a new dynamin-like protein, adl6, is involved in trafficking from the trans-golgi network to the
TRANSCRIPT

The Plant Cell, Vol. 13, 1511–1525, July 2001, www.plantcell.org © 2001 American Society of Plant Biologists
RESEARCH ARTICLE
A New Dynamin-Like Protein, ADL6, Is Involved in Trafficking
from the
trans
-Golgi Network to the Central Vacuolein Arabidopsis
Jing Bo Jin,
a
Young A Kim,
b
Soo Jin Kim,
b
Sung Hoon Lee,
b
Dae Heon Kim,
b
Gang-Won Cheong,
c
and Inhwan Hwang
b,1
a
Department of Molecular Biology, Gyeongsang National University, Chinju 660-701, Korea
b
Center for Plant Intracellular Trafficking and Division of Molecular and Life Sciences, Pohang University of Science and Technology, Pohang 790-784, Korea
c
Department of Biochemistry, Gyeongsang National University, Chinju 660-701, Korea
Dynamin, a high-molecular-weight GTPase, plays a critical role in vesicle formation at the plasma membrane during en-docytosis in animal cells. Here we report the identification of a new dynamin homolog in Arabidopsis named Arabidopsisdynamin-like 6 (ADL6). ADL6 is quite similar to dynamin I in its structural organization: a conserved GTPase domain at theN terminus, a pleckstrin homology domain at the center, and a Pro-rich motif at the C terminus. In the cell, a majority ofADL6 is associated with membranes. Immunohistochemistry and in vivo targeting experiments revealed that ADL6 is lo-calized to the Golgi apparatus. Expression of the dominant negative mutant ADL6[K51E] in Arabidopsis protoplasts inhib-
ited trafficking of cargo proteins destined for the lytic vacuole and caused them to accumulate at the
trans
-Golgi
network. In contrast, expression of ADL6[K51E] did not affect trafficking of a cargo protein, H
�
-ATPase:green fluorescentprotein, destined for the plasma membrane. These results suggest that ADL6 is involved in vesicle formation for vacuolartrafficking at the
trans
-Golgi network but not for trafficking to the plasma membrane in plant cells.
INTRODUCTION
In eukaryotic cells, a large number of proteins are trans-ported to their final destination by a process called intracel-lular trafficking. During the past two decades, intracellulartrafficking has been studied extensively in yeast, animal,and plant cells (Rothman, 1994; Hawes et al., 1999; Jahnand Südhof, 1999; Bassham and Raikhel, 2000). As in thecases of animal and yeast cells, the endoplasmic reticulum(ER), Golgi apparatus, plasma membrane, and vacuoles arethe main components of the endomembrane systems inplant cells, and it is generally assumed that the mechanismof intracellular trafficking between compartments in plantcells would be similar to that in other systems (Hawes et al.,1999). Recent advances in the understanding of intracellulartrafficking in plant cells support this notion. For example,many plant genes have been shown to complement muta-tions in corresponding genes in yeast cells (Bassham et al.,1995; Takeuchi et al., 1998). Furthermore, many proteinsknown to be involved in trafficking, such as clathrin, thecoatomer subunits of coat protein 1 (COPI) vesicles, various
soluble
N
-ethylmaleimide–sensitive factor attachment pro-tein receptors, and many small GTP binding proteins, havebeen identified in plant cells (Lebas and Axelos, 1994;Blackbourn and Jackson, 1996; Movafeghi et al., 1999; Pimplet al., 2000; Sanderfoot et al., 2000; Takeuchi et al., 2000).
AtVPS45, a homolog of yeast Sec1p that is thought to beinvolved in vesicle fusion, has been identified in Arabidopsis(Bassham et al., 2000), and Kim et al. (2001) recently demon-strated that phosphatidylinositol 3-phosphate [PI(3)P] playsan important role in vacuolar trafficking in plant cells, as itdoes in animal and yeast cells (Schu et al., 1993; Jones andHowell, 1997). However, recent studies also have revealedmany differences between plant and animal cells in intracellu-lar trafficking (Baba et al., 1997; Bassham and Raikhel, 2000;Toyooka et al., 2000). An example of this is the presence inplant cells of at least two different types of vacuoles: lytic andstorage vacuoles (Neuhaus and Rogers, 1998). The lytic vac-uole is thought to be similar to the vacuole of yeast cells orthe lysosome of animal cells, whereas an organelle that issimilar to the plant storage vacuole is not present in yeast oranimal cells, suggesting that trafficking between the storagevacuole and other organelles is likely unique to plant cells
1
To whom correspondence should be addressed. E-mail [email protected]; fax 82-54-279-8159.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

1512 The Plant Cell
Figure 1. Amino Acid Sequence Alignment of ADL6.
The deduced amino acid sequence of ADL6 was aligned with the sequences of dynamin I, Vps1p, and aG68 using the multiple alignment pro-gram of DNASIS (Hitachi, Tokyo, Japan). Gaps were introduced to maximize identity. Identical amino acid residues are indicated by dark shad-ing. The GTPase domain, pleckstrin homology (PH) domain, and Pro-rich domain (PRD) are indicated according to the domains of dynamin I.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

ADL6 in Trafficking from the TGN to the Vacuole 1513
(Jiang et al., 2000). Interestingly, precursor-accumulating ves-icles have been demonstrated to transport storage proteinsto the storage vacuole directly from the ER in pumpkin seeds(Hara-Nishimura et al., 1998). In addition, storage proteinssuch as vicilin have been shown to be sorted to the storagevacuole at the
cis
-Golgi (Hillmer et al., 2001).Although recent efforts to unravel the mechanism of intra-
cellular trafficking in plant cells have made significantprogress toward understanding the trafficking of cargo mol-ecules to the lytic vacuole (Baba et al., 1997; Frigerio et al.,1998; Zheng et al., 1999; Ahmed et al., 2000; Jiang et al.,2000), the vast majority of molecular players involved in thevarious steps of intracellular trafficking are still unknown. Tounderstand the mechanism of intracellular trafficking inplant cells, we decided to investigate the proteins involvedin vesicle formation. In animal cells, dynamin, a high-molec-ular-weight GTPase, has been shown to play a critical role invesicle formation during endocytosis (Obar et al., 1990). Themechanism by which dynamin plays a role in vesicle forma-tion has been well characterized in animal cells (Gout et al.,1993; Herskovits et al., 1993; Damke et al., 1994; Hinshawand Schmid, 1995; Sweitzer and Hinshaw, 1998; Marks etal., 2001). Vps1p, a homolog of dynamin, also has beenshown to be involved in vacuolar targeting of proteins inyeast (Rothman et al., 1990; Bensen et al., 2000). Now, dy-namin and related proteins have been isolated from a varietyof eukaryotic cells, ranging from yeast to human cells, andhave been found to constitute a family of high-molecular-weight-GTP binding proteins (Obar et al., 1990; Rothman etal., 1990; Chen et al., 1991; Dombrowski and Raikhel, 1995;Gu and Verma, 1996; Kang et al., 1998). We previously iden-tified two proteins in Arabidopsis, ADL1 and ADL2, thathave a high degree of amino acid sequence homology withdynamin (Kang et al., 1998; Park et al., 1998). However, theyare not likely to be involved in vesicle formation during intracel-lular trafficking because both proteins have been shown to belocalized within the chloroplast (Kang et al., 1998; Park etal., 1998). Also, another dynamin homolog, phragmoplastin,found in soybean has been shown to be involved in the forma-tion of the cell division plate (Gu and Verma, 1996, 1997).
Here we report that a new dynamin homolog, Arabidop-sis dynamin-like 6 (ADL6), is localized to the Golgi appa-ratus and is involved in the vacuolar trafficking of cargoproteins from the
trans
-Golgi network (TGN) to the lyticvacuole.
RESULTS
Isolation of the cDNA
ADL6
EncodingDynamin-Like Protein
A search through the expressed sequence tag (EST) analy-sis files (University of Minnesota, Plant Molecular Informat-ics Center) resulted in the identification of 12 EST clones
that had significant sequence homology with dynamin. Anal-ysis of these EST clones suggested that there were at leastsix dynamin-like protein genes in Arabidopsis. A genomicDNA fragment corresponding to the EST clone W43823 wasamplified by polymerase chain reaction (PCR) using specificprimers corresponding to the 5
�
and 3
�
ends of the ESTclone and used as a hybridization probe to screen an Arabi-dopsis ZAPII
�
cDNA library. Five positive clones were ob-tained from the screening, and pBluescript SK
�
clones wereexcised from these clones. The cDNA clone was named
ADL6
. The cDNA clone with the largest insert was selected,and the nucleotide sequence was determined using a dyeterminator sequencing kit. The nucleotide sequence wasdeposited in GenBank with the accession number AF180732.The size of the
ADL6
cDNA was 3.21 kb. The first Metcodon was located at nucleotide 205 and was followed byan open reading frame of 2.74 kb and an untranslated re-gion of 263 bp.
Sequence Analysis of ADL6
ADL6
has an open reading frame of 2742 bp, which wouldencode a protein of 914 amino acid residues with a calcu-lated molecular mass of 100 kD. The amino acid sequenceof ADL6 reveals the presence of a GTP binding domain atthe N terminus, a pleckstrin homology domain in the center,and a Pro-rich SH3 binding domain at the C terminus, indi-cating that it belongs to the dynamin family of proteins(Obar et al., 1990; Chen et al., 1991). Thus, it appears thatADL6 is more closely related to dynamin I than to othermembers of the dynamin family with regard to its structuralorganization. The deduced amino acid sequence of ADL6also showed a high degree of similarity with other membersof the dynamin family; that is, it shared 26% amino acid se-quence identity with ADL2 (Kang et al., 1998), 20% with dy-namin I (Obar et al., 1990), and 19% with Vps1p (Rothmanet al., 1990). The sequence alignments of ADL6 and otherdynamin-like proteins are shown in Figure 1.
Subcellular Localization of ADL6
To understand the biological role of ADL6, we next exam-ined its subcellular distribution by protein gel blot analysis.To perform this experiment, we raised a polyclonal antibodyagainst the C-terminal region (amino acid residues 521 to914) of ADL6 expressed in
Escherichia coli
in rabbits. Inwhole cell extracts, the antibody specifically recognized oneband of
�
100 kD, a size that was in good agreement withthe calculated molecular mass of the ADL6 gene, whereasthe control serum did not detect any protein bands (Figure2A), suggesting that the antibody was specific to ADL6.However, we often observed an additional weak band rightbelow the major band, and the intensity of the weak band
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

1514 The Plant Cell
(lower band) varied from experiment to experiment. Interest-ingly, the lower band was present preferentially in the solu-ble fraction (Figure 2B).
To examine the subcellular distribution of ADL6, wholeextracts of Arabidopsis seedlings were fractionated into sol-uble, membrane, and pellet fractions by Suc step gradientcentrifugation, and ADL6 was detected in the various frac-tions by protein gel blot analysis using the polyclonal anti-ADL6 antibody. Also, to control the quality of fractionationby the Suc step gradient, we used other antibodies, such aspolyclonal anti-PIP and anti-SPK4 antibodies (Park et al.,1997), which detect plasma membrane intrinsic protein (PIP)and a soluble soybean protein kinase 4 homolog (SPK-4),respectively. As shown in Figure 2B, ADL6 was present in allfractions. However, the protein species in the upper fractionof the gradient was different from that detected in the mem-brane and pellet fractions; that is, the lower migrating bandpredominated in the upper fraction and the upper migratingband predominated in the membrane and pellet fractions.
ADL6 Is Localized to the Golgi Apparatus
To understand the biological role of ADL6, we attempted tolocalize ADL6 using the anti-ADL6 antibody and a fluores-cein isothiocyanate (FITC)–conjugated secondary antibody.
Cryosections were prepared from root tips and used for im-munolocalization studies. As shown in Figure 3A, the greenfluorescent signals of FITC were observed as punctatestaining in cortical cells of the root tips (left). However, whenthe root tips were treated before fixing with brefeldin A (BFA),a Golgi-disrupting agent (Misumi et al., 1986; Fujiwara et al.,1988; Driouich et al., 1993), the punctate staining pattern dis-appeared and was replaced instead by a diffuse stainingpattern (middle), raising the possibility that ADL6 may be as-sociated with the Golgi apparatus. To unequivocally demon-strate its localization, we attempted colocalization withJIM84, a monoclonal antibody that was shown to detect N-gly-cans with an oligosaccharide sequence of Gal
�
(1-3)[Fuc
�
(1-4)]GlcNAc at the Golgi apparatus in plant cells (Satiat-Jeunemaitreand Hawes, 1992; Fitchette et al., 1999).
Previously, it was thought that Arabidopsis cells werenegative for the JIM84 epitope in the Golgi apparatus. Re-cently, however, we learned that this epitope can be usedas a Golgi marker in Arabidopsis (C. Hawes, personal com-munication). As shown in Figure 3B, the JIM84 antibodygave a punctate staining pattern in the cortical cells of Ara-bidopsis root tips (left), indicating that the JIM84 antibodyrecognizes an epitope in the Golgi apparatus of Arabidop-sis, as in other plant cells. In addition, when the tissues weretreated with BFA before fixation, the punctate staining patterndetected by JIM84 aggregated into larger spots (middle),
Figure 2. Subcellular Distribution of ADL6.
(A) Specificity of the polyclonal anti-ADL6 antibody. A polyclonal anti-ADL6 antibody was generated in a rabbit using the C-terminal region(amino acids 521 to 914) of ADL6 expressed in E. coli as a recombinant protein. Total protein (20 �g) obtained from leaf tissues was separatedon an SDS-polyacrylamide gel and transferred onto nylon (polyvinylidene difluoride) membranes. The blots were probed with either the poly-clonal anti-ADL6 antibody (A) or a control serum (C).(B) Subcellular distribution of ADL6. Total protein was fractionated on a Suc step gradient (15, 26, and 35%) by ultracentrifugation. Top, mem-brane (26/35%), and pellet fractions were collected separately. Twenty micrograms (Total and Top) or 5 �g (Membrane and Pellet) of protein wasseparated by SDS-PAGE, and protein gel blot analysis was performed with the polyclonal anti-ADL6 antibody. Polyclonal anti-PIP and anti-SPK4 antibodies were used as controls for the fractionation of membrane and soluble fractions, respectively.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022
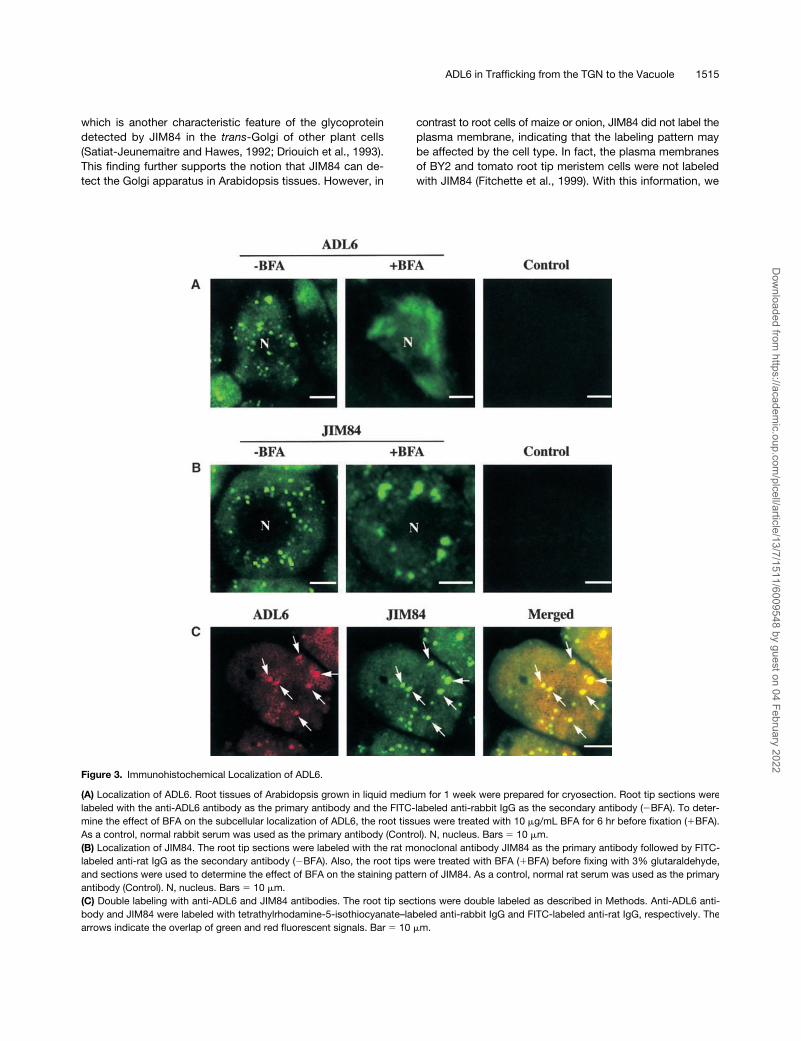
ADL6 in Trafficking from the TGN to the Vacuole 1515
which is another characteristic feature of the glycoproteindetected by JIM84 in the
trans
-Golgi of other plant cells(Satiat-Jeunemaitre and Hawes, 1992; Driouich et al., 1993).This finding further supports the notion that JIM84 can de-tect the Golgi apparatus in Arabidopsis tissues. However, in
contrast to root cells of maize or onion, JIM84 did not label theplasma membrane, indicating that the labeling pattern maybe affected by the cell type. In fact, the plasma membranesof BY2 and tomato root tip meristem cells were not labeledwith JIM84 (Fitchette et al., 1999). With this information, we
Figure 3. Immunohistochemical Localization of ADL6.
(A) Localization of ADL6. Root tissues of Arabidopsis grown in liquid medium for 1 week were prepared for cryosection. Root tip sections werelabeled with the anti-ADL6 antibody as the primary antibody and the FITC-labeled anti-rabbit IgG as the secondary antibody (�BFA). To deter-mine the effect of BFA on the subcellular localization of ADL6, the root tissues were treated with 10 �g/mL BFA for 6 hr before fixation (�BFA).As a control, normal rabbit serum was used as the primary antibody (Control). N, nucleus. Bars 10 �m.(B) Localization of JIM84. The root tip sections were labeled with the rat monoclonal antibody JIM84 as the primary antibody followed by FITC-labeled anti-rat IgG as the secondary antibody (�BFA). Also, the root tips were treated with BFA (�BFA) before fixing with 3% glutaraldehyde,and sections were used to determine the effect of BFA on the staining pattern of JIM84. As a control, normal rat serum was used as the primaryantibody (Control). N, nucleus. Bars 10 �m.(C) Double labeling with anti-ADL6 and JIM84 antibodies. The root tip sections were double labeled as described in Methods. Anti-ADL6 anti-body and JIM84 were labeled with tetrathylrhodamine-5-isothiocyanate–labeled anti-rabbit IgG and FITC-labeled anti-rat IgG, respectively. Thearrows indicate the overlap of green and red fluorescent signals. Bar 10 �m.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

1516 The Plant Cell
addressed the question of whether the two punctate stain-ing patterns detected by anti-ADL6 antibody and JIM84overlapped each other. As shown in Figure 3C, the red(ADL6) and green (JIM84) punctate staining patterns clearlyoverlapped each other (arrowheads), as indicated by theyellow areas in the merged image, suggesting that ADL6 islocalized to the Golgi apparatus.
To obtain independent evidence for the localization ofADL6, we attempted in vivo targeting using an ADL6:greenfluorescent protein (GFP) fusion protein. As a marker for theGolgi apparatus, we used rat sialyltransferase fused to redfluorescent protein (ST:RFP) (Wee et al., 1998; Kim et al.,2001). As shown in Figure 4, both ADL6:GFP and ST:RFP gavepunctate staining patterns (Figures 4a and 4b), and the ma-jority of green fluorescent signals closely overlapped the redfluorescent signals (Figure 4c). Interestingly, some of the redpunctate stains did not overlap the green fluorescent signalsof ADL6. However, when we amplified the green signals ofADL6 using image analysis software, we were able to seeweak green fluorescent signals at these red punctate stains(data not shown). The difference in the signal intensity ofADL6 at the Golgi apparatus may be attributable to the dy-namic nature of ADL6. As in the case of dynamin I, ADL6also appeared to cycle between soluble dimeric and mem-brane-associated multimeric forms (J.B. Jin and I. Hwang,unpublished data). Also, when GFP was placed at the N ter-minus of ADL6, ADL6:GFP gave punctate staining patternsthat overlapped ST:RFP (data not shown). Thus, the in vivotargeting experiments further support the notion that ADL6is localized to the Golgi apparatus.
ADL6 Is Involved in the Trafficking of Cargo Molecules from the TGN to the Central Vacuole
Previously, it was shown that a mutation (K44A) located inthe first GTPase binding motif of dynamin I inhibits endocy-tosis in animal cells by preventing the formation of vesicles
from the plasma membrane (Herskovits et al., 1993; Damkeet al., 1994). We decided to take a similar mutation-basedapproach to investigate the biological role of ADL6 bymeans of an in vivo trafficking assay that uses protoplaststhat transiently express reporter proteins destined for variousorganelles (Kim et al., 2001). An ADL6 mutant, ADL6[K51E],was generated for use as a dominant negative mutant, andthe wild-type and mutant proteins were tagged with the T7epitope at the N terminus (Zheng et al., 1999). BecauseADL6 appeared to be localized to the Golgi apparatus, weselected sporamin, a storage protein in the tubers of sweetpotato (Hattori et al., 1989), as a cargo protein because ithas been shown to be transported to the central vacuolethrough the Golgi apparatus (Matsuoka et al., 1995; Kim etal., 2001).
We fused GFP to sporamin to facilitate monitoring of thetrafficking in the cells. First, we examined the effect ofADL6[K51E] on the trafficking of sporamin:GFP to the cen-tral vacuole in protoplasts. As shown in Figure 5A, we usu-ally observed two different patterns of green fluorescent signalswhen protoplasts were transformed with
sporamin
:
GFP
alone or together with
ADL6
; green fluorescent signals in thecentral vacuole (Figure 5A, b) indicated that sporamin:GFPhad been targeted to its final destination, whereas green flu-orescent signals in the ER (Figure 5A, a) indicated thatsporamin:GFP had been translated but not yet translocatedto its final destination. The localization of untargeted spor-amin:GFP to the ER was confirmed by colocalization ofsporamin:GFP with chaperone binding protein:red fluores-cent protein (BiP:RFP), an ER marker protein (Figure 5A, c)(Kim et al., 2001).
Next, we examined the effect of ADL6[K51E] on the traf-ficking of sporamin:GFP. When
ADL6[K51E]
was cotrans-formed into protoplasts, we observed a new punctatestaining pattern for sporamin:GFP (Figure 5B, c) in additionto the two patterns of staining seen in the control protoplasts(Figure 5B, a and b). When we carefully examined the wholeprotoplast population at various times after transformation,
Figure 4. In Vivo Targeting of ADL6.
ADL6:GFP was cotransformed into protoplasts with ST:RFP, and the localization of these proteins was examined at various times after transfor-mation. (a), (b), and (c) are images of ADL6:GFP, ST:RFP, and the overlap of ADL6:GFP and ST:RFP, respectively. Arrowheads indicate overlapsof green and red fluorescent signals. Bar 20 �m.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

ADL6 in Trafficking from the TGN to the Vacuole 1517
Figure 5. ADL6[K51E] Inhibits Vacuolar Trafficking of Sporamin:GFP.
(A) Trafficking of sporamin:GFP to the central vacuole. Protoplasts were transformed with sporamin:GFP plus ADL6 ([a] and [b]) or sporamin:GFPplus ADL6 plus BiP:RFP (c). (a) and (b) Two patterns of GFP signals at the ER and central vacuole (V), respectively, 24 hr after transformation. (c)Colocalization of sporamin:GFP with BiP:RFP. Yellow color indicates the overlap between green and red fluorescent signals. Bars 20 �m.(B) Inhibition of vacuolar trafficking of sporamin:GFP by ADL6[K51E]. Protoplasts were transformed with sporamin:GFP plus ADL6[K51E]. (a), (b), and(c) Three types of protoplasts with GFP signals at the ER, the central vacuole, and punctate stains, respectively. V, central vacuole. Bars 20 �m.(C) Trafficking efficiency of sporamin:GFP to the central vacuole in the presence of ADL6[K51E]. The whole population of transformed proto-plasts was divided into three groups based on GFP signal localization: protoplasts with an ER staining pattern, a punctate staining pattern, or avacuolar staining pattern. A minimum of 200 transformed protoplasts in each group was counted each time. At least three independent experi-ments were performed to determine the trafficking efficiency. The first, second, and third columns for each time point indicate protoplasts trans-formed with sporamin:GFP alone, sporamin:GFP plus ADL6, and sporamin:GFP plus ADL6[K51E], respectively. The numbers are means, and theerror bars indicate SD (n 3).
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

1518 The Plant Cell
the percentage of protoplasts with GFP signals in the centralvacuole was reduced by as much as 50% compared withprotoplasts transformed with
sporamin
:
GFP
alone or with
sporamin
:
GFP
plus
ADL6
, although the targeting efficiencyrecovered slowly with time (Figure 5C). However, the per-centage of transformed protoplasts with GFP signals in theER was not affected by the presence of ADL6[K51E] (Figure5C). Thus, these results indicate that the protoplasts with apunctate staining pattern were produced at the expense ofthe protoplasts with the vacuolar staining pattern.
One possible explanation for this finding would be thatADL6[K51E] inhibits the trafficking of sporamin:GFP to thecentral vacuole and, as a result, sporamin:GFP is retained inan organelle situated between the ER and the central vacu-
ole. In addition, the punctate staining pattern raised the in-triguing possibility that sporamin:GFP may accumulate atthe Golgi apparatus in the presence of ADL6[K51E]. To ad-dress this possibility, the two constructs,
sporamin
:
GFP
and
ADL6[K51E]
, were introduced into protoplasts together withanother construct,
ST
:RFP (Kim et al., 2001). As shown inFigure 6A, the red fluorescent signals of ST:RFP at the Golgiapparatus (Figure 6A, b) appeared as punctate stains andclearly overlapped the punctate green fluorescent signals ofsporamin:GFP (Figure 6A, c, arrowheads), suggesting thatADL6[K51E] caused the accumulation of sporamin:GFP atthe Golgi apparatus, possibly in the region corresponding tothe trans-Golgi or the TGN. However, to conclude thatsporamin:GFP is accumulated at the Golgi apparatus in the
Figure 6. Colocalization of Punctate Staining with ST:RFP at the Golgi Apparatus.
(A) Protoplasts were cotransformed with three constructs, sporamin:GFP, ST:RFP, and ADL6[K51E], and the localization of sporamin:GFP (a)and ST:RFP (b) were examined 24 to 40 hr after transformation. GFP, RFP, and Merged indicate green fluorescent signals, red fluorescent sig-nals, and the overlap of red and green fluorescent signals, respectively. Arrowheads indicate the overlap of red and green fluorescent signals.Bar 20 �m.(B) Localization of ST:RFP in the presence of ADL6 and ADL6[K51E]. Protoplasts were transformed with ST:RFP plus ADL6 plus BiP:GFP ([a],[b], and [c]) or ST:RFP plus ADL6[K51E] plus BiP:GFP ([d], [e], and [f]), and localization of ST:RFP was examined in the absence ([a] and [d]) orthe presence of BFA (10 �g/mL) ([b], [c], [e], and [f]). (b) and (e) Images of ST:RFP. (a), (c), (d), and (f) Merged images between ST:RFP andBiP:GFP. Bars 20 �m.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

ADL6 in Trafficking from the TGN to the Vacuole 1519
presence of ADL6[K51E], it is necessary to show that theexpression of ADL6[K51E] does not affect the localization ofST:RFP. To address this question, we compared the stain-ing patterns of ST:RFP in protoplasts expressing ADL6 orADL6[K51E] and also examined whether ST:RFP can be re-located to the ER in the presence of BFA. Two reporter pro-teins, ST:RFP and BiP:GFP, were expressed in the presenceof ADL6 or ADL6[K51E], and their localization was examinedin the absence and presence of BFA. As shown in Figure6B, in both cases ST:RFP gave punctate staining patterns inthe absence of BFA (Figure 6B, a and d), whereas ST:RFPgave diffuse patterns (Figure 6B, b and e) that overlappedBiP:GFP in the presence of BFA (Figure 6B, c and f), indicat-ing that ADL6[K51E] does not affect the localization ofST:RFP at the Golgi apparatus.
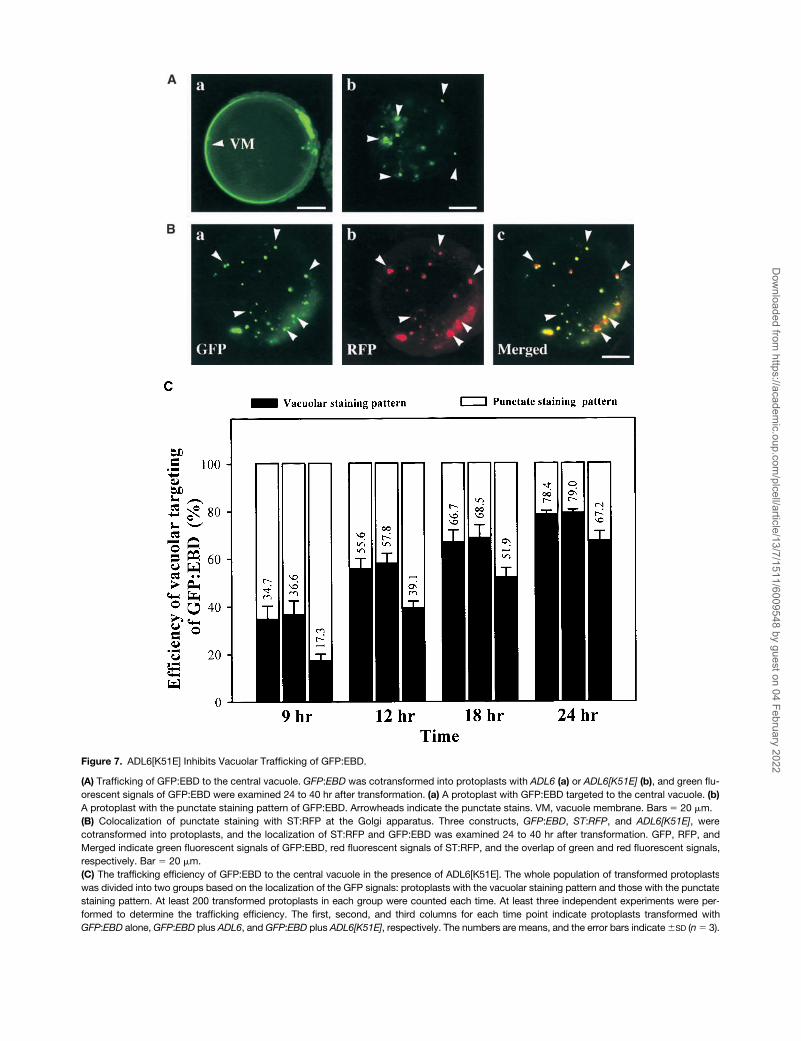
To obtain additional evidence for the role of ADL6 in traf-ficking from the Golgi apparatus to the central vacuole, weselected another cargo protein, endosome binding domain(EBD; the C-terminal region ranging from amino acid resi-dues 1257 to 1411) of human early endosome antigen 1(Stenmark et al., 1996). The C-terminal EBD that has anRab5 binding motif and an FYVE zinc finger binding domainhas been shown to be responsible for the PI(3)P-dependentlocalization of early endosome antigen 1 to the endosome.Interestingly, when EBD was expressed as a GFP fusionprotein in plant cells, GFP:EBD was transported from theTGN to the central vacuole in a PI(3)P-dependent manner(Kim et al., 2001). As shown in Figure 7A, GFP:EBD wastransported to the central vacuole in the presence of ADL6(Figure 7A, a). When ADL6[K51E] was cotransformed withGFP:EBD, the targeting efficiency of GFP:EBD was de-creased by as much as 50% at 9 hr after transformationcompared with that of protoplasts transformed with GFP:EBD alone or with GFP:EBD plus ADL6 (Figure 7C). At thesame time, we observed an increase in the number of proto-plasts with a punctate staining pattern for GFP:EBD (Figure7A, b). The punctate signals of the green fluorescence ofGFP:EBD (Figure 7B, a) clearly overlapped the red fluores-cence of ST:RFP (Figure 7B, c, arrowheads), as was the casefor sporamin:GFP, indicating that ADL6[K51E] also inhibitedtrafficking of GFP:EBD to the central vacuole at the TGN. Thisresult was very similar to that found with sporamin:GFP,and, as in that case, the efficiency recovered slowly at latertimes. This would be expected if the trafficking had not beeninhibited completely. Together, these results suggested thatADL6[K51E] had the same inhibitory effect on the two cargoproteins, sporamin:GFP and GFP:EBD, at the TGN.
ADL6[K51E] Does Not Inhibit Trafficking to thePlasma Membrane
To further investigate the specificity of ADL6 involvementin intracellular trafficking, we examined the effect ofADL6[K51E] on the destiny of the integral plasma membraneprotein H�-ATPase:GFP (Kim et al., 2001). As shown in Fig-
ure 8, H�-ATPase:GFP was transported to the plasmamembrane in the control protoplasts as expected (Figure8b). Also, in protoplasts expressing ADL6[K51E] (Figure 8c),H�-ATPase:GFP was transported to the plasma membraneas efficiently as in the control protoplasts and did not accu-mulate as a punctate pattern in the cytosol, suggesting thatADL6 is not involved in the trafficking of cargo proteins tothe plasma membrane.
To confirm that the difference in the targeting efficiencywas attributable to ADL6[K51E], we examined the expres-sion levels of the constructs by performing protein gel blotanalysis of protein extracts obtained from the transformedprotoplasts. The monoclonal anti-T7 and anti-GFP antibod-ies were used to detect ADL6 and reporter proteins, respec-tively. As shown in Figure 9, wild-type and mutant ADL6were expressed at equal levels. Also, the reporter proteinsGFP:EBD (Figure 9A) and sporamin:GFP (Figure 9B) wereexpressed at nearly equal levels. Thus, this result excludedthe possibility that the differential effects of the wild-typeand mutant ADL6 proteins on trafficking were caused by adifference in their expression levels.
DISCUSSION
The structural similarity of ADL6 to dynamin I strongly sug-gests that ADL6 may play a role in vesicle formation at cer-tain membranes. If this is the case, localization of ADL6could reveal important information about the pathways inwhich it plays a role. Thus, we first determined the subcellu-lar distribution of ADL6 by subcellular fractionation experi-ments. When the products of the subcellular fractionationwere examined by protein gel blot analysis, the majority ofADL6 was found to be associated with membranes, as in thecase of dynamin (Tuma et al., 1993). Interestingly, however,the soluble form of ADL6 migrated slightly ahead of themembrane-bound form. One possible explanation for the differ-ence in the migration rate between the two forms is that ADL6may be subject to post-translational modification such asphosphorylation. Previously, it was shown that dynaminand dynamin-like proteins are modified by phosphorylationand that they appear as doublets on protein gel blots. Thephosphorylated form is found in the soluble fraction and theunmodified form is found in the membrane fraction (Tuma etal., 1993; Park et al., 1997). However, further studies arenecessary to understand the difference in the migration ratebetween the soluble and membrane-bound forms of ADL6.
Next, we established that ADL6 is localized to the Golgiapparatus. This conclusion was based on immunohisto-chemistry and in vivo targeting. These techniques, however,were not able to reveal the exact location of ADL6 within theGolgi apparatus because of their limited resolution. One pos-sibility is that ADL6 may be localized to the trans-Golgi orTGN because N-glycans with an oligosaccharide sequenceof Gal�(1-3)[Fuc�(1-4)] GlcNAc, the epitope recognized by
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022
1520 The Plant Cell
Figure 7. ADL6[K51E] Inhibits Vacuolar Trafficking of GFP:EBD.
(A) Trafficking of GFP:EBD to the central vacuole. GFP:EBD was cotransformed into protoplasts with ADL6 (a) or ADL6[K51E] (b), and green flu-orescent signals of GFP:EBD were examined 24 to 40 hr after transformation. (a) A protoplast with GFP:EBD targeted to the central vacuole. (b)A protoplast with the punctate staining pattern of GFP:EBD. Arrowheads indicate the punctate stains. VM, vacuole membrane. Bars 20 �m.(B) Colocalization of punctate staining with ST:RFP at the Golgi apparatus. Three constructs, GFP:EBD, ST:RFP, and ADL6[K51E], werecotransformed into protoplasts, and the localization of ST:RFP and GFP:EBD was examined 24 to 40 hr after transformation. GFP, RFP, andMerged indicate green fluorescent signals of GFP:EBD, red fluorescent signals of ST:RFP, and the overlap of green and red fluorescent signals,respectively. Bar 20 �m.(C) The trafficking efficiency of GFP:EBD to the central vacuole in the presence of ADL6[K51E]. The whole population of transformed protoplastswas divided into two groups based on the localization of the GFP signals: protoplasts with the vacuolar staining pattern and those with the punctatestaining pattern. At least 200 transformed protoplasts in each group were counted each time. At least three independent experiments were per-formed to determine the trafficking efficiency. The first, second, and third columns for each time point indicate protoplasts transformed withGFP:EBD alone, GFP:EBD plus ADL6, and GFP:EBD plus ADL6[K51E], respectively. The numbers are means, and the error bars indicate SD (n 3).
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

ADL6 in Trafficking from the TGN to the Vacuole 1521
the JIM84 antibody, are known to be synthesized at thetrans-most part of the Golgi apparatus (Fitchette et al.,1999) (see below for further discussion). This location is sim-ilar to that of animal dynamin II, which has been shown to belocalized to the TGN and to play roles in post-Golgi traffick-ing (Henley and McNiven, 1996; Maier et al., 1996; Cao etal., 1998; Jones et al., 1998). Also, Vps1p, which plays a rolein vacuolar trafficking, has been shown to be localized to theGolgi apparatus in yeast (Wilsbach and Payne, 1993). How-ever, this is in contrast to dynamin I, which is localized to theplasma membrane in animal cells (Herskovits et al., 1993;Robinson et al., 1993; Sweitzer and Hinshaw, 1998).
Among the members of the dynamin family, the molecularmechanism by which dynamin I plays a role during endocy-tosis has been best characterized. Dynamin I is recruited andassembled into rings at the necks of clathrin-coated buds atthe plasma membrane, and a conformational change of thedynamin rings results in the severance of the necks to re-lease the buds as vesicles during endocytosis (Herskovits etal., 1993; Robinson et al., 1993; Sweitzer and Hinshaw,1998). Given its homology with dynamin I, ADL6 may play arole at the Golgi apparatus, and possibly at the TGN, similarto that which dynamin I plays at the plasma membrane. Infact, some vacuolar cargo proteins have been shown toleave the TGN in clathrin-coated vesicles in plant cells (Hohlet al., 1996; Robinson et al., 1998).
To address this possibility, we examined the effect of adominant negative mutant of ADL6 on the vacuolar traffick-ing of cargo proteins in protoplasts, a strategy that hadalready been successful in revealing the role of human dy-namin I (Herskovits et al., 1993; Damke et al., 1994). Thedominant negative mutant of dynamin I causes inhibition ofendocytosis in animal cells. Similarly, in the presence ofADL6[K51E], we observed significant inhibition of the vacu-olar trafficking of two reporter proteins: sporamin:GFP, a re-porter protein transported to the lytic vacuole through theER and the Golgi apparatus (Matsuoka et al., 1995; Kim etal., 2001), and GFP:EBD, a reporter protein that is trans-
ported from the TGN to the lytic vacuole (Kim et al., 2001).At the same time, we observed a concomitant increase inthe punctate distribution of these reporters, which wasfound to represent the Golgi apparatus.
The accumulation of cargo proteins at the Golgi appara-tus was in good agreement with the proposed role of ADL6.In animal cells expressing the dominant negative mutant,transferrin accumulated in a punctate fashion at the plasmamembrane, as indicated by the fact that vesicles were notreleased from it (Herskovits et al., 1993; Damke et al., 1994).Similarly, vesicles may not be released from the Golgi appa-ratus in the presence of the dominant negative mutant ofADL6. The inhibition of both reporter proteins at the Golgiapparatus strongly suggests that ADL6 plays a role at theTGN and that the cargo proteins may have accumulated atthe TGN in the presence of the mutant protein. This conclu-sion is based on the fact that, unlike sporamin:GFP,GFP:EBD is expressed in the cytosol, is recruited to theTGN in a PI(3)P-dependent manner, and then is transportedto the central vacuole but is not transported through theGolgi apparatus (Kim et al., 2001).
In plant cells, there are other routes that vesicles derivedfrom the Golgi apparatus can take to transport cargo pro-teins. One example is the trafficking pathway from the Golgiapparatus to the plasma membrane (Hawes et al., 1999). Incontrast to vacuolar trafficking, ADL6[K51E] did not affectthe trafficking of H�-ATPase:GFP to the plasma membrane,suggesting that ADL6 is not involved in trafficking from theGolgi apparatus to the plasma membrane.
Recently, protein components involved in trafficking fromthe TGN to the lytic vacuole have been identified. These in-clude BP80, AtELP, AtVTI1a, AtPEP12p, AtVAMP3p, andAtVPS45 (Paris et al., 1997; Bassham and Raikhel, 1999;Zheng et al., 1999; Ahmed et al., 2000; Bassham et al., 2000).Among these proteins, AtELP, a putative cargo receptor,has been shown to be localized to the TGN and is thoughtto be involved in the vacuolar trafficking of cargo proteinsfrom the TGN (Ahmed et al., 2000). Thus, one possibility is
Figure 8. ADL6[K51E] Does Not Inhibit the Trafficking of H�-ATPase:GFP to the Plasma Membrane.
Protoplasts were transformed with H�-ATPase:GFP alone (a), H�-ATPase:GFP plus ADL6 (b), or H�-ATPase:GFP plus ADL6[K51E] (c), andgreen fluorescent signals of H�-ATPase:GFP were examined at various times. PM and CH indicate the plasma membrane and chloroplasts, re-spectively. Bars 25 �m.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

1522 The Plant Cell
that ADL6 may play a role in the formation of vesicles thatare induced by AtELP at the TGN. Further studies will benecessary to prove the exact relationship between AtELPand ADL6 during vacuolar trafficking in plant cells.
In this study, we used sporamin:GFP and GFP:EBD ascargo proteins for the in vivo trafficking assay in Arabidopsisprotoplasts. Sporamin, a storage protein found in the tubersof sweet potato, has been shown to be transported to thelytic vacuole in heterologous systems such as tobacco(Matsuoka et al., 1995; Kim et al., 2001). Also, GFP:EBD hasbeen shown to be transported to the lytic vacuole from theTGN in a PI(3)P-dependent manner in Arabidopsis (Kim etal., 2001). When we expressed the sporamin:GFP in Arabi-dopsis protoplasts, the targeting efficiency to the lytic vacu-ole was �50% at 40 hr after transformation. However, theefficiency was increased to 70 to 80% at 72 hr (data notshown). In contrast, the targeting efficiency of GFP:EBD andH�-ATPase:GFP was nearly 80 and 95%, respectively, at 24hr after transformation (data not shown for H�-ATPase:GFP),indicating that the targeting efficiency is dependent on thereporter proteins. One possible explanation for the slowertrafficking of sporamin is that protoplasts prepared from Ar-abidopsis leaf cells (a heterologous system for the expres-sion of sporamin) may have a limited supply of thenecessary machinery or of one of its components. In fact,the targeting efficiency was increased to nearly 70% at 12hr after transformation when Seh1h, a polypeptide that in-teracts with ADL6, was expressed in the protoplasts (S.H.Lee and I. Hwang, unpublished data).
METHODS
Growth of Plants
Arabidopsis thaliana (ecotype Columbia) was grown either on Murashigeand Skoog (1962) plates at 20°C in a culture room or in a greenhouseunder conditions of 70% RH and a 16-hr-light/8-hr-dark cycle.
Screening of a cDNA Encoding Dynamin-Like Protein
A probe for screening of cDNAs encoding dynamin-like proteins inArabidopsis was prepared by polymerase chain reaction (PCR) am-plification using primers designed from the nucleotide sequence in-formation deposited in the expressed sequence tag (EST) database(GenBank accession number W43823). The primers were as follows:ADL6-5, 5�-TGCTAGTCGCGATGGAG-3�; ADL6-3, 5�-CTCGTGTAG-CACCATTC-3�. PCR amplification and library screening were per-formed as described previously (Kang et al., 1998).
Preparation of Membrane Fractions
Protein extracts were prepared and fractionated by a Suc step gradi-ent according to Park et al. (1997). The presence of Arabidopsis dy-namin-like 6 (ADL6) in fractions collected from the gradient wasdetected using the polyclonal anti-ADL6 antibody.
Preparation of Antibody and Protein Gel Blot Analysis
For protein gel blot analysis, a polyclonal antibody was raisedagainst a truncated form of ADL6. The C-terminal region of ADL6(393 amino acid residues) was expressed in Escherichia coli as a re-combinant protein using the expression vector pRSET-B (Invitrogen,Carlsbad, CA). The recombinant protein was expressed and purifiedaccording to the manufacturer’s protocol. The purified protein wasinjected into a rabbit to raise antibody according to a published pro-tocol (Harlow and Lane, 1988). The polyclonal antibody was purifiedand used for protein gel blot analysis according to Park et al. (1997).The blots were developed using an enhanced chemiluminescencedetection system for protein gel blots (Amersham).
Immunocytohistochemistry
Cryosections were prepared from root tissues of Arabidopsis grownin liquid Murashige and Skoog (1962) medium as described previ-ously (Wick, 1993) and then immunolabeled as described previously(Gindullis and Meier, 1999). The root tissues also were treated withbrefeldin A (BFA) (10 �g/mL) before fixation. The purified anti-ADL6
Figure 9. Expression Levels of the Constructs in Protoplasts.
Total protein extracts were prepared from protoplasts transformed with various constructs and separated on an SDS-polyacrylamide gel for pro-tein gel blot analysis. The blots were probed with monoclonal anti-T7 (T7 mAb) and anti-GFP (GFP mAb) antibodies to detect the T7 epitope-tagged ADLs and GFP-tagged reporter proteins, respectively. The expected sizes of proteins are indicated.(A) C, 1, and 2 indicate untransformed protoplasts, protoplasts transformed with T7:ADL6 plus GFP:EBD, and protoplasts transformed withT7:ADL6[K51E] plus GFP:EBD, respectively.(B) C, 1, and 2 indicate untransformed protoplasts, protoplasts transformed with T7:ADL6 plus sporamin:GFP, and protoplasts transformed withT7:ADL6[K51E] plus sporamin:GFP, respectively.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

ADL6 in Trafficking from the TGN to the Vacuole 1523
antibody and fluorescein isothiocyanate (FITC)-labeled anti-rabbitIgG were used as primary and secondary antibodies, respectively.For double labeling, a mixture of a monoclonal antibody, JIM84 (Sa-tiat-Jeunemaitre and Hawes, 1992), and an anti-ADL6 antibody wasused as the primary antibody, and then the sections were stained se-quentially with FITC-labeled anti-rat IgG and tetrathylrhodamine-5-isothiocyanate–labeled anti-rabbit IgG. Images were observed us-ing a Zeiss (Jena, Germany) Axioplan fluorescence microscope.
Generation of Constructs
ADL6[K51E] was generated by PCR using primers 5�-CTGCAG-ACTCTCCAGCTCCCACATTTCC-3� and 5�-TGCTAGTCGCGATGG-AG-3�. The T7 epitope tag also was added to the N terminus ofADL6 by PCR using primers 5�-ACTGGTGGACAGCAAATGGGT-CGCGGATCCATGGAGGCGATCGATGAG-3� and 5�-AAAGAGTAA-AGAAGAACAATGGCTAGCATGACTGGTGGACAGCAAATG-3�. ThePCR products were confirmed by nucleotide sequencing. The ADL6constructs were placed under the control of the 35S cauliflower mo-saic virus promoter in a pUC vector. The ADL6:green fluorescentprotein (GFP) fusion construct was generated by placing the ADL6coding region in frame to the C terminus of the GFP coding regionwithout the termination codon.
In Vivo Targeting of Green and Red Fluorescent ProteinFusion Constructs
Plasmids were purified using Qiagen (Valencia, CA) columns accord-ing to the manufacturer’s protocol. The fusion constructs were intro-duced into Arabidopsis protoplasts prepared from whole seedlingsby polyethylene glycol–mediated transformation (Kang et al., 1998).Briefly, leaf tissues (5 g) of 3- to 4-week-old Arabidopsis plantsgrown on soil in a greenhouse were cut into small squares (5 to 10mm2) with a new razor blade and incubated with 50 mL of enzymesolution (0.25% Macerozyme (Yakult Honsha Co., Ltd., Tokyo, Japan)R-10, 1.0% Cellulase (Yakult Honsha Co., Ltd.) R-10, 400 mM man-nitol, 8 mM CaCl2, and 5 mM Mes-KOH, pH 5.6) at 22°C for 5 hr withgentle agitation (50 to 75 rpm). After incubation, the protoplast sus-pension was filtered through 100-�m mesh and protoplasts werecollected by centrifugation at 46g for 5 min. The pelleted protoplastswere resuspended in 5 to 10 mL of W5 solution (154 mM NaCl, 125mM CaCl2, 5 mM KCl, 5 mM glucose, and 1.5 mM Mes-KOH, pH5.6), overlaid on top of 20 mL of 21% Suc, and centrifuged for 10 minat 78g. The intact protoplasts at the interface were transferred to anew Falcon tube containing 20 mL of W5 solution. The protoplastswere pelleted again by centrifugation at 55g for 5 min and resus-pended in 20 mL of W5 solution. The protoplasts were incubated onice for 30 min.
To transform DNA into protoplasts, protoplasts were pelletedagain at 46g for 5 min and resuspended in MaMg solution (400 mMmannitol, 15 mM MgCl2, and 5 mM Mes-KOH, pH 5.6) at a density of5 � 106 protoplasts/mL. Plasmid DNA (20 to 50 �g total at a concen-tration of 2 mg/mL) was added to 300 �L of protoplast suspensionfollowed by 325 �L of PEG solution [400 mM mannitol, 100 mMCa(NO3)2, and 40% polyethylene glycol 4000]. The mixture wasmixed gently and incubated for 30 min at room temperature. Afterincubation, the mixture was diluted with 10 mL of W5 solution. Pro-toplasts were recovered by centrifugation at 50g for 5 min, resus-pended in 3 mL of W5 solution, and incubated at 22°C in the dark.Expression of protein was monitored at various times after transfor-
mation, and images were captured with a cooled charge-coupleddevice camera using a Zeiss Axioplan fluorescence microscope. The fil-ter sets used were XF116 (exciter, 474AF20; dichroic, 500DRLP; emit-ter, 510AF23), XF33/E (exciter, 535DF35; dichroic, 570DRLP;emitter, 605DF50), and XF137 (exciter, 540AF30; dichroic, 570DRLP;emitter, 585ALP) (Omega, Inc., Brattleboro, VT) for green fluores-cent protein, red fluorescent protein, and autofluorescence of chlo-rophyll, respectively. Data were then processed using Adobe(Mountain View, CA) Photoshop software, and the images were ren-dered in pseudocolor.
ACKNOWLEDGMENTS
We thank C. Hawes (Oxford Brookes University, Oxford, UK) for themonoclonal antibody JIM84 and for the personal communication re-garding the localization of the epitope detected by JIM84. Also, wethank Dr. Soon-Ok Eun (Pohang University of Science and Technol-ogy, Pohang, Korea) for technical help in immunohistochemicallabeling. This work was supported by a grant from the National Cre-ative Research Initiatives of the Ministry of Science and Technologyin Korea.
Received December 15, 2000; accepted April 29, 2001.
REFERENCES
Ahmed, S.U., Rojo, E., Kovaleva, V., Venkataraman, S.,Dombrowski, J.E., Matsuoka, K., and Raikhel, N.V. (2000). Theplant vacuolar sorting receptor AtELP is involved in transport ofNH(2)-terminal propeptide-containing vacuolar proteins in Arabi-dopsis thaliana. J. Cell Biol. 149, 1335–1344.
Baba, M., Osumi, M., Scott, S.V., Klionsky, D.J., and Ohsumi, Y.(1997). Two distinct pathways for targeting proteins from the cyto-plasm to the vacuole/lysosome. J. Cell Biol. 139, 1687–1695.
Bassham, D.C., and Raikhel, N.V. (1999). The pre-vacuolar t-SNAREAtPEP12p forms a 20S complex that dissociates in the presenceof ATP. Plant J. 19, 599–603.
Bassham, D.C., and Raikhel, N.V. (2000). Unique features of the plantvacuolar sorting machinery. Curr. Opin. Cell Biol. 12, 491–495.
Bassham, D.C., Gal, S., da Silva Conceicao, A., and Raikhel, N.V.(1995). An Arabidopsis syntaxin homologue isolated by functionalcomplementation of a yeast pep12 mutant. Proc. Natl. Acad. Sci.USA 92, 7262–7266.
Bassham, D.C., Sanderfoot, A.A., Kovaleva, V., Zheng, H., andRaikhel, N.V. (2000). AtVPS45 complex formation at the trans-Golgi network. Mol. Biol. Cell. 11, 2251–2265.
Bensen, E.S., Costaguta, G., and Payne, G.S. (2000). Syntheticgenetic interactions with temperature-sensitive clathrin in Saccha-romyces cerevisiae: Roles for synaptojanin-like Inp53p anddynamin-related Vps1p in clathrin-dependent protein sorting atthe trans-Golgi network. Genetics 154, 83–97.
Blackbourn, H.D., and Jackson, A.P. (1996). Plant clathrin heavychain: Sequence analysis and restricted localisation in growingpollen tubes. J. Cell Sci. 109, 777–786.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

1524 The Plant Cell
Cao, H., Garcia, F., and McNiven, M.A. (1998). Differential distribu-tion of dynamin isoforms in mammalian cells. Mol. Biol. Cell 9,2595–2609.
Chen, M.S., Obar, R.A., Schroeder, C.C., Austin, T.W., Poodry,C.A., Wadsworth, S.C., and Vallee, R.B. (1991). Multiple forms ofdynamin are encoded by shibire, a Drosophila gene involved inendocytosis. Nature 351, 583–586.
Damke, H., Baba, T., Warnock, D.E., and Schmid, S.L. (1994).Induction of mutant dynamin specifically blocks endocytic coatedvesicle formation. J. Cell Biol. 127, 915–934.
Dombrowski, J.E., and Raikhel, N.V. (1995). Isolation of a cDNAencoding a novel GTP-binding protein of Arabidopsis thaliana.Plant Mol. Biol. 28, 1121–1126.
Driouich, A., Zhang, G.F., and Staehelin, L.A. (1993). Effect ofbrefeldin A on the structure of the Golgi apparatus and on thesynthesis and secretion of proteins and polysaccharides insycamore maple (Acer pseudoplatanus) suspension-culturedcells. Plant Physiol. 101, 1363–1373.
Fitchette, A.C., Cabanes-Macheteau, M., Marvin, L., Martin, B.,Satiat-Jeunemaitre, B., Gomord, V., Crooks, K., Lerouge, P.,Faye, L., and Hawes, C. (1999). Biosynthesis and immunolocal-ization of Lewis a-containing N-glycans in the plant cell. PlantPhysiol. 121, 333–344.
Frigerio, L., de Virgilio, M., Prada, A., Faoro, F., and Vitale, A.(1998). Sorting of phaseolin to the vacuole is saturable andrequires a short C-terminal peptide. Plant Cell 10, 1031–1042.
Fujiwara, T., Oda, K., Yokota, S., Takatsuki, A., and Ikehara, Y.(1988). Brefeldin A causes disassembly of the Golgi complex andaccumulation of secretory proteins in the endoplasmic reticulum.J. Biol. Chem. 263, 18545–18552.
Gindullis, F., and Meier, I. (1999). Matrix attachment region bindingprotein MFP1 is localized in discrete domains at the nuclear enve-lope. Plant Cell 11, 1117–1128.
Gout, I., Dhand, R., Hiles, I.D., Fry, M.J., Panayotou, G., Das, P.,Truong, O., Totty, N.F., Husan, J., Booker, G.W., Campbell,I.D., and Waterfield, M.D. (1993). The GTPase dynamin binds toand is activated by a subset of SH3 domains. Cell 75, 25–36.
Gu, X., and Verma, D.P.S. (1996). Phragmoplastin, a dynamin-likeprotein associated with cell plate formation in plants. EMBO J. 15,695–704.
Gu, X., and Verma, D.P. (1997). Dynamics of phragmoplastin in liv-ing cells during cell plate formation and uncoupling of cell elonga-tion from the plane of cell division. Plant Cell 9, 157–169.
Hara-Nishimura, I., Shimada, T., Hatano, K., Takeuchi, Y., andNishimura, M. (1998). Transport of storage proteins to proteinstorage vacuoles is mediated by large precursor-accumulatingvesicles. Plant Cell 10, 825–836.
Harlow, E., and Lane, D. (1988). Antibodies: A Laboratory Manual.(Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).
Hattori, T., Yoshida, N., and Nakamura, K. (1989). Structural rela-tionship among the members of a multigene family coding for thesweet potato tuberous root storage protein. Plant Mol. Biol. 13,563–572.
Hawes, C.R., Brandizzi, F., and Andreeva, A.V. (1999). Endomem-branes and vesicle trafficking. Curr. Opin. Plant Biol. 2, 454–461.
Henley, J.R., and McNiven, M.A. (1996). Association of a dynamin-
like protein with the Golgi apparatus in mammalian cells. J. CellBiol. 133, 761–775.
Herskovits, J.S., Burgess, C.C., Obar, R.A., and Vallee, R.B.(1993). Effects of mutant rat dynamin on endocytosis. J. Cell Biol.122, 565–578.
Hillmer, S., Movafeghi, A., Robinson, D.G., and Hinz, G. (2001).Vacuolar storage proteins are sorted in the cis-cisternae of thepea cotyledon Golgi apparatus. J. Cell Biol. 152, 41–50.
Hinshaw, J.E., and Schmid, S.L. (1995). Dynamin self-assemblesinto rings suggesting a mechanism for coated vesicle budding.Nature 374, 190–192.
Hohl, I., Robinson, D.G., Chrispeels, M.J., and Hinz, G. (1996).Transport of storage proteins to the vacuole is mediated by vesi-cles without a clathrin coat. J. Cell Sci. 109, 2539–2550.
Jahn, R., and Südhof, T.C. (1999). Membrane fusion and exocyto-sis. Annu. Rev. Biochem. 68, 863–911.
Jiang, L., Phillips, T.E., Rogers, S.W., and Rogers, J.C. (2000).Biogenesis of the protein storage vacuole crystalloid. J. Cell Biol.150, 755–770.
Jones, S.M., and Howell, K.E. (1997). Phosphatidylinositol 3-kinaseis required for the formation of constitutive transport vesicles fromthe TGN. J. Cell Biol. 139, 339–349.
Jones, S.M., Howell, K.E., Henley, J.R., Cao, H. and McNiven,M.A. (1998). Role of dynamin in the formation of transport vesiclesfrom the trans-Golgi network. Science 279, 573–577.
Kang, S.G., Jin, J.B., Piao, H.L., Pih, K.T., Jang, H.J., Lim, J.H.,and Hwang, I. (1998). Molecular cloning of an Arabidopsis cDNAencoding a dynamin-like protein that is localized to plastids. PlantMol. Biol. 38, 437–447.
Kim, D.H., Eu, Y.-J., Yoo, C.M., Kim, Y.W., Pih, K.T., Jin, J.B., Kim,S.J., Stenmark, H., and Hwang, I. (2001). Trafficking of phosphati-dylinositol 3-phosphate from the trans-Golgi network to the lumenof the central vacuole in plant cells. Plant Cell 13, 287–301.
Lebas, M., and Axelos, M. (1994). A cDNA encoding a new GTP-binding protein of the ADP-ribosylation factor family from Arabi-dopsis. Plant Physiol. 106, 809–810.
Maier, O., Knoblich, M., and Westermann, P. (1996). Dynamin IIbinds to the trans-Golgi network. Biochem. Biophys. Res. Com-mun. 223, 229–233.
Marks, B., Stowell, M.H., Vallis, Y., Mills, I.G., Gibson, A., Hopkins,C.R., and McMahon, H.T. (2001). GTPase activity of dynaminand resulting conformation change are essential for endocytosis.Nature 410, 231–235.
Matsuoka, K., Watanabe, N., and Nakamura, K. (1995). O-Glyco-sylation of a precursor to a sweet potato vacuolar protein,sporamin, expressed in tobacco cells. Plant J. 8, 877–889.
Misumi, Y., Misumi, Y., Miki, K., Takatsuki, A., Tamura, G., andIkehara, Y. (1986). Novel blockade by brefeldin A of intracellulartransport of secretory proteins in cultured rat hepatocytes. J. Biol.Chem. 261, 11398–11403.
Movafeghi, A., Happel, N., Pimpl, P., Tai, G.H., and Robinson,D.G. (1999). Arabidopsis Sec21p and Sec23p homologs: Proba-ble coat proteins of plant COP-coated vesicles. Plant Physiol.119, 1437–1446.
Murashige, T., and Skoog, F. (1962). A revised medium for rapidgrowth and bioassays with tobacco tissue culture. Physiol. Plant.15, 473–497.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022

ADL6 in Trafficking from the TGN to the Vacuole 1525
Neuhaus, J.M., and Rogers, J.C. (1998). Sorting of proteins to vac-uoles in plant cells. Plant Mol. Biol. 38, 127–144.
Obar, R.A., Collins, C.A., Hammarback, J.A., Shpetner, H.S., andVallee, R.B. (1990). Molecular cloning of the microtubule-associ-ated mechanochemical enzyme dynamin reveals homology with anew family of GTP-binding proteins. Nature 347, 256–261.
Paris, N., Rogers, S.W., Jiang, L., Kirsch, T., Beevers, L., Phillips,T.E., and Rogers, J.C. (1997). Molecular cloning and further char-acterization of a probable plant vacuolar sorting receptor. PlantPhysiol. 115, 29–39.
Park, J.M., Kang, S.G., Pih, K.T., Jang, H.J., Piao, H.L., Yoon,H.W., Cho, M.J., and Hwang, I. (1997). A dynamin-like protein,ADL1, is present in membranes as a high-molecular-mass com-plex in Arabidopsis thaliana. Plant Physiol. 115, 763–771.
Park, J.M., Cho, J.H., Kang, S.G., Jang, H.J., Pih, K.T., Piao, H.L.,Cho, M.J., and Hwang, I. (1998). A dynamin-like protein in Arabi-dopsis thaliana is involved in biogenesis of thylakoid membranes.EMBO J. 17, 859–867.
Pimpl, P., Movafeghi, A., Coughlan, S., Denecke, J., Hillmer, S.,and Robinson, D.G. (2000). In situ localization and in vitro induc-tion of plant COPI-coated vesicles. Plant Cell 12, 2219–2236.
Robinson, D.G., Hinz, G., and Holstein, S.E. (1998). The molecularcharacterization of transport vesicles. Plant Mol. Biol. 38, 49–76.
Robinson, P.J., Sontag, J.-M., Liu, J.-P., Fykse, E.S., Slaughter,C., McMahon, H., and Sudhof, T.C. (1993). Dynamin GTPaseregulated by protein kinase C phosphorylation in nerve terminals.Nature 365, 163–166.
Rothman, J.E. (1994). Mechanisms of intracellular protein import.Nature 372, 55–63.
Rothman, J.H., Raymond, C.K., Gilbert, T., O’Hara, P.J., andStevens, T.H. (1990). A putative GTP binding protein homologousto interferon-inducible Mx proteins performs an essential functionin yeast protein sorting. Cell 61, 1063–1074.
Sanderfoot, A.A., Assaad, F.F., and Raikhel, N.V. (2000). The Ara-bidopsis genome: An abundance of soluble N-ethylmaleimide-sensitive factor adaptor protein receptors. Plant Physiol. 124,1558–1569.
Satiat-Jeunemaitre, B., and Hawes, C. (1992). Redistribution of aGolgi glycoprotein in plant cells treated with brefeldin A. J. CellSci. 103, 1153–1166.
Schu, P.V., Takegawa, K., Fry, M.J., Stack, J.H., Waterfield, M.D.,
and Emr, S.D. (1993). Phosphatidylinositol 3-kinase encoded byyeast VPS34 gene essential for protein sorting. Science 260, 88–91.
Sever, S., Muhlberg, A.B., and Schmid, S.L. (1999). Impairment ofdynamin’s GAP domain stimulates receptor-mediated endocyto-sis. Nature 398, 481–486.
Stenmark, H., Aasland, R., Toh, B.H., and D’Arrigo, A. (1996).Endosomal localization of the autoantigen EEA1 is mediated by azinc-binding FYVE finger. J. Biol. Chem. 271, 24048–24054.
Sweitzer, S.M., and Hinshaw, J.E. (1998). Dynamin undergoes aGTP-dependent conformational change causing vesiculation. Cell93, 1021–1029.
Takeuchi, M., Tada, M., Saito, C., Yashiroda, H., and Nakano, A.(1998). Isolation of a tobacco cDNA encoding Sar1 GTPase andanalysis of its dominant mutations in vesicular traffic using a yeastcomplementation system. Plant Cell Physiol. 39, 590–599.
Takeuchi, M., Ueda, T., Sato, K., Abe, H., Nagata, T., andNakano, A. (2000). A dominant negative mutant of sar1 GTPaseinhibits protein transport from the endoplasmic reticulum to theGolgi apparatus in tobacco and Arabidopsis cultured cells. PlantJ. 23, 517–525.
Toyooka, K., Okamoto, T., and Minamikawa, T. (2000). Masstransport of proform of a KDEL-tailed cysteine proteinase (SH-EP)to protein storage vacuoles by endoplasmic reticulum-derivedvesicle is involved in protein mobilization in germinating seeds. J.Cell Biol. 148, 453–464.
Tuma, P.L., Stachniak, M.C., and Collins, C.A. (1993). Activationof dynamin GTPase by acidic phospholipids and endogenous ratbrain vesicles. J. Biol. Chem. 268, 17240–17246.
Wee, E.G., Sherrier, D.J., Prime, T.A., and Dupree, P. (1998). Tar-geting of active sialyltransferase to the plant Golgi apparatus.Plant Cell 10, 1759–1768.
Wick, S.M. (1993). Immunolabeling of antigens in plant cells. InMethods in Cell Biology, Vol. 37, Antibodies in Cell Biology, D.J.Asai, ed (San Diego: Academic Press), pp. 171–200.
Wilsbach, K., and Payne, G.S. (1993). Vps1p, a member of thedynamin GTPase family, is necessary for Golgi membrane proteinretention in Saccharomyces cerevisiae. EMBO J. 12, 3049–3059.
Zheng, H., von Mollard, G.F., Kovaleva, V., Stevens, T.H., andRaikhel, N.V. (1999). The plant vesicle-associated SNAREAtVTI1a likely mediates vesicle transport from the trans-Golgi net-work to the prevacuolar compartment. Mol. Biol. Cell 10, 2251–2264.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/13/7/1511/6009548 by guest on 04 February 2022