abcc2 is involved in the hepatocyte perinuclear barrier for small organic compounds
TRANSCRIPT

Biochemical Pharmacology 84 (2012) 1651–1659
ABCC2 is involved in the hepatocyte perinuclear barrier for small organiccompounds
Ruben Rosales, Maria J. Monte, Alba G. Blazquez, Oscar Briz, Jose J.G. Marin *
Laboratory of Experimental Hepatology and Drug Targeting, University of Salamanca, IBSAL, CIBERehd, Spain
A R T I C L E I N F O
Article history:
Received 15 August 2012
Accepted 18 September 2012
Available online 3 October 2012
Keywords:
ABC proteins
Bilirubin
Mitoxantrone
MRP2
Liver toxicity
A B S T R A C T
Small organic molecules are believed to freely diffuse across nuclear pores but this may not be so if this
route is blocked during protein and nucleic acid transfer. Here we have investigated the existence of
transport mechanisms across the nuclear envelope (NE) of hepatocytes. Using nuclei isolated from rat
liver cells, evidence for the existence of ATP-dependent transporters of organic compounds was found. In
rat hepatocyte NE, with negligible contamination by other membranes, the presence of mature and
glycosylated ABCC2, but not other ABC export pumps, was detected. ABCC2 was localized in the same
membranes as the conjugating enzyme UGT1A1. Human ABCC2 ORF was tagged with V5 and transfected
to human hepatoma cells. ABCC2-V5 protein was detected at perinuclear ER vesicles and at the NE. Both
compartments expressing ABCC2-V5 were able to exclude calcein. ABCC2 abundance at the NE of rat
hepatocytes was modified by treatments able to increase or reduce the expression of canalicular ABCC2.
The sensitivity to mitoxantrone was higher for hepatocytes obtained from TR- rats whose NE lacked
ABCC2. Incubation with mitoxantrone after depletion of ATP resulted in a marked accumulation of
mitoxantrone in the nucleus of wild-type, but not TR-, hepatocytes. In sum, ABCC2 is present at the NE
and perinuclear ER where, in combination with the activity of conjugating enzymes, this pump may be
involved in the perinuclear barrier for small organic molecules, playing a role in protecting DNA from
genotoxic compounds and in the control of intranuclear concentrations of ligands for nuclear receptors.
� 2012 Elsevier Inc. All rights reserved.
Contents lists available at SciVerse ScienceDirect
Biochemical Pharmacology
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/b io c hem p har m
1. Introduction
Many liver functions are regulated by modification in geneexpression profiles to adjust the phenotype of hepatocytes andliver cells to continuously changing conditions. This is accountedfor by a complex network of nuclear receptors able to respond tochanges in the chemical composition of extracellular environmentand the intracellular metabolic profile. Accordingly, a large varietyof small organic molecules must reach the nucleus of hepatocytesto interact with their nuclear receptors. This is for instance the caseof bile acids able to activate the farnesoid X receptor (FXR), which
Abbreviations: ABC, ATP-binding cassette; BDL, bile duct ligation; CAT, chloram-
phenicol acetyltransferase; CDDP, cisplatin; CGamF, cholylglycylamidofluorescein;
FITC, fluorescein isothiocyanate; MRP, multidrug resistance-associated protein;
MTX, mitoxantrone; NE, nuclear envelope; NPC, nuclear pore complex; UDCGamF,
ursodeoxycholylglycylamidofluorescein.
* Corresponding author at: Department of Physiology and Pharmacology,
Campus Miguel de Unamuno E.I.D. S-09, 37007 Salamanca, Spain.
Tel.: +34 923 294674; fax: +34 923 294669.
E-mail addresses: [email protected] (R. Rosales), [email protected] (M.J. Monte),
[email protected] (A.G. Blazquez), [email protected] (O. Briz), [email protected]
(Jose J.G. Marin).
0006-2952/$ – see front matter � 2012 Elsevier Inc. All rights reserved.
http://dx.doi.org/10.1016/j.bcp.2012.09.029
specifically activates the transcription of genes involved in bileacid homeostasis, but which has been recently suggested to be alsoinvolved in very different functions, such as the activation ofmechanisms of chemoresistance [1]. Translocation to the nucleusafter binding of bile acids in the cytosol to carrier proteins, such asthe glucocorticoid receptors [2–4] and probably histone deacety-lases [5] has been suggested as a mechanism that permits theentrance of bile acids to the nucleus. However, in previous studies,we have demonstrated that the access of small organic molecules(�1 kDa) to the nucleus is not strictly dependent on cytoplasmicproteins and could be partly accounted for by simple diffusion,probably through the nuclear pore complexes (NPCs) [6]. These arestructures formed by 8 lateral channels and 1 central pore [7,8]. Formolecules smaller than 10 kDa transit across the NPC probablyoccurs across the lateral channels, whereas the central poreprobably permits the passage of molecules of up to 40 kDa [6,9].Nevertheless, previous findings also revealed that there is a certainmolecular selectivity in the ability of isolated nuclei to retainorganic compounds due to mechanisms that are not merelydependent upon the molecular weight or the charge of thecompound [6]. Thus, although small organic molecules arebelieved to freely diffuse across nuclear pores this may not beso if this route is blocked during protein and nucleic acid transfer.

R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–16591652
Additionally, regarding the ability of the whole nucleus to take up,retain and export small molecules, the nuclear envelope (NE)constitutes an important compartment that could affect the netexchange of these compounds with the cytoplasm.
A simple explanation for the molecular selectivity in thenucleus–cytoplasm exchange would be the existence of carrierproteins involved in the trafficking across the membranousboundary between both compartments. This is not unexpectedbecause among the more than 70 proteins that have beenidentified in the NE the functions of only few more than 10 ofthem have been elucidated [10]. An important type of primaryactive transporters involved in the barrier function of severalepithelia are ATP-binding cassette (ABC) proteins which are able tocarry out ATP-dependent pumping of a large variety of substrates[11]. Several of the 48 human genes belonging to the ABCsuperfamily have been involved in the multidrug resistance (MDR)phenotype characteristic of many tumor cells. One of these genes(ABCC2) encodes ABCC2 protein, initially described as thecanalicular multiple organic anion transporter (cMOAT) [12] andnow usually called the multidrug resistance-associated protein-2(MRP2). This pump is expressed at the apical membranes ofhepatocytes and epithelial cells of renal proximal tubules,gallbladder, small intestine, colon, and lung [13]. Some membersof the ABC superfamily of proteins, such as these of family D, areexpressed in intracellular membranes and play a role in limitingthe net trafficking between different subcellular compartments. Inthis respect, using immunohistochemical techniques some evi-dence for the presence of ABCC2 at the nucleus, presumably at theNE, of cells from ovarian [14] and breast [15] carcinoma, andABCC1, not at the NE but inside the nuclei, of mucoepidermoidcarcinoma cells [16] have been suggested but not unambiguouslydemonstrated.
Since, on one hand, the liver is an important organ regardingdrug detoxification, and hence the hepatocyte nuclei are particu-larly exposed to genotoxic compounds and, on the other hand,ABCC2 is highly expressed in these cells the present study wasaimed to investigate the presence of this ABC protein at the NE andthe closely connected perinuclear ER compartment of hepatocytes.Moreover, differential sensitivity to genotoxicity of the ABCC2substrate mitoxantrone of hepatocytes from normal and ABCC2-deficient rats was investigated.
2. Materials and methods
2.1. Chemicals
Glycoursodeoxycholic acid (GUDCA) was from Calbiochem(Darmstadt, Germany). Sodium salt of glycocholic acid (GCA),fluorescein isothiocyanate (FITC), doxorubicin, calcein, rhodamine123, dexamethasone, cisplatin (CDDP) and mitoxantrone (MTX)were from Sigma–Aldrich (Madrid, Spain). Calcein acetoxymethylester (calcein-AM) was from Invitrogen (Carlsbad, CA). Cholylgly-cyl amidofluorescein (CGamF) and ursodeoxycholylglycyl amido-fluorescein (UDCGamF) were synthesized by coupling the amidogroup of FITC to the carboxyl group of the glycine moiety of GCA orGUDCA, respectively [17].
2.2. Animals and cells
Male Wistar (Animal House, University of Salamanca, Spain)and Wistar TR- (HsdAmc:TR-Abcc2; Harlan, OR) received humanecare according to the criteria outlined in the ‘‘Guide for the Careand Use of Laboratory Animals’’ prepared by the National Academyof Sciences and published by the National Institutes of Health (NIHpublication vol. 2, 2nd ed., 2002). The experimental protocols wereapproved by the University of Salamanca Ethical Committee for the
Use of Laboratory Animals. Rat hepatocytes were isolated fromWistar or TR- rats, seeded in collagen-coated plates and incubatedin Williams’ Medium E as described elsewhere [18]. Alexander(PLC/PRF/5) human hepatoma cells from the American TypeCulture Collection (ATCC: CRL-8024, LGC Standards S.L.U., Barce-lona, Spain) were cultured with an appropriate medium in ahumidified CO2:air (5:95%) atmosphere at 37 8C.
2.3. Studies on isolated nuclei
Nuclei were obtained from rat liver homogenates as previouslydescribed [19,6], purified by serial centrifugation [20], suspendedin STM solution (250 mM sucrose, 50 mM Tris HCl, and 5 mMMgSO4, pH 7.4), and counted in a Neubauer chamber. Thesepreparations consisted of clean suspensions of nuclei free of celldebris (Fig. 1), which were kept at 4 8C until use within thefollowing 24 h. Flow cytometry using a FACSort flow cytometer(BD Biosciences, Madrid, Spain) was used to determine nuclearuptake or efflux (after loading) of fluorescent compounds aspreviously described [6]. In some experiments, the incubationbuffer also contained 3 mM ATP plus an ATP-regenerating system(3 mM phosphocreatine plus 100 mg/ml creatine phosphokinase)[21].
2.4. Experiments on isolated hepatocytes
Freshly isolated hepatocytes from Wistar or TR- rats werepreincubated with PBS (control) or with ATP-depleting medium(PBS containing 15 mM sodium azide and 50 mM 2-deoxyglucose)[22] at 37 8C in 5% CO2 for 20 min and then incubated with 5 mMMTX in PBS or in ATP-depleting medium for 30 min. Afterincubation, cells were analyzed on the flow cytometer. Dead cells,whose identification was based on propidium iodide staining, wereexcluded from the data analysis. To visualize the subcellularlocalization of MTX the same treatment was also performed inhepatocytes seeded in collagen-coated plates. After the incubationwith MTX, cells were fixed and observed by confocal microscopy.Cell viability in wild type or TR- hepatocytes was determined usingthe formazan test (CellTiter 96-AQUEOUS Non-Radioactive CellProliferation Assay, Promega, Madrid, Spain) after incubation withMTX for 72 h.
2.5. Western blot analyses
Western blot analyses were performed on rat liver homo-genates, crude liver membranes, isolated hepatocyte nuclei andmembrane nuclear ghosts derived from NE obtained from isolatednuclei by treatment with DNAse I and RNAse A [23]. In some casessamples were incubated with 500 U of PNGase F (New EnglandBioLabs, Ipswich, UK) for 60 min to deglycosylate proteins. In someexperiments rats were treated with dexamethasone (50 mg/kg/day, i.p., 4 days) or CDDP (2 mg/kg/day, i.p., 4 days) or subjected tobile duct ligation (BDL) 2 days prior to being used as donors forhepatocyte nuclei isolation.
Immunoblotting analyses were carried out in 7.5% sodiumdodecyl sulfate-polyacrylamide gels, loading 25 mg of protein perlane, and then were transferred onto a nitrocellulose membrane(Bio-Rad, Hercules, CA). The primary antibodies were as follows:rabbit polyclonal antibodies against rat calnexin (ab22595 Abcam,Cambridge, UK), lamin B1 (ab16048, Abcam), ABCC2 (K13,generously donated by Bruno Stieger from Zurich University),ABCC3 (M0318, Sigma–Aldrich), ABCC6 (sc-25505, Santa CruzBiotechnology, CA) and UGT1A1 (AB10339, Millipore Iberica,Madrid); and mouse monoclonal antibodies against rat ABCG2(BXP21, Abcam), GAPDH (SC32233, Santa Cruz Biotechnology),ABCB1 (Mdr1/Mdr2, ab3364, Abcam), ABCC2 (M2III5 and M2III6,

R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–1659 1653
Alexis Axxora, Exeter, UK), ABCC4 (ab15602, Abcam), NPC(Mab414, Abcam), Na+/K+-ATPase (M7-PB-E9, Abcam) andTGN38 (Thermo Scientific, Madrid, Spain). Anti-rabbit and anti-mouse IgG horseradish peroxidase-linked secondary antibodies(Amersham Pharmacia Biotech, Freiburg, Germany) were used.
2.6. Immunofluorescence assays
Immunofluorescence studies were carried out on fixed rathepatocytes, rat hepatocyte nuclei or human hepatoma Alexandercells by using ab22595 antibody for calnexin, ab16048 antibody forlamin B1, Mab414 antibody for NPC (all of them from Abcam),M2III6 antibody for ABCC2 (Alexis Axxora), AB10339 antibody forUGT1A1 (Millipore), R961 antibody for V5 (Invitrogen) and cyanin-5-conjugated anti-rabbit secondary antibody (Invitrogen), AlexaFluor 488 or Alexa Fluor 594-conjugated anti-rabbit or anti-mousesecondary antibody (Molecular Probes, Leiden, The Netherlands),as appropriate. Negative controls were obtained by omitting theprimary antibody.
2.7. Cloning and expression of human ABCC2
From total RNA extracted from healthy human liver, the ORF ofhuman ABCC2 was amplified by RT-PCR using the high-fidelityAccuPrime Pfx DNA polymerase (Invitrogen), and specific oligo-nucleotide primers (forward: 50-GGGGA CAAGT TTGTA CAAAAAAGCA GGCTG CATGC TGGAG AAGTT CTGCAAC; reverse: 50-GGGGA CCACT TTGTA CAAGA AAGCT GGGTG GAATT TTGTG CTGTTCACATTCTC), to which adapters for HindIII and BglII and attB sitesto obtain cDNA adapted for Gateway1 cloning (Invitrogen) were
Fig. 1. (A) Representative image of phase-contrast microscopy of rat hepatocyte nuclei
confocal microscopy of isolated rat hepatocyte nuclei. The nuclear envelope was labele
stained with propidium iodide (B) or DAPI (C). (D) Representative Western blot analysis o
loading 25 mg protein/lane from three liver homogenates (lanes 1–3) and three prepa
expected: 68 kDa for lamin B1, 37 kDa for GAPDH, 110 kDa for Na+/K+-ATPase, 90 kDa
added. Multicloning site region of pcDNA3.1-hygro(+) wasamplified by PCR and recombined with the pDONR207 vector(Invitrogen) to generate Entry plasmid, which was digested withHindIII and BglII. cDNA of ABCC2 was then ligated with T4-ligase(Promega). Entry plasmid containing ABCC2 was recombined withthe pcDNA6.2/V5-DEST destination vector (Invitrogen), containingthe C-terminal V5 reporter gene, to generate an expression vector.As a control of non-specific subcellular localization, the bacterialprotein chloramphenicol acetyltransferase (CAT) (Invitrogen) wasused. The obtained ABCC2-V5 expression vector was used totransfect Alexander cells by using lipofectamine (Invitrogen). After3 days in culture, transfected cells were used for immunofluores-cence assays. In some cases, preincubation with calcein-AM for30 min was performed before cells were fixed and processed forimmunofluorescence assays.
2.8. Analytical and statistical methods
DNA measurements were carried out using Hoechst-33258 as afluorescent probe [24] and calf thymus DNA as standard. Tocalculate the statistical significance of differences among groups,the paired Student’s t-test or the Bonferroni method of multiple-range testing was used, as appropriate.
3. Results
To investigate whether the molecular selectivity in thenucleus–cytoplasm exchange of small organic molecules wouldinvolve carrier proteins playing a role in the overall traffic acrossthe membranous boundary between both compartments, isolated
isolated from liver homogenate. (B and C) Representative images obtained by laser
d with antibodies against the nuclear pore complex (B) or lamin B1 (C). DNA was
f typical markers of intracellular organelles and cellular membranes, carried out by
rations of isolated rat hepatocyte nuclei (lanes 4–6). Molecular weights were as
for calnexin and 38 kDa for the trans golgi network (TGN38).

R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–16591654
nuclei were obtained from rat hepatocytes as previously described[6]. Light microscopy revealed that these preparations consisted ofa clean suspension of nuclei with no debris or contamination byother organelles (Fig. 1A). Immunofluorescence labelling of theNPC (Fig. 1B) or lamin B1 located in the inner membrane of the NE(Fig. 1C) indicated the structural integrity of the isolated nuclei.
The absence of contamination of isolated nuclei by othercellular regions was determined by Western blot (Fig. 1D). GAPDH(used here as a marker of cytoplasmic proteins), TGN38 (a markerof the trans-golgi network) and Na+/K+-ATPase (a marker of plasmamembrane), were detected in the homogenate but not in thenuclei. As expected [25] lamin B1 and calnexin were seen in bothhomogenate and nuclei.
Flow cytometry analysis of propidium iodide labeled nucleirevealed that these were distributed into two populations ofdiploid (small) and tetraploid (large) nuclei (data not shown).When incubated with FITC for 30 min both groups were able totake up the fluorescent compound and reached final values higherin larger than in smaller nuclei (Fig. 2A). When the ability to takeup several compounds, such as FITC, CGamF, UDCGamF, calcein,rhodamine-123 and doxorubicin was evaluated in the absence andin the presence of ATP plus an ATP-regenerating system the overallload by the nuclei was reduced under energy supplementation
Fig. 2. Uptake (A and B) and efflux (C and D) of fluorescent compounds by isolated rat
isothiocyanate (FITC) uptake by isolated rat hepatocyte nuclei incubated with 20 mM F
observed, large nuclei (closed circles) and small nuclei (open circles). (B) Effect of ATP o
incubated with 20 mM FITC, cholylglycyl amidofluorescein (CGamF) or ursodeoxycholy
doxorubicin at 37 8C for 5 min, in the absence (control) or the presence of 3 mM ATP p
phosphokinase). (C) Efflux of UDCGamF from isolated rat hepatocyte nuclei that were pre-
were diluted with 100 volumes of UDCGamF-free medium. Samples were taken over the
absence (�ATP) or the presence (+ATP) of 3 mM ATP plus an ATP-regenerating system o
values obtained by flow cytometry analysis in 5 experiments performed in duplicate on
experiment are also shown. *p < 0.05, significantly different from the values obtained in t
conditions (Fig. 2B). Efflux of UDCGamF from nuclei preloaded for30 min displayed two components: (i) a rapid efflux, presumablydue to diffusion across nuclear pores that once isolated from cellsare free of protein/nucleic acids transport functions, followed by(ii) a slower component that we hypothesized might be due to theefflux of UDCGamF from the NE compartment (Fig. 2C). The slopeof the first component was not affected by the presence of ATP(data not shown) but that of the second one was consistently andsignificantly steeper when ATP plus an ATP-regenerating systemwere present in the medium (Fig. 2D).
To determine whether the activity of an ATP-dependent pumpcould be involved in diminishing the net nuclear uptake of thesefluorescent compounds, the presence in the NE of several ABCproteins was investigated. The presence of P-glycoproteins (theprimary antibody recognizes Mdr1, Mdr2 and probably othersmembers of ABCB family), ABCG2, ABCC2, ABCC3, ABCC4 andABCC6 was determined in rat liver homogenate, crude membranepreparations, isolated nuclei and NE. NPC and lamin B1 wereassayed as markers of NE. All ABC proteins were detected in liverhomogenate and crude membrane preparations but only ABCC2gave a clear band in isolated nuclei and NE (Fig. 3A). Since amongthe fluorescent compounds assayed only calcein is a knownsubstrate of ABCC2, these results suggested that the observed
hepatocyte nuclei as measured by flow cytometry. (A) Time course of fluorescein
ITC during the indicated times. Two nuclear populations with different size were
n the uptake of fluorescent compounds by the large nuclei population. Nuclei were
lglycyl amidofluorescein (UDCGamF), or with 5 mg/ml calcein, rhodamine 123 or
lus an ATP-regenerating system (3 mM phosphocreatine plus 100 mg/ml creatine
incubated at 37 8C with 20 mM UDCGamF for 30 min before the nuclear suspensions
following 30 min to analyze the fluorescence retained in the nuclei. (D) Effect of the
n UDCGamF content at minute 10. Values are means � SD of the mean fluorescence
5 different nuclear preparations. In (D) individual paired values obtained for each
he absence of ATP by the paired t-test. AUF, arbitrary units of fluorescence.

Fig. 3. (A) Representative Western blot analysis of ABC proteins carried out by loading 25 mg protein/lane from rat liver homogenates (lane 1), crude membrane preparations
(lane 2), isolated rat hepatocyte nuclei (lane 3) and isolated nuclear envelopes (NE) from rat hepatocyte nuclei (lane 4). Molecular weights were as expected for each carrier:
�140 kDa for MDRs, 72 kDa for ABCG2, 190 kDa for ABCC2, 180 kDa for ABCC3, 159 kDa for ABCC4 and 170 kDa for ABCC6, and for the NE markers nuclear pore complex (NPC;
62 kDa) and lamin B1 (68 kDa). (B) Representative Western blot analysis of ABCC2 carried out in rat liver homogenates (lane 1), crude membrane preparations (lane 2),
isolated rat hepatocyte nuclei (lane 3) and NE from rat hepatocyte nuclei (lane 4). Two different antibodies against ABCC2 were used (K13 and M2III6). Mab414 antibody
against the NPC was used as a NE marker. (C) Representative Western blot analysis of ABCC2 in samples treated with (DG) or without (control) PNGase F enzyme to
deglycosylate glycoproteins. Upper panel shows rat liver homogenates (lanes 1 and 2) and crude membrane preparations (lanes 3 and 4). Lower panel shows isolated rat
hepatocyte nuclei (lanes 1 and 2) and NE from rat hepatocyte nuclei (lanes 3 and 4). In all cases 25 mg protein/lane were loaded. ABCC2 molecular weight was 190 kDa in
control and 180 kDa in deglycosylated samples. All results were confirmed in at least two additional experiments.
Fig. 4. (A) Representative Western blot analysis of UDP glucuronosyltransferase 1A1 (UGT1A1) carried out by loading 25 mg protein/lane from liver homogenates (lane 1),
crude membrane preparations (lane 2), isolated rat hepatocyte nuclei (lane 3) and isolated nuclear envelopes (NE) from rat hepatocyte nuclei (lane 4). Lamin B1 was used as a
marker for NE. (B) Representative image obtained by laser confocal microscopy of rat hepatocytes in primary cultured showing immunolocalization of UGT1A1 (red
fluorescence) and ABCC2 (green fluorescence). DNA was stained with DAPI (blue fluorescence). (C–F) Representative images obtained by laser confocal microscopy of
Alexander human hepatoma cells transfected with a V5-tagged ABCC2 construct. Immunolocalization of (C) V5 (green fluorescence) as a marker of exogenous ABCC2, and (D)
UGT1A1 (red fluorescence). (E) Merge fluorescence; DNA was stained with DAPI (blue). (F) A magnification of (E).
R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–1659 1655

R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–16591656
ATP-dependent effect was due to the involvement of other exportpumps not investigated here or to the fact that conjugation of thesecompounds occurs at the NE and their derivatives are transportedby ABCC2.
To elucidate whether the signal of ABCC2 obtained with themouse monoclonal antibody M2III5 was a specific one, similaranalyses were carried out with two additional different primaryantibodies K13 (rabbit polyclonal) and M2III6 (mouse monoclonal).All these gave similar results (Fig. 3B). Moreover, N-deglycosyla-tion using PNGase F [26] reduced the molecular weight of ABCC2detected in liver homogenate and NE at a similar extent (from 190to 180 kDa) (Fig. 3C).
Since a known role of ABCC2 in hepatocytes is the canalicularsecretion of glucuronated bilirubin we investigated whether theconjugating enzyme uridine 50-diphosphoglucoronosyl transferase1A1 (UGT1A1) was also expressed in the same sub-cellularcompartment. Western blot analysis revealed that UGT1A1 wasindeed present in isolated nuclei and NE (Fig. 4A). Immunoflor-escence revealed the presence of this protein mainly in the nuclearand perinuclear compartment of rat hepatocytes (red fluorescencein Fig. 4B). When ABCC2 was analyzed in these cells (greenfluorescence in Fig. 4B) the protein was mainly detected at theplasma membrane and perinuclear compartment. Detection ofABCC2 at the NE of freshly isolated hepatocytes or cryosections ofhuman and rat liver tissue gave not consistent results (data notshown). This is in agreement with the weak signal for endogenous
Fig. 5. Representative images obtained by laser confocal microscopy of Alexander human
or ABCC2 (B–I). (B) Immunolocalization of V5 (green fluorescence) as a marker of exo
envelope (NE). (C) Magnification of (B). (D–I) ABCC2-mediated exclusion of calcein from
construct 48 h before been incubated with 10 mM calcein-AM (green fluorescence) fo
exogenous ABCC2. Immunolocalization of calnexin (purple fluorescence) was used as
endoplasmic reticulum vesicles. DNA was stained with DAPI (blue fluorescence).
ABCC2 at the NE of ovarian carcinoma cells found by others usingelectron microscopy [14]. The presence at the NE in situ ofassociated proteins able to hide specific ABCC2 epitopes inimmunofluorescence analysis and/or the effect of the denaturationprocess used in Western blotting may account for the differentimmunoreactivity of anti-ABCC2 antibodies observed in bothtechniques.
To enhance the ability to in situ detect ABCC2 in the subcellularcompartment, human ABCC2 was cloned and tagged with V5. Thisconstruct was transfected to human hepatoma Alexander cells.Using this approach ABCC2 could be clearly detected at the NE inclose association with UGT1A1 (Fig. 4C–F). When in a similarconstruct the ORF of ABCC2 was replaced by that of the non-eukaryotic protein CAT the fluorescence was detected dispersewithin the cells (Fig. 5A) in contrast with ABCC2 that was located atthe NE and perinuclear membranes (Fig. 5B and C). When lamin B1was detected in the same cells both lamin B1 and ABCC2 werelocated at the NE (Fig. 5B and C) but at a different membrane; i.e.,lamin B1, as has been described (for review see [27]), was bound tothe inner face of the internal nuclear membrane whereas ABCC2was located at the outer membrane (Fig. 5C).
To elucidate whether subcellular ABCC2 was a functional pumpthe cells were incubated with calcein. Fig. 5D–I show a cellpositively transfected surrounded by cells not expressing ABCC2.Immunofluorescence images revealed the co-localization of ABCC2together with calnexin at the NE and perinuclear vesicles.
hepatoma cells transfected with a construct containing the ORF of V5-tagged CAT (A)
genous ABCC2, and lamin B1 (red fluorescence) as a marker for the inner nuclear
compartments expressing ABCC2. Cells were transfected with a V5-tagged ABCC2
r 30 min. Immunolocalization of V5 (red fluorescence) was used as a marker of
a marker of ER. Exclusion of calcein (green fluorescence) from the nucleus and
R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–1659 1657
Moreover, NE and ER vesicles, where ABCC2 was expressed, wereseen to be able to prevent calcein accumulation (Fig. 5G and I).
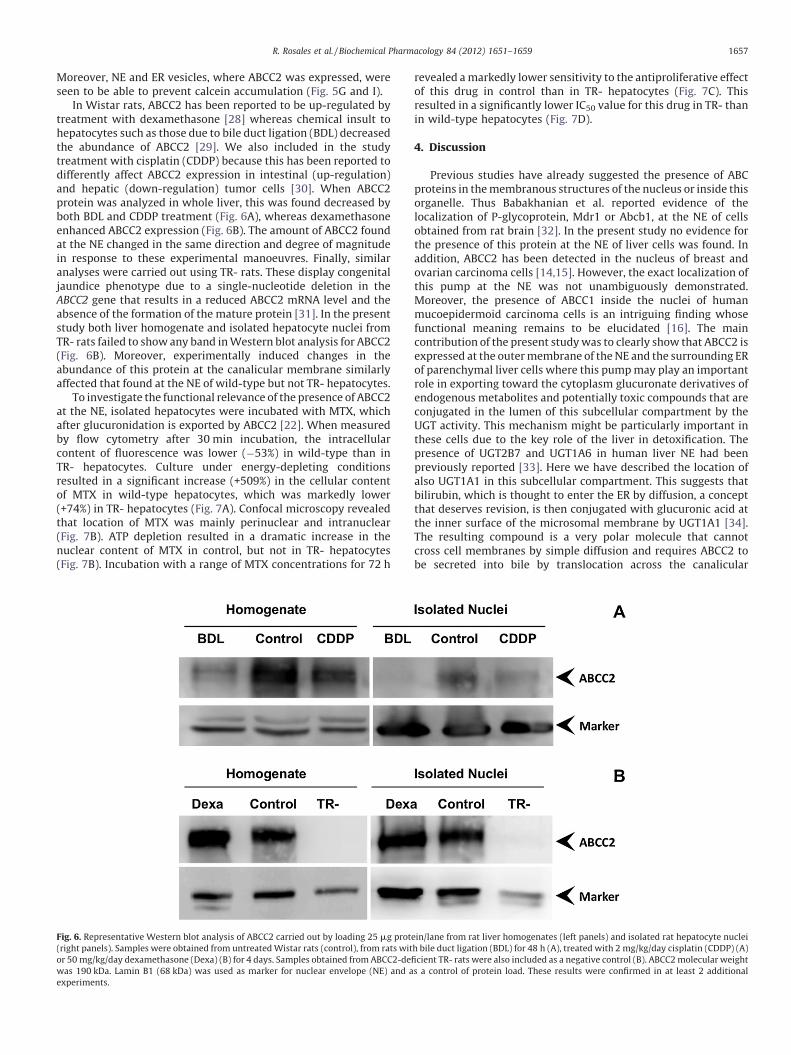
In Wistar rats, ABCC2 has been reported to be up-regulated bytreatment with dexamethasone [28] whereas chemical insult tohepatocytes such as those due to bile duct ligation (BDL) decreasedthe abundance of ABCC2 [29]. We also included in the studytreatment with cisplatin (CDDP) because this has been reported todifferently affect ABCC2 expression in intestinal (up-regulation)and hepatic (down-regulation) tumor cells [30]. When ABCC2protein was analyzed in whole liver, this was found decreased byboth BDL and CDDP treatment (Fig. 6A), whereas dexamethasoneenhanced ABCC2 expression (Fig. 6B). The amount of ABCC2 foundat the NE changed in the same direction and degree of magnitudein response to these experimental manoeuvres. Finally, similaranalyses were carried out using TR- rats. These display congenitaljaundice phenotype due to a single-nucleotide deletion in theABCC2 gene that results in a reduced ABCC2 mRNA level and theabsence of the formation of the mature protein [31]. In the presentstudy both liver homogenate and isolated hepatocyte nuclei fromTR- rats failed to show any band in Western blot analysis for ABCC2(Fig. 6B). Moreover, experimentally induced changes in theabundance of this protein at the canalicular membrane similarlyaffected that found at the NE of wild-type but not TR- hepatocytes.
To investigate the functional relevance of the presence of ABCC2at the NE, isolated hepatocytes were incubated with MTX, whichafter glucuronidation is exported by ABCC2 [22]. When measuredby flow cytometry after 30 min incubation, the intracellularcontent of fluorescence was lower (�53%) in wild-type than inTR- hepatocytes. Culture under energy-depleting conditionsresulted in a significant increase (+509%) in the cellular contentof MTX in wild-type hepatocytes, which was markedly lower(+74%) in TR- hepatocytes (Fig. 7A). Confocal microscopy revealedthat location of MTX was mainly perinuclear and intranuclear(Fig. 7B). ATP depletion resulted in a dramatic increase in thenuclear content of MTX in control, but not in TR- hepatocytes(Fig. 7B). Incubation with a range of MTX concentrations for 72 h
Fig. 6. Representative Western blot analysis of ABCC2 carried out by loading 25 mg prot
(right panels). Samples were obtained from untreated Wistar rats (control), from rats wit
or 50 mg/kg/day dexamethasone (Dexa) (B) for 4 days. Samples obtained from ABCC2-de
was 190 kDa. Lamin B1 (68 kDa) was used as marker for nuclear envelope (NE) and a
experiments.
revealed a markedly lower sensitivity to the antiproliferative effectof this drug in control than in TR- hepatocytes (Fig. 7C). Thisresulted in a significantly lower IC50 value for this drug in TR- thanin wild-type hepatocytes (Fig. 7D).
4. Discussion
Previous studies have already suggested the presence of ABCproteins in the membranous structures of the nucleus or inside thisorganelle. Thus Babakhanian et al. reported evidence of thelocalization of P-glycoprotein, Mdr1 or Abcb1, at the NE of cellsobtained from rat brain [32]. In the present study no evidence forthe presence of this protein at the NE of liver cells was found. Inaddition, ABCC2 has been detected in the nucleus of breast andovarian carcinoma cells [14,15]. However, the exact localization ofthis pump at the NE was not unambiguously demonstrated.Moreover, the presence of ABCC1 inside the nuclei of humanmucoepidermoid carcinoma cells is an intriguing finding whosefunctional meaning remains to be elucidated [16]. The maincontribution of the present study was to clearly show that ABCC2 isexpressed at the outer membrane of the NE and the surrounding ERof parenchymal liver cells where this pump may play an importantrole in exporting toward the cytoplasm glucuronate derivatives ofendogenous metabolites and potentially toxic compounds that areconjugated in the lumen of this subcellular compartment by theUGT activity. This mechanism might be particularly important inthese cells due to the key role of the liver in detoxification. Thepresence of UGT2B7 and UGT1A6 in human liver NE had beenpreviously reported [33]. Here we have described the location ofalso UGT1A1 in this subcellular compartment. This suggests thatbilirubin, which is thought to enter the ER by diffusion, a conceptthat deserves revision, is then conjugated with glucuronic acid atthe inner surface of the microsomal membrane by UGT1A1 [34].The resulting compound is a very polar molecule that cannotcross cell membranes by simple diffusion and requires ABCC2 tobe secreted into bile by translocation across the canalicular
ein/lane from rat liver homogenates (left panels) and isolated rat hepatocyte nuclei
h bile duct ligation (BDL) for 48 h (A), treated with 2 mg/kg/day cisplatin (CDDP) (A)
ficient TR- rats were also included as a negative control (B). ABCC2 molecular weight
s a control of protein load. These results were confirmed in at least 2 additional
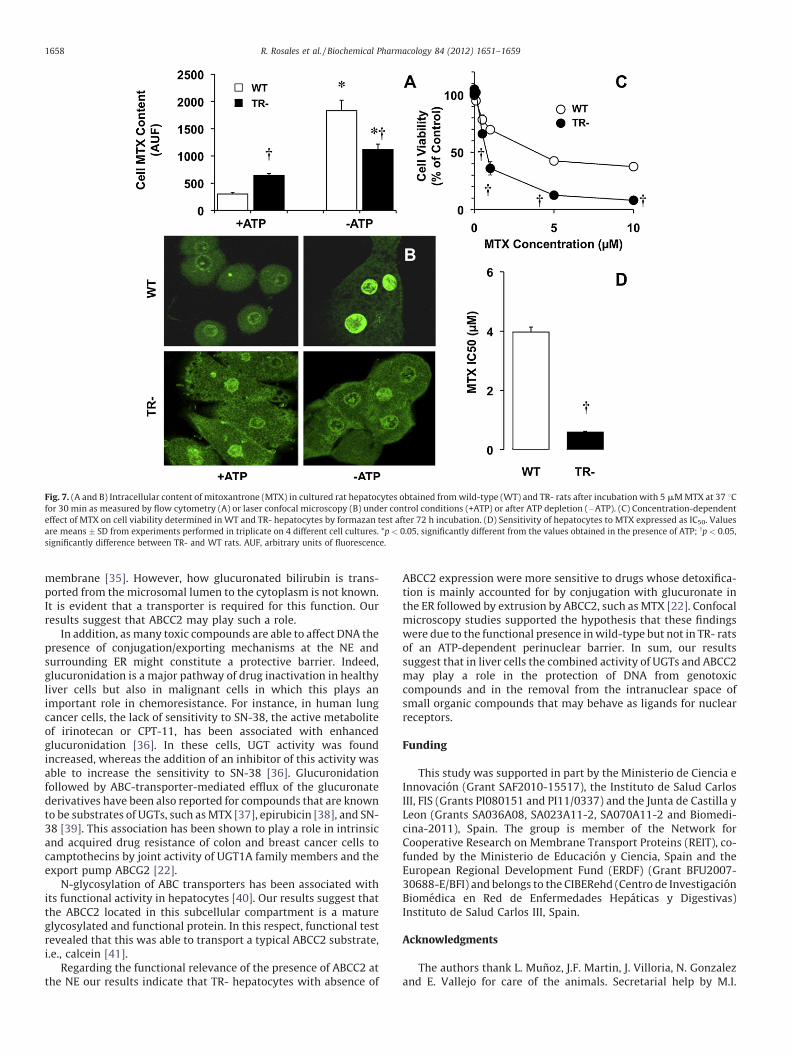
Fig. 7. (A and B) Intracellular content of mitoxantrone (MTX) in cultured rat hepatocytes obtained from wild-type (WT) and TR- rats after incubation with 5 mM MTX at 37 8Cfor 30 min as measured by flow cytometry (A) or laser confocal microscopy (B) under control conditions (+ATP) or after ATP depletion (�ATP). (C) Concentration-dependent
effect of MTX on cell viability determined in WT and TR- hepatocytes by formazan test after 72 h incubation. (D) Sensitivity of hepatocytes to MTX expressed as IC50. Values
are means � SD from experiments performed in triplicate on 4 different cell cultures. *p < 0.05, significantly different from the values obtained in the presence of ATP; yp < 0.05,
significantly difference between TR- and WT rats. AUF, arbitrary units of fluorescence.
R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–16591658
membrane [35]. However, how glucuronated bilirubin is trans-ported from the microsomal lumen to the cytoplasm is not known.It is evident that a transporter is required for this function. Ourresults suggest that ABCC2 may play such a role.
In addition, as many toxic compounds are able to affect DNA thepresence of conjugation/exporting mechanisms at the NE andsurrounding ER might constitute a protective barrier. Indeed,glucuronidation is a major pathway of drug inactivation in healthyliver cells but also in malignant cells in which this plays animportant role in chemoresistance. For instance, in human lungcancer cells, the lack of sensitivity to SN-38, the active metaboliteof irinotecan or CPT-11, has been associated with enhancedglucuronidation [36]. In these cells, UGT activity was foundincreased, whereas the addition of an inhibitor of this activity wasable to increase the sensitivity to SN-38 [36]. Glucuronidationfollowed by ABC-transporter-mediated efflux of the glucuronatederivatives have been also reported for compounds that are knownto be substrates of UGTs, such as MTX [37], epirubicin [38], and SN-38 [39]. This association has been shown to play a role in intrinsicand acquired drug resistance of colon and breast cancer cells tocamptothecins by joint activity of UGT1A family members and theexport pump ABCG2 [22].
N-glycosylation of ABC transporters has been associated withits functional activity in hepatocytes [40]. Our results suggest thatthe ABCC2 located in this subcellular compartment is a matureglycosylated and functional protein. In this respect, functional testrevealed that this was able to transport a typical ABCC2 substrate,i.e., calcein [41].
Regarding the functional relevance of the presence of ABCC2 atthe NE our results indicate that TR- hepatocytes with absence of
ABCC2 expression were more sensitive to drugs whose detoxifica-tion is mainly accounted for by conjugation with glucuronate inthe ER followed by extrusion by ABCC2, such as MTX [22]. Confocalmicroscopy studies supported the hypothesis that these findingswere due to the functional presence in wild-type but not in TR- ratsof an ATP-dependent perinuclear barrier. In sum, our resultssuggest that in liver cells the combined activity of UGTs and ABCC2may play a role in the protection of DNA from genotoxiccompounds and in the removal from the intranuclear space ofsmall organic compounds that may behave as ligands for nuclearreceptors.
Funding
This study was supported in part by the Ministerio de Ciencia eInnovacion (Grant SAF2010-15517), the Instituto de Salud CarlosIII, FIS (Grants PI080151 and PI11/0337) and the Junta de Castilla yLeon (Grants SA036A08, SA023A11-2, SA070A11-2 and Biomedi-cina-2011), Spain. The group is member of the Network forCooperative Research on Membrane Transport Proteins (REIT), co-funded by the Ministerio de Educacion y Ciencia, Spain and theEuropean Regional Development Fund (ERDF) (Grant BFU2007-30688-E/BFI) and belongs to the CIBERehd (Centro de InvestigacionBiomedica en Red de Enfermedades Hepaticas y Digestivas)Instituto de Salud Carlos III, Spain.
Acknowledgments
The authors thank L. Munoz, J.F. Martin, J. Villoria, N. Gonzalezand E. Vallejo for care of the animals. Secretarial help by M.I.

R. Rosales et al. / Biochemical Pharmacology 84 (2012) 1651–1659 1659
Hernandez and revision of the English spelling, grammar and styleof the manuscript by N. Skinner is also gratefully acknowledged.
References
[1] Herraez E, Gonzalez-Sanchez E, Vaquero J, Romero MR, Serrano MA, Marin JJ,et al. Cisplatin-induced chemoresistance in colon cancer cells involves FXR-dependent and FXR-independent up-regulation of ABC proteins. Mol Pharm2012;9:2565–76.
[2] Sola S, Amaral JD, Castro RE, Ramalho RM, Borralho PM, Kren BT, et al. Nucleartranslocation of UDCA by the glucocorticoid receptor is required to reduceTGF-beta1-induced apoptosis in rat hepatocytes. Hepatology 2005;42:925–34.
[3] Tanaka H, Makino I. Ursodeoxycholic acid-dependent activation of the gluco-corticoid receptor. Biochem Biophys Res Commun 1992;188:942–8.
[4] Weitzel C, Stark D, Kullmann F, Scholmerich J, Holstege A, Falk W. Ursodeoxy-cholic acid induced activation of the glucocorticoid receptor in primary rathepatocytes. Eur J Gastroenterol Hepatol 2005;17:169–77.
[5] Mitro N, Godio C, De Fabiani E, Scotti E, Galmozzi A, Gilardi F, et al. Insights inthe regulation of cholesterol 7alpha-hydroxylase gene reveal a target formodulating bile acid synthesis. Hepatology 2007;46:885–97.
[6] Monte MJ, Rosales R, Macias RI, Iannota V, Martinez-Fernandez A, Romero MR,et al. Cytosol-nucleus traffic and colocalization with FXR of conjugated bileacids in rat hepatocytes. Am J Physiol Gastrointest Liver Physiol 2008;295:G54–62.
[7] Aebi U, Pante N, Jarnik M. Structure and function of the nuclear pore complex, asupramolecular machine mediating molecular trafficking across the nuclearenvelope. Verh Dtsch Zool Ges 1992;85:285–96.
[8] Shahin V, Danker T, Enss K, Ossig R, Oberleithner H. Evidence for Ca2+- and ATP-sensitive peripheral channels in nuclear pore complexes. FASEB J 2001;15:1895–901.
[9] Peters R. Fluorescence microphotolysis to measure nucleocytoplasmic trans-port and intracellular mobility. Biochim Biophys Acta 1986;864:305–59.
[10] D’Angelo MA, Hetzer MW. The role of the nuclear envelope in cellularorganization. Cell Mol Life Sci 2006;63:316–32.
[11] Holland IB. ABC transporters, mechanisms and biology: an overview. EssaysBiochem 2011;50:1–17.
[12] Wada M, Toh S, Taniguchi K, Nakamura T, Uchiumi T, Kohno K, et al. Mutationsin the canilicular multispecific organic anion transporter (cMOAT) gene, anovel ABC transporter, in patients with hyperbilirubinemia II/Dubin–Johnsonsyndrome. Hum Mol Genet 1998;7:203–7.
[13] Taniguchi K, Wada M, Kohno K, Nakamura T, Kawabe T, Kawakami M, et al. Ahuman canalicular multispecific organic anion transporter (cMOAT) gene isoverexpressed in cisplatin-resistant human cancer cell lines with decreaseddrug accumulation. Cancer Res 1996;56:4124–9.
[14] Surowiak P, Materna V, Kaplenko I, Spaczynski M, Dolinska-Krajewska B,Gebarowska E, et al. ABCC2 (MRP2, cMOAT) can be localized in the nuclearmembrane of ovarian carcinomas and correlates with resistance to cisplatinand clinical outcome. Clin Cancer Res 2006;12:7149–58.
[15] Maciejczyk A, Jagoda E, Wysocka T, Matkowski R, Gyorffy B, Lage H, et al.ABCC2 (MRP2, cMOAT) localized in the nuclear envelope of breast carcinomacells correlates with poor clinical outcome. Pathol Oncol Res 2012;18:331–42.
[16] Cai BL, Xu XF, Fu SM, Shen LL, Zhang J, Guan SM, et al. Nuclear translocation ofMRP1 contributes to multidrug resistance of mucoepidermoid carcinoma. OralOncol 2011;47:1134–40.
[17] Sherman IA, Fisher MM. Hepatic transport of fluorescent molecules: in vivostudies using intravital TV microscopy. Hepatology 1986;6:444–9.
[18] Martinez-Diez MC, Serrano MA, Monte MJ, Marin JJ. Comparison of the effectsof bile acids on cell viability and DNA synthesis by rat hepatocytes in primaryculture. Biochim Biophys Acta 2000;1500:153–60.
[19] Monte MJ, Martinez-Diez MC, El-Mir MY, Mendoza ME, Bravo P, Bachs O, et al.Changes in the pool of bile acids in hepatocyte nuclei during rat liverregeneration. J Hepatol 2002;36:534–42.
[20] Kaufmann SH, Shaper JH. A subset of non-histone nuclear proteins reversiblystabilized by the sulfhydryl cross-linking reagent tetrathionate. Polypeptidesof the internal nuclear matrix. Exp Cell Res 1984;155:477–95.
[21] Marin JJ, Bravo P, el-Mir MY, Serrano MA. ATP-dependent bile acid transportacross microvillous membrane of human term trophoblast. Am J Physiol1995;268:G685–94.
[22] Brangi M, Litman T, Ciotti M, Nishiyama K, Kohlhagen G, Takimoto C, et al.Camptothecin resistance: role of the ATP-binding cassette (ABC), mitoxan-trone-resistance half-transporter (MXR), and potential for glucuronidation inMXR-expressing cells. Cancer Res 1999;59:5938–46.
[23] Matunis MJ. Isolation and fractionation of rat liver nuclear envelopes andnuclear pore complexes. Methods 2006;39:277–83.
[24] Labarca C, Paigen K. A simple, rapid, and sensitive DNA assay procedure. AnalBiochem 1980;102:344–52.
[25] Dreger M, Bengtsson L, Schoneberg T, Otto H, Hucho F. Nuclear envelopeproteomics: novel integral membrane proteins of the inner nuclear mem-brane. Proc Natl Acad Sci USA 2001;98:11943–48.
[26] Tretter V, Altmann F, Marz L. Peptide-N4-(N-acetyl-beta-glucosaminyl)aspar-agine amidase F cannot release glycans with fucose attached alpha 1!3 to theasparagine-linked N-acetylglucosamine residue. Eur J Biochem 1991;199:647–52.
[27] Dechat T, Pfleghaar K, Sengupta K, Shimi T, Shumaker DK, Solimando L, et al.Nuclear lamins: major factors in the structural organization and function ofthe nucleus and chromatin. Genes Dev 2008;22:832–53.
[28] Courtois A, Payen L, Guillouzo A, Fardel O. Up-regulation of multidrug resis-tance-associated protein 2 (MRP2) expression in rat hepatocytes by dexa-methasone. FEBS Lett 1999;459:381–5.
[29] Trauner M, Arrese M, Soroka CJ, Ananthanarayanan M, Koeppel TA, SchlosserSF, et al. The rat canalicular conjugate export pump (Mrp2) is down-regulatedin intrahepatic and obstructive cholestasis. Gastroenterology 1997;113:255–64.
[30] Briz O, Macias RI, Vallejo M, Silva A, Serrano MA, Marin JJ. Usefulness ofliposomes loaded with cytostatic bile acid derivatives to circumvent chemo-therapy resistance of enterohepatic tumors. Mol Pharmacol 2003;63:742–50.
[31] Paulusma CC, Bosma PJ, Zaman GJ, Bakker CT, Otter M, Scheffer GL, et al.Congenital jaundice in rats with a mutation in a multidrug resistance-associ-ated protein gene. Science 1996;271:1126–8.
[32] Babakhanian K, Bendayan M, Bendayan R. Localization of P-glycoprotein at thenuclear envelope of rat brain cells. Biochem Biophys Res Commun 2007;361:301–6.
[33] Radominska-Pandya A, Pokrovskaya ID, Xu J, Little JM, Jude AR, Kurten RC, et al.Nuclear UDP-glucuronosyltransferases: identification of UGT2B7 and UGT1A6in human liver nuclear membranes. Arch Biochem Biophys 2002;399:37–48.
[34] Meech R, Mackenzie PI. Determinants of UDP glucuronosyltransferase mem-brane association and residency in the endoplasmic reticulum. Arch BiochemBiophys 1998;356:77–85.
[35] Jedlitschky G, Leier I, Buchholz U, Barnouin K, Kurz G, Keppler D. Transport ofglutathione, glucuronate, and sulfate conjugates by the MRP gene-encodedconjugate export pump. Cancer Res 1996;56:988–94.
[36] Takahashi T, Fujiwara Y, Yamakido M, Katoh O, Watanabe H, Mackenzie PI. Therole of glucuronidation in 7-ethyl-10-hydroxycamptothecin resistance invitro. Jpn J Cancer Res 1997;88:1211–7.
[37] Blanz J, Mewes K, Ehninger G, Proksch B, Greger B, Waidelich D, et al. Isolationand structure elucidation of urinary metabolites of mitoxantrone. Cancer Res1991;51:3427–33.
[38] Morris RG, Kotasek D, Paltridge G. Disposition of epirubicin and metaboliteswith repeated courses to cancer patients. Eur J Clin Pharmacol 1991;40:481–7.
[39] Gupta E, Lestingi TM, Mick R, Ramirez J, Vokes EE, Ratain MJ. Metabolic fate ofirinotecan in humans: correlation of glucuronidation with diarrhea. CancerRes 1994;54:3723–5.
[40] Draheim V, Reichel A, Weitschies W, Moenning U. N-glycosylation of ABCtransporters is associated with functional activity in sandwich-cultured rathepatocytes. Eur J Pharm Sci 2010;41:201–9.
[41] Feller N, Kuiper CM, Lankelma J, Ruhdal JK, Scheper RJ, Pinedo HM, et al.Functional detection of MDR1/P170 and MRP/P190-mediated multidrug re-sistance in tumour cells by flow cytometry. Br J Cancer 1995;72:543–9.