a new species of pythium isolated from the burgundy region in france
TRANSCRIPT

A new species of Pythium isolated from the Burgundy regionin France
Bernard Paul *, Delphine Galland, Tej Bhatnagar, Hubert DulieuLaboratoire de Mycologie, U.F.R. Sciences et techniques, 9, avenue Alain Savary, Universiteè de Bourgogne, B.P. 400, 21011 Dijon, France
Received 5 November 1997; revised 12 November 1997; accepted 12 November 1997
Abstract
Pythium nodosum sp. nov. has been isolated from a soil sample taken in the Burgundy region in France. The fungus hasspherical to variously shaped proliferating sporangia, smooth-walled oogonia which are crowded with different antheridialbranches making a complicated knot around the former, and aplerotic oospores. Morphological and reproductive aspects ofPythium nodosum as well as the PCR of the internal transcribed spacer (ITS1) of the ribosomal nuclear DNA coupled withrestriction fragment length polymorphism analysis are described here. The nucleotide sequences of ITS1 encoding 5.8S rRNAis also given. z 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V.
Keywords: Antheridium; Nuclear ribosomal DNA; Oogonium; Pythium ; Restriction fragment length polymorphism
1. Introduction
The genus Pythium with more than 120 describedspecies is distributed throughout the world and oc-cupies a wide range of terrestrial and aquatic habi-tats [1,2]. It is well known for its pathogenic natureon a wide variety of plants. The pathogenic membersof the genus Pythium infect mainly juvenile or suc-culent tissues, thus restricting their parasitic behaviorto seedlings or to the feeder roots or root tips ofolder plants, and to watery fruits or stem tissues[3]. Although these fungi are mainly known for theirparasitic behavior on plants, one species is reportedto be a mammalian parasite [4] and several species
are under study for their production of human diet-ary supplements [5,6].
The members of the genus Pythium are true fungihaving a coenocytic branched mycelium, but phylo-genetic comparisons place them close to algae andhigher plants [7,8]. Unlike most of the eumycetes, themembers of this genus remain diploid throughouttheir life cycles with meiosis occurring in the game-tangia before fertilization [9].
The literature reveals that very little work has beendone on the genus Pythium in France. AlthoughRoze and Cornu [10] described Pythium pythioides(Cystosiphon pythioides) as early as in 1869 fromthis country, later works were mainly on the patho-logical aspects of this genus. In 1907, Butler [11]reported Pythium rostratum from a garden soil inFrance. Paul [12,13] described a new species,Pythium radiosum, in 1992 and a new variety,
0378-1097 / 98 / $19.00 ß 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V.PII S 0 3 7 8 - 1 0 9 7 ( 9 7 ) 0 0 5 2 4 - 7
FEMSLE 7952 9-1-98
* Corresponding author. Tel/Fax: +33 (3) 80-39-61-28.E-mail: [email protected]
FEMS Microbiology Letters 158 (1998) 207^213

Pythium pachycaule var. rami¢catum, in 1993, fromthe north of this country. Eight species of Pythiumwere also isolated and described from the same re-gion [14]. In 1995, Mugnier and Grosjean gave arestriction fragment length polymorphism (RFLP)analysis of some 35 species of Pythium some ofwhich were isolated in France.
The morphological description of di¡erent speciesof Pythium can be complemented by the polymerasechain reaction coupled to restricted fragment lengthpolymorphism analysis (PCR-RFLP). This methodhas become a useful tool in fungal taxonomy[15,16] and is currently used to identify di¡erent spe-cies of Pythium [17^19]. Ampli¢cation of ribosomalgenes is used for the genetic identi¢cation of manyorganisms because they comprise both sequenceshighly conserved during evolution and sequencesthat are highly variable among species and evenwithin species. The ribosomal nuclear DNA consistsof transcribed and non-transcribed regions [17]. TheITS1 (internal transcribed spacer) is a non-conservedregion and has been ampli¢ed with the PCR methodusing primers MC1 and MC3 [20]. Mugnier andGrosjean obtained restriction digestion pro¢les byusing DraI, MboI, AluI, RsaI, HhaI, and TaqI re-striction enzymes. These enzymes gave satisfactoryresults for 35 species of Pythium.
A new species, Pythium nodosum, has been isolatedfrom a soil sample taken in Longvic in the Burgundyregion of France. The fungus is characterized by itssmooth-walled proliferating sporangia, its antheridiacrowding and making a knot around the oogonia,and its antheridial cell being attached in a campanu-late manner to the oogonia. In this paper the RFLPpro¢le of the PCR-ampli¢ed ITS1 region of Pythiumnodosum ribosomal nuclear DNA is presented to-gether with its morphological details.
2. Materials and methods
2.1. Fungus
The fungus was isolated from a soil sample takenin Longvic in the Burgundy region of France bymeans of the usual baiting techniques using boiledhemp seed halves [2,21^23]. A bit of the fungal my-celium was inoculated upon solid media, i.e. potato
carrot agar (PCA) and corn meal agar (CMA). Allthe cultures were incubated at 25³C. The fungus wasidenti¢ed by the help of keys provided by Middleton[24], Van der Plaats-Niterink [2], Dick [1] andthrough PCR-RFLP [17,19].
2.2. Culture conditions and DNA isolation
The fungus was grown in a liquid medium (10 g ofpeptone, 20 g of glucose, 8 g of yeast extract, 5 g ofNaCl per liter of distilled water) in 30-ml £asks con-taining 10 ml of medium. After a 5-day incubationon a rotary shaker at 25³C, the mycelium waswashed on TE bu¡er (10 mM Tris-HCl pH 8,1 mM EDTA). The mycelium was kept at 320³Cfor 24 h and DNA was isolated from this using themethod described by Chen et al. [18].
2.3. Polymerase chain reaction
Primers MC1 and MC3 [20] were synthesized byOligo Express (Paris). MC1 (AAGTTAAAGTCG-TAACAAGG) is at the 3P end of the 17S rDNAgene and MC3 (CATCCACTGCTGAAAGTTG) isat the 5P end of the 5.8S rDNA gene (Fig. 1).
The primer concentrations were estimated spectro-photometrically at 260 nm. PCR reactions were per-formed in 50-Wl volumes containing 100 pmol ofeach of the primers; 200 WM of each of the fourdNTPs; 1.5 U of Taq polymerase (Pharmacia Bio-tech) and 300 ng DNA template in a PCR bu¡er (50mM KCl, 1.5 mM MgCl2, 10 mM Tris-HCl). Thetemperature cycling parameters were: denaturationat 95³C for 3 min for the ¢rst cycle and 1 min forsubsequent cycles, primer annealing for 1 min at55³C and primer extension at 72³C for 1 min witha total of 35 cycles and a ¢nal extension at 72³C for3 min [25]. The temperature cycles were controlledusing a DNA thermal cycler (Progene of TechneCambridge). A negative control with all reaction re-agents except DNA templates was included in each
FEMSLE 7952 9-1-98
Fig. 1. MC1 and MC3 primer positions on nuclear ribosomicDNA of Pythium.
B. Paul et al. / FEMS Microbiology Letters 158 (1998) 207^213208

set of reactions. The ampli¢cation was checked byrunning 4 Wl of the reaction solution through a 1.6%agarose gel prepared with Tris-acetate-EDTA bu¡er(40 mM Tris-HCl pH 8, 20 mM acetate and 1 mMEDTA), staining with ethidium bromide and visual-ization and photography under UV light.
2.4. Restriction fragment length polymorphism
The ampli¢ed DNA was used for restriction en-zyme analysis. Six restriction enzymes (DraI, MboI,AluI, RsaI, HhaI, TaqI) (Gibco BRL, Life Technol-ogies) were used according to the manufacturer's in-structions. The enzyme-digested DNA was runthrough a 1.6% agarose gel, stained with ethidiumbromide, visualized and photographed under UVlight.
2.5. DNA sequence
The DNA sequence was graciously determined byDr. Denisot of the Institut de biologie structurale etmicrobiologie, Marseille, France.
3. Results
3.1. Morphological description: Pythium nodosum sp.nov.
Mycelium bene rami¢catum, sine loculis, hyphaprincipalis 6^7 Wm diam. Sporangia globosa, subglo-bosa vel ovata, intercalaria, interdum catenaria, 10^25Wm diam. Oogonia terminalia vel intercalaria, globosa,11^27 Wm diam. Antheridia plurima circa singulumoogonium formans nodum circum, post fecundationemreliquum cellae antheridialis saepe simile est campa-nae. Oosporae saepissime apleuroticae, globulus, 10^21 Wm diam. Incrementum radiale quotidianum 17 mmad 25³C in agaro Solani tuberosi et Dauci carotae(PCA). Secretum ex terra in Longvic (Bourgogne)France. Holotypus in herbario Universitatis Bour-gogne conservatus (F-918.1).
Mycelium hyaline, main hyphae up to 7 Wm wide.On PCA submerged colonies exhibit a vague chrys-anthemum pattern. On CMA the colonies are sub-merged with a radiate pattern. The average dailygrowth of the fungus on PCA and CMA is 17 mm,
however colonies on PCA are denser than those onCMA. The fungus grows well on hemp seed halves inwater producing whitish colonies.
Sporangia and zoospores are readily formed at 20^25³C (Fig. 3). The sporangia are spherical, globose,subglobose, ovoid, pyriform and at times peanut-shaped, measuring 10^25 (av. 19) Wm diam., mostlyintercalary and catenulate, at times terminal (Fig.2a1^a11Fig. 3a); encysted zoospores measure 7^10Wm diam. (Fig. 2b; Fig. 3d). The sporangia produce
FEMSLE 7952 9-1-98
Fig. 2. Pythium nodosum. a1^a3 : Terminal sporangia. a4^a11 : In-tercalary sporangia. b: Encysted zoospores. c: Oogonia sur-rounded by antheridia. d1^d5 : Oogonia with a campanulate an-theridial cell. e1^e6 : Single oospore in oogonia. f1^f4 : Singleoospores. g: Double oospore. Bars=20 Wm.
B. Paul et al. / FEMS Microbiology Letters 158 (1998) 207^213 209

short germ tubes on which sporangial vesicles areformed (Fig. 3b).
Oogonia are spherical, smooth-walled, terminaland intercalary, measuring 11^27 (av. 20.8) Wm
FEMSLE 7952 9-1-98
Fig. 4. Microscopic study of sexual reproduction of Pythium nodosum. a, b: Oogonia surrounded by antheridia forming a knot. c: Oogo-nia with a campanulate antheridial cell. d: Thin-walled oospore. Bars = 20 Wm.
Fig. 3. Microscopic study of asexual reproduction of Pythium nodosum. a: Sporangia. b: Germinating sporangia. c: Sporangia with zoo-spores forming in vesicle. d: Encysted zoospores. Bars (a, b, d) = 20 Wm, bar (c) = 40 Wm.
B. Paul et al. / FEMS Microbiology Letters 158 (1998) 207^213210
diam., and are mostly unisporous but at times havetwo oospores (Fig. 2g).
Antheridia one to many, crowding and forming acomplicated knot around the oogonia (Fig. 2cFig.4a,b). This knot makes it di¤cult to observe the ex-act number, position and type of contact with theoogonia. Soon after fertilization the knot disappearsleaving one antheridial cell attached to the oogoniain a campanulate fashion (Fig. 2d1^d5, e1^e6Fig.4c,d).
Oospores are aplerotic, smooth-walled, measuring10^21 (av. 16.9) Wm diam., and have a moderatelythin wall (Fig. 2f1^f4Fig. 4d).
FEMSLE 7952 9-1-98
Fig. 6. Comparision of the ITS1 sequence of Pythium nodosum with Phytophthora megakarya (S 40356).
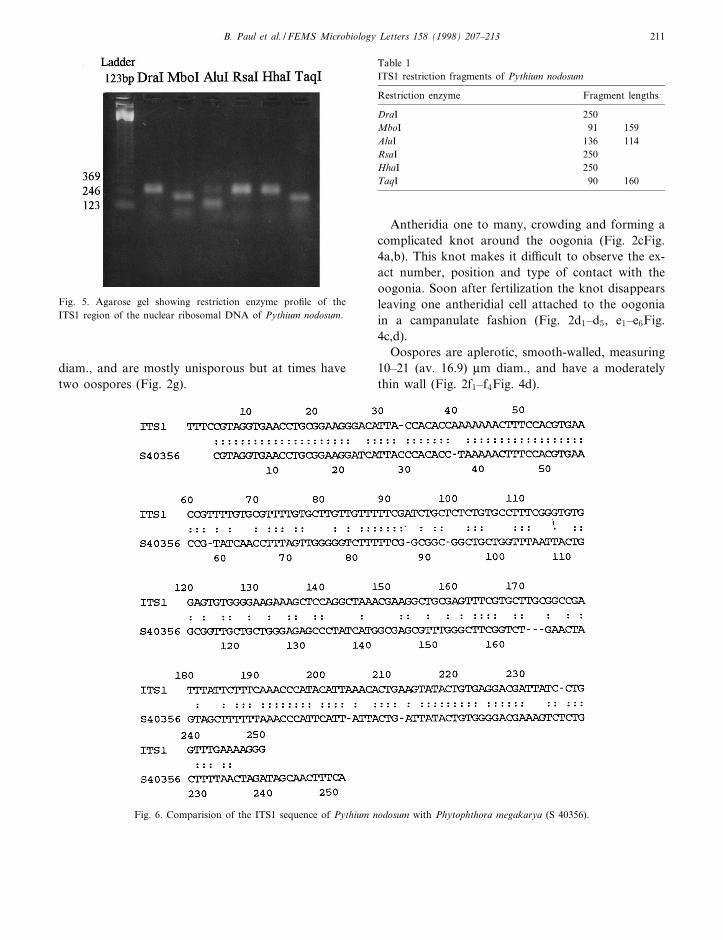
Fig. 5. Agarose gel showing restriction enzyme pro¢le of theITS1 region of the nuclear ribosomal DNA of Pythium nodosum.
Table 1ITS1 restriction fragments of Pythium nodosum
Restriction enzyme Fragment lengths
DraI 250MboI 91 159AluI 136 114RsaI 250HhaI 250TaqI 90 160
B. Paul et al. / FEMS Microbiology Letters 158 (1998) 207^213 211

3.2. Restriction fragment length polymorphismanalysis
The ampli¢cation of the ITS1 region of the ribo-somal DNA of the fungus and its subsequent diges-tion by restriction enzymes DraI, MboI, AluI, RsaI,HhaI, and TaqI gives the pro¢le as indicated in Fig.5.
Table 1 and Fig. 6 show that the ITS1 region ofPythium nodosum is about 250 bp long. This lengthvariant shows a distinct pro¢le from the other spe-cies when digested with restriction enzymes. EnzymesDraI, RsaI and HhaI have no cutting sites, whileMboI, AluI and TaqI have one cutting site each giv-ing two fragments. MboI gives 159-bp and 91-bpfragment lengths. With AluI, two bands appears:one at 136 bp and the other at 114 bp. The lastenzyme TaqI cuts ITS1 into two fragments: of 160and 90 bp.
4. Discussion
Pythium nodosum is characterized by the knotformed around the oogonia by the antheridial ¢la-ments, the campanulate antheridial cell being at-tached to the oogonia, and proliferating sporangiaof di¡erent shapes and sizes. The complicated knotaround the oogonia has been reported in some spe-cies of Pythium : P. sylvaticum, P. heterothallicum, P.scleroteichum, P. capillosum. However, Pythium no-dosum is quite di¡erent from the former two hetero-thallic species as it reproduces readily in single cul-tures. This species di¡ers from all four species as itproduces proliferating globose sporangia and zoo-spores abundantly which is not the case with theformer three fungi, and sporangia of Pythium capil-
losum are of the ¢lamentous, non-in£ated type.Pythium nodosum is closely related to Pythium vexansand Pythium perplexum as these species have thecampanulate antheridial cell attached to their oogo-nia. However, these species do not produce compli-cated antheridia like those of Pythium nodosum. In1986, Paul reported a new species, Pythium toru-loides, from Algeria. This species closely resemblesPythium nodosum as it has the antheridial knotaround its oogonia, its hyphal bodies are very similarto the sporangia of the latter, and it has the campa-nulate antheridial cell attached to the oogonium.Unfortunately the Algerian isolate was not depositedin a culture collection and subsequently was lost dur-ing the transfer of the senior author's fungal collec-tion from Algeria to France. The only di¡erence be-tween the two isolates is that Pythium nodosumproduces zoospores while Pythium toruloides doesnot. Since the latter is lost, its comparison with theformer is based only on its description in the liter-ature. The isolate F-918.1 of Pythium nodosum isbeing deposited at `Centraalbureau voor Schimmel-cultures' in Baarn (Netherlands) and at the `Interna-tional Mycological Institute' in Egham (UK).
The PCR-RFLP analysis indicates that Pythiumnodosum belongs to the genus Pythium as the ITS1region of the fungus is about 250 bp long and thusfalls within the prescribed length of 151^365 bp [19].The ITS1 pro¢le of this fungus is di¡erent from allothers recently described by Mugnier and Grosjean.The ITS1 region of Pythium nodosum is entirely dif-ferent from that of Pythium vexans. It is longer (311bp) than the ITS1 of P. nodosum (250 bp); theirrestriction enzyme sites can be compared in Table 2.
FastA sequence analysis shows a high percentageof identity with the 18S rRNA-5.8S rRNA ITS1. Ahigh degree of resemblance was shown in a 243-nu-
FEMSLE 7952 9-1-98
Table 2Comparison of ITS1 restriction fragment lengths of Pythium nodosum with Pythium vexans
Species ITS1 (bp) Principal restriction enzyme fragment lengths (bp)
DraI MboI AluI RsaI HhaI TaqI
Pythium nodosum 250 250 91 136 250 250 90159 114 160
Pythium vexans 311 226 44 145 9885 267 311 311 166 66
147
B. Paul et al. / FEMS Microbiology Letters 158 (1998) 207^213212

cleotide overlap of Phytophthora megakarya(S40356) (Fig. 6). As morphologically the genus Phy-tophthora is very close to the genus Pythium, thesequence analysis further strengthens the closenessof these two genera.
Acknowledgments
The authors would like to thank Dr. F. Denisot ofthe Institut de biologie structurale et microbiologie,Marseille, France for graciously doing the ITS1 se-quences of Pythium nodosum and Miss D. Bazireaufor help in preparing the Latin diagnosis.
References
[1] Dick, M.W. (1990) Keys to Pythium. University of ReadingPress, Reading.
[2] Van der Plaats-Niterink, A.J. (1981) Monograph of the genusPythium. Studies in Mycology, Vol. 21, pp. 1^242. Centraal-bureau voor Schimmelcultures, Baarn.
[3] Hendrix, F.F. and Campbell, W. (1973) Pythiums as plantpathogens. Annu. Rev. Phytopathol. 11, 77^98.
[4] DeCock, A.W.A.M., Mendoza, L., Padhye, A.A., Ajello, L.and Kaufman, L. (1987) Pythium insidiosum sp. nov., the etio-logic agent of pythiosis. J. Clin. Microbiol. 25, 344^349.
[5] Gandhi, S.R. and Weete, J.D. (1991) Production of polyun-saturated fatty acids arachidonic acid and eicosapentaenoicacid by the fungus Pythium ultimum. J. Gen. Microbiol. 137,1825^1830.
[6] Stinson, E.E., Kwoczak, R. and Kurantz, M.J. (1991) E¡ectof cultural conditions on production of eicosapentaenoic acidby Pythium irregulare. J. Ind. Microbiol. 8, 171^178.
[7] Baldauf, S.L. and Palmer, J.D. (1993) Animals and fungi areeach other's closest relatives: congruent evidence from multi-ple proteins. Proc. Natl. Acad. Sci. USA 90, 11558^11562.
[8] Knoll, H.A. (1992) The early evolution of eucaryotes : a geo-logical perspective. Science 256, 622^627.
[9] Martin, F. (1995) Meiotic instability of Pythium sylvaticum asdemonstrated by inheritance of the nuclear markers and kar-yotype analysis. Genetics 139, 1233^1246.
[10] Roze, E. and Cornu, M. (1869) Cystosiphon pythioides Rozeand Cornu. Ann. Sci. Nat. Bot. 11, 72^91.
[11] Butler, E.J. (1907) An account of the genus Pythium and someChytridiaceae. Mem. Dept. Agr. India Bot. 1, 1^160.
[12] Paul, B. (1992) Pythium radiosum, a new species with orna-mented oogonia from France. Mycol. Helv. 5, 1^8.
[13] Paul, B. (1993) A new variety of Pythium isolated from culti-vated soil in France. Mycol. Helv. 5, 149^156.
[14] Paul, B. (1994) Some species of Pythium isolated from culti-vated soils in northern France. Cryptogamie Mycol. 15, 263^271.
[15] Lee, S.B. and Taylor, J.W. (1991) Molecular phylogeny ofPhytophthora species. Phytophthora Newslett.
[16] White, T.J., Bruns, T., Lee, S.B. and Taylor, J.W. (1990)Ampli¢cation and direct sequencing of fungal ribosomalRNA genes for phylogenetics. In: PCR Protocols: A Guideto Methods and Application (Innis M.A., Ed.), pp. 315^322.
[17] Chen, W. (1992) Restriction fragment length polymorphismsin enzymatically ampli¢ed ribosomal DNAs of 3 heterothallicPythium species. Phytopathology 82, 1467^1472.
[18] Chen, W., Schneider, R.W. and Hoy, J.W. (1992) Taxonomicand phylogenetic analyses of ten Pythium species using iso-zyme polymorphisms. Phytopathology 82, 1234^1244.
[19] Mugnier, J. and Grosjean, M.C. (1995) PCR Catalogue inPlant Pathology: Pythium. Rhoêne-Poulenc Agro, Lyon.
[20] Grosjean, M.C. (1992) Classi¢cation et identi¢cation des es-peéces du genre Pythium, champignons phytopathogeénes dusol, par l'analyse de l'espace interne transcrit de l'operon ri-bosomique. Ph.D. Thesis, University of Lyon.
[21] Paul, B. (1986) An aquatic species Pythium torulo|ëdes sp. nov.from Algeria. Hydrobiologia 140, 233^236.
[22] Paul, B. (1987) A new species of Pythium with ¢lamentoussporangia from Algeria. Trans. Br. Mycol. Soc. 89, 195^198.
[23] Paul, B. (1987) A new species of Pythium with ornamentedoogonia from Algeria. Mycologia 79, 797^802.
[24] Middleton, J.T. (1943) The taxonomy, host range and geo-graphic distribution of the genus Pythium. Mem. TorreyBot. Club 20, 1^171.
[25] Chen, W., Hoy, J.W. and Schneider, R.W. (1992) Species-spe-ci¢c polymorphisms in transcribed ribosomal DNA of ¢vePythium species. Exp. Mycol. 16, 22^34.
FEMSLE 7952 9-1-98
B. Paul et al. / FEMS Microbiology Letters 158 (1998) 207^213 213