a novel nickel resistance determinant found in sewage-associated bacteria
TRANSCRIPT

Microb Ecol (1997) 33:230–239
q 1997 Springer-Verlag New York Inc.
A Novel Nickel Resistance Determinant Found in Sewage-Associated Bacteria
R.W. Pickup,1 H.E.H. Mallinson,1 G. Rhodes,1 L.K. Chatfield2
1 Institute of Freshwater Ecology, Windermere Laboratory, Far Sawrey, Ambleside, Cumbria, LA22 0LP, UK2 Department of Applied Biology, Univesity of Central Lancashire, Preston, Lancashire PR1 2TQ, UK
Received: 30 January 1996; Revised: 8 May 1996
A B S T R A C T
Nickel-resistant bacteria were isolated from effluent discharged from a sewage treatment outfall
over an 18-month period. One of these strains, Enterobacter cloacae FBA30, was found to harbor
a narrow host range conjugative plasmid, designated pFBA30, which confers nickel resistance on
its host. A 10.2-kb SstI restriction fragment was cloned from pFBA30 and was shown to specify
inducible nickel resistance, both in its original host and in laboratory strains of Escherichia coli.
This DNA fragment, and a 1.75-kb SmaI fragment derived from it, were used as probes to examine
other bacterial strains isolated during the study. Homologous nickel resistance genes were detected
in enteric bacteria emerging directly from the treatment plant, but not in strains isolated from
sediments downstream of the outfall, or in strains harboring well-characterized determinants such
as ncc/nre, cnr, or Klebsiella oxytoca type. Thus, this element constitutes a new nickel resistance
determinant and was named nrf.
Nickel-resistant strains were sorted into two groups, based on the sampling date and the size of
restriction fragments homologous to probes developed from pFBA30. Group A isolates all carried
a homologous 6.5-kb PvuII restriction fragment and expressed nickel resistance constitutively.
Group B strains, isolated independently from the same outfall, were different from group A in that
the nickel resistance was associated with a 4.7-kb PvuII fragment, and group B strains displayed a
slower growth rate on nickel salts media. Each group contained a range of enteric bacterial species,
including K. oxytoca, Citrobacter freundii, and Enterobacter spp. Localized genetic exchange probably
occurs within wastewater treatment effluents, and, once effluents are discharged into the environ-
ment, enteric bacteria carrying pFBA30-like determinants do not persist downstream.
Correspondence to: R.W Pickup. E-mail: [email protected].
79050E$P18 03-31-97 14:35:19

Nickel Resistance in Sewage-Associated Bacteria 231
treatment increases the proportion of bacteria that carryIntroductioneither single or multiple antibiotic resistances; between 0.1
and 1% of total coliforms in fecal material were resistant,Plasmid-encoded nickel resistance in bacteria has been char-and this reached frequencies of 10% in urban wastewater,acterized most extensively in Alcaligenes eutrophus CH3450% in river water, and 80% in potable water [16]. Bacteria[21]. This strain harbors the conjugative plasmid pMOL28that carry resistance to multiple antibiotics have also beenwhich confers resistance to a range of metals including nickelfound in both drinking water and wastewater [2, 3]. Further-and cobalt. Resistance is mediated by an inducible energy-more, chlorination of water has been shown to select fordependent specific efflux system [11, 25, 34, 35, 36, 41].multiple antibiotic resistance [23]. Resistance to heavy met-Extensive molecular characterization of the nickel efflux sys-als, linked to that of antibiotics, has been found not only intem was achieved by cloning the resistance gene (cnr) frombacteria isolated from sewage effluent [41] but also in bacteriapMOL28 [15, 37]. Most related studies have used nickel-from hospitals [19], metal-contaminated estuaries [1], andresistant strains isolated from anthropogenically pollutedsoils [18]. Bacteria affected by either water or wastewatersites, particularly those associated with the metal-reclamationtreatment are eventually released into aquatic environments.industries [20, 39], and those where nickel percolates natu-Almost all investigations of antibiotic resistance in the aquaticrally through an ecosystem [39]. Resistance levels conferredhabitat have concentrated on its incidence in bacteria of fecal
by these plasmid- or chromosomally located genes rangeorigin [5, 12–14]. However, a positive correlation was noted
from 1 to 40 mM in Tris-gluconate media [31, 33].between the occurrence of heavy metal and multiple antibiotic
There is some diversity to be found among nickel resis-resistance in a range of bacteria isolated from water distribu-
tance determinants. Some nickel resistance genes show a hightion and treatment systems [7]. The aim of this study was to
degree of homology with cnr of pMOL28 and ncc of A.develop nucleic acid probes to examine the nickel-resistant
xylosoxidans 31A [32, 33], while others have been found tobacterial community being dispersed from a wastewater treat-
carry a second resistant determinant (nre) that has only lim-ment works, and to assess whether metal resistance transfers
ited homology with pMOL28. The nre determinant expressesto the bacterial community downstream of the outfall pipe.
in E. coli and is thought to be widespread among nickel-
resistant bacteria [32]. Comparative analysis of a large num-
ber of nickel-resistant isolates enabled Stoppel and SchlegelMaterials and Methods[39] to distinguish four groups of nickel resistance. TheseBacterial Strains and Plasmidscomprised (1) those showing homology with cnr of pMOL28
[17] and ncc from pTOM (33); (2) a combination of cnr, The strains used in this study are described in Table 1. All nickel-
resistant strains were maintained on normal-strength nutrient agarncc, and nre [32]; and (3) a determinant found in K. oxytoca,(Oxoid, Basingstoke UK) supplemented with 5.0 mM nickel chlo-showing weak homology with nre [40]. These loci were notride. Escherichia coli HB101 and JM103 containing recombinantubiquitous, however, as indicated by the existence of a fourthplasmids were maintained on nutrient agar supplemented with 50
group of resistant strains exhibiting no sequence homologymg ml21 ampicillin. All other strains were maintained on nutrient
with either cnr/ncc or nre [32]. agar.The occurrence of metal resistant bacteria in anthropo-
genically polluted sites is well documented [10, 38]. Studies Sampling and Isolation of Bacteriaof bacteria emerging from a sewage treatment plant in the
Church Beck is a freshwater stream in the Coniston Fells, Cumbria,English Lake District showed the presence of a range of metal
UK and has been described previously [26, 27]. The sample siteresistances, which included nickel [26] and copper [27]. No was the sewage outfall in the lower section of Church Beck whichselection for resistance due to metal contamination was ap- feeds directly into Coniston (National Grid Reference SD 308971).
Bacteria were isolated from 11 samples of water, as described byparent at this site, since this treatment system received onlyPickup [26], except that filters were placed on nutrient agar supple-domestic inputs [26]. It has been noted previously that resis-mented with 5 mM NiCl2•6H2O (pH 7.6). Sediments were collectedtance to heavy metals occurs at a surprisingly high frequencydownstream of the outfall pipe in glass containers, vigorously shaken
in the absence of sources of direct selection [38]. in the laboratory, and allowed to settle. The supernatant was thenSewage effluent, wastewater, and water treatments have filtered and strains were isolated in the same way as the water
samples. Plates were incubated at 208C until single colonies ap-been shown to have several effects on bacteria. Wastewater
79050E$P18 03-31-97 14:35:19

232 R.W. Pickup et al.
Table 1. Bacterial strains and plasmids
Strain designationused in this study Species name and strain number Plasmid name/(content) Descriptiona Origin
FBA30 Enterobacter cloacae pFBA30/pFBA31 (2) inducible Nir This studyFBA30-1 Escherichia coli HB101 pFBA30 (1) inducible Nir This studyFBA30-2 Enterobacter cloacae NCIB 8151 pFBA30 (1) inducible Nir This studyFBA30-3 Enterobacter aerogenes pFBA30 (1) inducible Nir This studyFBA30-4 Serratia rubidae pFBA30 (1) inducible Nir This studyFBA13 Klebsiella oxytoca (group A) 0 constitutive Nir This studyFBA17B Enterobacter sakazakii (group A) not named (2) constitutive Nir This studyFBA28 Enterobacter cloacae (group A) not named (2) constitutive Nir This studyFBA1.3.21 Citrobacter freundii (group B) not named (1) inducible Nir This studyFBA1.4.2 ndb (group B) not named (1) inducible Nir This studyFBA1.5.43 Enterobacter agglomerans (group B) not named (1) inducible Nir This studyFBA30-10 Escherichia coli JM103 pFBA30-10 (1) inducible Nir This studyFBA30-11 Escherichia coli JM103 pFBA30-11 (1) inducible Nir This studyFBA30-12 Escherichia coli JM103 pFBA30-12 (1) constitutive Nir This studyNCIMBc8151 Enterobacter cloacae (0) Nis NCIMBHB101 Escherichia coli HB101 (0) Smr Nis 6JM103 Escherichia coli JM103 (0) Nis 43PaW340 Pseudomonas putida PaW340 (0) Smr trp2 42M695 Escherichia coli JM109 pSK2::KOHI4-1 Nir K. oxytoca 40M696 Escherichia coli JM109 pSK2::TBA9a Nir nre/ncc 32,33M697 Escherichia coli JM109 pHLI4 Nir cnr 17
a trp, Tryptophan; Nir, nickel resistance; Smr, streptomycin resistanceb nd, Not determinedc NCIMB, National Collection of Industrial and Marine Bacteria, Aberdeen
peared. All colonies were purified and retained. API identification ml21) were use as the selective medium. Single colonies were isolated,
purified, and examined for the presence of plasmids.strips (Biomerieux Ltd., Basingstoke, UK) were used, as directed
by the manufacturer.
DNA ManipulationsBacterial Conjugation
Plasmids from environmental isolates and transconjugants wereExogenous Plasmid Isolation. Exogenous plasmid isolation was
extracted using the method of Wheatcroft and Williams [42]. Totalmodified from the method of Bale et al. [4]. Water/effluent samples
genomic DNA was extracted using the method of Pitcher et al.of 50, 100, and 500 ml were mixed with 1 ml of recipient strain
[29], and recombinant plasmids were prepared by the method ofculture (PaW340 or HB101) and filtered through a 0.22-mm Milli-
Close and Rodriguez [8]. Plasmids used for genetic manipulationpore HC filter under vacuum. Filters were incubated on nutrient
were purified by SDS lysis/CsCl centrifugation [30].agar at 308C overnight. The accumulated growth was resuspended
Restriction endonucleases were obtained from Gibco BRL (Pais-in sterile distilled water, serially diluted, and spread onto nutrient
ley, UK). Agarose gel electrophoresis was carried out by establishedagar plates supplemented with NiCl2 (5 mM) and streptomycin (100
procedures. Reaction conditions for DNA ligation with T4 DNAmg ml21). Plates were examined for single colonies after incubation
ligase were as recommended by the supplier (Gibco BRL). Transfor-at 308C. Appropriate donor (water/effluent sample) and recipient
mation into E. coli JM103 was achieved by the procedure of Cohencontrols were performed simultaneously.
et al. [9]. DNA probes (either from purified DNA or from restriction
fragments excised from low melting temperatures gels, as directedEndogenous Plasmid Transfer. Conjugation experiments were per-
formed using a plate mating method with purified donor strains by the manufacturer) were labeled with [32P]dCTP using a Random
Primed DNA Labelling Kit (Boehringer, Lewes UK). pUC19 geneisolated from the sewage outfall, as described by Pickup [28], using
both PaW340 and HB101 as recipients. To facilitate screening a banks were screened by the method of Close and Rodrigues [9].
DNA hybridization was carried out as described previously [22].large number of isolates, streak matings were used. The recipient
was streaked vertically down the left side of the selection plate.
Donors were then streaked horizontally, in one movement, from Induction Studiesthe left to the right side through the recipient streak. The donor
streak to the left of the recipient served as a control. Nutrient agar Bacteria were cultured overnight in 50 ml normal-strength nutrient
broth and in 50 ml nutrient broth supplemented with 0.5 mMplates supplemented with NiCl2 (5 mM) and streptomycin (100 mg
79050E$P18 03-31-97 14:35:19

Nickel Resistance in Sewage-Associated Bacteria 233
100 isolates, in total, were retained from sediments down-
stream of the outfall during sample trips 2 and 3. The concen-
tration of nickel-resistant isolates in the sediments was not
determined. Of the sediment isolates, 60% were confirmed
as oxidase negative.
Conjugation Experiments
An attempt was made to isolate conjugative nickel-resistant
plasmids from environmental samples by transfer to labora-
tory strains of Pseudomonas putida (PaW340) and E. coli
(HB101). No transconjugants were isolated by this method
when filtered bacteria from sewage sample 1 or either of the
sediment samples were used as donor populations, so purified
strains isolated from sample 1 were examined further by
individual streak matings. Again, no transconjugants were
observed when PaW340 was used as a recipient, but a single
isolate regularly yielded nickel-resistant transconjugants in
matings with HB101. This strain was designated FBA30. API
identification and analysis of plasmid DNA confirmed that
the colonies were true transconjugants. The donor isolate
was identified as Enterobacter cloacae (probability .99%)
using API test strips. Plasmid analysis of the donor strainFig. 1. HindIII digestion of plasmids found in FBA30 and trans-
conjugants. Lanes: 1, l (HindIII); 2, FBA30(pFBA30:pFBA31); 3, (FBA30) revealed two plasmids (designated pFBA30 andE. coli JM103(pFBA30); 4, E. cloacae NCIMB 8151(pFBA30); 5, E. pFBA31; Fig. 1). Comparison of the plasmid content ofaerogenes(pFBA30); 6, Serratia rubidae(pFBA30).
FBA30 and the transconjugants obtained by plate matings
revealed that one plasmid (pFBA30) had transferred with a
frequency of 1.9 3 1023 transconjugants per donor cell, andNiCl2 (previously determined to be a nonlethal concentration), with
was stably maintained (Fig. 1). FBA30-1 was retained asshaking (100 rpm) at 308C. The optical density at 600 nm was thenrepresentative of the transconjugants. Plate matings withmeasured (Cecil Instruments CE292 Digital UV Spectrophotome-
ter), with nutrient broth as a blank. Each culture was diluted in FBA30 and FBA30-1 showed that plasmid pFBA30 was trans-nutrient broth to an optical density of 1.0 at 600 nm. After adjust- missible between enteric bacteria (E. cloacae NCIB8151, E.ment, 1 ml of each culture was inoculated into duplicate flasks aerogenes, Serratia rubidae), but not to nonenteric strains (P.containing 50 ml nutrient both supplemented with 2.5 mM NiCl2. putida PaW340, P. fluorescens). Transconjugants harboringThese were incubated at 308C with shaking (100 rpm). The optical
pFBA30 were isolated and purified from these matings. Thesedensity of each was measured hourly for a total of 8 h after inocu-strains were designated FBA30-2, FBA30-3, and FBA30-4,lation.
respectively (Table 1). Restriction endonuclease digestion
with a range of enzymes revealed pFBA30 to be approxi-Results mately 55 kb in size (data not presented).Isolation of Nickel-Resistant Strains
Cloning of Nickel Resistance Determinant from pFBA30The sewage outfall was sampled three times (samples 1, 2,
and 3) at intervals of 6 months. Bacteria resistant to 5 mM Plasmid DNA from pFBA30 was isolated and digested with
SstI. Resultant DNA fragments were ligated into pUC19,NiCl2 were readily isolated on each occasion at a concentra-
tion of approximately 1,000 cells l21. In the first instance, and recombinant plasmids were transformed into JM103. All
clones were tested for nickel resistance, and a single, nickel-over 100 isolates were retained from sample 1. A total of 100
isolates from samples 2 and 3 were also retained for further resistant colony was obtained. The recombinant plasmid in
this strain was found to carry an inserted 10.2-kb SstI restric-analysis. All were confirmed as oxidase negative. A further
79050E$P18 03-31-97 14:35:19
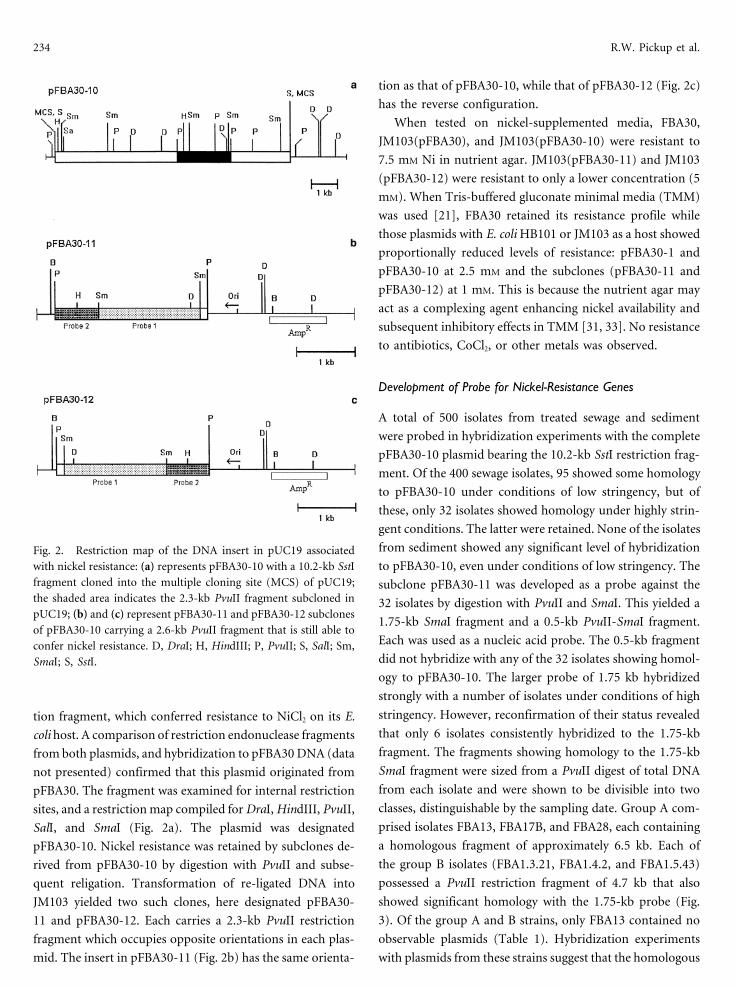
234 R.W. Pickup et al.
tion as that of pFBA30-10, while that of pFBA30-12 (Fig. 2c)
has the reverse configuration.
When tested on nickel-supplemented media, FBA30,
JM103(pFBA30), and JM103(pFBA30-10) were resistant to
7.5 mM Ni in nutrient agar. JM103(pFBA30-11) and JM103
(pFBA30-12) were resistant to only a lower concentration (5
mM). When Tris-buffered gluconate minimal media (TMM)
was used [21], FBA30 retained its resistance profile while
those plasmids with E. coli HB101 or JM103 as a host showed
proportionally reduced levels of resistance: pFBA30-1 and
pFBA30-10 at 2.5 mM and the subclones (pFBA30-11 and
pFBA30-12) at 1 mM. This is because the nutrient agar may
act as a complexing agent enhancing nickel availability and
subsequent inhibitory effects in TMM [31, 33]. No resistance
to antibiotics, CoCl2, or other metals was observed.
Development of Probe for Nickel-Resistance Genes
A total of 500 isolates from treated sewage and sediment
were probed in hybridization experiments with the complete
pFBA30-10 plasmid bearing the 10.2-kb SstI restriction frag-
ment. Of the 400 sewage isolates, 95 showed some homology
to pFBA30-10 under conditions of low stringency, but of
these, only 32 isolates showed homology under highly strin-
gent conditions. The latter were retained. None of the isolates
from sediment showed any significant level of hybridizationFig. 2. Restriction map of the DNA insert in pUC19 associated
with nickel resistance: (a) represents pFBA30-10 with a 10.2-kb SstI to pFBA30-10, even under conditions of low stringency. Thefragment cloned into the multiple cloning site (MCS) of pUC19; subclone pFBA30-11 was developed as a probe against thethe shaded area indicates the 2.3-kb PvuII fragment subcloned in 32 isolates by digestion with PvuII and SmaI. This yielded apUC19; (b) and (c) represent pFBA30-11 and pFBA30-12 subclones
1.75-kb SmaI fragment and a 0.5-kb PvuII-SmaI fragment.of pFBA30-10 carrying a 2.6-kb PvuII fragment that is still able to
Each was used as a nucleic acid probe. The 0.5-kb fragmentconfer nickel resistance. D, DraI; H, HindIII; P, PvuII; S, SalI; Sm,did not hybridize with any of the 32 isolates showing homol-SmaI; S, SstI.
ogy to pFBA30-10. The larger probe of 1.75 kb hybridized
strongly with a number of isolates under conditions of high
stringency. However, reconfirmation of their status revealedtion fragment, which conferred resistance to NiCl2 on its E.that only 6 isolates consistently hybridized to the 1.75-kbcoli host. A comparison of restriction endonuclease fragmentsfragment. The fragments showing homology to the 1.75-kbfrom both plasmids, and hybridization to pFBA30 DNA (dataSmaI fragment were sized from a PvuII digest of total DNAnot presented) confirmed that this plasmid originated fromfrom each isolate and were shown to be divisible into twopFBA30. The fragment was examined for internal restrictionclasses, distinguishable by the sampling date. Group A com-sites, and a restriction map compiled for DraI, HindIII, PvuII,
prised isolates FBA13, FBA17B, and FBA28, each containingSalI, and SmaI (Fig. 2a). The plasmid was designated
a homologous fragment of approximately 6.5 kb. Each ofpFBA30-10. Nickel resistance was retained by subclones de-
the group B isolates (FBA1.3.21, FBA1.4.2, and FBA1.5.43)rived from pFBA30-10 by digestion with PvuII and subse-
possessed a PvuII restriction fragment of 4.7 kb that alsoquent religation. Transformation of re-ligated DNA into
showed significant homology with the 1.75-kb probe (Fig.JM103 yielded two such clones, here designated pFBA30-
3). Of the group A and B strains, only FBA13 contained no11 and pFBA30-12. Each carries a 2.3-kb PvuII restriction
observable plasmids (Table 1). Hybridization experimentsfragment which occupies opposite orientations in each plas-
mid. The insert in pFBA30-11 (Fig. 2b) has the same orienta- with plasmids from these strains suggest that the homologous
79050E$P18 03-31-97 14:35:19
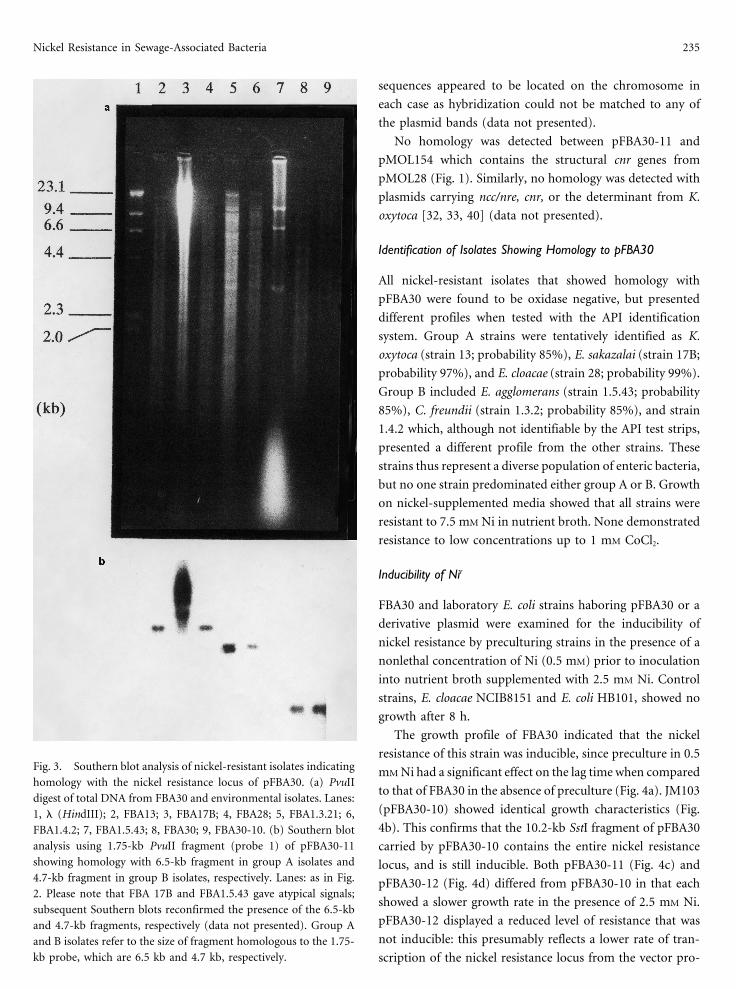
Nickel Resistance in Sewage-Associated Bacteria 235
sequences appeared to be located on the chromosome in
each case as hybridization could not be matched to any of
the plasmid bands (data not presented).
No homology was detected between pFBA30-11 and
pMOL154 which contains the structural cnr genes from
pMOL28 (Fig. 1). Similarly, no homology was detected with
plasmids carrying ncc/nre, cnr, or the determinant from K.
oxytoca [32, 33, 40] (data not presented).
Identification of Isolates Showing Homology to pFBA30
All nickel-resistant isolates that showed homology with
pFBA30 were found to be oxidase negative, but presented
different profiles when tested with the API identification
system. Group A strains were tentatively identified as K.
oxytoca (strain 13; probability 85%), E. sakazalai (strain 17B;
probability 97%), and E. cloacae (strain 28; probability 99%).
Group B included E. agglomerans (strain 1.5.43; probability
85%), C. freundii (strain 1.3.2; probability 85%), and strain
1.4.2 which, although not identifiable by the API test strips,
presented a different profile from the other strains. These
strains thus represent a diverse population of enteric bacteria,
but no one strain predominated either group A or B. Growth
on nickel-supplemented media showed that all strains were
resistant to 7.5 mM Ni in nutrient broth. None demonstrated
resistance to low concentrations up to 1 mM CoCl2.
Inducibility of Nir
FBA30 and laboratory E. coli strains haboring pFBA30 or a
derivative plasmid were examined for the inducibility of
nickel resistance by preculturing strains in the presence of a
nonlethal concentration of Ni (0.5 mM) prior to inoculation
into nutrient broth supplemented with 2.5 mM Ni. Control
strains, E. cloacae NCIB8151 and E. coli HB101, showed no
growth after 8 h.
The growth profile of FBA30 indicated that the nickel
resistance of this strain was inducible, since preculture in 0.5Fig. 3. Southern blot analysis of nickel-resistant isolates indicating mM Ni had a significant effect on the lag time when comparedhomology with the nickel resistance locus of pFBA30. (a) PvuII
to that of FBA30 in the absence of preculture (Fig. 4a). JM103digest of total DNA from FBA30 and environmental isolates. Lanes:
(pFBA30-10) showed identical growth characteristics (Fig.1, l (HindIII); 2, FBA13; 3, FBA17B; 4, FBA28; 5, FBA1.3.21; 6,4b). This confirms that the 10.2-kb SstI fragment of pFBA30FBA1.4.2; 7, FBA1.5.43; 8, FBA30; 9, FBA30-10. (b) Southern blot
analysis using 1.75-kb PvuII fragment (probe 1) of pFBA30-11 carried by pFBA30-10 contains the entire nickel resistanceshowing homology with 6.5-kb fragment in group A isolates and locus, and is still inducible. Both pFBA30-11 (Fig. 4c) and4.7-kb fragment in group B isolates, respectively. Lanes: as in Fig. pFBA30-12 (Fig. 4d) differed from pFBA30-10 in that each2. Please note that FBA 17B and FBA1.5.43 gave atypical signals;
showed a slower growth rate in the presence of 2.5 mM Ni.subsequent Southern blots reconfirmed the presence of the 6.5-kb
pFBA30-12 displayed a reduced level of resistance that wasand 4.7-kb fragments, respectively (data not presented). Group Anot inducible: this presumably reflects a lower rate of tran-and B isolates refer to the size of fragment homologous to the 1.75-
kb probe, which are 6.5 kb and 4.7 kb, respectively. scription of the nickel resistance locus from the vector pro-
79050E$P18 03-31-97 14:35:19

236 R.W. Pickup et al.
moter. pFBA30-11, with its insert in the same orientation as resistance systems [21, 24, 32, 37], that of pFBA30 does not
provide resistance to metals such as cobalt, zinc, mercury,that of pFBA30-10, displayed a slow growth rate and a re-
duced form of inducibility. chromate, or cadmium. Therefore, the nickel resistance
gene(s) of pFBA30 represents another, unrelated gene familyEnvironmental isolates were also examined for inducibil-
ity of Nir. Group A strains 13, 17B, and 28 demonstrated a and was designated the nrf determinant.
We also report herein that this nickel resistance was induc-common growth profile indicative of a noninducible consti-
ible, both in E. cloacae and in other enteric species. Inducibil-tutive resistance (Fig. 4e,f,g). In contrast, group B strains all
ity was retained by the 10.2-kb SstI restriction fragment ofshowed a clear inducible response (Fig. 4h,i,j). Growth rates
pFBA30-10, but not by the 2.3-kb PvuII fragment derivedfor these strains in the presence of nickel were consistently
from this plasmid. pFBA30-11 and pFBA30-12, each carryingslower than that for FBA30.
the PvuII fragment in opposite orientations, displayed consti-
tutive expression and slower growth in the presence of nickel.
This may stem from the loss of regulatory genes locatedDiscussion outside the fragment, but this has yet to be confirmed by
complementation studies. It is considered unlikely that theTransfer of nickel resistance from environmental samples to effect is due to vector promoters overriding the inducibleHB101 or PaW340 was not detected by the exogenous plas- expression of the cloned nickel resistance gene(s), since con-mid isolation procedure. Endogenous matings, again using struction of these plasmids removes the regulated plac pro-the same recipients, identified only a single strain out of 100 moter of pUC19, and the only other available promoters lieisolates that could transfer nickel resistance by conjugation. in the ori region of the plasmid.This may indicate the inability of these laboratory strains to Use of the 10.2-kb SstI restriction fragment as a hybridiza-act as recipients in matings with recently isolated bacteria tion probe against DNA extracted from environmental iso-under the conditions used here. It is significant that all nickel- lates of the Church Beck site identified 95 strains (24% ofresistant strains were enteric species, and this, in itself, may nickel-resistant isolates) sharing limited homology with thepreclude PaW340 from stably inheriting narrow host range nickel resistance locus of pFBA30. Of these, 32 showed strongplasmids from these sources. However, previous studies have homology with the probe fragment under conditions of highisolated metal resistant strains by these methods (silver resis- stringency. These 32 isolates were further challenged by hy-tant plasmids, Pickup, unpublished data), and the results bridization experiments using a 1.75-kb SmaI fragment frommay instead indicate a very low frequency of plasmid-borne pFBA30-10. Six of these demonstrated extensive homologyresistance in these bacterial populations. with this probe and were retained for further study. The
FBA30 was tentatively identified as E. cloacae and was nickel resistance of these six strains appeared to be locatedfound to contain two plasmids. One of these was designated on the bacterial chromosome of each, rather than on a plas-pFBA30 and is described as a conjugative nickel-resistance mid. The majority of previously reported nickel resistanceplasmid of approximately 55 kb. It is considered to have a genes have been shown to reside on plasmids [39], althoughnarrow host range since it was readily introduced into enteric chromosomal resistance has been reported for Alcaligenesrelatives such as E. coli, E. aerogenes, and S. rubidae, but not denitrificans 4a-2 and K. oxytoca [15, 40].into oxidase-positive strains such as P. putida or P. fluorescens. These six strains were classified into two groups, desig-The plasmid pFBA31 found in FBA30 is smaller than pFBA30 nated A and B, distinguishable by the date on which eachand was not transmissible to other strains; it was not consid- was sampled from the environment. The two groups canered further. be further distinguished by the size of chromosomal PvuII
The nickel-resistance locus was cloned on a 10.2-kb SstI restriction fragments sharing homology with the nickel resis-
restriction fragment of pFBA30 DNA, and localized to an tance gene(s) of pFBA30. Group A and B strains carry 6.5-
internal 2.3-kb PvuII fragment. Preliminary characterization and 4.7-kb PvuII fragments, respectively. which are homolo-
of the nickel resistance determinant carried by pFBA30 indi- gous with the 1.75-kb SmaI probe fragment from pFBA30.
cates that it is not related to any of the previously described All these strains are enteric species associated with sewage
nickel resistance families such as ncc/nre, cnr, or the determi- discharge. It is worth noting that one of these species (K.
nant from K. oxytoca [32, 33, 39, 40], and is apparently oxytoca; strain 13 of group A) has already been associated
with nickel resistance [39, 40]. All three group A strainsconfined to enteric bacteria. Furthermore, unlike other nickel
79050E$P18 03-31-97 14:35:19
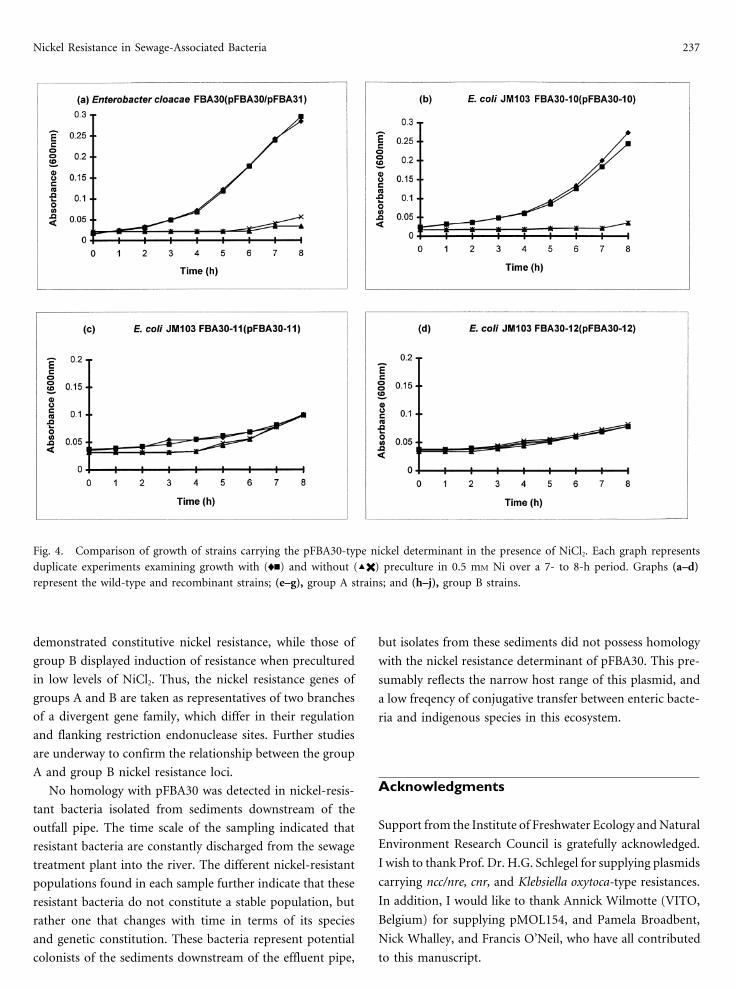
Nickel Resistance in Sewage-Associated Bacteria 237
Fig. 4. Comparison of growth of strains carrying the pFBA30-type nickel determinant in the presence of NiCl2. Each graph represents
duplicate experiments examining growth with (ln) and without (X✖) preculture in 0.5 mM Ni over a 7- to 8-h period. Graphs (a–d)represent the wild-type and recombinant strains; (e–g), group A strains; and (h–j), group B strains.
demonstrated constitutive nickel resistance, while those of but isolates from these sediments did not possess homology
group B displayed induction of resistance when precultured with the nickel resistance determinant of pFBA30. This pre-
in low levels of NiCl2. Thus, the nickel resistance genes of sumably reflects the narrow host range of this plasmid, and
groups A and B are taken as representatives of two branches a low freqency of conjugative transfer between enteric bacte-
of a divergent gene family, which differ in their regulation ria and indigenous species in this ecosystem.and flanking restriction endonuclease sites. Further studies
are underway to confirm the relationship between the group
A and group B nickel resistance loci.AcknowledgmentsNo homology with pFBA30 was detected in nickel-resis-
tant bacteria isolated from sediments downstream of theSupport from the Institute of Freshwater Ecology and Naturaloutfall pipe. The time scale of the sampling indicated thatEnvironment Research Council is gratefully acknowledged.resistant bacteria are constantly discharged from the sewageI wish to thank Prof. Dr. H.G. Schlegel for supplying plasmidstreatment plant into the river. The different nickel-resistantcarrying ncc/nre, cnr, and Klebsiella oxytoca-type resistances.populations found in each sample further indicate that theseIn addition, I would like to thank Annick Wilmotte (VITO,resistant bacteria do not constitute a stable population, but
Belgium) for supplying pMOL154, and Pamela Broadbent,rather one that changes with time in terms of its species
Nick Whalley, and Francis O’Neil, who have all contributedand genetic constitution. These bacteria represent potential
colonists of the sediments downstream of the effluent pipe, to this manuscript.
79050E$P18 03-31-97 14:35:19
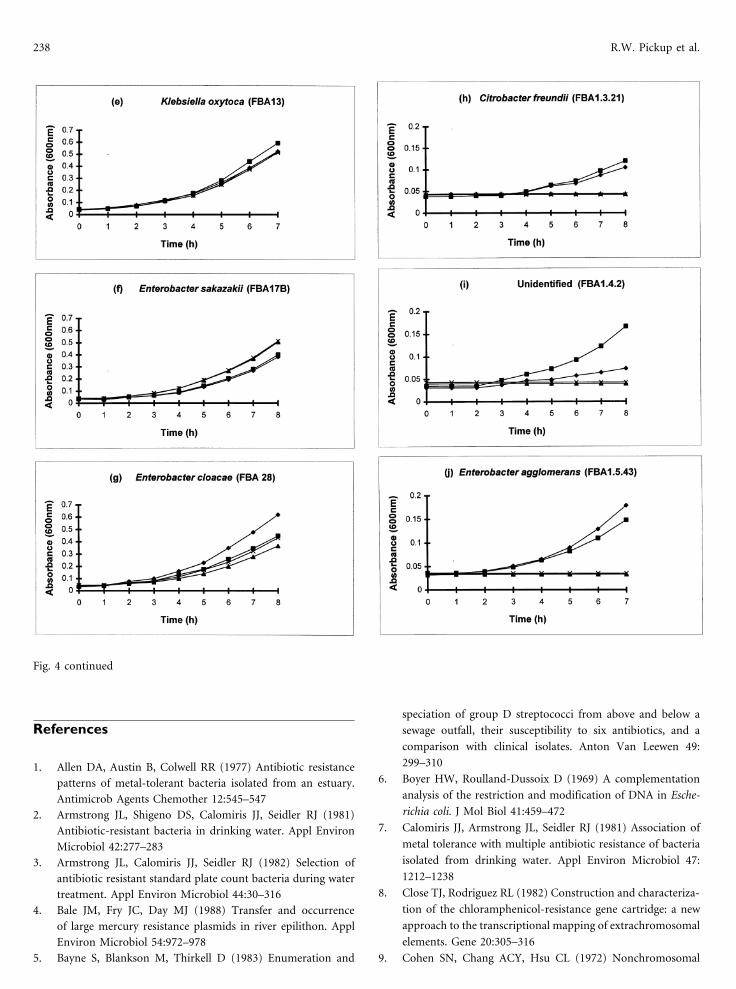
238 R.W. Pickup et al.
Fig. 4 continued
speciation of group D streptococci from above and below a
References sewage outfall, their susceptibility to six antibiotics, and a
comparison with clinical isolates. Anton Van Leewen 49:
299–3101. Allen DA, Austin B, Colwell RR (1977) Antibiotic resistance6. Boyer HW, Roulland-Dussoix D (1969) A complementationpatterns of metal-tolerant bacteria isolated from an estuary.
analysis of the restriction and modification of DNA in Esche-Antimicrob Agents Chemother 12:545–547richia coli. J Mol Biol 41:459–4722. Armstrong JL, Shigeno DS, Calomiris JJ, Seidler RJ (1981)
7. Calomiris JJ, Armstrong JL, Seidler RJ (1981) Association ofAntibiotic-resistant bacteria in drinking water. Appl Environmetal tolerance with multiple antibiotic resistance of bacteriaMicrobiol 42:277–283isolated from drinking water. Appl Environ Microbiol 47:3. Armstrong JL, Calomiris JJ, Seidler RJ (1982) Selection of1212–1238antibiotic resistant standard plate count bacteria during water
8. Close TJ, Rodriguez RL (1982) Construction and characteriza-treatment. Appl Environ Microbiol 44:30–316tion of the chloramphenicol-resistance gene cartridge: a new4. Bale JM, Fry JC, Day MJ (1988) Transfer and occurrenceapproach to the transcriptional mapping of extrachromosomalof large mercury resistance plasmids in river epilithon. Applelements. Gene 20:305–316Environ Microbiol 54:972–978
5. Bayne S, Blankson M, Thirkell D (1983) Enumeration and 9. Cohen SN, Chang ACY, Hsu CL (1972) Nonchromosomal
79050E$P18 03-31-97 14:35:19

Nickel Resistance in Sewage-Associated Bacteria 239
antibiotic resistance in bacteria: genetic transformation of Esch- 25. Nies DH (1992) Resistance to cadmium, cobalt, zinc, and
nickel in microbes. Plasmid 27:17–28erichia coli by R factor DNA. Proc Natl Acad Sci USA 69:
2110–2114 26. Pickup RW (1989a) The discharge of bacteria resistant to heavy
metals from sewage into freshwater. AQUA 38:230–23510. Diels L, Mergeay M (1990) DNA probe-mediated detection of
resistant bacteria from soils highly polluted by heavy metals. 27. Pickup RW (1989b) A common plasmid type found in an
English Lake District stream. Microb Ecol 18:211–230Appl Environ Microbiol 56:1485–1491
11. Eberz G, Eitinger T, Friedrich B (1989) Genetic determinants 28. Pickup RW (1992) Detection of gene transfer in aquatic envi-
ronments. In: Wellington EH, Van Elsas JD (eds) Geneticof a nickel-specific transport system are part of the plasmid-
encoded hydrogenase gene cluster in Alcaligenes eutrophus. J interactions among microorganisms in the natural environ-
ment. Pergamon Press, Oxford, pp 145–164Bacteriol 171:1340–1345
12. Grabow WOK (1977) South African experience on indicator 29. Pitcher DG, Saunders NA, Owen RJ (1989) Rapid extraction
of bacterial genomic DNA with guanidium thiocyanate. Lettbacteria, Pseudomonas aeruginosa, and R1 coliforms in water
quality control. In: Hoadley AW, Dukta BJ (eds), Bacterial Appl Microbiol 8:151–156
30. Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning:indicators health hazards associated with water. ASTM STP
635, American Society for Testing Materials, pp 168–181 a laboratory manual. Cold Spring Harbor Laboratory Press,
Cold Spring Harbor,13. Jones JG, Gardener S, Simon BM, Pickup RW. (1986a) Antibi-
otic resistant bacteria in Windermere and two remote upland 31. Schmidt T, Schlegel HG (1989) Nickel and cobalt resistance of
various bacteria isolated from soil and highly polluted domestictarns in the English Lake District. J Appl Bacteriol 60:443–453
14. Jones JG, Gardener S, Simon BM, Pickup RW (1986b) Factors and industrial wastes. FEMS Microbiol Ecol 62:315–328
32. Schmidt T, Schlegel HG (1994) Combined nickel-cobalt-cad-affecting the measurement of antibiotic resistance in bacteria
isolated from lake water. J Appl Bacteriol 60:454–462 mium resistance encoded by the ncc locus of Alcaligenes xylosox-
idans 31A. J Bacteriol 176:7045–705415. Kaur P, Roß K, Siddiqui RA, Schlegel HG (1990) Nickel resis-
tance of Alcaligenes denitrificans strain 4a-2 is chromosomally 34. Sensfuss C, Schlegel HG (1988) Plasmid pMOL28-encoded
resistance to nickel is due to specific efflux. FEMS Microbiolcoded. Arch Microbiol 154:133–138
16. LeClerc H, Mizon F (1978) Eaux d’alimentation et bacteriies Lett 55:295–298
35. Siddiqui RG, Schlegel HG (1987) Plasmid pMOL28-mediatedresistantes aux antibiotiques incidences sur les normes. Rev
Epidemiol Med Soc Sante Publique 26:137–116 inducible nickel resistance in Alcaligenes eutrophus CH34.
FEMS Microbiol Lett 43:9–1317. Liesegang H, Lemke K, Siddiqui RA, Schlegel HG (1993) Char-
acterization of the inducible nickel and cobalt resistance deter- 36. Siddiqui RA, Schlegel HG, Meyer M (1988) Inducible and
constitutive expression of pMOL28-encoded nickel resistanceminant cnr from pMOL28 of Alcaligenes eutrophus CH34. J
Bacteriol 175:767–778 in Alcaligenes eutrophus N9A. J Bacteriol 170:4188–4193
37. Siddiqui RA, Benthin K, Schlegel HG (1989) Cloning of18. Marques AM, Congregado F, Simon-Pujol DM (1979) Antibi-
otic and heavy metal resistance of Pseudomonas aeruginosa pMOL28-encoded nickel resistance genes and expression of
the genes in Alcaligenes eutrophus and Pseudomonas spp. J.isolated from soils. J Appl Bacteriol 47:347–350
19. McHugh GL, Hopkins CC, Mocllering RC, Swartz MN (1975) Bacteriol 171:5071–5078
38. Silver S, Misra TK (1988) Plasmid-mediated heavy metal resis-Salmonella typhimurium resistant to silver nitrate, chloram-
phenicol, and ampicillin. Lancet ii:235–240 tances. Annu Rev Microbiol 42:717–743
39. Stoppel RD, Schlegel HG (1995) Nickel-resistant bacteria from20. Mergeay M, Houba C, Gerits J (1978) Extrachromosomal in-
heritance controlling resistance to cadmium, cobalt, and zinc anthropogenically nickel-polluted and naturally nickel-perco-
lated ecosystems. Appl Environ Microbiol 61:2276–2285ions: evidence from curing in a Pseudomonas. Arch Int Physiol
Biochim 86:328–334 40. Stoppel RD, Meyer M, Schlegel HG (1995) The nickel resistance
determinant cloned from enterobacterium Klebsiella oxytoca:21. Mergeay M, Nies D, Schlegel HG, Gerits J, Charles P, Gijsegem
Van F (1985) Alcaligenes eutrophus CH34 is a facultative che- conjugational transfer, expression, regulation, and DNA ho-
mologies to various nickel-resistant bacteria. Biometals 8:molithotroph with plasmid-bound resistance to heavy metals.
J Bacteriol 162:328–334 70–79
41. Varma MM, Thomas WA, Prasad C (1976) Resistance to inor-22. Morgan JAW, Rhodes G, Pickup RW (1993) Survival of non-
culturable Aeromonas salmonicida in lake water. Appl Environ ganic salts and antibiotics among sewage borne enterobacteria-
ceae and archromobacteriaceae. J Appl Bacteriol 41:347–349Microbiol 59:874–880
23. Murray GE, Tobin RS, Junkins B, Kushner DJ (1984) Effect of 42. Wheatcroft R, Williams PA (1981) Rapid methods for the
study of both stable and unstable plasmids in Pseudomonas. Jchlorination on antibiotic resistance profiles of sewage related
bacteria. Appl Environ Microbiol 48:73–77 Gen Microbiol 124:433–437
43. Yanisch-Perron C, Vieira J, Messing J (1985) Improved M1324. Nies A, Nies DH, Silver S (1989) Cloning and expression of
plasmid genes encoding resistances to chromate and cobalt in cloning vectors and host strains: nucleotide sequences of the
M13mp18 and pUC19 vectors. Gene 33:103–119Alcaligenes eutrophus. J Bacteriol 117:5065–5070
79050E$P18 03-31-97 14:35:19