adaptation to day length and yield stability of families from 4x×2x crosses in potato
TRANSCRIPT

Euphytica 56: 187-195, 1991. (~) 1991 Kluwer Academic Publishers. Printed in the Netherlands.
Adaptation to day length and yield stability of families from 4x × 2x crosses in potato
Rodomiro Ortiz 1, Rosanna Freyre 2, Stanley J. Peloquin 3 & Masaru Iwanaga 1 1 Department of Horticulture, University of Wisconsin-Madison, 1575 Linden Drive, Madison, WI 53706, USA; 2 International Potato Center (CIP). Present address: Department of Crop and Soil Sciences, Michigan State University, East Lansing, MI 48915, USA; 3 Corresponding author: Departments of Genetics and Horticulture, University of Wisconsin-Madison, 1575 Linden Drive, Madison, WI 53706, USA; 4 CIP. present address: CIAT, Apartado Aereo 6713, Cali, Colombia
Received 17 June 1991; accepted 3 September 1991
Key words: allelic diversity, FDR 2n pollen, genotype x environment interaction, parental selection, response to day length, yield stability, potato (Solanum tuberosum)
Summary
The objectives of this research were to evaluate the response to day length and the yield stability of environments of 4x families from 4x x 2x crosses in potato. Twenty families from crosses between 4x female parents with different genetic backgrounds and 2x parents, which formed 2n pollen by first division restitution and were selected under short days, were evaluated in four locations in Peru (short days) and during two years at Rhinelander, USA (long days). Results from the USA indicate that the 4x progeny from 4x × 2x crosses were very late maturing and poor yielding.
Further, vine maturity, tuber size and tuber number are highly correlated with the percentage of Tuberosum germplasm in the parentage. Results from Peru indicate that most 4x × 2x families had high tuber yield, stability for yield and high dry matter content. The lack of correlation between rank order of the parental value, based on progeny testing, of the male and female parents among short day locations and between short and long day environments indicates that selection of parents must be done in each location in which the final product, either a family or a clone, will be used.
Introduction
The cultivated potato of short day environments from South America exhibits late vine maturity and very poor yields under long day conditions. Long days promote elongation of stems and long stolons while the opposite effects are observed under short days. Likewise, the number of tubers increases and the tuber weight decreases as daylength increases. Mendoza & Haynes (1976, 1977) indicated that photoperiod strongly affects both tuberization and
maturity. This suggests that both traits could be under control of either a) the same major gene(s) controlling photoperiod reaction or b) modifier genes for each trait.
Ross (1986) indicated that more than 50 traits should be combined in a modem potato variety. As in other crops new potato cultivars must primarily combine yield, yield stability and quality traits. Allard (1966) suggested that the degree of genetic diversity (allelic diversity) in a population deter- mines the extent to which stable yields can be

188
achieved over a long period of time and over differ- ent environments. In this regard, Amoros & Men- doza (1979) used five different populations of pota- to with different genetic backgrounds (group Tu- berosum, group Andigena - Neotuberosum, and group Phureja, and crosses between them), and compared these populations in six locations in Pe- ru. They found a correlation between the level of genetic diversity and yield, and highest yield stabil- ity in the population with the most genetic diversi- ty.
The use of 2n gametes in potatoes provides the opportunity to increase the allelic diversity through the use of wild 2x relatives. This approach could lead to yield improvement and yield stability. In effect de Jong et al. (1981) and Darmo & Peloquin (1990) found that the more highly heterozygous parallel spindle derived 4x clones from 4x × 2x crosses had higher yield and better yield stability than standard 4x cultivars in different environ- ments. A simple mendelian recessive meiotic mu- tant, ps, is responsible for the parallel orientation of the spindles at the second meiotic division, which result in the formation of 2n pollen. This mode of 2n pollen formation is genetically equiv- alent to a first division restitution (FDR) mecha- nism. It has been estimated that approximately 80% of the heterozygosity and a large fraction of the epistatic interactions are transmitted from par- ent to offspring through 4x x 2x crosses using FDR 2n gametes (Douches & Quiros, 1987; Peloquin et al., 1989).
A large genotype by environment interaction, especially for tuber yield, has been a major prob- lem in clonal selection in potatoes. It results in an increase in the number of seasons and locations needed for advanced clonal testing. Concilio (1985) indicated that there could be a causal relationship between the level of heterozygosity and homeosta- sis, and that this could be responsible for better performance of the 4x × 2x clones than the 4x × 4x clones evaluated in more unfavorable environ- ments. Therefore, as indicated by Peloquin et al. (1989), selection for yield in single hills may be more effective in progeny from 4x × 2x crosses than in progeny from 4x × 4x crosses. This could be due to the homeostasis resulting from the high
level of heterozygosity, and also a higher percent- age of high yielding 4x progeny from 4x × 2x cross- es as compared to the progeny from 4x × 4x cross- es.
The objectives of the present research were to: a) evaluate the yield of 4x progeny from 4x x 2x crosses in long day environments using 2x parents selected under short days, and b) determine the yield stability of 4x × 2x families, with wide genetic backgrounds, in four short day locations in Peru.
Materials and methods
The pedigrees, species background and geograph- ical origin of the 4x and 2x parents are listed in Table 1. The four FDR 2x male parents from the International Potato Center Breeding Program (selected under short days) were crossed with six females: Serrana, LT-1, HP-278.22, Katahdin, At- lantic and AVRDC-1287.19 following a line × test- er mating design. The male parents were 84.36.29 and 85.37.38, both with resistance to root-knot nematode (Iwanaga et al., 1989), and MI-49.10 and 84.194.30, bacterial wilt resistant clones.
The environments used were located in Peru (short days) and the USA (long days). Short day environments were characterized as having ap- proximately 12 hours photoperiod throughout the growing season; the long day environment was characterized as having approximately 16 hours photoperiod in early growth: which gradually de- creased to 12 hours at harvest.
Twenty families from 24 possible combinations, were obtained and evaluated during 1988 and 1989 under long day conditions at Rhinelander, WI (USA). The two US cultivars used as parents, At- lantic and Katahdin, were also included in the yield trial at Rhinelander. Fifteen families (five families of those used at Rhinelander were not included) were evaluated under short day conditions at four locations in Peru during 1989 and 1990. The loca- tions in Peru represent different agroecological re- gions of the country. La Molin a (240 m altitude) is located in a coastal desert. Huancayo (3260 m alti- tude) and Huaraz are in the cool Andean high- lands. San Ramon (800 m altitude) is in the mid-

elevation jungle of Peru on the eastern slope of the Andes.
During the first year, 75 true seeds from each family were planted in flats (in both short and long day locations) and later the seedlings were trans- planted to the field using two replications of 20 seedlings each in a randomized complete block design (RCBD). Each family was planted in one row. At harvest time, one medium size tuber was saved from 15 seedlings of the two replications for the clonal trial the following year. These tubers were planted in two replications of 15 plants in each location the next year. The experiments were har- vested at 90 days in Rhinelander and San Ramon, and after 120 days in La Molina, Huancayo and Huaraz.
Data recorded for all the Peruvian locations on a plot basis were: (1) Plant vigor, scored 50 days after planting using
a 1 to 9 scale, where 1 indicated the poorest and 9 the most vigorous;
(2) Plant uniformity, scored 50 days after planting using a (very nonuniform) to 9 (very uniform);
(3) Haulm earliness, scored 30 days before har- vest, using a scale 1 (earliest) to 9 (latest);
189
(4-5) Total and marketable tuber yield (tubers with more than 50 g) per plant in kilograms;
(6) Number of tubers per plant; (7) Average tuber weight (in kg), by dividing the
total tuber yield per plant by the number of tubers per plant;
(8) General tuber appearance (GTA), using a scale 1 (excellent) to 5 (poor); and
(9) Specific gravity (total solids), by weighing the tubers in air (x) and in water (y). Thus, specific gravity = x / (x - y).
Plant vigor and uniformity, and earliness were evaluated at Rhinelander using the same scale. The families from crosses between parents developed in short days were too late for the growing season at Rhinelander (long days). Therefore, an accurate estimation of total tuber yield and its components, to distinguish between parents, was not possible in either the seedling (1988) or the tuber (1989) gener- ations. Thus, a visual index score was used for the evaluation of tuber traits in both years. The scale for total yield ranged from 1 (poor yield with many small tubers) to 3 (high yield with few large tubers). The scale for tuber set ranged from 1 (less than 15 big sized tubers) to 3 (more than 30 small sized tubers).
Table I. Pedigrees, species background and geographical origin of 4x and 2x parents
Clone Pedigree Species ~ (Origin)
4x females: Serrana MPI 59703121 x (Selecta Balcarce 1 x {Saranac x Eariaine))
Katahdin
Atlantic
S40568 (Busola x Rural New Yorker) x $24642 (Sutton Flowerball x Aroostook Wonder) Lenape x Wauseon.
AVRDC-1287.19 (adg x {chc, rap, sgr)) x tbr LT-1 Katahdin x Aquila
HP-278.22 tbr x (2x-tbr x phu) FDR 2x male parents selected in Peru. 84.36.29 (2x of (T-169 x PVY bulk} x HT bulk) 381 bulk 84.194.30 (80M x BW) x MI-49.10 85.37.38 84.194;71 x (2x of {T-169 x PVY bulk} x 381 bulk) MI-49.10 (2x of (T-169 x PVY bulk) x BW.11
sto, tbr (Argentina) tbr (USA) tbr, ehc (USA) (Taiwan) tbr, dins, adg (Peru) (Peru)
adg, tbr, phu, stn, spl adg, phu, stn, spl, chc, tbr adg, phu, tbr, stn, spl, chc adg, tbr, spl, phu
1 Species abbreviations according to Huaman and Ross (1985).
190
Analyses of variance for RCBD at each location and combined over short day environments (four locations evaluated during 1989-1990) and long day environments (two years at one location) were per- formed using MSTATC (1989). The parental value of males and females was estimated using the sub- program NONORTHO, which is recommended in cases of missing results for a factorial analysis. NONORTHO calculated the means in a two-way table, computing corrected averages in both direc- tions, along with an analysis of variance, giving corrected mean squares for both groupings, inter- actions, and within cells. Likewise, NONORTHO computed an accurate F value and probability per- centage with missing values. The interaction de- gree of freedom was computed as the product of the two group variable degrees of freedom minus a degree of freedom for each missing combination. The interaction mean square was used for the error term. The mean squares for males and females obtained from the analysis of variance of the line x tester design were equivalent to the GCA of males and females respectively, meanwhile the expected mean square of the interaction source was equiv- alent to the SCA.
Phenotypic correlations between traits were cal- culated for all possible pair combinations under long and short day environments.
The analysis of tuber yield stability for family performance under short day conditions was done following the model proposed by Eberhart & Rus- sel (1966). The genotype means obtained for sever- al environments are regressed against the envi- ronmental means for all the genotypes tested. The parameters of the model were Y = a + bX, where Y was the family tuber yield, a the intercept, b the regression coefficient or slope, and X the envi- ronmental mean yield. Families with b = 1, r 2 (coefficient of determination) > 0.50, and sd (de- viation mean squares)= 0 were considered as a stable genotype (Stofella et al., 1984). The estimate of the stability parameter for yield, b, was consid- ered to be consistent across environments if it had a minimum variance, i.e., Sb = 0. Thus, the families were grouped according to the tuber yield stability parameters as (a) stable and consistent if b = 1, r > 0.7, and Sb = 0, (b) unstable and consistent if b <
> 1, r < 0.7, and s b = 0, (C) stable but inconsistent if b = 1, r > 0.7 and Sb > 0, and (d) unstable and inconsistent if b < > 1, r < 0.7 and %> 0.
Spearman's rank correlations between the pa- rental performance of males and females in the different locations were calculated. Regression analyses using the percentage of adapted germ- plasm as the independent variable and the traits evaluated as the dependent variables were per- formed to estimate the effect of adaptation to long days at Rhinelander.
Results
Long days
None of the 4x progenies from 4x x 2x crosses had tuber yields with more than 0.7 kg/plant and most families had significantly less tuber yield per plant.
The analysis of variance for several traits eval- uated in 20 families during two years under long
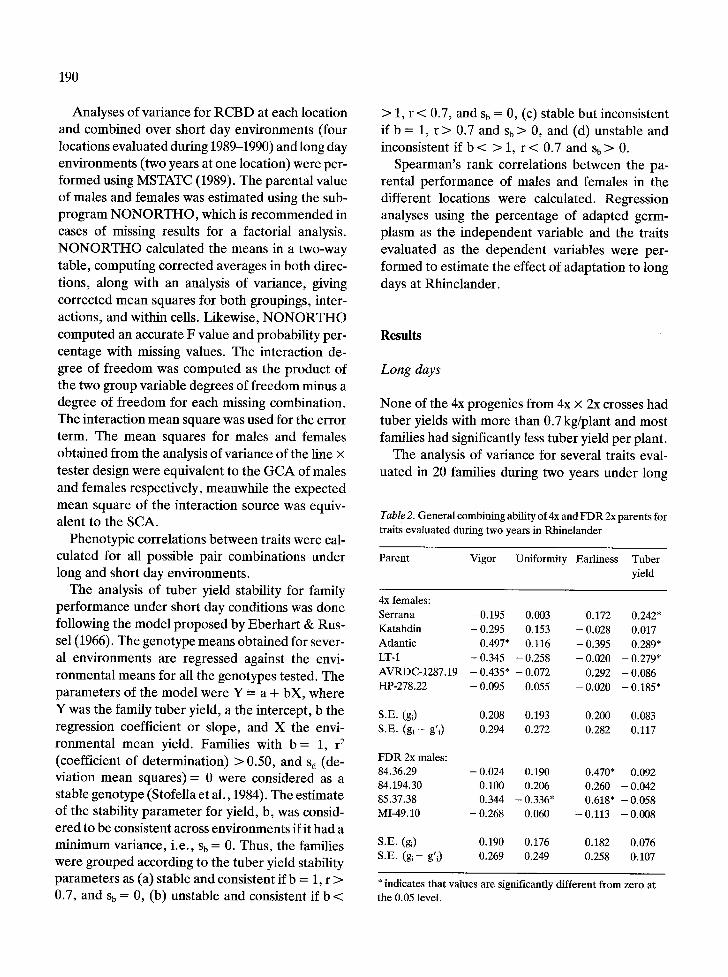
Table 2. G e n e r a l c o m b i n i n g abi l i ty o f 4x a n d F D R 2x p a r e n t s f o r
t ra i t s e v a l u a t e d d u r i n g t w o y e a r s in R h i n e l a n d e r
P a r e n t V i g o r U n i f o r m i t y E a r l i n e s s T u b e r
yield
4x f ema le s :
S e r r a n a - 0 .195 0 .003 0 .172 0 .242*
K a t a h d i n - 0 .295 0 .153 - 0 .028 0 . 0 1 7
A t l a n t i c 0 .497* 0 .116 - 0 .395 0 .289*
LT-1 - 0 .345 - 0 .258 - 0 .020 - 0 .279*
A V R D C - 1 2 8 7 . 1 9 - 0 .435* - 0 .072 0 .292 - 0 . 0 8 6
H P - 2 7 8 . 2 2 - 0 .095 0 .055 - 0 .020 - 0 . 1 8 5 "
S .E . (gi) 0 .208 0 .193 0 . 2 0 0 0 .083
S .E . ( g l - g ' i) 0 .294 0 .272 0 . 2 8 2 0 . 1 1 7
F D R 2x males :
84 .36 .29 - 0 .024 0 .190 0 .470* 0 . 0 9 2
84 .194 .30 0 .100 0 .206 0 . 2 6 0 - 0 .042
85 .37 .38 0 .344 - 0 . 3 3 6 " 0 . 6 1 8 " - 0 . 0 5 8
M I - 4 9 . 1 0 - 0 .268 0 .060 - 0 . 1 1 3 - 0 .008
S .E . (gi) 0 .190 0 .176 0 . 1 8 2 0 . 0 7 6
S .E . (gi-- g'i) 0 . 2 6 9 0 .249 0 .258 0 A 0 7
* ind ica t e s t h a t va lue s a r e s ign i f i can t ly d i f f e r e n t f r o m ze ro a t
t he 0 .05 level .
days indicated that there were only significant dif- ferences between female parents for plant vigor and total tuber yield. Males were only significantly different for earliness. The interaction male × fe- male was not significant for the traits evaluated. The female × year and male × year interactions were significant for plant vigor and total yield, while the year × male interaction was significant only for earliness.
The best female 4x parents for total yield were Serrana and Atlantic, whose GCA estimates were significantly different from zero (Table 2). The US cultivars were significantly earlier than their re- spective progenies. The progenies also had higher tuber set with smaller tubers than their respective 4x US parents. However, there was an improve- ment in the earliness of the progenies when adapt- ed: US females were used as parents with CIP males (data not shown).
Significant phenotypic correlation was found be- tween earliness/tuber set (0.639"*). Further, prog- eny with low yield had many small sized tubers (data not shown). These findings indicated that the
191
early maturity types had few but larger sized tu- bers.
Short days
Significant differences between the 15 families evaluated under short days were found for all the traits evaluated. There were highly significant dif- ferences between females, based on progeny test- ing, for traits such as general tuber appearance (GTA), specific gravity, average tuber weight, to- tal and marketable tuber yield. The interaction male x female was significant for tuber set, specif- ic gravity, total and marketable tuber yield. The genotype (family) x environment (location) inter- action was significant for all but one (specific grav- ity) trait evaluated.
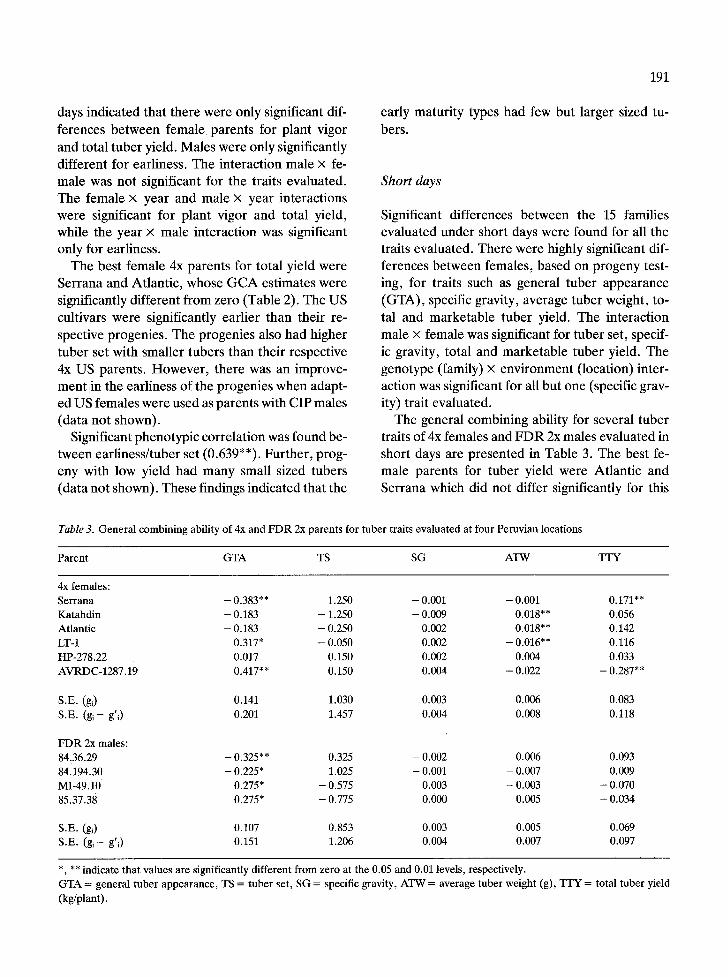
The general combining ability for several tuber traits of 4x females and FDR 2x males evaluated in short days are presented in Table 3. The best fe- male parents for tuber yield were Atlantic and Serrana which did not differ significantly for this
Table 3. G e n e r a l c o m b i n i n g abi l i ty of 4x and F D R 2x pa ren t s for t u b e r t ra i ts e v a l u a t e d at four P e r u v i a n loca t ions
P a r e n t G T A TS SG A T W T r Y
4x females :
S e r r a n a - 0.383** 1.250 - 0.001 - 0.001 0.171"*
K a t a h d i n - 0.183 - 1.250 - 0.009 0.018"* 0.056
A t l a n t i c - 0.183 - 0.250 0.002 0.018"* 0.142
LT-1 0.317" - 0.050 0.002 - 0.016"* 0.116
HP-278.22 0.017 0.150 0.002 0.004 0.033
A V R D C - 1 2 8 7 . 1 9 0.417"* 0.150 0.004 - 0.022 - 0.287"*
S .E. (gi) 0.141 1.030 0.003 0.006 0.083
S .E. ( g i - g' i) 0.201 1.457 0.004 0.008 0.118
F D R 2x males :
84.36.29 - 0.325** 0.325 - 0.002 0.006 0.093
84.194.30 - 0.225* 1.025 - 0.001 - 0.007 0.009
MI -49 .10 0.275* - 0.575 0.003 - 0.003 - 0.070
85.37.38 0.275* - 0.775 0.000 0.005 - 0.034
S .E. (gi) 0.107 0.853 0.003 0.005 0.069
S .E. ( g i - g'i) 0.151 1.206 0.004 0.007 0.097
*, ** ind ica te t h a t va lues a re s ign i f ican t ly d i f fe ren t f rom ze ro at the 0.05 and 0.01 levels , respec t ive ly .
G T A = gene ra l t u b e r a p p e a r a n c e , TS = t u b e r set , S G = specif ic g rav i ty , A T W = ave rage t ube r we igh t (g), T T Y = to ta l t u b e r y ie ld
(kg /p lan t ) .

192
trait. Atlantic produced fewer but larger tubers than Serrana, which resulted in a higher market-
able tuber yield in its progenies than in Serrana progenies. There were no significant differences between 2x parents for all but one (general tuber appearance) trait measured.
The family performance for tuber yield, specific
gravity and stability parameters for yield of the 15 families evaluated under short days is given in Ta-
ble 4. The 4x × 2x family Serrana × 85.37.38 com- bined acceptable tuber yield, yield stability and
good specific gravity. Moreover, it produced tu-
bers with adequate size (data not shown).
The Spearman's rank correlation coefficients for yield across locations, among male and female par- ents were not significant.
Discussion
The low tuber yield of 4x x 2x progenies under
long days was mainly the consequence of lack of adaptation of this germplasm to long days. More-
over, late vine maturity of these progenies evaluat- ed in Rhinelander indicates that proper selection of parents for early maturity type and adaptation to day length should be done in the 2x population. Thus, an improvement in both earliness and tuber
yield in the 4x × 2x progeny should be expected through this selection technique.
Stability and allelic diversity
Stability analysis results indicate that 11 of 15 fam-
ilies were stable, but 6 of these were inconsistent in yield over environments. The four remaining fam-
ilies were unstable and low yielding. This high fre-
quency of stable families could result from the fact that F D R 2x parents transmitted a large fraction of
their heterozygosity and epistasis to their 4x proge-
ny. In this regard, Lerner (1954) found that hetero-
Table 4. Tuber yield (TTY, kg/hill), yield stability parameters and specific gravity (SG) of 4x families evaluated at four Peruvian locations
4x x 2x family TTY Stability parameters SG
b 1 sb 2 sd 3 r2, 4 HP278.22 × 84.36.29 0.987 1.59 0.49* 0.108" 0.84** 1.071 Serrana x 84.36.29 0.977 0.90 0.49* 0.104" 0.63* 1.071 Katahdin × 84.194.30 0.864 1.38 0.31' 0.042* 0.91"* 1.066 Serrana × 85.37.38 0.835 0.89 0.20 0.018 0.91"* 1.079 Atlantic x 81.194.30 0.822 1.17 0.30* 0.039* 0.88** 1.083 Serrana × 84.194.30 0.745 1.25 0.38* 0.062* 0.85** 1.077 Katahdin × MI-49.10 0.694 1.18 0.12 0.006 0.97** 1.068 LT-1 x 85.37.38 0.619 1.10 0.60* 0.156" 0.63* 1.070 HP278.22 × 85.37.38 0.566 0.87 0.17 0.013 0.93** 1.085 HP278.22 x MI-49.10 0.546 1.06 0.06 0.002 0.99** 1.078 Katahdin × 85.37.38 0.525 0.92 0.16 0.001 0.94** 1.065 LT-1 × 84.36.29 0.513 0.64* 0.45* 0.090* 0.49 1.074 AVRDC x MI-49.10 0.467 0.35* 0.33* 0.046* 0.36 1.93 AVRDC x 84.36.29 0.382 0.11" 0.14 0.008 0.23 1.086 AVRDC × 84.194.30 0.370 0.11" 0.20 0.018 0.12 1.069
LSD0.05 0.258 0.017
*, ** indicates significance at the 5 and 1 percent levels, respectively. 1 b, regression coefficient. 2sb, standard error for b. 3 sd, deviation mean squares from regression. 41.2, coefficient of determination.

zygous genotypes were more resistant to the unsta- bilizing effect of climatic variation than homozy- gous genotypes. Further, results reported in maize by Adams & Shank (1959) demonstrated that an increase in levels of heterozygosity in groups of lines and hybrids resulted in higher levels of ho- meostasis in these groups. Recently Darmo & Pe- loquin (1990) found that three 4x clones, from 4x - 2x crosses with good processing traits, were more stable and not significantly different for yield than Atlantic. This is very important since tuber yield and yield stability must be achieved for the success of a potential genotype as a cultivar.
Specific gravity and tuber yield
A potato cultivar should have high specific gravity for processing, since it is highly correlated with the dry matter content of the raw product. High specif- ic gravity is required by the potato chip industry, since it results in an increase in chip yield and reduced production costs. Stevenson et al. (1954) indicated that specific gravity is influenced by dif- ferent environmental factorss. For example, i n these results highly significant differences were found among the four short day locations. How- ever, as was concluded by Killick & Simmonds (1974), specific gravity is insensitive to genotype by environment interactions. Similar results were ob- tained in this experiment, in which the family x locations interaction for specific gravity was not significant. This means that the relative ranking for specific gravity of the 15 families was similar across the four locations.
High estimates of heritability for specific gravity have been reported either in the progeny of 2x x 2x crosses (Landeo, 1979; Ruttencutter et al., 1979) or that of 4x x 4x crosses (Peterson & Plaisted, 1960; Sanford, 1960; Tai, 1976). However, accord- ing to Tai! (1976) and Killick (1977) SCA effects for specific gravity are also important in 4x progenies. Moreover, Masson (1985) estimated heritability values o f 58% and 72% for two sets of 4x x 2x crosses. Despite this estimation he indicated that SCA was also significant for specific gravity. Buso (1986) also found heterosis for: specific gravity in 4x
193
families from 4x x 2x crosses (12% higher than the 4x parents). The significant male x female interac- tion explains the previous findings, indicating that nonadditive genetic effects are also important for specific gravity. Therefore, the performance of FDR 2x clones in transmitting high specific gravity to their 4x offspring was expected.
Adaptation and the choice of parents
Significant negative regressions were found for tu- ber set (p = 0.002) and earliness, (p < 0.001) when the percentage of adapted germplasm was used as an independent variable in regression analysis for the traits studied in the long day environments (dependent variables). The results indicate that unadapted germplasm produced many small tu- bers. Likewise, an increase in percentage of adapt- ed Tuberosum background will result in early ma- turing types, which would be an indication that the lack of adaptation might be related to late maturity and initiation of tuberization.
The best 4x parent based on progeny testing under long day environments was the cultivar At- lantic (Table 2), indicating that a cultivar or ad- vanced selection adapted to the target environment should be used was the female in 4x x 2x crosses. In this regard, Plaisted (1985) mentioned that es- sentially all US and Canadian cultivars are the re- sult of pedigree type crossing programs. He point- ed out that the choice of parents can be measured by the frequency that named varieties enter into pedigrees, e.g. of 135 varieties released during 1950-1980, 12 were the product of crossing two varieties, and 73 had a variety as one of the parents. Only 50 were the result of intermating two breed- ing lines.
Mendoza & Haynes (1976) indicated that Tuber- osum varieties (selected under long days) yield more under long than under short days. However, the performance of the US varieties as parents, in crosses with FDR 2x parents selected under short days, suggests that they could be used along with those FDR 2x parents to produced high yielding progenies under short days, e.g. Katahdinx 84.194.30 or Atlantic x 84.194.30. The high yield

194
of these families could be the result of both early maturity, provided by the Tuberosum parent, and improved tuberization provided by the adapted FDR 2x parent.
The lack of correlation between the rank order of the parental performance for tuber yield of the male and female parents across the environments used (short and long days) clearly indicates that the selection of parents must be done in each location in which the final product (either a family or a clone) will be used. The parent must have high general combining ability for its use as a good cross- ing partner. However, since yield also depends on nonadditive gene action, the search for parents which in crosses have good specific combing ability is also important to maximize heterosis for tuber yield. The best 4x candidates seems to be the highly adapted cultivars. However, the lack of correlation between FDR 2x clones and their 4x progeny for tuber yield (Schroeder, 1983; Buso, 1986) clearly reinforces the need for progeny testing of potential FDR parents before their utilization as regular par- ents in a breeding program. Moreover, the FDR 2x parents must also be adapted to the target area in which its 4x progeny will be used.
Acknowledgments
The authors thanks to Mr. Don Kichefski and Mr. Simon Perez who took care of the plant material in USA and Peru, respectively. Paper No. 3211 from the Laboratory of Genetics. Research supported by the College of Agricultural and Life Sciences; International Potato Center; USDA-CRGO-88- 37234 3619, and Frito-Lay, Inc.
References
Adams, M.W. & D.B. Shank, 1959. Relation of heterozygosity to homeostasis in maize hybrids. Genetics 44: 777-785.
Allard, R.W. 1966. Relationship between genetic diversity and consistency of performance in different environments. Crop Sci. 6: 127-133.
Amoros, W.A. & H.A. Mendoza, 1979. Relationships between heterozygosity and yield in autotetraploid potatoes. Amer. Potato J. 45:455 (abstract).
Buso, J.A. 1986. Evaluation of families and clones from the 4x × 2x breeding scheme in potato. PhD Thesis, University of Wisconsin-Madison. 211 pp.
Concilio, L. 1985. Evaluation of yield and other agronomic characteristics of TPS families and advanced clones from different breeding schemes. MS Thesis, University of Wis- consin-Madison, 63 pp.
Darmo, E. & S.J. Peloquin, 1990. Performance and stability of nine 4x clones from 4 x - 2x crosses and four commercial cultivars. Potato Research 33: 357-364.
De Jong, H., G.C.C. Tai, W.A. Russell, G.R. Johnston & K.G. Proudfoot, 1981. Yield potential and genotype-environment interactions of tetraploid-diploid ( 4 x - 2x) potato hybrids. Amer. Potato Journal 58: 191-199.
Douches, D.S. & C.F. Quiros, 1987. Use of 4x - 2x crosses to determine gene-centromere map distances of isozyme loci in Solanum species. Genome 29: 519-527.
Eberhart, S.A. & W.A. Russell, 1966. Stability parameters for comparing varieties. Crop Sci. 6: 36--40.
Huaman, Z. & R.W. Ross, 1985. Updated listing of potato species names, abbreviations and taxonomic status. Amer. Potato J. 62: 629-641.
Iwanaga, M., P. Jatala, R. Ortiz & E. Guevara, 1989. Use of FDR 2n pollen to transfer resistance to Root-Knot nematodes into cultivated 4x potatoes. J. Amer. Soc. Hort. Sci. 114: 1008-1013.
Killick, R.J. & N.W. Simmonds, 1974. Specific gravity in di- haploid potato tubers as a character showing small-environ- mental interactions. Heredity 32: 109-112.
Killick, R.J. 1977. Genetic analysis of several traits in potatoes by means of a diallel cross. Ann. Appl. Biol. 86: 279-289.
Landeo, J.A. 1979. Breeding potential of Group Andigena haploid potatoes. PhD Thesis, University of Wisconsin-Madi- son. 124 pp.
Lerner, I.M. 1954. Genetic homeostasis. Oliver and Boyd, Edinburgh. 134 pp.
Masson, M.F. 1985. Mapping, combining abilities, heritabilities and heterosis with 4x x 2x crosses in potatoes. PhD Thesis, University of Wisconsin-Madison. 119 pp.
MSTATC. 1989. A microcomputer program for the design, management and analysis of agronomic research experi- ments. Michigan State University.
Mendoza, H.A. & F.L. Haynes, 1976. Variability for photope- riod reaction among diploid and tetraploid potato clones from three taxonomic groups. Amer. Potato J. 53: 319-332.
Mendoza, H.A. & F.L. Haynes, 1977. Inheritance of tuber initiation in tuber-bearing Solanum as influenced by photope- riod. Amer. Potato J. 54: 243-252.
Peloquin, S.J., G.L. Yerk, J.E. Werner & E. Darmo, 1989. Potato Breeding with haploids and 2n gametes. Genome 31: 1000-1004.
Peterson, L.C. & R.L. Plaisted, 1960. Recurrent selection for high specific gravity. In: 30th annual report for cooperators. The National Potato Breeding Program. USA. p. 154-156.
Plaisted, R.L. 1985. Population breeding applied to improve- ment of unadapted Solanurn cultivated species. In: Report of

CIP Planning Conference on Present and Future Strategies for Potato Breeding. International Potato Center, Lima-Pe- ru. December 12-14, 1983. p. 143-162.
Ross, H. 1986. Potato Breeding-problems and perspectives. Verlag Paul Parey. 132 pp.
Ruttencutter, G., F.L. Haynes & R.H. Moll, 1979. Estimation of narrow-sense heritability for specific gravity in diploid potatoes (Solanum tuberosum spp. phureja and tuberosum). Amer. Potato J. 56: 447-453.
Sanford, L.L. 1960. Comparative evaluation of clones as testers for yield, specific gravity and tuber appearance in potato. PhD thesis, Iowa State University, Ames, Iowa.
195
Schroeder, S.H. 1983. Parental value of 2x, 2n pollen clones and 4x cultivars in 4x x 2x crosses in potatoes. PhD Thesis, Uni- versity of Wisconsin-Madison. 192 pp.
Stevenson, P.J., R.V. Akeley & J.G. McLean, 1954. Potato utilization in relation to variety (heredity) and environment. Amer. Potato J. 38: 105-113.
Stofella, P.J., H.H. Bryan, T.H. Howe, J.W. Scott, S.F. Lo- cascio & S.M. Olson, 1984. Stability differences among fresh market tomato genotypes. 1. Fruit yields. J. Amer. Soc. Hort. Sci. 109: 615-618.
Tai, G.C.C. 1976. Estimation of general and specific combining ability in potatoes. Can J. Genet. Cytol. 18: 463-440.