells or subcellular fractions (cell-free extract, cell membrane fraction, and purified aggregation factor) as source (Table II). After incubation of intact cells in the standard glycosidase assay a ,B-glucuronidase activity of 10.4 pkat/107 cells was determined. From this finding alone, it was not possible to draw the conclusion, that a considerable amount of the en- zyme is localized on the outer cell surface because of possible leakage phenomena which might occur during the incubation at pH 5.0 in the standard assay. Therefore, the main subcel- lular fractions, the cell-free extract, the cell membranes, and the aggregation factor were tested for their enzyme activity. The experiment revealed that the aggregation factor is devoid
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

Controlled Intercellular Adhesion
TABLE II
1283
Localization of P-glucuronidase and /3-galactosidase in Geodia cells
The homogenate was prepared by suspending 10’ cells (dissociated (cell-free extract) was assayed; it contained 1.6 mg of protein/ml. The in ASW) in 1 ml of 10 mM Tris-HCl buffer (pH 7.5, containing 5 mM procedures for isolation of cells (in CMF medium), isolation of cell 2-mercaptoethanol and 0.1% Nonidet NP-40) and subsequently ho- membranes, and isolation of aggregation factor are mentioned under mogenizing in Dounce homogenizer with 10 strokes. The homogenate “Materials and Methods.” was then centrifuged (20,000 x g; 30 min; O’C) and the supernatant
SOUrCS
Intact cells
Input per assay
5 X 10’ cells
Specific P-glucuronidase activity Specific P-galactosidase activity
10.4 pkat/107 cells 3.0 pkat/107 cells
CelI extract
Ceil membrane
Aggregation factor
80 pg protein 26.1 pkat/mg protein 9.4 pkat/mg protein
45 pg protein 3.7 pkat/mg protein 2.1 pkat/mg protein
75 pg protein 0.1 pkat/mg protein 0.1 pkat/mg protein
of detectable /3-glucuronidase activity. The enzyme is predom- inantly located in the cell-free fraction; however, the mem- brane fraction (which has been pretreated (1 h; O’C) with 0.1% Nonidet NP-40 to obtain full enzyme activity; Ref. 25) contains /3-glucuronidase at a level of 3.7 pkat/mg of protein. From these data, we have strong indications that the ,B-glu- curonidase is not only located in the cytoplasm but also in the cell membrane.
Localization of ,&Galactosidase-In analogous studies, as outlined for ,&glucuronidase, the localization of P-galactosid- ase was determined (Table II). Again, the enzyme is absent in the aggregation factor complex. fi-Galactosidase activity was detected not only after incubation of intact cells and a cell- free extract in the standard assay but also after testing the cell membrane fraction. Therefore, it seems to be obvious that also the P-galactosidase is associated with cell membranes from Geodia.
Alterations of ,B-Glucuronidase- and /3-Galactosidase Ac- tivity during Reaggregation-As a first approach to the ques- tion whether the two glycosidases, studied in the present work, might be involved in the aggregation process, the amount of membrane-associated enzymes was determined during the early phase of formation of aggregates from single cells in the presence of the aggregation factor. Under the incubation conditions used, the kinetics of aggregation was almost identical with the one described previously (26). Ag- gregation factor-mediated reaggregation process starts after a lag phase of about 60 min and is completed 4 h after addition of the factor (formation of aggregates with a diameter of 2 to 3 mm).
After incubation for different periods of time, both the single cells and the cell aggregates were harvested (total, 25 + 5 lo7 cells) and then treated to isolate the cell membranes. Subsequently, the membrane-associated ,&glucuronidase- and P-galactosidase activity was determined. The results, plotted in Fig. 3, show for the two enzymes a strong increase of the respective activities between 3 and 5 h after the addition of the aggregation factor; during this period of time, the j?- glucuronidase activity increases by 92% and the ,8-galactosid- ase activity by 67%. After that, the activities of the glycosi- dases remain at this high level (average for ,&glucuronidase 0.48 pkat/lOO pg of cell membrane-protein and for /?-galacto- sidase 0.35 pkat/lOO ag). It should be mentioned that the strong increase of the glycosidase activities occurs during the same period of time at which the rate of reaggregation reaches a maximum.
Changes of ,L?-Glucuronosyltransferase- and j&Galactosyl- transferase Activity during Reaggregation-In experiments parallel to those determining the activity of glycosidase, the activity of the two glycosyltransferases, P-giucuronosyltrans- ferase and P-galactosyltransferase, which are associated with the extracellular high molecular weight particle carrying also
0 5 10
Time of incubation lh)
FIG. 3. Alterations of membrane-associated /3-glucuronidase and /3-galactosidase activity during reaggregation. A suspension (10 ml) of dissociated single cells was incubated in the presence of the aggrega- tion factor as described under “Materials and Methods.” At the time indicated on abscissa, both the aggregates and the nonaggregated cells were harvested. Subsequently, cell membranes were isolated, pretreated with 0.1% Nonidet (see “Materials and Methods”), and then assayed for /I-glucuronidase- (ordinate, left, +---+ and /I- galactosidase activity (ordinate, right, 0- - -0). The activity is re- ferred to 100 pg of cell membrane protein.
the aggregation factor (9), was determined in dependence on the aggregation process. From previous studies (3) it is known that the glycosyltransferases, as well as the aggregation factor, carrying high molecular weight particles remain integer and stable throughout the aggregation process. The particles can be reisolated after reaggregation by incubation of the aggre- gates in EGTA with a yield greater than 94%. Due to the fact that EGTA chelates selectively Ca’+, the activation ion for the transferases, magnesium remains in the ASW which is used as aggregation medium. Therefore, a determination of the transferase activity can be performed without a possible interference by the chelating agent. The experiments revealed (Fig. 4) that both the fl-glucuronosyltransferase and the ,5- galactosyltransferase activity remains constant during the early aggregation phase.
Influence of ,&Glucuronidase on Aggregation Receptor- From cytological experiments, it is known that the D-ghCU-
ronic acid moieties of the aggregation receptor play an impor- tant role in its binding to the aggregation factor (11). Analyt- ical studies revealed that 8.9% of the aggregation receptor consists of D-glucuronic acid (11). Hence, it seemed promising to check whether the P-glucuronidase, partially purified from the sponge Geodia, can use the aggregation receptor isolated from the same source as a substrate. Therefore, the aggrega- tion receptor was labeled in uiuo with u-[‘4C]glucuronic acid and subsequently partially purified (see “Materials and Meth- ods”). Analysis of the aggregation receptor by polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate revealed that the main radioactivity was observed at a relative
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from
1284 Controlled Intercellular Adhesion
FIG. 4. Influence of the aggregation process on the activity of the glycosyltransferases, associated with the extracellulsr high molecular weight particle. Dissociated single cells were incubated in the presence of the high molecular weight particle, also carrying the aggregation factor, as described in Fig. 3 and under “Materials and Methods.” At different time points, the assays, containing the aggregates, were treated with 20 mM EGTA (final concentration). After 20 min, the aggregates were completely redissociated into single cells. The sus- pension was centrifuged (20,000 x g; 10 min; 2°C) and the cell-free supernatant was dialyzed (12 h, 2°C) against saline followed by subsequent lyophilization. The residue was dissolved in 50 ,nl of distilled water and a 30-~1 aliquot was assayed for both P-glucurono- syltransferase activity (A----A) and /3-galactosyltransferase activity (A---A).
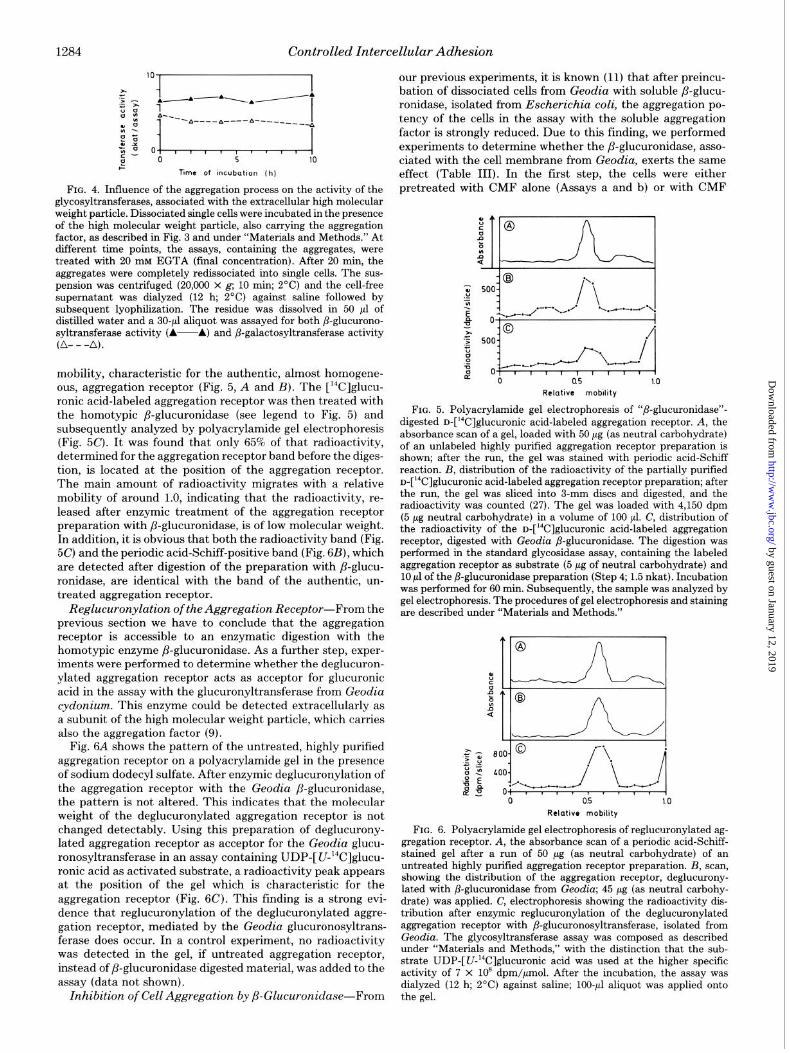
mobility, characteristic for the authentic, almost homogene- ous, aggregation receptor (Fig. 5, A and B). The [r4C]glucu- ronic acid-labeled aggregation receptor was then treated with the homotypic P-glucuronidase (see legend to Fig. 5) and subsequently analyzed by polyacrylamide gel electrophoresis (Fig. 5C). It was found that only 65% of that radioactivity, determined for the aggregation receptor band before the diges- tion, is located at the position of the aggregation receptor. The main amount of radioactivity migrates with a relative mobility of around 1.0, indicating that the radioactivity, re- leased after enzymic treatment of the aggregation receptor preparation with P-glucuronidase, is of low molecular weight. In addition, it is obvious that both the radioactivity band (Fig. 5C) and the periodic acid-Schiff-positive band (Fig. 6B), which are detected after digestion of the preparation with p-glucu- ronidase, are identical with the band of the authentic, un- treated aggregation receptor.
Reglucuronylation of the Aggregation Receptor-From the previous section we have to conclude that the aggregation receptor is accessible to an enzymatic digestion with the homotypic enzyme ,8-glucuronidase. As a further step, exper- iments were performed to determine whether the deglucuron- ylated aggregation receptor acts as acceptor for glucuronic acid in the assay with the glucuronyltransferase from Geodia cydonium. This enzyme could be detected extracellularly as a subunit of the high molecular weight particle, which carries also the aggregation factor (9).
Fig. 6A shows the pattern of the untreated, highly purified aggregation receptor on a polyacrylamide gel in the presence of sodium dodecyl sulfate. After enzymic deglucuronylation of the aggregation receptor with the Geodia ,&glucuronidase, the pattern is not altered. This indicates that the molecular weight of the deglucuronylated aggregation receptor is not changed detectably. Using this preparation of deglucurony- lated aggregation receptor as acceptor for the Geodia glucu- ronosyltransferase in an assay containing UDP-[ U-“‘C]glucu- ronic acid as activated substrate, a radioactivity peak appears at the position of the gel which is characteristic for the aggregation receptor (Fig. 6C). This finding is a strong evi- dence that reglucuronylation of the deglucuronylated aggre- gation receptor, mediated by the Geodia glucuronosyltrans- ferase does occur. In a control experiment, no radioactivity was detected in the gel, if untreated aggregation receptor, instead of ,&glucuronidase digested material, was added to the assay (data not shown).
Inhibition of Cell Aggregation by ,L?-Glucuronidase-From
our previous experiments, it is known (11) that after preincu- bation of dissociated cells from Geodia with soluble ,8-glucu- ronidase, isolated from Escherichia coli, the aggregation po- tency of the cells in the assay with the soluble aggregation factor is strongly reduced. Due to this finding, we performed experiments to determine whether the fi-glucuronidase, asso- ciated with the cell membrane from Geodia, exerts the same effect (Table III). In the fist step, the cells were either pretreated with CMF alone (Assays a and b) or with CMF
Relatwe mobdtty
FIG. 5. Polyacrylamide gel electrophoresis of “/&glucnronidase”- digested D-[‘4C]glucuronic acid-labeled aggregation receptor. A, the absorbance scan of a gel, loaded with 50 pg (as neutral carbohydrate) of an unlabeled higbly purified aggregation receptor preparation is shown; after the run, the gel was stained with periodic acid-Schiff reaction. B, distribution of the radioactivity of the partially purified n-[W]glucuronic acid-labeled aggregation receptor preparation; after the run, the gel was sliced into 3-mm discs and digested, and the radioactivity was counted (27). The gel was loaded with 4,150 dpm (5 pg neutral carbohydrate) in a volume of 100 ~1. C, distribution of the radioactivity of the D-[‘4C]glUCUrOniC acid-labeled aggregation receptor, digested with Geodia /3-glucuronidase. The digestion was performed in the standard glycosidase assay, containing the labeled aggregation receptor as substrate (5 pg of neutral carbohydrate) and 10 ~1 of the /3-glucuronidase preparation (Step 4; 1.5 nkat). Incubation was performed for 60 min. Subsequently, the sample was analyzed by gel electrophoresis. The procedures of gel electrophoresis and staining are described under “Materials and Methods.”
Rclotlve mobMy
FIG. 6. Polyacrylamide gel electrophoresis of reglucuronylated ag- gregation receptor. A, the absorbance scan of a periodic acid-Schiff- stained gel after a run of 50 pg (as neutral carbohydrate) of an untreated highly purified aggregation receptor preparation. B, scan, showing the distribution of the aggregation receptor, deglucnrony- lated with /3-glucuronidase from Geodia; 45 pg (as neutral csrbohy- drate) was applied. C, electrophoresis showing the radioactivity dis- tribution after enzymic reglucuronylation of the deglucuronylated aggregation receptor with /?-glucuronosyltransferase, isolated from Geodia. The glycosyltransferase assay was composed as described under “Materials and Methods,” with the distinction that the sub- strate UDP-[U-‘4C]glucuronic acid was used at the higher specific activity of 7 x lo8 dpm/pmol. After the incubation, the assay was dialyzed (12 h; 2°C) against saline; 100~~1 aliquot was applied onto the gel.
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

Controlled Intercellular Adhesion 1285
plus Nonidet (Assay c). The mild, nonionic surface active agent, Nonidet, was used to remove the ,LLglucuronidase from the cell membrane (25), without destroying its lipid layer (28). Subsequently, the cells were transferred into media which are either optimal for /3-glucuronidase reaction (phosphate buffer, pH 5.0) or which suppress the action of P-glucuronidase (CMF). The determination of the cell membrane-bound ,0- glucuronidase activity of the cells after the pretreatment and incubation procedures revealed that those cells, which had been pretreated with Nonidet (Assay c), have lost 80% of their ,f3-glucuronidase activity.
In the crucial set of experiments, the aggregation potency of cells, either containing (Table III, Assay a and b) or lacking (Assay c) /3-glucuronidase activity, was determined. ,f3-Glucu- ronidase-containing cells, which had been pretreated and in- cubated under assay conditions (CMF, Assay a), at which this glycosidase is inactive, have not lost any aggregation potency during the incubation period. Cells, which are lacking cell surface-bound P-glucuronidase, (pretreated with CMF plus Nonidet) show the same aggregation potency as the controls (Assay a), even though they are incubated under assay con- ditions, found to be optimal for &glucuronidase (Assay c). In contrast, those cells which have not lost their membrane- bound ,&glucuronidase during the pretreatment period (in CMF) but were incubated in the phosphate buffer (pH 5.0, NaCl and 2-mercaptoethanol), which allows optimal ,6-glucu- ronidase reactivity, show a strongly reduced aggregation po- tency (Assay b). The latter result led us to assume that the cell membrane-bound ,&glucuronidase reduces under optimal assay conditions the extent of the aggregation potency of the cells. The next section presents the evidence that the reason for this reduced ability of the cells to reaggregate is indeed due to a /3-glucuronidase-mediated deglucuronylation of a cell membrane component.
TABLE III
Inhibitorypotency of the cell membrane-bound Geodia /3- glucuronidase on the aggregation potency of Geodia cells
Geodia cells, 2.5 x 10’ (dissociated in CMF), were pretreated for 12 h at 0°C in (a) 10 ml of CMF, (b) 10 ml of CMF, and (c) 10 ml of CMF plus 0.1% Nonidet NP-40. Subsequently, the cells from Assay a were transferred by centrifugation (2,000 x g; 10 min; 2’C) into 10 ml of CMF, the ones from Assays b and c into 10 ml of 10 rnM sodium phosphate buffer (pH 5.0, containing 200 mM NaCl and 5 mrvr 2. mercaptoethanol) and incubated for 2 h at 20°C. After incubation of the cells, the membrane-bound ,&glucuronidase activity was deter- mined after transfer of 5 X 10’ cells into 50 ~1 of the above-mentioned phosphate buffer and assayed in the standard glycosidase assay with p-nitrophenyl-P-n-glucuronide as substrate. The aggregation potency of the cell was measured before and after incubation of the cells in CMF or phosphate buffer. For this determination, the standard aggregation assay, containing lo4 AU/ml of Geodia aggregation fac- tor, was used; the diameter of the aggregates was estimated 60 min after addition of the aggregation factor. Further details are given under “Materials and Methods.”
Diameter of aggre- /3-Glucuronidase gates
Assay Pretreatment Incubation of activity aftrr in- of the cells the cells cub&ion of the Before After in-
cells incuba- cub&ion tion
pkat/lO’ cells w
a CMF CMF 9.3 1,150 1,200
b CMF Phosphate 9.1 1,100 350 buffer (pH 5.0)
C CMF plus Phosphate 1.7 1,350 1,250 Nonidet buffer (pH
5.0)
TABLE IV
Aggregation potency of reglucuronylated Geodia cells
Geodia cells 2.5 x 10’ (dissociated in CMF) were pretreated for 2 h at 20°C in 10 ml of sodium phosphate buffer (pH 5.0, containing 206 mix NaCl and 5 mM 2-mercaptoethanol) (Assays a to c). Then the cells were transferred by centrifugation (2,000 x g; 10 min; 2°C) into a 50 mM Tris-HCl buffer (pH 8.2; 300 mM NaCl, 0.5 mM UDP- glucuronic acid, 10 mM EGTA containing either 10 mM MgCle (Assay a) or 10 mM MgC12 and 150 pg of glucuronosyltransferase (referred to protein)/ml (Assay b), or 10 mM MgC12, 150 pg of glucuronosyltrans- ferase/ml and 10 mM DTNB (Assay c); the cell density was adjusted to 25 -t 5 x 10” single cells/ml. Incubation was performed for 2 h at 22°C. The ability of the cells to reaggregate was determined before and after the incubation period of the cells by transferring them into the standard aggregation assay, containing lo4 AU/ml of Geodia aggregation factor; the diameter of the aggregates was determined 60 min after addition of the aggregation factor.
Diameter of aggre- gates
Assay Pretreatment of the cells Incubation of the cells Before
incuba- After in-
tion cubation
Km
a Phosphate buffer Tris (pH 8.2) plus 350 400 (PH 5.0) Mg*+
b Phosphate buffer Tris (pH 8.2) plus 350 1,550 (PH 5.0) Mg*+ and transfer-
ase
C Phosphate buffer Tris (pH 8.2) plus 350 450 (PH 5.0) Mg”, DTNB and
transferase
Role of Glucuronosyltran.sferase during Cell Aggrega-
tion-For this series of experiments (Table IV), the disso- ciated single cells were pretreated in the phosphate buffer (pH 5.0) for 2 h at 20°C to deglucuronylate cell membrane components by the membrane-bound ,&glucuronidase (see previous chapter). Subsequently, the cells were transferred into the Tris buffer (pH 8.2) which has been shown to be optimal for glucuronosyltransferase (9) but which suppresses the activity of ,&glucuronidase (this work). The cells were incubated for 2 h at 22°C either in the absence (Assay a) or presence of the glucuronosyltransferase (Assays b and c). Due to the fact that the transferase is associated with the aggre- gation factor, the biological activity of the latter component was suppressed by addition of the selective calcium-chelating agent EGTA. In a previous study, it was established that, only in the presence of Ca’+, can the aggregation factor exert its aggregation promoting activity; Mg”+ was found to be inactive in this reaction (7). In Assay c, DTNB, a thiol-alkylating reagent (29), was added to inactivate glucuronosyltransferase; 10 mM DTNB abolishes the transferase activity by more than
90%. The results, summarized in Table IV, show that the aggre-
gation potency of cells, carrying on their cell surface deglu- curonylated molecules, can be restored after incubation with glucuronosyltransferase and UDP-glucuronic acid (Assay b). Cells incubated in the absence of the transferase (Assay a) or in the presence of DTNB-inactivated glucuronosyltransferase (Assay c) do not show a marked increase in their ability to form large aggregates in the presence of the aggregation factor. Due to the finding that the aggregation receptor, a glycopro- tein with glucuronic acid termini, is a prerequisite for aggre- gation factor-mediated reaggregation (10, 11, SO), we have to assume that the aggregation receptor molecules have been reglucuronylated during the incubation of the cells in the presence of the glucuronosyltransferase and UDP-glucuronic acid.
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

1286 Controlled Intercellular Adhesion
DISCUSSION
The studies with embryonic retina (1, 2), brain (31), and sponge cells (4, 5) have provided experimental evidence that molecules with ligand activity (aggregation factors) cross-link cells and thereby enhance cell attachment and cell aggrega- tion. The data available indicate that they may be glycopro- teins which the aggregation factors attach to the cell mem- brane (10, 30, 32). Actually, three hypotheses are offered explaining the involvement of carbohydrates in the intercel- lular adhesion: antibody-antigen hypothesis (33), the hydro- gen bond concept (34), and the substrate-enzyme mechanism proposed by Roseman (35). In the latter interesting theory, it is suggested that a glycosyltransferase on one cell surface binds to its receptor molecule on the surface of another cell. I f the sugar nucleotide appropriate for the enzyme is then provided, glycosylation takes place, and this sugar is added to the growing chain. When no sugar nucleotide is present, the reaction does not complete and the enzyme on one cell surface is supposed to remain bound to the substrate on the other cell surface. Thus, the addition of the nucleotide sugar allows completion of the enzymic reaction, with the subsequent loss of the bond between the two cells. Objections have arisen over Roseman’s theory because of the lack of direct enzymic data demonstrating the existence of glycosyltransferases on the outer cell surface of mammalian cells (36). In addition, the hypothesis does not take into account the presence of glyco- sidases on the cell surface. At least since the studies of Bos- mann (37, 38), it seems to be established that the glycosidase activity alters with the growth state and possibly with the aggregation potency of the cell. The data of Aoyagi et al. (13) conclusively demonstrate that some glycosidases are also as- sociated with the cell membrane.
In the sponge (Geodia cydonium) model, used for the present study, several glycosyltransferases (sialyltransferase, galactosyltransferase, and glucuronyltransferase; Refs. 8 and 9) could be clearly localized in the extracellular space. In addition, and in contrast to the Roseman theory, the aggre- gation factor can be separated from glycosyltransferases (8), which indicates that the glycosyltransferases might modulate the aggregation potency of the cells, but that they do not ultimately provoke adhesiveness between two cells. The latter process is, at least in the Geodia system, a result of an interaction between the aggregation factor present in the intercellular space and the aggregation receptor bound to the cell surface (10). In the presented study, it could be elucidated for the first time, that cell surface-associated P-glucuronidase can modulate the aggregation potency of sponge cells. It was found that the activity of both the cell surface-associated /?- glucuronidase and the cell surface-bound ,C?-galactosidase in- crease during the reaggregation process. In addition, the sol- uble /?-glucuronidase was found to hydrolyze the terminal glucuronic acid moiety of the aggregation receptor under appropriate incubation conditions. On the other side, incuba- tion of the intact cells under assay conditions, optimal for membrane-bound fl-glucuronidase, reduces markedly the ag- gregation potency of the cells. The strong hint, that this reduction is due to the hydrolyzing action of the bound ,C glucuronidase, resulting in a liberation of glucuronic acid from the aggregation receptor, came from the finding that after incubation of these cells with glucuronosyltransferase (iso- lated from the same sponge) in the presence of UDP-glucu- ronic acid, the original aggregation potency of the cells is restored. The indirect evidence that it is the aggregation receptor, bound to the cell surface which is deglucuronylated or reglucuronylated, is supported by the presented data, ob- tained from the experiments with isolated aggregation recep- tor molecules. These studies showed that the receptor deglu-
curonylated in the presence of isolated Geodia P-glucuroni- dase can be reglucuronylated in the presence of Geodia /3- glucuronyltransferase and UDP-glucuronic acid. From the comparison of the mobility pattern of the deglucuronylated aggregation receptor in polyacrylamide gels with those of the untreated or reglucuronylated aggregation receptor prepara- tions, which are almost identical, we have to conclude that the ,&glucuronidase is an ectoglycosidase.
As a direct approach to demonstrate that under the influ- ence of P-glucuronidase the aggregation potency of the Geodia cells is abolished, preformed aggregates were treated with heterotypic ,&glucuronidase (from E. coli) at assay conditions described elsewhere (11). The results revealed that, at an enzyme concentration of 0.00025 unit/ml, the size of the aggregates is reduced by more than 50% during an incubation period of 30 min (11).
Even if the purity of the membrane fractions was checked by chemical analysis and by the measurement of a marker enzyme (5’-nucleotidase), the possibility cannot be excluded that the membrane preparations used in the present study are free of cytoplasmic contaminations. Already from the studies of Warren et al. (15) it is known that cell membranes, obtained by the Tris method which was applied in our investigation, are associated with vesicles of undetermined origin. Therefore, future studies must answer the question, whether these vesi- cles contain glycosidases. As suggested by Reutter et al. (39), vesicles which arise by blebbing of the Go&i cisternae or of the lysosomes migrate to the cell periphery carrying glycosyl- transferases and glycosidases on their inner surface. When fusing with the plasma membrane, the vesicles might be turned inside out, thus exposing the enzyme molecules. There- fore, the observed increase in membrane-associated enzyme activity during aggregation (Fig. 3) may be the result because membranes prepared from aggregates are “contaminated” with vesicles to a greater extent than membranes from indi- vidual cells (15).
From our data, obtained in the Geodia system, we postulate a working hypothesis to explain cell aggregation and cell separation on the level of an interaction between glycosidases and glucuronidases in the following series: (a) activation of the aggregation receptor by its enzymic glucuronylation; (6) adhesive recognition of two cells, mediated by the aggregation factor and the glucuronylated aggregation receptor; (c) inac- tivation of the aggregation receptor by its deglucuronylation with the membrane-associated ,&glucuronidase; and (d) cell separation due to the loss of the recognition site (glucuronic acid) of the aggregation receptor for the aggregation factor. Steps c and d are consequences of an “activation” of the membrane-associated ,&glucuronidase, which is inactive un- der the pH and ion conditions of the sea water. No data are available which give direct experimental evidence for a pH or ion change of the sponge cell membrane during the process of cell separation. However, from previous studies (40), it is known that the aggregation factor from Geodia alters the cell surface charge after its binding to homologous cells. Due to the fact that an alteration of the cell surface charge is also caused by a pH change at the cell surface (41), it is quite likely that pH fluctuations govern the activity of both the glycosi- dase and the glycosyltransferase. In addition to the described aggregation-dissociation hypothesis based on the interrelation between glycosyltransferases and glycosidases, a further mechanism (reversible inactivation of the aggregation factor by an antiaggregation receptor) exists which controls cell adhesion ( 12).
REFERENCES
1. Thiery, J.-P., Brackenbury, R., Rutishauser, U., and Edelman, G. M. (1977) J. Biol. Chem. 252,6841-6845
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

Controlled Intercellular Adhesion 1287
2. Hausman, R. E., and Moscona, A. A. (1976) Proc. Natl. Acad. Sci. U. S. A. 73, 3594-3598
3. Muller, W. E. G., Muller, I., and Zahn, R. K. (1978) Aggregation in Sponges, Research in Molecular Biology, Vol. 8, pp. l-87, Akademie der Wissenschaften und der Literatur, Maim
4. Moscona, A. A. (1962) J. Cell. Comp. Physiol. 60, Suppl. 1, 65-80 5. Miller, W. E. G., and Zahn, R. K. (1973) Exp. Cell Res. 80,95-104 6. Zahn, R. K., Miiller, W. E. G., Geisert, M., RemmulIer, J., Ml-
chaelis, M., Pondeljak, V., and Beyer, R. (1976) Cell Diff 5, 129-137
Ga.Miiller, W. E. G., Zahn, R. K., Arendes, J., Kurelec, B., Steffen, R.. and Miiller. I. (1979) Biochim. Bioohys. Acta. in mess
7. Muller, W. E. G.; Muller, I., and Zahn, k k. (1974) Eiperientia 30,899-902
8. Muller, W. E. G., Arendes, J., Kurelec, B., Zahn, R. K., and Miller, I. (1977) J. Biol. Chem. 252, 3836-3842
9. Muller, W. E. G., Zahn, R. K., Kurelec, B., Mdller, I., Uhlenbruck, G., and Vaith, P. (1978) Hoppe-Seyler’s Z. Physiol. Chem. 359, 529-537
10. Muller, W. E. G., Muller, I., Zahn, R. K., and Kurelec, B. (1976) J. Cell Sci. 21, 227-241
11. Vaith, P., Miiller, W. E. G., and Uhlenbruck, G. (1979) Dev. Comp. Immunol., in press
12. Miiller, W. E. G., Zahn, R. K., Kurelec, B., and Miiller, I. (1979) Proc. Natl. Acad. Sci. U. S. A., in press
13. Aoyagi, T., Suda, H., Nagai, M., Ogawa, K., Suzuki, J., Takeuchi, T., and Umezawa, H. (1976) Biochim. Biophys. Acta 452, 131-143
14. Muller, W. E. G., Muller, I., Kurelec, B., and Zahn, R. K. (1976) Exp. Cell Res. 98, 31-40
15. Warren, L., Glick, M. C., and Nass, N. K. (1967) J. Cell Physiol. 68,269-288
16. Determann, H. (1967) Gelchromatographie, Springer, New York 17. Bosmann, H. B., and Bernacki, R. J. (1970) Exp, Cell Res. 61,
379-386 18. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
(1951) J. Biol. Chem. 193, 265-275 19. Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., and
Smith, F. (1956) Anal. Chem. 28, 350-356
20. Hinsberg, K., and Lang, K. (1951) Medizinische Chemie, pp. 245-307, Urban and Schwarzenberg, Munchen
21. Muller, W. E. G., Schroder, H. C., Steffen, R., Zahn, R. K., and Dose, K. (1977) Mol. Biol. Rep. 3, 331-337
22. Laemmli, U. K. (1970) Nature 227,680-685 23. Zacharius, R. M., Zell, T. E., Morrison, J. H., and Woodlock, J. J.
(1969) Anal. Biochem. 30, 148-152 24. Netter, H. (1959) Theoretische Biochemie-Physihalisch chem-
ische Grundlagen der Lebensvorgiinge, p. 557, Springer-Ver- lag, Berlin
25. de Duve, C., Pressman, B. C., Gianetto, R., Wattiaux, R., and Appelmans, F. (1955) Biochem. J. 60,604-617
26. Muller, W. E. G., Muller. I., Pondellak. V.. Kurelec. B.. and Zahn. R. K. (1978) Differentiation 10,45-53
27. Miiller. W. E. G.. Seibert. G.. Bever. R.. Breter. H. J.. Maidhof. A., and Zahn, k. K. (1977) Can’cer’Res. 37, 3824-3833 ’
28. Helenius, A., and Simons, K. (1975) Biochim. Biophys. Acta 415, 29-79
29. Weitzman, P. D. J. (1975) Anal. Biochem. 64,628-630 30. Muller, W. E. G., Zahn, R. K., Kurelec, B., and Muller, I. (1978)
Differentiation 10, 55-60 31. Merrell, R., Gottlieb, D. I., and Glaser, L. (1975) J. Biol. Chem.
250, 5655-5659 32. Turner, R. S., and Burger, M. M. (1973) Nature New Biol. 244,
509-510 33. Tyler, A. (1946) Growth 10, 7-19 34. Percival, E. G. V., and Percival, E. (1962) Structural Carbohy-
drate Chemistry, J. Garnet Miller, Ltd., London 35. Roseman, S. (1970) Chem. Phys. Lipids 5, 270-297 36. Deppert, W., Wechaw, H., and Walter, G. (1974) Proc. Natl.
Acad. Sci. U. S. A. 71, 3068-3072 37. Bosmann. H. B. (1969) Exp. Cell Res. 54. 217-221 38. Bosmann; H. B. (1972) B&him. Biophys. Acta 264, 339-343 39. Reutter, W., and Bauer, C. (1978) in Morris Hepatomas (Morris,
H. P., and Criss, W. E., eds) pp. 405-437, Plenum, New York 40. Muller, W. E. G., Zahn, R. K., Kurelec, B., and Muller, I. (1978)
Wilhelm Roux’Arch. Dev. Biol. 184,29-40 41. Curtis, A. S. G. (1967) The Cell Surface: Its Molecular Role in
Morphogenesis, p. 70, Academic Press, New York
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

W E Müller, R K Zahn, B Kurelec, I Müller, G Uhlenbruck and P Vaithglycosidases.
adhesion, basing on the interrelation between glycosyltransferases and Aggregation of sponge cells. A novel mechanism of controlled intercellular
1979, 254:1280-1287.J. Biol. Chem.
http://www.jbc.org/content/254/4/1280.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/4/1280.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from