a myb/zml complex regulates wound-induced lignin genes … · a myb/zml complex regulates...
TRANSCRIPT

A MYB/ZML Complex Regulates Wound-Induced LigninGenes in Maize
Isabel-Cristina Vélez-Bermúdez,a,b,1 Jorge E. Salazar-Henao,a,b,1 Silvia Fornalé,a Irene López-Vidriero,c
José-Manuel Franco-Zorrilla,c Erich Grotewold,d John Gray,e Roberto Solano,f Wolfgang Schmidt,b
Montserrat Pagés,a Marta Riera,a,2 and David Caparros-Ruiza,2
a Centre de Recerca en Agrigenòmica, Consortium CSIC-IRTA-UAB-UB, Cerdanyola del Vallès, 08193 Barcelona, Spainb Institute of Plant and Microbial Biology, Academia Sinica, 11529 Taipei, TaiwancGenomics Unit, Centro Nacional de Biotecnología, Consejo Superior de Investigaciones Científicas, 28049 Madrid, SpaindCenter for Applied Plant Sciences and Department of Molecular Genetics, The Ohio State University, Columbus, Ohio 43210eDepartment of Biological Sciences, University of Toledo, Toledo, Ohio 43606f Department of Plant Molecular Genetics, Centro Nacional de Biotecnología, Consejo Superior de Investigaciones Científicas, 28049Madrid, Spain
ORCID IDs: 0000-0001-7586-8830 (S.F.); 0000-0001-9362-8662 (I.L.-V.); 0000-0003-1352-2900 (J.G.); 0000-0002-7850-6832 (W.S.);0000-0001-8431-5605 (M.P.); 0000-0003-1007-5324 (M.R.); 0000-0002-7461-8888 (D.C.-R.)
Lignin is an essential polymer in vascular plants that plays key structural roles in vessels and fibers. Lignification is induced byexternal inputs such as wounding, but the molecular mechanisms that link this stress to lignification remain largely unknown.In this work, we provide evidence that three maize (Zea mays) lignin repressors, MYB11, MYB31, and MYB42, participate inwound-induced lignification by interacting with ZML2, a protein belonging to the TIFY family. We determined that the threeR2R3-MYB factors and ZML2 bind in vivo to AC-rich and GAT(A/C) cis-elements, respectively, present in a set of lignin genes.In particular, we show that MYB11 and ZML2 bind simultaneously to the AC-rich and GAT(A/C) cis-elements present in thepromoter of the caffeic acid O-methyl transferase (comt) gene. We show that, like the R2R3-MYB factors, ZML2 also acts asa transcriptional repressor. We found that upon wounding and methyl jasmonate treatments, MYB11 and ZML2 proteins aredegraded and comt transcription is induced. Based on these results, we propose a molecular regulatory mechanism involvinga MYB/ZML complex in which wound-induced lignification can be achieved by the derepression of a set of lignin genes.
INTRODUCTION
Lignin is essential in vascular plants and its deposition in thesecondary cell walls improves the hydrophobicity of vessels,reinforcesthefibers,andactsasaphysicalbarrieragainstpathogenattacks (Sarkanen and Ludwig, 1971; Carpita and Gibeaut, 1993;Jouanin et al., 2000; Boerjan et al., 2003). Several studies haveidentified different regulators of lignin biosynthesis in grasses (re-viewed in Gray et al., 2012), such as maize (Zea mays) MYB31 andMYB42, two subgroup 4 R2R3-MYB transcription factors that actas repressors of the lignin pathway (Fornalé et al., 2006, 2010;Sonbol et al., 2009). Despite their different roles in lignin regulation,both factors repress the maize caffeic acid O-methyl transferase(comt) gene (AC196475). Mutations in this gene have been shownto produce the brown midrib3 phenotype (Vignols et al., 1995;Fornaléetal., 2006, 2010). Theexpressionofmaizecomt is inducedby wounding (Capellades et al., 1996) through a mechanism thatremains unknown, while it has been demonstrated that the wound
induction of the Arabidopsis thaliana COMT is CORONATINE-INSENSITIVEPROTEIN1COI1 dependent (Reymond et al., 2000).Plants respond to wounding by triggering the induction of lig-
nification in tissues that normally do not accumulate this polymer(Vance et al., 1980; Lawton and Lamb, 1987; Rogers andCampbell, 2004). Consequently, several genes of the ligninpathway are induced by wounding and/or methyl jasmonate(MeJA) (Ellard-Ivey and Douglas, 1996; Bell-Lelong et al., 1997;Mizutani et al., 1997; Ehlting et al., 1999; Reymond et al., 2000;Devoto et al., 2005;Pauwels et al., 2008). In addition, the inductionof MeJA cascade has been positively correlated with an increasein lignin content in several species, such as Bryonia dioica (Kaiseret al., 1994), Cassia tora (Xue et al., 2008), and rice (Oryza sativa;Tianpei et al., 2015).One of the most important groups of proteins involved in
wounding/MeJA responses is the TIFY family (Vanholme et al.,2007). TIFYs comprise four subgroups of proteins: ZML, TIFY,PPD, and JASMONATE ZIM domain (JAZ) proteins. JAZ proteinsare coreceptors of the hormone and targets of the SCF(COI1)
complex and function as negative regulators of the jasmonatesignaling pathway (Chini et al., 2007; Thines et al., 2007). Sincetheir discovery in Arabidopsis, JAZ proteins have been implicatedin protein-protein interactions with multiple transcription factors,such as the basic helix-loop-helix factor MYC2 (Lorenzo et al.,2004; Chini et al., 2007). More recent studies in Arabidopsis haveshown that the activity of someR2R3-MYB transcription factors isregulated through their interaction with JAZ proteins. UponMeJA
1 These authors contributed equally to this work.2 Address correspondence to [email protected] or [email protected] authors responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy describedin the Instructions for Authors (www.plantcell.org) are: Erich Grotewold([email protected]) and David Caparros-Ruiz ([email protected]).www.plantcell.org/cgi/doi/10.1105/tpc.15.00545
The Plant Cell, Vol. 27: 3245–3259, November 2015, www.plantcell.org ã 2015 American Society of Plant Biologists. All rights reserved.

treatment, R2R3-MYBs are released by the MeJA-induced deg-radation of the JAZ proteins, and the target genes are sub-sequently activated (Qi et al., 2011; Song et al., 2011). Within theZML subfamily, Arabidopsis ZML1 and ZML2 can bind DNA,suggesting that these proteins can act as transcriptional regu-lators (Shaikhali et al., 2012). How ZML factors cooperate withother regulators to activate or repress gene expression and towhatextentproteindegradation ispartof the regulatory functionofZML factors remain unknown.
Here,wepresentaregulatorymechanismbywhichtheexpressionof lignin genes can be induced upon wounding/MeJA by the actionof MYB/ZML transcriptional repressors in maize. In particular, weshowed that the jasmonate signaling cascade in maize triggers thedegradationofMYB11andZML2, leading to thederepressionof thelignin comt gene. The regulatory mechanism described for MYB11can be extended to the lignin repressors MYB31 and MYB42. Ourdata further show that theseMYB/ZML proteins also regulate othergenes of the lignin biosynthetic pathway.
RESULTS
Maize MYB11 Binds to the comt Promoter in Vivo andRepresses Its Expression
Based on sequence similarity with MYB31 and MYB42, twotranscriptional repressors of comt (Fornalé et al., 2006, 2010), weidentified myb11 (GRMZM2G000818) as a gene encodinga subgroup 4 R2R3-MYB factor (Supplemental Figure 1). Geneexpression analyses revealed thatmyb11 is expressed at variousstages of maize development, both in roots and in the aerial partsof the plant. In addition, myb11 is induced 1 h after wounding(Supplemental Figure 1).
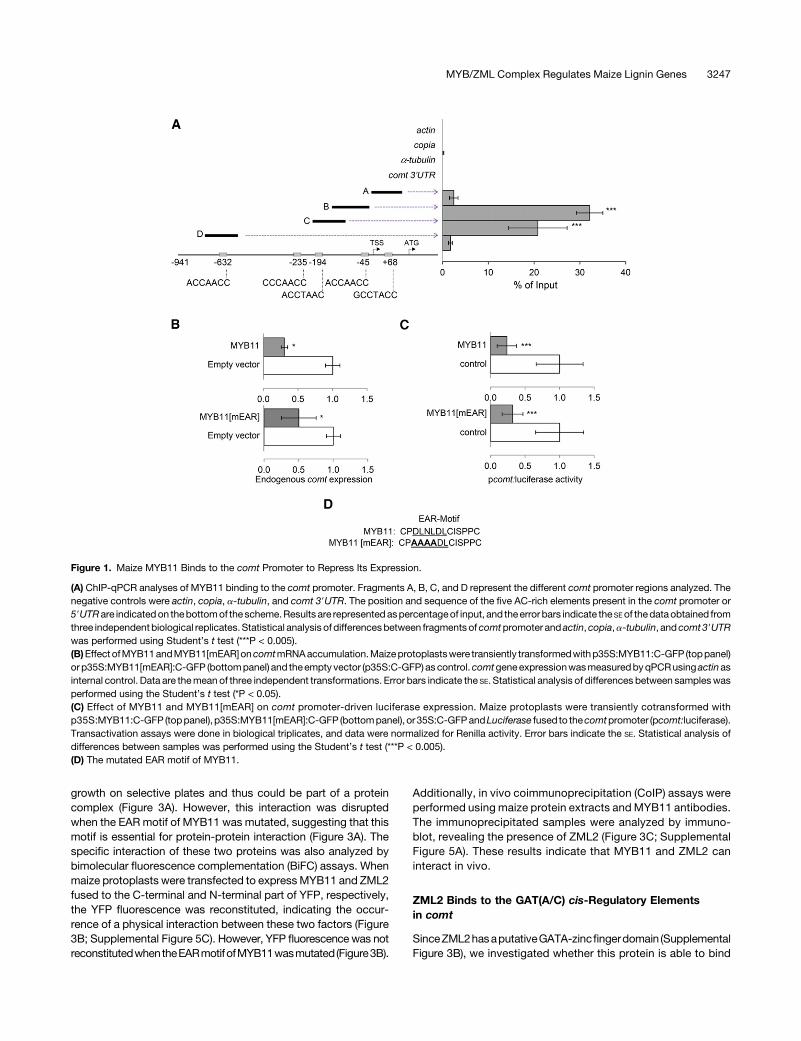
MYB31 and MYB42 repress comt expression in vivo (Fornaléet al., 2006, 2010). In the case of MYB31, this repression occursthrough its binding to a typical AC-rich MYB motif present in thecomt promoter (Fornalé et al., 2006). Therefore, we raised specificantibodies against MYB11 (Supplemental Figure 2) to investigatewhether this factor is also able to bind to comt by chromatinimmunoprecipitation (ChIP) experiments using actin, copia, anda-tubulin as negative controls. Our results demonstrate thatMYB11 binds in vivo to the comt gene promoter, preferentially tosequences within the first 250 bp from the transcription start sitecontaining the AC-rich MYB binding motifs (Figure 1A).
To elucidate whether MYB11 regulates the expression of comt,maize protoplasts were transformed to transiently express theMYB11protein fused toGFP. The subsequent qRT-PCRanalysesshowed that comtmRNA accumulation was reduced (Figure 1B).Similar repression was observed when maize protoplasts werecotransformed with MYB11-GFP and the luciferase gene underthe control of the comt promoter (pcomt:luciferase) (Figure 1C).
MYB11 belongs to subgroup 4 of the R2R3-MYB family, whosecharacteristic feature is the presence of an ethylene-responsiveelement binding factor-associated amphiphilic repression (EAR)motif in the C-terminal portion of the protein (Kagale andRozwadowski,2011).Toestablishwhether theEARmotif isessentialfor MYB11 repression activity, we mutated this motif (MYB11-mEAR, Figure 1D) and investigated the capacity of the mutated
protein to repress the expression of comt in maize protoplasts.Analyses of endogenous comt and pcomt:luciferase expressionindicate that MYB11-mEAR maintains its capacity to represscomt expression (Figures 1B and 1C), suggesting that this mu-tated EAR motif does not significantly affect the transcriptionalrepression activity of MYB11.
MYB11 and ZML2 Are Degraded upon Wounding
AsMYB11 represses comt gene expression by directly binding itspromoter andcomt is also inducedbywounding (Capelladesetal.,1996; Fornalé et al., 2006), we investigated whether the mainfunction of MYB11 could be associated with the regulation oflignin biosynthesis in response to this stress.Some proteins belonging to the TIFY family can interact with
R2R3-MYB factors tomodulate their function (Qi et al., 2011;Songet al., 2011). As members of the TIFY family regulate wounding/MeJA responses (Vanholme et al., 2007), we investigatedwhethermaize TIFY proteins can regulate the function of MYB11 uponwounding. Within the maize TIFY family, we found three proteinsthat belong to theZMLsubfamily, ZML1, ZML2, andML3 (Bai et al.,2011) (SupplementalFigure3A).Wesurveyedtheexpressionprofileof each of the zml genes using the qTeller bioinformatics tool andfocused on zml2 (GRMZM2G058479), as it was the one with themost similar gene expression pattern to myb11 (SupplementalFigure 3C) (Sekhon et al., 2013). In addition, as ChIP-qPCR withMYB11 was done in 9-d-old maize samples, we also tested theexpression of these three genes in this developmental stage. Ourresults indicate that zml2 is the main (if not the only) zml geneexpressed at this stage (Supplemental Figure 3D).We raised specific antibodies against ZML2 (Supplemental
Figure 4) and investigated whether MYB11 and ZML2 proteinlevels are affected by wounding or MeJA treatment in 9-d-oldmaize leaves. Immunoblot analysis showed that MYB11 andZML2 levels decreased 1 h after treatments (Figure 2A). However,the decrease observed after MeJA treatment was not due toa reduced mRNA accumulation of these two genes (Figure 2B),indicating that MeJA may trigger the degradation of MYB11 andZML2. Therefore, we transiently expressed MYB11 and ZML2fused to GFP in maize protoplasts (MYB11-GFP and ZML2-GFP,respectively) andGFPaloneasacontrol (Figure 2C;SupplementalFigure 5B). The fluorescence from these two fusion proteins wasseverelydecreasedafterMeJA treatment,and thisdecreasedidnotoccur in the presence of the proteasome inhibitor MG132 (Figure2C). These results suggest that thedecrease in the accumulationofMYB11 and ZML2 is due to degradation, likely mediated by theproteasome. Interestingly, when we assayed MYB11 with themutated EAR domain fused to GFP (MYB11-mEAR-GFP) in maizeprotoplasts, we found that the MeJA-induced degradation wasabolished, suggesting that the EARmotif plays an important role inthe proteasomal degradation of MYB11 (Figure 2C).
MYB11 Interacts with ZML2
Since myb11 and zml2 have similar expression patterns, andMeJApromotes the degradation of both proteins, we investigatedwhether they could physically interact in yeast two-hybrid matingassays. MYB11 and ZML2 proteins interacted to allow yeast
3246 The Plant Cell
growth on selective plates and thus could be part of a proteincomplex (Figure 3A). However, this interaction was disruptedwhen the EAR motif of MYB11 was mutated, suggesting that thismotif is essential for protein-protein interaction (Figure 3A). Thespecific interaction of these two proteins was also analyzed bybimolecular fluorescence complementation (BiFC) assays. Whenmaize protoplasts were transfected to expressMYB11 and ZML2fused to the C-terminal and N-terminal part of YFP, respectively,the YFP fluorescence was reconstituted, indicating the occur-rence of a physical interaction between these two factors (Figure3B; Supplemental Figure 5C). However, YFP fluorescence was notreconstitutedwhentheEARmotifofMYB11wasmutated (Figure3B).
Additionally, in vivo coimmunoprecipitation (CoIP) assays wereperformed using maize protein extracts and MYB11 antibodies.The immunoprecipitated samples were analyzed by immuno-blot, revealing the presence of ZML2 (Figure 3C; SupplementalFigure 5A). These results indicate that MYB11 and ZML2 caninteract in vivo.
ZML2 Binds to the GAT(A/C) cis-Regulatory Elementsin comt
SinceZML2hasaputativeGATA-zincfingerdomain (SupplementalFigure 3B), we investigated whether this protein is able to bind
Figure 1. Maize MYB11 Binds to the comt Promoter to Repress Its Expression.
(A) ChIP-qPCR analyses of MYB11 binding to the comt promoter. Fragments A, B, C, and D represent the different comt promoter regions analyzed. Thenegative controls were actin, copia, a-tubulin, and comt 39UTR. The position and sequence of the five AC-rich elements present in the comt promoter or59UTRare indicatedon thebottomof the scheme.Results are representedaspercentageof input, and theerror bars indicate the SEof thedataobtained fromthree independentbiological replicates. Statistical analysis of differencesbetween fragments ofcomtpromoter and actin,copia,a-tubulin, andcomt39UTRwas performed using Student’s t test (***P < 0.005).(B)Effect ofMYB11andMYB11[mEAR]oncomtmRNAaccumulation.Maizeprotoplastswere transiently transformedwithp35S:MYB11:C-GFP (toppanel)or p35S:MYB11[mEAR]:C-GFP (bottompanel) and theempty vector (p35S:C-GFP) ascontrol.comtgeneexpressionwasmeasuredbyqPCRusing actinasinternal control. Data are themean of three independent transformations. Error bars indicate the SE. Statistical analysis of differences between sampleswasperformed using the Student’s t test (*P < 0.05).(C) Effect of MYB11 and MYB11[mEAR] on comt promoter-driven luciferase expression. Maize protoplasts were transiently cotransformed withp35S:MYB11:C-GFP (toppanel), p35S:MYB11[mEAR]:C-GFP (bottompanel), or35S:C-GFPandLuciferase fused to thecomtpromoter (pcomt:luciferase).Transactivation assays were done in biological triplicates, and data were normalized for Renilla activity. Error bars indicate the SE. Statistical analysis ofdifferences between samples was performed using the Student’s t test (***P < 0.005).(D) The mutated EAR motif of MYB11.
MYB/ZML Complex Regulates Maize Lignin Genes 3247

DNA. Using an in vitro protein binding microarray (PBM) assaycontaining all possible double-stranded 6-mers (Godoy et al.,2011), we determined the DNA binding specificity of ZML2. Weobtainedsignificantdifferences for the sequencescontaining theconsensus sequence GAT(A/C). The highest signal intensitiesand E-scores were obtained for the core 59-GATC-39, a mediumto low affinity was observed for the 59-GATA-39 element, and no
bindingaffinitywasobserved for an irrelevant element (59-GTAC-39) (Figure 4A).The maize comt promoter contains two GATA cis-elements
(Figure 4B). Thus, we investigated whether ZML2 can binddirectly to oneor both of these elements in this promoter in vivo.ChIP assays showed that ZML2 binds to the comt gene pro-moter in a region containing a GATA cis-element (fragments B
Figure 2. MYB11 and ZML2 Proteins Are Degraded by Wounding and MeJA.
(A) Immunodetection of MYB11 and ZML2 in protein extracts from 9-d-old maize leaves using anti-MYB11 and anti-ZML2 antibodies. Protein levels weredetermined 1 h after wounding (W, wounded samples; C, control samples) and after 1 h of 0.01% MeJA treatments using 0.01% DMSO as a control (C).b-Actin was used as protein loading control.(B)qRT-PCRanalysis ofmyb11and zml2expression in 9-d-oldmaize leaves fromplants treated for 1hwith25µMMeJAor 0.01%DMSO.actinwasusedasinternal control gene.White and gray bars refer to control andMeJA-treated samples, respectively. Error bars indicate the SE of the data obtained from threeindependent biological replicates.(C)Effect ofMeJAonMYB11 andZML2 stability inmaize protoplasts transiently transformedwith p35S:MYB11:C-GFP, p35S:MYB11[mEAR]:C-GFP, andp35:ZML2:C-GFP. Samples were treated 1 hwith 25 µMMeJA or 25 µMMeJA plus 5 µMMG132. GFP quantification is shown at the bottom of each panel.Assays were done in biological triplicates and data were normalized to T0. Error bars indicate the SD (n = 3). Statistical analysis of differences betweensamples was performed using the Student’s t test (***P < 0.005 and *P < 0.05). Bars = 10 mm.
3248 The Plant Cell

and C), while no enrichment was observed in other fragmentsof comt lacking this cis-element or in the negative controls(Figure 4B).
To elucidate whether ZML2 regulates the expression of comt,maize protoplasts were transiently transformed to express GFPalone or ZML2 fused to GFP and the expression of comt wasmeasured by qRT-PCR. The results indicated that the expres-sionof the endogenouscomtgene is reduced inpresenceof highlevels of ZML2 (Figure 4C). Similar results were obtained whenmaize protoplasts were cotransformed with ZML2-GFP andpcomt:luciferase as a reporter gene (Figure 4D), indicating thatZML2, like MYB11, acts as a transcriptional repressor of themaize comt gene.
MYB11 and ZML2 Bind Simultaneously to thecomt Promoter
Matrix-Scan software (http://rsat.ulb.ac.be/) was used to pre-dict putative cis-regulatory modules (CRMs) to establishwhether the 59 region of the comt genes of different plantspecies were enriched in the AC-rich (Supplemental Figure 6A)
and GAT(A/C) motifs recognized by MYB11 and ZML2, re-spectively. Our results revealed that the comt promoters ofgrasses are enriched in AC-GAT(A/C) cis-regulatory elements,suggesting the existence of a conserved regulatory module(Figure 5A).Since our protein-protein interaction results indicate thatMYB11
and ZML2 can be part of the same protein complex and both bindthe comt promoter in vivo, we tested whether MYB11 and ZML2bind to these promoter regions in the same cells simultaneouslyusing serial ChIP analyses. In a first step,we performed two controlassays by sequentially immunoprecipitating chromatin frommaizeleaves using (1) anti-MYB11 antibodies (ChIP) and then the pre-immune serum (reChIP) and (2) anti-ZML2 antibodies to performboth ChIP and reChiP (Supplemental Figure 6B). These resultswere compared with those from sequential immunoprecipitationperformed using anti-MYB11 (ChIP) and anti-ZML2 (reChIP)antibodies by analyzing the relative enrichment of fragment C ofthe maize comt promoter by qPCR, using actin as a control (Xieand Grotewold, 2008). No enrichment was observed whenreChIP was performed using preimmune serum, indicating that theC fragment ChIPed by anti-MYB11 antibodies was not retained
Figure 3. MYB11 Interacts with ZML2 through Its EAR Motif.
(A) Yeast two-hybrid mating assays were used to detect interaction between ZML2 and MYB11 or MYB11[mEAR] proteins. pGBKT7 vector and pGADT7empty vectors were used as controls.(B) Determination of the interactions between MYB11 or MYB11[mEAR] and ZML2 by BiFC assay. MYB11 or MYB11[mEAR] was fused to the C-terminalfragment of YFP (YFPC), and ZML2was fused to the N-terminal fragment of YFP (YFPN). Both constructs were coinfiltrated intomaize protoplasts. Emptyvectors (YFPN and YFPC) were used as controls. Bars = 10 mm.(C)CoIPofZML2andMYB11.ThepresenceofZML2andMYB11wasdetectedby immunoblot analyses usingZML2andMYB11antibodies. Input extractsfrom 9-d-old maize leaves were immunoprecipitated with MYB11 antibodies, and ZML2 was detected by immunoblot. The presence of MYB11 in theimmunoprecipitated samples was also detected as a control.
MYB/ZML Complex Regulates Maize Lignin Genes 3249

when the sample was unspecifically reChIPed. By contrast, thesignificant enrichment observed when anti-ZML2 antibodies wereused for ChIP and reChIP suggests that these antibodies aresuitable to perform the reChIP step of this assay. Finally, the sig-nificant enrichment obtained using anti-MYB11 and anti-ZML2antibodies gave a first indication that these two factors bind si-multaneously to the maize comt promoter (Supplemental Figure6B).
Thus, we investigated the simultaneous binding of MYB11and ZML2 to all the AC-rich and GATA cis-elements presentwithin the first kilobase of the maize comt promoter by nano-fluidic qPCR (Fluidigm) using actin, a-tubulin, copia, and the
comt-39UTR as controls. While no enrichment was observedwhen using the control genes, a high enrichment of the comtpromoter fragments containing both the AC-rich and GATAcis-elements was observed (Figure 5B), indicating that MYB11and ZML2 can co-occupy these promoter regions in vivo. Inparticular, our results suggest that MYB11 and ZML2 bindcooperatively to DNA, given the higher enrichment in ChIPassays of the fragment C, that contains both AC (coordinate2195) and GATA/C boxes (coordinate 2165) (Figure 5B).Additionally, we investigated whether the interaction between
the two factors played a role in the repression of the comt gene.Thus, maize protoplasts were cotransformed with the comt
Figure 4. ZML2 Binds the GATA cis-Elements of the Maize comt Promoter in Vivo.
(A)Determination of the DNA binding specificity of ZML2 in vitro using protein bindingmicroarrays. Left: box plot of enrichment scores (E-scores) of all thepossible 6-mers containing the 4-mer coreGATA (gray) or GATC (white). UnderlinedMcorresponds toCor A. Right: box plot of E-scores of all the possible6-merscontainingthe4-mercore indicated.Boxes representquartiles25to75%.Horizontal line represents themedianof thedistribution (quartile50%).Barsindicate quartiles 1 to 25% (above) and 75 to 100% (below), and dots denote outliers of the distribution. PBM indicates the distribution of E-scores of all thepossible 6-mers represented in the microarray.(B)ChIP-qPCRanalyses of ZML2binding to thecomtgenepromoter. FragmentsA,B,C, andD represent the comtpromoter regions analyzed. actin,copia,a-tubulin, and comt 39UTR were used as a controls. The position of the two GATA-rich elements is indicated at the bottom. Results are represented aspercentage of input. Error bars indicate the SE of the data obtained from three biological replicates. Statistical analysis of differences between fragments ofcomt promoter and internal negative controls was performed using Student’s t test (**P < 0.01 and ***P < 0.005).(C) Effect of ZML2 on the expression of the endogenous comt gene. Maize protoplasts were transformed with p35S:ZML2:C-GFP and the empty vector(p35S:C-GFP) as control. comt expression was measured by qPCR using actin as control. Data are the means of three independent transformations. Theerror bars indicate the SE. Statistical analysis of differences between samples was performed using the Student’s t test (*P < 0.05).(D)EffectofZML2onthecomtgenepromoter-driven luciferaseexpression (pcomt:luciferase).Maizeprotoplastswerecotransformedwithp35S:ZML2:C-GFPor 35S:C-GFP and pcomt:luciferase. Transactivation assays were done in biological triplicates, and data were normalized for Renilla activity. Error barsindicate the SE. Statistical analysis of differences between samples was performed using the Student’s t test (*P < 0.05 and ***P < 0.005).
3250 The Plant Cell

promoter fused to luciferase and zml2 or myb11 fused to GFP.The resulting luciferase activity confirmed that both transcriptionfactors could individually represscomtexpression.However, theextent of comt repression was higher when ZML2 and MYB11were simultaneously present (Figure 6A). However, when thesetransient expression assays were performed with ZML2 andMYB11-mEAR, the repression of comt was similar to the ob-served when both proteins were assayed separately (Figures 6Aand 6B). These results are in line with previous results showingthat the mutated MYB11 protein cannot interact with ZML2(Figure 3).
Finally, when both the AC-rich and GATA binding motifs weremutated in the comtgene promoter,MYB11 andZML2 could notrepress the expression of the reporter gene (SupplementalFigure 7).
MeJA Derepresses comt through MYB11 andZML2 Degradation
As already shown, both MYB11 and ZML2 repress comt and aredegraded in aMeJA-dependentmanner. Therefore, to investigatewhether the degradation of these proteins affects the expression
Figure 5. MYB11 and ZML2 Bind Simultaneously to a AC-GAT(A/C) CRM That Is Present in the comt Promoter of Maize and Other Plant Species.
(A) Phylogenetic tree of COMT from several species. On the right, predictions of AC and GAT(A/C) elements and CRERs in the comt promoter of differentspecies. AC elements are represented by lines in different shades of blue, andGAT(A/C) boxes are shown as purple lines. Predicted cis-regulatorymodulesare shaded in yellow.(B)Sequential chromatin immunoprecipitation (ChIP-reChIP) using antibodies againstMYB11andZML2. qPCRassayswere performed for fragmentsA,B,C,C-II,C-III, andDof thecomtpromoterand foractin,copia,a-tubulin, andcomt39UTRasacontrol. AC (grayboxes) andGATA (blackboxes) elements in themaize comt promoter or 59UTR are indicated at the bottom. Gray bars refer to ChIP-reChIP performed using MYB11 (ChIP) and ZML2 antibodies (reChIP).White bars refer to control ChIP-reChIPusingZML2antibodiesonly. Results are representedaspercentageof input. Error bars indicate the SEobtained fromthree independent biological replicates. Statistical analysis was performed using Student’s t test (***P < 0.005).
MYB/ZML Complex Regulates Maize Lignin Genes 3251

of comt, we treatedmaize leaveswithMeJAanddetermined comtmRNA accumulation at different time points (1 and 4 h). The ex-pression of comt was induced 1 h after the addition of the elicitorand the effect of MeJA on comt expression reverted after 4 h oftreatment (Figure 6C).
To investigate whether MeJA affects the binding capacity ofMYB11 and ZML2 to comt gene promoter, we performed ChIPassays using MYB11 and ZML2 antibodies to immunoprecip-itate chromatin fromcontrol andMeJA-treatedmaize leaves. ByqPCR analyses, we determined and compared the enrichmentof fragment C of the comt promoter from treated and untreatedsamples. By fixing the fragment C enrichment of untreatedsamples as 1, and using copia and actin as control genes, weobserved that the enrichment of fragmentC in theMeJA-treatedsamples usingMYB11 or ZML2 antibodies was reduced (Figure6D). This result is in line with the degradation of these twotranscription factors upon MeJA treatment (Figure 2) sug-gesting that the induced expression of comt gene upon MeJA
exposure is, at least partially, caused by the derepressionachieved by the release of MYB11 and ZML2 from the comtpromoter.
pal1, 4cl2, and comt3 Lignin Genes Are Targets of MYB11and ZML2
To determine whether MYB11 and ZML2 act on other genesinvolved in lignin biosynthesis, we developed a catalog of pu-tative genes involved in this process. Based onphylogenetic andexpression analyses of maize databases, 40 putative ligningenes were selected (Supplemental Table 1) to perform ChIP-qPCR experiments using antibodies against ZML2 and MYB11and actin, copia, a-tubulin, and the 39UTR region of the comtgene as negative controls. In addition to comt, 10 gene pro-moters were in vivo targets of MYB11. Among them, pal1, 4cl2,and comt3 genes were also recognized by ZML2 (Figure 7;Supplemental Figure 8) and their expression was induced (or
Figure 6. comt Repression Is Enhanced by the Simultaneous Binding of MYB11 and ZML2 and Is Relieved by MeJA.
(A) Effect of ZML2 andMYB11 on the comt gene promoter-driven luciferase expression.Maize protoplasts were transiently cotransformedwith Luciferasefusedto thecomtpromoter (pcomt:luciferase)withp35S:C-GFP(control)orwithp35S:ZML2:C-GFP+p35S:MYB11:C-GFP,withp35S:MYB11[mEAR]:C-GFP,with p35S:ZML2:C-GFP, or with p35S:MYB11:C-GFP.(B) Transient expression assays using maize protoplasts transformed with pcomt:Luc and p35S:C-GFP (control) or p35S:MYB11:C-GFP + p35S:ZML2:C-GFPorp35S:MYB11[mEAR]:C-GFP+p35S:ZML2:C-GFP.The transient transactivationassaysof (A)and (B)weredone inbiological triplicates, anddatawerenormalized forRenilla activity. Letters indicate significantdifferences incomt-driven luciferase intensity usingaone-wayANOVA followedbyaTukey’stest (P < 0.05; Tukey’s LSD).(C) qRT-PCR analysis of comt expression in 9-d-oldmaize leaves after 1 or 4 h ofMeJA (gray bars) treatments or after 1 or 4 h of 0.01%DMSO (white bars)treatments. The fold change in comt expression was normalized using actin. The error bars indicate the SE of the data obtained from three independentbiological replicates.(D)qPCRanalyses comparing the enrichment of fragment Cof the comt promoter fromMeJA-treated (gray bars) and untreated (white bars) samples. copiaand actin were used as reference genes. The difference in fold enrichment between treated and untreated samples was determined by dividing the foldenrichment ofMeJA-treated samples by the fold enrichment of untreated samples. The error bars indicate the SE of the data obtained from two independentbiological replicates. Statistical analysis of differences between samples was performed using the Student’s t test (*P < 0.05).
3252 The Plant Cell

derepressed) by MeJA (Supplemental Figure 9), suggesting thatthese three genes can be regulated in a way similar to that de-scribed for comt.
The Lignin Repressors MYB31 and MYB42 Interact withZML2 and Share Common Targets among the Lignin Genes
As previously mentioned, MYB11 is evolutionarily closely relatedto MYB31 and MYB42, which are already described as ligninrepressors (Fornalé et al., 2006, 2010; Sonbol et al., 2009). Weperformed ChIP-qPCR using specific antibodies against MYB31andMYB42 and determined that they bind in vivo to 7 and 5 ligningenes, respectively (Figure 7; Supplemental Figure 8). Amongthem, only4cl2and comtwere recognizedbyboth these twoMYBfactors and ZML2. However, we did not detect lignin genes thatwere recognized solely by ZML2 (Figure 7).
We also analyzed the capacity of these transcription factors tobind to several cellulose synthase (cesa) genes. MYB42 was ableto interact invivowith thecesa7promoter (Supplemental Figure8).This is in line with previous results showing that MYB42 alters cell
wall polysaccharides biosynthesis when expressed in Arabi-dopsis (Sonbol et al., 2009).In addition, yeast two-hybrid and BiFC assays indicate that both
MYB31 and MYB42 interact with ZML2 (Supplemental Figure 10),suggesting for MYB31 and MYB42 the existence of a mechanismsimilar to the one described for comt regulation by the MYB11/ZML2complex.Thus,using theconsensussequenceof thebindingsite of the threeMYB factorsderived fromChIP-qPCRdata assays,the binding site of ZML2 obtained from PBM assay and a pattern-matching procedure, we predicted putative cis-regulatorymodulescontaining the AC-rich and GAT(A/C) motifs in 15 lignin genepromoters (Figure 7). Interestingly, the region of 4cl2 and comt3promoters in which MYBs and ZML2 bound corresponded to theregion in which the AC-rich and GAT(A/C) module was predicted.
DISCUSSION
Lignin biosynthesis is affected by biotic and abiotic stresses.However, themolecularmechanisms governing these processes,
Figure 7. Schematic Representation of the DNA Binding Sites of MYB11, MYB31, MYB42, and ZML2 and Prediction of an AC-GAT(A/C) CRM in All theLignin Genes Analyzed.
Blue, red, and green filled circles refer to MYB11, MYB31, and MYB42 binding sites identified by ChIP-qPCR assays, respectively. Predictions of AC andGAT(A/C) elements and CRERs in the promoter of different lignin genes were performed using the onlineMatrix-Scan software (seeMethods) by scanningthe upstream sequences with matrices representing binding motifs for theMYB transcription factors and ZML2. The AC elements are represented by blue(forMYB11), red (MYB31), and green (MYB42) lines, andGAT(A/C) boxes are shownas purple lines. Predicted cis-regulatorymodules are shaded in yellow.
MYB/ZML Complex Regulates Maize Lignin Genes 3253
from the stress perception to the alteration of the lignin geneexpression, are largely unknown. In this study, we identifiedand characterized two new regulators that act as repressors ofthe maize comt gene. One of them, MYB11, belongs to thesubgroup 4 of the R2R3-MYB family, and the second, ZML2,belongs to the TIFY family. MYB11 and ZML2 bind to the AC-rich and GATA cis-elements of the comt promoter in vivo,respectively. MYB11 constitutes the third R2R3-MYB factor,after MYB31 and MYB42 (Fornalé et al., 2006), that repressesmaizecomtgene,while ZML2 is amember of theZMLsubfamilyof TIFY factors that we propose as a newly described regulatorof a lignin gene.
Members of the TIFY family regulate wounding/MeJA re-sponses (Vanholme et al., 2007). In line with this, we showed thatboth treatments trigger upregulation of the maize comt genethrough the simultaneous degradation of ZML2 and MYB11, in-dicating thatboth regulatorscanparticipate in thewound-inducedactivation of lignin biosynthesis. The wound-induced expressionof MYB11 and ZML2 can be considered as a part of the alreadydescribedmechanismaimedtoadjust theabundanceof these twotranscription factors after their protein degradation (Vom Endtet al., 2002).
We show that ZML2 interacts with MYB11, MYB31, andMYB42. Similar interactions have been described for R2R3-MYBfactors and TIFY proteins belonging to the JAZ-domain subfamily(Qi et al., 2011; Song et al., 2011). In these cases, when the JAZprotein is degraded through the jasmonate signaling cascade, theR2R3-MYBpartner is released to trigger theactivationof the targetgenes (Qi et al., 2011). In the case of MYB11 and ZML2, bothfactors are degraded through the jasmonate signaling cascade,and this could lead to the derepression of comt. Such a mecha-nism of gene regulation by transcription factor degradation hasbeen described previously (Catic et al., 2013; McShane andSelbach, 2014).
Regarding the mechanism by which the EAR motif inhibitstranscription, the JAZproteins inArabidopsis recruit theGroucho/Tup1-type corepressor TOPLESS8 (TPL8) and TPL-related pro-teins (TPRs) through NOVEL INTERACTOR OF JAZ (NINJA),whose transcriptional repressor activity is mediated by a func-tional TPL binding EAR repression motif. Both NINJA and TPLproteins function as negative regulators of jasmonate responses(Pauwels et al., 2010). Although the mutation of the EAR motif ofMYB11doesnot seemtoaffect its repressionactivity significantly,it is critical for its interaction with ZML2, as these two factorscannot interact when the EAR motif of MYB11 is mutated. Theimportance of the EAR motif for protein-protein interactions hasbeen reported for EAR-containing proteins, such as a MADS-domain protein (Hill et al., 2008) and a poplar zinc finger protein(Hamel et al., 2011). We show that mutation of the EAR motif ofMYB11 prevents the MeJA-mediated degradation of the protein.This result suggests that the EAR motif of MYB11 is essential toconnect this protein to the MeJA signaling cascade through itsinteraction with ZML2. In that manner, MYB11 can be physicallyremoved from comt gene promoter, allowing the action of tran-scriptional activators. The AC and GATA cis-elements present inthe maize comt promoter constitute a regulatory module, sug-gesting thatMYB11andZML2canbind andact simultaneously torepress comt expression. This was demonstrated by serial ChIP
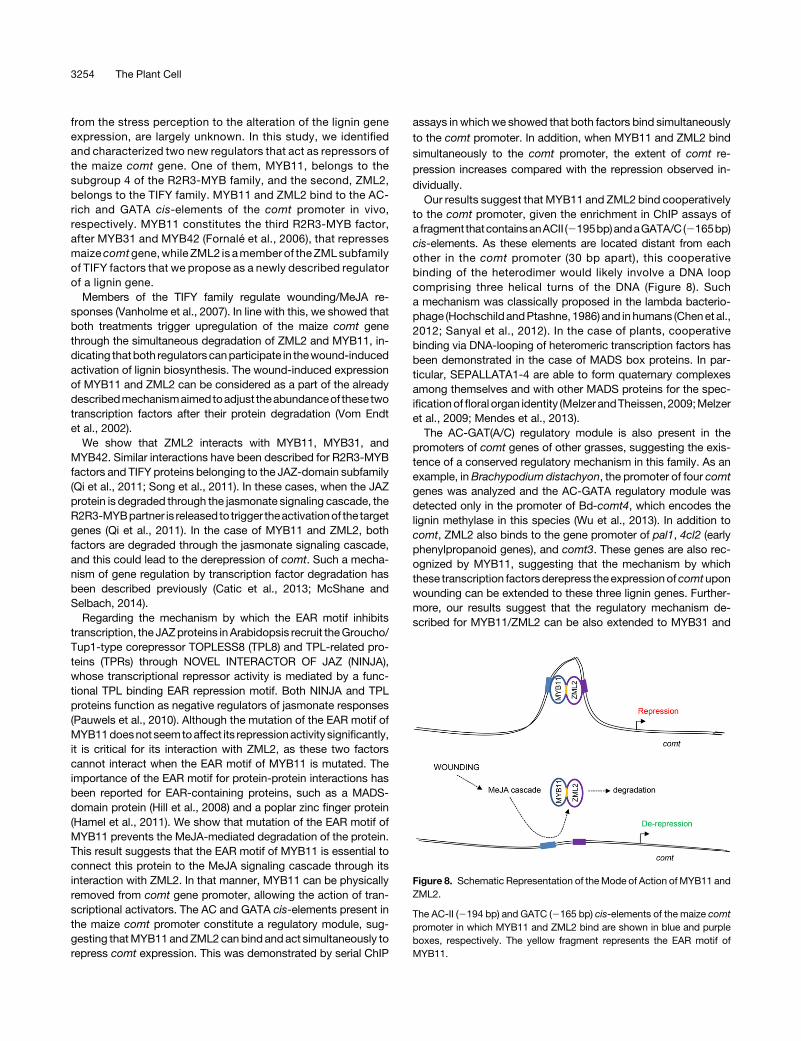
assays in whichwe showed that both factors bind simultaneouslyto the comt promoter. In addition, when MYB11 and ZML2 bindsimultaneously to the comt promoter, the extent of comt re-pression increases compared with the repression observed in-dividually.Our results suggest that MYB11 and ZML2 bind cooperatively
to the comt promoter, given the enrichment in ChIP assays ofa fragment thatcontainsanACII (2195bp)andaGATA/C(2165bp)cis-elements. As these elements are located distant from eachother in the comt promoter (30 bp apart), this cooperativebinding of the heterodimer would likely involve a DNA loopcomprising three helical turns of the DNA (Figure 8). Sucha mechanism was classically proposed in the lambda bacterio-phage (Hochschild andPtashne, 1986) and in humans (Chenet al.,2012; Sanyal et al., 2012). In the case of plants, cooperativebinding via DNA-looping of heteromeric transcription factors hasbeen demonstrated in the case of MADS box proteins. In par-ticular, SEPALLATA1-4 are able to form quaternary complexesamong themselves and with other MADS proteins for the spec-ificationoffloral organ identity (Melzer andTheissen, 2009;Melzeret al., 2009; Mendes et al., 2013).The AC-GAT(A/C) regulatory module is also present in the
promoters of comt genes of other grasses, suggesting the exis-tence of a conserved regulatory mechanism in this family. As anexample, in Brachypodium distachyon, the promoter of four comtgenes was analyzed and the AC-GATA regulatory module wasdetected only in the promoter of Bd-comt4, which encodes thelignin methylase in this species (Wu et al., 2013). In addition tocomt, ZML2 also binds to the gene promoter of pal1, 4cl2 (earlyphenylpropanoid genes), and comt3. These genes are also rec-ognized by MYB11, suggesting that the mechanism by whichthese transcription factorsderepress theexpressionofcomtuponwounding can be extended to these three lignin genes. Further-more, our results suggest that the regulatory mechanism de-scribed for MYB11/ZML2 can be also extended to MYB31 and
Figure 8. Schematic Representation of theMode of Action of MYB11 andZML2.
The AC-II (2194 bp) and GATC (2165 bp) cis-elements of the maize comtpromoter in which MYB11 and ZML2 bind are shown in blue and purpleboxes, respectively. The yellow fragment represents the EAR motif ofMYB11.
3254 The Plant Cell

MYB42, as these twoMYB factors interact with ZML2 and bind tothe comt and 4cl2 gene promoters.
In summary, we show that the regulation of the maize comtexpression is at least partially mediated by the interaction ofsubgroup 4 R2R3-MYB transcription factors and ZML2. Thisprotein complex directly binds to comt promoter. The in-teraction of MYB11 with ZML2 occurs through the EAR motif,and when wounding occurs, the jasmonate signaling cascadetriggers the degradation not only of ZML2 but also of MYB11,leading to the derepression of comt (Figure 8). This regulatorymechanism suggested for MYB11 and ZML2 can be extendedto the lignin repressors MYB31 and MYB42, as both factorsbind in vivo to the comt promoter and physically interact withZML2.
Finally, as MYB11, MYB31, and MYB42 repressors and ZML2share common targets within the lignin biosynthetic genes, it canbe proposed that the induction of lignin biosynthesis that is re-quired upon wounding can rely on the coordinated activation ofthe lignin genes through the action of a MYB/ZML protein com-plex. Thus, this regulatory mechanism would be part of the reg-ulatorycascade triggering thedenovo ligninbiosynthesis requiredupon wounding.
METHODS
Plant Materials, Growth Conditions, and Treatments
Maize (Zeamays)B73 inbredplantsweregrown in soil during9d inagrowthchamber (28°C, 70% relative humidity). For protoplast assays, kernels ofB73xMo17 plants were grown for 13 d on soil in a growth chamber in darkconditions and at 28°C.
Maizeplantswere sprayedwith 100mMMeJA (Sigma-Aldrich) diluted in0.01%DMSO, and control plantswere treatedwith 0.01%DMSOonly andkept inclosedplasticcontainers.Formechanicalwoundtreatments, leavesof 9-d-oldmaize plantswerewounded several timeswith a razor blade anddamaged leaveswere harvested, immediately immersed in liquid nitrogen,and stored at 280°C until use.
DNA Constructs
The full-length cDNAs encoding MYB11, MYB31, ZmMYB42, MYB11[mEAR], ZML1, and ZML2 were cloned into pDONR207 (Invitrogen) usingthe specific primers detailed in Supplemental Table 2. The cDNA clones inthe pDONR207 vector were recombined by LR Gateway reaction into thefinal pDestTH1 (bacterial MBP fusion expression vector) for MYB11 andZML2 antibody testing, in pYFPN43 and pYFPC43 (Belda-Palazón et al.,2012) for BiFC, and in pGBKT7 and pGADT7 Gateway vectors for yeasttwo-hybrid mating assays.
The MYB11[mEAR] was created by primer extension mutagenesisusingMYB11 pDNOR221DNA as a template and the oligonucleotidesZmMYB11_GatewayFW, ZmMYB11[mEAR]A39RV (A), ZmMYB11[mEAR]B39RV (B; synthetic fragment), ZmMYB11[mEAR]A+B39RV (A+B),and ZmMYB11[mEAR]C59FW (C).
First, PCR was performed using ZmMYB11_GatewayFW andZmMYB11[mEAR]A39RV (A) to produce fragment A; then, ZmMYB11[mEAR]B39RV (B) was annealed with fragment A over seven cycles andZmMYB11_GatewayFW and ZmMYB11[mEAR]A+B39RV (A+B) were addedto the mix containing fragments A and B for PCR using 25 cycles. Afterthat, PCR with ZmMYB11[mEAR]C59FW (C) and ZmMYB11_GatewayRVwas performed to obtain the fragment C. Finally, fragments A+Band C were annealed and amplified as above and the MYB11[mEAR]
full-length cDNA clone was generated. The primer sequences areavailable in the Supplemental Table 2.
Production of MYB11, MYB42, and ZML2 Antibodies
Antibodies against MYB31 were obtained as already described (Fornaléet al., 2010) and anti-MYB11 and anti-MYB42 antibodies were raisedagainst specific fragments spanning from amino acids 128 to 201 and 128to 205, respectively (Supplemental Figure 2A; see Supplemental Table 2for primer sequences). In the case of ZML2, the antibodies were obtainedagainst the full-length protein. In all cases, purified proteins (or fragments)were injected in rabbits to produce polyclonal antibodies that were sub-sequentlypurifiedaccording to (Bar-PeledandRaikhel, 1996;Fornaléetal.,2010).
ChIP, Serial ChIP, and Gene Expression Analyses
ChIP experiments were performed as described previously (Morohashiet al., 2007; Morohashi and Grotewold, 2009) with minor modifications(Supplemental Methods).
Serial ChIP for sequential chromatin immunoprecipitations was per-formed as previously described (Xie and Grotewold, 2008) with minormodifications. Cross-linking between Dynabeads/Protein A (Invitrogen)and theMYB11 and ZML2 antibodies was performed to avoid nonspecificbinding to the beads. The first elution was performed according to theprevious protocol.
Nano-quantitative (real-time) PCR analyses of ChIP and serial ChIPsamples were performed using the BioMark System based on Fluidigmmicrofluidic chips at the Genomics and NGSCore Facility of the Centre forResearch in Agricultural Genomics (CRAG), according to the BioMarkSystem protocol (Supplemental Methods).
RNAextractionofmaize leavesorprotoplastswasperformedwithTrizolreagent (Invitrogen) following the manufacturer’s protocol. The cDNA wassynthesized from 1mg total RNA usingQuantiTect reverse transcription kit(Qiagen).
Quantitative PCR of ChIP samples and gene expression analyses wereperformed using LightCycler 480 and DNA SYBR Green I Master Mix(Roche Diagnostics) (Supplemental Methods).
GFP Visualization and BiFC Assays in Maize Protoplasts
Maize protoplasts were isolated from 13-d-old etiolated plants as pre-viously described (Vélez-Bermúdez et al., 2015). For BiFC experiments, allconstructs were transformed by electroporation in maize protoplasts asdescribed (Vélez-Bermúdez et al., 2015). Visualization of the YFP wasperformed 22 h after electroporation, and analysis of single fluorescenceimages was done as described below. Quantification of fluorescenceimages acquired with high magnification objective lens of 603 (BiFCsignals) was performed as described previously (Walter et al., 2004) usingImageJ software (Schneider et al., 2012). The filter sets used for excitationand emissionwere the following: yellow fluorescent protein (EYFP), 514nm(excitation)/ 490 to 500 nm (emission); bright-field, 633 nm. Images wereanalyzed and processed with an FV10-ASW 2.0 viewer.
Maize protoplasts were treated with 25 mM MeJA (Sigma-Aldrich)dissolved in DMSO 18 h after electroporation. The elicitor was added to100mLprotoplastsuspensionand incubatedfordifferent timeperiodsat roomtemperature. For the treatments with MeJA+MG132, the protoplasts weretreated with the same concentration of MeJA (25 mM) and 5 mM MG132. Inboth cases, DMSO-treated protoplasts were used as control. The GFP wasvisualized at 0 h and after 1 h in each sample with a 488-nm argon-ion laser(500- to 545-nm emission filter). The different captured images of maize pro-toplasts harboring p35S:MYB11:C-GFP, p35S:ZML2:C-GFP, p35S:MYB11[mEAR]:C-GFP, and p35S:C-GFP treated with MeJA (25 mM), 5 mMMG132, and DMSO are presented as 3D projected stacks made of
MYB/ZML Complex Regulates Maize Lignin Genes 3255

14 layers of 1 mm. The images were processed with the Olympus FVsoftware and analyzed and quantified using the image processingsoftware Image J (Schneider et al., 2012). The images of proteins labelingwere obtained using a FV 1000 confocal microscope (Olympus) witha 360 oil immersion lens.
Protein Purification and Determination of ZML2 DNA Binding Motifs
TheDNAbindingspecificity of ZML2wasdeterminedusingprotein bindingmicroarrays (PBM11) as previously described (Godoy et al., 2011). ThecDNA of zml2 cloned in fusion to MBP in the pDESTH1 vector usingGateway technology was used for this experiment. Donor template wasobtained through PCR amplification of zml2 with gene-specific oligonu-cleotides (Supplemental Table 1). Recombinant insert was verified bysequencing and plasmid introduced into BL-21 strain. Expression of re-combinant protein was performed as described for the pMAL purificationsystem (New England Biolabs).
CoIP Assays
Maize leaves extracts resuspended in immunoprecipitation buffer supple-mentedwith33plant inhibitor cocktail (Sigma-Aldrich), 50mMMG132,1mMphenylmethylsulfonyl fluoride, 100 mM NaCl, and 2 mM DTT were used forCoIP experiments using the Dynabeads coimmunoprecipitation kit (In-vitrogen) following the supplier’s instructions with modifications and addedto 50 mL Dynabeads previously incubated with MYB11 antibodies. The Dyna-beads were washed and eluted with 30mL SDS-PAGE 1.53 sample buffer(62.5 mM Tris-HCl, pH 6.8, 2 mM b-mercaptoethanol, 10% glycerol, 2%SDS, and 0.01% bromophenol blue) for the immunoblotting analysis. TheZML2 andMYB11 antibodieswere used at a 1:100 dilution. The secondaryantibody (GE Healthcare; NA934-100UL) was used at a 1:5000 dilution.
Luciferase Assays
Maize protoplast transformation efficiency for the luciferase assayswas estimated based on GFP expression. For transient expressionassays, the Dual-Luciferase Reporter Assay System (Promega) wasused (Supplemental Methods).
Phylogenetic Analysis
Dendrograms were assembled from protein-coding sequences by theneighbor-joining method using ClustalW software, version 1.83 (http://www.genome.jp/tools/clustalw/) (Saitou and Nei, 1987; Chenna et al.,2003) (Supplemental Data Sets 1 to 3). The parameters used were as al-ready described for a slow, accurate tree with gap open penalty of 10, gapextension penalty of 0.05, and a Gonnet weight matrix for proteins formultiple alignments; agapopenpenaltyof 10,gapextensionpenalty of 0.1,and a Gonnet weight matrix for proteins for pairwise alignments; andbootstrapped 1000 times (Penning et al., 2009). At-MYB6 was used as theoutgroup for phylogenetic tree ofMYBproteins. Dendrogramswere drawnusingTreeDynavailable at http://www.phylogeny.fr. For the caseofMYBs,we used the online tool http://www.trex.uqam.ca for the tree represen-tation. The protein sequences were obtained from maize sequencing,version 5b.60 (Schnable et al., 2009; http://www.maizesequence.org), andGRASSIUS database (Yilmaz et al., 2009).
Analysis of Promoter Motif Enrichment within Modules
Matrix-Scansoftware (http://rsat.ulb.ac.be/)wasusedtoperformcomputer-based predictions of CRMs to establish whether the 59 region of thecomt gene of different plant species was enriched in the AC-GAT(A/C)CRM. This software detects cis-regulatory element enriched regions
(CRERs; short sequence regionswithasignificanthighdensityofpredictedsites). APvalue is associated toeachCRER,using thebinomial distributionof probability (Turatsinze et al., 2008). The parameters used were as fol-lows: CRERs, background model: Markov Chain Order 0 (the model wastrained from the input sequences), CRER size: 10 to 500, site P value: 1e-2and CRER sig:2 (Turatsinze et al., 2008). We used TRANSFAC formatmatrices that represent the binding site affinity of MYB11 and ZML2.MYB11 ChIP experiment data and ZML2 PMB11 data were used toconstruct thematrices. We usedMEME software available with theMEMEsuite (Bailey, 2011) to identify themotif within the genomic regions that thetranscription factors can bind according to ChIP-qPCR data. The pa-rameters usedwere the same as described above (Turatsinze et al., 2008).In addition, we used DNA pattern software for identification the AC andGAT(A/C) box in the promoter regions (Thomas-Chollier et al., 2008).
Matrix-Scan softwarewasused toestablishwhether the59 regionof thelignin genes was enriched in the regulatory module AC-GATA(C).TRANSFAC matrices for the four transcription factors were employed toidentify CRMs in the 59 region of the genes. The TRANSFAC matrices forMYB31andMYB42weregeneratedusing thesameprotocolas forMYB11.
The parameters used in Matrix-Scan were as follows: CRERs, back-ground model: Markov Chain Order 3 (constructed with the sequenceregion corresponding to 1500 bp upstream of all genes identified in themaize genome), CRER size: 30 to 500, site P value: 1e-3, CRER sites: 1e-3,CRER P-value: 1e-3 y CRER sig: 1. In addition, we used the DNA patternsoftware for identification the ACandGATA(C) box in the promoter regions(Thomas-Chollier et al., 2008).
Accession Numbers
Sequence data from this article can be found in the GenBank/EMBL data-bases under the following accession numbers: myb11 (GRMZM2G000818),myb31 (GRMZM2G050305), myb42 (GRMZM2G419239), zml2(GRMZM2G058479), and comt (AC196475.3_FGT004).
Supplemental Data
Supplemental Figure 1.MYB11 belongs to the R2R3MYB subgroup 4family and is induce by wounding.
Supplemental Figure 2. Production of specific antibodies againstMYB11.
Supplemental Figure 3. Structure and phylogenetic analysis of ZML2and gene expression analyses.
Supplemental Figure 4. Production of specific antibodies againstZML2.
Supplemental Figure 5. MYB11 interacts with ZML2.
Supplemental Figure 6. Consensus binding sequences for MYB11,MYB31, and MYB42 and ChIP-reChIP of MYB11 and ZML2.
Supplemental Figure 7. Effect of ZML2 and MYB11 on the luciferaseexpression driven by comt promoter mutated in AC and GAT(A/C)boxes.
Supplemental Figure 8. ChIP-qPCR analyses of MYB11, MYB31,Zm-MYB42, and ZML2 binding to the comt promoter.
Supplemental Figure 9. pal1, 4cl2, and comt3 expression is inducedby MeJA.
Supplemental Figure 10. MYB31 and MYB42 interact with ZML2.
Supplemental Table 1. List of genes used for ChIP-qPCR analyses.
Supplemental Table 2. Primers used in this work.
Supplemental Methods. Biochemical (immunoblot and yeast twohybrid) and molecular (ChIP and serial ChIP) techniques.
3256 The Plant Cell

Supplemental Data Set 1. Text file of the alignment used for thephylogenetic analysis shown in Supplemental Figure 1A.
Supplemental Data Set 2. Text file of the alignment used for thephylogenetic analysis shown in Supplemental Figure 3A.
Supplemental Data Set 3. Text file of the alignment used for thephylogenetic analysis shown in Figure 5A.
ACKNOWLEDGMENTS
We thank Kengo Morohashi, Maria Katherine Mejía-Guerra, and MariaIsabel Casas (Center for Applied Plant Sciences, Ohio State University) forassistance with ChIP assays, for suggestions, and for providing B73 andMo17xB73 seeds. We also thank Maria José Molina (CRAG, ConsortiumCSIC-IRTA-UAB-UB) for her assistance with yeast two-hybrid assays. Wethank Tsuyoshi Nakagawa (Shimane University) for providing Gatewaybinary vectors that contain thebargene,whichwas identifiedbyMeiji SeikaKaisha. We thank Chung-Ju Rachel Wang (Academia Sinica) for providingtheB73andMO17xB73maizeseeds.Research inD.C.-R.’s laboratorywassupported by a grant from the Spanish Ministry of Science and Education(AGL2011-30545-C02-01), the “Xarxa de Referència de Biotecnologia”(XarBa) fromtheAutonomousGovernmentofCatalonia, theCONSOLIDER-INGENIO program (CSD2007-00036) from the Spanish Ministry of Sci-ence and Innovation, and the SGR programs (SGR2009-GRC703).Research in M.P.’s laboratory was supported by two grants from theSpanish Ministry of Science and Education (BIO2009-13044-C02-01and BIO2012-31860), the framework of the XarBa, and the SGR pro-grams (SGR2009-GRC626) from the Autonomous Government of Cata-lonia. Research in R.S.’s laboratory was supported by grants from theMinistry of Science and Innovation to R.S. (BIO2013-44407). M.P. andR.S. received financial support from the CONSOLIDER-INGENIO pro-gram (CSD2007-00057-B) from the Spanish Ministerio de Ciencia eInnovación. Research in the W.S. laboratory is supported by grantsfrom the Ministry of Science and Technology and Academia Sinica.Research in phenylpropanoid gene regulation in the laboratories ofE.G. and J.G. was supported by a grant from the National ScienceFoundation (IOS-1125620). I.-C.V.-B. was supported by a Spanish FPIFellowship (BES-2007-17316). J.E.S.-H. was supported by the Depart-ment of Innovation, Universities and Enterprise of the GeneralitatdeCatalunya, the European Social Fund FI Fellowship (AGAUR: FI-2006,Resolució EDU/3600/2006; FI-2008, Resolució IUE/2658/2007 andBE-DGR2010), and CRAG.
AUTHOR CONTRIBUTIONS
D.C.-R., M.P., E.G., R.S., J.G., M.R., S.F., I.-C.V.-B., and J.E.S.-H.designed the research. I.-C.V.-B., J.E.S.-H., S.F., I.L.-V., J.-M.F.-Z., andM.R. performed research. D.C.-R., M.P., E.G., R.S., M.R., S.F., I.-C.V.-B.,J.E.S.-H., I.L.-V., W.S., and J.-M.F.-Z. analyzed data. D.C.-R., S.F., M.R.,and M.P. wrote the article. All authors read and approved the article.
Received June 17, 2015; revised October 5, 2015; accepted October 28,2015; published November 13, 2015.
REFERENCES
Bai, Y., Meng, Y., Huang, D., Qi, Y., and Chen, M. (2011). Origin andevolutionary analysis of the plant-specific TIFY transcription factorfamily. Genomics 98: 128–136.
Bailey, T.L. (2011). DREME: motif discovery in transcription factorChIP-seq data. Bioinformatics 27: 1653–1659.
Bar-Peled, M., and Raikhel, N.V. (1996). A method for isolation andpurification of specific antibodies to a protein fused to the GST.Anal. Biochem. 241: 140–142.
Belda-Palazón, B., Ruiz, L., Martí, E., Tárraga, S., Tiburcio, A.F.,Culiáñez, F., Farràs, R., Carrasco, P., and Ferrando, A. (2012).Aminopropyltransferases involved in polyamine biosynthesis local-ize preferentially in the nucleus of plant cells. PLoS One 7: e46907.
Bell-Lelong, D.A., Cusumano, J.C., Meyer, K., and Chapple, C.(1997). Cinnamate-4-hydroxylase expression in Arabidopsis. Reg-ulation in response to development and the environment. PlantPhysiol. 113: 729–738.
Boerjan, W., Ralph, J., and Baucher, M. (2003). Lignin biosynthesis.Annu. Rev. Plant Biol. 54: 519–546.
Capellades, M., Torres, M.A., Bastisch, I., Stiefel, V., Vignols, F.,Bruce, W.B., Peterson, D., Puigdomènech, P., and Rigau, J.(1996). The maize caffeic acid O-methyltransferase gene promoteris active in transgenic tobacco and maize plant tissues. Plant Mol.Biol. 31: 307–322.
Carpita, N.C., and Gibeaut, D.M. (1993). Structural models of primarycell walls in flowering plants: consistency of molecular structure with thephysical properties of the walls during growth. Plant J. 3: 1–30.
Catic, A., Suh, C.Y., Hill, C.T., Daheron, L., Henkel, T., Orford, K.W.,Dombkowski, D.M., Liu, T., Liu, X.S., and Scadden, D.T. (2013).Genome-wide map of nuclear protein degradation shows NCoR1turnover as a key to mitochondrial gene regulation. Cell 155: 1380–1395.
Chen, Y., Bates, D.L., Dey, R., Chen, P.-H., Machado, A.C., Laird-Offringa, I.A., Rohs, R., and Chen, L. (2012). DNA binding by GATAtranscription factor suggests mechanisms of DNA looping and long-range gene regulation. Cell Reports 2: 1197–1206.
Chenna, R., Sugawara, H., Koike, T., Lopez, R., Gibson, T.J.,Higgins, D.G., and Thompson, J.D. (2003). Multiple sequencealignment with the Clustal series of programs. Nucleic Acids Res.31: 3497–3500.
Chini, A., Fonseca, S., Fernández, G., Adie, B., Chico, J.M.,Lorenzo, O., García-Casado, G., López-Vidriero, I., Lozano,F.M., Ponce, M.R., Micol, J.L., and Solano, R. (2007). The JAZfamily of repressors is the missing link in jasmonate signalling.Nature 448: 666–671.
Devoto, A., Ellis, C., Magusin, A., Chang, H.S., Chilcott, C., Zhu, T.,and Turner, J.G. (2005). Expression profiling reveals COI1 to bea key regulator of genes involved in wound- and methyl jasmonate-induced secondary metabolism, defence, and hormone inter-actions. Plant Mol. Biol. 58: 497–513.
Ehlting, J., Büttner, D., Wang, Q., Douglas, C.J., Somssich, I.E.,and Kombrink, E. (1999). Three 4-coumarate:coenzyme A ligasesin Arabidopsis thaliana represent two evolutionarily divergent clas-ses in angiosperms. Plant J. 19: 9–20.
Ellard-Ivey, M., and Douglas, C.J. (1996). Role of jasmonates in theelicitor- and wound-inducible expression of defense genes inparsley and transgenic tobacco. Plant Physiol. 112: 183–192.
Fornalé, S., Sonbol, F.M., Maes, T., Capellades, M., Puigdomènech, P.,Rigau, J., and Caparrós-Ruiz, D. (2006). Down-regulation of the maizeand Arabidopsis thaliana caffeic acid O-methyl-transferase genes bytwo new maize R2R3-MYB transcription factors. Plant Mol. Biol. 62:809–823.
Fornalé, S., et al. (2010). ZmMYB31 directly represses maize ligningenes and redirects the phenylpropanoid metabolic flux. Plant J. 64:633–644.
Godoy, M., Franco-Zorrilla, J.M., Pérez-Pérez, J., Oliveros, J.C.,Lorenzo, O., and Solano, R. (2011). Improved protein-binding
MYB/ZML Complex Regulates Maize Lignin Genes 3257

microarrays for the identification of DNA-binding specificities oftranscription factors. Plant J. 66: 700–711.
Gray, J., Caparrós-Ruiz, D., and Grotewold, E. (2012). Grass phe-nylpropanoids: regulate before using! Plant Sci. 184: 112–120.
Hamel, L.P., Benchabane, M., Nicole, M.C., Major, I.T., Morency,M.J., Pelletier, G., Beaudoin, N., Sheen, J., and Séguin, A. (2011).Stress-responsive mitogen-activated protein kinases interact withthe EAR motif of a poplar zinc finger protein and mediate its deg-radation through the 26S proteasome. Plant Physiol. 157: 1379–1393.
Hill, K., Wang, H., and Perry, S.E. (2008). A transcriptional repressionmotif in the MADS factor AGL15 is involved in recruitment of histonedeacetylase complex components. Plant J. 53: 172–185.
Hochschild, A., and Ptashne, M. (1986). Cooperative binding of l
repressors to sites separated by integral turns of the DNA helix. Cell44: 681–687.
Jouanin, L., Goujon, T., de Nadaï, V., Martin, M.T., Mila, I., Vallet,C., Pollet, B., Yoshinaga, A., Chabbert, B., Petit-Conil, M., andLapierre, C. (2000). Lignification in transgenic poplars with ex-tremely reduced caffeic acid O-methyltransferase activity. PlantPhysiol. 123: 1363–1374.
Kagale, S., and Rozwadowski, K. (2011). EAR motif-mediated tran-scriptional repression in plants: an underlying mechanism for epi-genetic regulation of gene expression. Epigenetics 6: 141–146.
Kaiser, I., Engelberth, J., Groth, B., and Weiler, E.W. (1994). Touch-induced and methyl jasmonate-induced lignification in tendrils ofBryonia dioica Jacq. Bot. Acta 107: 24–29.
Lawton, M.A., and Lamb, C.J. (1987). Transcriptional activation ofplant defense genes by fungal elicitor, wounding, and infection.Mol. Cell. Biol. 7: 335–341.
Lorenzo, O., Chico, J.M., Sánchez-Serrano, J.J., and Solano, R.(2004). JASMONATE-INSENSITIVE1 encodes a MYC transcriptionfactor essential to discriminate between different jasmonate-regulateddefense responses in Arabidopsis. Plant Cell 16: 1938–1950.
McShane, E., and Selbach, M. (2014). Gene expression: degrade toderepress. EMBO J. 33: 407–408.
Melzer, R., and Theissen, G. (2009). Reconstitution of ‘floral quartets’in vitro involving class B and class E floral homeotic proteins. Nu-cleic Acids Res. 37: 2723–2736.
Melzer, R., Verelst, W., and Theissen, G. (2009). The class E floralhomeotic protein SEPALLATA3 is sufficient to loop DNA in ‘floralquartet’-like complexes in vitro. Nucleic Acids Res. 37: 144–157.
Mendes, M.A., Guerra, R.F., Berns, M.C., Manzo, C., Masiero, S.,Finzi, L., Kater, M.M., and Colombo, L. (2013). MADS domaintranscription factors mediate short-range DNA looping that is es-sential for target gene expression in Arabidopsis. Plant Cell 25:2560–2572.
Mizutani, M., Ohta, D., and Sato, R. (1997). Isolation of a cDNA anda genomic clone encoding cinnamate 4-hydroxylase from Arabi-dopsis and its expression manner in planta. Plant Physiol. 113: 755–763.
Morohashi, K., and Grotewold, E. (2009). A systems approach re-veals regulatory circuitry for Arabidopsis trichome initiation by theGL3 and GL1 selectors. PLoS Genet. 5: e1000396.
Morohashi, K., Zhao, M., Yang, M., Read, B., Lloyd, A., Lamb, R.,and Grotewold, E. (2007). Participation of the Arabidopsis bHLHfactor GL3 in trichome initiation regulatory events. Plant Physiol.145: 736–746.
Pauwels, L., Morreel, K., De Witte, E., Lammertyn, F., VanMontagu, M., Boerjan, W., Inzé, D., and Goossens, A. (2008).Mapping methyl jasmonate-mediated transcriptional reprogram-ming of metabolism and cell cycle progression in cultured Arabi-dopsis cells. Proc. Natl. Acad. Sci. USA 105: 1380–1385.
Pauwels, L., et al. (2010). NINJA connects the co-repressorTOPLESS to jasmonate signalling. Nature 464: 788–791.
Penning, B.W., et al. (2009). Genetic resources for maize cell wallbiology. Plant Physiol. 151: 1703–1728.
Qi, T., Song, S., Ren, Q., Wu, D., Huang, H., Chen, Y., Fan, M., Peng,W., Ren, C., and Xie, D. (2011). The Jasmonate-ZIM-domain pro-teins interact with the WD-Repeat/bHLH/MYB complexes to regu-late Jasmonate-mediated anthocyanin accumulation and trichomeinitiation in Arabidopsis thaliana. Plant Cell 23: 1795–1814.
Reymond, P., Weber, H., Damond, M., and Farmer, E.E. (2000).Differential gene expression in response to mechanical woundingand insect feeding in Arabidopsis. Plant Cell 12: 707–720.
Rogers, L.A., and Campbell, M.M. (2004). The genetic control oflignin deposition during plant growth and development. New Phytol.164: 17–30.
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406–425.
Sanyal, A., Lajoie, B.R., Jain, G., and Dekker, J. (2012). The long-range interaction landscape of gene promoters. Nature 489: 109–113.
Sarkanen, K.V., and Ludwig, C.H. (1971). Lignins: Occurrence, For-mation, Structure, and Reactions. (New York: Wiley-Interscience).
Schnable, P.S., et al. (2009). The B73 maize genome: complexity,diversity, and dynamics. Science 326: 1112–1115.
Schneider, C.A., Rasband, W.S., and Eliceiri, K.W. (2012). NIH Image toImageJ: 25 years of image analysis. Nat. Methods 9: 671–675.
Sekhon, R.S., Briskine, R., Hirsch, C.N., Myers, C.L., Springer,N.M., Buell, C.R., de Leon, N., and Kaeppler, S.M. (2013). Maizegene atlas developed by RNA sequencing and comparative evalu-ation of transcriptomes based on RNA sequencing and microarrays.PLoS One 8: e61005.
Shaikhali, J., de Dios Barajas-Lopéz, J., Ötvös, K., Kremnev, D.,Garcia, A.S., Srivastava, V., Wingsle, G., Bako, L., and Strand, Å.(2012). The CRYPTOCHROME1-dependent response to excesslight is mediated through the transcriptional activators ZINCFINGER PROTEIN EXPRESSED IN INFLORESCENCE MERISTEMLIKE1 and ZML2 in Arabidopsis. Plant Cell 24: 3009–3025.
Sonbol, F.M., Fornalé, S., Capellades, M., Encina, A., Touriño, S.,Torres, J.L., Rovira, P., Ruel, K., Puigdomènech, P., Rigau, J.,and Caparrós-Ruiz, D. (2009). The maize ZmMYB42 represses thephenylpropanoid pathway and affects the cell wall structure, com-position and degradability in Arabidopsis thaliana. Plant Mol. Biol.70: 283–296.
Song, S., Qi, T., Huang, H., Ren, Q., Wu, D., Chang, C., Peng, W.,Liu, Y., Peng, J., and Xie, D. (2011). The Jasmonate-ZIM domainproteins interact with the R2R3-MYB transcription factors MYB21and MYB24 to affect jasmonate-regulated stamen development inArabidopsis. Plant Cell 23: 1000–1013.
Thines, B., Katsir, L., Melotto, M., Niu, Y., Mandaokar, A., Liu, G.,Nomura, K., He, S.Y., Howe, G.A., and Browse, J. (2007). JAZrepressor proteins are targets of the SCF(COI1) complex duringjasmonate signalling. Nature 448: 661–665.
Thomas-Chollier, M., Sand, O., Turatsinze, J.V., Janky, R.,Defrance, M., Vervisch, E., Brohée, S., and van Helden, J.(2008). RSAT: regulatory sequence analysis tools. Nucleic AcidsRes. 36: W119–W127.
Tianpei, X., Li, D., Qiu, P., Luo, J., Zhu, Y., and Li, S. (2015). Scor-pion peptide LqhIT2 activates phenylpropanoid pathways viajasmonate to increase rice resistance to rice leafrollers. Plant Sci.230: 1–11.
Turatsinze, J.V., Thomas-Chollier, M., Defrance, M., and vanHelden, J. (2008). Using RSAT to scan genome sequences for
3258 The Plant Cell

transcription factor binding sites and cis-regulatory modules. Nat.Protoc. 3: 1578–1588.
Vance, C.P., Kirk, T.K., and Sherwood, R.T. (1980). Lignification asa mechanism of disease resistance. Annu. Rev. Phytopothol. 18:259–288.
Vanholme, B., Grunewald, W., Bateman, A., Kohchi, T., and Gheysen,G. (2007). The tify family previously known as ZIM. Trends Plant Sci. 12:239–244.
Vélez-Bermúdez, I.C., Carretero-Paulet, L., Legnaioli, T., Ludevid,D., Pagès, M., and Riera, M. (2015). Novel CK2a and CK2b sub-units in maize reveal functional diversification in subcellular locali-zation and interaction capacity. Plant Sci. 235: 58–69.
Vignols, F., Rigau, J., Torres, M.A., Capellades, M., and Puigdomènech,P. (1995). The brown midrib3 (bm3) mutation in maize occurs in the geneencoding caffeic acid O-methyltransferase. Plant Cell 7: 407–416.
Vom Endt, D., Kijne, J.W., and Memelink, J. (2002). Transcriptionfactors controlling plant secondary metabolism: what regulates theregulators? Phytochemistry 61: 107–114.
Walter, M., Chaban, C., Schütze, K., Batistic, O., Weckermann, K.,Näke, C., Blazevic, D., Grefen, C., Schumacher, K., Oecking, C.,Harter, K., and Kudla, J. (2004). Visualization of protein inter-actions in living plant cells using bimolecular fluorescence com-plementation. Plant J. 40: 428–438.
Wu, X., Wu, J., Luo, Y., Bragg, J., Anderson, O., Vogel, J., and Gu,Y.Q. (2013). Phylogenetic, molecular, and biochemical character-ization of caffeic acid o-methyltransferase gene family in Brachypodiumdistachyon. Int. J. Plant Genomics 2013: 423189.
Xie, Z., and Grotewold, E. (2008). Serial ChIP as a tool to investigate the co-localization or exclusion of proteins on plant genes. Plant Methods 4: 25.
Xue, Y.J., Tao, L., and Yang, Z.M. (2008). Aluminum-induced cell wallperoxidase activity and lignin synthesis are differentially regulatedby jasmonate and nitric oxide. J. Agric. Food Chem. 56: 9676–9684.
Yilmaz, A., Nishiyama, M.Y., Jr., Fuentes, B.G., Souza, G.M., Janies, D.,Gray, J., and Grotewold, E. (2009). GRASSIUS: a platform for com-parative regulatory genomics across the grasses. Plant Physiol. 149:171–180.
MYB/ZML Complex Regulates Maize Lignin Genes 3259

DOI 10.1105/tpc.15.00545; originally published online November 13, 2015; 2015;27;3245-3259Plant Cell
Montserrat Pagés, Marta Riera and David Caparros-RuizJosé-Manuel Franco-Zorrilla, Erich Grotewold, John Gray, Roberto Solano, Wolfgang Schmidt, Isabel-Cristina Vélez-Bermúdez, Jorge E. Salazar-Henao, Silvia Fornalé, Irene López-Vidriero,
A MYB/ZML Complex Regulates Wound-Induced Lignin Genes in Maize
This information is current as of October 19, 2018
Supplemental Data /content/suppl/2015/11/02/tpc.15.00545.DC1.html
References /content/27/11/3245.full.html#ref-list-1
This article cites 61 articles, 19 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists