a novel regulatory mechanism of pim-3 kinase stability and...
TRANSCRIPT

A Novel Regulatory Mechanism of Pim-3 Kinase Stability and its Involvement in
Pancreatic Cancer Progression
Fei Zhang1, Bin Liu1, Zhen Wang1, Xian-Jun Yu2, Quan-Xing Ni2 Wen-Tao Yang3,
Naofumi Mukaida4, Ying-Yi Li1,4*
1Cancer Research Institute, 2Department of Pancreas and Hepatobiliary, Pancreatic
Cancer Institute, 3Department of Pathology, Fudan University Shanghai Cancer Center,
Department of Oncology, Shanghai Medical College, Fudan University, Shanghai, China
and 4Division of Molecular Bioregulation, Cancer Microenvironment Research Program,
Cancer Research Institute, Kanazawa University, Kanazawa, Japan
Running Title: Stabilization of Pim-3 by TCTP
Key words: Serine/threonine kinase; TCTP; Stabilization; Apoptosis; Cell cycle.
Grant support: This work was supported in part by the National Science Foundation of
China (NSFC) (30973476, 812727), the Shanghai Pujiang Program (KW201028464),
Fudan University ‘985 Project’ Phase III Cancer Research Projects II (985 -YFX0102),
and Shanghai Committee of Science and Technology (12DZ2260100).
*Correspondence should be addressed to: Ying-Yi Li, Room 1216, 2nd Building,
Cancer Research Institute, Fudan University Shanghai Cancer Center, 270 DongAn Road,
Shanghai 200032, China
Phone: +86-21-64175590-5220; Fax: +86-21-64172585; Email: [email protected]
Competing interests: The authors declare no competing interests.
Word count of the manuscript including abstract, introduction, materials and methods,
results and discussion sections is 4,601. Total number of figures is 6.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 2
Abstract Translationally-controlled tumor protein (TCTP/TPT1) was identified from a yeast
two-hybrid screen and shown to interact with Pim-3, a member of the proto-oncogene
Pim family with serine/threonine kinase activity. TCTP was aberrantly expressed in
human pancreatic cancer cells and malignant ductal epithelial cells, but not in normal
pancreatic duct epithelial cells adjacent to tumor foci of human pancreatic cancer tissue.
Moreover, TCTP co-localized with Pim-3 both in human pancreatic cancer cells and
clinical tissues. Mapping studies revealed that the interaction between Pim-3 and TCTP
occurred through the C-terminal region of Pim-3 and N-terminal region of TCTP.
Although Pim-3 had no effect on TCTP expression or phosphorylation, overexpression of
TCTP increased the amount of Pim-3 in a dose-dependent manner. Interestingly,
RNAi-mediated ablation of TCTP expression reduced Pim-3 protein but not mRNA,
through a mechanism involving the ubiquitin-proteasome degradation system. As a
consequence of Pim-3 instability and subsequent degradation, tumor growth in vitro and
in vivo was inhibited by arresting cell cycle progression and enhancing apoptosis.
Furthermore, TCTP and Pim-3 expression were significantly correlated in pancreatic
adenocarcinoma specimens, and patients with highly expressed TCTP and Pim-3
presented with a more advanced tumor stage. These observations indicate that TCTP
enhances Pim-3 stability to simultaneously promote and prevent cell cycle progression
and apoptosis, respectively. Hence, TCTP and Pim-3 serve a pivotal role in human
pancreatic cancer with important ramifications for clinical diagnostic and therapeutic
implications.
Implications: The present study provides a new idea and experimental evidence
for recognizing TCTP/Pim-3 pathway as a target for therapy in human pancreatic cancer.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 3
Introduction
Pim-3, a member of the proto-oncogene Pim family with serine/threonine kinase
activity, was originally identified as a depolarization-induced gene, KID-1, in the rat
pheochromocytoma cell line PC12 (1). Later, KID-1 was renamed Pim-3 because of its
high sequence similarity with Pim family proteins, which belong to the group of
calcium/calmodulin-regulated kinase (1). Subsequently, Deneen and colleagues
demonstrated that Pim-3 gene transcription was enhanced in EWS/ETS-induced
malignant transformation of NIH3T3 cells (2), suggesting the involvement of Pim-3 in
tumorigenesis. In line with these observations, we demonstrated that Pim-3 expression
was enhanced in malignant lesions, but not normal tissues of endoderm-derived organs
such as the liver (3), pancreas (4), colon (5), and stomach (6). Hepatocellular carcinoma
development was accelerated in mice expressing the Pim-3 transgene selectively in liver,
when these mice were treated with a hepatocarcinogen (7). We observed that Pim-3 can
inactivate Bad in human pancreas and colon carcinoma cell lines by phosphorylating
Ser112, but not Ser136, and ultimately promoting their survival (4, 5) as observed for
Pim-1 and Pim-2 (8, 9). Moreover, it has been reported that Pim-3 can promote cell cycle
progression by modulating the functions of molecules that regulate cell cycle progression
and augment protein synthesis through the regulation of PGC-1α, eventually contributing
to carcinogenesis (10). Baines and colleagues recently reported that Pim-3 suppression
can sensitize pancreatic cancer cells to gemcitabine (11). We also demonstrated that
Pim-3 can promote tumor growth and angiogenesis by stimulating the VEGF
pathway(12). Furthermore, Pim-3 modulates Myc activity to promote tumorigenesis (13).
Thus, Pim-3 is a key player in tumorigenesis, and therefore, an ideal target for cancer
therapy.
The translationally controlled tumor protein (TCTP) is a highly conserved
hydrophilic protein (14) that has been identi�ed in a wide range of eukaryotic organisms,
including fungi, yeast, insects, plants, and mammals (15). TCTP is also known as
IgE-dependent histamine releasing factor (HRF), fortilin, P21, P23, and TPT-1 (16-18).
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 4
This protein was named TCTP because its mRNA was found to be controlled at the
translational level (19). Although TCTP is found ubiquitously in tissues and cell types,
its expression levels vary depending on the tissue type, growth, stress factors, and
cytotoxic signals (20-22). A series of recent reports proved that TCTP plays important
roles in a number of cell physiological events in cancer, cell proliferation, apoptosis
regulation, stress response, gene regulation, heat shock response, and allergic response
(23). TCTP can also interact with many cellular proteins, including translation elongation
factors eEF1A and eEF-B-�, tubulin, actin, myeloid cell leukemia protein-1 (MCL1),
Bcl-XL, p53, and Na- and K-ATPase (24, 25). However, the roles of TCTP in
tumorigenesis remain largely unknown.
Considering the critical role of Pim-3 in tumor development and progression,
defining regulatory mechanisms of Pim-3 signaling networks is important. In this study,
in order to identify potential novel regulators of Pim-3, we performed yeast two-hybrid
screening using human HeLa matchmaker cDNA library. We observed that TCTP
specifically interacts with Pim-3 and enhances Pim-3 protein stability by blocking the
ubiquitin–proteasome-mediated degradation of Pim-3 protein, thereby promoting tumor
growth in vitro and in vivo.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 5
Materials and Methods
Cell culture and antibodies
Human pancreatic carcinoma cell lines, PCI35 and PCI55, were gifts from Prof.
Mukaida Naofumi of Kanazawa University (Kanazawa, Japan). Human pancreatic
carcinoma cell lines SW1990, MiaPaca-2, PANC-1, and BxPC-3 and the human
embryonic kidney (HEK) cell line 293T were purchased from the American Type Culture
Collection (ATCC). Among them, SW1990, MiaPaca-2 and PANC-1 cells were cultured
in RPMI 1640 supplemented with 10% FBS (Biowest, Inc., Loire Valley, France).
BxPC-3 and 293T cells were cultured in DMEM supplemented with 10% FBS at 37°C in
a humidi�ed 5% CO2 incubator. The authenticity of all the cell lines was confirmed by
determining DNA profiling of Short Tandem Repeat (STR) while mycoplasma
contamination was excluded with the help of Amelogenin (Beijing Microread Genetics
Co.,Ltd) The antibodies used in this study are described in Supplementary Materials &
Methods.
Yeast two-hybrid screening
Human Pim-3 cDNA fragment (121-326aa) was cloned into pGBKT7 vector and used
to screen a pACT2-human HeLa matchmaker cDNA library in a yeast two-hybrid system
(Clontech, Mountain View, CA), because we did not detect Pim-3 protein in Hela cells
(4). �-galactosidase activities were measured using o-nitrophenyl-galactoside as a
substrate. Clones activating the �-galactosidase reporter gene were sequenced and
analyzed.
Generation of expression vectors
An expression plasmid for TCTP tagged with the His epitope at the COOH- terminus
was constructed to detect TCTP protein by the anti-His antibody. A TCTP cDNA
fragment, which has an XhoI restriction site in place of the stop codon, was generated by
PCR using TCTP cDNA cloned in the pACT2 plasmid as the template and two primers
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 6
(Supplementary Table S1). The ampli�ed fragments were digested with HindIII and XhoI
(Takara, Dalian, China) and inserted into HindIII and XhoI sites of the pcDNA4 vector
(Invitrogen). The Pim-3 expression vector (pcDNA4-Pim-3) and the Pim-3-shRNA
vector (pSilencer-Pim-3-shRNA) were constructed as previously described (4).
Three deletion mutants of TCTP and two deletion mutants of Pim-3 were isolated by
PCR using a combination of primers (Supplementary Table S1) for His-tagged constructs.
PCR products spanning each fragment were cloned into the EcoRI and XhoI sites of the
pcDNA4 vector, and named �TCTP-1 (Met1-Gly69), �TCTP-2 (Val70-Ala119), �TCTP-3
(Glu120-Cys172), �Pim-3A(met1-pro126), and �Pim-3B (leu121-leu326), respectively. All
mutated nucleotides were con�rmed by sequencing.
Co-immunoprecipitation and immunoblotting
HEK293T cells were co-transfected with the Pim-3 expression and
pcDNA4-TCTP-His vectors using Lipofectamine 2000 (Invitrogen) according to the
manufacturer’s protocol. Forty-eight hours after transfection, the cells were collected and
solubilized with 1 ml of NP-40 lysis buffer [50 mM Tris-HCl, 150 mM NaCl, 1% NP-40,
1 mM EDTA, pH 7.4, with complete protein inhibitor cocktail (Roche)]. Pre-cleared cell
lysates were incubated with 2 μg rabbit anti-Pim-3 antibody or mouse anti-His antibody
overnight at 4°C and precipitated with 20 μl protein G-Sepharose 4 Fast flow (GE
Healthcare Bio-sciences AB) for 2 to 4 h at 4°C. The beads were washed three times with
cell lysis buffer. Materials bound to the beads were eluted with SDS-PAGE loading
buffer containing 1% �-mercaptoethanol, boiled for 5 min, and separated on 12%
SDS-polyacrylamide gels and transferred to polyvinylidene di�uoride membranes
(Millipore, Billerica, MA). Immunoblotting was performed as previously described (4),
Immunofluorescence analysis
PCI55 cells were cultured for 48 h, washed and fixed with 4% paraformaldehyde/PBS,
and hybridized with a combination of goat polyclonal anti-Pim-3 antibodies (1:100,
Santa Cruz) and rabbit monoclonal anti-TCTP antibodies (1:100, Epitomics). Next, the
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 7
cells were sequentially incubated with Alexa Fluor 594 donkey anti-goat IgG and Alexa
Fluor 488-conjugated goat anti-rabbit IgG antibodies. The signals were visualized using
immuno�uorescence confocal microscopy (Leica, Switzerland). Immunofluorescence
analysis was performed similarly on human pancreatic cancer tissues.
Real-time quantitative (q)RT-PCR
Total RNA was extracted using the Trizol LS reagent (Invitrogen). mRNA was
reverse-transcribed using the SuperScript First-Strand Synthesis System (Invitrogen).
Real-time PCR was performed using the Applied Biosystems HT7900 PCR system with
2× QuantiFast SYBR Green PCR Master Mix (Qiagen), 0.2 μM primers (Supplementary
Table S2), and <100 ng cDNA in a 25-μl reaction mixture. Relative expression of target
genes was analyzed by the ��Ct method. Results are expressed as means ± SD.
Knockdown of TCTP expression
The selected short interfering RNA target sequence in TCTP
(5�-AAGGTACCGAAAGCACAGT-3� corresponded to 179–197 residues) and
nonspecific control short interfering RNA duplexes
(5�-UUCUCCGAACGUGUCACGUTT-3�) were designed and synthesized by
GenePharma (Shanghai Co., Ltd.). For transient knockdown of TCTP expression, PCI55
cells were transiently transfected with the resultant siRNA using Lipofectamine 2000.
The small hairpin RNA (shRNA)-encoding oligonucleotides for TCTP and scramble were
prepared by Sangon (Shanghai). The annealed shRNA were inserted into the AgeI and
EcoRI sites of the lentiviral plasmid pLKO.1-TRC cloning vector (Addgene, Cambridge,
USA). For stable knockdown of TCTP, HEK293T cells were plated in 75-cm2 culture
flasks and transfected with 10 μg TCTP shRNA or scramble shRNA lentiviral vectors.
The medium was changed the next day and viral supernatant was harvested 48 h later. All
viral containing medium was collected, passed through 0.45-μm syringe filters. PCI55,
Miapaca-2, and SW1990 cells were incubated with the lentivirus supernatant for 24 h and
selected with puromycin (4 μg/ml).
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 8
Cell cycle, cell apoptosis, and cell viability
Flow cytometric analysis was conducted to examine cell cycle and apoptosis with the
help of propidium iodine (Invitrogen) and human Annexin V-FITC Kit (Invitrogen),
respectively, according to the manufacturer’s protocol. Cell viability was determined at 0,
24, 48, 72, 96, 120 h using the Cell Counting Kit-8 reagent (Dojindo) according to the
manufacturer’s protocol. All observations were reproduced at least three times in
independent experiments.
Mouse xenografts
Female Balb/c nude mice (6–8 weeks of age, weighing 18–20 g, and specific
pathogen free) were obtained from Shanghai SLAC Laboratory Animals (Shanghai,
China). Before the experiment, mice were divided into four groups (Mipaca-2-scramble
shRNA, Miapaca-2-TCTP shRNA, SW1990-scramble shRNA, and SW1990-TCTP
shRNA). Each cancer cell line (5 × 106/site) was injected subcutaneously into the right
flank of a nude mouse. After establishment of the nude mice xenograft model, tumor
sizes were measured every 3 to 4 days using micrometer calipers. Tumor volumes were
calculated using the following formula: Volume � 1/2 a × b2, where a and b represent
the larger and smaller tumor diameters, respectively. Tumor growth was followed for 28
days from the first injection. All animal experiments were performed in compliance with
the Guidelines for the Care and Use of Laboratory Animals of Fudan University. The
protocol was approved by the Committee on the Ethics of Animal Experiments of Fudan
University (Permit Number, SYXK(Hu)2009-0082).
Patient and tissue samples
The study included 148 patients who underwent surgery at Fudan University
Shanghai Cancer Center from 2004 to 2012 after histological verification of pancreatic
ductal adenocarcinoma. None of these patients received preoperative chemotherapy or
radiotherapy. The patients provided written consent for the use of tumor tissue for
clinical research, and the Fudan University Shanghai Cancer Center Ethical Committee
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 9
approved the research protocol. Detailed description of patients and tissue samples is
provided in Supplementary Materials & Methods.
Immunohistochemical analysis of human pancreatic cancer tissues
Following deparaffinization and quenching of endogenous peroxidase, sections were
incubated with 1% bovine serum albumin (BSA) in PBS. Subsequently, the slides were
treated with rabbit anti-TCTP (1:500) and goat anti-Pim-3 (1:25) antibodies followed by
incubation with donkey anti-goat IgG and goat anti-rabbit IgG antibodies, respectively.
Pim-3 and TCTP immunoreactivities were visualized using the GT vision DAB kit
(GeneTech). The slides were counterstained with ChemMate Hematoxylin
(DakoCytomation) and mounted and observed under a microscope (Olympus). The
proportion of Pim-3- or TCTP-positive cells in human pancreatic carcinoma tissues were
evaluated by 2 independent pathologists without a prior knowledge on the clinical
information. The scoring of TCTP and Pim-3 was performed as described in
Supplementary Materials & Methods.
Statistical analysis
The χ2 test or Fisher’s exact probability test was used to compare clinicopathological
features of the 148 patients with TCTP and Pim-3 expression. Correlation between TCTP
and Pim-3 expression was evaluated using Spearman correlation analysis. Statistical
analysis was performed with SPSS statistical software (IBM SPSS Statistics 20). Data
were reported as the means ± SD when appropriate and p < 0.05 was considered
statistically significant.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 10
Results
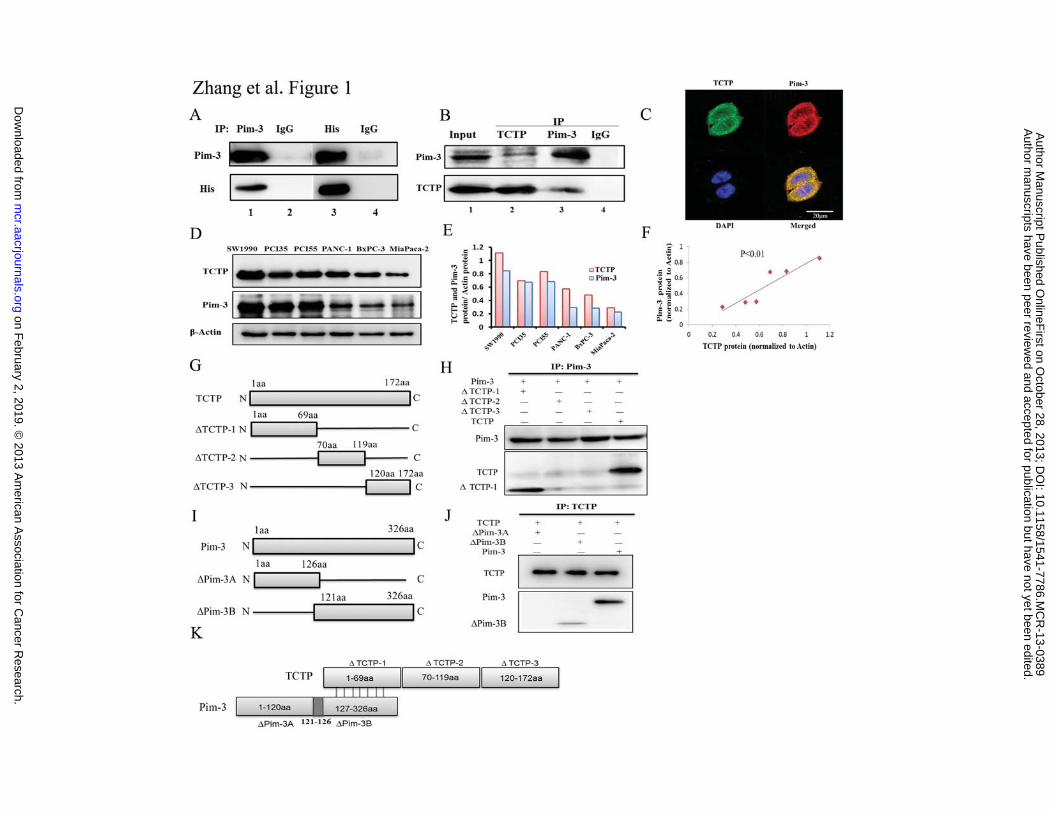
Interaction between Pim-3 and TCTP
We initially screened a cDNA library to identify the gene products that can modulate
Pim-3 activity as well as serve as its substrate. Using the yeast two-hybrid system with
Pim-3 as bait, six candidate genes were identi�ed (data not shown). Among them, we
characterized human TCTP protein (NM_003295.2) at first. To validate the speci�city of
the interaction between Pim-3 and TCTP, His-tagged TCTP protein was co-expressed
with Pim-3 in HEK293T cells. Immunoprecipitation with anti-Pim-3 antibodies but not
control IgG, co-precipitated His-tagged TCTP (Fig. 1A, compare lanes 1 and 2).
Moreover, immunoprecipitation with anti-His antibodies but not control IgG also
speci�cally co-precipitated Pim-3 protein (Fig. 1A, lane 3 and 4). Next, we examined the
interaction between Pim-3 and TCTP in PCI55 cells at endogenous levels.
Immunoprecipitation with anti-TCTP antibodies, but not control IgG, co-precipitated
endogenous Pim-3 (Fig. 1B, compare lanes 2 and 4). In a reciprocal co-IP experiment,
the TCTP protein was found to be present in the immune complexes pulled down by the
anti-Pim-3 antibodies but not by the control IgG (Fig. 1B, compare lanes 3 and 4).
Furthermore, immunofluorescence analysis showed that TCTP and Pim-3 were
predominantly localized in the cytoplasm and co-localized in human pancreatic cancer
cell lines (Fig. 1C). These observations indicate a physical interaction between Pim-3
and TCTP, and prompted us to examine TCTP expression in human pancreatic cancer cell
lines. We detected TCTP mRNA (data not shown) and protein expression in all six human
pancreatic cancer cell lines that we examined (Fig. 1D and 1E). Moreover, TCTP
expression correlated with Pim-3 expression in these cell lines (Spearman correlation
coef�cient, 1.0; p < 0.01) (Fig. 1F). To map the region necessary for the interaction
between Pim-3 and TCTP, we constructed truncation vectors of TCTP and Pim-3. As
shown in Fig. 1G and 1H, Pim-3 was co-precipitated with �TCTP-1 deletion mutants but
not with the other deletion mutant (�TCTP-2 and �TCTP-3). Moreover, the Pim-3
mutant lacking the C-terminal domain was not co-precipitated by TCTP, whereas the
Pim-3 mutant lacking N-terminal domain was (Fig. 1I and 1J). Taken together, these
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 11
results indicate that the interaction between Pim-3 and TCTP is immediately mediated
through the C-terminal region of Pim-3 and N-terminal region of TCTP (Fig. 1K).
Pim-3 has no effect on TCTP expression or phosphorylation
We previously observed that Pim-3 can prevent cell apoptosis and eventually lead to
progression of human pancreatic cancer (4). Hence, we postulated that functional
interaction of Pim-3 and TCTP can promote human pancreatic carcinogenesis. When
HEK293T cells with endogenous TCTP protein were transiently transfected with the
Pim-3 expression vector, TCTP protein expression did not change (Fig. 2A). Moreover,
TCTP protein levels were marginally affected even when the amounts of Pim-3 (Fig. 2B)
or incubation periods were increased (data not shown). Furthermore, when MiaPaca-2
cells with a low level of Pim-3 protein, were transiently transfected with Pim-3, TCTP
expression was not enhanced (Fig. 2C). When PCI55 cells with a high level of Pim-3
protein was depleted of endogenous Pim-3, TCTP expression did not change discernibly
(Fig. 2D). It was previously reported that TCTP can be phosphorylated mainly at Ser46,
but not at Thr65 (26). Pim-3 overexpression and ablation of Pim-3 expression increased
and decreased the amount of phospho-Ser112 Bad, respectively, but with few effects on
the levels of phospho-TCTPser46 or total amount of Bad in human pancreatic cancer cells
(Fig. 2E and 2F). These observations indicate that Pim-3 has minimal effect on TCTP
expression or phosphorylation states.
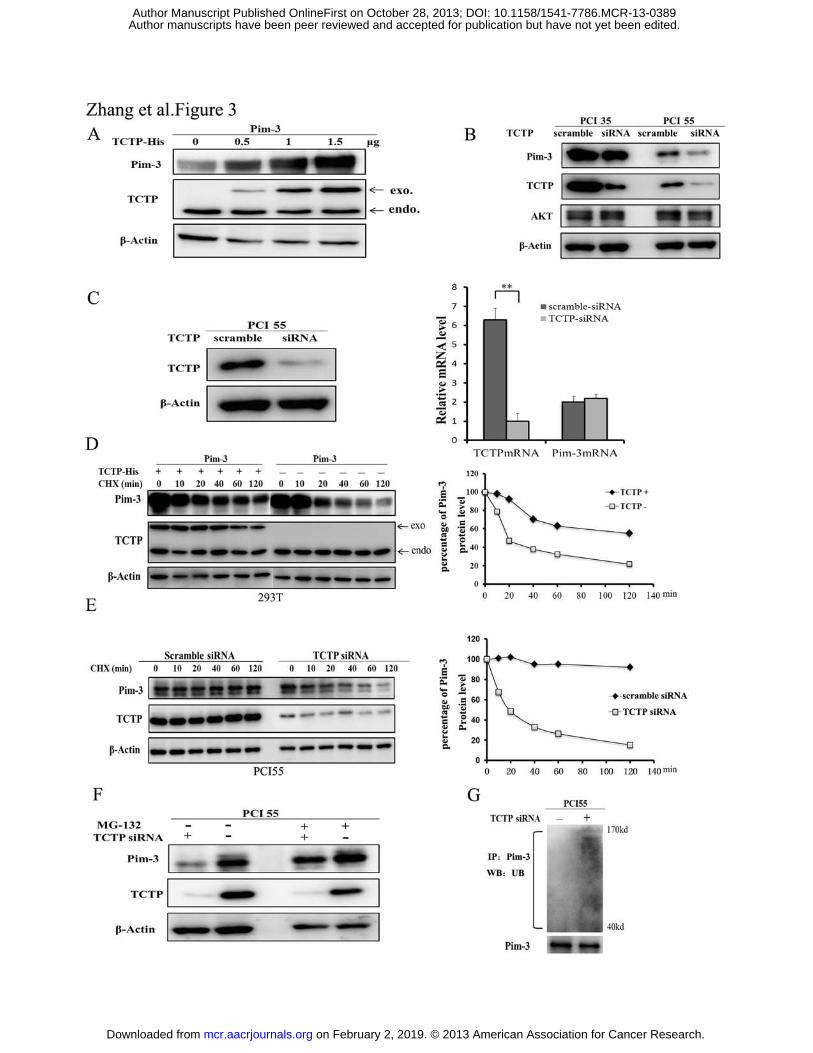
TCTP enhances the protein stability of Pim-3 by blocking ubiquitin–proteasome
degradation of Pim-3
As TCTP can stabilize Mcl-1 protein (15), we assumed that TCTP may enhance
Pim-3 protein levels by stabilizing Pim-3. To address this possibility, we transiently
co-transfected HEK293T cells with a fixed amount of Pim-3 and increasing quantities of
the His-tagged TCTP expression vectors. Pim-3 protein levels increased as the amounts
of the transfected His-tagged TCTP vectors were increased (Fig. 3A). Moreover, ablation
of endogenous TCTP protein significantly decreased the amount of Pim-3 protein but not
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 12
Akt, indicating the specificity of the TCTP-mediated effect on Pim-3 levels (Fig. 3B).
The elevation of these protein levels could be either due to enhanced mRNA expression,
protein synthesis, or inhibition of protein degradation. We found that TCTP siRNA
treatment did not affect Pim-3 mRNA levels (Fig. 3C). On the contrary, transient
transfection of the TCTP expression vector in HEK293T cells delayed the degradation of
Pim-3 protein (Fig. 3D). Conversely, ablation of TCTP with siRNA enhanced the
degradation of Pim-3 protein, compared to scrambled siRNA in PCI55 cells (Fig. 3E).
Moreover, TCTP siRNA-mediated enhancement in Pim-3 protein degradation was
abrogated by a proteasomal inhibitor, MG132 (Fig. 3F). Furthermore, TCTP siRNA
treatment promoted Pim-3 ubiquitination in PCI55 cells (Fig. 3G). Taken together, these
results indicate that TCTP can increase Pim-3 protein stability by blocking the
ubiquitin–proteasome-mediated degradation of Pim-3 in human pancreatic cancer cells.
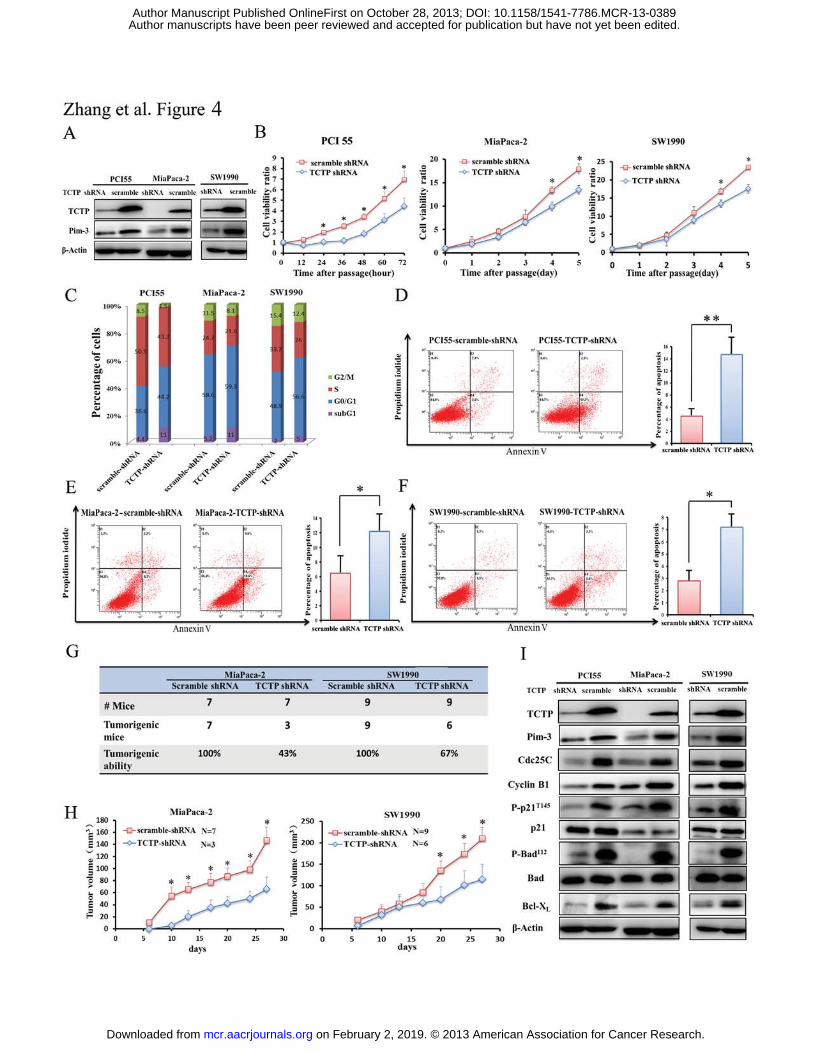
Ablation of TCTP protein prevents Pim-3-mediated tumor growth by arresting cell
cycle progression and enhancing apoptosis
Ablation of TCTP can destabilize Pim-3 protein in human pancreatic cancer cells
while reduction of Pim-3 expression can arrest cell cycle progression and promote
apoptosis (4). Hence, we investigated whether ablation of endogenous TCTP would
affect the cell cycle and apoptosis in human pancreatic cancer cells. TCTP shRNA, but
not scrambled shRNA treatment markedly diminished Pim-3 proteins in PCI55,
MiaPaca-2, and SW1990 cell lines (Fig. 4A) and signi�cantly retarded cell proliferation
compared to scramble shRNA (Fig. 4B). TCTP shRNA treatment increased the ratio of
G0/G1 phase cell population with reduced S and G2/M phase population, compared with
scrambled shRNA treatment (Fig. 4C). Moreover, the stable transfection of TCTP shRNA
resulted in a markedly higher ratio of apoptotic cells as evidenced by enhanced
phosphatidylserine externalization (Fig. 4D–4F). Furthermore, in comparison to a
subcutaneous injection with control shRNA-transfected cells, subcutaneous injection
with TCTP shRNA-transfected cells resulted in a lower tumor formation frequency and a
smaller tumor mass in nude mice 4 weeks after injection, (43% for MiaPaca-2-TCTP
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 13
shRNA cells and 67% for SW1990-TCTP shRNA cells, respectively; 100% for both of
MiaPaca-2-scramble shRNA and SW1990-scramble shRNA cells) (Fig. 4G and 4H).
Consistent with these observations, ablation of TCTP decreased Pim-3 expression and
the amounts of phosphorylated p21, the molecules that participate in cell progression at
G0/G1 phase at the tumor sites in the injected mice (27) (Fig. 4I). Cyclin B1 and Cdc25C,
molecules that participate in cell progression at G2/M phase, were also decreased (Fig.
4I). Likewise, ablation of TCTP decreased Pim-3 expression, and eventually diminished
the amounts of phosphor-Ser112-Bad and Bcl-XL at the tumor sites in the injected mice,
without any effects on the amounts of Bad (Fig. 4I) and some other proteins related with
cell cycle (data not show). Collectively, these observations indicate that TCTP can
stabilize Pim-3 and promote cell cycle progression and cell viability, eventually
promoting human pancreatic carcinogenesis.
Clinical relevance of TCTP and Pim-3 expression in pancreatic ductal
adenocarcinoma (PDAC)
Finally, to investigate the clinical relevance of TCTP and Pim-3 expression, we
performed immunohistochemistry on 148 resected PDAC tissues. TCTP and Pim-3
staining patterns were cytoplasmic, and both TCTP and Pim-3 expression were positive
in more than 90% of PDAC tissues. Immunofluorescence analysis showed that TCTP and
Pim-3 localized to the cytoplasm and co-localized in human pancreatic cancer tissues
(Fig. 5A). Moreover, TCTP protein was abundantly detected in malignant ductal
epithelium cells and some pancreatic acinar cells, but not in normal pancreatic duct
epithelial cell adjacent to tumor foci (Fig. 5B). Positive reactions were not observed
when control IgG was used as the primary antibody instead of the anti-TCTP antibody
(data not shown), indicating the specificity of the reaction. Furthermore, the expression
patterns of TCTP and Pim-3 exhibited perfect concordance in serial sections of human
pancreatic cancer tissues (Fig. 5C). TCTP expression signi�cantly correlated with Pim-3
expression in 148 human pancreatic ductal adenocarcinoma specimens (Spearman
correlation coef�cient, 0.518; p < 0.01) (Fig. 5D). Furthermore, higher TCTP and Pim-3
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 14
expression in PDAC was signi�cantly associated with an advanced tumor stage (p =
0.001 and p = 0.008; Table 1). Finally, higher TCTP expression also correlated with
nodal metastasis (p = 0.024), but neither TCTP nor Pim-3 expression did show any
significant correlation with age, gender, tumor location, tumor size, and tumor
differentiation (Table 1). These observations suggest that enhanced TCTP and Pim-3
expression may be involved in the malignant progression of human pancreatic cancer.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 15
Discussion
We previously observed that a proto-oncogene with serine/threonine kinase activity,
Pim-3, is aberrantly expressed in various malignant lesions, but not normal tissues of
endoderm-derived organs such as the liver, pancreas, colon, and stomach (3-5, 28).
Moreover, it can contribute to tumorigenesis by inhibiting the apoptosis of tumor cells
and promoting their cell cycle progression. Pim kinase family consists of three members,
Pim-1, Pim-2, and Pim-3, which exhibit marked sequence similarity, especially in their
kinase domains. As Pim kinases do not possess a regulatory domain and are
constitutively active when they are expressed (29), the activity of Pim kinases is largely
regulated at transcriptional, translation, and post-translational levels (30). Furthermore, a
very short half-life of their mRNA and protein (31) suggests the importance of regulation
of Pim kinase protein levels.
Pim-1 and Pim-3 have been shown to bind to the serine/threonine protein
phosphatase 2A (PP2A), resulting in their dephosphorylation, ubiquitination, and
proteasomal degradation (32, 33). Moreover, heat shock 70-kDa protein 1A/B (HsP70)
binds ubiquitylated Pim-1 and promotes its proteasomal degradation. On the contrary,
heat shock protein 90� (HsP90) can stabilize Pim-1 protein by binding to it, and the
inhibition of HsP90 induced rapid degradation of Pim-1 (34, 35). However, the molecular
mechanisms of Pim-3 expression in carcinogenesis still remain largely unknown. To
de�ne the detailed regulatory machinery, we conducted yeast two-hybrid screening to
identify Pim-3-interacting proteins and demonstrated for the first time that TCTP directly
interacts with Pim-3. TCTP is a multi-functional protein and can interact with many
cellular proteins. TCTP binds to p53 to promote its proteasomal degradation (24, 25),
whereas TCTP interacts with Mcl-1 to enhance its stability (15). Indeed, Pim-3 protein
stability is enhanced by its interaction with TCTP, which can block the
ubiquitin-proteasome-mediated degradation of Pim-3.
Comparisons of TCTP sequences in 24 eukaryotes (36) revealed the presence of two
highly conserved regions in TCTP protein; one region from 45 to 55 residues and another
region from 129 to 147 residues. Several proteins can interact with TCTP by binding to
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 16
either region (Supplementary Table S3) (26, 36-40). We proved that the N-terminal
portion of TCTP (residues 1–69) and the C-terminal portion of Pim-3 were required for
their physical interaction. Although the crystal structure of Pim-3 has not yet been
determined, Pim-3 seems to exhibit a similar structure to Pim-1, due to its extraordinarily
high amino acid sequence similarity with Pim-3. The Pim-1 kinase structure adopts a
two-lobe kinase fold with a deep and intervening cleft (29). The N-terminal and
C-terminal lobes are composed of β-sheets and α-helices, respectively, while the two
domains are connected through the hinge region (residues 121-126). TCTP could interact
with the C-terminal but not the N-terminal portion-possessing Pim-3 mutant, indicating
that the α-helices in the C-terminal portion are mainly involved in binding to TCTP.
Pim kinases can phosphorylate p27Kip1 and regulate its expression at both
transcriptional and post-translational levels, to promote tumorigenesis (41). Moreover,
Pim-2 overexpression in HCT116 cells leads to enhanced phosphorylation of p21 to
increase its stability (42). Furthermore, Pim-3 can augment protein synthesis (13) and
regulate transcriptional activity of Myc (31). However, overexpression or ablation of
Pim-3 failed to induce any changes in TCTP protein expression or phospho-TCTPser46
levels. Thus, Pim-3 has few effects on TCTP expression or phosphorylation levels, even
though Pim-3 can bind to TCTP.
TCTP was overexpressed in many human cancer tissues including liver (43),
colorectal (44), prostate (45), breast (46), and lung cancer (47). Likewise, TCTP was
detected in pancreatic cancer cell lines, and malignant duct epithelial cells and some
normal acinar cells but not normal pancreatic duct epithelial cells adjacent to tumor foci.
Moreover, TCTP expression positively correlated with Pim-3 in pancreatic cancer cell
lines and tissues. Furthermore, patients with high TCTP and Pim-3 expression often had
advanced tumor stage. Thus, TCTP may promote pancreatic cancer by preserving Pim-3
expression.
All Pim kinase members can bind to and phosphorylate CDK inhibitor, p27, at its
threonine residues and induce the binding of p27 to 14-3-3 protein, resulting in its
nuclear export and proteasome-dependent degradation (41). Moreover, Pim-1 can
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 17
promote cell cycle progression by phosphorylating and modulating the functions of
molecules involved in cell cycle progression such as Cdc25A, cyclinD1-associated
kinases (48), Cdc25C-associated kinase 1 (C-TAK1)(49), and G1-specific inhibitor p21
(Waf) (50). Given the high sequence identity between Pim-1 and Pim-3, it is likely that
Pim-3 can also modulate these molecules like Pim-1. Indeed, cell cycle progression is
accelerated in hepatocytes of transgenic mice, which express human Pim-3 cDNA
selectively in hepatocytes (7). Moreover, a small-molecule Pim-3 kinase inhibitor
markedly retarded in vitro growth of human pancreatic cancer cell lines by inducing
G2/M arrest (51), suggesting a potential role for Pim-3 in cell cycle progression.
Consistently, ablation of TCTP decreased the amounts of Pim-3, Cdc25C, cyclin B1, and
phospho-p21T145, but not the total amounts of p21. Furthermore, TCTP ablation arrested
cell cycle progression at the G1 phase in human pancreatic cancer cells, similar to when
the cells were depleted of Pim-3.
Diraison and colleagues recently reported that increased TCTP expression reduced
the sensitivity of pancreatic β cells to apoptosis (52). We previously observed that Pim-3
shRNA treatment in vitro enhanced the apoptosis of various types of cancer cells (3-5).
Likewise, TCTP shRNA destabilized and induced the degradation of Pim-3, and
promoted the apoptosis of human pancreatic cancer cells. Moreover, TCTP
shRNA-treated human pancreatic cancer cells exhibited a weaker tumorigenic ability
than scrambled shRNA-treated cells, when they were injected into nude mice.
Furthermore, we detected a much lower level of Pim-1 and Pim-2 mRNA than Pim-3
mRNA in human pancreatic cancer cells that we examined (Supplementary Fig. S1).
Thus, it is likely that TCTP can regulate the cell cycle and cell apoptosis mainly by
Pim-3-mediated in human pancreatic cancer cells.
Several independent groups developed small-molecule inhibitors against Pim kinases
including flavonol quercetargetin, imidazole[1,2-b]pyridazines,
bezylindene-thiazolidine-2,4-dione, 3,5-disubstituted indole derivatives,
pyrazolo[3,4-g]quinoxaline derivatives, 1,6-dihydropyrazolo[4,3-c]carbazoles and
3,6-dihydropyrazolo[3,4-c]carbazoles derivatives (53), and pyrrolo[2,3-a]carbazole and
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 18
pyrrolo[2,3-g]indazoles derivatives (54, 55). Among them,
1,6-dihydropyrazolo[4,3-c]carbazoles, 3,6-dihydropyrazolo[3,4-c]carbazoles and
pyrrolo[2,3-g]indazoles could be used as an interesting molecular tool to study Pim-3
biological functions (53). Consistently, we also demonstrated that stemonamide synthetic
intermediates derivative can inhibit Pim-3 as well as Pim-1 and Pim-2 activities and can
reduce tumor growth in vivo xenograft models using a human pancreatic cancer cell line
without causing major adverse effects (56, 57). TCTP but not Pim-3 expression was well
correlated with nodal metastasis in human pancreatic cancer patients. Thus, given the
fact that TCTP can bind and modulate several molecules, TCTP can contribute to
pancreatic cancer development and progression besides its effects on Pim-3. If so, the
targeting of TCTP may supplement Pim-3 inhibitors for the treatment of various types of
cancer, which exhibit enhanced Pim-3 and TCTP expression.
Acknowledgments
The authors thank Dr. Yi Qin, Hong-yu Gu, Ping Zhang, Yu-hu Xin and Yue Cao of
the Cancer Research Institute, Fudan University Shanghai Cancer Center for their
technical support.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 19
References: 1. Konietzko U, Kauselmann G, Scafidi J, Staubli U, Mikkers H, Berns A, et al.. Pim kinase
expression is induced by LTP stimulation and required for the consolidation of enduring LTP.
Embo J 1999;18(12):3359-69.
2. Deneen B, Welford SM, Ho T, Hernandez F, Kurland I, Denny CT. PIM3 proto-oncogene
kinase is a common transcriptional target of divergent EWS/ETS oncoproteins. Mol Cell Biol
2003;23(11):3897-908.
3. Fujii C, Nakamoto Y, Lu P, Tsuneyama K, Popivanova BK, Kaneko S, et al.. Aberrant
expression of serine/threonine kinase Pim-3 in hepatocellular carcinoma development and
its role in the proliferation of human hepatoma cell lines. Int J Cancer 2005;114(2):209-18.
4. Li YY, Popivanova BK, Nagai Y, Ishikura H, Fujii C, Mukaida N. Pim-3, a proto-oncogene
with serine/threonine kinase activity, is aberrantly expressed in human pancreatic cancer and
phosphorylates bad to block bad-mediated apoptosis in human pancreatic cancer cell lines.
Cancer Res 2006;66(13):6741-7.
5. Popivanova BK, Li YY, Zheng H, Omura K, Fujii C, Tsuneyama K, et al.. Proto-oncogene,
Pim-3 with serine/threonine kinase activity, is aberrantly expressed in human colon cancer
cells and can prevent Bad-mediated apoptosis. Cancer Sci 2007;98(3):321-8.
6. Zheng HC, Tsuneyama K, Takahashi H, Miwa S, Sugiyama T, Popivanova BK, et al..
Aberrant Pim-3 expression is involved in gastric adenoma-adenocarcinoma sequence and
cancer progression. J Cancer Res Clin Oncol 2008;134(4):481-8.
7. Wu Y, Wang YY, Nakamoto Y, Li YY, Baba T, Kaneko S, et al.. Accelerated hepatocellular
carcinoma development in mice expressing the Pim-3 transgene selectively in the liver.
Oncogene 2010;29(15):2228-37.
8. Zha J, Harada H, Yang E, Jockel J, Korsmeyer SJ. Serine phosphorylation of death agonist
BAD in response to survival factor results in binding to 14-3-3 not BCL-X(L). Cell
1996;87(4):619-28.
9. Yang E, Zha J, Jockel J, Boise LH, Thompson CB, Korsmeyer SJ. Bad, a heterodimeric
partner for Bcl-XL and Bcl-2, displaces Bax and promotes cell death. Cell
1995;80(2):285-91.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 20
10. Mukaida N, Wang YY, Li YY. Roles of Pim-3, a novel survival kinase, in tumorigenesis.
Cancer Sci 2011;102(8):1437-42.
11. Xu D, Cobb MG, Gavilano L, Witherspoon SM, Williams D, White CD, et al.. Inhibition of
oncogenic Pim-3 kinase modulates transformed growth and chemosensitizes pancreatic
cancer cells to gemcitabine. Cancer Biol Ther 2013;14(6):492-501.
12. Wang C, Li HY, Liu B, Huang S, Wu L, Li YY. Pim-3 promotes the growth of human
pancreatic cancer in the orthotopic nude mouse model through vascular endothelium growth
factor. J Surg Res 2013.
13. Beharry Z, Mahajan S, Zemskova M, Lin YW, Tholanikunnel BG, Xia Z, et al.. The Pim
protein kinases regulate energy metabolism and cell growth. Proc Natl Acad Sci U S A
2011;108(2):528-33.
14. Li F, Zhang D, Fujise K. Characterization of fortilin, a novel antiapoptotic protein. J Biol
Chem 2001;276(50):47542-9.
15. Liu H, Peng HW, Cheng YS, Yuan HS, Yang-Yen HF. Stabilization and enhancement of the
antiapoptotic activity of mcl-1 by TCTP. Mol Cell Biol 2005;25(8):3117-26.
16. Bohm H, Benndorf R, Gaestel M, Gross B, Nurnberg P, Kraft R, et al.. The growth-related
protein P23 of the Ehrlich ascites tumor: translational control, cloning and primary structure.
Biochem Int 1989;19(2):277-86.
17. Gross B, Gaestel M, Bohm H, Bielka H. cDNA sequence coding for a translationally
controlled human tumor protein. Nucleic Acids Res 1989;17(20):8367.
18. Chitpatima ST, Makrides S, Bandyopadhyay R, Brawerman G. Nucleotide sequence of a
major messenger RNA for a 21 kilodalton polypeptide that is under translational control in
mouse tumor cells. Nucleic Acids Res 1988;16(5):2350.
19. Thomas G, Thomas G, Luther H. Transcriptional and translational control of cytoplasmic
proteins after serum stimulation of quiescent Swiss 3T3 cells. Proc Natl Acad Sci U S A
1981;78(9):5712-6.
20. Oikawa K, Ohbayashi T, Mimura J, Fujii-Kuriyama Y, Teshima S, Rokutan K, et al.. Dioxin
stimulates synthesis and secretion of IgE-dependent histamine-releasing factor. Biochem
Biophys Res Commun 2002;290(3):984-7.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 21
21. Nielsen HV, Johnsen AH, Sanchez JC, Hochstrasser DF, Schiotz PO. Identification of a
basophil leukocyte interleukin-3-regulated protein that is identical to IgE-dependent
histamine-releasing factor. Allergy 1998;53(7):642-52.
22. Teshima S, Rokutan K, Nikawa T, Kishi K. Macrophage colony-stimulating factor stimulates
synthesis and secretion of a mouse homolog of a human IgE-dependent histamine-releasing
factor by macrophages in vitro and in vivo. J Immunol 1998;161(11):6356-66.
23. Koziol MJ, Gurdon JB. TCTP in development and cancer. Biochem Res Int
2012;2012:105203.
24. Rho SB, Lee JH, Park MS, Byun HJ, Kang S, Seo SS, et al.. Anti-apoptotic protein TCTP
controls the stability of the tumor suppressor p53. Febs Lett 2011;585(1):29-35.
25. Amson R, Pece S, Lespagnol A, Vyas R, Mazzarol G, Tosoni D, et al.. Reciprocal repression
between P53 and TCTP. Nat Med 2012;18(1):91-9.
26. Yarm FR. Plk phosphorylation regulates the microtubule-stabilizing protein TCTP. Mol Cell
Biol 2002;22(17):6209-21.
27. Zhang Y, Wang Z, Magnuson NS. Pim-1 kinase-dependent phosphorylation of
p21Cip1/WAF1 regulates its stability and cellular localization in H1299 cells. Mol Cancer
Res 2007;5(9):909-22.
28. Zheng HC, Tsuneyama K, Takahashi H, Miwa S, Sugiyama T, Popivanova BK, et al..
Aberrant Pim-3 expression is involved in gastric adenoma-adenocarcinoma sequence and
cancer progression. J Cancer Res Clin Oncol 2008;134(4):481-8.
29. Qian KC, Wang L, Hickey ER, Studts J, Barringer K, Peng C, et al.. Structural basis of
constitutive activity and a unique nucleotide binding mode of human Pim-1 kinase. J Biol
Chem 2005;280(7):6130-7.
30. Amaravadi R, Thompson CB. The survival kinases Akt and Pim as potential pharmacological
targets. J Clin Invest 2005;115(10):2618-24.
31. Nawijn MC, Alendar A, Berns A. For better or for worse: the role of Pim oncogenes in
tumorigenesis. Nat Rev Cancer 2011;11(1):23-34.
32. Ma J, Arnold HK, Lilly MB, Sears RC, Kraft AS. Negative regulation of Pim-1 protein kinase
levels by the B56beta subunit of PP2A. Oncogene 2007;26(35):5145-53.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 22
33. Losman JA, Chen XP, Vuong BQ, Fay S, Rothman PB. Protein phosphatase 2A regulates the
stability of Pim protein kinases. J Biol Chem 2003;278(7):4800-5.
34. Shay KP, Wang Z, Xing PX, McKenzie IF, Magnuson NS. Pim-1 kinase stability is regulated
by heat shock proteins and the ubiquitin-proteasome pathway. Mol Cancer Res
2005;3(3):170-81.
35. Mizuno K, Shirogane T, Shinohara A, Iwamatsu A, Hibi M, Hirano T. Regulation of Pim-1
by Hsp90. Biochem Biophys Res Commun 2001;281(3):663-9.
36. Thaw P, Baxter NJ, Hounslow AM, Price C, Waltho JP, Craven CJ. Structure of TCTP
reveals unexpected relationship with guanine nucleotide-free chaperones. Nat Struct Biol
2001;8(8):701-4.
37. Yang Y, Yang F, Xiong Z, Yan Y, Wang X, Nishino M, et al.. An N-terminal region of
translationally controlled tumor protein is required for its antiapoptotic activity. Oncogene
2005;24(30):4778-88.
38. Cans C, Passer BJ, Shalak V, Nancy-Portebois V, Crible V, Amzallag N, et al..
Translationally controlled tumor protein acts as a guanine nucleotide dissociation inhibitor on
the translation elongation factor eEF1A. Proc Natl Acad Sci U S A 2003;100(24):13892-7.
39. Yoon T, Jung J, Kim M, Lee KM, Choi EC, Lee K. Identification of the self-interaction of rat
TCTP/IgE-dependent histamine-releasing factor using yeast two-hybrid system. Arch
Biochem Biophys 2000;384(2):379-82.
40. Kim M, Jung Y, Lee K, Kim C. Identification of the calcium binding sites in translationally
controlled tumor protein. Arch Pharm Res 2000;23(6):633-6.
41. Morishita D, Katayama R, Sekimizu K, Tsuruo T, Fujita N. Pim kinases promote cell cycle
progression by phosphorylating and down-regulating p27Kip1 at the transcriptional and
posttranscriptional levels. Cancer Res 2008;68(13):5076-85.
42. Wang Z, Zhang Y, Gu JJ, Davitt C, Reeves R, Magnuson NS. Pim-2 phosphorylation of
p21(Cip1/WAF1) enhances its stability and inhibits cell proliferation in HCT116 cells. Int J
Biochem Cell Biol 2010;42(6):1030-8.
43. Chan TH, Chen L, Liu M, Hu L, Zheng BJ, Poon VK, et al.. Translationally controlled tumor
protein induces mitotic defects and chromosome missegregation in hepatocellular carcinoma
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 23
development. Hepatology 2012;55(2):491-505.
44. Slaby O, Sobkova K, Svoboda M, Garajova I, Fabian P, Hrstka R, et al.. Significant
overexpression of Hsp110 gene during colorectal cancer progression. Oncol Rep
2009;21(5):1235-41.
45. Arcuri F, Papa S, Carducci A, Romagnoli R, Liberatori S, Riparbelli MG, et al..
Translationally controlled tumor protein (TCTP) in the human prostate and prostate cancer
cells: expression, distribution, and calcium binding activity. Prostate 2004;60(2):130-40.
46. Deng SS, Xing TY, Zhou HY, Xiong RH, Lu YG, Wen B, et al.. Comparative proteome
analysis of breast cancer and adjacent normal breast tissues in human. Genomics Proteomics
Bioinformatics 2006;4(3):165-72.
47. Kim JE, Koo KH, Kim YH, Sohn J, Park YG. Identification of potential lung cancer
biomarkers using an in vitro carcinogenesis model. Exp Mol Med 2008;40(6):709-20.
48. Mochizuki T, Kitanaka C, Noguchi K, Muramatsu T, Asai A, Kuchino Y. Physical and
functional interactions between Pim-1 kinase and Cdc25A phosphatase. Implications for the
Pim-1-mediated activation of the c-Myc signaling pathway. J Biol Chem
1999;274(26):18659-66.
49. Bachmann M, Hennemann H, Xing PX, Hoffmann I, Moroy T. The oncogenic
serine/threonine kinase Pim-1 phosphorylates and inhibits the activity of Cdc25C-associated
kinase 1 (C-TAK1): a novel role for Pim-1 at the G2/M cell cycle checkpoint. J Biol Chem
2004;279(46):48319-28.
50. Wang Z, Bhattacharya N, Mixter PF, Wei W, Sedivy J, Magnuson NS. Phosphorylation of the
cell cycle inhibitor p21Cip1/WAF1 by Pim-1 kinase. Biochim Biophys Acta
2002;1593(1):45-55.
51. Wang YY, Taniguchi T, Baba T, Li YY, Ishibashi H, Mukaida N. Identification of a
phenanthrene derivative as a potent anticancer drug with Pim kinase inhibitory activity.
Cancer Sci 2012;103(1):107-15.
52. Diraison F, Hayward K, Sanders KL, Brozzi F, Lajus S, Hancock J, et al.. Translationally
controlled tumour protein (TCTP) is a novel glucose-regulated protein that is important for
survival of pancreatic beta cells. Diabetologia 2011;54(2):368-79.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 24
53. Suchaud V, Gavara L, Saugues E, Nauton L, Thery V, Anizon F, et al.. Identification of
1,6-dihydropyrazolo[4,3-c]carbazoles and 3,6-dihydropyrazolo[3,4-c]carbazoles as new Pim
kinase inhibitors. Bioorg Med Chem 2013;21(14):4102-11.
54. Akue-Gedu R, Letribot B, Saugues E, Debiton E, Anizon F, Moreau P. Kinase inhibitory
potencies and in vitro antiproliferative activities of N-10 substituted pyrrolo[2,3-a]carbazole
derivatives. Bioorg Med Chem Lett 2012;22(11):3807-9.
55. Gavara L, Suchaud V, Nauton L, Thery V, Anizon F, Moreau P. Identification of
pyrrolo[2,3-g]indazoles as new Pim kinase inhibitors. Bioorg Med Chem Lett
2013;23(8):2298-301.
56. Li YY, Wang YY, Taniguchi T, Kawakami T, Baba T, Ishibashi H, et al.. Identification of
stemonamide synthetic intermediates as a novel potent anticancer drug with an
apoptosis-inducing ability. Int J Cancer 2010;127(2):474-84.
57. Wang Z, Li XM, Shang K, Zhang P, Wang CF, Xin YH, et al.. T-18, a stemonamide synthetic
intermediate inhibits Pim kinase activity and induces cell apoptosis, acting as a potent
anticancer drug. Oncol Rep 2013;29(3):1245-51.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389
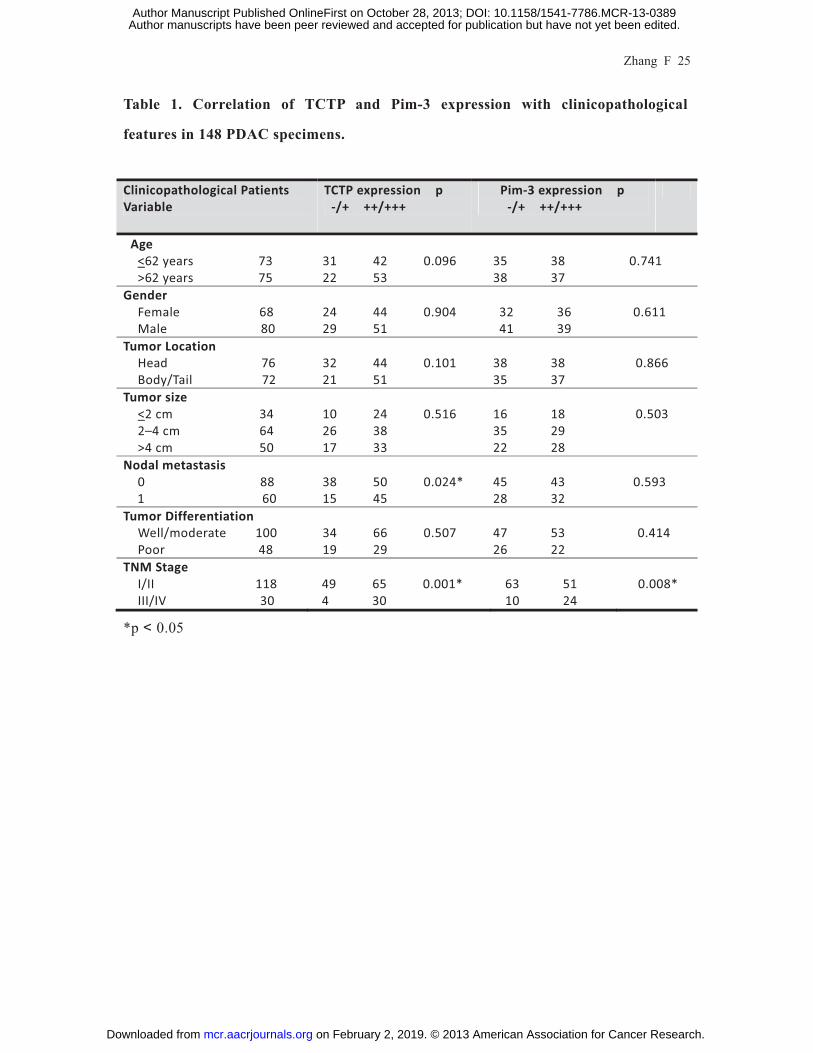
Zhang F 25
Table 1. Correlation of TCTP and Pim-3 expression with clinicopathological
features in 148 PDAC specimens.
Clinicopathological Patients Variable
TCTP expression p -/+ ++/+++
Pim-3 expression p -/+ ++/+++
Age <62 years 73
>62 years 75
31 42 0.096 22 53
35 38 38 37
0.741
Gender Female 68 Male 80
24 44 0.904 29 51
32 36 41 39
0.611
Tumor Location Head 76 Body/Tail 72
32 44 0.101 21 51
38 38 35 37
0.866
Tumor size <2 cm 34 2–4 cm 64 >4 cm 50
10 24 0.516 26 38 17 33
16 18 35 29 22 28
0.503
Nodal metastasis 0 88
1 60
38 50 0.024*15 45
45 43
28 32
0.593
Tumor Differentiation Well/moderate 100 Poor 48
34 66 0.507 19 29
47 53 26 22
0.414
TNM Stage I/II 118 III/IV 30
49 65 0.001* 4 30
63 51 10 24
0.008*
*p 0.05
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 26
Legends to Figures
Figure 1. Association of TCTP with Pim-3 in human pancreatic cancer cell lines. A.
Co-immunoprecipitation of TCTP and Pim-3 in transiently transfected HEK293T cells.
Cell lysates were obtained from HEK293T cells transiently transfected with
pcDNA4-Pim-3 and pcDNA4-TCTP-His. The resultant lysates were subjected to
immunoprecipitation and immunoblotting. B. Co-immunoprecipitation of endogenous
TCTP and Pim-3 in PCI55 cells. Cell lysates were obtained from PCI55 cells. The
resultant lysates were subjected to immunoprecipitation and immunoblotting. One tenth
of the cell lysates from PCI55 were subjected to immunoblotting with anti-TCTP or
anti-Pim-3 antibodies as positive controls. C. Co-localization of TCTP and Pim-3 in
human pancreatic cancer cells. PCI55 cells were immunostained with a combination of
anti-TCTP and anti-Pim-3 antibodies as described in Materials and Methods. The
fluorescent images were digitally merged. Yellow coloration in overlay panels indicates
co-localization of Pim-3 and TCTP. Nuclei were counterstained with DAPI. D. TCTP and
Pim-3 expression in human pancreatic cancer cell lines was detected by western blotting.
�-actin was used as an internal control. E. The expression levels of TCTP, Pim-3, and
�-actin were quantified using NIH ImageJ software. TCTP/�-actin and Pim-3/�-actin
ratios were calculated for each cell line. F. Correlation between TCTP and Pim-3
expression in human pancreatic cancer cell lines, with a linear regression line and
Spearman correlation significance (Spearman correlation coefficient 1, p < 0.01). G.
Schematic representation of cDNA constructs for each TCTP deletion mutant. H. Cell
lysates were obtained from HEK293T cells transiently co-transfected with Pim-3 and
TCTP deletion mutants. The resultant lysates were immunoprecipitated and
immunoblotted. I. Schematic representation of cDNA constructs for each Pim-3 deletion
mutant. J. Cell lysates were obtained from HEK293T cells transiently co-transfected
with TCTP and Pim-3 deletion mutants. The resultant lysates were immunoprecipitated
and immunoblotted. K. Schematic representation of the presumed interaction between
Pim-3 and TCTP proteins.
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 27
Figure 2. Pim-3 has no effect on TCTP expression or phosphorylation. A and B. Cell
lysates were obtained from HEK293T cells transiently transfected with pcDNA4-Pim-3
expression vector, empty pcDNA4 vector (A), or the indicated amounts of Pim-3
expression vector (B). The resultant lysates were subjected to immunoblotting with the
indicated antibodies. C and D. Cell lysates were obtained from MiaPaca-2 cells
transiently transfected with pcDNA4-Pim-3 expression or empty pcDNA4 vector (C) or
from PCI55 cells transiently transfected with Pim-3 shRNA or scrambled shRNA (D).
The resultant lysates were subjected to immunoblotting with the indicated antibodies. E
and F. Cell lysates were obtained from MiaPaca-2 cells stably overexpressing
pMEI-5-Pim-3 or empty pMEI-5 vector (E) or from PCI55 cells stably transfected with
Pim-3 or scramble shRNA vectors (F). The resultant lysates were subjected to
immunoblotting with the indicated antibodies.
Figure 3. TCTP enhances the protein stability of Pim-3 by blocking the
ubiquitin-proteasome degradation of Pim-3. A. pcDNA4-Pim-3 expression vector (0.5
μg) was co-transfected with the indicated amount of pcDNA4-TCTP-His or empty
pcDNA4 vector into HEK293T cells. After 48 h, cell lysates were analyzed by western
blotting with the indicated antibodies. B. The cell lysates were obtained from PCI35 or
PCI55 cells transiently transfected with TCTP or scramble siRNA. The resultant lysates
were subjected to immunoblotting with the indicated antibodies. C. Immunoblotting for
TCTP and real-time qRT-PCR for TCTP and Pim-3 in TCTP siRNA-treated PCI55 cells
(left). Cell lysates were obtained from PCI55 cells transiently transfected with TCTP or
scramble siRNA. After 48 h, the cell lysates were obtained and subjected to
immunoblotting. Real-time RT-PCR analysis was performed to quantify TCTP and Pim-3
mRNA levels as described in Materials and Methods. The results are expressed as mean
± SD **, p < 0.01 vs scrambled siRNA (right). D. pcDNA4-Pim-3 expression vector (0.5
μg) was co-transfected with pcDNA4-TCTP-His (+; 1.5 μg) or empty pcDNA4 (-;
1.5 μg) vector into HEK293T cells. At 24 h after transfection, the cells were treated with
cycloheximide (CHX, 30μg/ml) for various time intervals. Cell lysates were then
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 28
analyzed by immunoblotting with indicated antibodies (left). The percentage of Pim-3
protein level was determined using densitometry scanning (NIH ImageJ software) (right).
E. TCTP or control scramble siRNA was transfected transiently into PCI55 cells. At 24 h
after transfection, the cells were treated with cycloheximide for various times as
indicated. Cell lysates were then analyzed by immunoblotting with the indicated
antibodies (left). The percentage of Pim-3 protein level was determined using
densitometry scanning (NIH ImageJ software) (right). F. PCI55 cells were transiently
transfected with TCTP (+) or scrambled siRNA (-). At 24 h after transfection, the cells
were treated with either DMSO or MG132 (1 μg/ml) for an additional 4 h. Cell lysates
were obtained and subjected to the immunoblotting with the indicated antibodies. G.
PCI55 cells were transiently transfected with TCTP siRNA (+) or scrambled siRNA (-).
At 24 h after the transfection, the cell lysates were prepared and immunoprecipitated
with anti-Pim-3 antibodies. The immune complex was then analyzed by immunoblotting
with anti-ubiquitin (UB) or Pim-3 antibodies.
Figure 4. The effects of ablation of endogenous TCTP on cell viability and cell
apoptosis of human pancreatic cancer cell lines. A. TCTP or scramble shRNA was
stably infected into PCI55, MiaPaca-2, or SW1990 cells using a lentivirus vector. TCTP
and Pim-3 expression was detected by immunoblotting. �-actin was used as an internal
control. B. PCI55, MiaPaca-2, or SW1990 cells were stably transfected with TCTP
shRNA or scramble shRNA. Three thousand of the resultant cells were plated in a
96-well microplate. After 12 h, when the cells were adhered to the microplate, was
designated as time 0. The cell numbers were determined as the indicated time intervals
using the Cell Counting Kit-8 reagent and the ratios were compared to time 0. Point,
mean (n = 5); bars, SD. C. PCI55, MiaPaca-2, or SW1990 cells were stably transfected
with TCTP shRNA or scrambled shRNA. The proportion of cells in each cell cycle phase
was determined as described in Meterials and Methods. D–F. Apoptosis of PCI55,
MiaPaca-2, and SW1990 cells, which were stably transfected with TCTP or scrambled
shRNA. Cells were stained with a combination of propidium iodide and Annexin V as
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Zhang F 29
described in Materials and Methods. The number in each quadrant indicates the
proportion of the cells present in the quadrant. G. Incidence of tumor formation in nude
mice induced by injection of MiaPaca-2 or SW1990 cells, which were stably transfected
with TCTP or scramble shRNA. H. Tumor sizes were measured twice a week. The mean
and SD were calculated and are shown here. I. Cell lysates were obtained from PCI55,
MiaPaca-2, and SW1990 cells, which were stably transfected with TCTP or scramble
shRNA. The resultant lysates were subjected to immunoblotting with the indicated
antibodies as described in Material and Methods.
Figure 5. Clinicopathological analysis of TCTP and Pim-3 expression in pancreatic
ductal adenocarcinoma (PDAC). A. Co-localization of TCTP and Pim-3 in human
pancreatic cancer tissue. Human pancreatic cancer tissues were immunostained with a
combination of anti-TCTP and anti-Pim-3 antibodies as described in Materials and
Methods. The fluorescent images were digitally merged. B. TCTP expression was
detected on the section of normal pancreatic duct epithelium adjacent to tumor foci (left
and middle panels) and pancreatic adenocarcinoma (right panel) by
immunohistochemistry. C. TCTP and Pim-3 expression was analyzed by
immunohistochemistry on serial sections of different human pancreatic cancer tissues. D.
Cross-tabulation of TCTP and Pim-3 expression detected by immunohistochemistry in
148 pancreatic ductal adenocarcinoma tissues. Correlation between TCTP and Pim-3 was
significant on Spearman correlation analysis (Spearman correlation coefficient 0.518, p <
0.01).
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389
on February 2, 2019. ©
2013 Am
erican Association for C
ancer Research.
mcr.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 28, 2013; DO
I: 10.1158/1541-7786.MC
R-13-0389

on February 2, 2019. ©
2013 Am
erican Association for C
ancer Research.
mcr.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 28, 2013; DO
I: 10.1158/1541-7786.MC
R-13-0389
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389

Published OnlineFirst October 28, 2013.Mol Cancer Res Fei Zhang, Bin Liu, Zhen Wang, et al. Involvement in Pancreatic Cancer ProgressionA Novel Regulatory Mechanism of Pim-3 Kinase Stability and its
Updated version
10.1158/1541-7786.MCR-13-0389doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2013/10/28/1541-7786.MCR-13-0389.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/early/2013/10/26/1541-7786.MCR-13-0389To request permission to re-use all or part of this article, use this link
on February 2, 2019. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2013; DOI: 10.1158/1541-7786.MCR-13-0389