abai final poster may 27 6am - personal homepage...
TRANSCRIPT


Introduction
� Events with aversive function (AF) are those that organisms evade, avoid, or escape from (e.g., a bright light, loud noises, or electric shocks). Aversive Control (AC) is the study of how aversive contingencies regulate behaviour (e.g., negative reinforcement or punishment) 2, 4, 6, 7, 11, 15.
� Until recently, Electric Shock (ES) has been employed as the main aversive stimulus in AC studies13, 14, 9.
� Changes in ethical protocols and the Institutional Review Board’s guidelines regulating the use of ES, may explain the decline of AC research 3 .
� Preliminary research at the U. of Manitoba’s Fish Operant Behaviour Laboratory (UMFOBL) has confirmed, for the first time, the AF of Water Disturbance (WD) in the behaviour of Betta splendens, and has successfully developed techniques for studying AC phenomena in this species 8 .
� The exploration of a novel and less controversial form of AF is, therefore, merited. Moreover, the preliminary findings of WD’s AF calls for the exploration of the generality of behavioural phenomena that has been previously confirmed with the use of ES.
� B. splendens has typically been utilized in positive reinforcement studies involving aggressive display or social interactions. However, there is a lack of recent AC studies on this species 1, 5, 12 .

Statement of the problem
� Previous research at the UMFOBL was focused on identifying the AF of different forms of WD (i.e., Water Flows - WF and air bubbles). These studies did not provide direct evidence of AC learning in B. splendens.
� Consequently, the present study was intended to:
� Systematically replicate the findings of preliminary studies that provided evidence of the aversive function of WF. Accordingly, it was proposed to utilize a new subject, implement an improved experimental design, and introduce a new dependent variable to the analysis (i.e., switches).
� Gather direct evidence of learning in B. splendens with WF as a punishment.

Method
Subject
� One male B. splendens.
Apparatus
� A 40 x 40 x 20 experimental tank with 10 cm of water divided into two equal areas with an opening in the centre (3 x 10 cm)(see Figure 1). Water pumps located on each side of the tank were installed to create WF.
� A video tracking system17 recorded the position and amount of time the fish spent swimming in different areas of the tank. Specifically, the fish’s time allocation in pre-designated areas of the tank (i.e., disturbed and non-disturbed areas) and number of entries in those areas were recorded.
Water pumps Opening
Figure 1. Arrangement of the fish tank during the experiment (top and side view). Two equal areas with an opening in the center (3 x 10 cm). Water pumps located on each side of the tank were used to create WF.

Method
Design Conditions (A) Base Line (B) WF in the Right Side of the Tank (C) WF in the Left Side of the Tank (D) Punishment Sequence A-B-A-C-A-B-A-C-A-D-A
Procedure
� During the first nine treatments of the
study (i.e., sessions 1 to 99), the fish was exposed to 20-minute daily sessions. The last two phases (session 100 to 117) were 45 min each.
� Alternations between sides of the disturbance in the tank. In each session, continuous WF was presented on one
side of the tank, leaving the other area of the tank without disturbance. WD was continuously introduced until a stable state of preference was observed across sessions. WD was then alternated to the other side of the experimental tank.
� Punishment Condition (PC). Every time the fish entered the left area of the tank, the WF was presented. The presentation of the WF was constant and turned off only when the fish crossed back to the right side of the tank. The WF remained off when the fish remained on the right side of the tank.

Method
Variables/Measurements
Preference Ratio (PR) This measurement consisted of the difference between the amount of time the fish spent on the right and left side of the tank over the total session time:
PR = [Time on right side – Time on left side]/Session time If the difference of time allocated on the right and left side of the tank increased, the ratio tended to approach 1.0 or -1.0. A positive value of the ratio indicated a preference for the right side of the tank. A negative ratio indicated a preference for the left side. Conversely, a ratio near zero indicated no preference for either side of the tank. Switches This measurement consisted of the number of times the fish crossed from one side of the tank to the other during the session.

Results
Preference ratio (Figure 2a)
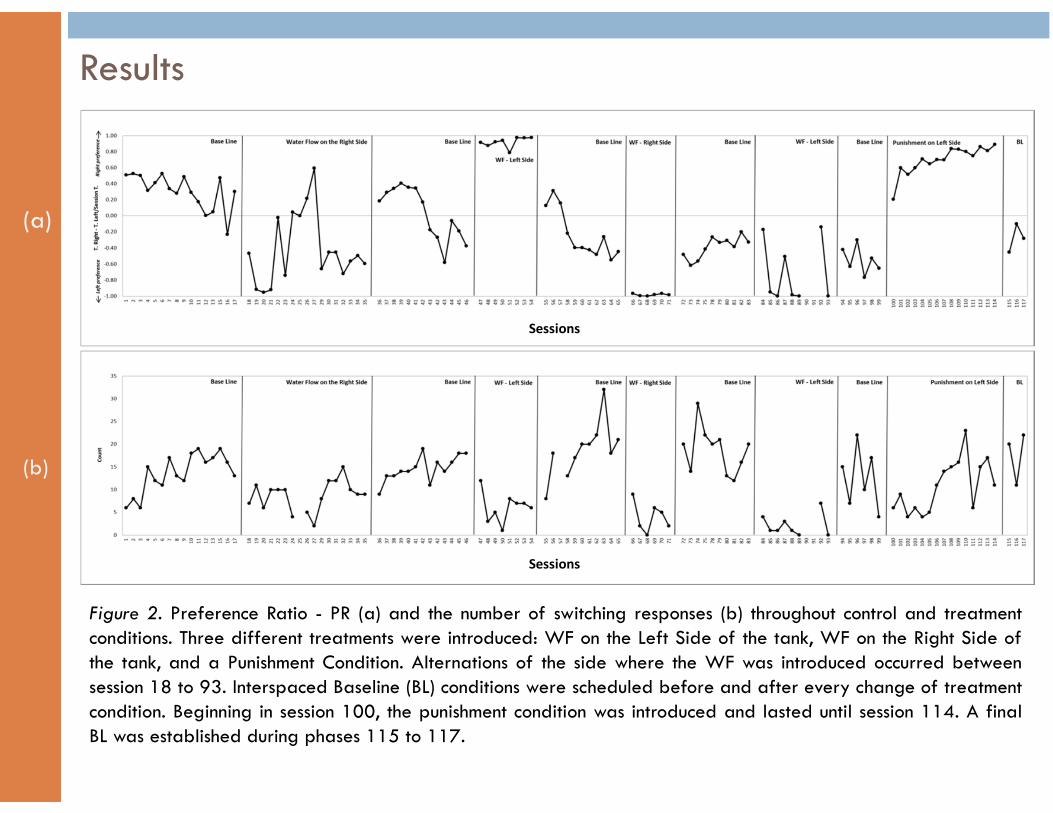
� Although the fish showed a clear preference for the non-disturbed (ND) side of the tank during the first sessions of the initial treatment (i.e., sessions 18 - 21), later sessions were characterized by an increase of the time the fish spent in the disturbed area (DA) (i.e., sessions 22 - 27) (see Figure 2a).
� This unexpected response pattern ceased drastically in session 29. During the remaining seven sessions of the first treatment (30 to 36) the fish’s PR remained between -0.6 and -0.8 (i.e., a preference for the ND-left side).
� During the two subsequent treatment conditions (session 47 to 54 and 66 to 71) the fish showed a clear preference for the corresponding ND area (PR =1.0 or -1.0).
� During the final scheduled alternation of the WF (i.e., sessions 84 to 93) the fish showed erratic PR scores, but overall, longer stances in the DA (i.e., left side). Moreover, this consistent preference for the left side of the tank was considered to be an opportunity to introduce the Punishment Contingency (PC).

Results
� Beginning in session 100, the PC was introduced. An increasing preference for the non-punished side of the tank (i.e., right side) was observed across the 14 sessions of this phase. The final PR values reached 0.9 and 1.0 during the last sessions of this treatment. A final baseline (i.e., sessions 111 - 117) showed the immediate fish’s return to a preference for the left side when the aversive contingency was removed.
Switches (Figure 2b)
� Having compared the number of switches displayed during the alternating WF treatments versus the BL conditions, it is clear that the fish displayed a higher rate of switches during BL.
� A different pattern was observed during the PC (sessions 100 to 114). During this condition the fish displayed a similar rate of switching responses to the rate observed during BL (see Figure 2b).
Results
(a)
(b)
Sessions
Sessions
Figure 2. Preference Ratio - PR (a) and the number of switching responses (b) throughout control and treatment conditions. Three different treatments were introduced: WF on the Left Side of the tank, WF on the Right Side of the tank, and a Punishment Condition. Alternations of the side where the WF was introduced occurred between session 18 to 93. Interspaced Baseline (BL) conditions were scheduled before and after every change of treatment condition. Beginning in session 100, the punishment condition was introduced and lasted until session 114. A final BL was established during phases 115 to 117.

Discussion � The aversive function of WF was
systematically replicated (i.e., different design and B. splendens subject).
� Regarding the unexpected high time allocation of the fish in the DA, the session videos suggested that this behaviour may be explained by the presence of uncontrolled weak WD areas in the tank.
� New evidence of AC in B. splendens
(i.e. punishment) was found. The fish showed a dramatic reduction in the time allocation as a result of the contingent presentations of WF.
� The introduction of a new measure (i.e. switches) provided interesting dissociations. Whereas the time allocation on the target side of the tank was reduced, the visits to this
same area were not suppressed. Conversely, the switching response decreased during the continuous WF conditions and increased every time the BL were re-established.
� The dissociations between time allocation and switching responses suggest a differential sensibility of the temporal and repeatability dimensions 10 of B. splendens
responding under a PC.
� Presently, a research project at the UMFOBL aims to replicate the observed PC effects. Additionally, an adaptation of a Sidman-avoidance procedure and non-contingent presentations of WF will be tested in this project.

Conclusions
� The recently developed model for studying B. splendens at the UMFOBL represents an alternative preparation for investigating AC. Its value lies in the exclusion of painful/harmful stimuli, the ecological relevance with respects to the aversive function of the WD, along with low costs and easy maintenance of the experimental subjects.
� The AC model with B. splendens may be implemented in different research programs, such as the replication of behavioural phenomena observed with different forms of aversive stimulation. Furthermore, given the controversy regarding the aversive effect observed when increases in required response effort (or force) have been implemented16, 18, the WD model may contribute to this debate because the introduction of WF can be interpreted as an increase in the force requirement for swimming.

References 1. Adler, N., & Hogan, J. A. (1963). Classical conditioning and punishment of an instinctive response in Betta splendens. Animal Behaviour, 11(2-3), 351-
354.
2. Catania, A. C. (1998). Learning (4th ed.). Upper Saddle River, NJ: Prentice Hall.
3. Catania, A. C. (2007). The journal of the experimental analysis of behavior at zero, fifty, and one hundred. The Journal of the Experimental Analysis of
Behavior, 89(1), 111–118.
4. Crosbie, J. (1998). Negative reinforcement and punishment. In K. A. Lattal & M. Perone, Handbook of research methods in human operant behaviour (pp.
163-189). New York: Plenum Press.
5. Fantino, E., Weigele, S., & Lancy, D. (1972). Aggressive display in the siamese fighting fish (Betta splendens). Learning and Motivation, 3(4), 457-468.
6. Hineline, P. N. (1977). Negative reinforcement and avoidance. In Werner K. Honig, J. E. R. Staddon, Handbook of operant behavior (pp. 364-414).
Englewood Cliffs, N.J.: Prentice-Hall.
7. Hineline, P. N. (1984). Aversive control: a separate domain. Journal of the Experimental Analysis of Behavior, 42(3), 495-509.
8. Hurtado-Parrado, H. C., Pear, J., & Froese, K. (2009). Water disturbance as a form of aversive stimulation for The Siamese Fighting Fish (Betta splendens).
Poster presented at the International Conference of the Association for Behavior Analysis, Oslo, Norway (August 7-9, 2009).
9. Hurtado-Parrado, H. C. (2010 – in press). Neuronal mechanisms of learning in teleost fish. Universitas Psychologica, 9, xxx-xxx.
10. Johnston J. M. & Pennypacker, H. S. (2009). Strategies and tactics of behavioral research. New York: Lawrence Erlbaum Associates. 11. Malott, R. (2008). Principles of behaviour. Upper Saddle River, NJ: Prentice Hall.
12. Melvin, K. B., & Ervey, D. H. (1973). Facilitative and suppressive effects of punishment on species-typical aggressive display in Betta splendens. Journal
of Comparative and Physiological Psychology, 83(3), 451-457.
13. Overmier, J. B., & Hollis, K. L. (1990). Fish in the think tank: Learning, memory and integrated behaviour. In R. P. Kesner & D. S. Olson (Eds). Neurobiology of Comparative Cognition, (pp 205-236). Hillsdale, NJ: Erlbaum.
14. Pear, J. J. (2001). The science of learning. Philadelphia, PA: Psychology Press.
15. Pierce, W. D. & Cheney, C. D. (2004). Behavior analysis and learning. Mahwah, N. J.: L. Erlbaum Associates.
16. Friman, P. C., & Poling, A. (1995). Making life easier with effort: Basic findings and applied research on response effort. Journal of Applied Behavior
Analysis, 28(4), 583-590.
17. Silva, F., & Pear, J. (1995). Stereotypy of spatial movements during noncontingent and contingent reinforcement. Animal Learning & Behavior, 23(3),
245-255.
18. Zarcone, T. J., Chen, R., & Fowler, S. C. (2007). Effects of differing response-force requirements on food-maintained responding in CD-1 mice. Journal of
the Experimental Analysis of Behavior, 88(3), 381-393.
* Contact information Camilo Hurtado-Parrado
Ph.D. Student
University of Manitoba, Canada.
E-mail: [email protected]