aba and sugar interactions regulating development: cross-talk or voices in a crowd?
TRANSCRIPT

26
Plant growth and development are controlled by the concertedaction of many signaling pathways that integrate information fromenvironmental signals with that from developmental and metaboliccues. Physiological studies have demonstrated that abscisic acidand sugars have both similar and antagonistic effects on diverseprocesses, including seed development, germination, and seedlinggrowth. Recent genetic studies have identified several loci thatare involved in both sugar and hormonal responses. It is rarelyclear whether these apparent linkages reflect direct or indirectinteractions between sugar and hormone signaling pathways,but the identification of gene products that are encoded at theseloci is allowing these possibilities to be tested.
Addresses*Department of Molecular, Cellular and Developmental Biology,University of California at Santa Barbara, Santa Barbara, California 93106,USA; e-mail: [email protected]†Department of Biochemistry and Cell Biology — MS140, Rice University,6100 Main Street, Houston, Texas 77005, USA;e-mail: [email protected]
Current Opinion in Plant Biology 2001, 5:26–32
1369-5266/01/$ — see front matter© 2001 Elsevier Science Ltd. All rights reserved.
AbbreviationsABA abscisic acidABI4 ABA INSENSITIVE4ctr1 constitutive triple response1DET1 DE-ETIOLATED1fus3 fusca3GA gibberellic acidGARE GA-response elementgin6 glucose insensitive6isi3 impaired sucrose induction3lec1 leafy cotyledon1 mig mannose-insensitive germinationprl1 pleiotropic regulatory locus1Ramy1A rice α-amylase gene1Asis5 sugar insensitive5sun6 sucrose uncoupled6
IntroductionThe autotrophic growth of plants depends on their abilityto produce sugars photosynthetically, then transport, consumeor store sugars and their derivatives within the plant. Becausesugars are important as sources of energy and carbon-skeletons, access to sugars is an important determinant of aplant’s potential for growth. Consequently, plants have avariety of mechanisms to regulate the production of andaccess to sugars in response to assorted environmental factors(e.g. light intensity), developmental signals (e.g. phytohor-mones or stage-specific regulators), and metabolic status(e.g. C:N balance). In recent years, it has become clear thatmany of these regulatory mechanisms interact to form asignaling web (reviewed in [1,2•–5•]).
Deciphering the relationships among the pathways thatcomprise this web will require a more complete under-standing of the relevant signals, of their sensors andsignaling intermediates, and of the processes affected bythose signals. Although sugars were initially assumed tocontrol growth via strictly metabolic effects, hexose- and/orsucrose-specific signaling mechanisms have also beenshown to regulate gene expression [2•,6]. Recent studiesshow that the disaccharide trehalose, or its biosyntheticintermediates, may also be involved in sugar sensing [7,8].A variety of proteins that are involved in photosynthesisand in carbon metabolism, transport and uptake are encodedby sugar-regulated genes, providing opportunities forfeedback regulation of carbon fixation or use. In somecases, one or more isoforms of an enzyme may be inducedby high sugar concentrations (i.e. ‘feast’ conditions) insome tissues, whereas different isoform(s) are inducedby low sugar concentrations (i.e. ‘famine’ conditions) inother tissues [6].
The characterization of sugar-response pathways is complexbecause plants appear to respond to sugars via multiplepathways, and the identities of the molecules that triggersugar responses are not clear [2•–4•]. Also, some experimentssuggest that sugar-response pathways may respond to therate of flux through a particular pathway, rather than to theabsolute level of a sugar or sugar metabolite. Furthermore, itis possible that sugars modulate some processes via mech-anisms that do not involve traditional response pathways.For example, glucose alters the activities of some proteinsfrom diverse organisms via glycation, the non-enzymaticaddition of glucose moieties [9].
Sugar signaling systems have been well-characterized inbacteria, yeast and animals. Some elements of thesepathways — for example, hexokinases, SUCROSE NON-FERMENTING1 (SNF1)-related protein kinases,phosphatases, and secondary messengers such as Ca2+ — areevolutionarily conserved. The roles of these conservedfactors in sugar signaling in plants are being tested bybiochemical and reverse-genetic experiments. Suchstudies have shown that although yeast and Arabidopsishexokinases have functionally similar catalytic activities,overexpression of these proteins in plants produces effectson sugar sensitivity that are opposite to those in yeast,indicating that their regulatory functions are not conserved.Similarly, although genes resembling those encoding yeasttransporter-like sensors have been identified in plants,their roles in sugar sensing remain questionable. Ourcurrent understanding of plant sugar-sensing and signalingmechanisms has been discussed extensively in severalrecent reviews [2•–4•].
ABA and sugar interactions regulating development: cross-talkor voices in a crowd?Ruth R Finkelstein* and Susan I Gibson†

ABA and sugar interactions regulating development Finkelstein and Gibson 27
Additional putative sugar-response components have beenidentified using genetic screens that selected for eitheraltered growth on high concentrations of sugar (sucrose,glucose or mannose) or alterations in sugar-regulated geneexpression [2•–5•]. These screens identified a largenumber of sugar-response loci, mutations in many of whichalso cause defects in hormone synthesis or response. Inparticular, new alleles of two loci that are required forabscisic acid (ABA) biosynthesis, and one locus involvedin ABA response (ABA INSENSITIVE4 [ABI4]), havebeen isolated on the basis of strong their sugar-resistantphenotypes. In addition, mutants have been isolated thathave defects in ethylene response (i.e. new constitutivetriple response1 [ctr1] alleles) or pleiotropic hypersensitivityto sugars, ABA, ethylene, auxin, cytokinin and cold(i.e. pleiotropic regulatory locus1 [prl1]). The prl1 mutantdiffers from the others in that its responses to sugars,ABA and ethylene are similarly affected. In other mutants,decreased sugar/ABA response is often accompanied byincreased ethylene synthesis or response. Given that thesephytohormones are unique to plants, and that the ABI4protein is a plant-specific transcription factor with no closehomologs [10•], such studies are likely to uncover novelaspects of sugar signaling in plants.
The term ‘cross-talk’ is frequently used to refer to inter-actions among signaling pathways. However, it may beover-used in the absence of evidence for either direct orindirect interactions among the pathways affecting a givenprocess. Evidence of direct interaction might include theidentification of shared signaling intermediates, of alteredhormone levels or sensitivity mediating the response toanother factor, of direct protein–protein interactionsbetween components of different pathways, or of theregulation of the activity of a component of one pathwayby a component of another pathway. This review attemptsto discriminate between the direct and indirect interactionsof the sugar and abscisic acid response pathways thatcontrol three developmental processes: seed development,germination, and seedling growth.
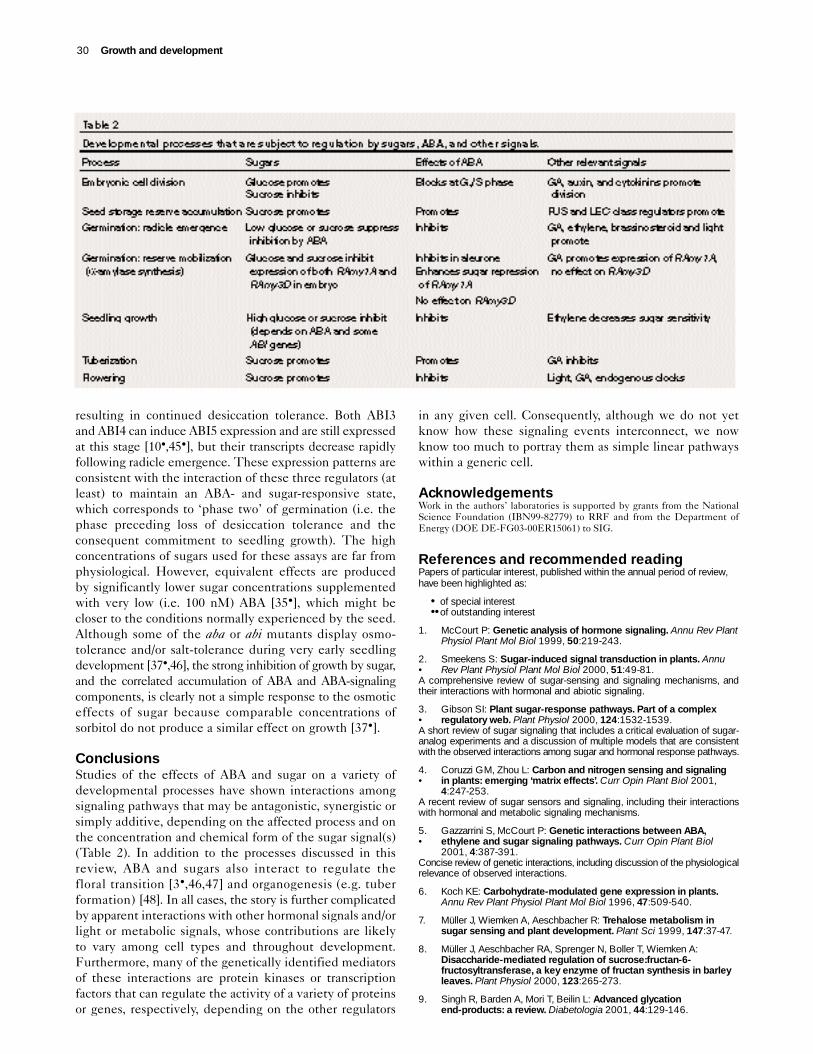
Seed developmentDeveloping embryos undergo a transition from growth bycell division to growth by cell enlargement as they begin toaccumulate storage reserves. Both sugars and ABA havebeen implicated in the control of this transition (reviewedin [11,12]). Early in embryo development, invertase levelsare high, promoting a high ratio of hexoses to sucrose. Asdevelopment progresses, invertase activity decreases whilethe activities of sucrose-phosphate synthase and sucrosesynthase increase, as does the sucrose:hexose ratio. Inlegumes, the reduction in glucose concentration spreadsacross the cotyledons in a wave, and correlates withdecreased mitotic activity and increased starch accumulation[11,12]. At the same time, the levels of ABA in the seedincrease and are correlated with cell cycle arrest at theG1→S transition (reviewed in [10•]). Presumably, ABAinduces the expression of ICK1, a cyclin-dependent kinase
inhibitor that decreases Cdc2a-like activity [10•].Manipulation of soluble sugar levels in cotyledons, eitherin culture experiments or by transgenic expression of ayeast-derived invertase, has shown that glucose promotescell division. In contrast, increased sucrose concentration iscorrelated with increased cell expansion and accumulationof storage reserves [11]. Although sugar levels have alsobeen shown to affect cyclin activity [13,14•], both glucoseand sucrose promote cyclin expression, leaving open thequestion of how glucose and sucrose have opposite effectson mitotic activity in seeds.
Increased ABA levels in mid-embryogenesis also promotethe accumulation of storage reserves, as shown by embryoculture studies and analysis of ABA-response mutants ortransgenic seeds expressing anti-ABA antibodies (reviewedin [10•,12,15]). Although there is no evidence of a directinteraction between the antagonistic effects of glucose andABA on the cell cycle, the similar effects of sucrose andABA on reserve accumulation appear to be linked. Thephaseolin promoter is induced by ABA; this response isenhanced by the combination of sucrose and CaCl2 but,surprisingly, inhibited by sucrose alone [12]. Similarly, recentstudies of sugar-induced gene expression in Arabidopsisleaves have shown that ABA greatly enhances the sucrose-induction of two genes that are involved in starchbiosynthesis, ADP-glucose pyrophosphorylase (subunitApL3) and starch-branching enzyme Sbe2.2, despitehaving no effect on their expression in the absence ofsucrose [16•]. Furthermore, the seeds of severe abi3 mutantslack any significant storage protein or lipid accumulation,but accumulate 3–6-fold more sucrose than wildtype seedsduring the reserve accumulation phase of development [17].These findings indicate that high sucrose is not sufficientto promote reserve accumulation in the absence of afunctional ABA signaling mechanism.
In legumes, the accumulation of starch is generally coupledto the accumulation of storage protein [11]. However, twotypes of exception to this rule have been described. First, inVicia narbonensis, reduction of starch synthesis due toantisense expression of ADP-glucose pyrophosphorylase isaccompanied by increased total protein accumulation,possibly reflecting an increased period of seed-filling ordelayed maturation [18]. Second, the leafy-cotyledon classof Arabidopsis mutants (i.e. fusca3 [fus3], leafy cotyledon1 [lec1],and lec2) have excessive starch accumulation but almostno storage protein or lipid accumulation [19]. It is not yetknown whether soluble sugar metabolism or signaling isaltered in these mutants. The FUS3 and LEC genes arecentral regulators of embryogenesis that interact geneticallywith ABA-dependent regulatory system(s) to suppressvivipary at mid-embryogenesis [20], and with several of theABI loci to mediate ABA sensitivity at seed maturity [10•].The DE-ETIOLATED1 (DET1)/FUS2, FUS3 and LEC1 lociare also negative regulators of some light responses that areenhanced by sugars, for example anthocyanin accumulation,such that the (de-repressed) mutant seeds are abnormally

28 Growth and development
pigmented [10•]. This effect is greatly enhanced by some ofthe abi mutations. Additional interactions between ABA andlight signaling during seed development are reflected inthe dependence on DET1, FUS3 and LEC1 for normalaccumulation of ABI3 protein [10•]. Taken together, theseresults suggest that the transcription factors encoded by theABI3, ABI4, ABI5, DET1, FUS3, and LEC1 loci [10•] may allparticipate in integrating ABA, light, and possibly sugarsignaling during embryogenesis.
The disaccharide trehalose may also be involved in theregulation of embryo maturation. Although trehalose doesnot accumulate significantly in higher plants, except inhighly desiccation tolerant species, recent studies showthat trehalose biosynthetic enzymes are present in diversespecies and can both regulate carbohydrate allocation andinteract with sugar-sensing mechanisms [7]. A recentlyisolated Arabidopsis mutant that is defective in the firststep of trehalose synthesis, trehalose-6-phosphate synthase1(tps1), is an embryo lethal (PJ Eastmond et al., Abstract 130,12th International Conference on Arabidopsis Research,23–27 June 2001, Madison, WI). Although tps1 embryomorphogenesis is normal, the development of this mutantis retarded and eventually arrests early in the phase of cellexpansion and storage-reserve accumulation, suggesting thattrehalose metabolism is essential during embryo maturation.It will be interesting to learn whether there is any interactionbetween trehalose and ABA signaling.
Germination and seedling growthHormonal and environmental controls that regulate germi-nation have been extensively characterized with respect totheir effects on visible germination, i.e. radicle emergence,and on changes in gene expression associated withmobilization of storage reserves (especially in cerealgrains). These studies have shown that light, pre-chilling,gibberellic acid (GA), ethylene, and brassinosteroidspromote germination in many species, whereas ABAinhibits germination. Sugars appear to interact with thehormonal effects in various ways, depending on theconcentration of sugar and the response assayed.
During seed germination, the mobilization of storagereserves must be tightly controlled to ensure an adequatesupply of energy and carbon skeletons to the developingseedling. In germinating cereal seeds, the expression of keygenes involved in the mobilization of energy reserves such asstarch appears to be regulated by interactions among sugar,ABA and gibberellin response pathways [21]. Cerealα-amylases are encoded by multigene families that aresubject to differential spatial, temporal, hormonal andmetabolic regulation that affects expression at multiplelevels including transcription, mRNA stability, and secretion(reviewed in [21,22]). For example, gibberellins promoteα-amylase expression in both the aleurone and the scutellarepithelium tissues, but sugars repress GA-induced expressiononly in the scutellum [23]. This sugar-repression cannot becounteracted by additional GA or by the constitutive GA
response conferred by the slender (sln) mutant, suggesting thatsugars act downstream of GA in a SLN signaling pathway [23].In contrast, ABA represses α-amylase expression in bothaleurone and embryonic tissues, but this can be partiallyreversed by additional GA, implying that ABA and sugars actat different signaling steps or via different pathways [23].
The GA-response element (GARE) is required for GA-induction by the transcription factor GAMyb and is alsopartially involved in repression by both sugar and ABA[24–26]. Comparison of the promoter regions of theembryo-expressed α-amylase genes has shown, however,that the GARE is present in only some α-amylase genes,for example, the rice α-amylase gene1A (RAmy1A) [21]. Inaleurone, ABA repression is at least partially mediated byan ABA-induced protein kinase (PKABA1) that leads todecreased GAMyb expression [27•]; it is not clear whetherthis mechanism is functional in embryos. In isolatedscutellar tissues, a potent degradation-resistant ABAanalog ([+]-8′,8′,8′-trifluoroabscisic acid) slightly repressedRAmy1A expression and slightly enhanced the repressiveeffect of glucose [28]. Furthermore, this analog stimulatedglucose uptake and sucrose synthesis in scutellar tissues,suggesting that the repressive effects of ABA on α-amylaseexpression in embryos [23] might be potentiated byABA-induced increases in sugar concentrations. In contrast,RAmy3D is induced by starvation early in germination,lacks the GARE sequence and does not respond to GA orABA [21]. In addition, sugar-repression of RAmy3D is notaccompanied by a decrease in GAMyb expression [29•],indicating that alternate mechanisms of repression areinvolved. Surprisingly, treatment with glucose (25–90 mM)decreases endogenous ABA and suppresses ABA-inducibleexpression of the dehydrin Rab16A [23,30•], demonstratingthat some of the effects of ABA and glucose in embryo tissues are antagonistic. Sugar repression of both RAmy3D andRab16A involves decreases in both transcription and mRNAstability [30•,31•]. At least two signaling pathways appearto mediate sugar-repression of α-amylase expression: glucosedestabilizes the transcripts whereas several non-metabolizeddisaccharides repress the GA-induced accumulation ofα-amylase mRNA without affecting its stability [31•].
Antagonistic effects of sugars and ABA have also beenobserved in germinating Arabidopsis seeds that were exposedto low concentrations of sugars (30–90 mM) [32,33•].These seeds are able to complete radicle emergence in thepresence of up to 100-fold more ABA than is tolerated byseeds germinating on glucose-free medium. Nevertheless,upon completion of radicle emergence, seedlings grownon ABA supplemented with 30–90 mM sugar arrest andaccumulate anthocyanins [33•], as do seedlings germinatedon high sugar (>300 mM) but no ABA. It has been suggestedthat the exogenous sugar permits germination by supplyingenergy and nutrients, thereby relieving a metabolic blockimposed by ABA [32]. This view is consistent with thewell-characterized inhibition of reserve mobilization byexogenous ABA in both starch-storing seeds and some
oilseeds [10•]. However, recent studies have shown thatthe glyoxylate cycle is not essential for germination, andpost-germination growth can be supported by either photo-synthesis or exogenous sugar in the absence of a functionalglyoxylate cycle [34•]. Furthermore, when exposed to ABA,mature Arabidopsis seeds express genes encoding enzymesthat are required for lipid breakdown (SL Pritchard et al.,Abstract 149, 12th International Conference on ArabidopsisResearch, 23–27 June 2001, Madison, WI) despite showingno visible signs of germination. Although the optimalconcentration of glucose required for ABA-resistant seedgermination was too low to be consistent with a purelynutritional effect [33•], studies with glucose analogs didnot provide clear evidence of glucose signaling in thisresponse (RR Finkelstein, TJ Lynch, unpublished data).
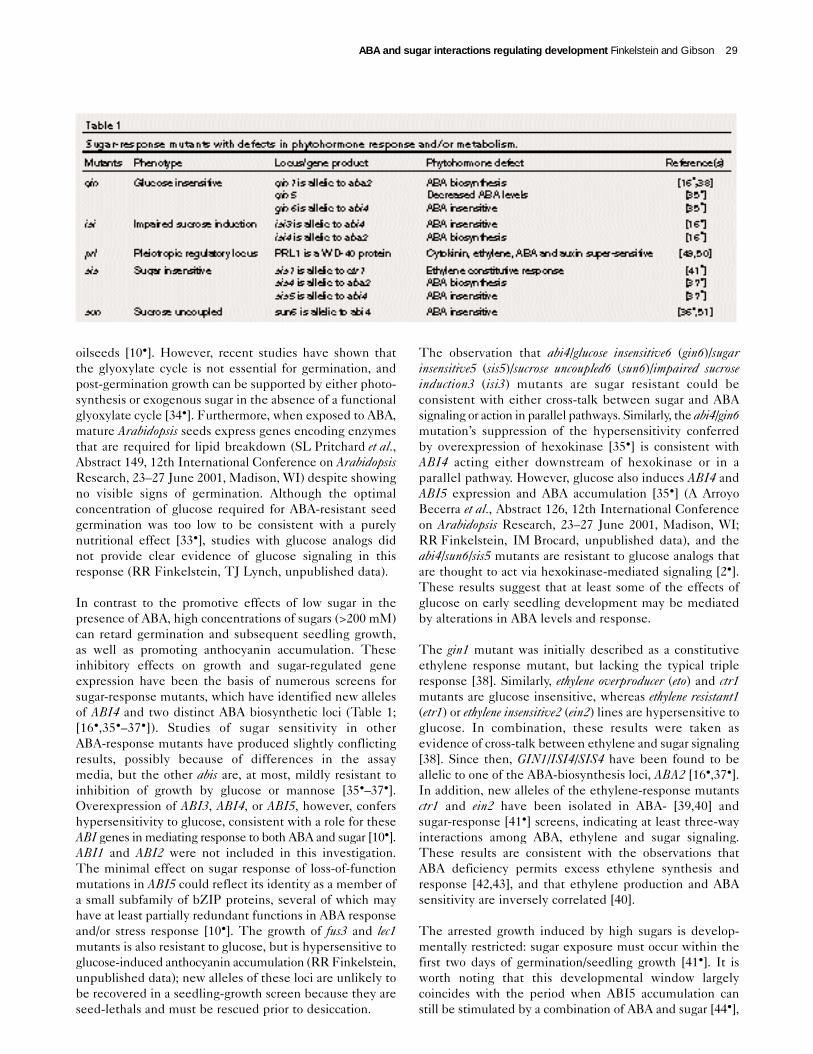
In contrast to the promotive effects of low sugar in thepresence of ABA, high concentrations of sugars (>200 mM)can retard germination and subsequent seedling growth,as well as promoting anthocyanin accumulation. Theseinhibitory effects on growth and sugar-regulated geneexpression have been the basis of numerous screens forsugar-response mutants, which have identified new allelesof ABI4 and two distinct ABA biosynthetic loci (Table 1;[16•,35•–37•]). Studies of sugar sensitivity in otherABA-response mutants have produced slightly conflictingresults, possibly because of differences in the assaymedia, but the other abis are, at most, mildly resistant toinhibition of growth by glucose or mannose [35•–37•].Overexpression of ABI3, ABI4, or ABI5, however, confershypersensitivity to glucose, consistent with a role for theseABI genes in mediating response to both ABA and sugar [10•].ABI1 and ABI2 were not included in this investigation.The minimal effect on sugar response of loss-of-functionmutations in ABI5 could reflect its identity as a member ofa small subfamily of bZIP proteins, several of which mayhave at least partially redundant functions in ABA responseand/or stress response [10•]. The growth of fus3 and lec1mutants is also resistant to glucose, but is hypersensitive toglucose-induced anthocyanin accumulation (RR Finkelstein,unpublished data); new alleles of these loci are unlikely tobe recovered in a seedling-growth screen because they areseed-lethals and must be rescued prior to desiccation.
The observation that abi4/glucose insensitive6 (gin6)/sugarinsensitive5 (sis5)/sucrose uncoupled6 (sun6)/impaired sucroseinduction3 (isi3) mutants are sugar resistant could beconsistent with either cross-talk between sugar and ABAsignaling or action in parallel pathways. Similarly, the abi4/gin6mutation’s suppression of the hypersensitivity conferredby overexpression of hexokinase [35•] is consistent withABI4 acting either downstream of hexokinase or in aparallel pathway. However, glucose also induces ABI4 andABI5 expression and ABA accumulation [35•] (A ArroyoBecerra et al., Abstract 126, 12th International Conferenceon Arabidopsis Research, 23–27 June 2001, Madison, WI;RR Finkelstein, IM Brocard, unpublished data), and theabi4/sun6/sis5 mutants are resistant to glucose analogs thatare thought to act via hexokinase-mediated signaling [2•].These results suggest that at least some of the effects ofglucose on early seedling development may be mediatedby alterations in ABA levels and response.
The gin1 mutant was initially described as a constitutiveethylene response mutant, but lacking the typical tripleresponse [38]. Similarly, ethylene overproducer (eto) and ctr1mutants are glucose insensitive, whereas ethylene resistant1(etr1) or ethylene insensitive2 (ein2) lines are hypersensitive toglucose. In combination, these results were taken asevidence of cross-talk between ethylene and sugar signaling[38]. Since then, GIN1/ISI4/SIS4 have been found to beallelic to one of the ABA-biosynthesis loci, ABA2 [16•,37•].In addition, new alleles of the ethylene-response mutantsctr1 and ein2 have been isolated in ABA- [39,40] andsugar-response [41•] screens, indicating at least three-wayinteractions among ABA, ethylene and sugar signaling.These results are consistent with the observations thatABA deficiency permits excess ethylene synthesis andresponse [42,43], and that ethylene production and ABAsensitivity are inversely correlated [40].
The arrested growth induced by high sugars is develop-mentally restricted: sugar exposure must occur within thefirst two days of germination/seedling growth [41•]. It isworth noting that this developmental window largelycoincides with the period when ABI5 accumulation canstill be stimulated by a combination of ABA and sugar [44•],
ABA and sugar interactions regulating development Finkelstein and Gibson 29
resulting in continued desiccation tolerance. Both ABI3and ABI4 can induce ABI5 expression and are still expressedat this stage [10•,45•], but their transcripts decrease rapidlyfollowing radicle emergence. These expression patterns areconsistent with the interaction of these three regulators (atleast) to maintain an ABA- and sugar-responsive state,which corresponds to ‘phase two’ of germination (i.e. thephase preceding loss of desiccation tolerance and theconsequent commitment to seedling growth). The highconcentrations of sugars used for these assays are far fromphysiological. However, equivalent effects are producedby significantly lower sugar concentrations supplementedwith very low (i.e. 100 nM) ABA [35•], which might becloser to the conditions normally experienced by the seed.Although some of the aba or abi mutants display osmo-tolerance and/or salt-tolerance during very early seedlingdevelopment [37•,46], the strong inhibition of growth by sugar,and the correlated accumulation of ABA and ABA-signalingcomponents, is clearly not a simple response to the osmoticeffects of sugar because comparable concentrations ofsorbitol do not produce a similar effect on growth [37•].
ConclusionsStudies of the effects of ABA and sugar on a variety ofdevelopmental processes have shown interactions amongsignaling pathways that may be antagonistic, synergistic orsimply additive, depending on the affected process and onthe concentration and chemical form of the sugar signal(s)(Table 2). In addition to the processes discussed in thisreview, ABA and sugars also interact to regulate thefloral transition [3•,46,47] and organogenesis (e.g. tuberformation) [48]. In all cases, the story is further complicatedby apparent interactions with other hormonal signals and/orlight or metabolic signals, whose contributions are likelyto vary among cell types and throughout development.Furthermore, many of the genetically identified mediatorsof these interactions are protein kinases or transcriptionfactors that can regulate the activity of a variety of proteinsor genes, respectively, depending on the other regulators
in any given cell. Consequently, although we do not yetknow how these signaling events interconnect, we nowknow too much to portray them as simple linear pathwayswithin a generic cell.
AcknowledgementsWork in the authors’ laboratories is supported by grants from the NationalScience Foundation (IBN99-82779) to RRF and from the Department ofEnergy (DOE DE-FG03-00ER15061) to SIG.
References and recommended readingPapers of particular interest, published within the annual period of review,have been highlighted as:
• of special interest••of outstanding interest
1. McCourt P: Genetic analysis of hormone signaling. Annu Rev PlantPhysiol Plant Mol Biol 1999, 50:219-243.
2. Smeekens S: Sugar-induced signal transduction in plants. Annu • Rev Plant Physiol Plant Mol Biol 2000, 51:49-81.A comprehensive review of sugar-sensing and signaling mechanisms, andtheir interactions with hormonal and abiotic signaling.
3. Gibson SI: Plant sugar-response pathways. Part of a complex • regulatory web. Plant Physiol 2000, 124:1532-1539.A short review of sugar signaling that includes a critical evaluation of sugar-analog experiments and a discussion of multiple models that are consistentwith the observed interactions among sugar and hormonal response pathways.
4. Coruzzi GM, Zhou L: Carbon and nitrogen sensing and signaling • in plants: emerging ‘matrix effects’. Curr Opin Plant Biol 2001,
4:247-253.A recent review of sugar sensors and signaling, including their interactionswith hormonal and metabolic signaling mechanisms.
5. Gazzarrini S, McCourt P: Genetic interactions between ABA, • ethylene and sugar signaling pathways. Curr Opin Plant Biol
2001, 4:387-391.Concise review of genetic interactions, including discussion of the physiologicalrelevance of observed interactions.
6. Koch KE: Carbohydrate-modulated gene expression in plants.Annu Rev Plant Physiol Plant Mol Biol 1996, 47:509-540.
7. Müller J, Wiemken A, Aeschbacher R: Trehalose metabolism insugar sensing and plant development. Plant Sci 1999, 147:37-47.
8. Müller J, Aeschbacher RA, Sprenger N, Boller T, Wiemken A:Disaccharide-mediated regulation of sucrose:fructan-6-fructosyltransferase, a key enzyme of fructan synthesis in barleyleaves. Plant Physiol 2000, 123:265-273.
9. Singh R, Barden A, Mori T, Beilin L: Advanced glycationend-products: a review. Diabetologia 2001, 44:129-146.
30 Growth and development

10. Finkelstein RR, Gampala SSL, Rock CD: Abscisic acid signaling in • seeds. Plant Cell 2002, in press. A recent review of ABA signaling that emphasizes genetic and molecularinteractions among ABA-response loci, developmental regulators, and othersignaling components. The discussion includes a demonstration of sugarhypersensitivity in ABI-overexpression lines.
11. Wobus U, Weber H: Sugars as signal molecules in plant seeddevelopment. Biol Chem 1999, 380:937-944.
12. Wobus U, Weber H: Seed maturation: genetic programmes andcontrol signals. Curr Opin Plant Biol 1999, 2:33-38.
13. Soni R, Carmichael JP, Shah ZH, Murray JAH: A family of cyclin Dhomologs from plants differentially controlled by growthregulators and containing the conserved retinoblastoma proteininteraction motif. Plant Cell 1995, 7:85-103.
14. Riou-Khamlichi C, Menges M, Healy JMS, Murray JAH: Sugar control • of the plant cell cycle: differential regulation of Arabidopsis D-type
cyclin gene expression. Mol Cell Biol 2000, 20:4513-4521.The authors demonstrate that the expression of cyclins D2 and D3 is notdependent on growth phase but is correlated with sucrose availability, providinga mechanism for sucrose participation in cell cycle control. Cytokinin treatmentenhances sucrose-induction of CycD3 expression but is ineffective in theabsence of sucrose.
15. Rock C, Quatrano R: The role of hormones during seeddevelopment. In Plant Hormones: Physiology, Biochemistry andMolecular Biology, edn 2. Edited by Davies PJ. Dordrecht: KluwerAcademic Publishers; 1995:671-697.
16. Rook F, Corke F, Card R, Munz G, Smith C, Bevan MW: Impaired • sucrose-induction mutants reveal the modulation of
sugar-induced starch biosynthetic gene expression by abscisicacid signalling. Plant J 2001, 26:421-433.
A negative selection marker was used to isolate mutants that are defectivein sucrose-induced gene expression. Two of the loci identified were ABAbiosynthesis and response loci that had been previously identified insugar-resistant seedling growth screens: isi3/abi4/sis5/gin6/sun6 andisi4/aba2/gin1/sis4. This study demonstrated that the apparent defect inethylene signaling described for the gin1 mutants was caused by a defect inABA biosynthesis. ABA enhanced the sucrose-induced expression of somegenes, suggesting that ABA can potentiate some sugar responses.
17. Ooms JJJ, Léon-Kloosterziel KM, Bartels D, Koornneef M,Karssen CM: Acquisition of desiccation tolerance and longevity inseeds of Arabidopsis thaliana. Plant Physiol 1993, 102:1185-1191.
18. Weber H, Rolletschek H, Heim U, Golombek S, Gubatz S, Wobus U:Antisense-inhibition of ADP-glucose pyrophosphorylase indeveloping seeds of Vicia narbonensis moderately decreasesstarch but increases protein content and affects seed maturation.Plant J 2000, 24:33-43.
19. Harada J: Role of Arabidopsis LEAFY COTYLEDON genes in seeddevelopment. J Plant Physiol 2001, 158:405-409.
20. Raz V, Bergervoet J, Koornneef M: Sequential steps for developmentalarrest in Arabidopsis seeds. Development 2001, 128:243-252.
21. Thomas BR, Rodriguez RL: Metabolite signals regulate geneexpression and source/sink relations in cereal seedlings. PlantPhysiol 1994, 106:1235-1239.
22. Jacobsen JV: Gibberellin action in germinated cereal grains. In PlantHormones: Physiology, Biochemistry and Molecular Biology, edn 2. Editedby Davies PJ. Dordrecht: Kluwer Academic Publishers; 1995:246-271.
23. Perata P, Matsukura C, Vernieri P, Yamaguchi J: Sugar repression ofgibberellin-dependent signaling pathway in barley embryos. PlantCell 1997, 9:2197-2208.
24. Skriver K, Olsen FL, Rogers JC, Mundy J: Cis-acting DNA elementsresponsive to gibberellin and its antagonist abscisic acid. ProcNatl Acad Sci USA 1991, 88:7266-7270.
25. Morita A, Umemura TA, Kuroyanagi M, Futsuhara Y, Perata P, Yamaguchi J:Functional dissection of a sugar-repressed alpha-amylase gene(RAmy1A) promoter in rice embryos. FEBS Lett 1998, 423:81-85.
26. Toyofuku K, Umemura TA, Yamaguchi J: Promoter elements requiredfor sugar-repression of the RAmy3D gene for alpha-amylase inrice. FEBS Lett 1998, 428:275-280.
27. Gómez-Cadenas A, Zentella R, Walker-Simmons MK, Ho THD: • Gibberellin/abscisic acid antagonism in barley aleurone cells:
site of action of the protein kinase PKABA1 in relation to gib-berellin signaling molecules. Plant Cell 2001, 13:667-679.
This work demonstrates a direct interaction between ABA and GA signaling:ABA induction of the PKABA1 protein kinase suppresses the expression ofa transcription factor that is required for GA-induced gene expression.Interestingly, PKABA1 is related to SNF1 protein kinases, which are postulatedto play an important role in sugar response in diverse organisms.
28. Kashem MA, Hori H, Itoh K, Hayakawa T, Todoroki Y, Hirai N,Ohigashi H, Mitsui T: Effects of (+)-8′′,8′′,8′′-trifluoroabscisic acid onαα-amylase expression and sugar accumulation in rice cells. Planta1998, 205:319-326.
29. Loreti E, Matsukura CA, Gubler F, Alpi A, Yamaguchi J, Perata P: • Glucose repression of alpha-amylase in barley embryos is
independent of GAMYB transcription. Plant Mol Biol 2000, 44:85-90.The authors demonstrate that glucose repression of α-amylase expressioneither is independent of GA signaling or affects a step downstream ofGA-induced GAMyb transcription.
30. Toyofuku K, Loreti E, Vernieri P, Alpi A, Perata P, Yamaguchi J: • Glucose modulates the abscisic acid-inducible Rab16A gene in
cereal embryos. Plant Mol Biol 2000, 42:451-460.This work confirms that glucose-treated cereal embryos have reduced ABAlevels. This results in the inhibited expression of an ABA-regulated gene, butdoes not directly affect the expression of a sugar-repressed α-amylase gene.Hence, an antagonistic interaction between glucose and ABA signaling earlyin cereal germination is demonstrated.
31. Loreti E, Alpi A, Perata P: Glucose and disaccharide-sensing • mechanisms modulate the expression of αα-amylase in barley
embryos. Plant Physiol 2000, 123:939-948.Mechanisms that sense and regulate glucose and disaccharides independentlyare shown to exist in barley embryos. The authors investigate the structuralrequirements for disaccharide sensing, leading them to suggest thatdisaccharides may be perceived at the plasma membrane in a way that isindependent of sucrose transport.
32. Garciarrubio A, Legaria JP, Covarrubias AA: Abscisic acid inhibitsgermination of mature Arabidopsis seeds by limiting theavailability of energy and nutrients. Planta 1997, 203:182-187.
33. Finkelstein RR, Lynch TJ: Abscisic acid inhibition of radicle · emergence but not seedling growth is suppressed by sugars.
Plant Physiol 2000, 122:1179-1186.Antagonistic interactions between low exogenous sugars and ABA signalingare shown to occur early in Arabidopsis germination.
34. Eastmond PJ, Germain V, Lange PR, Bryce JH, Smith SM, Graham IA: • Postgerminative growth and lipid catabolism in oilseeds lacking
the glyoxylate cycle. Proc Natl Acad Sci USA 2000, 97:5669-5674.This work demonstrates that the glyoxylate cycle is not essential for Arabidopsisseedling establishment in light, indicating that the inhibition of germinationby ABA cannot be fully explained by carbon starvation due to ABA inhibitionof the glyoxylate cycle.
35. Arenas-Huertero F, Arroyo A, Zhou L, Sheen J, Léon P: Analysis of • Arabidopsis glucose insensitive mutants, gin5 and gin6, reveals a
central role of the plant hormone ABA in the regulation of plantvegetative development by sugar. Genes Dev 2000, 14:2085-2096.
ABA-deficient and ABA-insensitive mutants are selected on the basis of theirglucose-insensitive seedling growth. The allelism of gin6/abi4 was determinedby the cloning of gin6, which was based on T-DNA insertion. Growth on highsugar concentrations is shown to increase ABA levels, and gin/abi mutationsare shown to block hexokinase-induced signaling. The authors characterizethree classes of ABA-deficient mutants, as well as abi4 and abi5, as havinggin phenotypes; other abis were described as having a wildtype sugar response.
36. Huijser C, Kortstee A, Pego J, Weisbeek P, Wisman E, Smeekens S: • The Arabidopsis SUCROSE UNCOUPLED-6 gene is identical to
ABSCISIC ACID INSENSITIVE-4: involvement of abscisic acid insugar responses. Plant J 2000, 23:577-585.
A transposon-tagged allele of sun6/abi4 is selected by screening for mannose-insensitive germination and glucose-insensitive seedling growth. The authorscharacterize three classes of ABA-deficient mutants as having strong gin,but wildtype or weak mannose-insensitive germination (mig) phenotypes. Ofthe ABA-insensitive mutants, only abi4 has both gin and mig phenotypesthat are significant, but abi1 and abi3 also have mild mig phenotypes.
37. Laby RJ, Kincaid MS, Kim D, Gibson SI: The Arabidopsis• sugar-insensitive mutants sis4 and sis5 are defective in abscisic
acid synthesis and response. Plant J 2000, 23:587-596.ABA-deficient (sis4/aba2) and ABA-insensitive (sis5/abi4) mutants areselected on the basis of their sucrose-insensitive seedling growth. sis4/aba2and sis5/abi4 mutants showed glucose- and sorbitol-resistant early seedlingdevelopment; sis5/abi4 mutants were also mannose-resistant. Among otherabi mutants, abi5 was weakly resistant to these sugars, whereas abi1, abi2and abi3 showed little or no resistance.
ABA and sugar interactions regulating development Finkelstein and Gibson 31

38. Zhou L, Jang JC, Jones TL, Sheen J: Glucose and ethylene signaltransduction crosstalk revealed by an Arabidopsisglucose-insensitive mutant. Proc Natl Acad Sci USA 1998,95:10294-10299.
39. Beaudoin N, Serizet C, Gosti F, Giraudat J: Interactions betweenabscisic acid and ethylene signaling cascades. Plant Cell 2000,12:1103-1115.
40. Ghassemian M, Nambara E, Cutler S, Kawaide H, Kamiya Y,McCourt P: Regulation of abscisic acid signaling by the ethyleneresponse pathway in Arabidopsis. Plant Cell 2000,12:1117-1126.
41. Gibson SI, Laby RJ, Kim D: The sugar-insensitive1 (sis1) mutant of • Arabidopsis is allelic to ctr1. Biochem Biophys Res Comm 2001,
280:196-203.This work links ethylene and sugar response by the identification of a newallele of ctr1 in a screen for sucrose-resistant seedling growth. It alsodemonstrates gin, mig and osmotolerant phenotypes for both sis1/ctr1 andan ethylene overproducer (eto1), as well as sugar hypersensitivity for twoethylene-resistant mutants. In addition, this study identifies a limiteddevelopmental window (up to 40–50 h post-imbibition) during which sucroseinhibits seedling growth.
42. Sharp RE, LeNoble ME, Else MA, Thorne ET, Gherardi F:Endogenous ABA maintains shoot growth in tomatoindependently of effects on plant water balance: evidence for aninteraction with ethylene. J Exp Bot 2000, 51:1575-1584.
43. Spollen WG, LeNoble ME, Samuels TD, Bernstein N, Sharp RE:Abscisic acid accumulation maintains maize primary rootelongation at low water potentials by restricting ethyleneproduction. Plant Physiol 2000, 122:967-976.
44. Lopez-Molina L, Mongrand S, Chua NH: A postgermination • developmental arrest checkpoint is mediated by abscisic acid and
requires the ABI5 transcription factor in Arabidopsis. Proc NatlAcad Sci USA 2001, 98:4782-4787.
Accumulation of the transcription factor ABI5 is shown to be induced by ABA,drought-stress or salt-stress, and depends on stabilization by post-translational
modification. This stimulation is limited to the first 48–60 h post-stratification,a window similar to that of sugar-sensitive growth (see [41•]). ABI5 over-expression confers hypersensitivity to ABA, as described for overexpressionof ABI3 and ABI4 (see [10•,45•,46]), and ABI5 levels are correlated with thearrest of seedling growth and maintenance of a desiccation-tolerant state.
45. Söderman E, Brocard I, Lynch T, Finkelstein R: Regulation and • function of the Arabidopsis ABA-insensitive4 (ABI4) gene in seed
and ABA response signaling networks. Plant Physiol 2000,124:1752-1765.
ABI4 is shown to be expressed and to have a functional role in vegetativetissues as well as in seeds. A comparison of an allelic series shows that aweakly ABA-resistant phenotype probably reflects a complete loss of functionat this locus. ABI3, ABI4, and ABI5 function and expression are shown to becross-regulated, suggesting that these transcription factors act combinatorially.
46. Finkelstein RR, Rock CD: Abscisic acid biosynthesis and signaling.In The Arabidopsis Book. Edited by Somerville CR, Meyerowitz EM.2001, in press.
47. Blázquez MA, Green R, Nilsson O, Sussman MR, Weigel D:Gibberellins promote flowering of Arabidopsis by activating theLEAFY promoter. Plant Cell 1998, 10:791-800.
48. Xu X, Lammeren AAMV, Vermeer E, Vreugdenhil D: The role ofgibberellin, abscisic acid, and sucrose in the regulation of potatotuber formation in vitro. Plant Physiol 1998, 117:575-584.
49. Németh K, Salchert K, Putnoky P, Bhalerao R, Koncz-Kálmán Z,Stankovic-Stangeland B, Bakó L, Mathur J, Ökrész L, Stabel S et al.:Pleiotropic control of glucose and hormone responses by PRL1, anuclear WD protein, in Arabidopsis. Genes Dev 1998, 12:3059-3073.
50. Bhalerao RP, Salchert K, Bakó L, Ökrész L, Szabados L, Muranaka T,Machida Y, Schell J, Koncz C: Regulatory interaction of PRL1 WDprotein with Arabidopsis SNF1-like protein kinases. Proc NatlAcad Sci USA 1999, 96:5322-5327.
51. Dijkwel PP, Huijser C, Weisbeek PJ, Chua NH, Smeekens SCM:Sucrose control of phytochrome A signaling in Arabidopsis. PlantCell 1997, 9:583-595.
32 Growth and development