a pyrethroid-treated bed net increases host attractiveness for
TRANSCRIPT

1
Apyrethroïd-treatedbednetincreaseshostattractiveness1
forAnophelesgambiaes.s.carryingthekdralleleinadual-2
choiceolfactometer3
Angélique PORCIANI1,2,*,¶, Malal DIOP1,2,¶, Nicolas MOIROUX3, Tatiana KADOKE-LAMBI2, Anna4
COHUET3,FabriceCHANDRE3,LaurentDORMONT4,CédricPENNETIER1,25
1: MiVEGEC (UM1-UM2-CNRS 5290-IRD 224), Institut de Recherche pour le Développement (IRD)6
Cotonou,Bénin.7
2:CentredeRechercheEntomologiquedeCotonou(CREC),Bénin8
3:MiVEGEC (UM1-UM2-CNRS 5290-IRD 224), Institut de Recherche pour le Développement (IRD),9
Montpellier,France.10
4:CNRSUMR5175,Centred’EcologieFonctionnelleetEvolutive(CEFE),Montpellier,France.11
*Correspondingauthors12
Email:[email protected]
¶Theseauthorscontributedequallytothiswork14
15
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

2
Abstract16
Theuseoflonglastinginsecticidenets(LLINs)treatedwithpyrethroïdisknownforitsmajor17
contributioninmalariacontrol.However,LLINsaresuspectedtoinducebehavioralchangesin18
malariavectors,whichmayinturndrasticallyaffecttheirefficacyagainstPlasmodiumsp.19
transmission.InsubSaharanAfrica,wheremalariaimposestheheaviestburden,themainmalaria20
vectorsarewidelyresistanttopyrethroïds,theinsecticidefamilyusedonLLINs,whichalso21
threatensLLINefficiency.Thereisthereforeacrucialneedfordecipheringhowinsecticide-22
impregnatedmaterialsmightaffectthehost-seekingbehaviorofmalariavectorsinregardsto23
insecticideresistance.Inthisstudy,weexploredtheimpactofpermethrin-impregnatednetonthe24
hostattractivenessforAnophelesgambiaemosquitoes,eithersusceptibletoinsecticides,orcarrying25
theinsecticideresistanceconferringallelekdr.Groupsoffemalemosquitoeswerereleasedinadual-26
choiceolfactometerandtheirmovementstowardsanattractiveodorsource(arabbit)protectedby27
insecticide-treated(ITN)oruntreatednets(UTN)weremonitored.Kdrhomozygousmosquitoes,28
resistanttoinsecticides,weremoreattractedbyahostbehindanITNthananUTN,whilethe29
presenceofinsecticideonthenetdidnotaffectthechoiceofsusceptiblemosquitoes.Theseresults30
suggestthatpermethrin-impregnatednetisdetectablebymalariavectorsandthatthekdrmutation31
impactstheirresponsetoaLLINprotectedhost.Wediscusstheimplicationoftheseresultsfor32
malariavectorcontrol.33
34
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

3
Introduction35
Anophelesgambiaeisoneofthemajormosquitovectorsofhumanmalariaparasitesinsub-36
SaharanAfrica.Itsremarkablevectorialcapacity[1]mainlyreliesonitshighdegreeofanthropophily.37
Moreover,An.gambiaepreferstobitehumansindoorsandoftenrestsinsidehousesafterblood38
feeding[2–4].Thesebehavioralpreferencesledtothedevelopmentofinsecticide-basedindoor39
vectorcontrolmeasures,suchasinsecticide-treatedbednets(ITNs)andindoorresidualspraying40
(IRS),tolimitthehuman-vectorcontactsandreducemosquitosurvival.Todate,fourinsecticide41
familiesareavailableforIRS(organochlorides,organophosphates,carbamatesandpyrethroïds),42
whereasonlypyrethroïdsarerecommendedformosquitonetsbecauseoftheirlowmammalian43
toxicityandhighinsecticidalpotency[5].44
Tokill,insecticidemoleculesmustcontactandpenetratethroughthemosquitocuticle/gut45
tothenreachandinteractwiththeirtargetbeforebeingdegraded.Anyphysiologicalorbehavioral46
mechanismthatmayinterferewithoneofthesestepscanleadtoinsecticideresistance.The47
widespreaduseofpyrethroïd(PYR)insecticidesinmalariavectorcontrolandagriculturehasfavored48
thedevelopmentofresistanceinmalariavectorspecies[6].Oneofthemoststudiedphysiological49
mechanismsinvolvedinPYRresistanceisthereducedsensitivityofthevoltage-gatedsodium50
channelstoPYRbindingcausedbynon-silentmutations,knownasknockdownresistance(kdr)51
mutations[7].BehavioralresistanceisanothermechanisminvolvedinPYRresistance.Thiscanbe52
definedasamodificationofthemosquitobehaviortoavoidcontactwithalethaldoseofinsecticide53
[8].Todate,behavioralresistancetoinsecticidesremainspoorlydocumented,despiteofitshuge54
potentialimpactonmalariatransmission.55
Behavioraladaptationstopesticidescanbeclassifiedasstimulus-dependentor-independent56
[9].Stimulus-independentadaptationsarenotassociatedwiththeperceptionofchemicals,butmore57
probablywithmodificationsofthevectorintrinsicbehavior,suchaschangesinhost-seeking58
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

4
behaviorpreferences(levelofanthropophily,endophagy,endophilyorhourlybitingactivities).Such59
behavioralmodificationshaverecentlybeenobservedinthecontextofITNwidespreaduse:60
mosquitovectorsmaypostponetheirbloodfeedinguntilthemorning,whenhumanhostsare61
protectedbyITNsanymore[10–12].Thesechangesmaylimitthecontactbetweenaggressive62
malariavectorsandtreatedsurfaces,thusthreateningtheefficiencyofindoorvectorcontroltools.63
Conversely,stimulus-dependentbehavioraladaptationsarespecificallylinkedtothedetectionof64
chemicals.Stimulus-dependentinsecticideavoidancecanbedefinedasa“flyaway”behaviorto65
leavetheimmediatetoxicenvironmentaftercontact(irritancy)ornot(repellence)withthetreated66
surface[13–15].AvoidancebehaviorfollowingcontactwithPYRhasbeenreportedinsomecases67
[16–20],butsimilarbehaviorintheabsenceofdirectcontactwiththeinsecticidehasbeenpoorly68
documented.OnlyindirectobservationssuggestadetectionandavoidanceofITNsbymalaria69
vectors:mosquitoentryrateswerefoundreducedinexperimentalhutscontaininginsecticide-70
treatednetscomparedtoentryratesincontrolhuts,moreovertheobservedratesweredependent71
onthekdrallelepresenceinthemosquitoes[21–23].Althoughtheeffectsofpyrethroïdson72
differentpartofhostseekingbehaviorhasbeenalreadystudied[20,24–26],theirinfluenceonthe73
relativehostattractivenesshasbeenneglecteddespiteitsimportanceinhostchoiceandonmalaria74
transmission.Therefore,inordertoadequatelyevaluateanduseITNs,ithasbecomeurgentto75
investigatethepossiblemodulationofthehost-seekingbehaviorinpresenceofindoorvectorcontrol76
toolsinregardstootherinsecticideresistancemechanisms77
Inthisstudy,weexaminedthelong-rangehost-seekingbehaviorofAn.gambiaemosquitoes78
todeterminewhethertheattractivenessofavertebratehost(arabbit)inadual-choiceolfactometer79
wasinfluencedbyphysicaland/orchemicalbarriers(insecticide-treatedanduntreatednets)andby80
themosquitokdr(L1014F)genotype.81
82
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

5
Methods83
Ethicsstatement84
RabbitswerehandledandblooddrawninaccordancetotheprotocolapprovedbyNationalComity85
forEthicandResearch(CNERS)andHealthministryofBenin(N°023).Thisstudywascarriedoutin86
strictaccordancewiththerecommendationsofAnimalCareandUseCommitteenamed“Comité87
d’éthiquepourl’expérimentationanimale;LanguedocRoussillon”andtheprotocolwasapprovedby88
theCommitteeontheEthicsofAnimalExperiments(CEEA-LR-13002fortherabbits).Rabbitswere89
notsubjectedtoanesthesia,analgesiaorsacrifice.90
Mosquitoes91
TwolaboratoryreferencestrainsofAnophelesgambiaesensustricto(formerlycalledSmolecular92
form)(20)wereusedinthisstudy.TheKisumureferencestrain,isolatedinKenyain197593
(VectorBase,http://www.vectorbase.org,KISUMU1),isfreeofanydetectableinsecticideresistance94
mechanism.Thekdr-kisstrainwasobtainedbyintrogressionintotheKisumugenomeofthekdr-west95
allele(L1014F)[27]thatoriginatedfromaPYR-resistantpopulationcollectedinKouValley,Burkina96
Faso,whichwasusedtoestablishastrainnamedVKPer.Introgressionwasobtainedthrough1997
successiveback-crossesbetweenKisumuandVKPer[28].VKPerstraindisplayedthesameexpression98
levelofmetabolicresistanceenzymeasKisumu[29].Kisumuandkdr-kismosquitoesaretherefore99
homozygoussusceptible(SS)andhomozygousresistant(RR)atthekdrlocus,respectively. The100
heterozygousgenotypeRSwasobtainedbycrossingKisumuSSfemaleswithkdr-kisRRmales.101
Mosquitoeswererearedininsectaryconditions(27±3°C,60-80%relativehumidityanda12:12light102
anddarkcycle).Groundcatfoodwasusedtofeedlarvaeand10%sucrosesolution(withrabbit103
bloodtwiceperweek)tofeedadultfemales.Forbehavioralexperiments,5-12dayoldfemales,104
withoutprioraccesstoabloodmeal,werestarvedfor4hbeforetheassay.105
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

6
Experimentalset-up106
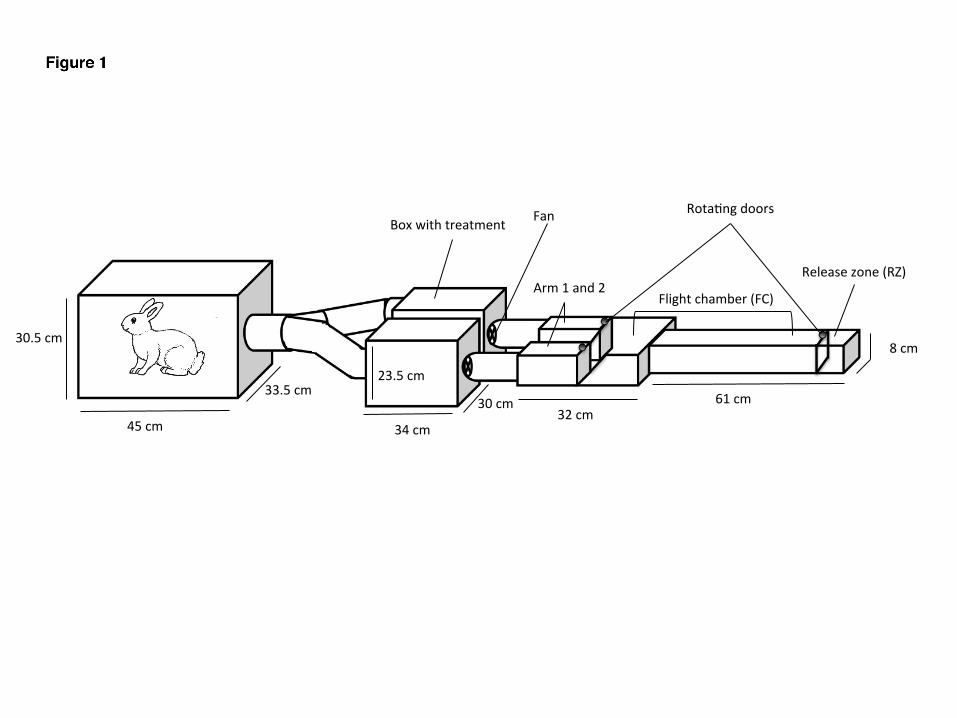
Thedual-choiceolfactometerwasadaptedfromGeierandBoeckh(1999)[30].Itwasmadeof107
Plexiglasandwasdividedinfourparts:releasezone(RZ),flightchamber(FC)andonecollectingzone108
ineachofthetwoarms(A1orA2)(Fig1).RotatingdoorsmadefrommeshgauzeintheRZandin109
botharmsallowedmosquitoreleaseorcapture.Theupwindpartoftheexperimentalset-upwas110
composedofawidechamberwhereanattractivehost(arabbit)canbeplaced,andthatwas111
connectedtotwotreatmentboxesthatcontainedornotthenets.Eachtreatmentboxwas112
connectedtoonearmoftheolfactometer.Inordertoavoidanyperturbationontheairflowbythe113
treatment,fanswereplacedonthedownwindfacesoftheexperimentboxesandextractedtheair114
fromthetreatmentboxestotheolfactometer,providingtheodor-ladenaircurrent.Atthebeginning115
ofeachexperiment,theairflowwasmeasuredinarm1and2andinthereleasezoneusingaTesto©116
435-1multifunctionalmeter(Testo,Forbach,France)andthermo-anemometricprobe(m.s-1)and117
adjustedat0.20±0.03m.s-1.Duringtheexperiment,athickblacktarpaulincoveredtheolfactometer118
tokeepallthesystemindarknessandavoidvisualdisturbance.119
Fig1.Experimentalset-up.120Dual-choiceolfactometer(rightside)connectedtothetreatmentboxes(middle)andthewide121chamber(leftside).122123
Experimentaldesign124
Fourexperiments,summarizedinTable1,wereperformedusingSS,RRandRSmosquitoes.The125
treatmentboxesandthewidechamberwereemptyduringthefirstexperiment.Fortheother126
experiments,thewidechambercontainedarabbitasodorsource.Thetreatmentboxescontained,127
dependingontheexperiment,nothingor2m2ofuntreated(UTN)orinsecticide-treatednet(ITN,128
Olyset®Netimpregnatedwith1000mg/m2ofpermethrin).Netsweredividedin50piecesof129
20x20cmandhungonametallicstructureperpendicularlytotheairflow.Thesamenetswereused130
duringallexperiment,theOlyset®wasconservedat4°Cbetweeneachdayofexperiment.Thenets131
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

7
wereplacedinboxesthatcouldnotbevisibleformosquitoes,sothatnovisualclueswereavailable132
tomosquitoesduringtheexperiments.133
Table1:Descriptionoftheexperimentaldesign134
135
136
137
138
(UTN:untreatednet,ITN:insecticide-treatednet)139
Assaysforthefourexperimentswereperformedeverydayfor20daysbetween10:00amand140
14:00pm(correspondingtomosquitostrainfeedingtimeinlaboratory).Wealwaysstartedwith141
assaysofexperiment1,tocheckpossibleodororinsecticidecontaminations.Whenpossible(i.e.,142
whentheinsectaryproductionwassufficient),femalesofthethreegenotypesweretestedthesame143
dayforthefourexperiments,otherwiseatleasttwogenotypesweretestedthesameday(a144
summaryoftheassaysispresentedinsupplementarydata).Eachday,inassaysforexperiments3145
and4,treatmentswererotatedonetimebetweenboxestopreventanyarmeffect.Between146
rotations,theboxeswerecarefullycleanedwithethanoltoavoidanyresidualinsecticideeffect.147
Moreover,theolfactometerwascleanedwithethanoleveryday.Theexperimenterworelatex148
glovestoavoidcontamination.Thesamerabbitwasusedasodorsourceduringallthestudy.Itwasa149
1-yearoldfemalerearedinthesameconditionsasthoseusedininsectariestofeedmosquitoes.CO2150
concentrationandrelativehumidity(RH)weremonitoredineacharmswithaTesto©435-1151
multifunctionalmeter(Testo,Forbach,France)equippedwithanIndoorAirQuality(IAQ)probe152
[%RH;range:0to+100%RH;accuracy:±2%RH(+2to+98%RH)],[CO2;range:0to+10000ppm;153
accuracy:(±75ppmCO2±3%ofmv)(0to+5000ppmCO2)].Theroomwaskeptataconstant154
temperatureof25°Cduringthestudy.155
Experimentno.
Experimentname
Odorsource Treatment box1
Treatment box2
1 Empty None Empty Empty2 Rabbitalone Rabbit Empty Empty3 Rabbit+UTN Rabbit Empty UTN UTN Empty4 Rabbit+ITN Rabbit ITN UTN UTN ITN
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

8
Foreachassay,abatchof20-23femaleswasreleasedintheRZfor5minforacclimation.The156
rotatingdoorswerethenopenedandfemaleswerefreetoflyintheolfactometer.After5minutes,157
therotatingdoorswereclosedandthenumbersofmosquitoesinRZ(NRZ),FC(NFC),A1andA2(NA1158
andNA2)wererecorded(Figure1).159
Behavioralindicators160
Theindicatorsusedinthisstudydescribethemosquitoprogressinsidetheolfactometerand161
therelativeattractiveness(RA)oftreatmentsorarms.162
Twoindicatorsoftheprogressioninsidetheolfactometerwerecalculated.First,upwind163
flight(UF)thatistheproportionoffemalethatleftthereleasezone(i.e.collectedinFC,A1andA2)164
relativetothetotalnumberofreleasedmosquitoes(N).Secondisthelocalization(L)ofodorsource165
thatistheproportionoffemalethatreachedA1andA2(NA1andNA2),relativetothenumberof166
mosquitoesthatlefttheRZ(N-NRZ).Theseindicatorswerecalculatedforeachreleaseandforeach167
odorsource(none,rabbitwithoutITNandrabbitwithITN).168
Theupwindflightandlocalizationvaluesmeasuredinexperiment1(emptyset-up,cleanair)are169
baselineindicatorsoftheanemotacticresponseofthethreemosquitogenotypestoairflow.The170
influenceofrabbitodoronmosquito’sprogressionwasdeterminedbycomparingthevaluesof171
upwindflightandlocalizationrecordedintheemptysystem(experiment1)withthoserecordedin172
thesystemwithoutITN(mergedUFandLvaluesofexperiments2and3).Themergedupwindflight173
andlocalizationvaluesrecordedinexperiments2and3(rabbitodor,noITN)werecomparedto174
thoserecordedinexperiment4(rabbitodorandITN)todetermineITNodorinfluenceonmosquito175
behavior.176
Therelativeattractiveness(RA)ofonearmversustheotherwascalculatedastheproportionof177
mosquitoesinA1orA2(NA1orNA2)relativetothesumofthemosquitoescollectedinbotharms.In178
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

9
ordertoverifythesymmetryoftheexperimentalset-up,wemeasuredRAexp2inexperiment2(rabbit179
asanodorsource,emptyboxes)asfollowandexpectedittonotbedifferentthan0.5:180
!"#$%& =()*
(()* + ()&)181
RelativeattractivenessofUTNversusemptybox(RAexp3)andITNversusUTN(RAexp4)werealso182
calculatedfromexperiments3and4,respectively,usingthefollowingequations:183
!"#$%. =(/01
(23%45 + (/01184
!"#$%6 =(701
(/01 + (701185
whereNUTNisthenumberofmosquitoescollectedinthearmwiththeboxcontainingtheUTN186
(experiment3or4),NEmptyisthenumberofmosquitoesinthearmwiththeemptybox(experiment187
3)andNITNisthenumberofmosquitoescollectedinthearmwiththeboxcontainingtheITN188
(experiment4).ThemeasureofRAexp3allowedustoassessthepossibleeffectoftheUTNasa189
physicalbarrierforthediffusionofodorcomingfromtherabbittotheolfactometer.190
Statisticalanalysis191
AllanalyseswereperformedusingtheRsoftware,version3.0.2[31],withthelme4package[32].We192
analyzedupwindflightandlocalizationusingbinomiallogisticmixed-effectmodels.Thedayof193
releasewassetasrandominterceptbecausereleasesperformedonasamedaymightnotbe194
independentandbecauseallthreegenotypeshavenotbeentestedeachday.Thekdrgenotypes(SS,195
RSorRR),thedifferentodorsources(none,RabbitwithoutITN,andRabbit+ITN)andinteractions196
betweenthemwereincludedinthemodelsasexplanatoryvariables.Upwindflight(UF)and197
localization(L)modelswerewrittenasfollow:198
logit(=>@ABCDE) = FG +FCH#IJ45%# + FD
KLJM + FCH#IJ45%#×FD
KLJOM + PE 199
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

10
,whereUForLijkistheproportionUForLrecordedforgenotypeiwithodorsourcejonday200
k,FCH#IJ45%#denotestheeffectonthelogitoftheclassificationincategoryi(SS,RSorRR)of201
Genotype;FDKLJMdenotestheeffectoftheclassificationincategoryjofOdorsource:Empty202
(experiment1),RabbitwithoutITN(experiment2and3),orRabbit+ITN(experiment4);anddk203
representstherandominterceptfordayk.Eachcombinationofcategoriesiandjoftheexplanatory204
variableswassuccessivelyusedasreferenceclassformultiplecomparisonsamonggenotypesand205
odorsources.Oddsratiosandtheir95%confidenceintervals(CI)werecomputed.206
Weverifiedthesymmetryoftheexperimentalset-upbymodellingtherelativeattractiveness207
measuredinexperiment2(RAexp2)usingabinomialmixed-effectmodelwiththereleasedayas208
randomeffect:209
logit(!"QRS&,CE) = FG +FCH#IJ45%# + PE 210
,whereRAexp2,ikistheproportionRAinA1forgenotypeiinexperiment2ondayk,FCH#IJ45%# isthe211
effectonthelogitoftheclassificationincategoryi(SS,RSorRR)ofGenotype;anddk,therandom212
interceptfordayk.213
RelativeattractivenessofUTNvs.emptyboxandITNvs.UTNwereanalyzedusingasimilarmodel214
that,inaddition,allowedforrandomeffectsoftheboxthatreceivedthetreatment:215
logit(!"CEU) = FG +FCH#IJ45%# + VU + PE 216
,whereRAiklistheproportionRAexp3orRAexp4forgenotypeiondaykwiththetreatmentplacedin217
boxl,FCH#IJ45%# indicatestheeffectonthelogitoftheclassificationincategoryi(SS,RSorRR)of218
Genotype;bl,theeffectonthelogitofthetheboxlthatreceivedthetreatment(UTNorITNfor219
RAexp3andRAexp4,respectively)anddk,therandominterceptfordayk.Eachgenotypewas220
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

11
successivelyusedasreferenceclassformultiplecomparisons.Oddsratiosandtheir95%CIwere221
computed.222
CO2concentrationswerecomparedbetweenarmsusingtheWilcoxonsigned-ranktestforpaired223
data.RHvalueswerecomparedbetweenarmsusingthepairedTtest.224
Results225
Overall,6286mosquitoeswereincludedintheassays(2621SS,1268RSand2397RR)during226
47,49,84and98releasesforexperiments1to4respectively(Table2).227
Table2:Numberofreleasesperformedpergenotypeandexperiment228
Experiment GenotypesSS RS RR Total
1-Empty 19 9 19 472-Rabbitalone 20 10 19 493-Rabbit+UTN 34 18 32 844-Rabbit+ITN 40 20 38 98Total 113 57 108 278
(UTN:Untreatednet,ITN:Insecticide-treatednet)229
DoAn.gambiaefemalesrespondtotheairflow?230
Wefirstinvestigatedtheresponsetotheairflow(anemotacticresponse)bycalculatingthe231
proportionofupwindflight(UF)femalesandthoselocated(L)inarmsintheemptyset-up232
(Experiment1).Overall,theprobabilitytoleaveRZ(UF)was0.43(95%CI[0.38–0.48];Fig2A).233
Amongtheactivatedmosquitoes,10%(95%CI[6–17])reachedA1orA2(Fig2B).Inspiteofsimilar234
upwindflightproportionamonggenotypes,theprobabilityoflocalization(L)forRSanopheleswas235
higherthanthoseofRRmosquitoes(Figure2B;ORL=2.15,95%CI[1.04,4.41]).236
Fig2:Upwindflightandlocalizationindicatorsforthethreegenotypesinrelationtotreatment237(Mean±95%ConfidenceInterval).***p<0.001,**p≤0.01,*p≤0.05,ns=notsignificant.238
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

12
DoAn.gambiaefemalesrespondtoanattractiveodorsource?239
Thepresenceofarabbitasanattractiveodorsource(experiments2and3)didnotchangethe240
proportionofupwindflightmosquitoescomparedtotheexperimentswithoutattractantodor241
(experiment1),independentlyoftheirgenotype(Table3).However,thecomparisonoftheupwind242
flightprobabilitybetweengenotypesshowthatforRSmosquitoes,UFprobabilitiesbecame243
significantlyhigherthanforSSandRRindividuals(Fig2A;ORRSvsSS=1.2495%CI[1.01,1.54];ORRSvsRR=244
1.2995%CI[1.04,1.59]).Moreover,thelocalizationprobabilitysignificantlyincreasedforall245
genotypesinthepresenceofanodorstimuluscomparedtonoodor(Table3),independentlyof246
genotypes(Fig2B).Therabbitodorhadaneffectonmosquitobehavioronlywhentheywereclose247
toarmslikelybecauseoftheodorconcentrationthatwasmoreimportantinarmsthaninthe248
releasezone.249
250
Behavioralindicator OdorsourcescomparisonsGenotypefor
thekdrmutation
OddsRatios[95%ConfidenceInterval] p-value
Upwindflight(UF)
Rabbitavsnoodorb
SS 1.09[0.87,1.37] ns
RS 1.27[0.91,1.77] ns
RR 1.08[0.85,1.37] ns
Rabbit+ITNcvsRabbita
SS 1.02[0.85,1.21] ns
RS 1.05[0.82,1.34] ns
RR 1.12[0.94,1.35] ns
Localization(L)
Rabbitvsnoodorb
SS 2.63[1.67,4.15] ***RS 1.96[1.14,3.36] *RR 4.63[2.67,8.02] ***
Rabbit+ITNcvsRabbita
SS 1.3[0.99,1.69] ns
RS 1.01[0.72,1.42] ns
RR 1.01[0.76,1.33] ns
Table3:ResultsoftheUpwindflight(UF)andlocalization(L)generalizedlinearmodels.Comparison251ofmosquitoes’progressfirstwhentherabbitwasaddedasanodorsource(vs.noodor)andthenwhenITN252waspresent(vs.rabbitalone).aexperiments2and3,bexperiment1,cexperiment4(seeTable1);***p<0.001,253**p≤0.01,*p≤0.05,ns:notsignificant.ITN:insecticide-treatednet.SS:homozygousforthesusceptibleallele,254RS:heterozygous,RR:homozygousfortheresistantallele.255
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

13
Ismosquitoresponseinfluencedbyinsecticide-treatednets?256
Totestwhethertheinsecticideonnetfibersaffectedmosquitoprogression,wecompared257
upwindflightandlocalizationprobabilitiesinthepresence(experiment4)orabsence(experiments2258
and3)oftheITN.TheprobabilitiesweresimilarinpresenceorabsenceoftheITN,regardlessofthe259
genotype(Table3;Fig2A,2B).Nevertheless,thecomparisonbetweengenotypesshowedthat260
upwindflightprobabilitiesforheterozygousRSmosquitoesremainedhigherthanthoseofthetwo261
othergenotypes,bothinthepresenceorabsenceofinsecticide(Fig2A;ORRSvsSS=1.2895%CI[1.01,262
1.62],ORRSvsRR=1.2095%CI[0.94,1.53]).263
Istheexperimentalsetupsymmetric?264
Analysisofthearms’relativeattractivenessdatafromexperiment2(Rabbitodor;twoempty265
boxes)showednosignificantdifferencesbetweenthenumberofmosquitoescollectedinA1vs.A2,266
regardlessofthegenotypes(Fig3A;RAexp2,SS=0.58,95%CI[0.34,0.79];RAexp2,RS=0.62,95%CI[0.34,267
0.83];RAexp2,RR=0.54,95%CI[0.30,0.76]).NodifferenceinRAexp2wasobservedamonggenotypes268
(ORSSvsRS=1.1695%CI[0.46,2.94];ORSSvsRR=0.8595%CI[0.38,1.94],ORRSvsRR=0.7395%CI[0.28,1.90]).269
Moreover,CO2concentrationandRHwerenotdifferentbetweenarms(p>0.05;S1).Theseresults270
demonstratedthattheolfactometerwassymmetrical.Moreover,analysesofRAexp3andRAexp4,271
(resultsdescribedbelow),showednoeffectrelativetotheboxreceivingthetreatment(i.e.variable272
nosignificantinthemodel),indicatingthatsymmetrywasmaintainedduringexperiments3and4.273
Figure3:Relativeattractivenessrates274RA:numberofmosquitoesfoundinonearmrelativetothetotalnumberofmosquitoesfoundin275botharms.(A)Experiment2(rabbitonly).(B)Experiment3(rabbit+UTNoremptybox).(C)276Experiment4(Rabbit+UTNorITN).Asterisksshowdifferenceto0.5,traducingachoiceforone277treatmentratherthantheother.Errorbarsshowthe95%CI;**p≤0.01,*p≤0.05.UTN:Untreated278net,ITN:Insecticide-treatednet.SS:homozygotefortheL1014Sallele(insecticide-susceptible),RS:279heterozygousfortheL1014Fallele,RR:homozygousforL1014Fallele(insecticide-resistant).280
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

14
IstheattractivenessoftheodorsourceinfluencedbytheUTN?281
Inexperiment3(oneemptyboxandoneboxwith2m²ofUTN,bothinpresenceofrabbitodor),the282
emptyboxwasmoreattractiveforSSandRRmosquitoesbutnotforRS(RAexp3,SS=0,3195%CI283
[0.20,0.46],p-value=0.013;RAexp3,RR=0.27,95%CI[0.17,0.42],p-value=0.002)(Fig3B).Nosignificant284
differenceofmosquito’sproportioninarmswasevidencedbetweengenotypes(ORSSvsRS=1.24,285
95%CI[0.61,2.50];ORSSvsRR=0.83,95%CI[0.41,1.66],ORRSvsRR=0.67,95%CI[0.31,1.47]).CO2286
concentrationwasnotdifferentbetweenarms,whileasignificant1%differenceinRHwasobserved287
(63.9%intheUTNarmand64.9%intheemptyarm,pairedTtestp-value=0.007).288
IstheattractivenessoftheodorsourceinfluencedbytheITN?289
AnalysisofRAexp4fromexperiment4(Rabbitodor;oneboxwith2m²ofUTNandonewith2m²of290
ITN)showedthatRRmosquitoespreferablychosetheITNarmwithprobability0.63(95%CI[0.53-291
0.73],p-value=0.01;Fig3C).ThisprobabilitywassignificantlyhigherthanthoseobservedbothforRS292
(RAexp4,RS=0.4795%CI[0.34-0.60];ORRRvsRS=1.95,95%CI[1.06,3.57],p-value=0.03)andSSgenotypes293
(RAexp4,SS=0.595%CI[0.40-0.61];ORRRvsSS=1.71,95%CI[1.03,2.83],p-value=0.04).CO2concentration294
andRHwerenotdifferentbetweenarms.295
Discussion296
Thehost-seekingbehaviorofmosquitoestowardshumanssleepingunderabednetispoorly297
understood.Particularly,itisnotknownwhetherspecificvolatilechemicalsemanatingfromtreated298
netsmightmodulatethisbehavior.Here,weusedadual-choiceolfactometertoassesswhetherthe299
presenceofpermethrin-treatednetsmayinfluencethehostattractivenessformosquitoesof300
differentkdrgenotypes.Wefoundthatnetsrepresentbothaphysicalandachemicalsignalthat301
modulatemosquitoactivationand/orchoice.Moreover,thethreekdrgenotypesbehaveddifferently302
inresponsetohostodorsinthepresenceofITNsorUTNs.303
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

15
Physicalbarrier&environmentalcues 304
Inexperiment3,mosquitoespreferablychosethearmconnectedtotheemptyboxrather305
thantheboxwithUTNs.NodifferenceinCO2quantitywasnotedbetweenarms.However,the306
humiditylevelwasslightlyhigherinthearmconnectedwiththeemptybox.Ashumidityisknownto307
attractmosquitoes[33],theobservedpreferencefortheemptybox(higherhumidity)wasnot308
surprising.ThisdifferencecouldhavebeencausedbythephysicalbarrierformedbyUTNsthatmay309
absorbhumiditycomingfromtherabbitbox.Inaddition,thenetstructurecouldalsohaveretained310
volatilechemicalsemanatingfromrabbitwhichareimportantinmosquitoorientationandchoice311
[34,35].312
Chemicalecology&ITNs313
Ourresultsraisedthequestionofthevolatilechemicalsemanatingfromnetsthatmaydrive314
aspecificbehaviorinresistantmosquitoes.Permethrinisnotknownasaclassicalvolatilecompound315
becauseofitslowvaporpressure(5.18x10-8mmHgat25°c).Nevertheless,Bouvieretal[41]recently316
detectedpermethrininindoorairsamples(11and18.8ng/m3fortrans-permethrinandcis-317
permethrinrespectively)indicatedthatsuchpyrethroidmightbepresentintheaireventheyare318
semi-volatileorganiccompounds.Moreaccurately,astudybyBomannetal.[42]fromtheBayer319
companymeasuredameanconcentrationofcyfluthrin(apyrethroidwithamolecularstructureclose320
tothepermethrin)of0.000021mg/m3at1mawayfromatreatednet.Suchconcentration321
correspondsto3.46x109molecules/cm3.Angioyetal[36]foundthatonly6moleculesofa322
pheromoneenteredincontactwiththeolfactorysensillumofmothspeciesmayinducea323
physiologicalresponse.Wethereforehypothesizethatmosquitomaydetectverylowconcentration324
ofpyrethroidintheair.325
Moreover,somenasaltrouble(i.erunnynose)havebeenrecordedwhenLNswereusedfor326
thefirsttime[37].SuchobservationsreinforcethehypothesisthatLNsemitchemicalsintheair.327
Regardlessthesechemicalsareinsecticideitself,additivechemicals(i.e.fragrances),degradation328
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

16
products,thatcomposedthenet,theymaybedetectedbymosquitoesandelicitbehavioral329
modulation.330
Thebehaviorofinsects,suchasmosquitoes,isdrivenbytheperceptionandintegrationof331
odorantsignalsinantennaeandhigherbraincenter.Inourstudy,weobservedthatkdrresistant332
mosquitoesweremoreattractedbyhostodorsemanatingbehindapermethrin-treatednetthan333
hostodorsbehindanuntreatednet(Fig3C),itindicatesthattheyperceivedatdistanceadifference334
betweenITNandUTNandbehaveddifferentlyinresponse.Wethenhypothesizethatmosquitoes335
areabletodetectchemicalsreleasedbynetwitholfactoryreceptors(Ors)tunedtorespondtothese336
chemicals.Asanexample,authorsrecentlyidentifiedoneolfactoryreceptoractivatedandanother337
inhibitedbysyntheticpyrethroïdinAedesaegypti[38],suggestingthatsuchORmayalsoexistin338
Anophelesmosquitoes.Themajorresearchperspectiveraisedbyourresultsisthereforetostudythe339
chemicalandbehavioralecologyrelativetovectorcontroltoolsalreadywidespreadinendemic340
country.341
Insecticideresistance&hostseekingbehavior342
Ourresultsalsoclearlyindicatedthatthekdrmutation,orcloselylinkedpolymorphisms[39],343
modulatedthehostchoiceofAn.gambiaemosquitoesinthepresenceofaITN.Thestrainsusedin344
ourstudysharethesamegeneticbackground.ExceptifgenescodingforORssensitivetoLN-related345
odorantsarelocatedinflankingregionofKdrmutation,,onlyapleiotropiceffectofthekdrmutation346
affectingthetransmissionorintegrationoftheneuronalsignalinhomozygousmosquitoescould347
explainthedifferentbehaviorsbetweengenotypes.Thekdrmutationmayinfluencethetransmission348
ofanodorantsignaltowardshigherbrainregionsbyenhancingtheclosed-stateinactivationofthe349
voltage-gatedsodiumchannel,whichplaysacentralroleinmessagepropagationinthenervous350
system.Asaconsequence,areductionofneuronalexcitabilitycouldbeobservedinkdrmutantsin351
comparisontosusceptibleindividuals[40].Allchemicalsignalsaretransducedbyspikefrequencies352
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

17
intheolfactorysensoryneurons[41]andtheinformationsentbystimulatedorinhibitedneuronsis353
treatedintheantennallobe[42].Therefore,iftheneuronalexcitabilitydiffersinhomozygouskdr354
genotypes,theresponsepatternoftheolfactoryneuronsandsubsequentlythesignalintegration355
andprocessinginthecentralnervoussystemcouldbealtered,leadingtoamodifiedmotorresponse,356
inthiscase,adifferenceinhostchoice.357
Thepresentstudysuggeststheexistenceofinteractionsbetweenthephysiological358
mechanismsthatallowmosquitoestosurviveacontactwithinsecticideandthebehavioralresponse359
toolfactorycues.Theseinteractionsmayhavemajorimplicationsinmalariacontrol.Asanexample,360
chemicalsemanatingfromtheITNsarestronglyrelatedtothepresenceofhumanbeings.Shouldit361
beintegratedasanattractivecueforresistantmosquitoes?Thismaydramaticallyaffectsthe362
personalandcommunityprotectiongivenbythemassiveuseofITNs.Ourstudyonlyfocusedonthe363
Kdrmutation,buttheresistancepatterninwildAnophelespopulationsisfarmorecomplex[43].It364
wouldbeinterestingtoinvestigatetheinteractionbetweeneachresistancemechanismsisolatedin365
specificstrainsbeforegoingtostudythisinteractionbetweenresistance,behaviorandITNsinsemi-366
fieldandnaturalconditions.Recentpapersweremodelingandquestioningtheriskconferredby367
resistance,basedonsurvivaltoinsecticideexposure[44],buttheimpactofsuchresistanceon368
behaviorisalsotobeinvestigatedurgently[45].369
Weusedrabbitasanodorsourcebecausemosquitoeswerefedonrabbitsatthelaboratory,370
andwerelikely“selected”torespondtorabbit’sodor.Butinthefield,Anophelesgambiaeprefersto371
bitehumanwhenavailable[46].Whetherthesameexperimentconductedwithhumansasanodor372
sourcewillprovidesimilarresultsremaintobeexperimentallyevaluated.Ifweusedahumaninstead373
oftherabbitwechangethecompositionofodorplume(quantityandqualityofsemiochemicals).374
ThereforetheinteractionbetweenchemicalsreleasedbyLLINandhumanodorshouldinducea375
differentbehavior.Nevertheless,ourexperimenthighlightedtheinvolvementofLLINinhostseeking376
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

18
behaviorandemphasizedtheneedtostudyingtherelationbetweenLLIN,hostodorsandmosquito377
hostseekingbehavior.378
Kdrgenotypes&behavior379
HeterozygousRSmosquitoesweremoreactivethanSSandRRmosquitoes.Theadditionof380
anattractantdidnotchangetheproportionofRSleavingtheRZ,suggestingthatthisbehaviormight381
berelatedtoabetteranemotacticresponse(i.eresponsetoairflow)orspontaneousflightactivity382
thanabetterperceptionofodorantsinRSmosquitoes.Thishypothesisisstrengthenedbythe383
absenceofdifferenceintheprogressiontowardstheolfactometerarmsamonggenotypes.Inother384
words,heterozygousmosquitoesflymore,butmightnotsmellbetter.Ontheonehand,byflying385
moretheymightincreasetheprobabilityofencounteringahostodorantplumewhichmightbe386
advantageous.SuchheterozygousadvantageforthekdrlocusinAn.gambiaes.s.hasbeenrecently387
documentedalsoforanotherbehavioraltrait:theabilitytofindaholeinapieceofbednet[24]and388
formalemating[47].Inotherhand,itcouldrepresentacostformosquitoesifenergyspentduring389
flightisnomoreavailableforothertraitscloselyrelatedtofitnessasfertility,fecundityand390
longevity.Thistradeoffmustbedeeplyinvestigatedasthismighthavegreatinfluenceon391
Plasmodiumtransmission.392
Thebehaviorofkdrheterozygousindividualsinourstudymustbeinterpretedwithcaution393
becauseotherloci,distinctfromthekdrlocus,couldalsoinfluencethisbehavioraltrait.Introgression394
andselectionthekdralleletoproducethehomozygousresistantstrainwasindeedlikelytoalsohave395
selectedlinkedpolymorphisms[45].396
Conclusion397
Inconclusion,ourstudyshowedthattheAnophelesmosquitoesdetectedthepresenceof398
bothphysicalandchemicalbarriersofITNS.Facetothisresults,iturgestodecipherwiththe399
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

19
interactionbetweenhost-seekingbehavior,insecticideresistanceandvectorcontroltools.Themost400
overlookedpartofthepuzzleisthechemicalecologyinacontextoflargevectorcontrolmeasure401
deployment.Thisresearchavenuewillbechallengingforthevectorcontrolcommunitybutiscrucial402
nottowasteforcesinwrongdirections.403
Acknowledgement404
We thank TeunDekker for helpful discussion.Wewould like to thank the IEMTV staff in Abomey405
Calavi(Benin)fortechnicalassistance.406
407
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

20
References408
[1] CohuetA,HarrisC,RobertV,FontenilleD.EvolutionaryforcesonAnopheles:whatmakesa409
malariavector?TrendsParasitol2010;26:130–6.doi:10.1016/j.pt.2009.12.001.410
[2] Garrett-Jones C, Shidrawi GR. Malaria vectorial capacity of a population of Anopheles411
gambiae:anexerciseinepidemiologicalentomology.BullWorldHealthOrgan1969;40.412
[3] Garrett-JonesC,BorehamPFL,PantCP.Feedinghabitsofanophelines(Diptera:Culicidae)in413
1971–78,withreferencetothehumanbloodindex:areview.BullEntomolRes1980;70:165–85.414
[4] ColuzziM,SabatiniA,PetrarcaV,DecoMAD.Chromosomaldifferentiationandadaptationto415
humanenvironments intheAnophelesgambiaecomplex.TransRSocTropMedHyg1979;73:483–416
97.doi:10.1016/0035-9203(79)90036-1.417
[5] ZaimM,AitioA,NakashimaN.Safetyofpyrethroid-treatedmosquitonets.MedVetEntomol418
2000;14:1–5.419
[6] Ranson H, N’Guessan R, Lines J, Moiroux N, Nkuni Z, Corbel V. Pyrethroid resistance in420
African anopheline mosquitoes: what are the implications for malaria control? Trends Parasitol421
2011;27:91–8.422
[7] Ranson H, Jensen B, Vulule J, Wang X, Hemingway J, Collins F. Identification of a point423
mutation in thevoltage-gatedsodiumchannelgeneofKenyanAnophelesgambiaeassociatedwith424
resistancetoDDTandpyrethroids.InsectMolBiol2001;9:491–7.425
[8] GattonML,ChitnisN,ChurcherT,DonnellyMJ,GhaniAC,GodfrayHCJ,etal.TheImportance426
of Mosquito Behavioural Adaptations to Malaria Control in Africa. Evolution 2013;67:1218–30.427
doi:10.1111/evo.12063.428
[9] GeorghiouGP.Theevolutionofresistancetopesticides.AnnuRevEcolSyst1972;3:133–68.429
[10] CorbelV,AkogbetoM,DamienGB,DjenontinA,ChandreF,RogierC,etal.Combinationof430
malaria vector control interventions in pyrethroid resistance area in Benin: a cluster randomised431
controlledtrial.LancetInfectDis2012.432
[11] Moiroux N, GomezMB, Pennetier C, Elanga E, Djènontin A, Chandre F, et al. Changes in433
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

21
AnophelesfunestusBitingBehaviorFollowingUniversalCoverageofLong-LastingInsecticidalNetsin434
Benin.JInfectDis2012.435
[12] NdiathMO,Mazenot C, Sokhna C, Trape J-F. How theMalaria Vector Anopheles gambiae436
AdaptstotheUseofInsecticide-TreatedNetsbyAfricanPopulations.PloSOne2014;9.437
[13] Lockwood JA, Sparks TC, Story RN. Evolution of Insect Resistance to Insecticides: A438
ReevaluationoftheRolesofPhysiologyandBehavior.BullESA1984;30:41–51.439
[14] ChareonviriyaphapT,BangsMJ,SuwonkerdW,KongmeeM,CorbelV,Ngoen-KlanR.Review440
ofinsecticideresistanceandbehavioralavoidanceofvectorsofhumandiseasesinThailand.Parasit441
Vectors2013;6:280.doi:10.1186/1756-3305-6-280.442
[15] Miller JR, Siegert PY, Amimo FA, Walker ED. Designation of Chemicals in Terms of the443
LocomotorResponsesTheyElicit From Insects:AnUpdateofDethieretal. (1960). JEconEntomol444
2009;102:2056–60.doi:10.1603/029.102.0606.445
[16] Chareonviriyaphap T, Roberts D, Andre RG, Harlan HJ, Manguin S, Bangs MJ. Pesticide446
avoidancebehavior inAnopheles albimanus, amalaria vector in theAmericas. JAmMosqControl447
Assoc-MosqNews1997;13:171–83.448
[17] Malima R, Oxborough R, Tungu P,Maxwell C, Lyimo I,Mwingira V, et al. Behavioural and449
insecticidal effects of organophosphate, carbamate and pyrethroid treatedmosquito nets against450
Africanmalariavectors.MedVetEntomol2009;23:317–25.451
[18] Achee NL, Sardelis MR, Dusfour I, Chauhan KR, Grieco JP. Characterization of Spatial452
Repellent,ContactIrritant,andToxicantChemicalActionsofStandardVectorControlCompounds1.453
JAmMosqControlAssoc2009;25:156–67.454
[19] DeletreE,MartinT,CampagneP,BourguetD,CadinA,MenutC,etal.Repellent,Irritantand455
ToxicEffectsof20PlantExtractsonAdultsoftheMalariaVectorAnophelesgambiaeMosquito.PLoS456
ONE2013;8:e82103.doi:10.1371/journal.pone.0082103.457
[20] Siegert PY, Walker E, Miller JR. Differential behavioral responses of Anopheles gambiae458
(Diptera: Culicidae) modulate mortality caused by pyrethroid-treated bednets. J Econ Entomol459
2009;102:2061–71.460
[21] CorbelV,ChabiJ,DabireRK,EtangJ,NwaneP,PigeonO,etal.Fieldefficacyofanewmosaic461
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

22
long-lastingmosquitonet(PermaNet3.0)againstpyrethroid-resistantmalariavectors:amulticentre462
studyinWesternandCentralAfrica.MalarJ2010;9.doi:10.1186/1475-2875-9-113.463
[22] Lindsay SW, Adiamah JH, Miller JE, Armstrong JRM. Pyrethroid-treated bednet effects on464
mosquitoes of the Anopheles gambiae complex in TheGambia.MedVet Entomol 1991;5:477–83.465
doi:10.1111/j.1365-2915.1991.tb00576.x.466
[23] PleassRj.,ArmstrongJr.m.,CurtisCf.,JawaraM,LindsaySw.Comparisonofpermethrin467
treatments for bednets in The Gambia. Bull Entomol Res 1993;83:133–9.468
doi:10.1017/S0007485300041870.469
[24] DiopMM,MoirouxN, Chandre F,Martin-HerrouH,Milesi P, Boussari O, et al. Behavioral470
Cost & Overdominance in Anopheles gambiae. PLoS ONE 2015;10:e0121755.471
doi:10.1371/journal.pone.0121755.472
[25] Parker JEA, Angarita-Jaimes N, Abe M, Towers CE, Towers D, McCall PJ. Infrared video473
tracking of Anopheles gambiae at insecticide-treated bed nets reveals rapid decisive impact after474
brieflocalisednetcontact.SciRep2015;5:13392.doi:10.1038/srep13392.475
[26] Grieco JP,AcheeNL,ChareonviriyaphapT, SuwonkerdW,ChauhanK, SardelisMR,etal.A476
NewClassification System for theActionsof IRSChemicals TraditionallyUsed ForMalaria Control.477
PLOSONE2007;2:e716.doi:10.1371/journal.pone.0000716.478
[27] Martinez-Torres D, Chandre F, Williamson M, Darriet F, Berge JB, Devonshire AL, et al.479
Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector480
Anophelesgambiaess.InsectMolBiol1998;7:179–84.481
[28] Djegbé I. Modification physiologique et comportementale induites par la résistance aux482
insecticideschezlesvecteursdupaludismeauBénin.Universitéd’AbomeyCalavi,2013.483
[29] N’GuessanR,CorbelV,AkogbetoM,RowlandM.Reducedefficacyofinsecticide-treatednets484
andindoorresidualsprayingformalariacontrolinpyrethroidresistancearea,Benin.EmergentInfect485
Dis2007;13:199–206.486
[30] GeierM,BoeckhJ.AnewY-tubeolfactometerformosquitoestomeasuretheattractiveness487
ofhostodours.EntomolExpAppl1999;92:9–19.488
[31] R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R489
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

23
FoundationforStatisticalComputing;2014.490
[32] BatesD,MaechlerM,BolkerB,WalkerS.lme4:Linearmixed-effectsmodelsusingEigenand491
S4.Rpackageversion1.1-72014.492
[33] TakkenW,KnolsBGJ,OttenH.Interactionsbetweenphysicalandolfactorycuesinthehost493
seeking behaviour of mosquitoes: the role of relative humidity. Ann Trop Med Parasitol494
1997;91:119–20.doi:10.1080/00034989761427.495
[34] DekkerT,TakkenW,CardeRT.Structureofhost-odourplumesinfluencescatchofAnopheles496
gambiaessandAedesaegyptiinadual-choiceolfactometer.PhysiolEntomol2001;26:124–34.497
[35] CardéRT,GibsonG.Hostfindingbyfemalemosquitoes:mechanismsoforientationtohost498
odours and other cues. Olfaction Vector-Host Interact., Willem Takken and Bart G.J. Knols.499
WageningenAcademicPublishers;2010.500
[36] Angioy AM, Desogus A, Barbarossa IT, Anderson P, Hansson BS. Extreme sensitivity in an501
olfactorysystem.ChemSenses2003;28:279–84.502
[37] WHO.Guidelinesforlaboratoryandfield-testingoflong-lastinginsecticidalnets.2013.503
[38] Bohbot JD, Fu L, LE TC, Chauhan KR, Cantrell CL, Dickens JC. Multiple activities of insect504
repellents on odorant receptors in mosquitoes. Med Vet Entomol 2011;25:436–44.505
doi:10.1111/j.1365-2915.2011.00949.x.506
[39] Mitri C, Markianos K, Guelbeogo WM, Bischoff E, Gneme A, Eiglmeier K, et al. The kdr-507
bearing haplotype and susceptibility to Plasmodium falciparum in Anopheles gambiae: genetic508
correlationandfunctionaltesting.MalarJ2015;14.doi:10.1186/s12936-015-0924-8.509
[40] Vais H, Williamson MS, Goodson SJ, Devonshire AL, Warmke JW, Usherwood PN, et al.510
Activation of Drosophila sodium channels promotes modification by deltamethrin. Reductions in511
affinitycausedbyknock-downresistancemutations.JGenPhysiol2000;115:305–18.512
[41] Guidobaldi F,May-Concha IJ,GuerensteinPG.Morphology andphysiologyof theolfactory513
systemofblood-feedinginsects.JPhysiolParis2014.doi:10.1016/j.jphysparis.2014.04.006.514
[42] HansonB,ChristensenTA. Functional characteristicsof theantennal lobe. InsectOlfaction.515
Springer,Berlin:B.S.Hansson;1999,p.126–57.516
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

24
[43] Corbel V, N’guessan R, Brengues C, Chandre F, Djogbenou L, Martin T, et al. Multiple517
insecticide resistance mechanisms in Anopheles gambiae and Culex quinquefasciatus from Benin,518
WestAfrica.ActaTrop2007;101:207–16.519
[44] VianaM,HughesA,MatthiopoulosJ,RansonH,FergusonHM.Delayedmortalityeffectscut520
the malaria transmission potential of insecticide-resistant mosquitoes. Proc Natl Acad Sci521
2016;113:8975–80.doi:10.1073/pnas.1603431113.522
[45] ThomasMB,ReadAF.Thethreat (ornot)of insecticideresistanceformalariacontrol.Proc523
NatlAcadSci2016;113:8900–2.doi:10.1073/pnas.1609889113.524
[46] LefèvreT,GouagnaL-C,DabiréKR,ElgueroE,FontenilleD,RenaudF,etal.Beyondnature525
andnurture:phenotypicplasticityinblood-feedingbehaviorofAnophelesgambiaesswhenhumans526
arenotreadilyaccessible.AmJTropMedHyg2009;81.527
[47] Platt N, Kwiatkowska RM, Irving H, Diabaté A, Dabire R,Wondji CS. Target-site resistance528
mutations (kdr and RDL), but not metabolic resistance, negatively impact male mating529
competiveness in the malaria vector Anopheles gambiae. Heredity 2015;115:243–52.530
doi:10.1038/hdy.2015.33.531
[48] BerticatC,BoquienG,RaymondM,ChevillonC.Insecticideresistancegenesinduceamating532
competitioncostinCulexpipiensmosquitoes.GenetRes2002;79:41–7.533
534
Supportinginformation535
S1:Effectoftreatmentonenvironmentvariables536537538539
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;
61#cm#
Rota*ng#doors#
8#cm#
32#cm#45#cm#
30.5#cm#
33.5#cm#
34#cm#
30#cm#
23.5#cm#
Fan#Box#with#treatment#
Release#zone#(RZ)#
Flight#chamber#(FC)#Arm#1#and#2#
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

No-odor (Experiment 1)
Rabbit (Experiment 2+3)
Rabbit+ITN (Experiment 4)
0.0
0.2
0.4
0.6
0.8
1.0P
ropo
rtion
of f
emal
es
SSRS
RR
* * *ns
ns nsns ns
ns
Upwind flight
No-odor (Experiment 1)
Rabbit (Experiment 2+3)
Rabbit+ITN (Experiment 4)
0.0
0.2
0.4
0.6
0.8
1.0
Pro
porti
on o
f fem
ales
nsns *
ns nsnsns ns ns
Localisation
A
B
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;

0.00.20.40.60.81.0
0.00.20.40.60.81.0
0.00.20.40.60.81.0
0.0 0.2 0.4 0.6 0.8 1.0
0.0 0.2 0.4 0.6 0.8 1.0
0.0 0.2 0.4 0.6 0.8 1.0
Proportion of female found in arm
A
B
C
Empty box 1 Empty box 2
UTN Empty box
ITN UTN
*
**
**
RS
SS
RR
.CC-BY-NC-ND 4.0 International licensenot peer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was. http://dx.doi.org/10.1101/077552doi: bioRxiv preprint first posted online Sep. 26, 2016;