activity - bmjmp.bmj.com/content/molpath/49/6/m331.full.pdf ·...
TRANSCRIPT

J Clin Pathol:Mol Pathol 1996;49:M331-M339
Papers
Inhibitors of collagenase but not of gelatinasereduce cartilage explant proteoglycan breakdowndespite only low levels of matrix metalloproteinaseactivity
Carol J Brown, Shamim Rahman, Allison C Morton, Claire L Beauchamp,Helena Bramwell, David J Buttle
Aims-To investigate the level of matrixmetalloproteinase activity during thetime-course of cartilage explant prote-oglycan breakdown; to determine theeffects of selective small-molecule inhibi-tors of matrix metalloproteinases on pro-teoglycan degradation.Methods-The levels of matrix metallo-proteinase activity in cartilage explantcultures and conditioned media weremonitored by use of a quenched fluores-cent substrate. The constants for inhibi-tion of certain matrix metalloproteinasesby a series of synthetic inhibitors weredetermined. Bovine and human cartilageexplant cultures were treated withinterleukin-1, tumour necrosis factor orretinoic acid and the amount ofproteogly-can released into the culture medium inthe absence and presence of the inhibitorswas quantified. Control experiments, ex-amining the inhibition of other protein-ases, and investigating possible toxic ornon-specific effects of the inhibitors, werecarried out.Results-The profile of inhibition of pro-teoglycan release suggested the involve-ment of interstitial collagenase-like,rather than gelatinase- or possiblystromelysin-like, proteinases. No evidencewas found for toxic or non-specificmechanisms of inhibition. Very low levelsof activity of the known matrix metallo-proteinases were present during the time-course of aggrecan breakdown.Conclusions-A novel collagenase-likeproteinase(s) may be involved in cartilageproteoglycan breakdown. Gelatinase-typematrix metalloproteinases do not seem tobe involved in this process. Specific colla-genase inhibitors may be therapeuticallyefficacious in the treatment of arthritis.( Clin Pathol: Mol Pathol 1996;49:M33 1-M339)
Keywords: cartilage, proteoglycan breakdown, pro-teases.
Aggregating proteoglycan (aggrecan) and type IIcollagen are the major extracellular protein com-ponents of cartilage. Loss of proteoglycan and itsassociated water can compromise the structureand function ofcartilage by reducing its resilienceto compressive loading and increasing frictionduring joint motion.' 2Thus, depletion of articu-lar cartilage proteoglycan (seen on x ray film asjoint space narrowing) may predispose the carti-lage to further mechanical damage. Preventingpathological cartilage proteoglycan breakdownmay therefore substantially protect the affectedjoint and help prevent the development of anarthritic lesion. The mechanism by which carti-lage proteoglycan is degraded is not fully under-stood, although recent evidence would suggestthe involvement of a number of proteolyticenzymes in a cascade reaction that may result inthe activation of an as yet undiscovered enzyme,"aggrecanase" (reviewed elsewhere3).There is evidence that members of the
matrix metalloproteinase (MMP), or matrixin,family of enzymes are involved in cartilage pro-teoglycan breakdown. The MMPs, like almostall metalloproteinases that have been charac-terised so far, utilise a zinc atom in the activesite.4 Based on their substrate specificity theMMPs can be divided into three groups: thecollagenase group, including interstitial colla-genase (EC 3.4.24.7); neutrophil collagenase(EC 3.4.24.34) and collagenase-3'; the strome-lysin group, which includes stromelysin 1 (EC3.4.24.17), stromelysin 2 (EC 3.4.24.22) andmatrilysin (EC 3.4.24.23); and the gelatinasegroup consisting of gelatinases A (EC3.4.24.24) and B (EC 3.4.24.35).6 Stromelysin3 actually represents a distinct group of theMMPs.4 In in vitro experiments matrilysinand stromelysin 1 have been shown to be themost efficient members of this subfamily atcatalysing aggrecan hydrolysis.8
Increased MMP concentrations have beenfound in both osteoarthritis' and rheumatoidarthritis."0 Amounts of the MMPs are raised bythe action of the proinflammatory cytokines,interleukin-1 (IL-1) and tumour necrosis
Institute for Bone andJoint Medicine,Department ofHumanMetabolism andClinical Biochemistry,University of SheffieldMedical School,Beech Hill Road,Sheffield SlO 2RXC J BrownS RahmanA C MortonH BramwellD J Buttle
British BiotechPharmaceuticals Ltd.,Watlington Road,Cowley,Oxford OX4 5LYC L Beauchamp
Correspondence to:Dr D J Buttle.email:[email protected]
Accepted for publication12 September 1996
M331
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from

Brown, Rahman, Morton, Beauchamp, Bramwell, Buttle
factor (TNF), that mediate catabolic responsesin cartilage."'l4The ability of synthetic inhibitors of the
MMPs to suppress cartilage proteoglycan deg-radation in model culture systems is welldocumented,'51" and it has been proposed thatinhibitors of these enzymes might be used fortherapeutic intervention in the arthritides." Todate, all of the inhibitors found to have a chon-droprotective effect are broad spectrum inhibi-tors that do not discriminate between the vari-ous MMPs. For instance, the most potentexample to date, the hydroxamate BB94, is atight binding inhibitor of all the MMPs investi-gated, with Kj(app) and IC50 values in the lownanomolar range and varying by only about afactor of 10 between the different MMPs.'520Such inhibitors therefore provide us with littleinformation about which members of this largegroup of enzymes may be involved in cartilageproteoglycan breakdown.We have now attempted to address the role
ofMMPs in cartilage proteoglycan breakdownby the use of a series of zinc chelating MMPinhibitors that demonstrate some discrimina-tion for the inhibition of various MMPs, andalso by the measurement of levels of MMPactivity present in cartilage explant culturesduring the time-course of aggrecan depletion.
MethodsHuman interstitial procollagenase, promatri-lysin and prostromelysin 1 were recombinantproteins purified from media conditioned byCHO cells transfected with cDNA coding forthe respective enzymes. Progelatinases A and Bwere isolated from media harvested from themalignant melanoma cell line RPMI 7591, andthe monocytic leukaemia cell line THP-1,respectively. The MMP inhibitors used in thisstudy, BB87, BB250, BB2001, BB2014 andBB3003*, were synthesised at British BiotechPharmaceuticals Ltd., Oxford, UK. Humancathepsin B (EC 3.4.22.1) was purified fromliver by affinity chromatography21 and humancathepsin D (EC 3.4.23.5) from liver was pre-pared by the published method.22 Peptidyldipeptidase A (angiotensin converting enzyme)(EC 3.4.15.1) was purified from rabbit lung.23Neprilysin (enkephalinase) (EC 3.4.24.11) wasfrom rabbit kidney, and was obtained in themembrane bound form essentially asdescribed.24 Recombinant human IL-i ox (rhIL-lc) (specific activity 5 x 107 U/mg) was a giftfrom Dr K Ray, Glaxo Group Research Ltd,Greenford Road, Greenford, Middlesex, UK.Recombinant human TNFo (rhTNFcx) (spe-cific activity 8 x 107 U/mg) was kindly providedby Dr E Amento, Genentech Inc. Boulder,Colorado, USA.'The recombinant human IL-i-receptor an-
tagonist protein was a gift from Synergen Inc.,*The structures of the MMP inhibitors are: BB87, N4-hydroxy-2R-isobutyl-N1- {2-phenyl-1S- [(pyrid-2-ylmethyl) -carbamoyl]-ethyl}-succinamide; BB250, N4-hydroxy - 2R - isobutyl - N1 - (1S- methyl -carbamoyl - 2 - phenyl-ethyl)-3S-thiophene-2- sulfonyl-methyl) succinamide; BB2001, 3R{1S(3 morpholin-4yl-propylcarbamoyl)-2-phenyl-ethyl-carbamoyl} - 6 - phenyl-hex-anoic acid; BB2014, 3R-(lS-methylcarbamoyl-2-phenyl -
ethylcarbamoyl) - 6 - phenylhexanoic acid; BB3003, 3R-(lS-methylcarbamoyl-2-phenyl-ethylcarbamoyl)-nonadecanoic acid,dicyclohexylamine salt.
Boulder, Colorado, USA. All-trans-retinoicacid (Ret), gentamicin, cortisol, chondroitinsulfate A, aminophenylmercuric acetate(APMA) and dimethyl sulfoxide were pur-chased from Sigma Chemical Co., Poole, Dor-set, UK. Dulbecco's modification of Eagle'smedium (DMEM), minimal essential medium(MEM), newborn-calf serum (NCS), fetal calfserum (FCS), amphotericin B, penicillin, andstreptomycin were purchased from Gibco LifeTechnologies Ltd, Paisley, UK. 1,9-Dimethylmethylene blue was from Aldrich,Gillingham, Dorset, UK. Cetylpyridiniumchloride, Triton X-100 and OCT embeddingmedium were from BDH Chemicals Ltd,Poole, Dorset, UK. 7-Methoxycoumarin-4-ylacetyl (Mca)-Pro-Leu-Gly-Leu-(3-[2,4-dini-trophenyl] - L - 2, 3 - diaminopropionyl)(Dpa)-Ala-Arg-NH2 was provided by Dr CGraham Knight, Strangeways Research Labo-ratory, Cambridge, UK. Human urokinase-type plasminogen activator (uPa) (EC3.4.21.73) was obtained from Calbiochem-Novabiochem Ltd, Nottingham, UK. Benzy-loxycarbonyl (Z)-Gly-Gly-Arg-aminomethyl-coumarylamide (NHMec) and Z-Arg-Arg-NHMec were the products of BachemFeinchemikalien AG, Bubendorf, Switzerland.35SO4 and 35S-methionine were products ofAmersham International plc, Little Chalfont,Buckinghamshire, UK. All buffers and saltswere of analytical grade.
PROTEINASE ASSAYSThe effects ofone oftheMMP inhibitors, BB250(10 FM), on the activities ofuPa, cathepsin B andcathepsin D, which have been implicated incartilage proteoglycan breakdown,3 25 weretested. In addition, we examined the capacity ofBB250 to inhibit two metallopeptidases that arenot MMPs- that is, peptidyl dipeptidase A andneprilysin.uPa activity was measured by continuous
rate assays in 50 mM Tris HCl, 0.1% TritonX-100, pH 7.5 at 37°C, with Z-Gly-Gly-Arg-NHMec as substrate (100 jM). The rate ofsubstrate hydrolysis was recorded before andafter the addition of the inhibitor to thecuvette. The effect of BB250 on human cathe-psin B activity was determined with Z-Arg-Arg-NHMec (5 jM) as substrate in 100 mMNa2H/KH2PO4, pH 6.0, containing 1 mMNa2EDTA and 2 mM dithiothreitol. Theactivity of cathepsin D in the presence andabsence ofBB250 was measured at 45°C using8%(w/v) haemoglobin as substrate in 0.1 Mformate buffer, pH 3.3. Following precipitationof undigested protein in 3% (w/v) trichloroace-tic acid and filtration the change in A280, ascompared with a blank without enzyme, wasdetermined. Peptidyl dipeptidase A activity wasassayed using N-benzoylglycine-His-Leu, asdescribed previously.23 Neprilysin was assayedusing the quenched fluorescent substratedansyl-D-Ala-Gly-Phe(pNO2)-Gly."2MMP activity in bovine nasal cartilage explant
cultures and conditioned media was assayed byuse of the quenched fluorescent substrateMca-Pro-Leu-Gly-Leu-Dpa-Ala-Arg-NH2. Thissubstrate is cleaved efficiently by all ofthe MMPs
M332
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from

Inhibitors of collagenase but not ofgelatinase reduce cartilage explant proteoglycan breakdown
tested so far, including interstitial collagenase,stromelysin 1, matrilysin, gelatinase A,26 gelati-nase B,'5 neutrophil collagenase, andcollagenase-3,27 and can therefore be used toassess the combined levels of activity of theseenzymes. The MMP activity in conditionedmedia from explants cultured with or withoutrhIL-lx (0.3 nM), rhTNFa (3.5 nM) or Ret(1 p1M) for up to five days, with and withoutAPMA (0.1 rmM; one hour) activation of proM-MPs, was assessed by continuous-rate assays at37°C. The buffer used was 0.1 M Tris HCl, pH7.5, containing 0.1 M NaCl, 10 mM CaCl2,0.2% Triton X-100, and Mca-Pro-Leu-Gly-Leu-Dpa-Ala-Arg-NH2 (5 giM) (ex. 328 nm, em. 393nm). The fluorimeter was standardised with aknown concentration of Mca-Pro-Leu.
In order to obviate potential problems offluorescence quenching, the substrate (10 gM)was added to explant cultures (two explants ofapproximate dimensions 2 x 2 x 2 mm/culture)in 0.4 ml phosphate buffered isotonic saline(PBS), pH 7.2, containing 4.5 mg/ml D-glu-cose and antibiotics (see later), for periods ofup to 24 hours. We have previously found thatcartilage explants can be successfully culturedfor at least two days under these conditions,during which time they behave in a similarmanner to explants cultured in DMEM, andresorb the proteoglycan component of theextracellular matrix upon treatment with cata-bolic stimuli. BB94'5 (10 jM) was also addedto some cultures; 0.3 ml of each culture wasdiluted with an equal part of Tris buffer(earlier), and the quantity of product formedwas determined by comparing the amount offluorescence with that of a known concentra-tion of standard.
DETERMINATION OF KINETIC PARAMETERS
Gelatinase A was activated overnight at 40C inlmM APMA. Gelatinase B was activated bythe same means at ambient temperature fortwo hours. Interstitial collagenase was activatedwith 0.66 mM APMA for 16 hours at 37°C,prior to purification. Purified stromelysin 1 wasactivated with trypsin (30 minutes, 370C), andmatrilysin was autoactivated by freeze-thawingprior to purification.
For the determination of IC50 values, allassays were performed at 370C for 16 hours in25 mM Hepes, pH 7.5, containing 5 mMCaC12, 0.05% (w/v) Brij 35 and 0.02% NaN3.Collagenase activity was determined in anassay28 utilising '4C-collagen as substrate.29Matrilysin and stromelysin 1 activities weremeasured in assays using '4C-casein as thesubstrate.30 Gelatinase A and B activities wereassayed by a modification of the previouslydescribed procedure,3' with the addition of 50gg/ml gelatin. IC50 values were found by com-paring enzyme activity in the presence of vary-ing concentrations of the inhibitors, and fittinga curve to the data thus obtained. The inhibitorconcentration that gave 50% inhibition wasthen acquired by interpolation.
CARTILAGE CULTURESMost of the work reported here has been doneon normal bovine cartilage, which has been
shown previously to behave catabolically in amanner similar to the human tissue.3 However,a limited number of experiments were doneusing two samples of non-arthritic human car-tilage. One sample was from the femoralcondyle of an 81 year old man with no historyof arthritic disease. The second sample wasfrom the interphalangeal joint of the left big toeof a 14 year old boy who underwent fusion ofthe joint in response to a non-arthritic footdeformity. Cartilage was sliced, to give anexplant size of approximately 3 x 3 x 2 mm.The slices were pre-incubated in MEM con-taining 10% (v/v) FCS, penicillin (100 u/ml)and streptomycin (100 jg/ml) for two days.They were then transferred individually towells of a 96-well plate containing 195 jl MEMcontaining 1% (v/v) FCS to which theinhibitors were added from 100-fold concen-trated stock solutions in methanol. After onehour Ret was added to a final concentration of1 gM. The cultures were maintained for fivedays after which the media and explants werestored frozen pending analysis.
Bovine nasal septum cartilage explants wereprepared and preincubated as describedpreviously.32 The cartilage discs were trans-ferred individually to wells of a 96-well plate,with 195 gl of serum-free DMEM to which themetalloproteinase inhibitors were added from100-fold stock solutions in methanol. A 1%(v/v) concentration of methanol did not affectcartilage catabolism (data not shown). Afterone hour, rhIL-la, rhTNFx, or Ret was addedto final concentrations of 250 U (0.3 nM),5000 U (3.5 nM), or 10-6 M, respectively. Thecultures were maintained for up to five dayswith a medium change after two days, the freshmedium containing exactly the same constitu-ents as the original. Conditioned media anddiscs were stored frozen pending analysis. Instudies examining the effects of the MMPinhibitors on stimulated proteoglycan releasefrom all types of cartilage examined, onlyexperiments in which the addition of thestimulus resulted in at least a doubling of theamount of proteoglycan release were includedin the analyses. This helped to ensure that thedetected inhibition was of stimulated, as well asbasal, release.
Bovine articular cartilage slices were dis-sected from metacarpophalangeal joints andpreincubated as reported previously,'5 afterwhich the explants were treated as describedearlier for the nasal cartilage discs. As shownelsewhere'5 articular cartilage was less respon-sive than nasal cartilage to the presence ofhuman cytokines. The concentrations ofrhIL- 1 ex and rhTNFax were therefore increasedto 1800 U (2 nM) and 10 000 U (7 nM),respectively. The cultures were maintained forfive days with a medium change on day 2, afterwhich the media and cartilage pieces werestored frozen.
DETERMINATION OF PROTEOGLYCANDEGRADATIONThe amount of proteoglycan in the culturemedium was determined as sulfated gly-cosaminoglycan by use of dimethylmethylene
M333
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from
Brown, Rahman, Morton, Beauchamp, Bramwell, Buttle
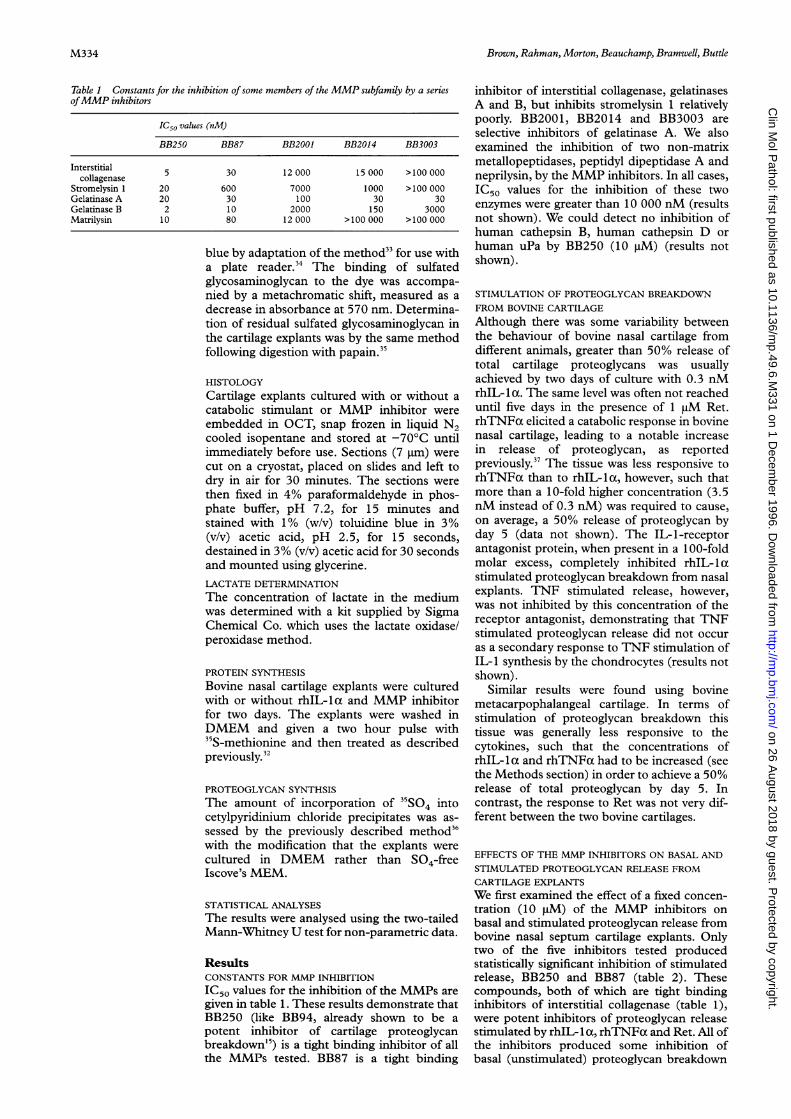
Table 1 Constants for the inhibition ofsome members of the MMP subfamily by a seriesofMMP inhibitors
IC50 values (nM)
BB250 BB87 BB2001 BB2014 BB3003
Interstitial 5 30 12 000 15 000 >100 000collagenase
Stromelysin 1 20 600 7000 1000 >100 000Gelatinase A 20 30 100 30 30Gelatinase B 2 10 2000 150 3000Matrilysin 10 80 12 000 >100 000 >100 000
blue by adaptation of the method33 for use witha plate reader.34 The binding of sulfatedglycosaminoglycan to the dye was accompa-nied by a metachromatic shift, measured as adecrease in absorbance at 570 nm. Determina-tion of residual sulfated glycosaminoglycan inthe cartilage explants was by the same methodfollowing digestion with papain.35
HISTOLOGYCartilage explants cultured with or without a
catabolic stimulant or MMP inhibitor were
embedded in OCT, snap frozen in liquid N2cooled isopentane and stored at -700C untilimmediately before use. Sections (7 im) were
cut on a cryostat, placed on slides and left todry in air for 30 minutes. The sections were
then fixed in 4% paraformaldehyde in phos-phate buffer, pH 7.2, for 15 minutes andstained with 1% (w/v) toluidine blue in 3%(v/v) acetic acid, pH 2.5, for 15 seconds,destained in 3% (v/v) acetic acid for 30 secondsand mounted using glycerine.LACTATE DETERMINATION
The concentration of lactate in the mediumwas determined with a kit supplied by SigmaChemical Co. which uses the lactate oxidase/peroxidase method.
PROTEIN SYNTHESIS
Bovine nasal cartilage explants were culturedwith or without rhIL-lca and MMP inhibitorfor two days. The explants were washed inDMEM and given a two hour pulse with35S-methionine and then treated as describedpreviously.32
PROTEOGLYCAN SYNTHSIS
The amount of incorporation of 31SO4 intocetylpyridinium chloride precipitates was as-sessed by the previously described method36with the modification that the explants were
cultured in DMEM rather than S04-freeIscove's MEM.
STATISTICAL ANALYSES
The results were analysed using the two-tailedMann-Whitney U test for non-parametric data.
ResultsCONSTANTS FOR MMP INHIBITION
IC50 values for the inhibition of the MMPs are
given in table 1. These results demonstrate thatBB250 (like BB94, already shown to be a
potent inhibitor of cartilage proteoglycanbreakdown'5) is a tight binding inhibitor of allthe MMPs tested. BB87 is a tight binding
inhibitor of interstitial collagenase, gelatinasesA and B, but inhibits stromelysin 1 relativelypoorly. BB2001, BB2014 and BB3003 areselective inhibitors of gelatinase A. We alsoexamined the inhibition of two non-matrixmetallopeptidases, peptidyl dipeptidase A andneprilysin, by the MMP inhibitors. In all cases,IC50 values for the inhibition of these twoenzymes were greater than 10 000 nM (resultsnot shown). We could detect no inhibition ofhuman cathepsin B, human cathepsin D orhuman uPa by BB250 (10 gM) (results notshown).
STIMULATION OF PROTEOGLYCAN BREAKDOWNFROM BOVINE CARTILAGEAlthough there was some variability betweenthe behaviour of bovine nasal cartilage fromdifferent animals, greater than 50% release oftotal cartilage proteoglycans was usuallyachieved by two days of culture with 0.3 nMrhIL- 1 a. The same level was often not reacheduntil five days in the presence of 1 jM Ret.rhTNFax elicited a catabolic response in bovinenasal cartilage, leading to a notable increasein release of proteoglycan, as reportedpreviously.37 The tissue was less responsive torhTNFox than to rhIL-la, however, such thatmore than a 10-fold higher concentration (3.5nM instead of 0.3 nM) was required to cause,on average, a 50% release of proteoglycan byday 5 (data not shown). The IL-1-receptorantagonist protein, when present in a 100-foldmolar excess, completely inhibited rhIL- 1 astimulated proteoglycan breakdown from nasalexplants. TNF stimulated release, however,was not inhibited by this concentration of thereceptor antagonist, demonstrating that TNFstimulated proteoglycan release did not occuras a secondary response to TNF stimulation ofIL-I synthesis by the chondrocytes (results notshown).
Similar results were found using bovinemetacarpophalangeal cartilage. In terms ofstimulation of proteoglycan breakdown thistissue was generally less responsive to thecytokines, such that the concentrations ofrhIL- 1 a and rhTNFa had to be increased (seethe Methods section) in order to achieve a 50%release of total proteoglycan by day 5. Incontrast, the response to Ret was not very dif-ferent between the two bovine cartilages.
EFFECTS OF THE MMP INHIBITORS ON BASAL ANDSTIMULATED PROTEOGLYCAN RELEASE FROMCARTILAGE EXPLANTSWe first examined the effect of a fixed concen-tration (10 gM) of the MMP inhibitors onbasal and stimulated proteoglycan release frombovine nasal septum cartilage explants. Onlytwo of the five inhibitors tested producedstatistically significant inhibition of stimulatedrelease, BB250 and BB87 (table 2). Thesecompounds, both of which are tight bindinginhibitors of interstitial collagenase (table 1),were potent inhibitors of proteoglycan releasestimulated by rhIL-lat, rhTNFax and Ret. All ofthe inhibitors produced some inhibition ofbasal (unstimulated) proteoglycan breakdown
M334
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from

Inhibitors of collagenase but not of gelatinase reduce cartilage explant proteoglycan breakdown
Table 2 Percentage inhibition ofproteoglycan release from bovine cartilage explants
Stimulant
Inhibitor (101M) Basal rhIL-lax rhTNFax Ret
NasalBB250 48 (3)t 105 (5)t 28 (7)* 78 (4)tBB87 61 (4)t 108 (2)t 97 (4)t 85 (5)tBB2001 12 (7) 5 (18) -13 (10) -4 (4)BB3003 22 (9) -15 (14) -14 (3) -6 (6)BB2014 32 (4)t -10 (17) -68 (12)* -19 (7)
ArticularBB87 56 (6)t 137 (10)* 146 (6)t 104 (9)*BB250 76 (3)t 110 (13)t 163 (4)t 91 (6)*
In the analyses of stimulated release basal levels of proteoglycan release were subtracted. Theresults are shown as percentage inhibition of proteoglycan release, expressed for purposes of clar-ity as mean (SEM). *p < 0.005; tP < 0.0005 compared with controls. Values of greater than 100%inhibition of stimulated release reflect total inhibition of stimulated release plus some inhibition ofthe basal level.
Table 3 Dose response for the inhibition of rhIL-lcxstimulated proteoglycan release frombovine nasal cartilage explants by BB87 and BB250
Per cent inhibition
Inhibitor concentration (,uM) BB250 BB87
0.001 27 (20)0.01 43 (17)0.1 26 (7)* 7 (6)1 77 (5)t 35 (6)t10 105 (5)t 108 (2)t
Cartilage explants were cultured for two days with or without rhIL-la and varying concentrationsof the MMP inhibitors before the culture media and explants were analysed for sulfatedglycosaminoglycan content. Results are expressed as mean (SEM). *p < 0.005; tp < 0.0005 com-pared with controls.
(table 2), though again BB250 and BB87 werethe most potent. Inhibition of basal release ofproteoglycan by the gelatinase inhibitorBB20 14 was also statistically significant.BB20 14 produced a statistically significantstimulation of proteoglycan release in the pres-ence of rhTNFax, however. Cells responsive toTNF are capable of shedding their TNFreceptors, a process dependent on a proteolyticcleavage.38 The soluble receptors then competewith signalling receptors for the ligand. Ourresult could thus be explained by the inhibitionby BB2014 of a proteinase involved in TNFreceptor shedding, or indeed of an enzyme thatis capable of degrading TNFa. These possibili-ties were not explored further.We next determined whether BB250 and
BB87 were capable of inhibiting proteoglycanbreakdown from bovine metacarpophalangealcartilage. As can be seen in table 2, a 10 jMconcentration of either inhibitor produced vir-tually complete inhibition of rhIL-1 a, rhTNFcxand Ret stimulated release, as well as asubstantial inhibition of basal proteoglycanbreakdown.
In cultures of rhIL-lax stimulated nasalcartilage explants a dose dependent inhibitionof proteoglycan release was seen in thepresence ofboth BB87 and BB250 (table 3). Ata concentration of 1 jM both inhibitorsproduced statistically significant reductions inproteoglycan release from the tissue, thoughBB250 seemed to be the more potent. This wasconfirmed at the lower concentration of 0.1 jiM,where BB250, but not BB87, produced statisti-cally significant inhibition.A small number of experiments were carried
out to investigate the inhibitory effects of BB87
and BB250 on human articular cartilageproteoglycan breakdown. As described in theMethods section, we obtained two samples ofnon-arthritic cartilage. Both of these re-sponded catabolically to the presence of 1 ,uMRet, releasing about 40% of total gly-cosaminoglycan by day 5 in the presence of thevitamin, and about 10% in its absence. BB250and BB87 at a dose of 10 jiM both stronglyinhibited the release of proteoglycan fromhuman cartilage. In one sample, basal levels ofrelease were inhibited by (mean (SEM)) 32%(12%) (p < 0.001) by BB250 and 41% (6%) (p< 0.0001) by BB87. Inhibition of Ret stimu-lated release was even more notable, the twoinhibitors giving 93% (5%) (p < 0.001) and99% (4%) (p < 0.001), respectively. In asecond experiment with cartilage from a differ-ent patient, BB250 at a concentration of 10 jiMproduced 38% (16%) (NS) inhibition of basalrelease and 102% (5%) (p < 0.001) inhibitionof Ret stimulated proteoglycan release.The effects of rhIL-lat and BB87 on bovine
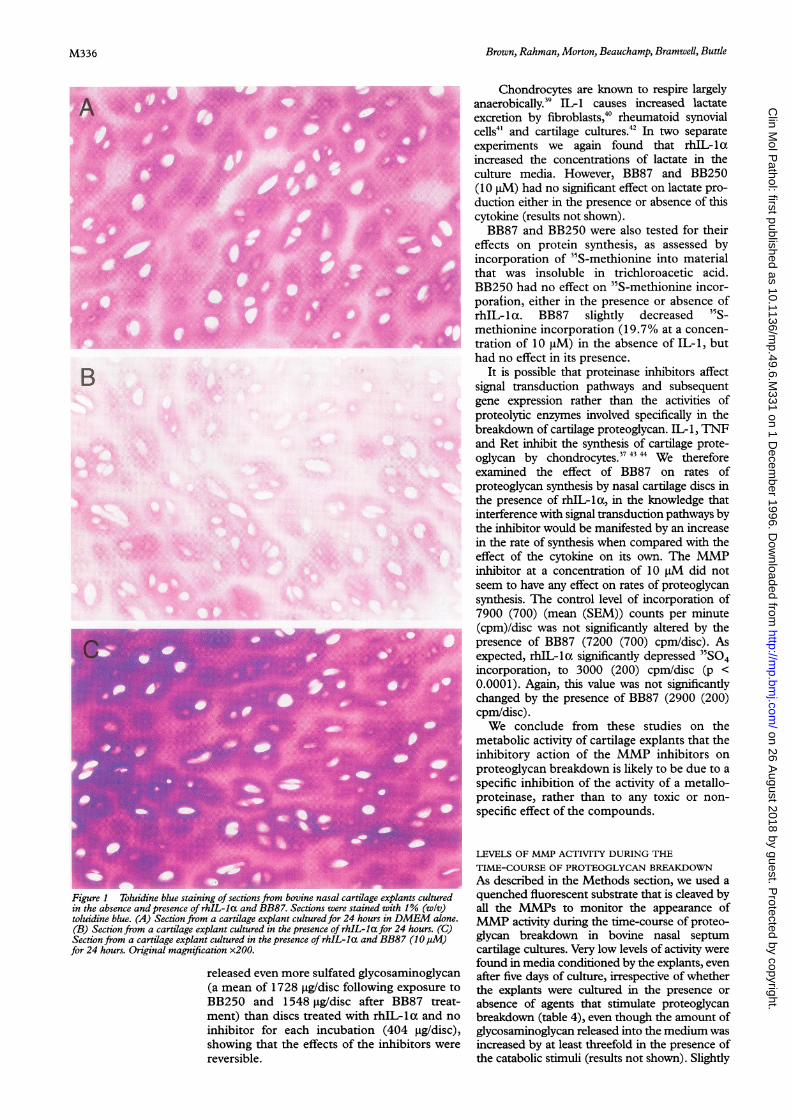
nasal cartilage explants were assessed histologi-cally, by use of toluidine blue, which binds tosulfated glycosaminoglycans. Cartilage cul-tured for 24 hours in the absence of catabolicstimulus retained most of its metachromasiaand hence its proteoglycan content (fig IA),the sections staining darker in the territorialthan in the interterritorial matrix. This was alsothe case with cartilage taken from freshlyslaughtered cattle and processed for histologywithout the culture period (not shown).Explants cultured in the presence of rhIL- 1 xfor 24 hours stained less intensely (fig 1 B). Thepresence of BB250 in rhIL-lx treated 24 hourcultures was chondroprotective, the histologi-cal appearance (fig 1 C) being very similar tothat of the unstimulated controls (fig 1A).
EFFECTS OF BB87 AND BB250 ON THEMETABOLIC ACTIVITY OF NASAL SEPTUM
CARTILAGE EXPLANTS
Proteoglycan release from cartilage explantscan be inhibited non-specifically by toxic com-pounds such as iodoacetate.32 We thereforeevaluated the effects of the inhibitory com-pounds on some metabolic activities ofchondrocytes, and determined whether theinhibitory effect on proteoglycan degradationwas reversible.The ability of nasal cartilage explants to
recover from the effects of BB87 and BB250was investigated. The explants were culturedfor two days with rhIL-1la (0.3 nM) and eitherof the inhibitors (10 gM) after which theexplants were allowed to recover in cytokine-free DMEM for 24 hours. The explants werethen placed in DMEM containing 0.3 nMrhIL-i1 . During the first incubation withinhibitor, BB87 and BB250 inhibited rhIL-1lastimulated release by 78% and 59%, respec-tively. Because of the inhibition ofproteoglycanbreakdown during the first incubation with theinhibitors, these explants retained a largeramount of glycosaminoglycan that could besubsequently lost from the tissue. Indeed, whenthese explants were thereafter stimulated withrhIL- 1 oC in the absence of the inhibitors they
M335
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from
Brown, Rahman, Morton, Beauchamp, Bramwell, Buttle
..:.
* X t :!k
*>:'. ghP. -g
....E,... ;t;<t i-t '...
... ,}.....
.. .......
T'.
''...:i' ':
., !- :B
'' X::.fl$ltn u '' ,:
,-u $:.>,> .:.
slbl s:::
.: .... ...: , ': ' ..... . :.... : .: .s -S.'',,. 1'b',.''. ' , .. :.'.'' . '' v* is:. . 8 ?.'0£ .. .... .. -. . w.: .: ........... .
*; .l's.
B
Figure1 Toluidine blue staining of sections from bovine nasal cartilage explants culturedin the absence and presence of rhIL-la and BB87. Sections were stained with 1% (w/v)toluidine blue. (A) Section from a cartilage explant cultured for 24 hours in DMEM alone.(B) Section from a cartilage explant cultured in the presence of rhIL-la for 24 hours. (C)Section from a cartilage explant cultured in the presence of rhIL-la and BB87 (10jM)for 24 hours. Original magnification x200.
released even more sulfated glycosaminoglycan(a mean of 1728 tggdisc following exposure toBB250 and 1548 ig/disc after BB87 treat-ment) than discs treated with rhIL-1la and no
inhibitor for each incubation (404 ig/disc),showing that the effects of the inhibitors were
reversible.
Chondrocytes are known to respire largelyanaerobically.39 IL-1 causes increased lactateexcretion by fibroblasts,40 rheumatoid synovialcells4" and cartilage cultures.42 In two separateexperiments we again found that rhIL-laincreased the concentrations of lactate in theculture media. However, BB87 and BB250(10 pM) had no significant effect on lactate pro-duction either in the presence or absence of thiscytokine (results not shown).BB87 and BB250 were also tested for their
effects on protein synthesis, as assessed byincorporation of 35S-methionine into materialthat was insoluble in trichloroacetic acid.BB250 had no effect on 35S-methionine incor-poration, either in the presence or absence ofrhIL-lac. BB87 slightly decreased 35S-methionine incorporation (19.7% at a concen-tration of 10 gM) in the absence of IL-1, buthad no effect in its presence.
It is possible that proteinase inhibitors affectsignal transduction pathways and subsequentgene expression rather than the activities ofproteolytic enzymes involved specifically in thebreakdown of cartilage proteoglycan. IL-1, TNFand Ret inhibit the synthesis of cartilage prote-oglycan by chondrocytes.374'34 We thereforeexamined the effect of BB87 on rates ofproteoglycan synthesis by nasal cartilage discs inthe presence of rhIL-ia, in the knowledge thatinterference with signal transduction pathways bythe inhibitor would be manifested by an increasein the rate of synthesis when compared with theeffect of the cytokine on its own. The MMPinhibitor at a concentration of 10 pM did notseem to have any effect on rates of proteoglycansynthesis. The control level of incorporation of7900 (700) (mean (SEM)) counts per minute(cpm)/disc was not significantly altered by thepresence of BB87 (7200 (700) cpmldisc). Asexpected, rhIL-lIx significantly depressed 35SO4incorporation, to 3000 (200) cpmldisc (p <0.0001). Again, this value was not significantlychanged by the presence of BB87 (2900 (200)cpmldisc).We conclude from these studies on the
metabolic activity of cartilage explants that theinhibitory action of the MMP inhibitors onproteoglycan breakdown is likely to be due to aspecific inhibition of the activity of a metallo-proteinase, rather than to any toxic or non-specific effect of the compounds.
LEVELS OF MMP ACTIVITY DURING THE
TIME-COURSE OF PROTEOGLYCAN BREAKDOWN
As described in the Methods section, we used aquenched fluorescent substrate that is cleaved byall the MMPs to monitor the appearance ofMMP activity during the time-course of proteo-glycan breakdown in bovine nasal septumcartilage cultures. Very low levels of activity werefound in media conditioned by the explants, evenafter five days of culture, irrespective of whetherthe explants were cultured in the presence orabsence of agents that stimulate proteoglycanbreakdown (table 4), even though the amount ofglycosaminoglycan released into the medium wasincreased by at least threefold in the presence ofthe catabolic stimuli (results not shown). Slightly
M336
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from

Inhibitors of collagenase but not ofgelatinase reduce cartilage explant proteoglycan breakdown
Table 4 Quenchedfluorescent substrate hydrolysing activity in media conditioned bybovine nasal cartilage explants
Days 0-2 Days 3-5
APMA
Stimulant - + - +
None 0.39 ± 0.12 0.75 ± 0.72 0.48 ± 0.30 2.61 ± 1.50rhIL-la 0.21 ± 0.21 1.14 ± 0.81 - -
rhTNFa 0.57 ± 0.33 1.29 ± 0.78 0.51 ± 0.33 2.04 ± 1.33Ret 0.60 ± 0.21 2.28 ± 1.05 0.36 ± 0.27 2.46 ± 1.74
Explants were cultured in serum-free DMEM with or without a catabolic stimulant for up to fivedays with one medium change on day 2. The conditioned media were assayed for Mca-Pro-Leu-Gly-Leu-Dpa-Ala-Arg-NH2-hydrolysing activity in continuous rate assays as described in Meth-ods. The results are expressed as pmol/min of product per explant.
Table S Quenched fluorescent substrate hydrolysing activity in bovine nasal explantcultures
Time (hours)
Stimulant BB94 1 2 4 6 24
None - 0.78 ± 0.04 0.66 ± 0.05 0.84 ± 0.06 0.78 ± 0.07 0.54 ± 0.03None + 0.66 ± 0.05 0.84 ± 0.08 0.60 ± 0.04 0.66 ± 0.05 0.54 ± 0.03rhIL-la - 0.78 ± 0.07 0.72 ± 0.06 0.84 ± 0.07 0.60 ± 0.04 0.48 ± 0.04rhIL-la + 0.72 ± 0.05 0.54 ± 0.06 0.66 ± 0.05 0.60 ± 0.04 0.48 ± 0.01
Explants were cultured in PBS and glucose, as described in Methods, in the presence of Mca-Pro-Leu-Gly-Leu-Dpa-Ala-Arg-NH2 (10,uM), with or without BB94 (10 p.M). The increase in fluo-rescence, as compared with the amount of fluorescence in unconditioned PBS, was thendetermined. The results are expressed as pmol/min of product per explant.
more activity was present following treatmentwith APMA, suggesting the presence of lowamounts of proMMPs. The low levels of activitygenerated during the time-course of proteo-glycan breakdown (< 0.6 pmol/min substratehydrolysed/explant, without APMA treatment)can be contrasted with the level of over 15 000pmol/min/explant found in week 3 medium fromexplants cultured in the presence of IL- lia, whencollagen breakdown is occurring.45 There is thusa difference in MMP activity of over four ordersof magnitude, between the times of proteoglycanand collagen breakdown.
It is possible that the bulk ofMMP activity inour cultures remained associated with theexplants rather than being secreted into thesurrounding medium. It has been reported thatactive stromelysin 146 and other MMPs bind tocomponents of the extracellular matrix, andindeed the enzymes can be extracted from solidtissues.47 In order to examine this possibility wedetermined the levels of MMP activity in thecultures themselves, as opposed to the activitypresent in isolated conditioned media (earlier).Once again, the levels of active enzyme werevery small, and there were no differences in theamounts of activity detected in cultures withand without rhIL- 1 a, or any increase ofenzyme activity with time (table 5). Mostimportant was the finding that the activity wasnot inhibited by BB94, a broad spectrumMMP inhibitor that also inhibits proteoglycanbreakdown.'5 We conclude from these experi-ments that MMP activity is very low during thefirst few days in cartilage explants cultured inthe presence of agents, such as IL-1 and TNF,that have been reported to promote the expres-sion of these enzymes"-'' as well as proteogly-can breakdown. Instead, we found that levels ofactive MMPs in cartilage explant cultures cor-related with collagen breakdown.45
DiscussionThe results presented here are in agreementwith earlier work demonstrating the efficacy ofMMP inhibitors at limiting the breakdown ofcartilage proteoglycan in experimentalsystems."'-"' Our findings also suggest that atleast one metalloproteinase may be involvedirrespective of the catabolic stimulus to carti-lage breakdown (the cytokines IL-1 and TNF,or the vitamin Ret) in both bovine and humansystems. As far as we are aware, this study is thefirst to investigate the effect ofMMP inhibitorson TNF stimulated breakdown and the first todemonstrate inhibition of proteoglycan releasefrom human cartilage by MMP inhibitors.Our results clearly show the lack of inhibi-
tion of stimulated proteoglycan breakdown bycompounds, BB2001, BB2014 and BB3003,that show a considerable degree of selectivityfor the inhibition of gelatinase activity. It there-fore seems unlikely that gelatinase(s) is in-volved in proteoglycan breakdown stimulatedby IL-1, TNF or Ret in our model systems.We found that inhibitors of collagenase-type
MMPs were effective at inhibiting cartilageproteoglycan breakdown, implicating the activ-ity of a collagenase-like MMP in proteoglycanloss. We must interpret this result with somecaution, because inhibitor concentrations wellin excess of the IC50 values determined in invitro experiments must be used to obtain inhi-bition of proteoglycan breakdown in tissue sys-tems. An example of this phenomenon was ourearlier work with BB94,'5 an inhibitor with lownanomolar inhibition constants with all theMMPs tested so far. Even so, in terms of theinhibition of proteoglycan breakdown, concen-trations of about 0.1 ,uM were required to effect50% inhibition. The reasons for this apparentdiscrepancy are not yet clear, but may berelated to problems of accessibility to the targetenzyme in cartilage, competition between theinhibitor and very high concentrations ofsubstrate within cartilage or, alternatively, maysuggest that the physiologically importantenzyme has yet to be identified. Our findingthat very low levels of activity of the knownMMPs were present in cultures during thetime-course of proteoglycan breakdownstrengthens the latter possibility. It is worthnoting that compounds designed to inhibit theMMPs are not necessarily entirely specific forthis family of enzymes. Some other metallopro-teinases share sufficient structural similarity,particularly around the active site, to begrouped together with the MMPs as themetzincins41 or proteinase clan MB.4 Thus, thestructural determinants that govern interac-tions with small molecule inhibitors may beshared features between these families of met-alloproteinases. An example is the binding withsimilar affinity of a small molecule inhibitor togelatinase A and to atrolysin C (EC 3.4.24.42),a snake venom proteinase of the reprolysinfamily.49 Mammalian representatives of thereprolysin family have been identified.4
Stromelysin 1 has long been considered to bethe most likelyMMP to be involved in cartilagebreakdown, either by a direct mechanism9 orvia activation of other proMMPs, such as
M337
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from

Brown, Rahman, Morton, Beauchamp, Bramwell, Buttle
procollagenase.50 However our data mightargue against a role for stromelysin 1 in thatBB87, a potent inhibitor of cartilage proteogly-can breakdown (table 2), is a 20-fold tighterbinding inhibitor of interstitial collagenasethan it is of stromelysin 1 (table 1). Although arecent independent study5" has also providedevidence against a role for stromelysin inchondrocyte mediated proteoglycan break-down, confirmation will await the results ofexperiments using probes that are moreselective for stromelysins.The loss of aggrecan from human cartilage,
both in various disease states and in tissue cul-ture models, predominantly occurs coincidentwith a specific proteolytic cleavage producedby the action of an as yet unidentified protein-ase referred to as "aggrecanase", at a Glu-Alabond within the interglobular region towardsthe N-terminus of aggrecan. This results in theloss from the matrix of almost the completeaggrecan molecule."5 52-55 The Glu-Ala bond isnot cleaved in vitro by any known MMP otherthan neutrophil collagenase (EC 3.4.24.34),56perhaps suggesting that neutrophil collagenaseis "aggrecanase" (reviewed elsewhere3). Al-though data presented here, and the fact thatneutrophil collagenase has been identifiedrecently in chondrocytes,57 would seem to sup-port this hypothesis, there are observations thatlend credence to the view that the enzymeresponsible for aggrecan cleavage is not neu-trophil collagenase. Neutrophil collagenasecleaves another bond in the interglobularregion of aggrecan, a Asn-Phe bond that is thefavoured cleavage site ofmost MMPs,58-60 moreefficiently than it cleaves the Glu-Ala bond.56Although the product of cleavage of theAsn-Phe bond can be found in humancartilage,6' the available evidence suggests thatthis is only a minor product and that cleavage atthe Glu-Ala bond prevails in the humanarthritic diseases54 and in cartilage culturesystems.55
Similarly, there is evidence that interstitial col-lagenase and collagenase-3 are not responsiblefor aggrecan breakdown. Although interstitialcollagenase is capable ofslowly cleaving aggrecanin the test tube,62 and cleaves the interglobularregion at two distinct sites,59 neither of thesecorrespond to the "aggrecanase" cleavage site.Most importantly we have demonstrated that vir-tually no active collagenase is produced by bovinecartilage explants during the first week ofculture.45 It is also worth noting that the naturalprotein inhibitor of the MMPs, Timpi, does notinhibit cartilage explant proteoglycan release.'6This observation, together with the very low lev-els of MMP activity present in our cultures,virtually precludes a role for any of the knownMMPs in aggrecan degradation. Our results doindicate, however, that a collagenase-like metallo-proteinase may be important in aggrecan break-down, and the previous demonstration of"aggrecanase"-like activity by neutrophilcollagenase56 may suggest that "aggrecanase" is ametalloproteinase with some active site featuressimilar to those of the neutrophil enzyme.The cascade nature of the events leading up
to aggrecan breakdown3 unfortunately do not
allow us to discriminate between the inhibitionof an enzyme acting directly on aggrecan asopposed to an enzyme involved in "proaggreca-nase" activation. The involvement of membersof the gelatinase group of MMPs in the eventsleading to aggrecan breakdown in our modelsystem does now seem unlikely, however.
In any event, the demonstration of potentinhibition of chondrocyte mediated aggrecanbreakdown by inhibitors of collagenase-likeMMPs may provide a lead for the developmentof selective inhibitors as therapeutic agents forthe treatment of arthritic diseases. The sameinhibitors also inhibit type II collagen break-down in cartilage explants45 and discriminatinginhibitors are less likely to produce unwantedside effects than those with a broader inhibitionprofile.
This work was supported by grants from the Arthritis & Rheu-matism Council (UK). We thank Mr D L Douglas, Orthopaed-ics Consultant, Royal Hallamshire and Children's Hospitals,Sheffield, for supplying us with post-operative cartilage samples.
1 Kempson GE, Tuke MA, Dingle JT, Barrett AJ, HorsfieldPH. The effects of proteolytic enzymes on the mechanicalproperties of adult human articular cartilage. Biochim Bio-phys Acta 1976;428:741-60.
2 Kempson GE, Muir H, Swanson SAV, Freeman MAR. Cor-relations between stiffness and the chemical constituents ofcartilage on the human femoral head. Biochim Biophys Acta1970;215:70-7.
3 Buttle DJ, Bramwell H, Hollander AP. Proteolytic mecha-nisms of cartilage breakdown - a target for arthritistherapy? Jf Clin Pathol: Mol Pathol 1 995;48:M 1 67-77.
4 Rawlings ND, Barrett AJ. Evolutionary families of metal-lopeptidases. Methods Enzymol 1995;248:183-228.
5 Freije JMP, Diez-Itza I, Balbin M, Sanchez LM, Blasco R,Tolivia J, et al. Molecular cloning and expression ofcollagenase-3, a novel human matrix metalloproteinaseproduced by breast carcinomas. Jf Biol Chem 1994;269:16766-73.
6 Murphy G, Reynolds JJ. Extracellular matrix degradation.In: Royce P, Steinmann B, eds. Connective tissue and its her-itable disorders. New York: Wiley-Liss Inc, 1993:287-316.
7 Murphy GJP, Murphy G, Reynolds JJ. The origin of matrixmetalloproteinases and their familial relationships. FEBSLett 1991 ;289:4-7.
8 Murphy G, Cockett MI, Ward RV, Docherty AJP. Matrixmetalloproteinase degradation of elastin, type IV collagenand proteoglycan. A quantitative comparison of the activi-ties of 95 kDa and 72 kDa gelatinases, stromelysins-1 and-2 and punctuated metalloproteinase (PUMP). Biochem J1991 ;277:277-9.
9 Dean DD, Martel-Pelletier J, Pelletier J-P, Howell DS,Woessner JF. Evidence for metalloproteinase and metallo-proteinase inhibitor imbalance in human osteoarthriticcartilage. J Clin Invest 1989;84:678-85.
10 Martel-Pelletier J, Cloutier J-M, Pelletier J-P. In vivo effectsof antirheumatic drugs on neutral collagenolytic proteasesin human rheumatoid arthritis cartilage and synovium.JRheumatol 1988;15:1198-204.
11 MacNaul KL, Chartrain N, Lark M, Tocci MJ, HutchinsonNI. Discordinate expression of stromelysin, collagenase,and tissue inhibitor of metalloproteinases-I in rheumatoidsynovial fibroblasts. Synergistic effect of interleukin-1 andtumor necrosis factor-a on stromelysin expression. J BiolChem 1990;265: 17238-45.
12 Hutchinson NI, Lark MW, MacNaul KL, Harper C, Hoer-rner LA, McDonnell J, et al. In vivo expression of strome-lysin in synovium and cartilage of rabbits injectedintraarticularly with interleukin-1p. Arthritis Rheum 1992;35:1227-33.
13 Dayer JM, Breard J, Chess L, Krane SM. Participation ofmonocyte-macrophages and lymphocytes in the produc-tion of a factor that stimulates collagenase and prostaglan-din release by rheumatoid synovial cells. J7 Clin Invest 1979;64:1386-92.
14 Murphy G, Hembry RM, Reynolds JJ. Characterization of aspecific antiserum to rabbit stromelysin and demonstrationof the synthesis of collagenase and stromelysin bystimulated rabbit articular chondrocytes. Collagen Relat Res1986;6:351-64.
15 Buttle DJ, Handley CJ, Ilic MZ, Saklatvala J, Murata M,Barrett AJ. Inhibition of cartilage proteoglycan release by aspecific inactivator of cathepsin B and an inhibitor ofmatrix metalloproteinases. Evidence for two convergingpathways of chondrocyte-mediated proteoglycan degrada-tion. Arthritis Rheum 1993;36:1709-17.
16 Andrews HJ, Plumpton TA, Harper GP, Cawston TE. Asynthetic peptide metalloproteinase inhibitor, but notTimp, prevents the breakdown of proteoglycan withinarticular cartilage in vitro. Agents Actions 1992;37:147-54.
M338
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from

Inhibitors of collagenase but not ofgelatinase reduce cartilage explant proteoglycan breakdown
17 Nixon JS, Bottomley KMK, Broadhurst MJ, Brown PA,Johnson WH, Lawton G, et al. Potent collagenase inhibitorsprevent interleukin 1-induced cartilage degradation invitro. IntJ Tiss Reac 1991;13:237-43.
18 Caputo CB, Sygowski LA, Wolanin DJ, Patton SP, CacceseRG, Shaw A, et al. Effect of synthetic metalloproteaseinhibitors on cartilage autolysis in vitro. 7 Pharmacol ExpTher 1987;240:460-5.
19 Cawston T. Blocking cartilage destruction with metallopro-teinase inhibitors: a valid therapeutic target? Ann RheumDis 1993;52:769-70.
20 Davies B, Brown PD, East N, Crimmin NJ, Balkwill FR. Asynthetic matrix metalloproteinase inhibitor decreasestumor burden and prolongs survival of mice bearinghuman ovarian carcinoma xenografts. Cancer Res 1993;53:2087-91.
21 Rich DH, Brown MA, Barrett AJ. Purification of cathepsinB by a new form of affinity chromatography. Biochem J1986;235:731-4.
22 Barrett AJ. Human cathepsin B 1. Purification and someproperties of the enzyme. Biochem3J 1973;131:809-22.
23 Piquilloud Y, Reinharz A, Roth M. Studies on theangiotensin converting enzyme with different substrates.Biochim BiophysActa 1970;206:136-42.
24 Florentin D, Sassi A, Roques BP. A highly sensitive fluoro-metric assay for "enkephalinase", a neutral metalloen-dopeptidase that releases tyrosine-glycine-glycine fromenkephalins. Anal Biochem 1984;141:62-9.
25 Poole AR, Hembry RM, Dingle JT. Cathepsin D incartilage: the immunohistochemical demonstration ofextracellular enzyme in normal and pathological condi-tions. J Cell Sci 1974;14:139-61.
26 Knight CG, Willenbrock F, Murphy G. A novel coumarin-labelled peptide for sensitive continuous assays of thematrix metalloproteinases. FEBS Lett 1992;296:263-6.
27 Knauper V, Lopez-Otin C, Smith B, Knight G, Murphy G.Biochemical characterization of human collagenase-3. JBiol Chem 1996;271:1544-50.
28 Cawston TE, Barrett AJ. A rapid and reproducible assay forcollagenase using [1-'4C] acetylated collagen. Anal Biochem1979;99:340-5.
29 Cawston TE, Murphy G. Mammalian collagenases. MethodsEnzymol 1981;80:711-22.
30 Cawston TE, Galloway WA, Mercer E, Murphy G,Reynolds JJ. Purification of rabbit bone inhibitor ofcollagenase. Biochem J 1981 ;195: 159-65.
31 Sellers A, Reynolds JJ, Meikle MC. Neutral metalloprotein-ases of rabbit bone. Separation in latent forms of distinctenzymes that when activated degrade collagen, gelatin andproteoglycans. Biochem I 1979;171:493-6.
32 Buttle DJ, Saklatvala J, Tamai M, Barrett AJ. Inhibition ofinterleukin 1-stimulated cartilage proteoglycan degrada-tion by a lipophilic inactivator of cysteine endopeptidases.Biochem 7 1992;281: 175-7.
33 Handley CJ, Buttle DJ. Assay of proteoglycan degradation.Methods Enzymol 1995;248:47-58.
34 Andrews HJ, Edwards TA, Cawston TE, Hazleman BL.Transforming growth factor-beta causes partial inhibitionof interleukin-l-stimulated cartilage degradation in vitro.Biochem Biophys Res Commun 1989;162:144-50.
35 Farndale RW, Buttle DJ, Barrett AJ. Improved quantitationand discrimination of sulphated glycosaminoglycans by useof dimethylmethylene blue. Biochim Biophys Acta 1986;883:173-7.
36 Buttle DJ, Saklatvala J, Barrett AJ. The inhibition ofinterleukin 1-stimulated cartilage proteoglycan degrada-tion by cysteine endopeptidase inactivators. Agents ActionsSuppl 1993;39:161-5.
37 Saklatvala J. Tumour necrosis factor a stimulates resorptionand inhibits synthesis of proteoglycan in cartilage. Nature1986;322:547-9.
38 Gullberg U, Lantz M, Lindvall L, Olsson I, Himmler A.Involvement of an Asn/Val cleavage site in the productionof a soluble form of a human tumor necrosis factor (TNF)receptor. Site-directed mutagenesis of a putative cleavagesite in the p55 TNF receptor chain. Eur . Cell Biol1992;58:307-12.
39 Stefanovic-Racic M, Stadler J, Georgescu HI, Evans CH.Nitric oxide and energy production in articular chondro-cytes. J Cell Physiol 1994;159:274-80.
40 Bird TA, Gearing AJH, Saklatvala J. Murine interleukin-lreceptor: differences in binding properties between fibro-blastic and thymoma cells and evidence for a two-chainreceptor model. FEBS Lett 1987;225:21-6.
41 Taylor DJ, Whitehead RJ, Evanson JM, Westmacott D,Feldmann M, Bertfield H, et al. Effect of recombinant
cytokines on glycolysis and fructose 2,6-bisphosphate inrheumatoid synovial cells in vitro. Biochem _7 1988;250:111-15.
42 Buttle DJ, Saklatvala J. Lysosomal cysteine endopeptidasesmediate interleukin 1-stimulated cartilage proteoglycandegradation. Biochem37 1992;287:657-61.
43 Tyler JA. Articular cartilage cultured with catabolin (piginterleukin 1) synthesizes a decreased number of normalproteoglycan molecules. Biochem. 1985;227:869-78.
44 Campbell MA, Handley CJ. The effect of retinoic acid onproteoglycan biosynthesis in bovine articular cartilage cul-tures. Arch Biochem Biophys 1987;253:462-74.
45 Kozaci LD, Buttle DJ, Hollander AP. Degradation of type IIcollagen, but not proteoglycan, correlates with matrix met-alloproteinase activity in cartilage explant cultures. ArthritisRheum (in press).
46 Allan JA, Hembry RM, Angal S, Reynolds JJ, Murphy G.Binding of latent and high Mr active forms of stromelysin tocollagen is mediated by the C-terminal domain. . Cell Sci1991 ;99:789-95.
47 Woessner JF. Quantification of matrix metalloproteinases intissue samples. Methods Enzymol 1995;248:510-28.
48 Bode W, Gomis-Ruth F-X, Stockler W. Astacins, serralysins,snake venom and matrix metalloproteinases exhibit identi-cal zinc-binding environments (HEXXHXXGXXH andMet-turn) and topologies and should be grouped into acommon family, the 'metzincins'. FEBS Lett 1993;313:134-40.
49 Zhang D, Botos I, Gomis-Riith F-X, Doll R, Blood C,Njoroge FG, et al. Structural interaction of natural andsynthetic inhibitors with the venom metalloproteinase,atrolysin C (form d). Proc Natl Acad Sci USA 1994;91:8447-51.
50 Murphy G, Cockett MI, Stephens PE, Smith BJ, DochertyAJP. Stromelysin is an activator of procollagenase. A studywith natural and recombinant enzymes. Biochem 7 1987;248:265-8.
51 Yocum SA, Lopresti-Morrow LL, Gabel CA, Milici AJ, Mitch-ell PG. Bafilomycin Al inhibits IL-1-stimulated proteoglycandegradation by chondrocytes without affecting stromelysinsynthesis. Arch Biochem Biophys 1995;316:827-35.
52 Sandy JD, Neame PJ, Boynton RE, Flannery CR. Catabo-lism of aggrecan in cartilage explants. _7 Biol Chem1991;266:8683-5.
53 Sandy JD, Flannery CR, Neame PJ, Lohmander LS. Thestructure of aggrecan fragments in human synovial fluid.Evidence for the involvement in osteoarthritis of a novelproteinase which cleaves the Glu373-Ala374 bond in theinterglobular domain. _7 Clin Invest 1992;89:1512-16.
54 Lohmander LS, Neame PJ, Sandy JD. The structure ofaggrecan fragments in human synovial fluid. Evidence thataggrecanase mediates cartilage degradation in inflamma-tory joint disease, joint injury, and osteoarthritis. ArthritisRheum 1993;36: 1214-22.
55 Ilic MZ, Handley CJ, Robinson HC, Mok MT. Mechanismof catabolism of aggrecan by articular cartilage. ArchBiochem Biophys 1992;294: 115-22.
56 Fosang AJ, Last K, Neame PJ, Murphy G, Knauper V,Tschesche H, et al. Neutrophil collagenase (MMP-8)cleaves at the aggrecanase site E373-A374 in theinterglobular domain of cartilage aggrecan. Biochem _71994;304:347-5 1.
57 Cole AA, Chubinskaya S, Schumacher B, Huch K, Cs-SzaboG, Yao J, et al. Chondrocyte matrix metalloproteinase-8.Human articular chondrocytes express neutrophil colla-genase.JBiol Chem 1996;271:11023-6.
58 Fosang AJ, Neame PJ, Hardingham TE, Murphy G, Hamil-ton JA. Cleavage of cartilage proteoglycan between GI andG2 domains by stromelysins. _7 Biol Chem 1991 ;266:15579-82.
59 Fosang AJ, Last K, Knauper V, Neame PJ, Murphy G,Hardingham TE, et al. Fibroblast and neutrophil colla-genases cleave at two sites in the cartilage aggrecaninterglobular domain. BiochemJ 1993;295:273-6.
60 Fosang AJ, Neame PJ, Last K, Hardingham TE, Murphy G,Hamilton JA. The interglobular domain of cartilage aggre-can is cleaved by PUMP, gelatinases and cathepsin B. . BiolChem 1992;267: 19470-4.
61 Flannery CR, Lark MW, Sandy JD. Identification of a strome-lysin cleavage site within the interglobular domain of humanaggrecan. Evidence for proteolysis at this site in vivo in humanarticular cartilage. _7 Biol Chem 1992;267:1008-14.
62 Hughes C, Murphy G, Hardingham TE. Metalloproteinasedigestion of cartilage proteoglycan. Pattern of cleavage bystromelysin and susceptibility to collagenase. Biochem _71991 ;279:733-9.
M339
on 26 August 2018 by guest. P
rotected by copyright.http://m
p.bmj.com
/C
lin Mol P
athol: first published as 10.1136/mp.49.6.M
331 on 1 Decem
ber 1996. Dow
nloaded from