a new species of mantidactylus (anura: mantellidae) from
TRANSCRIPT

372 A. CHANNING AND D. G. BROADLEY
Kafue National Park, 1525Dd, collected by D. Day 17March 1969). The Mongu specimens were taken atnight while the males were calling. The specimensfrom Ndanda were taken in pitfall traps 10 days later,on the eastern edge of the floodplain. The NdauSchool specimens come from pitfalls set in patches ofwoodland or scrub on the western edge of the flood-plain at the end of the rainy season.
Hemisus marmoratus.—Thirteen specimens NMZB13803 (Stepping Stones Farm, Kalomo, Zambia,1626Dc); NMZB 3511 (Sesheke, Zambia, 1724Ad);NMZB-UM 18061 (Ngoma, Kafue National Park,
Zambia, 1525Dd); NMZB-UM 30136, NMZB-UM30172–3 (Chunga, Kafue National Park, Zambia,1526Aa); NMZB-UM 31348 (Mfuwe, Luangwa valley,Zambia, 1331Bb); NMZB-UM9528 (Kalikali, Easternprovince, Zambia, 1332Bb); NMZB 4300 (KondoliloFalls, Zambia, 1330Bb); NMZB-UM 25086 (Lilongwe,Malawi, 1333Dd); NMZB-UM 26356 (Xiluvo, Moz-ambique, 1934Aa); NMZB-UM 19382 (NabaunamaDam, Mozambique, 1538Bd); NMZB-UM 3920 (Ma-gasso, Mozambique, 2132Db); NMZB-UM 28911(Massangana, Mozambique, 2132Db); NMZB-UM26061 (Muda-Lamego, Mozambique, 1934Ad).
Journal of Herpetology, Vol. 36, No. 3, pp. 372–378, 2002Copyright 2002 Society for the Study of Amphibians and Reptiles
A New Species of Mantidactylus (Anura: Mantellidae) from Andasibein Eastern Madagascar
FRANK GLAW1 AND MIGUEL VENCES2,3
1Zoologische Staatssammlung, Munchhausenstr. 21, D-81247 Munchen, Germany2Museum national d’Histoire naturelle, Labotoire des Reptiles et Amphibiens, 25 rue Cuvier, 75005 Paris, France
ABSTRACT.—A new scansorial frog species of Mantidactylus (subgenus Phylacomantis) is described fromAndasibe, a midaltitude locality in central, eastern Madagascar. The new species is similar to the sympatricMantidactylus cornutus but differs by smaller size, presence of a chevron-like dorsal ridge, more distinctsupraocular dermal spines, throat coloration, and advertisement call. Its discovery emphasizes the impor-tance of midaltitude rain forests in central, eastern Madagascar as a center of anuran diversity.
Madagascar harbors a high diversity of an-urans, with currently 182 described and numer-ous undescribed species (Glaw and Vences,2000). The genus Mantidactylus, endemic toMadagascar and Mayotte Island, belongs to thefamiliy Mantellidae (Blommers-Schlosser andBlanc, 1991; Vences and Glaw 2001) and is di-vided into 12 subgenera (Dubois, 1992; Glawand Vences, 1994). One of these subgenera, Phy-lacomantis, was erected to accommodate twogroups of scansorial species; the Mantidactyluspseudoasper group (defined by highly special-ized, territorial, and partly carnivorous tad-poles) and the Mantidactylus granulatus group.Species of Phylacomantis are similar to the direct-developing species of the Mantidactylus aspergroup in the subgenus Gephyromantis in externalmorphology but are distinguished from all Ge-phyromantis by behavior of calling along brooks(vs. calling far from water bodies). Including themost recent descriptions (Glaw and Vences,
3 Corresponding Author. Present address: Institutefor Biodiversity and Ecosystem Dynamics, Maurit-skade 61, 1090 GT Amsterdam, The Netherlands.E-mail: [email protected]
2001) eight Phylacomantis species have been de-scribed. In this paper, we describe a new speciesof this subgenus, which differs both morpholog-ically and bioacoustically from all other knownMantidactylus species.
MATERIALS AND METHODS
Frogs were primarily collected at dusk andnight by localization of calling males. Voucherspecimens were fixed in 96% ethanol, subse-quently stored in 70% ethanol, and deposited inthe herpetological collections of the Zoologisch-es Forschungsinstitut und Museum A. Koenig atBonn (ZFMK) and the Zoologische Staatssa-mmlung at Munchen (ZSM). Additionally, weexamined specimens in the Museo Regionale diScienze Naturali, Torino (MRSN) bearing pro-visional field numbers of Franco Andreone(FAZC). Vocalizations were recorded with a por-table Tensai tape recorder (model RCR-3222)and an external microphone (Vivanco EM 238).Tapes (TDK SA 90) were analyzed with the ME-DAV sound analyzing system Spektro 3.2. Thefollowing morphological measurements weretaken to the nearest 0.1 millimeter with a cali-

373NEW MANTIDACTYLUS FROM MADAGASCAR
FIG. 1. Holotype of Mantidactylus moseri in life, dor-solateral view.
FIG. 2. Holotype of Mantidactylus moseri in life,ventral view.
per: SVL (snout–vent length), HW (head width),HL (head length), ED (horizontal eye diameter),END (eye–nostril distance), NSD (nostril–snouttip distance), NND (nostril–nostril distance),TD (tympanum diameter), HAL (hand length),FORL (forelimb length), HIL (hind-limb length),FOL (foot length), FOTL (foot length includingtarsus); IMTL, IMTH (length and height of theinner metatarsal tubercle); FGL, FGW (lengthand width of femoral gland); TL1 (length of firsttoe).
Webbing formula follows Savage and Heyer(1967) as modified by Myers and Duellman(1982) and Savage and Heyer (1997). To facilitatecomparisons with other species of Mantidactylus,we also give the formula used by Blommers-Schlosser (1979) and most subsequent authorswho published accounts on Madagascan an-urans.
Mantidactylus moseri sp. n.Figures 1–2
Holotype.—ZSM 935/2000 (originally ZFMK60024), adult male, collected by F. Glaw and N.Rabibisoa on 18 December 1994 at Andasibe(18�55�3�S, 48�25�22�E; approximately 850 mabove sea level).
Paratypes.—ZFMK 60025–60026, two adultmales, same locality and collecting dates as ho-lotype.
Additional Specimens.—ZFMK 59896, adultmale, collected by F. Glaw and O. Ramilison on4 March 1995 at the Marojejy massif, Campsite1 (14�26�S, 49�46�E; approximately 300 m abovesea level); MRSN-FAZC 7040, 7010, 7011, 7370,7009, 7349 and 6786, six adult males and oneadult female, collected by F. Andreone, G. Apreaand J. E. Randrianirina in 1997 at Amboloko-patrika (14�32�S, 49�26�E); MRSN uncataloged,one adult male, collected by F. Andreone at Am-banizana. These specimens from northeasternMadagascar are larger and have shorter legs
than those from the type locality. Since theirconspecifity with the types is not confirmed bybioacoustic or genetic data, they are not includ-ed in the paratype series.
Diagnosis.—Assigned to the genus Mantidac-tylus based on the lack of nuptial pads and pres-ence of femoral glands in males. Distinguishedfrom all known species of this genus by com-bination of the following characters: (1) maleSVL 27–39 mm; (2) horizontal tympanum di-ameter about 50% of eye diameter; (3) lateralmetatarsalia largely separated; (4) feet distinctlybut not fully webbed; (5) dorsal surface withlarge granules and dermal spines, including oneconspicuous large spine above each eye and achevron-like ridge on the dorsum; (6) maleswith a grayish, largely distensible and slightlybilobed subgular vocal sac; (7) distinct, well de-fined femoral glands without external mediandepression; and (8) temporal and spectral pa-rameters of the advertisement call.
Description of the Holotype.—For measurementssee Table 1. Body relatively slender; head longerthan wide; body of same width as head in itsanterior two-thirds; snout pointed in dorsalview, truncate in lateral view; nostrils directedlaterally, slightly protuberant, nearer to tip ofsnout than to eye; canthus rostralis distinct,curved; loreal region concave; tympanum dis-tinct, vertically elliptical, 46% of eye diameter;supratympanic fold distinct, straight; tongueovoid, distinctly bifid posteriorly; vomerineteeth distinct, in two rounded aggregations, po-sitioned posterolateral to choanae; choanaelarge, rounded. Arms slender, subarticular tu-bercles single; inner and two outer metacarpaltubercles distinct; fingers without webbing;comparative finger length 1�2�4�3, secondfinger distinctly shorter than fourth finger; fin-

374F.G
LA
WA
ND
M.V
EN
CE
S
TABLE 1. Measurmeents (in millimeters) of adult specimens of Mantidactylus moseri. See Materials and Methods section for abbreviations of characters. M, male; F,female; HT, holotype; PT, paratype. RHL (relative hind-limb length) gives the position reached by the tibiotarsal articulation when the hind limb is adpressed alongthe body: 1, anterior eye corner; 2, between eye and nostril; 3, nostril; 4, between nostril and snout tip; 5, snout tip; 6, beyond snout tip; 7, widely beyond snout tip.
Cataloguenumber Sex Status SVL HW HL TD ED END NSD NND HAI FORI HII FOTL FOL IMTL IMTW FGL FGW RHL
AndasibeZSM 935/2000ZFMK 60025ZFMK 60026
MMM
HTPTPT
28.630.126.9
9.59.99.5
11.411.910.7
1.71.91.8
3.73.53.5
2.93.32.9
1.61.61.7
2.52.42.4
20.821.419.0
9.510.39.0
54.057.453.3
23.925.222.5
16.316.714.5
1.71.51.4
1.11.10.8
4.14.54.6
1.71.82.0
667
MarojejyZFMK 59896 M — 31.5 11.3 12.6 1.8 4.1 3.3 1.4 2.5 22.4 10.7 60.1 26.3 18.4 1.4 0.9 3.5 1.2 7
AmbanizanaMRSN-FAZCuncat. M — 39.1 13.6 14.8 2.8 5.1 4.4 2.1 3.1 11.0 23.3 59.3 26.0 17.6 1.9 1.4 4.6 1.8 2
AmbolokopatrikaMRSN-FAZC 7040MRSN-FAZC 7010MRSN-FAZC 7011MRSN-FAZC 7370MRSN-FAZC 7009MRSN-FAZC 7349MRSN-FAZC 6786
MMMMMMF
———————
35.134.934.330.935.530.736.7
11.412.211.010.511.810.211.4
13.313.512.812.113.711.313.9
2.02.42.01.92.12.02.1
4.44.64.24.44.53.94.1
3.73.73.63.04.03.23.9
2.11.71.91.92.01.71.9
2.82.82.82.82.82.72.9
10.010.110.09.2
10.19.9
10.9
21.321.821.419.221.320.023.5
57.159.359.651.858.852.565.9
24.525.626.022.726.224.228.6
16.617.017.214.917.116.318.6
1.51.71.71.61.51.81.2
1.11.21.21.00.90.90.6
4.74.35.04.65.03.9—
2.41.51.92.11.92.0—
3453516
375NEW MANTIDACTYLUS FROM MADAGASCAR
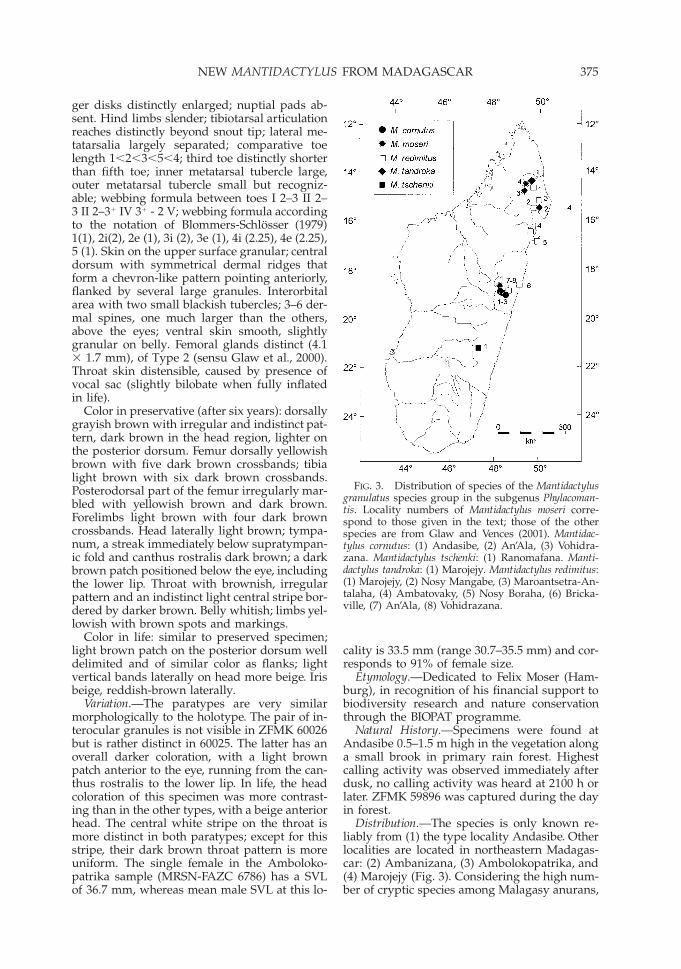
FIG. 3. Distribution of species of the Mantidactylusgranulatus species group in the subgenus Phylacoman-tis. Locality numbers of Mantidactylus moseri corre-spond to those given in the text; those of the otherspecies are from Glaw and Vences (2001). Mantidac-tylus cornutus: (1) Andasibe, (2) An’Ala, (3) Vohidra-zana. Mantidactylus tschenki: (1) Ranomafana. Manti-dactylus tandroka: (1) Marojejy. Mantidactylus redimitus:(1) Marojejy, (2) Nosy Mangabe, (3) Maroantsetra-An-talaha, (4) Ambatovaky, (5) Nosy Boraha, (6) Bricka-ville, (7) An’Ala, (8) Vohidrazana.
ger disks distinctly enlarged; nuptial pads ab-sent. Hind limbs slender; tibiotarsal articulationreaches distinctly beyond snout tip; lateral me-tatarsalia largely separated; comparative toelength 1�2�3�5�4; third toe distinctly shorterthan fifth toe; inner metatarsal tubercle large,outer metatarsal tubercle small but recogniz-able; webbing formula between toes I 2–3 II 2–3 II 2–3� IV 3� - 2 V; webbing formula accordingto the notation of Blommers-Schlosser (1979)1(1), 2i(2), 2e (1), 3i (2), 3e (1), 4i (2.25), 4e (2.25),5 (1). Skin on the upper surface granular; centraldorsum with symmetrical dermal ridges thatform a chevron-like pattern pointing anteriorly,flanked by several large granules. Interorbitalarea with two small blackish tubercles; 3–6 der-mal spines, one much larger than the others,above the eyes; ventral skin smooth, slightlygranular on belly. Femoral glands distinct (4.1� 1.7 mm), of Type 2 (sensu Glaw et al., 2000).Throat skin distensible, caused by presence ofvocal sac (slightly bilobate when fully inflatedin life).
Color in preservative (after six years): dorsallygrayish brown with irregular and indistinct pat-tern, dark brown in the head region, lighter onthe posterior dorsum. Femur dorsally yellowishbrown with five dark brown crossbands; tibialight brown with six dark brown crossbands.Posterodorsal part of the femur irregularly mar-bled with yellowish brown and dark brown.Forelimbs light brown with four dark browncrossbands. Head laterally light brown; tympa-num, a streak immediately below supratympan-ic fold and canthus rostralis dark brown; a darkbrown patch positioned below the eye, includingthe lower lip. Throat with brownish, irregularpattern and an indistinct light central stripe bor-dered by darker brown. Belly whitish; limbs yel-lowish with brown spots and markings.
Color in life: similar to preserved specimen;light brown patch on the posterior dorsum welldelimited and of similar color as flanks; lightvertical bands laterally on head more beige. Irisbeige, reddish-brown laterally.
Variation.—The paratypes are very similarmorphologically to the holotype. The pair of in-terocular granules is not visible in ZFMK 60026but is rather distinct in 60025. The latter has anoverall darker coloration, with a light brownpatch anterior to the eye, running from the can-thus rostralis to the lower lip. In life, the headcoloration of this specimen was more contrast-ing than in the other types, with a beige anteriorhead. The central white stripe on the throat ismore distinct in both paratypes; except for thisstripe, their dark brown throat pattern is moreuniform. The single female in the Amboloko-patrika sample (MRSN-FAZC 6786) has a SVLof 36.7 mm, whereas mean male SVL at this lo-
cality is 33.5 mm (range 30.7–35.5 mm) and cor-responds to 91% of female size.
Etymology.—Dedicated to Felix Moser (Ham-burg), in recognition of his financial support tobiodiversity research and nature conservationthrough the BIOPAT programme.
Natural History.—Specimens were found atAndasibe 0.5–1.5 m high in the vegetation alonga small brook in primary rain forest. Highestcalling activity was observed immediately afterdusk, no calling activity was heard at 2100 h orlater. ZFMK 59896 was captured during the dayin forest.
Distribution.—The species is only known re-liably from (1) the type locality Andasibe. Otherlocalities are located in northeastern Madagas-car: (2) Ambanizana, (3) Ambolokopatrika, and(4) Marojejy (Fig. 3). Considering the high num-ber of cryptic species among Malagasy anurans,

376 F. GLAW AND M. VENCES
FIG. 4. Sonagram (upper) and oscillogram (lower)of two notes from an advertisement call of Mantidac-tylus moseri, recorded on 18 December 1994 at the typelocality Andasibe (20�C air temperature).
assignment of these specimens to M. moseri re-mains tentative.
Calls.—Recorded at Andasibe on 18 Decem-ber 1994, 1920 h at 20�C air temperature. Thespecimens emitted regular series of unharmon-ious notes (Fig. 4). Each note corresponded toone expiration. Temporal parameters of the callsof one specimen (range with mean SD; andnumber of analyzed units in parentheses): noteduration 74–88 ms (83 3 ms; N � 20), dura-tion of intervals between notes 571–2492 ms(1104 505 ms; N � 19). Each note consistedof 23–25 pulses. Pulse rate was 315–333 (323 9; N � 4) pulses per second. Frequency was2700–6200 Hz, dominant frequency 4000–4650Hz.
Comparisons.—Mantidactylus moseri is assignedto the subgenus Phylacomantis based on vocalsac shape, behavior of calling along brooks, andgeneral phenetic resemblance to other species ofthe subgenus. Mantidactylus moseri is distin-guished from other Phylacomantis species as fol-lows: from M. pseudoasper and Mantidactylus cor-vus by the presence of enlarged dermal spineson eyes (vs. absence of such spines), vocal sacmorphology in males (single subgular or slight-ly bilobed and grayish vs. paired subgular withblackish folds along the lower jaw), and adver-tisement calls; from Mantidactylus granulatus andMantidactylus leucomaculatus by smaller SVL(adult males 27–39 mm vs. 40–47 mm, respec-tively, 32–41 mm), dermal folds and tubercles ondorsum and dermal spines above the eyes (vs.absence of these structures), vocal sac morphol-ogy (single subgular or slightly bilobed andgrayish vs. paired subgular with blackish foldsalong the lower jaw), and advertisement calls.On the basis of general appearance, M. moseri issimilar to Mantidactylus redimitus, Mantidactylustandroka and especially to Mantidactylus cornutusand Mantidactylus tschenki. All these species lackthe dorsal chevron ridge and generally have nodistinct median white stripe on the throat. Man-
tidactylus redimitus is much larger (males 42–50mm SVL), has generally shorter hind limbs (tib-iotarsal articulation never reaching beyondsnout tip), and a much smoother dorsal surfacewithout large dermal spines. Mantidactylus cor-nutus, which occurs sympatrically with M. mos-eri at Andasibe, has smaller spines above theeyes and generally a more distinct pair of black-ish tubercles between the eyes. The same ap-plies to M. tschenki, whereas M. tandroka is easilydistinguished by the presence of dorsolateralridges (absent in M. moseri). Furthermore, thenew species differs from M. redimitus, M. cor-nutus and M. tschenki by the chevron-shapedridge on the anterior dorsum, shorter note du-ration of advertisement calls (74–88 ms vs. 274–352 ms in M. redimitus, 90–113 in M. cornutus,and 274–335 im M. tschenki) and a much higherdominant frequency (4000–4650 Hz vs. 900–1500 Hz in M. redimitus and 1300–2100 Hz inM. cornutus). M. moseri of is also superficiallysimilar to M. asper and M. spinifer (subgenus Ge-phyromantis). It differs from these species, how-ever, by shape (slightly bilobed vs. distinctlypaired) and color (grayish vs. black) of the vocalsac in males. Mantidactylus asper furthermorehas a totally different advertisement call (Blom-mers-Schlosser, 1979).
KEY TO ADULT MALES OF Mantidactylus SPECIESIN THE SUBGENUS Phylacomantis
1a. Blackish paired subgular vocal sacs (easilyrecognizable as dark fold along the lower jaw);supraocular tubercles absent or indistinct ------- 2
1b. Grayish single or bilobed vocal sacs (not easilyrecognizable as dark fold along the lower jaw);supraocular tubercles distinct, often elevatedto spines ----------------------------------------------------------------- 6
2a. Tympanum large, about two-thirds of eye di-ameter; tibiotarsal articulation reaches be-tween eye and nostril; lateral metatarsaliapartly separated; no distinct border betweenlighter dorsal color and darker lateral color ---- 3
2b. Tympanum medium-sized, about one-half ofeye diameter; tibiotarsal articulation reachesbeyond snout tip; lateral metatarsalia largelyor completely separated; color border betweenlighter dorsal color and darker lateral colormore or less distinct --------------------------------------------- 4
3a. SVL 31–34 mm; advertisement call is a rapidseries of three melodious notes; northern,northeastern and northwestern Madagascar ---
--------------------------------------------------- Mantidactylus pseudoasper3b. SVL 37–38 mm; advertisement call is a series
of at least 13 slowly repeated unharmoniousnotes; only known from southwestern Mada-gascar (Isalo) ------------------------ Mantidactylus corvus
4a. Prominent interocular tubercles present; lon-gitudinal dorsolateral folds on the back pres-ent --------------------------------------- Mantidactylus tandroka
4b. Prominent interocular tubercles absent (black-ish spots can be present); longitudinal dorso-lateral folds on the back absent ------------------------- 5

377NEW MANTIDACTYLUS FROM MADAGASCAR
5a. Upper lip often with a white streak (if dark,always without yellow or beige patches); fem-oral glands indistinct, of less than 2 mmwidth; inner metatarsal tubercle large andprominent ----------------------- Mantidactylus granulatus
5b Upper lip usually dark (sometimes with yel-low or beige patches); femoral glands distinct,of at least 2 mm width; inner metatarsal tu-bercle small -------------- Mantidactylus leucomaculatus
6a. SVL large, 43–53 mm; hind limbs short (tib-iotarsal articulation reaches at most betweennostril and snout tip but usually between eyeand nostril when hind limbs are adpressedalong the body) supraocular tubercles mod-erately developed ------------- Mantidactylus redimitus
6b. SVL smaller, 27–40 mm; hind limbs long (tib-iotarsal articulation reaches at least betweeneye and nostril but often beyond the snout tipwhen hind limbs are adpressed along thebody); supraocular tubercles well developedand partly elevated to spines ----------------------------- 7
7a. SVL 27–39 mm; a chevron-shaped ridge onthe anterior dorsum ------------- Mantidactylus moseri
7b. SVL 35–40 mm; no chevron-shaped ridge onthe anterior dorsum --------------------------------------------- 8
8a. Vocal sac single subgular; note duration of ad-vertisement calls 90–113 ms; central, easternMadagascar ------------------------ Mantidactylus cornutus
8b. Vocal sac bilobate subgular; note duration ofadvertisement calls 274–335 ms; southeasternMadagascar ------------------------- Mantidactylus tschenki
DISCUSSION
The discovery of M. moseri adds another spe-cies to the herpetofauna of Andasibe, a midal-titude site in central, eastern Madagascar witha surprisingly high anuran species diversity(Blommers-Schlosser and Blanc, 1991; Glaw andVences, 1994). Despite of the fact that the forestsaround Andasibe have been rather intensivelystudied, we did not find any further specimensof M. moseri in the important collections of Paris(MNHN) and Amsterdam (ZMA). Around An-dasibe, we heard and found M. moseri only oncewithin many years of fieldwork along a singlesmall brook. This may indicate that its occur-rence in the speciose Malagasy anuran com-munity is very localized and that its mating ac-tivity is restricted to a short period or to specialweather conditions.
The ongoing discovery of additional anuranspecies in the forests around Andasibe (e.g.,Glaw and Vences, 1999) reinforces the conclu-sion that the central, eastern midaltitude rainforests are a biodiversity hotspot within Mada-gascar (Lees, 1996). Midaltitudes (700–800 m)were also found to harbour the highest herpe-tological species diversity in the Marojejy massifin northeastern Madagascar (Raselimanana etal., 2000).
Specimens attributed to Mantidactylus moseriare known from four localities with a maximum
distance of about 500 km from each other andwithin an altitudinal range of approximately300–850 m above sea level. Therefore, we canexpect that its distribution covers a significantpart of the Malagasy rain forests. In addition,two localities are located within protected areas(Analamazaotra Special Reserve and MarojejyNational Parc, respectively). Therefore, no im-mediate threats for its survival are discernibleas long as an effective protection of the reservenetwork of Madagascar is assured.
Acknowledgments.—We are indebted to N. Ra-bibisoa and O. Ramilison for their help in thefield and to the Malagasy authorities for re-search permission and the permit to exportvoucher specimens. The research in Madagascarwas made possible by cooperation between theUniversity of Antananarivo and the ‘‘Zoolo-gisches Forschungsinstitut und Museum A. Ko-enig,’’ Bonn. The work of both authors was fi-nancially supported by the ‘‘Deutscher Aka-demischer Austauschdienst’’ (DAAD).
LITERATURE CITED
BLOMMERS-SCHLOSSER, R. M. A. 1979. Biosystematicsof the Malagasy frogs. I. Mantellinae (Ranidae).Beaufortia 29:1–77.
BLOMMERS-SCHLOSSER, R. M. A., AND C. P. BLANC.1991. Amphibiens (premiere partie). Faune deMadagascar 75:1–379.
DUBOIS, A. 1992. Notes sur la classification des Ran-idae (Amphibiens Anoures). Bulletin mensuel dela Societe linneenne de Lyon 61:305–352.
GLAW, F., AND M. VENCES. 1994. A Fieldguide to theAmphibians and Reptiles of Madagascar. 2nd ed.Vences und Glaw Verlag, Koln, Germany.
. 1999. Resurrection and redescription of Man-tidactylus tricinctus from eastern Madagascar. Jour-nal of Herpetology 33:639–647.
. 2000. Current counts of species diversity andendemism of Malagasy amphibians and reptiles.In W. R. Lourenco and S. M. Goodman (eds.), Div-ersite et endemisme a Madagascar, pp. 243–248.Memoires de la Societe de Biogeographie, Paris.
. 2001. Two new sibling species of Mantidac-tylus cornutus from Madagascar (Amphibia, Anura,Ranidae). Spixiana 24:177–190.
GLAW, F., M. VENCES, AND V. GOSSMANN. 2000. Anew species of Mantidactylus (subgenus Guibeman-tis) from Madagascar, with a comparative surveyof internal femoral gland structure in the genus(Amphibia: Ranidae: Mantellinae). Journal of Nat-ural History 34:1135–1154.
LEES, D. C. 1996. The Perinet effect? Diversity gradi-ents in an adaptive radiation of Madagascan but-terflies (Satyrinae: Mycalesina) contrasted with oth-er species-rich rainforest taxa. In W. R. Lourenco(ed.), Actes du Colloque International Biogeogra-phie de Madagascar, pp. 479–490. Societe de Bio-geographie-Museum-ORSTOM, Paris.
MYERS, C. W., AND W. E. DUELLMAN. 1982. A newspecies of Hyla from Cerro Colorado, and othertree frog records and geographical notes from

378 F. GLAW AND M. VENCES
western Panama. American Museum Novitates2752:1–32.
RASELIMANANA, A. P., C. J. RAXWORTHY, AND R. A.NUSSBAUM. 2000. Herpetofaunal species diversityand elevational distribution within the Parc Na-tional de Marojejy, Madagascar. In S. M. Goodman(ed.), A floral and faunal inventory of the Parc Na-tional de Marojejy: with reference to elevationalvariation. Fieldiana Zoology (N.S.) 92:157–174.
SAVAGE, J. M., AND W. R. HEYER. 1967. Variation and
distribution in the tree frog genus Phyllomedusa inCosta Rica, Central America. Beitrage zur neotro-pischen Fauna 5:111–131.
. 1997. Digital webbing formulae for anurans:a refinement. Herpetological Review 28:131.
VENCES, M., AND F. GLAW. 2001. When moleculesclaim for taxonomic changes: new proposals on theclassification of Old World treefrogs (Amphibia,Anura, Ranoidea). Spixiana 24:85–92.
Accepted: 6 November 2001.
Journal of Herpetology, Vol. 36, No. 3, pp. 378–389, 2002Copyright 2002 Society for the Study of Amphibians and Reptiles
Population Characteristics and Activity Patterns of the NamaqualandSpeckled Padloper (Homopus signatus signatus) in the Early Spring
VICTOR J. T. LOEHR
Homopus Research Foundation, Nipkowplein 24, 3402 EC IJsselstein, Netherlands; E-mail: [email protected]
ABSTRACT.—Early spring population parameters and activity of wild Namaqualand speckled padlopers(Homopus signatus signatus) were monitored using mark-recapture and thread-trailing methods. Eighty-threetortoises were studied between 22 August and 22 September 2000. The maximum number of scute ringscounted was 14, with females appearing to have longer lives than males. Tortoises were encountered onrock slabs, on a rocky hill slope, and frequented adjacent, gently sloping fields with many flowering plants.The frequent use of shallow concealed crevices as hiding places may be, in part, caused by greater avail-ability of such refuges. Single specimens were usually found in refugia, but occasionally male-female pairswere observed. The shelter types identified (rock and spaces under shrubs) were rarely used for more thanone night. Activity was unimodal. Activity area and daily movements were lower in H. s. signatus than inother tortoise species, possibly because of their smaller body size, cooler ambient spring temperatures duringthis study, a short measurement period, smaller habitat requirement of H. s. signatus, or some combinationof the four.
The Namaqualand speckled padloper, Homo-pus signatus signatus, is endemic to Namaqua-land, South Africa (Ernst et al., 2000), an arid(150–250 mm rainfall per annum) region ofsmall shrubs (Succulent Karoo; Branch, 1998),and other succulents (Le Roux and Schelpe,1997). Homopus s. signatus appears plentiful insuitable habitat (i.e., rocky, granite hills, knownas Namaqualand Klipkoppe; Boycott and Bour-quin, 1988; Baard, 1994; Bayoff, 1995; Branch,1998), but the species’ range is restricted, em-phasizing its potential vulnerability. Apparentthreats to H. s. signatus include mining (causinghabitat destruction and dust production whichinhibits plant growth), overgrazing by goats andsheep, and road casualties (N. Bayoff and V.Loehr, pers. obs.; K. van Zijl, pers. comm.; J. vanAardt, pers. comm.); such threats have not beenevaluated rigorously. Most of the published in-formation for the species is from captive ani-mals (e.g., Palmer, 1994; Loehr, 1999a,b). In con-
trast, one study of wild H. s. signatus exists (Bay-off, 1995), and a few life-history notes are avail-able (Loehr and Harris-Smith, 1999; Licitra,2001). A basic knowledge of the ecology of H.s. signatus is critical for the long-term manage-ment and conservation of this species.
In 2000, a long-term project was initiated toincrease knowledge of the ecology of H. signatusin Namaqualand. This article evaluates the earlyspring population characteristics, activity pat-terns and movement of H. s. signatus.
MATERIALS AND METHODS
Study Site and Weather Conditions.—Homopus s.signatus was studied in the spring from 22 Au-gust to 22 September 2000 near Springbok, Na-maqualand, South Africa (2917DB). The site (ap-proximately 36,340 m2) overlapped a part of the6000 m2 site studied by Bayoff (1995) in 1991and 1992, and consisted of a southeasterly fac-ing rocky hill (altitude approximately 870–920