a unifying framework for modelling and analysing
TRANSCRIPT

BRANDENBURG UNIVERSITY OF TECHNOLOGY AT COTTBUS
Faculty of Mathematics, Natural Sciences and Computer Science
Institute of Computer Science
COMPUTER SCIENCE REPORTS
Report 02/07
July 2007
A UNIFYING FRAMEWORK FOR MODELLING AND ANALYSING BIOCHEMICAL PATHWAYS USING
PETRI NETS
DAVID GILBERT MONIKA HEINER
SEBASTIAN LEHRACK

Computer Science Reports Brandenburg University of Technology at Cottbus ISSN: 1437-7969 Send requests to: BTU Cottbus Institut für Informatik Postfach 10 13 44 D-03013 Cottbus

Computer Science Reports 02/07
July 2007
Brandenburg University of Technology at Cottbus
Faculty of Mathematics, Natural Sciences and Computer Science
Institute of Computer Science
David Gilbert, Monika Heiner, Sebastian Lehrack [email protected], [email protected],
[email protected] http://www-dssz.informatik.tu-cottbus.de
A Unifying Framework for Modelling and Analysing
Biochemical Pathways Using Petri Nets

Computer Science Reports Brandenburg University of Technology at Cottbus Institute of Computer Science Head of Institute: Prof. Dr. Winfried Kurth [email protected] BTU Cottbus Institut für Informatik Postfach 10 13 44 D-03013 Cottbus Research Groups: Headed by: Computer Engineering Prof. Dr. H. Th. Vierhaus Computer Network and Communication Systems Prof. Dr. H. König Data Structures and Software Dependability Prof. Dr. M. Heiner Database and Information Systems Prof. Dr. I. Schmitt Programming Languages and Compiler Construction Prof. Dr. P. Bachmann Software and Systems Engineering Prof. Dr. C. Lewerentz Theoretical Computer Science N.N. Graphics Systems Prof. Dr. W. Kurth Systems Prof. Dr. R. Kraemer Distributed Systems and Operating Systems Prof. Dr. J. Nolte Internet-Technology Prof. Dr. G. Wagner CR Subject Classification (1998): D.2.2, G.1.7, J.3 Printing and Binding: BTU Cottbus ISSN: 1437-7969

A Unifying Frameworkfor Modelling and Analysing
Biochemical Pathways Using Petri Nets
David Gilbert1, Monika Heiner2, and Sebastian Lehrack3
1 Bioinformatics Research Centre, University of GlasgowGlasgow G12 8QQ, Scotland, UK
[email protected] INRIA Rocquencourt, Projet Contraintes
BP 105, 78153 Le Chesnay CEDEX - [email protected], on sabbatical leave from 3
3 Department of Computer Science, Brandenburg University of TechnologyPostbox 10 13 44, 03013 Cottbus, Germany
Abstract. We give a description of a Petri net-based framework formodelling and analysing biochemical pathways, which unifies the qualita-tive, stochastic and continuous paradigms. Each perspective adds its con-tribution to the understanding of the system, thus the three approachesdo not compete, but complement each other. We illustrate our approachby applying it to an extended model of the three stage cascade, whichforms the core of the ERK signal transduction pathway. Consequentlyour focus is on transient behaviour analysis. We demonstrate how quali-tative descriptions are abstractions over stochastic or continuous descrip-tions, and show that the stochastic and continuous models approximateeach other. A key contribution of the paper consists in a precise defi-nition of biochemically interpreted stochastic Petri nets. Although ourframework is based on Petri nets, it can be applied more widely to otherformalisms which are used to model and analyse biochemical networks.
This Technical Report represents an extended version of [GHL07] andreplaces [Leh06].
1 Motivation
Biochemical systems are inherently governed by stochastic laws. However, dueto the computational efforts required to analyse stochastic models, two abstrac-tions are more popular: qualitative models, abstracting away from any timedependencies, and continuous models, commonly used to approximate stochas-tic behaviour by a deterministic one. The interrelationships between these threemodels are not always properly understood; for example, how the kinetics of abiochemical reaction, when described by a continuous model, is related to thestochastic nature of the underlying molecular mechanism.

2 Gilbert, Heiner, Lehrack
In a previous paper [GH06] we developed an approach for modelling andanalysing biochemical networks using discrete and continuous Petri nets. Ourcurrent work has taken this forward by considering stochastic Petri nets anddeveloping an overall framework to unify these three approaches, providing afamily of related models with high analytical power.
A key contribution of this paper is the precise definition of biochemicallyinterpreted stochastic Petri nets in a generic manner, and we demonstrate thebenefit of their incorporation into the model development process. We showhow the general definition can be tailored to very specific kinetic assumptionsby appropriate adjustments of the general hazard function. Also we discuss therelation of the stochastic Petri net to its time-free, purely qualitative abstrac-tion - the standard Petri net, as well as to its continuous approximation - thecontinuous Petri net (i.e., an ordinary differential equation system).
This paper is organised as follows. The following section provides an overviewof the biochemical context and introduces our running example. Next we outlineour framework, discussing the special contributions of the three individual anal-ysis approaches with special emphasis on the transient behaviour analysis, andexamining their interrelations. We then present the individual approaches anddiscuss mutually related properties in all three paradigms in the following order:we start off with the qualitative approach, which is conceptually the easiest, anddoes not rely on knowledge of kinetic information, but describes the networktopology and presence of the species. The qualitative modelling and analysisbasically adheres to the steps proposed in [GH06]. In addition, we show how tosystematically derive and interpret the partial order run of the signal responsebehaviour. We then demonstrate how the validated qualitative model can betransformed into the stochastic representation by addition of stochastic firingrate information. Next, the continuous model is derived from the qualitative orstochastic model by considering only deterministic firing rates. Suitable sets ofinitial conditions for all three models are constructed by qualitative analysis.Finally, we refer to related work, before concluding with a summary and outlookregarding further research directions.
2 Biochemical Context
We have chosen a model of the mitogen-activated protein kinase (MAPK) cas-cade published in [LBS00] as a running case study. This is the core of the ubiq-uitous ERK/MAPK pathway that can, for example, convey cell division anddifferentiation signals from the cell membrane to the nucleus. The model doesnot describe the receptor and the biochemical entities and actions immediatelydownstream from the receptor. Instead the description starts at the RasGTPcomplex which acts as a kinase to phosphorylate Raf, which phosphorylatesMAPK/ERK Kinase (MEK), which in turn phosphorylates Extracellular signalRegulated Kinase (ERK). This cascade (RasGTP → Raf → MEK → ERK) ofprotein interactions controls cell differentiation, the effect being dependent upon

A Unifying Framework, TR I-02/2007 3
the activity of ERK. We consider RasGTP as the input signal and ERKPP (ac-tivated ERK) as the output signal.
The bipartite graph in Figure 1 describes the typical modular structure forsuch a signalling cascade. Each layer corresponds to a distinct protein species.The protein Raf in the first layer is only singly phosphorylated. The proteinsin the two other layers, MEK and ERK respectively, can be singly as well asdoubly phosphorylated. In each layer, forward reactions are catalysed by kinasesand reverse reactions by phosphatases (Phase1, Phase2, Phase3). The kinases inthe MEK and ERK layers are the phosphorylated forms of the proteins in theprevious layer, see also [CKS07].
Raf
RasGTP
Raf_RasGTP
RafP
RafP_Phase1
MEK_RafP MEKP_RafP
MEKP_Phase2 MEKPP_Phase2
ERK
ERK_MEKPP ERKP_MEKPP
ERKP
MEKPP
ERKPP_Phase3ERKP_Phase3
MEKP
ERKPP
Phase2
Phase3
MEK
Phase1
k3
k6
k21
k18
k9 k12
k15
k24
k27k30
k7/k8
k1/k2
k4/k5
k10/k11
k16/k17
k22/k23k19/k20
k13/k14
k28/k29 k25/k26
Fig. 1. The bipartite graph for the extended ERK pathway model. The graph has beenderived by SBML import and automatic layout, manually improved, from the set of theODEs in [LBS00]. Circles stand for species (proteins, protein complexes). Protein com-plexes are indicated by an underscore “ ” between the constituent protein names. Thesuffixes P or PP indicate phosphorylated or doubly phosphorylated forms respectively.Squares stand for irreversible reactions, while two concentric squares specify reversiblereactions. The species that are read as input/output signals are given in grey.

4 Gilbert, Heiner, Lehrack
3 Overview of the framework
In the following we describe our overall framework, illustrated in Figure 2, thatrelates the three major ways of modelling and analysing biochemical networksdescribed in this paper: qualitative, stochastic and continuous.
Fig. 2. Conceptual framework
The most abstract representation of a biochemical network is qualitative andis minimally described by its topology, usually as a bipartite directed graph withnodes representing biochemical entities or reactions, or in Petri net terminologyplaces and transitions (see Figure 1). Arcs can be annotated with stoichiometricinformation, whereby the default stoichiometric value of 1 is usually omitted.
The qualitative description can be further enhanced by the abstract repre-sentation of discrete quantities of species, achieved in Petri nets by the use oftokens at places. These can represent the number of molecules, or the level ofconcentration, of a species, and a particular arrangement of tokens over a net-work is called a marking. The standard semantics for these qualitative Petrinets (QPN) does not associate a time with transitions or the sojourn of tokensat places, and thus these descriptions are time-free. The qualitative analysisconsiders however all possible behaviour of the system under any timing. Thebehaviour of such a net forms a discrete state space, which can be analysed inthe bounded case, for example, by a branching time temporal logic, one instanceof which is Computational Tree Logic (CTL), see [CGP01].
Timed information can be added to the qualitative description in two ways –stochastic and continuous. The stochastic Petri net (SPN) description preservesthe discrete state description, but in addition associates a probabilistically dis-tributed firing rate (waiting time) with each reaction. All reactions, which occurin the QPN, can still occur in the SPN, but their likelihood depends on theprobability distribution of the associated firing rates (waiting times). Special

A Unifying Framework, TR I-02/2007 5
behavioural properties can be expressed using e.g. Continuous Stochastic Logic(CSL), see [PNK06], a probabilistic counterpart of CTL. The QPN is an ab-straction of the SPN, sharing the same state space and transition relation withthe stochastic model, with the probabilistic information removed. All qualitativeproperties valid in the QPN are also valid in the SPN, and vice versa.
The continuous model replaces the discrete values of species with continuousvalues, and hence is not able to describe the behaviour of species at the levelof individual molecules, but only the overall behaviour via concentrations. Wecan regard the discrete description of concentration levels as abstracting over thecontinuous description of concentrations. Timed information is introduced by theassociation of a particular deterministic rate information with each transition,permitting the continuous model to be represented as a set of ordinary differentialequations (ODEs). The concentration of a particular species in such a model willhave the same value at each point of time for repeated experiments. The statespace of such models is continuous and linear. It can be analysed by, for example,Linear Temporal Logic with constraints (LTLc) in the manner of [CCRFS06].
The stochastic and continuous models are mutually related by approxima-tion. The stochastic description can be used as the basis for deriving a continu-ous Petri net (CPN) model by approximating rate information. Specifically, theprobabilistically distributed reaction firing in the SPN is replaced by a particularaverage firing rate over the continuous token flow of the CPN. This is achievedby approximation over hazard functions of type (1), described in more detail insection 5.1. In turn, the stochastic model can be derived from the continuousmodel by approximation, reading the tokens as concentration levels, as intro-duced in [CVGO06]. Formally, this is achieved by a hazard function of type (2),see again section 5.1.
It is well-known that time assumptions generally impose constraints on be-haviour. The qualitative and stochastic models consider all possible behavioursunder any timing, whereas the continuous model is constrained by its inherentdeterminism to consider a subset. This may be too restrictive when modellingbiochemical systems, which by their very nature exhibit variability in their be-haviour.
In the following the reader is assumed to be familiar with the standard Petrinet terminology as well as foundations of temporal logics, for an introductionsee, e.g., [Mur89] and [CGP01].
4 The Qualitative Approach
4.1 Qualitative Modelling
We interpret the graph given in Figure 1 as a place/transition Petri net, and callthe circles places , and the rectangles transitions . Reversible reactions have to bemodelled explicitly by two opposite transitions in the basic Petri net notation.However in order to retain the elegant graph structure of Figure 1, we use macro

6 Gilbert, Heiner, Lehrack
transitions, each of which stands here for a reversible reaction. The entire (flat-tened) place/transition Petri net consists of 22 places and 30 transitions, wherek1, k2, . . . stand for reaction (transition) labels.
We associate a discrete concentration with each of the 22 species. In thesimplest way, these concentrations can be thought of as being “high” or “low”(above or below a certain threshold), resulting in a two-level model where eachspecies can be read as a Boolean variable. More generally, we could apply amulti-level approach by differentiating between a finite number of discrete levels,each standing for an equivalence class of possibly infinitely many concentrations.Then species can be read as integer variables.
4.2 Qualitative Analysis
Analysis of general behavioural properties. The Petri net enjoys the threeorthogonal general properties of a discrete Petri net: boundedness, liveness andreversibility. The decision about the first two can be made for our running ex-ample in a static way, i.e. without construction of the state space, while the lastproperty requires dynamic analysis techniques, i.e., the explicit construction ofthe state space. The necessary steps of the systematic analysis procedure followbasically those given in [GH06]. We restrict ourselves here to the most essentialpoints. See Appendix A for the complete session protocol as produced by theIntegrated Net Analyser INA [SR99]
The net is strongly connected and thus self-contained, i.e. a closed system. Inorder to bring the net to life, we construct an initial marking using P-invariants.These are non-trivial non-negative integer solutions of the homogeneous linearequation system x · C = 0, where C stands for the incidence matrix of the net.There are seven minimal P-invariants covering the net, and consequently the netis bounded for any initial marking. All these P-invariants xi contain only entriesof 0 and 1, permitting a short-hand specification by just giving the names of theplaces involved.
x1 = (RasGTP, Raf RasGTP)x2 = (Raf, Raf RasGTP, RafP, RafP Phase1, MEK RafP, MEKP RafP)x3 = (MEK, MEK RafP, MEKP RafP, MEKP Phase2, MEKPP Phase2,
ERK MEKPP, ERKP MEKPP, MEKPP, MEKP)x4 = (ERK, ERK MEKPP, ERKP MEKPP, ERKP, ERKPP Phase3,
ERKP Phase3, ERK PP)x5 = (Phase1, RafP Phase1)x6 = (Phase2, MEKP Phase2, MEKPP Phase2)x7 = (Phase3, ERKP Phase3, ERKPP Phase3)
Each P-invariant stands for a reasonable conservation rule, the species preservedbeing given by the first name in the invariant. In signal transduction networksa P-invariant typically comprises all the different states of one species. In aBoolean approach, each species can be only in one state at any time, thus eachP-invariant gets exactly one token. Within a P-invariant, the species with themost inactive (i.e. non-phosphorylated) or the monomeric (non-complexed) state

A Unifying Framework, TR I-02/2007 7
is chosen. Following these criteria, the initial marking is: one token on each of Raf,RasGTP, MEK, ERK, Phase1, Phase2 and Phase3, while all remaining placesare empty. With this marking, the net is covered by 1-P-invariants (exactly onetoken in each P-invariant), and is therefore 1-bounded.
Generalising this reasoning to a multi-level concept, we could assign n tokensto each place, representing the most inactive state, in order to indicate thehighest concentration level for them in the initial state. The “abstract” massconservation within each P-invariant would then be n tokens, which could bedistributed fairly freely over the P-invariant’s places during the behaviour of themodel. This results in a dramatic increase of the state space, cf. the discussionin Section 5.2, while not improving the qualitative reasoning.
Model validation should include a check of all T-invariants for their biologicalplausibility. T-invariants are non-trivial non-negative integer solutions of thehomogeneous linear equation system C · y = 0. The entries of a T-invariantcan be read as the specification of a multiset of transitions, which reproduce agiven marking by their firing. If there are non-trivial solutions, then there areinfinitely many ones. Therefore, the plausibility check is usually restricted tothe consideration of all minimal solutions. The net representations of minimalT-invariants (their transitions plus their pre- and post-places and all arcs inbetween) characterise minimal self-contained subnetworks with an identifiablebiological meaning.
The net under consideration is covered by T-invariants, a necessary condi-tion for bounded nets to be live. Besides the expected ten trivial T-invariantsfor the ten reversible reactions, there are five non-trivial, but obvious minimalT-invariants, each corresponding to one of the five phosphorylation / dephos-phorylation cycles in the network structure:
y1 = (k1, k3, k4, k6),,y2 = (k7, k9, k16, k18),y3 = (k10, k12, k13, k15),y4 = (k19, k21, k28, k30),y5 = (k22, k24, k25, k27).
The interesting net behaviour, demonstrating how input signals cause finallyoutput signals, is contained in a non-negative linear combination of all five non-trivial T-invariants,
y1−5 = y1 + y2 + y3 + y4 + y5,
which is called an I/O T-invariant in the following. The I/O T-invariant is sys-tematically constructed by starting with the two minimal T-invariants, involvingthe input and output signal, which define disconnected subnetworks. Then weadd minimal sets of minimal T-invariants to get a connected subnet. For ourrunning example, the solution is unique, which is not generally the case.
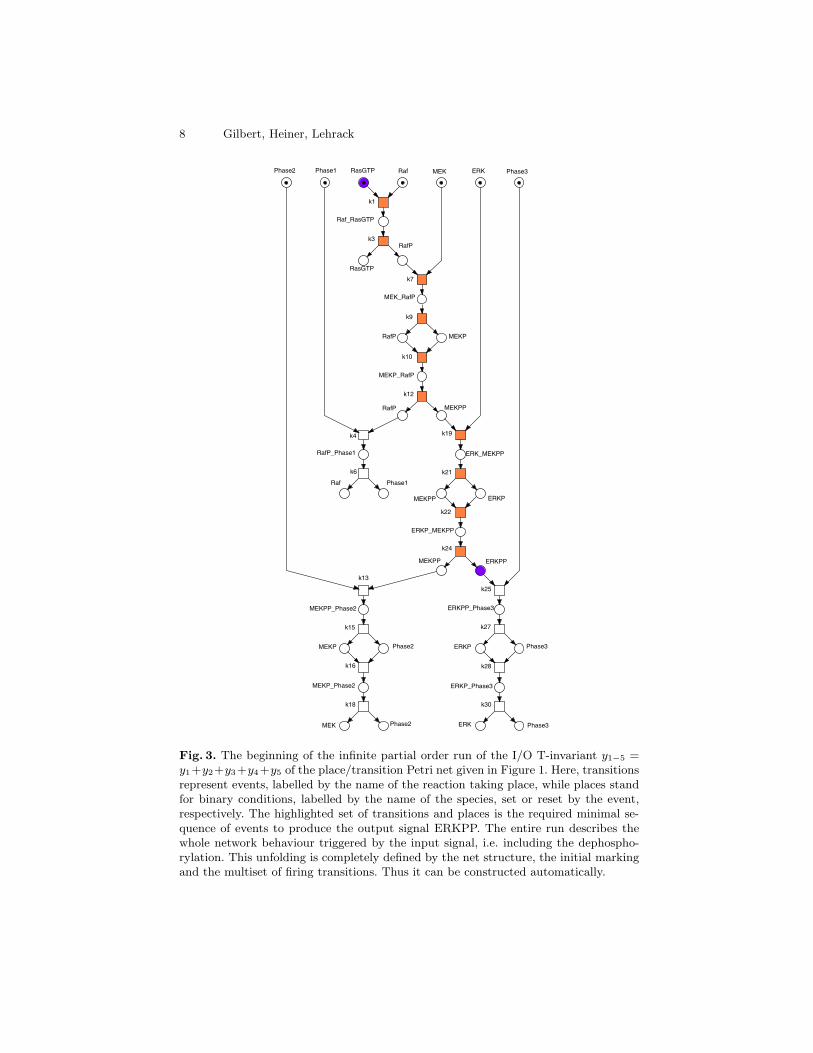
We check the I/O T-invariant for feasibility in the constructed initial mark-ing, which then involves the feasibility of all trivial T-invariants. We obtainan infinite partial order run, the beginning of which is given as labelled condi-tion/event net in Figure 3, and characterise this in a short-hand notation by the
8 Gilbert, Heiner, Lehrack
RasGTP Raf MEKPhase2 ERK Phase3
Raf_RasGTP
RasGTP
RafP
MEK_RafP
RafP MEKP
MEKP_RafP
RafP MEKPP
ERK_MEKPP
MEKPP ERKP
MEKPP ERKPP
ERKP_MEKPP
Phase1
RafP_Phase1
Raf Phase1
MEKPP_Phase2
MEKP Phase2
MEKP_Phase2
MEK Phase2 Phase3ERK
ERKP_Phase3
Phase3ERKP
ERKPP_Phase3
k1
k3
k7
k9
k10
k12
k19
k21
k22
k24
k4
k6
k13
k15
k16
k18
k25
k30
k28
k27
Fig. 3. The beginning of the infinite partial order run of the I/O T-invariant y1−5 =y1+y2+y3+y4+y5 of the place/transition Petri net given in Figure 1. Here, transitionsrepresent events, labelled by the name of the reaction taking place, while places standfor binary conditions, labelled by the name of the species, set or reset by the event,respectively. The highlighted set of transitions and places is the required minimal se-quence of events to produce the output signal ERKPP. The entire run describes thewhole network behaviour triggered by the input signal, i.e. including the dephospho-rylation. This unfolding is completely defined by the net structure, the initial markingand the multiset of firing transitions. Thus it can be constructed automatically.

A Unifying Framework, TR I-02/2007 9
following partially ordered word out of the alphabet of all transition labels (“;”stands for “sequentiality”, “‖” for “concurrency”):
( k1; k3; k7; k9; k10; k12;( ( k4; k6 ) ‖
( ( k19; k21; k22; k24 );( ( k13; k15; k16; k18 ) ‖ ( k25; k27; k28; k30 ) ) ) ) ).
This partial order run gives further insight into the dynamic behaviour ofthe network, which may not be apparent from the standard net representation,e.g. we are able to follow the (minimal) producing process of the proteins RafP,MEKP, MEKPP, ERKP and ERKPP, and we notice the clear independence ofthe dephosphorylation in all three levels.
The reachability graph of the net is finite because the net is bounded, and hasin the Boolean token interpretation 118 states out of 222 theoretically possibleones, forming one strongly connected component. Therefore, the Petri net isreversible, i.e. the initial system state is always reachable again, or in otherwords the system has the capability of self-reinitialization. Moreover, from theviewpoint of the discrete model, all of these 118 states are equivalent, and eachcould be taken as an initial state resulting in exactly the same total (discrete)system behaviour. This prediction will be confirmed by the observations gainedduring quantitative analyses, see Sections 5.2 and 6.2.
Model checking of special behavioural properties. Temporal logic is par-ticularly helpful in expressing special behavioural properties of the expectedtransient behaviour, whose truth can be determined via model checking. Weconfine ourselves here to two CTL properties, checking the generalizability ofthe insights gained by the partial order run of the I/O T-invariant. In the fol-lowing, places are interpreted as Boolean variables, in order to simplify notation.
property Q1: The signal sequence predicted by the partial order run of theI/O T-invariant is the only possible one. In other words, starting at the initialstate, it is necessary to pass through states RafP, MEKP, MEKPP and ERKPin order to reach ERKPP.
¬ [ E ( ¬ RafP U MEKP ) ∨ E ( ¬ MEKP U MEKPP ) ∨E ( ¬ MEKPP U ERKP ) ∨ E ( ¬ ERKP U ERKPP ) ]
property Q2: Dephosphorylation takes place independently. E.g., the du-ration of the phosphorylated state of ERK is independent of the duration of thephosphorylated states of MEK and Raf.
( EF [ Raf ∧ ( ERKP ∨ ERKPP ) ] ∧ EF [ RafP ∧ ( ERKP ∨ ERKPP ) ] ∧EF [ MEK ∧ ( ERKP ∨ ERKPP ) ] ∧EF [ ( MEKP ∨ MEKPP ) ∧ ( ERKP ∨ ERKPP ) ] )
In subsequent sections we will use Q1 as a basis to illustrate how the stochas-tic and continuous approaches provide complementary views of the system be-haviour.

10 Gilbert, Heiner, Lehrack
5 The Stochastic Approach
5.1 Stochastic Modelling
As with a qualitative Petri net, a stochastic Petri net maintains a discrete num-ber of tokens on its places. But contrary to the time-free case, a firing rate(waiting time) is associated with each transition t, which are random variablesXt ∈ [0,∞), defined by probability distributions. Therefore, all reaction timescan theoretically still occur, but the likelihood depends on the probability dis-tribution. Consequently, the system behaviour is described by the same discretestate space, and all the different execution runs of the underlying qualitativePetri net can still take place. This allows the use of the same powerful analysistechniques for stochastic Petri nets as already applied for qualitative Petri nets.
For a better understanding we describe the general procedure of a particularsimulation run for a stochastic Petri net. Each transition gets its own localtimer. When a particular transition becomes enabled, meaning that sufficienttokens arrive on its pre-places, then the local timer is set to an initial value,which is computed at this time point by means of the corresponding probabilitydistribution. In general, this value will be different for each simulation run. Thelocal timer is then decremented at a constant speed, and the transition will firewhen the timer reaches zero. If there is more than one enabled transition, a racefor the next firing will take place.
Technically, various probability distributions can be chosen to determine therandom values for the local timers. Biochemical systems are the prototype forexponentially distributed reactions. Thus, for our purposes, the firing rates ofall transitions follow an exponential distribution, which can be described bya single parameter λ, and each transition needs only its particular, generallymarking-dependent parameter λ to specify its local time behaviour. The follow-ing definition summarises this informal introduction.
Definition 1 (Stochastic Petri net, Syntax). A biochemically interpretedstochastic Petri net is a quintuple SPNBio = (P, T, f, v, m0), where
– P and T are finite, non empty, and disjoint sets. P is the set of places, andT is the set of transitions.
– f : ((P × T ) ∪ (T × P )) → IN0 defines the set of directed arcs, weighted bynon-negative integer values.
– v : T → H is a function, which assigns a stochastic hazard function ht toeach transition t, wherebyH :=
⋃t∈T
{ht |ht : IN|•t|
0 → IR+}
is the set of all stochastic hazard func-tions, and v(t) = ht for all transitions t ∈ T .
– m0 : P → IN0 gives the initial marking.
The stochastic hazard function ht defines the marking-dependent transitionrate λt(m) for the transition t. The domain of ht is restricted to the set of pre-places of t, i.e. •t := {p ∈ P |f (p, t) = 0}, to enforce a close relation betweennetwork structure and hazard functions. Therefore λt(m) actually depends onlyon a sub-marking.

A Unifying Framework, TR I-02/2007 11
Stochastic Petri net, Semantics. Transitions become enabled as usual, i.e.if all pre-places are sufficiently marked. However there is a time, which has toelapse, before an enabled transition t ∈ T fires. The transition’s waiting timeis an exponentially distributed random variable Xt with the probability densityfunction:
fXt(τ) = λt(m) · e(−λt(m)·τ), τ ≥ 0.
The firing itself does not consume time and again follows the standard firingrule of qualitative Petri nets. The semantics of a stochastic Petri net (withexponentially distributed reaction times for all transitions) is described by acontinuous time Markov chain (CTMC). The CTMC of a stochastic Petri netis isomorphic to the reachability graph of the underlying qualitative Petri net,while the arcs between the states are now labelled by the transition rates. Formore details see [MBC+95], [BK02].
Based on this general SPNBio definition, specialised biochemically inter-preted stochastic Petri nets can be defined by specifying the required kind ofstochastic hazard function more precisely. We give two examples, reading thetokens as molecules or as concentration levels. The stochastic mass-action haz-ard function tailors the general SPNBio definition to biochemical mass-actionnetworks, where tokens correspond to molecules:
ht := ct ·∏p∈•t
(m(p)f(p, t)
), (1)
where ct is the transition-specific stochastic rate constant, and m(p) is the cur-rent number of tokens on the pre-place p of transition t. The binomial coefficientdescribes the number of non-ordered combinations of the f(p, t) molecules, re-quired for the reaction, out of the m(p) available ones.
Tokens can also be read as concentration levels, as introduced in [CVGO06].The current concentration of each species is given as an abstract level. We as-sume the maximum molar concentration is M , and the amount of different levelsis N + 1. Then the abstract values 0, . . . , N represent the concentration inter-vals 0, (0, 1 ∗ M/N ], (1 ∗ M/N, 2 ∗ M/N ], . . . , (N − 1 ∗ M/N, N ∗ M/N ].Each of these (finite many) discrete levels stands for an equivalence class of (in-finitely many) continuous states. The stochastic level hazard function tailors thegeneral SPNBio definition to biochemical mass-action networks, where tokenscorrespond to concentration levels:
ht := kt · N ·∏p∈•t
(m(p)N
), (2)
where kt is the transition-specific deterministic rate constant, and N the num-ber of the highest level. The transformation rules between the stochastic anddeterministic rate constants are well-understood, see e.g. [Wil06]. In practice,

12 Gilbert, Heiner, Lehrack
kinetic rates are taken from literature, textbooks etc. or determined from bio-chemical experiments. Hazard function (2) is the means whereby the continuousmodel (see the framework in Figure 2 and Section 6) can be approximated by thestochastic model; this can generally be achieved by a limited number of levels –see Section 5.2.
5.2 Stochastic Analysis
Due to the isomorphy of the reachability graph and the CTMC, all qualitativeanalysis results obtained in Section 4 are still valid. The influence of time doesnot restrict the possible system behaviour. Specifically it holds that the CTMCof our case study is reversible, which ensures ergodicity; i.e. we could start thesystem in any of the reachable states, always resulting in the same CTMC withthe same steady state probability distribution.
Additionally, probabilistic analyses of the transient and steady state be-haviour are now available. Generally, this can be done in an analytic as wellas in a simulative manner. In the following our main focus is on the analyticmodel checking approach.
In Section 4.2 we employed CTL to express behavioural properties. Sincewe have now a stochastic model, we apply Continuous Stochastic Logic (CSL)[PNK06], which replaces the path quantifiers (E, A) in CTL by the probabilityoperator P��p, whereby �� p specifies the probability of the given formula. Forexample, introducing in CSL the abbreviation Fφ for trueUφ, the CTL formulaEFφ becomes the CSL formula P≥0[Fφ ], and AFφ becomes P≥1[Fφ ].
In order to use the probabilistic model checker PRISM [PNK06], we encodethe extended ERK pathway in its modelling language following the approachproposed in [DDS04], see Appendix B. This translation requires knowledge ofthe boundedness degree of all species involved, which we acquire by the structuralanalysis technique of P-invariants.
We only consider here the levelsemantics. Since the continuous con-centrations of proteins in the ERKpathway are all in the same range(0.1. . . 0.4 mMol in 0.1 steps), weemploy a model with only 4, and asecond version with 8 levels, com-pare Figure 4.The corresponding CTMCs (andreachability graphs) comprise24,065 states for the 4 level versionand 6,110,643 states for the 8 levelversion.
Fig. 4. The partitioning of the con-centration scale into discrete levels.

A Unifying Framework, TR I-02/2007 13
Equivalence check by transient analysis. We start with a transient analysisto prove the sufficient equivalence between the stochastic model in the levelsemantics and the corresponding continuous model, justifying the interpretationof the properties gained by the stochastic model also in terms of the continuousone. The probabilistic model checker PRISM permits the analysis of the transientbehaviour of the stochastic model, e.g., the concentration of RafP at time t isgiven by:
CRafP (t) = 0.1s ·
4s∑i=1
(i · P (LRafP (t) = i)
)︸ ︷︷ ︸expected value of LRafP (t)
.
The random variable LRafP (t) stands for the level of RafP at time t. We sets to 1 for the 4 level version, and to 2 for the 8 level version. The factor 0.1
scalibrates the expected value for a given level to the concentration scale. In the4 level version a single level represents 0.1 mMol and 0.05 mMol in the 8 levelversion. Figure 5 shows the simulation results for the species MEK and Ras-GTP in the time interval [0..100] according to the continuous and the stochasticmodels respectively. These results confirm that 4 levels are sufficiently adequateto approximate the continuous model, and that 8 levels are preferable if thecomputational expenses are acceptable.
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2
0 20 40 60 80 100
Con
cent
ratio
n
Time
MEK
ContinuousStochastic 4 levelStochastic 8 level
0.055
0.06
0.065
0.07
0.075
0.08
0.085
0.09
0.095
0.1
0 20 40 60 80 100
Con
cent
ratio
n
Time
RasGTP
ContinuousStochastic 4 levelStochastic 8 level
Fig. 5. Comparison of the concentration traces.
Probabilistic model checking of special behavioural properties. Wegive two properties related to the partial order run of the I/O T-invariant, seeSection 4.2 and qualitative property Q1 therein, from which we expect a con-secutive increase of RafP, MEKPP and ERKPP. Both properties are expressedas so-called experiments, which are analysed varying the parameter L over alllevels, i.e. 0 to N. For the sake of efficiency, we restrict the U operator to 100time steps. Note that places are read as integer variables in the following.
14 Gilbert, Heiner, Lehrack
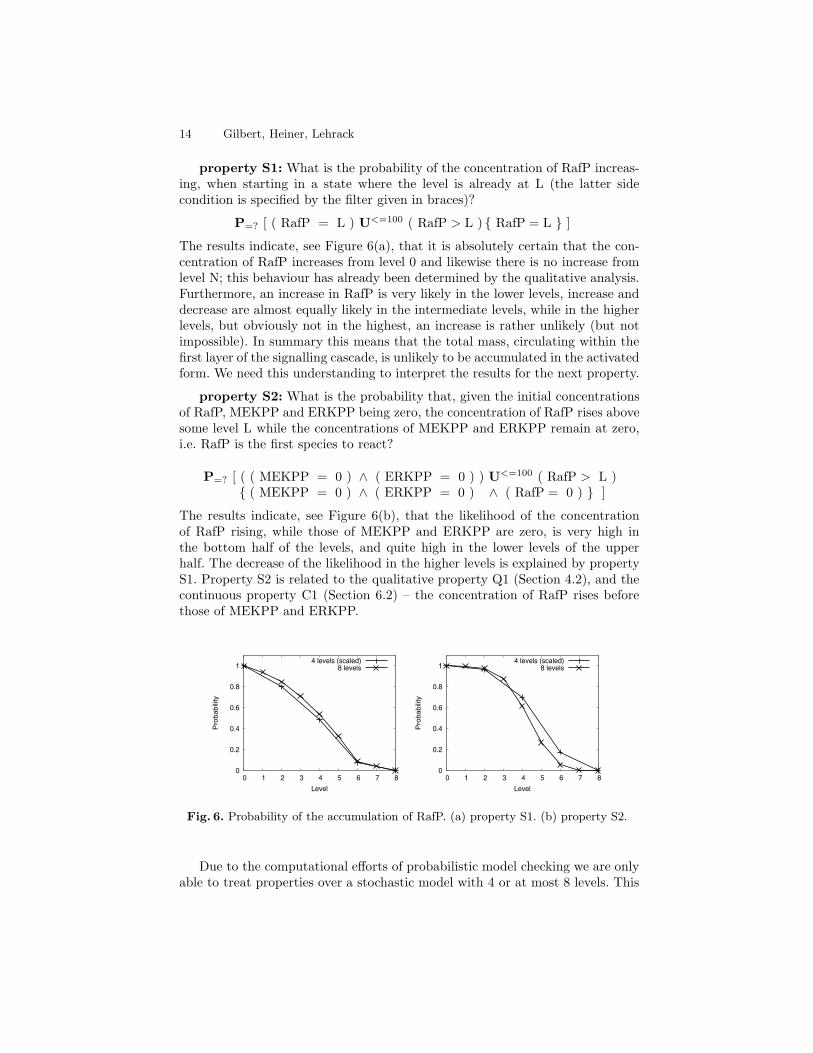
property S1: What is the probability of the concentration of RafP increas-ing, when starting in a state where the level is already at L (the latter sidecondition is specified by the filter given in braces)?
P=? [ ( RafP = L ) U<=100 ( RafP > L ) { RafP = L } ]
The results indicate, see Figure 6(a), that it is absolutely certain that the con-centration of RafP increases from level 0 and likewise there is no increase fromlevel N; this behaviour has already been determined by the qualitative analysis.Furthermore, an increase in RafP is very likely in the lower levels, increase anddecrease are almost equally likely in the intermediate levels, while in the higherlevels, but obviously not in the highest, an increase is rather unlikely (but notimpossible). In summary this means that the total mass, circulating within thefirst layer of the signalling cascade, is unlikely to be accumulated in the activatedform. We need this understanding to interpret the results for the next property.
property S2: What is the probability that, given the initial concentrationsof RafP, MEKPP and ERKPP being zero, the concentration of RafP rises abovesome level L while the concentrations of MEKPP and ERKPP remain at zero,i.e. RafP is the first species to react?
P=? [ ( ( MEKPP = 0 ) ∧ ( ERKPP = 0 ) ) U<=100 ( RafP > L ){ ( MEKPP = 0 ) ∧ ( ERKPP = 0 ) ∧ ( RafP = 0 ) } ]
The results indicate, see Figure 6(b), that the likelihood of the concentrationof RafP rising, while those of MEKPP and ERKPP are zero, is very high inthe bottom half of the levels, and quite high in the lower levels of the upperhalf. The decrease of the likelihood in the higher levels is explained by propertyS1. Property S2 is related to the qualitative property Q1 (Section 4.2), and thecontinuous property C1 (Section 6.2) – the concentration of RafP rises beforethose of MEKPP and ERKPP.
0
0.2
0.4
0.6
0.8
1
0 1 2 3 4 5 6 7 8
Pro
babili
ty
Level
4 levels (scaled)8 levels
0
0.2
0.4
0.6
0.8
1
0 1 2 3 4 5 6 7 8
Pro
babili
ty
Level
4 levels (scaled)8 levels
Fig. 6. Probability of the accumulation of RafP. (a) property S1. (b) property S2.
Due to the computational efforts of probabilistic model checking we are onlyable to treat properties over a stochastic model with 4 or at most 8 levels. This

A Unifying Framework, TR I-02/2007 15
restricts the kind of properties that we can prove; e.g., in order to check increasesof MEKPP and ERKPP – as suggested by the qualitative property Q1 and doneabove for RafP in the stochastic properties S1 and S2 – we would need 50 or 200levels respectively.
Analytic probabilistic model checking becomes more and more impracticablewith increasing size of the state space. Hence, the computation time of a prob-abilistic experiment, which typically consists of a series of probabilistic queries,can easily exceed several hours on a standard workstation. In order to avoidthe enormous computational power required for larger state spaces, the time-dependent stochastic behaviour can be simulated by dedicated algorithms, e.g.[Gil77], or approximated by a continuous one, see next section.
6 The Continuous Approach
6.1 Continuous Modelling
In a continuous Petri net the marking of a place is no longer an integer, buta positive real number, which can be read as the concentration of the speciesmodelled by the place. Transitions fire continuously, whereby the current deter-ministic firing rate generally depends on the current marking of the pre-places,i.e. of the current concentrations of the reactants. For our running case study,we derive the continuous model from the qualitative Petri net by associating amass action rate with each transition in the network, i.e., the reaction labels arenow read as the deterministic rate constants.
We can likewise derive the continuous Petri net from the stochastic Petri netby approximating over the hazard function of type (1), see for instance [Wil06].In both cases, we obtain a continuous Petri net, preserving the structure of thediscrete one, see Figure 2.
The semantics of a continuous Petri net is defined by a system of ODEs,whereby one equation describes the continuous change over time on the tokenvalue of a given place by the continuous increase of its pre-transitions’ flow andthe continuous decrease of its post-transitions’ flow, i.e., each place p subject tochanges gets its own equation:
dm (p)dt
=∑t∈•p
f (t, p) v (t) −∑t∈p •
f (p, t) v (t) ,
whereby •p stands for the set of pre-transitions of p, i.e. •p := {t ∈ T |f (t, p) = 0},and p • for the set of post-transitions of p, i.e. p • := {t ∈ T |f (p, t) = 0}. See[GH06] for more details. The complete system of non-linear ODEs generatedfrom the continuous Petri net of our running example is given in Appendix C.
The initial concentrations as suggested by the qualitative analysis correspondto those given in [LBS00], when mapping non-zero values to 1. For reasons ofbetter comparability we have also considered more precise initial concentrations,where the presence of a species is encoded by biologically motivated real valuesvarying between 0.1 and 0.4 in steps of 0.1, compare Table 2 in Appendix C.

16 Gilbert, Heiner, Lehrack
6.2 Continuous Analysis
Steady state analysis. Since there are 22 species, there are 222, i.e. 4,194,304possible initial states in the qualitative Petri net (Boolean token interpretation).Of these, 118 were identified by the reachability graph analysis (Section 4.2) toform one strongly connected component, and thus to be “good” initial states, seeTable 3 in Appendix C. These are ‘sensible’ initial states from the point of viewof biochemistry in that in all these 118 cases, and in none of the other 4,194,186states, each protein species is in a high initial concentration in only one of thefollowing states: uncomplexed, complexed, unphosphorylated or phosphorylated.These conditions relate exactly to the 1-P-invariant interpretation given in ourinitial marking construction procedure in Section 4.2.
We then computed the steady state of the set of species for each possibleinitial state, using the MatLab ODE solver ode45, which is based on an explicitRunge-Kutta formula, the Dormand-Prince pair [DP80], with 350 time steps.
In Figure 7 (a) we reproduce the computed behaviour of MEK for all 118good initial states, showing that despite differences in the concentrations atearly time points, the steady state concentration is the same in all 118 states.In Appendix C we reproduce two arbitrary chosen simulations of the model:State 1 (Figure 8) corresponding to the initial marking suggested by Section 4.2,and State 10 (Figure 9) corresponding to S10 in Table 3. The equivalence of thefinal states, compared with the difference in some intermediate states is clearlyillustrated in all these figures.
In summary, our results show that all of the ’good’ 118 states result in thesame set of steady state values for the 22 species in the pathway, within thebounds of computational error of the ODE solver. None of the remaining pos-sible initial states results in a steady state close to that generated by the 118markings in the reachability graph. See Table 4 in Appendix C for the steadystate concentrations of the 22 species.
0 50 100 150 200 250 300 3500
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Time (sec)
Con
cent
ratio
n (r
elat
ive
units
)
MEK
MEK
0
0.02
0.04
0.06
0.08
0.1
0.12
0 50 100 150 200 250 300 350
RasGTPRafP
MEKPPERKPP
Fig. 7. (a) Steady state analysis of MEK for all 118 ‘good’ states. (b) Continuoustransient analysis of the phosphorylated species RasP, MEKPP, ERKPP, triggered byRasGTP.

A Unifying Framework, TR I-02/2007 17
This is an interesting result, because the net considered here is not coveredby the class of net structures discussed in [ADLS06] with the unique steady stateproperty.
Continuous model checking of the transient behaviour. Correspondingto the partial order run of the I/O T-invariant, see Section 4.2, we expect aconsecutive increase of RafP, MEKPP, ERKPP, which we get confirmed by thetransient behaviour analysis, compare Figure 7 (b). To formalise the visual eval-uation of the diagram we use the continuous linear logic LTLc [CCRFS06], whichis interpreted over the continuous simulation trace of ODEs.
The following three queries confirm together the claim of the expected prop-agation sequence. In the queries we have to refer to absolute values. The steadystate values are obtained from the steady state analysis in the previous section;these are 0.12 mMol for RafP, 0.008 mMol for MEKPP and 0.002 mMol forERKPP, all of them being zero in the initial state. If a species’ concentrationis above half of its steady state value, we call this concentration level signifi-cant. Note that in order to simplify the notation, places are interpreted as realvariables in the following.
property C1: The concentration of RafP rises to a significant level, whilethe concentrations of MEKPP and ERKPP remain close to zero; i.e. RafP isreally the first species to react.
( (MEKPP < 0.001) ∧ (ERKPP < 0.0002) ) U (RafP > 0.06)
property C2: if the concentration of RafP is at a significant concentrationlevel and that of ERKPP is close to zero, then both species remain in thesestates until the concentration of MEKPP becomes significant; i.e. MEKPP isthe second species to react.
( (RafP > 0.06) ∧ (ERKPP < 0.0002) ) ⇒( (RafP > 0.06)∧ (ERKPP < 0.0002) ) U (MEKPP > 0.004)
property C3: if the concentrations of RafP and MEKPP are significant, theyremain so, until the concentration of ERKPP becomes significant; i.e. ERKPPis the third species to react.
( (RafP > 0.06) ∧ (MEKPP > 0.004) ) ⇒( (RafP > 0.06)∧ (MEKPP > 0.004) ) U (ERKPP > 0.0005)
Note that properties C1, C2 and C3 correspond to the qualitative propertyQ1, and that S2 is the stochastic counterpart of C1.

18 Gilbert, Heiner, Lehrack
7 Tools
The bipartite graph in Figure 1 and its interpretation as the three Petri net mod-els have been done using Snoopy [Sno], a tool to design and animate hierarchicalgraphs, including SBML import.
The qualitative analyses have been made using the Integrated Net AnalyserINA [SR99] and the Model Checking Kit [SSE04], supported by an IDD-basedmodel checker [Tov06]. We employed PRISM [PNK06] for probabilistic modelchecking, and Biocham [CCRFS06] for LTLc-based continuous model checking.
MATLAB [SR97] was used to produce the steady state analysis of all ini-tial states in the continuous model, and the transient analysis was done usingBioNessie [Bio], an SBML-based simulation and analysis tool for biochemicalnetworks.
8 Related Work
Stochastic Petri nets are an established concept for performance and depend-ability analysis of technical systems in the steady state, see e.g. [MBC+95],[BK02], recently extended by probabilistic model checking [DDS04]. The appli-cation of stochastic Petri nets to biochemical networks was first proposed in[GP98], where they were applied to a gene regulatory network. Further casestudies are discussed in [SPB01], [WMS05], [ST05], [SSW05], [Cur06]. Howevernone of these papers provides a general precise definition of the underlayingstochastic modelling concept.
Model checking is very popular for the verification of technical systems. Forbiochemical networks, qualitative model checking (Boolean semantics) has beenintroduced in [EKL+02], [CF03], probabilistic model checking in [CVGO06],[HKN+06], and continuous model checking in [APUM03], [CCRFS06].
The approximation of continuous behaviour by the discretisation of species’concentrations by a finite number of levels has been proposed in [CVGO06]. In[CGH05] it is shown how ODEs can be derived from stochastic process algebradescriptions using an activity matrix, related to stoichiometry, but they do notexplicitly discuss the interrelationship between the deterministic and stochasticmodels by the use of hazard functions. They have exploited this approach tocompare stochastic and continuous models via simulation traces [CDGH06].
The Biocham approach [CCRFS06], as it stands now, restricts itself to aBoolean semantics for qualitative models, defined over the Kripke structure,while we consider the more general case of integer semantics, which may collapseto the Boolean one. Petri net based model checking also supports the partial or-der semantics. Biocham uses PLTL to analyse stochastic models, where integersrepresent numbers of molecules. The stochastic view in Biocham does not givean interpretation in terms of the level semantics. To analyse continuous models,Biocham provides LTLc, which we used for continuous model checking of thetransient behaviour. Meanwhile, this step is facilitated substantially by the ex-tension introduced in [FR07], which allows to infer the variable values fulfillinga given temporal property from a (set of) continuous simulation run(s).

A Unifying Framework, TR I-02/2007 19
Finally, there is a lot of activity relating stochastic and continuous models –see [TSB04] for a review. Most work has ignored analysis but instead focussedon simulation, at the molecular, inherently stochastic, level and the population(continuous) level using differential equations, possibly stochastic.
9 Summary and Outlook
In this paper we have described an overall framework that relates the three majorways of modelling biochemical networks – qualitative, stochastic and continuous– and illustrated this in the context of Petri nets. In doing so we have given a pre-cise definition of biochemically interpreted stochastic Petri nets. We have shownthat the qualitative time-free description is the most basic, with discrete valuesrepresenting numbers of molecules or levels of concentrations. The qualitativedescription abstracts over two timed, quantitative models. In the stochastic de-scription, discrete values for the amounts of species are retained, but a stochasticrate is associated with each reaction. The continuous model describes amountsof species using continuous values and associates a deterministic rate with eachreaction. These two time-dependent models can be mutually approximated byhazard functions belonging to the stochastic world.
We have illustrated our framework by considering qualitative, stochastic andcontinuous Petri net descriptions of the ERK signalling pathway, based on themodel from Levchenko et al [LBS00]. We have focussed on analysis techniquesavailable in each of these three paradigms, in order to illustrate their complemen-tarity. Our special emphasis has been on model checking, which is especially use-ful for transient behaviour analysis, and we have demonstrated this by discussingrelated properties in the qualitative, stochastic and continuous paradigms. Al-though our framework is based on Petri nets, it can be applied more widely toother formalisms which are used to model and analyse biochemical networks.
We are now working on the incorporation of deterministic time into stochasticmodels, as well as the integration of continuous and stochastic aspects into onemodel.
Acknowledgements The running case study has been partly carried out bySebastian Lehrack during his study stay at the Bioinformatic Research Centreof the University of Glasgow. This stay was supported by the Max GruenebaumFoundation [MGF] and the UK Department of Trade and Industry Beacon Bio-science Programme.
We would like to thank Richard Orton and Xu Gu for the constructive dis-cussions as well as Vladislav Vyshermirsky for his support in the computationalexperiments.

20 Gilbert, Heiner, Lehrack
Appendix A: Qualitative Model
INA Analysis Protocol [SR99]
Integrated Net Analyzer [v2.2final-Jul 31 2003-win32] session report:
Current net options are:
token type: black (for Place/Transition nets)
time option: no times
firing rule: normal
priorities : not to be used
strategy : single transitions
line length: 80
Net read from levchenko_N1.pnt
Information on elementary structural properties:
Current name options are:
transition names to be written
place names to be written
The net is not statically conflict-free.
The net is pure.
The net is ordinary.
The net is homogenous.
The net is not conservative.
The net is not subconservative.
The net is not a state machine.
The net is not free choice.
The net is not extended free choice.
The net is not extended simple.
The net is marked.
The net is not marked with exactly one token.
The net is not a marked graph.
The net has a non-blocking multiplicity.
The net has no nonempty clean trap.
The net has no transitions without pre-place.
The net has no transitions without post-place.
The net has no places without pre-transition.
The net has no places without post-transition.
Maximal in/out-degree: 6
The net is connected.
The net is strongly connected.
ORD HOM NBM PUR CSV SCF CON SC Ft0 tF0 Fp0 pF0 MG SM FC EFC ES
Y Y Y Y N N Y Y N N N N N N N N N
DTP SMC SMD SMA CPI CTI B SB REV DSt BSt DTr DCF L LV L&S
? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Current name options are:
transition names to be written
place names to be written
Structural liveness properties of net nr. 0.t:
processed candidates: 124

A Unifying Framework, TR I-02/2007 21
There are 7 minimal deadlocks:
-------------------------------
1: 14.ERKPP_Phase3, 15.ERKP_Phase3, 19.Phase3,
2: 9.ERK, 10.ERK_MEKPP, 11.ERKP_MEKPP, 12.ERKP, 14.ERKPP_Phase3,
15.ERKP_Phase3, 17.ERKPP,
3: 7.MEKP_Phase2, 8.MEKPP_Phase2, 18.Phase2,
4: 5.MEK_RafP, 6.MEKP_RafP, 7.MEKP_Phase2, 8.MEKPP_Phase2,
10.ERK_MEKPP, 11.ERKP_MEKPP, 13.MEKPP, 16.MEKP, 20.MEK,
5: 1.RasGTP, 2.Raf_RasGTP,
6: 4.RafP_Phase1, 21.Phase1,
7: 0.Raf, 2.Raf_RasGTP, 3.RafP, 4.RafP_Phase1, 5.MEK_RafP,
6.MEKP_RafP,
Corresponding maximal traps:
----------------------------
1: 14.ERKPP_Phase3, 15.ERKP_Phase3, 19.Phase3,
2: 9.ERK, 10.ERK_MEKPP, 11.ERKP_MEKPP, 12.ERKP, 14.ERKPP_Phase3,
15.ERKP_Phase3, 17.ERKPP,
3: 7.MEKP_Phase2, 8.MEKPP_Phase2, 18.Phase2,
4: 5.MEK_RafP, 6.MEKP_RafP, 7.MEKP_Phase2, 8.MEKPP_Phase2,
10.ERK_MEKPP, 11.ERKP_MEKPP, 13.MEKPP, 16.MEKP, 20.MEK,
5: 1.RasGTP, 2.Raf_RasGTP,
6: 4.RafP_Phase1, 21.Phase1,
7: 0.Raf, 2.Raf_RasGTP, 3.RafP, 4.RafP_Phase1, 5.MEK_RafP,
6.MEKP_RafP,
The deadlock-trap-property is valid.
The net has no dead reachable states.
SM-allocatability of net nr. 0:
********************************
Some minimal place sets Q with QF=FQ, but all SCSM’s:
1: 14.ERKPP_Phase3, 15.ERKP_Phase3, 19.Phase3, SCSM
2: 9.ERK, 10.ERK_MEKPP, 11.ERKP_MEKPP, 12.ERKP, 14.ERKPP_Phase3,
15.ERKP_Phase3, 17.ERKPP, SCSM
3: 7.MEKP_Phase2, 8.MEKPP_Phase2, 18.Phase2, SCSM
4: 5.MEK_RafP, 6.MEKP_RafP, 7.MEKP_Phase2, 8.MEKPP_Phase2,
10.ERK_MEKPP, 11.ERKP_MEKPP, 13.MEKPP, 16.MEKP, 20.MEK, SCSM
5: 1.RasGTP, 2.Raf_RasGTP, SCSM
6: 4.RafP_Phase1, 21.Phase1, SCSM
7: 0.Raf, 2.Raf_RasGTP, 3.RafP, 4.RafP_Phase1, 5.MEK_RafP,
6.MEKP_RafP, SCSM
7 minimal component(s) found.
The net is state machine decomposable (SMD).
The net is coverable by state machines (SMC).
The net is covered by semipositive P-invariants.
The net is structurally bounded.
The net is bounded.
There are no proper semipositive T-surinvariants.

22 Gilbert, Heiner, Lehrack
The net is not state machine allocatable (SMA).
A pre-place allocation allocating no SCSM:
0==> 2, 1==> 4, 2==> 10, 3==> 7, 4==> 5,
5==> 6, 6==> 8, 7==> 11, 8==> 0, 9==> 2,
10==> 14, 11==> 15, 12==> 16, 13==> 7, 14==> 15,
15==> 19, 16==> 8, 17==> 13, 18==> 4, 19==> 3,
20==> 14, 21==> 17, 22==> 6, 23==> 5, 24==> 11,
25==> 10, 26==> 20, 27==> 3, 28==> 9, 29==> 12,
The net is covered by 1-token SCSM’s.
ORD HOM NBM PUR CSV SCF CON SC Ft0 tF0 Fp0 pF0 MG SM FC EFC ES
Y Y Y Y N N Y Y N N N N N N N N N
DTP SMC SMD SMA CPI CTI B SB REV DSt BSt DTr DCF L LV L&S
Y Y Y N Y ? Y Y ? N ? ? ? ? ? ?
Current options are:
computation of place-invariants
no computation of subinvariants
no computation of surinvariants
run automatically
output format = print non-zero entries with names
For net nr. 0.t :
7 place-invariants written to INVARI.HLP
0 rows lost!
Formatting lines from INVARI.HLP
Current reduction options are:
Delete repeated occurrences of the same support
Do not delete covered sub/sur/invariants
Delete strict invariants covering sub/sur/invariants
place sub/sur/invariants for net 0.t :
semipositive place invariants =
1 | 1.RasGTP : 1,
| 2.Raf_RasGTP : 1
2 | 4.RafP_Phase1 : 1,
| 21.Phase1 : 1
3 | 7.MEKP_Phase2 : 1,
| 8.MEKPP_Phase2 : 1,
| 18.Phase2 : 1
4 | 0.Raf : 1,
| 2.Raf_RasGTP : 1,
| 3.RafP : 1,
| 4.RafP_Phase1 : 1,
| 5.MEK_RafP : 1,
| 6.MEKP_RafP : 1
5 | 14.ERKPP_Phase3 : 1,
| 15.ERKP_Phase3 : 1,

A Unifying Framework, TR I-02/2007 23
| 19.Phase3 : 1
6 | 5.MEK_RafP : 1,
| 6.MEKP_RafP : 1,
| 7.MEKP_Phase2 : 1,
| 8.MEKPP_Phase2 : 1,
| 10.ERK_MEKPP : 1,
| 11.ERKP_MEKPP : 1,
| 13.MEKPP : 1,
| 16.MEKP : 1,
| 20.MEK : 1
7 | 9.ERK : 1,
| 10.ERK_MEKPP : 1,
| 11.ERKP_MEKPP : 1,
| 12.ERKP : 1,
| 14.ERKPP_Phase3 : 1,
| 15.ERKP_Phase3 : 1,
| 17.ERKPP : 1
The net is safe.
ORD HOM NBM PUR CSV SCF CON SC Ft0 tF0 Fp0 pF0 MG SM FC EFC ES
Y Y Y Y N N Y Y N N N N N N N N N
DTP SMC SMD SMA CPI CTI B SB REV DSt BSt DTr DCF L LV L&S
Y Y Y N Y ? Y Y ? N ? ? ? ? ? ?
Current options are:
computation of transition-invariants
no computation of subinvariants
no computation of surinvariants
run automatically
output format = print non-zero entries with names
For net nr. 0.t :
15 transition-invariants written to INVARI.HLP
0 rows lost!
Formatting lines from INVARI.HLP
Current reduction options are:
Delete repeated occurrences of the same support
Do not delete covered sub/sur/invariants
Delete strict invariants covering sub/sur/invariants
transition sub/sur/invariants for net 0.t :
The net is covered by semipositive T-invariants.
semipositive transition invariants =
1 | 8.k1 : 1,
| 9.k2 : 1
2 | 18.k5 : 1,
| 19.k4 : 1
3 | 0.k3 : 1,

24 Gilbert, Heiner, Lehrack
| 1.k6 : 1,
| 8.k1 : 1,
| 19.k4 : 1
4 | 23.k8 : 1,
| 26.k7 : 1
5 | 22.k11 : 1,
| 27.k10 : 1
6 | 12.k16 : 1,
| 13.k17 : 1
7 | 16.k14 : 1,
| 17.k13 : 1
8 | 25.k20 : 1,
| 28.k19 : 1
9 | 24.k23 : 1,
| 29.k22 : 1
10 | 5.k12 : 1,
| 6.k15 : 1,
| 17.k13 : 1,
| 27.k10 : 1
11 | 20.k26 : 1,
| 21.k25 : 1
12 | 2.k21 : 1,
| 11.k30 : 1,
| 15.k28 : 1,
| 28.k19 : 1
13 | 14.k29 : 1,
| 15.k28 : 1
14 | 7.k24 : 1,
| 10.k27 : 1,
| 21.k25 : 1,
| 29.k22 : 1
15 | 3.k18 : 1,
| 4.k9 : 1,
| 12.k16 : 1,
| 26.k7 : 1
ORD HOM NBM PUR CSV SCF CON SC Ft0 tF0 Fp0 pF0 MG SM FC EFC ES
Y Y Y Y N N Y Y N N N N N N N N N
DTP SMC SMD SMA CPI CTI B SB REV DSt BSt DTr DCF L LV L&S
Y Y Y N Y Y Y Y ? N ? ? ? ? ? ?
Current analysis options are:
no symmetrical reduction
no stubborn reduction
no depth restriction
do not use a ’bad’ predicate
Computation of the reachability graph
States generated: 118
Arcs generated: 468

A Unifying Framework, TR I-02/2007 25
Capacities needed:
Place 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Cap: 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
The net has no dead transitions at the initial marking.
Searching for dynamic conflicts:
State Conflict
2: 0.k3 # 9.k2
The net is not dynamically conflict-free.
Computing the strongly connected components
The computed graph is strongly connected.
The net is reversible (resetable).
The net is live.
The net is live, if dead transitions are ignored.
The net is live and safe.
ORD HOM NBM PUR CSV SCF CON SC Ft0 tF0 Fp0 pF0 MG SM FC EFC ES
Y Y Y Y N N Y Y N N N N N N N N N
DTP SMC SMD SMA CPI CTI B SB REV DSt BSt DTr DCF L LV L&S
Y Y Y N Y Y Y Y Y N ? N N Y Y Y
Current options written to options.ina
End of Analyzer session.

26 Gilbert, Heiner, Lehrack
State Space Sizes
– as computed by the IDD-based model checker iddctl [Tov06]# nodes in the idd data structure,# states of the reachability graph
– the figures in brackets refer to PRISM [PNK06]
levchenko_N1
#nodes 52 (5.2e+01)
#states 118 (2)
RS computation time: 0m0.40 sec
levchenko_N4 -> (k=1, prism model, immediate response)
#nodes 115 (1.2e+02)
#states 24,065 (4)
RS computation time: 0m0.81 sec
levchenko_N8 -> (k=2, prism model, a few seconds)
#nodes 269 (2.7e+02)
#states 6,110,643 (6)
RS computation time: 0m0.210 sec
levchenko_N40 -> (k=10, prism model, prism dies silently after 10’)
#nodes 3697 (3.7e+03)
#states 478,293,389,221,095 (14)
RS computation time: 0m12.808 sec
levchenko_N80
#nodes 13472 (1.3e+04)
#states 5,634,903,909,420,423,840 (18)
RS computation time: 4m2.589 sec
levchenko_N120
#nodes 29347 (2.9e+04)
#states 1,714,249,381,410,772,340,816 (21)
RS computation time: 64m10.407 sec
levchenko_N160
???

A Unifying Framework, TR I-02/2007 27
Appendix B: Stochastic Model
PRISM Format [PNK06]
ctmc
const int k;
const int n = k * 4;
const double r = 0.4 / n;
// SPN coding style
module levchenko
// improvements possible by exploiting the p-invariants details
RasGTP : [ 0..n ] init k * 1;
Raf : [ 0..n ] init k * 4;
Raf_RasGTP : [ 0..n ] init 0;
RafP : [ 0..n ] init 0;
RafP_Phase1 : [ 0..n ] init 0;
MEK : [ 0..n ] init k * 2;
MEK_RafP : [ 0..n ] init 0;
MEKP : [ 0..n ] init 0;
MEKP_RafP : [ 0..n ] init 0;
MEKPP : [ 0..n ] init 0;
MEKPP_Phase2 :[ 0..n ] init 0;
MEKP_Phase2 : [ 0..n ] init 0;
ERK : [ 0..n ] init k * 3;
ERK_MEKPP : [ 0..n ] init 0;
ERKP : [ 0..n ] init 0;
ERKP_MEKPP : [ 0..n ] init 0;
ERKPP : [ 0..n ] init 0;
ERKPP_Phase3 :[ 0..n ] init 0;
ERKP_Phase3 : [ 0..n ] init 0;
Phase1 : [ 0..n ] init k * 3;
Phase2 : [ 0..n ] init k * 2;
Phase3 : [ 0..n ] init k * 3;
// this version brings the right results
// but it’s actually to strong because the postplaces shouldn’t overflow
[ r1 ] ( Raf > 0 ) & ( RasGTP > 0 ) & ( Raf_RasGTP < n )
-> 1.0 * Raf * RasGTP * r :
( Raf’ = Raf - 1 ) & ( RasGTP’ = RasGTP - 1 ) &
( Raf_RasGTP’ = Raf_RasGTP + 1 );
[ r2 ] ( Raf_RasGTP > 0 ) & ( Raf < n ) & ( RasGTP < n )

28 Gilbert, Heiner, Lehrack
-> 0.4 * Raf_RasGTP :
( Raf_RasGTP’ = Raf_RasGTP - 1 ) & ( Raf’ = Raf + 1 ) &
( RasGTP’ = RasGTP + 1 );
[ r3 ] ( Raf_RasGTP > 0 ) & ( RasGTP < n ) & ( RafP < n )
-> 0.1 * Raf_RasGTP :
( Raf_RasGTP’ = Raf_RasGTP - 1 ) & ( RasGTP’ = RasGTP + 1 ) &
( RafP’ = RafP + 1 );
[ r4 ] ( RafP > 0 ) & ( Phase1 > 0 ) & ( RafP_Phase1 < n )
-> 0.5 * RafP * Phase1 * r :
( RafP’ = RafP - 1 ) & ( Phase1’ = Phase1 - 1 ) &
( RafP_Phase1’ = RafP_Phase1 + 1 );
[ r5 ] ( RafP_Phase1 > 0 ) & ( RafP < n ) & ( Phase1 < n )
-> 0.5 * RafP_Phase1 :
( RafP_Phase1’ = RafP_Phase1 - 1 ) & ( RafP’ = RafP + 1 ) &
( Phase1’ = Phase1 + 1 );
[ r6 ] ( RafP_Phase1 > 0 ) & ( Raf < n ) & ( Phase1 < n )
-> 0.1 * RafP_Phase1 :
( RafP_Phase1’ = RafP_Phase1 - 1 ) & ( Raf’ = Raf + 1 ) &
( Phase1’ = Phase1 + 1 );
[ r7 ] ( RafP > 0 ) & ( MEK > 0 ) & ( MEK_RafP < n )
-> 3.3 * RafP * MEK * r :
( RafP’ = RafP - 1 ) & ( MEK’ = MEK - 1 ) &
( MEK_RafP’ = MEK_RafP + 1 );
[ r8 ] ( MEK_RafP > 0 ) & ( RafP < n ) & ( MEK < n )
-> 0.42 * MEK_RafP :
( MEK_RafP’ = MEK_RafP - 1 ) & ( RafP’ = RafP + 1 ) &
( MEK’ = MEK + 1 );
[ r9 ] ( MEK_RafP > 0 ) & ( RafP < n ) & ( MEKP < n )
-> 0.1 * MEK_RafP :
( MEK_RafP’ = MEK_RafP - 1 ) & ( RafP’ = RafP + 1 ) &
( MEKP’ = MEKP + 1 );
[ r10 ] ( MEKP > 0 ) & ( RafP > 0 ) & ( MEKP_RafP < n )
-> 3.3 * RafP * MEKP * r:
( MEKP’ = MEKP - 1 ) & ( RafP’ = RafP - 1 ) &
( MEKP_RafP’ = MEKP_RafP + 1 );
[ r11 ] ( MEKP_RafP > 0 ) & ( RafP < n ) & ( MEKP < n )
-> 0.4 * MEKP_RafP :
( MEKP_RafP’ = MEKP_RafP - 1 ) & ( RafP’ = RafP + 1 ) &
( MEKP’ = MEKP + 1 );
[ r12 ] ( MEKP_RafP > 0 ) & ( RafP < n ) & ( MEKPP < n )

A Unifying Framework, TR I-02/2007 29
-> 0.1 * MEKP_RafP :
( MEKP_RafP’ = MEKP_RafP - 1 ) & ( RafP’ = RafP + 1 ) &
( MEKPP’ = MEKPP + 1 );
[ r13 ] ( MEKPP > 0 ) & ( Phase2 > 0 ) & ( MEKPP_Phase2 < n )
-> 10 * MEKPP * Phase2 * r :
( MEKPP’ = MEKPP - 1 ) & ( Phase2’ = Phase2 - 1 ) &
( MEKPP_Phase2’ = MEKPP_Phase2 + 1 );
[ r14 ] ( MEKPP_Phase2 > 0 ) & ( MEKPP < n ) & ( Phase2 < n )
-> 0.8 * MEKPP_Phase2 :
( MEKPP_Phase2’ = MEKPP_Phase2 - 1 ) & ( MEKPP’ = MEKPP + 1 ) &
( Phase2’ = Phase2 + 1 );
[ r15 ] ( MEKPP_Phase2 > 0 ) & ( MEKP < n ) & ( Phase2 < n )
-> 0.1 * MEKPP_Phase2 :
( MEKPP_Phase2’ = MEKPP_Phase2 - 1 ) & ( MEKP’ = MEKP + 1 ) &
( Phase2’ = Phase2 + 1 );
[ r16 ] ( MEKP > 0 ) & ( Phase2 > 0 ) & ( MEKP_Phase2 < n )
-> 10 * MEKP * Phase2 * r :
( MEKP’ = MEKP - 1 ) & ( Phase2’ = Phase2 - 1 ) &
( MEKP_Phase2’ = MEKP_Phase2 + 1 );
[ r17 ] ( MEKP_Phase2 > 0 ) & ( MEKP < n ) & ( Phase2 < n )
-> 0.8 * MEKP_Phase2 :
( MEKP_Phase2’ = MEKP_Phase2 - 1 ) & ( MEKP’ = MEKP + 1 ) &
( Phase2’ = Phase2 + 1 );
[ r18 ] ( MEKP_Phase2 > 0 ) & ( MEK < n ) & ( Phase2 < n )
-> 0.1 * MEKP_Phase2 :
( MEKP_Phase2’ = MEKP_Phase2 - 1 ) & ( MEK’ = MEK + 1 ) &
( Phase2’ = Phase2 + 1 );
[ r19 ] ( ERK > 0 ) & ( MEKPP > 0 ) & ( ERK_MEKPP < n )
-> 20 * ERK * MEKPP * r :
( ERK’ = ERK - 1 ) & ( MEKPP’ = MEKPP - 1 ) &
( ERK_MEKPP’ = ERK_MEKPP + 1 );
[ r20 ] ( ERK_MEKPP > 0 ) & ( ERK < n ) & ( MEKPP < n )
-> 0.6 * ERK_MEKPP :
( ERK_MEKPP’ = ERK_MEKPP - 1 ) & ( ERK’ = ERK + 1 ) &
( MEKPP’ = MEKPP + 1 );
[ r21 ] ( ERK_MEKPP > 0 ) & ( ERKP < n ) & ( MEKPP < n )
-> 0.1 * ERK_MEKPP :
( ERK_MEKPP’ = ERK_MEKPP - 1 ) & ( ERKP’ = ERKP + 1 ) &
( MEKPP’ = MEKPP + 1 );
[ r22 ] ( ERKP > 0 ) & ( MEKPP > 0 ) & ( ERKP_MEKPP < n )

30 Gilbert, Heiner, Lehrack
-> 20 * ERKP * MEKPP * r :
( ERKP’ = ERKP - 1 ) & ( MEKPP’ = MEKPP - 1 ) &
( ERKP_MEKPP’ = ERKP_MEKPP + 1 );
[ r23 ] ( ERKP_MEKPP > 0 ) & ( ERKP < n ) & ( MEKPP < n )
-> 0.6 * ERKP_MEKPP :
( ERKP_MEKPP’ = ERKP_MEKPP - 1 ) & ( ERKP’ = ERKP + 1 ) &
( MEKPP’ = MEKPP + 1 );
[ r24 ] ( ERKP_MEKPP > 0 ) & ( ERKPP < n ) & ( MEKPP < n )
-> 0.1 * ERKP_MEKPP :
( ERKP_MEKPP’ = ERKP_MEKPP - 1 ) & ( ERKPP’ = ERKPP + 1 ) &
( MEKPP’ = MEKPP + 1 );
[ r25 ] ( ERKPP > 0 ) & ( Phase3 > 0 ) & ( ERKPP_Phase3 < n )
-> 5 * ERKPP * Phase3 *r :
( ERKPP’ = ERKPP - 1 ) & ( Phase3’ = Phase3 - 1 ) &
( ERKPP_Phase3’ = ERKPP_Phase3 + 1 );
[ r26 ] ( ERKPP_Phase3 > 0 ) & ( ERKPP < n ) & ( Phase3 < n )
-> 0.4 * ERKPP_Phase3 :
( ERKPP_Phase3’ = ERKPP_Phase3 - 1 ) & ( ERKPP’ = ERKPP + 1 ) &
( Phase3’ = Phase3 + 1 );
[ r27 ] ( ERKPP_Phase3 > 0 ) & ( ERKP < n ) & ( Phase3 < n )
-> 0.1 * ERKPP_Phase3 :
( ERKPP_Phase3’ = ERKPP_Phase3 - 1 ) & ( ERKP’ = ERKP + 1 ) &
( Phase3’ = Phase3 + 1 );
[ r28 ] ( ERKP > 0 ) & ( Phase3 > 0 ) & ( ERKP_Phase3 < n )
-> 5 * ERKP * Phase3 * r :
( ERKP’ = ERKP - 1 ) & ( Phase3’ = Phase3 - 1 ) &
( ERKP_Phase3’ = ERKP_Phase3 + 1 );
[ r29 ] ( ERKP_Phase3 > 0 ) & ( ERKP < n ) & ( Phase3 < n )
-> 0.4 * ERKP_Phase3 :
( ERKP_Phase3’ = ERKP_Phase3 - 1 ) & ( ERKP’ = ERKP + 1 ) &
( Phase3’ = Phase3 + 1 );
[ r30 ] ( ERKP_Phase3 > 0 ) & ( ERK < n ) & ( Phase3 < n )
-> 0.1 * ERKP_Phase3 :
( ERKP_Phase3’ = ERKP_Phase3 - 1 ) & ( ERK’ = ERK + 1 ) &
( Phase3’ = Phase3 + 1 );
endmodule

A Unifying Framework, TR I-02/2007 31
Appendix C: Continuous Model
– as produced by Snoopy [Sno]
Table 1. ODE equation parameter
k1 = 1.0
k2 = 0.4
k3 = 0.1
k4 = 0.5
k5 = 0.5
k6 = 0.1
k7 = 3.3
k8 = 0.42
k9 = 0.1
k10 = 3.3
k11 = 0.4
k12 = 0.1
k13 = 10.0
k14 = 0.8
k15 = 0.1
k16 = 10.0
k17 = 0.8
k18 = 0.1
k19 = 20.0
k20 = 0.6
k21 = 0.1
k22 = 20.0
k23 = 0.6
k24 = 0.1
k25 = 5.0
k26 = 0.4
k27 = 0.1
k28 = 5.0
k29 = 0.4
k30 = 0.1
Table 2. Initial concentrations
Species μMLev μMPN
Raf 0.4 1RasGTP 0.1 1Raf RasGTP 0 0RafP 0 0RafP Phase1 0 0Phase1 0.3 1MEK 0.2 1MEKP 0 0MEKPP 0 0MEK RafP 0 0MEKP RafP 0 0
Species μMLev μMPN
MEKP Phase2 0 0MEKPP Phase2 0 0Phase2 0.2 1ERK 0.3 1ERKP 0 0ERKPP 0 0ERK MEKPP 0 0ERKP MEKPP 0 0ERKP Phase3 0 0ERKPP Phase3 0 0Phase3 0.3 1

32 Gilbert, Heiner, Lehrack
dRafdt
= k2 ∗ Raf RasGTP + k6 ∗ RafP Phase1 − k1 ∗ Raf ∗ RasGTP
dRasGTP
dt= k2 ∗ Raf RasGTP + k3 ∗ Raf RasGTP − k1 ∗ Raf ∗ RasGTP
dRaf RasGTPdt
= k1 ∗ Raf ∗ RasGTP − k2 ∗ Raf RasGTP − k3 ∗ Raf RasGTP
dRafPdt
= k3 ∗ Raf RasGTP + k12 ∗ MEKP RafP + k9 ∗ MEK RafP+k5 ∗ RafP Phase1 + k8 ∗ MEK RafP + k11 ∗ MEKP RafP−k7 ∗ RafP ∗ MEK − k10 ∗ MEKP ∗ RafP − k4 ∗ Phase1 ∗ RafP
dRafP Phase1dt
= k4 ∗ Phase1 ∗ RafP − k5 ∗ RafP Phase1 − k6 ∗ RafP Phase1
dMEK RafPdt
= k7 ∗ RafP ∗ MEK − k8 ∗ MEK RafP − k9 ∗ MEK RafP
dMEKP RafPdt
= k10 ∗ MEKP ∗ RafP − k11 ∗ MEKP RafP − k12 ∗ MEKP RafP
dMEKP Phase2dt
= k16 ∗ Phase2 ∗ MEKP − k18 ∗ MEKP Phase2 − k17 ∗ MEKP Phase2
dMEKPP Phase2dt
= k13 ∗ MEKPP ∗ Phase2 − k15 ∗ MEKPP Phase2 − k14 ∗ MEKPP Phase2
dERKdt
= k20 ∗ ERK MEKPP + k30 ∗ ERKP Phase3 − k19 ∗ MEKPP ∗ ERK
dERK MEKPPdt
= k19 ∗ MEKPP ∗ ERK − k20 ∗ ERK MEKPP − k21 ∗ ERK MEKPP
dERKP MEKPPdt
= k22 ∗ MEKPP ∗ ERKP − k24 ∗ ERKP MEKPP − k23 ∗ ERKP MEKPP
dERKPdt
= k21 ∗ ERK MEKPP + k23 ∗ ERKP MEKPP + k29 ∗ ERKP Phase3+k27 ∗ ERKPP Phase3 − k28 ∗ ERKP ∗ Phase3 − k22 ∗ MEKPP ∗ ERKP
dMEKPPdt
= k12 ∗ MEKP RafP + k20 ∗ ERK MEKPP + k23 ∗ ERKP MEKPP+k14 ∗ MEKPP Phase2 + k21 ∗ ERK MEKPP + k24 ∗ ERKP MEKPP−k19 ∗ MEKPP ∗ ERK − k22 ∗ MEKPP ∗ ERKP − k13 ∗ MEKPP ∗ Phase2
dERKPP Phase3dt
= k25 ∗ Phase3 ∗ ERKPP − k27 ∗ ERKPP Phase3 − k26 ∗ ERKPP Phase3
dERKP Phase3dt
= k28 ∗ ERKP ∗ Phase3 − k29 ∗ ERKP Phase3 − k30 ∗ ERKP Phase3
dMEKPdt
= k9 ∗ MEK RafP + k11 ∗ MEKP RafP + k17 ∗ MEKP Phase2+k15 ∗ MEKPP Phase2 − k16 ∗ Phase2 ∗ MEKP − k10 ∗ MEKP ∗ RafP
dERKPPdt
= k24 ∗ ERKP MEKPP + k26 ∗ ERKPP Phase3 − k25 ∗ Phase3 ∗ ERKPP
dPhase2dt
= k17 ∗ MEKP Phase2 + k14 ∗ MEKPP Phase2 + k18 ∗ MEKP Phase2+k15 ∗ MEKPP Phase2 − k16 ∗ Phase2 ∗ MEKP − k13 ∗ MEKPP ∗ Phase2
dPhase3dt
= k29 ∗ ERKP Phase3 + k26 ∗ ERKPP Phase3 + k30 ∗ ERKP Phase3+k27 ∗ ERKPP Phase3 − k28 ∗ ERKP ∗ Phase3 − k25 ∗ Phase3 ∗ ERKPP
dMEKdt
= k8 ∗ MEK RafP + k18 ∗ MEKP Phase2 − k7 ∗ RafP ∗ MEK
dPhase1dt
= k5 ∗ RafP Phase1 + k6 ∗ RafP Phase1 − k4 ∗ Phase1 ∗ RafP

A Unifying Framework, TR I-02/2007 33
Table 3. Initial ‘good’ state configurations ( 10 of 118 ).
S1 S2 S3 S4 S5 S6 S7 S8 S9 S10 . . .Raf 1 0 0 0 0 0 0 0 0 0RasGTP 1 0 1 1 1 1 1 1 1 1Raf RasGTP 0 1 0 0 0 0 0 0 0 0Rafx 0 0 1 0 1 0 1 1 1 1Rafx Phase1 0 0 0 0 0 0 0 0 0 0Phase1 1 1 1 1 1 1 1 1 1 1MEK 1 1 1 0 0 0 0 0 0 0MEKP 0 0 0 0 1 0 0 0 0 0MEKPP 0 0 0 0 0 0 1 0 1 0MEK Rafx 0 0 0 1 0 0 0 0 0 0MEKP Rafx 0 0 0 0 0 1 0 0 0 0MEKP Phase2 0 0 0 0 0 0 0 0 0 0MEKPP Phase2 0 0 0 0 0 0 0 0 0 0Phase2 1 1 1 1 1 1 1 1 1 1ERK 1 1 1 1 1 1 1 0 0 0ERKP 0 0 0 0 0 0 0 0 1 0ERKPP 0 0 0 0 0 0 0 0 0 0ERK MEKPP 0 0 0 0 0 0 0 1 0 0ERKP MEKPP 0 0 0 0 0 0 0 0 0 1ERKP Phase3 0 0 0 0 0 0 0 0 0 0ERKPP Phase3 0 0 0 0 0 0 0 0 0 0Phase3 1 1 1 1 1 1 1 1 1 1
0 50 100 150 200 250 300 3500
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Time (sec)
Con
cent
ratio
n (r
elat
ive
units
)
State1
Raf
RKIP
RasGTP
Raf_RasGTP
Rafx
Rafx_Phase1
MEK_Rafx
MEKP_Rafx
MEKP_Phase2
MEKPP_Phase2
ERK
ERK_MEKPP
ERKP_MEKPP
ERKP
MEKPP
ERKPP_Phase3
ERKP_Phase3
MEKP
ERKPP
Phase2
Phase3
MEK
Phase1
Fig. 8. Dynamic behaviour for state 1, compare Table 3.

34 Gilbert, Heiner, Lehrack
Table 4. Mean values for the steady states of the 118 ‘good’ initial states
Species Mean steady state concentrationRaf 0.101195RasGTP 0.831676Raf RasGTP 0.168324Rafx 0.242869Rafx Phase1 0.168324Phase1 0.831676MEK 0.170931MEK Rafx 0.263454MEKP 0.0348324MEKPP 0.00738116MEKP Rafx 0.0558341MEKP Phase2 0.263454MEKPP Phase2 0.0558344Phase2 0.680712ERK 0.686032ERK MEKPP 0.144696ERKP MEKPP 0.00358319ERKP 0.0169886ERKPP 0.000420698ERKPP Phase3 0.00358318ERKP Phase3 0.144696Phase3 0.851721
0 50 100 150 200 250 300 3500
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Time (sec)
Con
cent
ratio
n (r
elat
ive
units
)
State10
Raf
RKIP
RasGTP
Raf_RasGTP
Rafx
Rafx_Phase1
MEK_Rafx
MEKP_Rafx
MEKP_Phase2
MEKPP_Phase2
ERK
ERK_MEKPP
ERKP_MEKPP
ERKP
MEKPP
ERKPP_Phase3
ERKP_Phase3
MEKP
ERKPP
Phase2
Phase3
MEK
Phase1
Fig. 9. Dynamic behaviour for state 10, compare Table 3.

A Unifying Framework, TR I-02/2007 35
References
[ADLS06] D. Angeli, P. De Leenheer, and E.D. Sontag. On the structural mono-tonicity of chemical reaction networks. In Proc. 45th IEEE Conference onDecision and Control, pages 7–12, 2006.
[APUM03] M. Antoniotti, A. Policriti, N. Ugel, and B. Mishra. Model building andmodel checking for biochemical processes. Cell Biochemistry and Bio-physics, 38:271–286, 2003.
[Bio] BioNessie. A biochemical pathway simulation and analysis tool. Universityof Glasgow, www.bionessie.org.
[BK02] F. Bause and P.S. Kritzinger. Stochastic Petri Nets. Vieweg, 2002.
[CCRFS06] L. Calzone, N. Chabrier-Rivier, F. Fages, and S. Soliman. Machine learningbiochemical networks from temporal logic properties. Trans. on Computat.Syst. Biol. VI, LNBI 4220, pages 68–94, 2006.
[CDGH06] M. Calder, A. Duguid, S. Gilmore, and J. Hillston. Stronger computationalmodelling of signalling pathways using both continuous and discrete-statemethods. In Proc. CMSB 2006, pages 63–78. LNBI 4210, Springer, 2006.
[CF03] N. Chabrier and F. Fages. Symbolic model checking of biochemical net-works. In Proc. CMSB 2003, pages 149–162. LNCS 2602, Springer, 2003.
[CGH05] M. Calder, S. Gilmore, and J. Hillston. Automatically deriving ODEs fromprocess algebra models of signalling pathways. In Proc. CSMB 2005, pages204–215. LFCS, Univ. of Edinburgh, 2005.
[CGP01] E.M. Clarke, O. Grumberg, and D.A. Peled. Model checking. MIT Press1999, third printing, 2001.
[CKS07] V. Chickarmane, B. N. Kholodenko, and H. M. Sauro. Oscillatory dynamicsarising from competitive inhibition and multisite phosphorylation. Journalof Theoretical Biology, 244(1):68–76, January 2007.
[Cur06] E. Curry. Stochastic Simulation of the Entrained Circadian Rhythm. Mas-ter thesis, School of Informatics, Univ. of Edinburgh, 2006.
[CVGO06] M. Calder, V. Vyshemirsky, D. Gilbert, and R. Orton. Analysis of signallingpathways using continuous time Markov chains. Trans. on Computat. Syst.Biol. VI, LNBI 4220, pages 44–67, 2006.
[DDS04] D. D’Aprile, S. Donatelli, and J. Sproston. CSL model checking for theGreatSPN tool. In Proc. ISCIS 2004, LNCS 3280, 2004.
[DP80] J. R. Dormand and P. J. Prince. A family of embedded runge-kutta for-mulae. J. Comp. Appl. Math., 6:1–22, 1980.
[EKL+02] S. Eker, M. Knapp, K. Laderoute, P. Lincoln, J. Meseguer, and K. Sonmez.Pathway logic: Symbolic analysis of biological signaling. In Proc. SeventhPacific Symposium on Biocomputing, pages 400–412, 2002.
[FR07] F. Fages and A. Rizk. On the analysis of numerical data time series intemporal logic. In Proc. CMSB 2007. LNCS/LNBI 4695, Springer, 2007.
[GH06] D. Gilbert and M. Heiner. From Petri nets to differential equations - anintegrative approach for biochemical network analysis. In Proc. ICATPN2006, pages 181–200. LNCS 4024, Springer, 2006.
[GHL07] D. Gilbert, M. Heiner, and S. Lehrack. A unifying framework for modellingand analysing biochemical pathways using Petri nets. In Proc. CMSB 2007.LNCS/LNBI 4695, Springer, 2007.
[Gil77] D.T. Gillespie. Exact stochastic simulation of coupled chemical reactions.The Journal of Physical Chemistry, 81(25):2340–2361, 1977.

36 Gilbert, Heiner, Lehrack
[GP98] P. J. E. Goss and J. Peccoud. Quantitative modeling of stochastic systemsin molecular biology by using stochastic Petri nets. Proc. Natl. Acad. Sci.USA, 95(June):2340–2361, 1998.
[HKN+06] J. Heath, M. Kwiatkowska, G. Norman, D. Parker, and O. Tymchyshyn.Probabilistic model checking of complex biological pathways. In Proc.CMSB 2006, pages 32–47. LNBI 4210, Springer, 2006.
[LBS00] A. Levchenko, J. Bruck, and P.W. Sternberg. Scaffold proteins may bipha-sically affect the levels of mitogen-activated protein kinase signaling andreduce its threshold properties. Proc Natl Acad Sci USA, 97(11):5818–5823,2000.
[Leh06] S. Lehrack. Three Petri net approaches for biochemical network analysis.TR I-01, Dep. CS, BTU Cottbus, 2006.
[MBC+95] M. Ajmone Marsan, G. Balbo, G. Conte, S. Donatelli, and G. Franceschinis.Modelling with Generalized Stochastic Petri Nets. Wiley Series in ParallelComputing, John Wiley and Sons, 1995. 2nd Edition.
[MGF] Max-Gruenebaum-Foundation. www.max-gruenebaum-stiftung.de.[Mur89] T. Murata. Petri nets: Properties, analysis and applications. Proc.of the
IEEE 77, 4:541–580, 1989.[PNK06] D. Parker, G. Norman, and M. Kwiatkowska. PRISM 3.0.beta1 Users’
Guide, 2006.[Sno] Snoopy. A tool to design and animate hierarchical graphs. BTU Cottbus,
CS Dep., www-dssz.informatik.tu-cottbus.de.[SPB01] R. Srivastava, M.S. Peterson, and W.E. Bentley. Stochastic kinetic analysis
of the escherichia coli stress circuit using σ32-targeted antisense. Biotech-nology and Bioengineering, 75(1):120–129, 2001.
[SR97] L. F. Shampine and M. W. Reichelt. The MATLAB ODE Suite. SIAMJournal on Scientific Computing, 18:1–22, 1997.
[SR99] P.H. Starke and S. Roch. INA - The Intergrated Net Analyzer. HumboldtUniversity Berlin, www.informatik.hu-berlin.de/∼starke/ina.html, 1999.
[SSE04] C. Schroter, S. Schwoon, and J. Esparza. The Model Checking Kit. InProc. ICATPN, LNCS 2697, pages 463–472. Springer, 2004.
[SSW05] O.J. Shaw, L.J. Steggles, and A. Wipat. Automatic parameterisation ofstochastic Petri net models of biological networks. CS-TR-909, School ofCS, Univ. of Newcastle upon Tyne, 2005.
[ST05] O. Schulz-Trieglaff. Modelling the randomness in biological systems. Mas-ter thesis, School of Informatics, University of Edinburgh, 2005.
[Tov06] Tovchigrechko, Alex. Model checking using interval decision diagrams. PhDthesis, BTU Cottbus, Dep. of CS, 2006.
[TSB04] T. E. Turner, S. Schnell, and K. Burrage. Stochastic approaches for mod-elling in vivo reactions. Comp. Biology and Chemistry, 28(3):165–178, 2004.
[Wil06] D.J. Wilkinson. Stochastic Modelling for System Biology. CRC Press, NewYork, 1st Edition, 2006.
[WMS05] A. Sujathab W. Marwan and C. Starostzik. Reconstructing the regula-tory network controlling commitment and sporulation in Physarum poly-cephalum based on hierarchical Petri net modeling and simulation. J. ofTheoretical Biology, 236(4):349–365, 2005.
Remark The data files of the running case study in its three versions and the analysisresults are available at
www-dssz.informatik.tu-cottbus.de/examples/levchenko.