a leishmaniasis vaccine breakthrough: are we almost there? · pdf filea leishmaniasis vaccine...
TRANSCRIPT

A Leishmaniasis Vaccine Breakthrough: Are We Almost There?
1
Jude Uzonna, DVM, Ph.D.Professor & Research Manitoba Chair,Department of ImmunologyUniversity of Manitoba
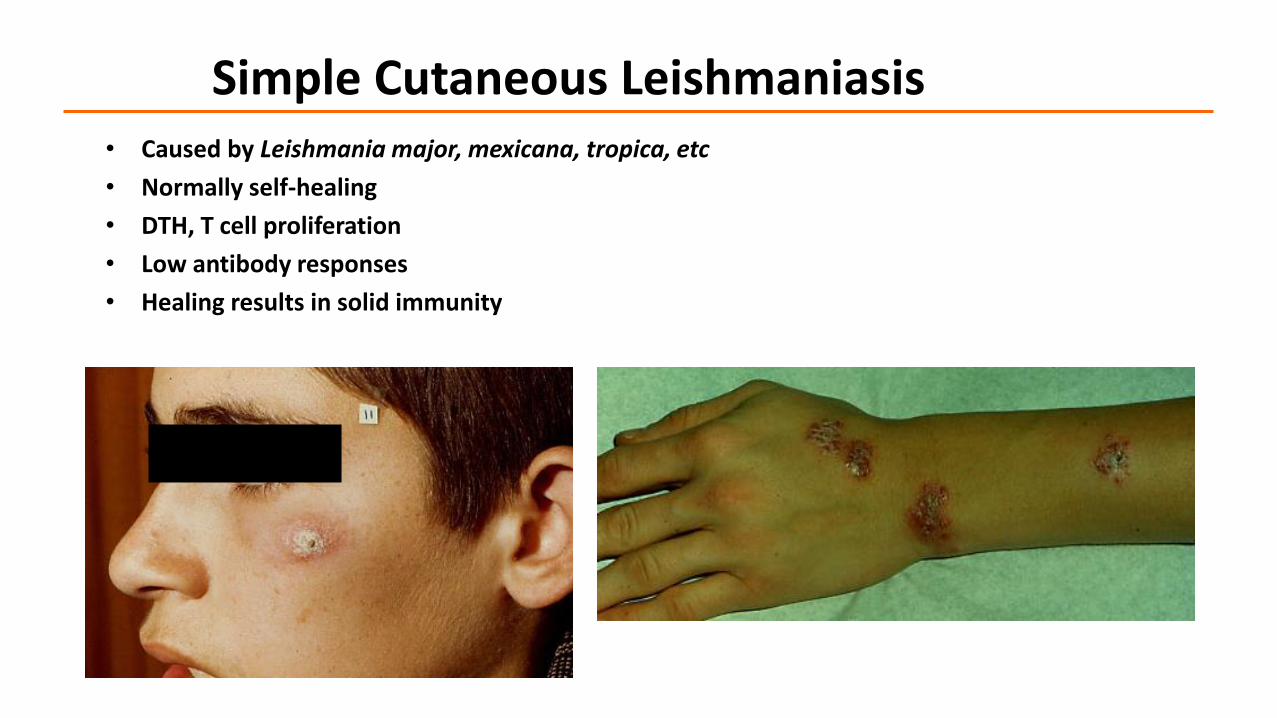
Simple Cutaneous Leishmaniasis• Caused by Leishmania major, mexicana, tropica, etc
• Normally self-healing
• DTH, T cell proliferation
• Low antibody responses
• Healing results in solid immunity
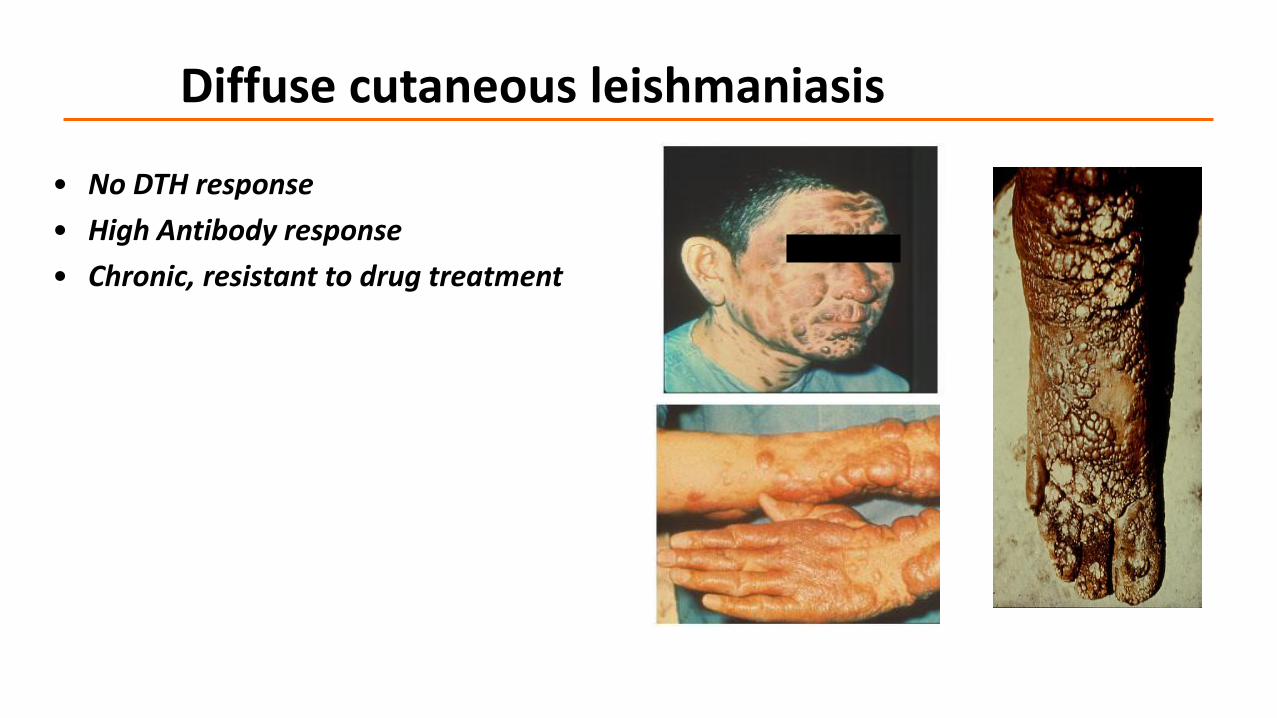
Diffuse cutaneous leishmaniasis
• No DTH response
• High Antibody response
• Chronic, resistant to drug treatment

Mucocutaneous leishmaniasis
• Very strong DTH
• Intense inflammatory response
• Chronic, resistant to drug treatment
• Very few parasites at inflammatory sites
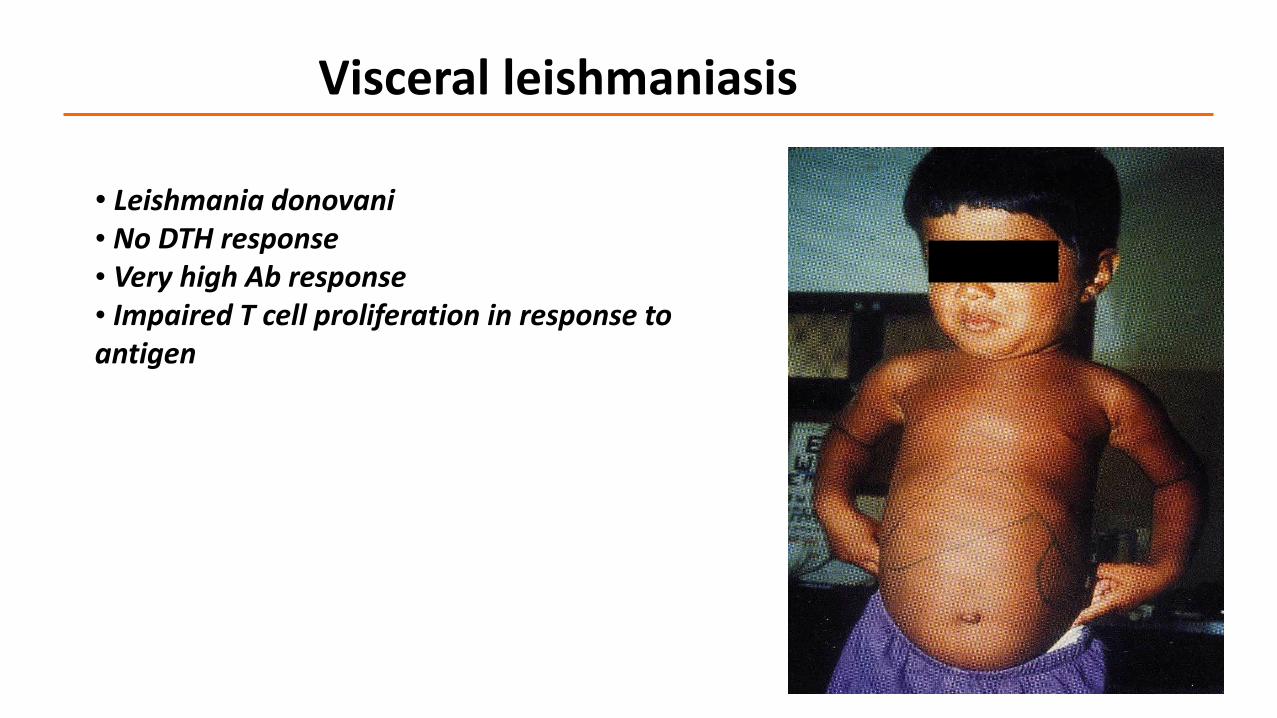
Visceral leishmaniasis
• Leishmania donovani• No DTH response• Very high Ab response• Impaired T cell proliferation in response to antigen
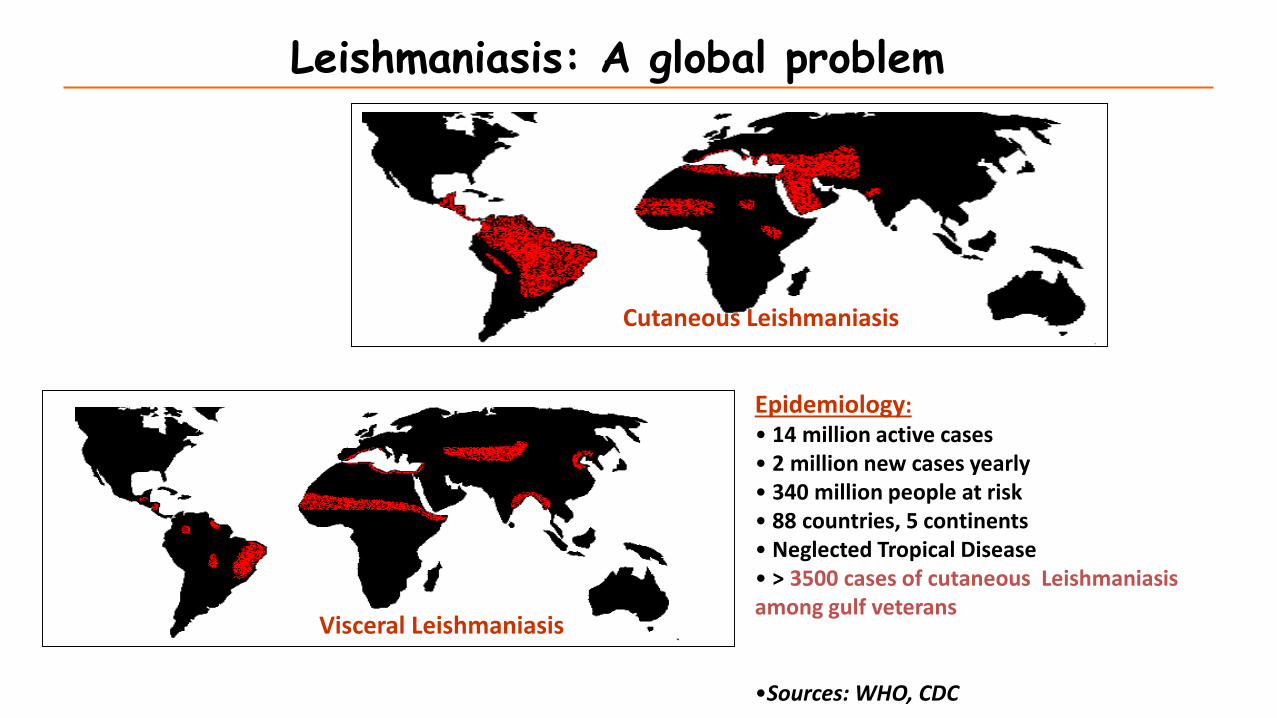
Cutaneous Leishmaniasis
Visceral Leishmaniasis
Leishmaniasis: A global problem
Epidemiology:• 14 million active cases• 2 million new cases yearly• 340 million people at risk• 88 countries, 5 continents• Neglected Tropical Disease• > 3500 cases of cutaneous Leishmaniasis among gulf veterans
•Sources: WHO, CDC
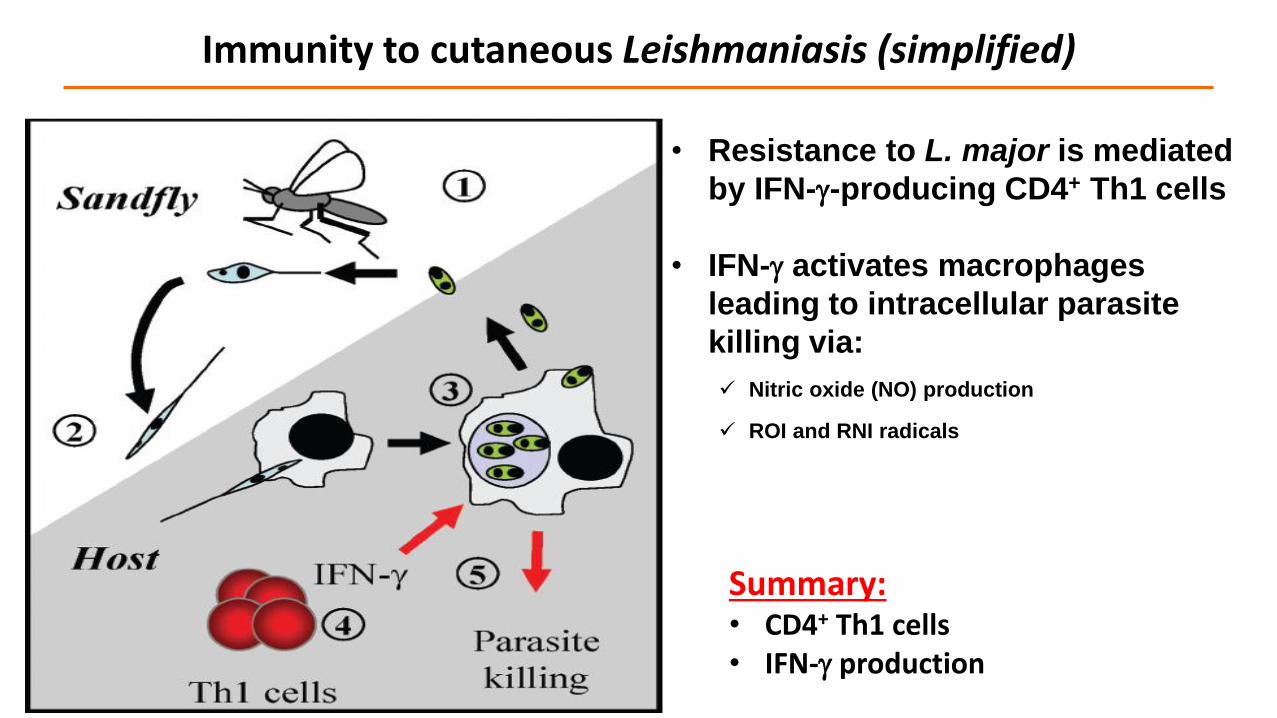
• Resistance to L. major is mediated
by IFN--producing CD4+ Th1 cells
• IFN- activates macrophages
leading to intracellular parasite
killing via:
Nitric oxide (NO) production
ROI and RNI radicals
Immunity to cutaneous Leishmaniasis (simplified)
Summary:• CD4+ Th1 cells• IFN- production
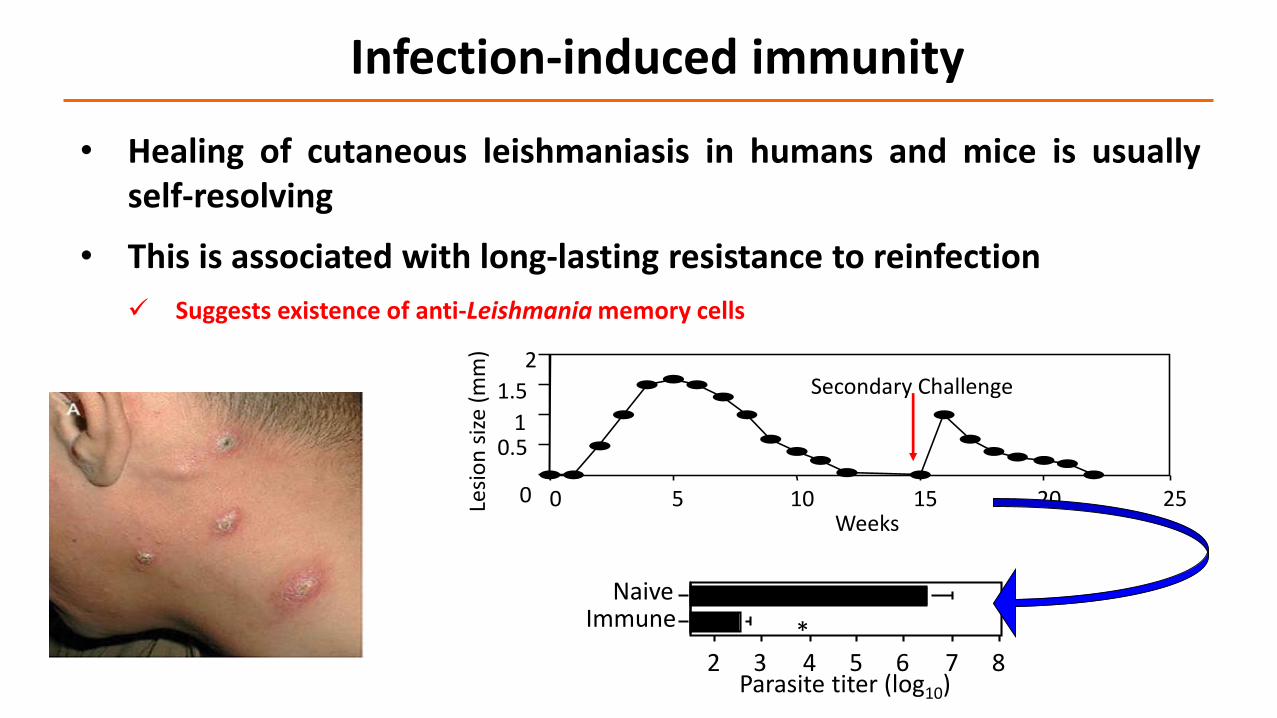
• Healing of cutaneous leishmaniasis in humans and mice is usuallyself-resolving
• This is associated with long-lasting resistance to reinfection
Suggests existence of anti-Leishmania memory cells
Infection-induced immunity
Lesi
on
siz
e (m
m)
0
0.5
1.5
2
0 5 10 15 20 25Weeks
1
Secondary Challenge
2 3 4 5 6 7 8Parasite titer (log10)
ImmuneNaive
*
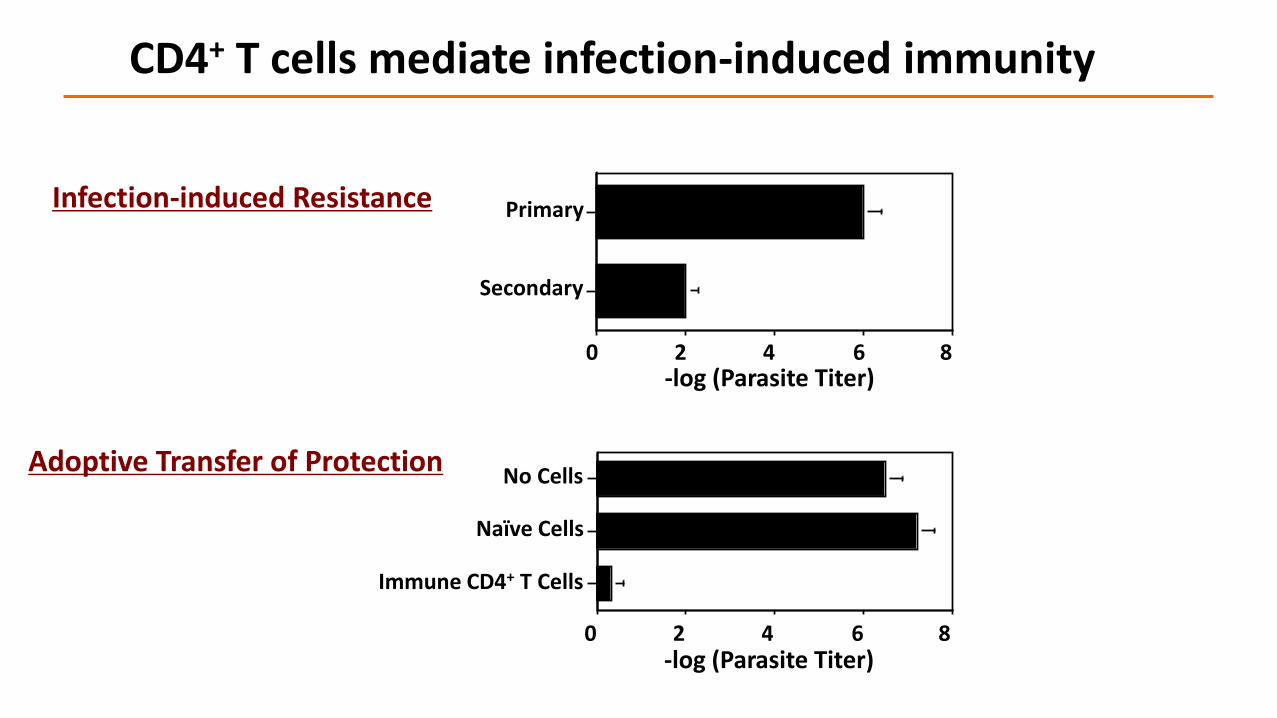
Infection-induced Resistance
Adoptive Transfer of Protection
Primary
Secondary
-log (Parasite Titer)0 2 4 6 8
No Cells
Naïve Cells
Immune CD4+ T Cells
-log (Parasite Titer)0 2 4 6 8
CD4+ T cells mediate infection-induced immunity

Is there a need for a Leishmania vaccines?
• Epidemiology: Over 14 million active cases 2 million new cases yearly 340 million people at risk 88 countries, 5 continents
• Chemotherapy until recently is very limited high cost, toxicity and route of administration
• Elimination can likely only be achieved through widespread vaccination
• Vaccination could provide longterm reductions in potential reservoirs
10

Pubmed Search!!
• Leishmania and immunity = 2606
• Leishmania and resistance = 2127
• Leishmania and vaccine = 1649
• Leishmania and vaccination = 689
• Leishmania and effective vaccine = 392
• Human Leishmaniasis and effective vaccine = 227
• Leishmania vaccination trials = 115
Approved Effective Human Leishmania Vaccine = 0

12
Scientific Issue: Why is there no effective human vaccine despite prolific
publications?

• We do not fully understand the correlates of
protective immunity.
Immunologic memory (development, maintenance and loss)
Antigens that mediate memory T cell responses
13

Is Vaccination Feasible?
• Infection-induced immunity: Recovery from virulent Leishmania infection leads to development of strong durable immunity against virulent challenge
• Leishmanization is very effective

• Heat Killed parasites
– First generation vaccines
• Live attenuated/genetically modified
– Second generation vaccines
• Defined protein/subunit vaccines
– Third generation vaccines
• Leishmanization
Leishmaniasis Vaccine Approaches

• Heat-inactivated (Autoclaved) Leishmania parasites
• Several doses or‘boosters’with with different adjuvants (BCG, Alum, CpG)
• Questionable efficacy
• Easy and cheap to make (minimal technology) and very safe
• Does not require understanding of complex immunologic correlates of protective immunity
• Reports of immunotherapeutic effects in New World cutaneous leishmaniasis
Heat-killed (ALM) vaccines

Problems with ALM Vaccine
• Standardization and scale-up issues
• Undesirable immune responses Qualitative differences in the early immune response to live and killed Leishmania major:
Implications for vaccination strategies against Leishmaniasis. Okwor I, Liu D, Uzonna JE.
Vaccine. 2009 Apr 28;27(19):2554-62.
Inoculation of killed Leishmania major into immune mice rapidly disrupts immunity to a
secondary challenge via IL-10-mediated process. Okwor I, Liu D, Beverley SM, Uzonna JE. Proc
Natl Acad Sci U S A. 2009 Aug 18;106(33):13951-6.
17

Leishmanization
• Deliberate inoculation of live (virulent) parasites to the hidden parts of the body with the aim of inducing protection from natural infection following recovery
• Very Effective
• Undesirable side effects!!!
18
Courtesy: Dr Ali Khamispour
Undesirable effects of Leishmanization

Rationale for Live Vaccine
• Infection-induced Immunity Recovery from infection leads to development of durable immunity
• Persistent parasites are important for maintaining infection-induced immunity Attenuated parasites persist for a long time in infected/vaccinated host
Clearance of parasites leads to loss of protective immunity
• Leishmanization works

Two major strategies for live-attenuated vaccine:
• Virulent parasites (Leishmanization)
• Attenuated parasites

Attenuated Parasites
• Gamma Irradiation– Rivier et al., 1993
• Serial in vitro passage with/without drugs– Daneshvar, et al 2003, Onyalo et al 2005
• Infection with non-pathogenic strain (L. tarentolae)– Breton et al 2005
• Genetically modified organisms– LPG1, LPG2, LPG5A/B, DHFR-Ts, GDP-MP, GP63, etc
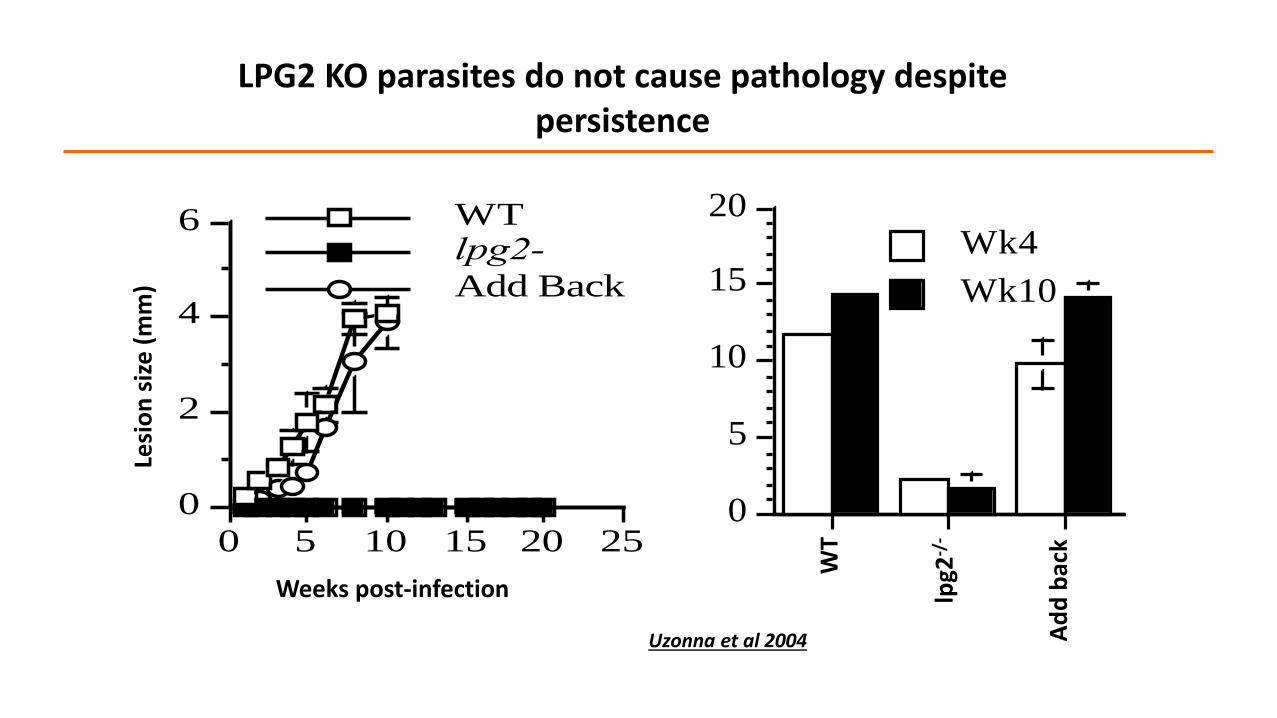
0
5
10
15
20
Wk10
Wk4
WT
lpg2
-/-
Ad
d b
ack
LPG2 KO parasites do not cause pathology despite persistence
0
2
4
6
0 5 10 15 20 25
Add Back
lpg2-WT
Weeks post-infection
Lesi
on
siz
e (
mm
)
Uzonna et al 2004
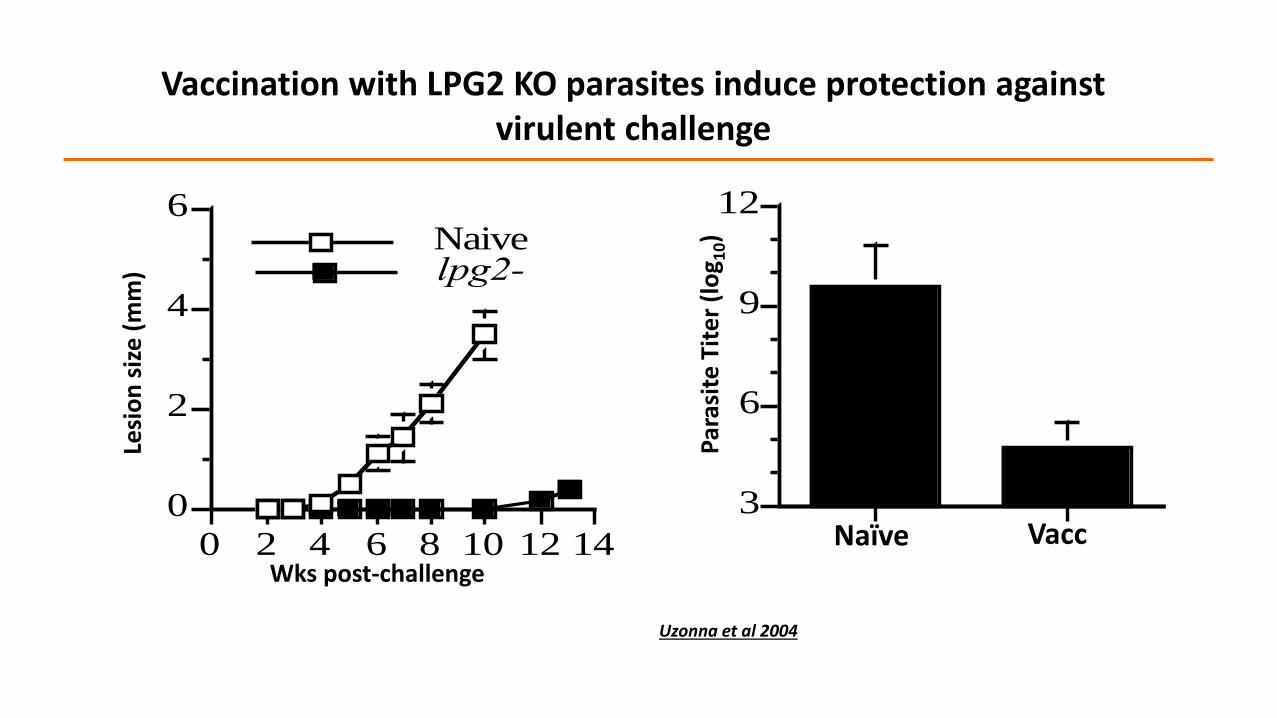
3
6
9
12
0
2
4
6
0 2 4 6 8 10 12 14
lpg2-Naive
Naïve VaccWks post-challenge
Par
asit
e T
ite
r (l
og 1
0)
Lesi
on
siz
e (
mm
)
Vaccination with LPG2 KO parasites induce protection against virulent challenge
Uzonna et al 2004

Problems of live-attenuated vaccine
• Reversion to virulence (due to long-term persistence)– Identification of a compensatory mutant (lpg2-REV) of Leishmania major able to survive as amastigotes
within macrophages without LPG2-dependent glycoconjugates and its significance to virulence and
immunization strategies. Späth GF, Lye LF, Segawa H, Turco SJ, Beverley SM. Infect Immun. 2004
Jun;72(6):3622-7.
• Contraindicated in immuno-compromised individuals
This vaccination protocol will most likely never be approved

3rd Generation Vaccines
• Subunit vaccines with refined products (recombinant proteins)
• Several recombinant proteins or polyproteins (e.g. Leish-111F, LmSTI1, KSAC, etc) with adjuvants such as MPL
• Some have progressed into clinical trials
• One approved for dogs in South America (Leishmune)
26

Why 3rd Generation Vaccines Fail
• Use of in silico models to predict “protective” vaccine candidates
• Selection of candidates based primarily on those that elicit strong primary immune response
• Skewed and biased immunization protocols: designs favor publication; not necessarily long-term protection– Animal model (BALB/c mice)
– Antigen dose
– Type of adjuvants
– Frequency of immunization
– Time of challenge etc.

28
Objectives:
• To use in vivo approach to identify Leishmania antigens that induce infection-induced immunity (memory CD4+ T cell responses?)
• Assess whether vaccination with these antigens will induce long-lasting protection against virulent challenge
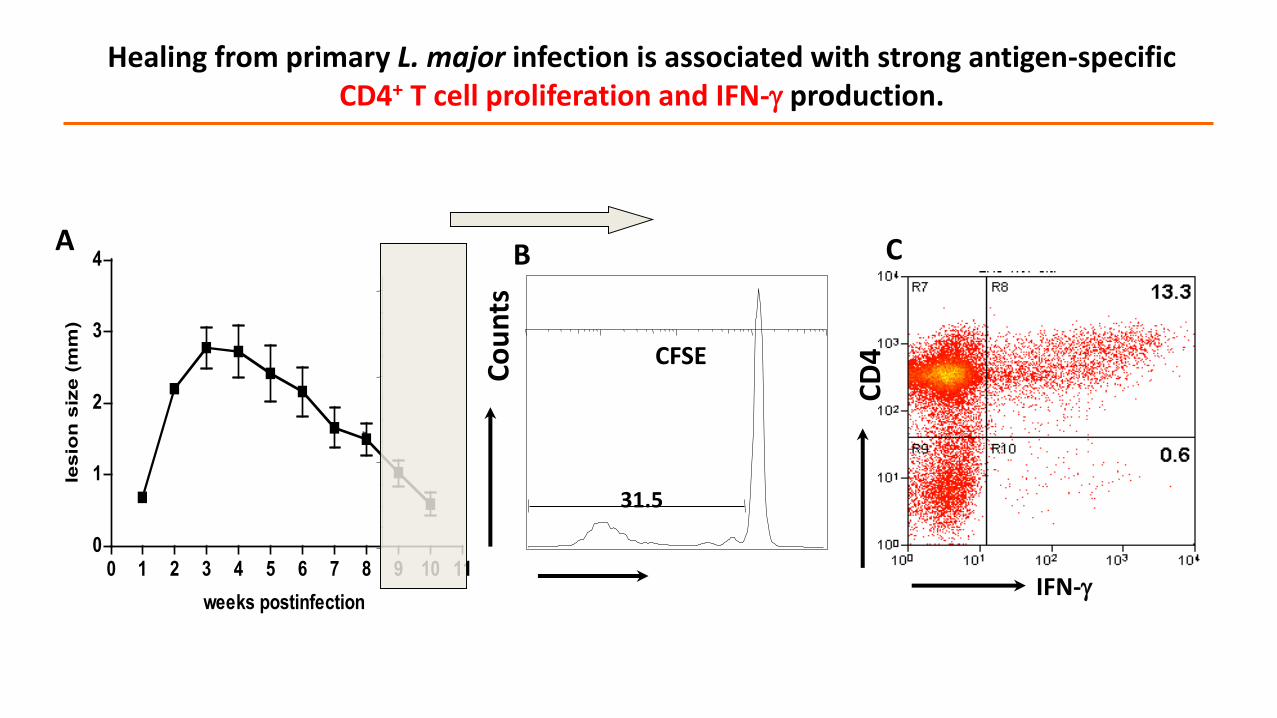
A B
CFSE#
31.5C
ou
nts
IFN-
CD
4
0 1 2 3 4 5 6 7 8 9 10 110
1
2
3
4
weeks postinfection
lesio
n s
ize (
mm
)
C
Healing from primary L. major infection is associated with strong antigen-specific CD4+ T cell proliferation and IFN- production.
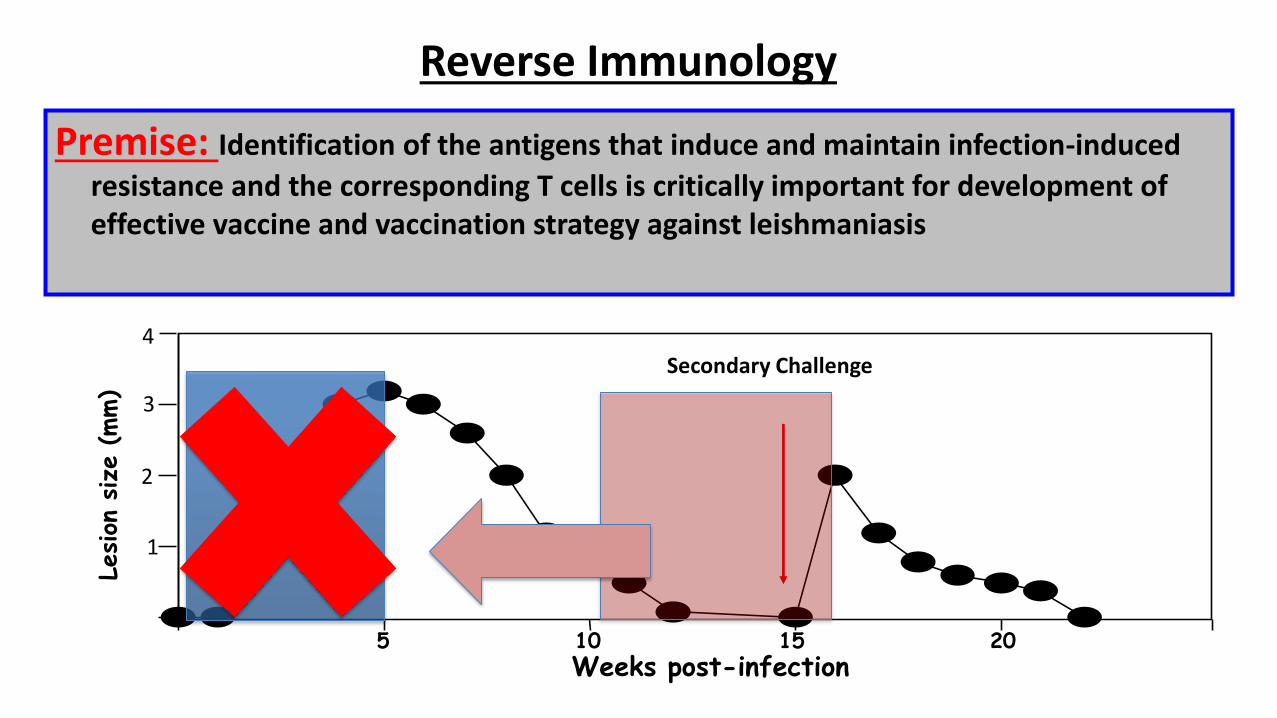
Premise: Identification of the antigens that induce and maintain infection-induced
resistance and the corresponding T cells is critically important for development of effective vaccine and vaccination strategy against leishmaniasis
Reverse Immunology
2
Secondary Challenge
1
3
4
Lesion
size (mm)
Weeks post-infection5 10 15 20
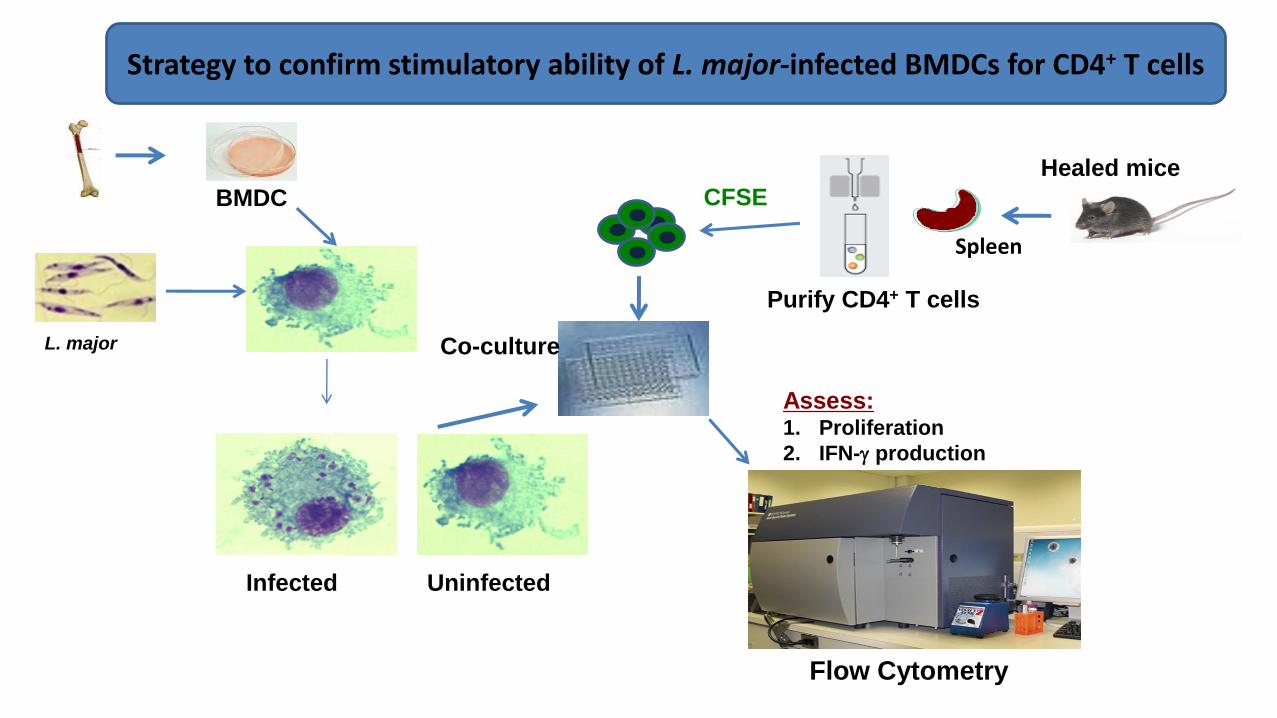
BMDC
L. major
Infected
Flow Cytometry
Purify CD4+ T cells
CFSE
Healed mice
Strategy to confirm stimulatory ability of L. major-infected BMDCs for CD4+ T cells
Uninfected
Co-culture
Assess:1. Proliferation
2. IFN- production
Spleen
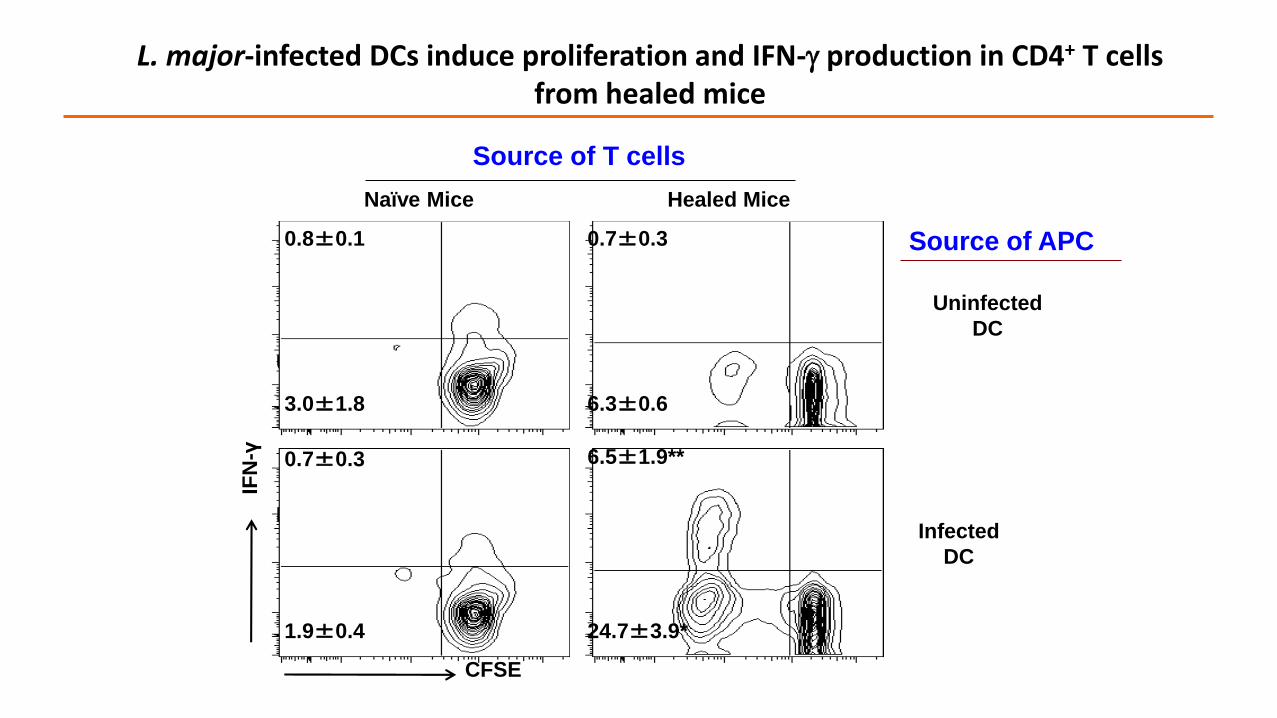
Naïve Mice
Infected BMDC
Uninfected BMDC
IFN
-γ
CFSE
Healed Mice
Source of T cells
0.8±0.1
3.0±1.8
0.7±0.3
1.9±0.4
0.7±0.3
6.3±0.6
6.5±1.9**
24.7±3.9*
Infected
DC
Uninfected
DC
L. major-infected DCs induce proliferation and IFN- production in CD4+ T cells from healed mice
Source of APC
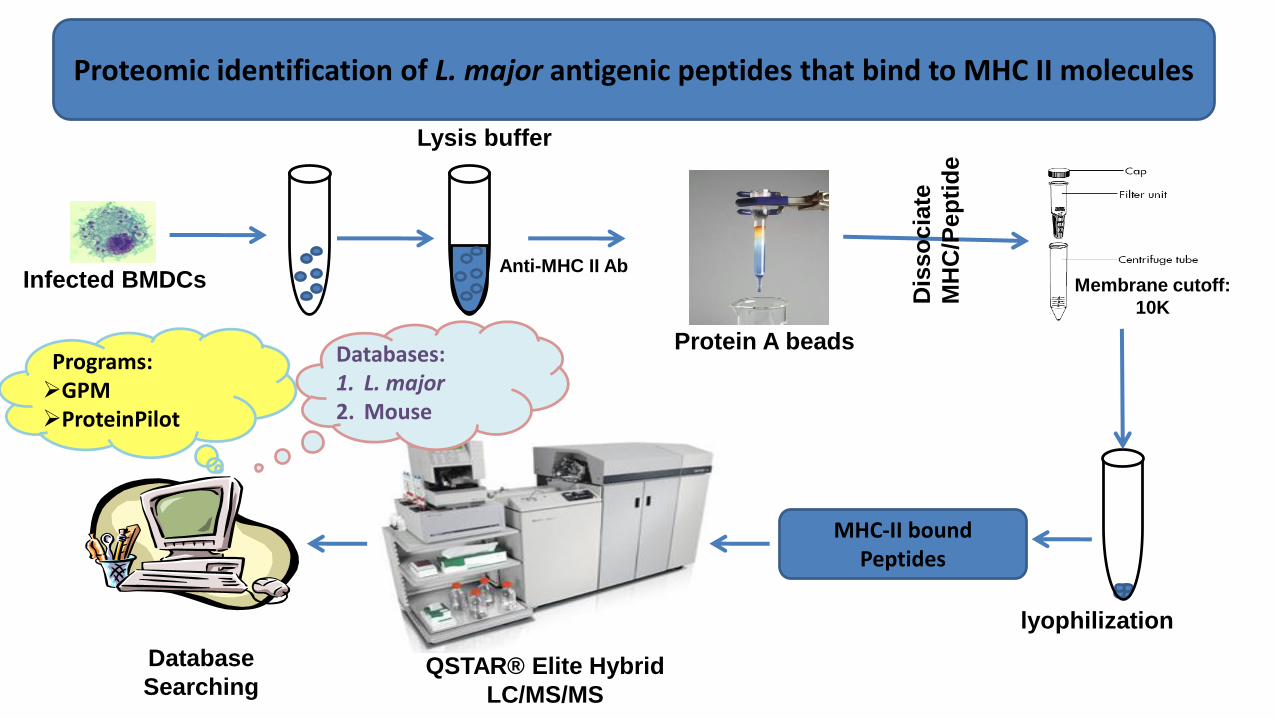
Membrane cutoff:
10K
Protein A beads
Anti-MHC II Ab
lyophilization
Lysis buffer
MHC-II bound Peptides
Programs:GPMProteinPilot
Databases:1. L. major2. Mouse
Infected BMDCs
QSTAR® Elite Hybrid
LC/MS/MS
Database
Searching
Proteomic identification of L. major antigenic peptides that bind to MHC II molecules
Dis
so
cia
te
MH
C/P
ep
tid
e
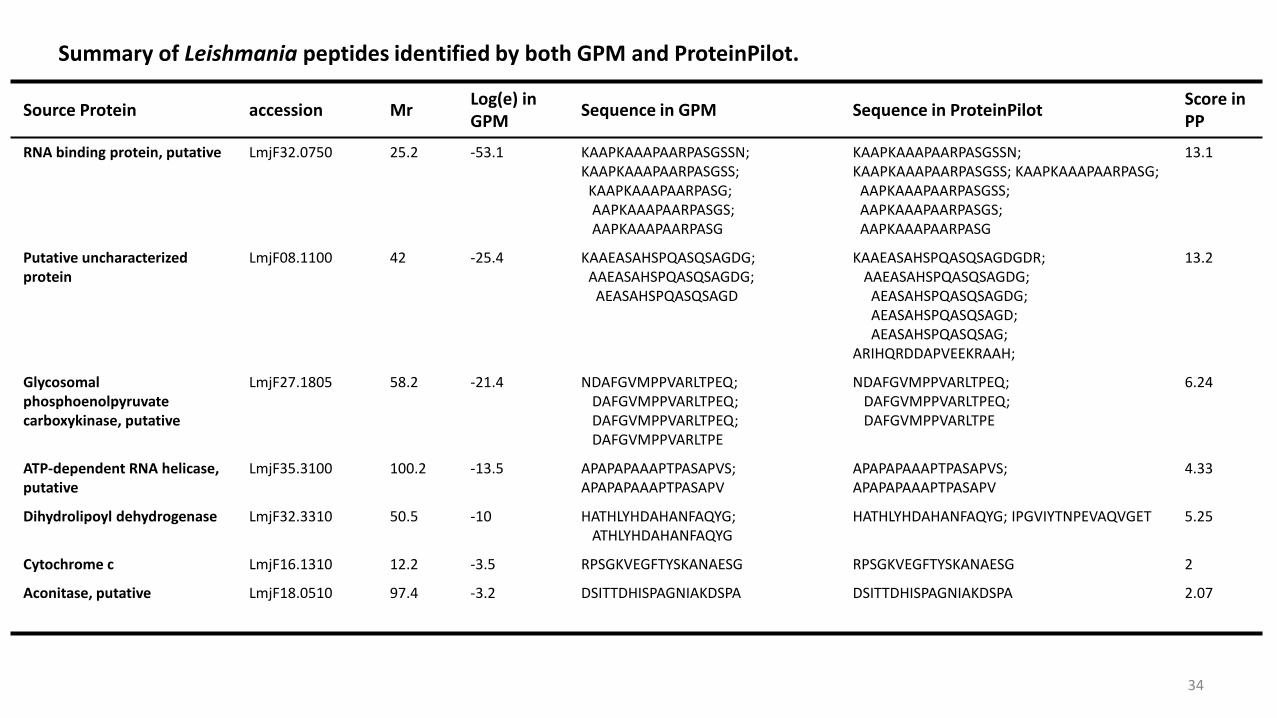
Source Protein accession MrLog(e) in GPM
Sequence in GPM Sequence in ProteinPilotScore in PP
RNA binding protein, putative LmjF32.0750 25.2 -53.1 KAAPKAAAPAARPASGSSN; KAAPKAAAPAARPASGSS;
KAAPKAAAPAARPASG;AAPKAAAPAARPASGS; AAPKAAAPAARPASG
KAAPKAAAPAARPASGSSN;KAAPKAAAPAARPASGSS; KAAPKAAAPAARPASG;
AAPKAAAPAARPASGSS;AAPKAAAPAARPASGS;AAPKAAAPAARPASG
13.1
Putative uncharacterized protein
LmjF08.1100 42 -25.4 KAAEASAHSPQASQSAGDG;AAEASAHSPQASQSAGDG;
AEASAHSPQASQSAGD
KAAEASAHSPQASQSAGDGDR;AAEASAHSPQASQSAGDG;
AEASAHSPQASQSAGDG; AEASAHSPQASQSAGD; AEASAHSPQASQSAG;
ARIHQRDDAPVEEKRAAH;
13.2
Glycosomalphosphoenolpyruvatecarboxykinase, putative
LmjF27.1805 58.2 -21.4 NDAFGVMPPVARLTPEQ;DAFGVMPPVARLTPEQ; DAFGVMPPVARLTPEQ; DAFGVMPPVARLTPE
NDAFGVMPPVARLTPEQ;DAFGVMPPVARLTPEQ; DAFGVMPPVARLTPE
6.24
ATP-dependent RNA helicase, putative
LmjF35.3100 100.2 -13.5 APAPAPAAAPTPASAPVS; APAPAPAAAPTPASAPV
APAPAPAAAPTPASAPVS;APAPAPAAAPTPASAPV
4.33
Dihydrolipoyl dehydrogenase LmjF32.3310 50.5 -10 HATHLYHDAHANFAQYG; ATHLYHDAHANFAQYG
HATHLYHDAHANFAQYG; IPGVIYTNPEVAQVGET 5.25
Cytochrome c LmjF16.1310 12.2 -3.5 RPSGKVEGFTYSKANAESG RPSGKVEGFTYSKANAESG 2
Aconitase, putative LmjF18.0510 97.4 -3.2 DSITTDHISPAGNIAKDSPA DSITTDHISPAGNIAKDSPA 2.07
Summary of Leishmania peptides identified by both GPM and ProteinPilot.
34
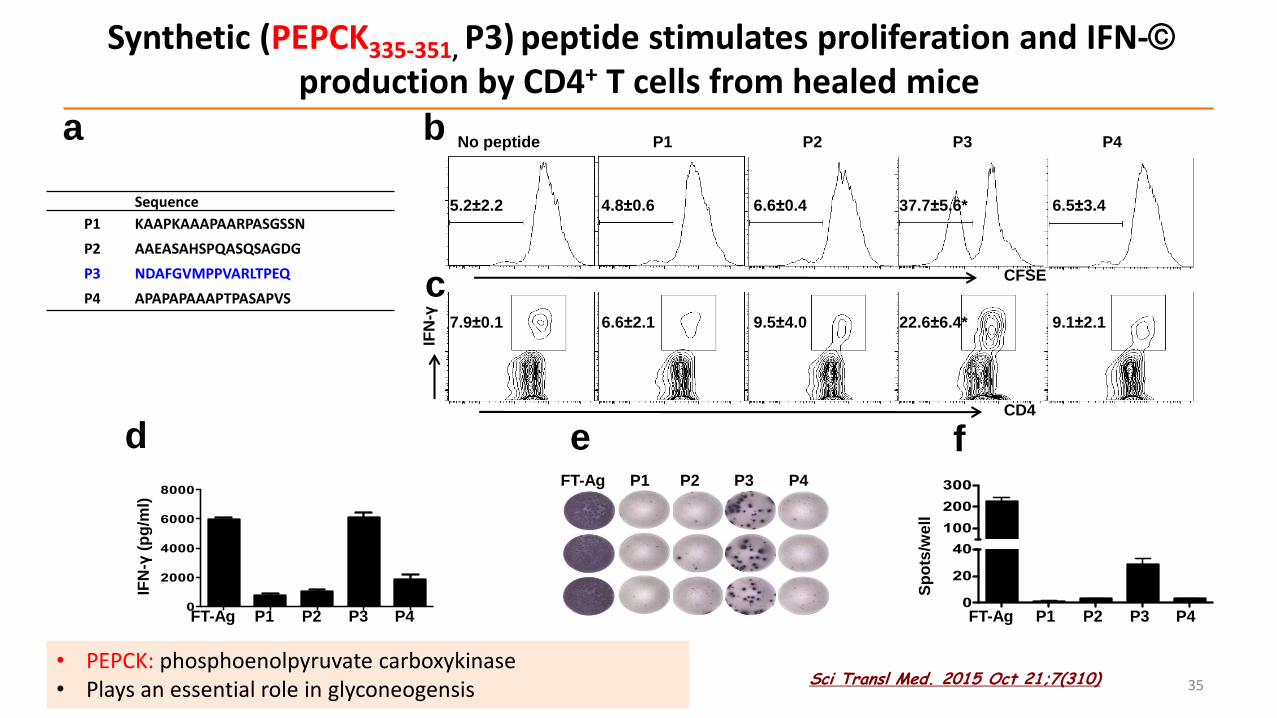
eP1 P2 P3 P4FT-Ag
CFSE
No peptide P1 P2 P3 P4
IFN
-γ
b
c7.9±0.1 6.6±2.1 9.5±4.0 22.6±6.4* 9.1±2.1
5.2±2.2 4.8±0.6 6.6±0.4 37.7±5.6* 6.5±3.4
d
0
2000
4000
6000
8000
FT-Ag P1 P2 P3 P4
IFN
-γ(p
g/m
l)
Sequence
P1 KAAPKAAAPAARPASGSSN
P2 AAEASAHSPQASQSAGDG
P3 NDAFGVMPPVARLTPEQ
P4 APAPAPAAAPTPASAPVS
a
CD4
FT-Ag P1 P2 P3 P4
Sp
ots
/we
ll
f
0
20
40
100
200
300
Synthetic (PEPCK335-351, P3) peptide stimulates proliferation and IFN-production by CD4+ T cells from healed mice
35
• PEPCK: phosphoenolpyruvate carboxykinase• Plays an essential role in glyconeogensis
Sci Transl Med. 2015 Oct 21;7(310)
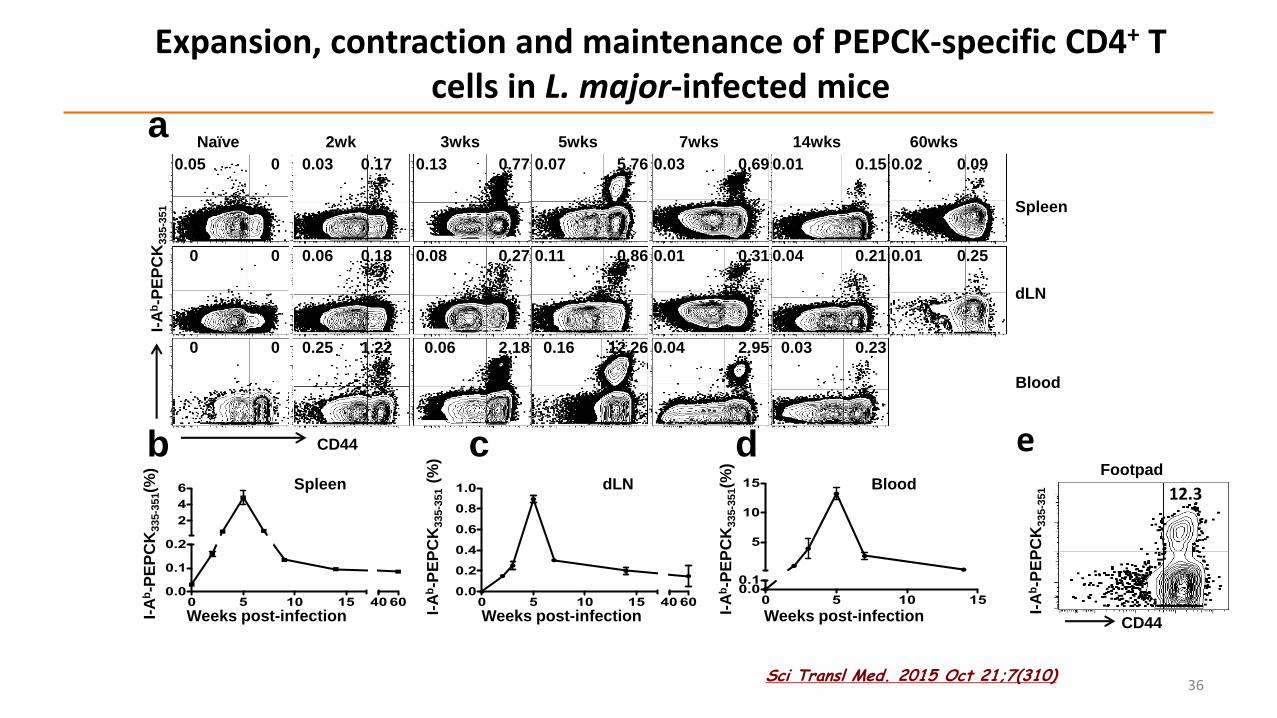
Expansion, contraction and maintenance of PEPCK-specific CD4+ T cells in L. major-infected mice
a
I-A
b-P
EP
CK
335-3
51
CD44
Footpad
Naïve 2wk 5wksI-
Ab-P
EP
CK
335-3
51
3wks 7wks
dLN
Spleen
14wks 60wks
0 0
0.05 0
0.06 0.18
0.03 0.17
0.08 0.86
0.13 5.76
0.01 0.31
0.03 0.69
0.04 0.21
0.01 0.15
0.01 0.25
0.02 0.09
0.27
0.77
0.11
0.07
CD44
Blood
0 0 0.25 1.22 0.06 2.18 0.16 12.26 0.04 2.95 0.03 0.23
0 5 10 150.00.1
5
10
15
0 5 10 15 40 600.0
0.2
0.4
0.6
0.8
1.0
0 5 10 15 40 600.0
0.1
0.2
2
4
6 Spleen
bdLN
Weeks post-infectionWeeks post-infection
cBlood
Weeks post-infection
d
I-A
b-P
EP
CK
335-3
51(%
)
I-A
b-P
EP
CK
335-3
51
(%)
I-A
b-P
EP
CK
335-3
51(%
)
e
12.3
36Sci Transl Med. 2015 Oct 21;7(310)
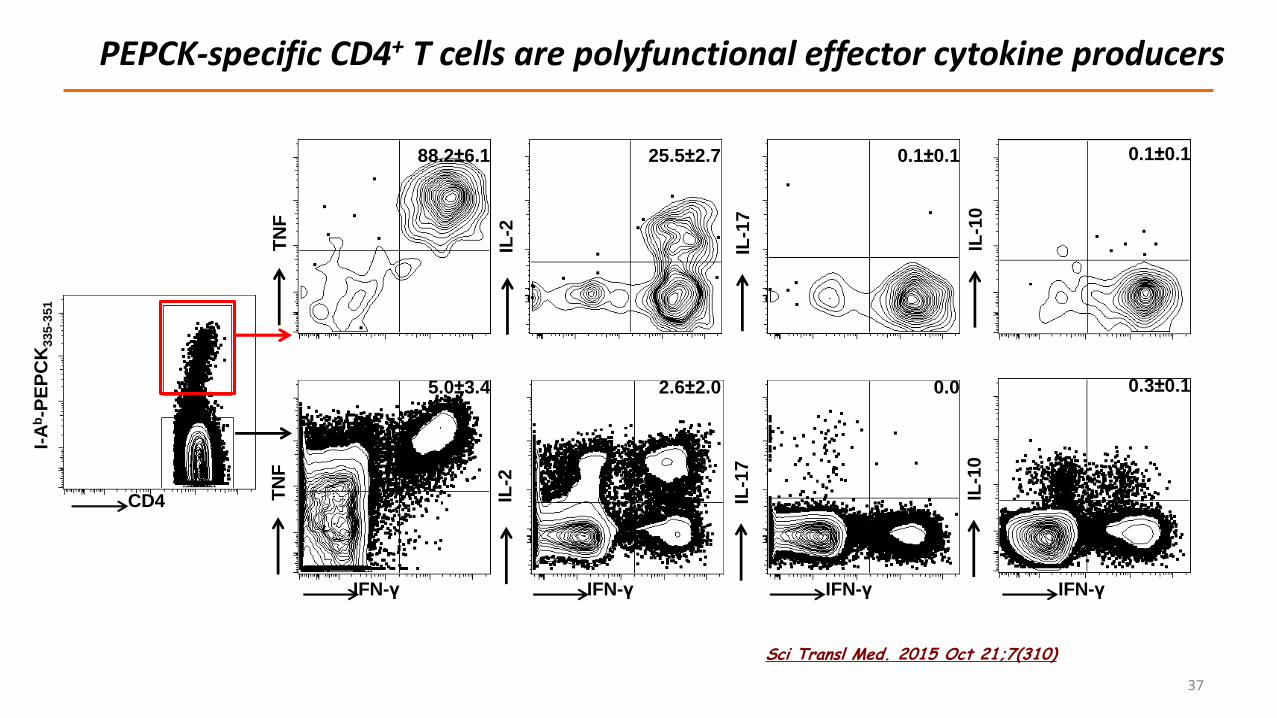
PEPCK-specific CD4+ T cells are polyfunctional effector cytokine producers
37
I-A
b-P
EP
CK
335-3
51
CD4 IL-2
TN
F
IL-1
7
88.2±6.1
5.0±3.4
25.5±2.7
2.6±2.0
0.1±0.1
0.0
IFN-γ IFN-γ IFN-γ
IL-1
0
IFN-γ
0.1±0.1
0.3±0.1
IL-2
TN
F
IL-1
7
IL-1
0
Sci Transl Med. 2015 Oct 21;7(310)
6
7
8
9
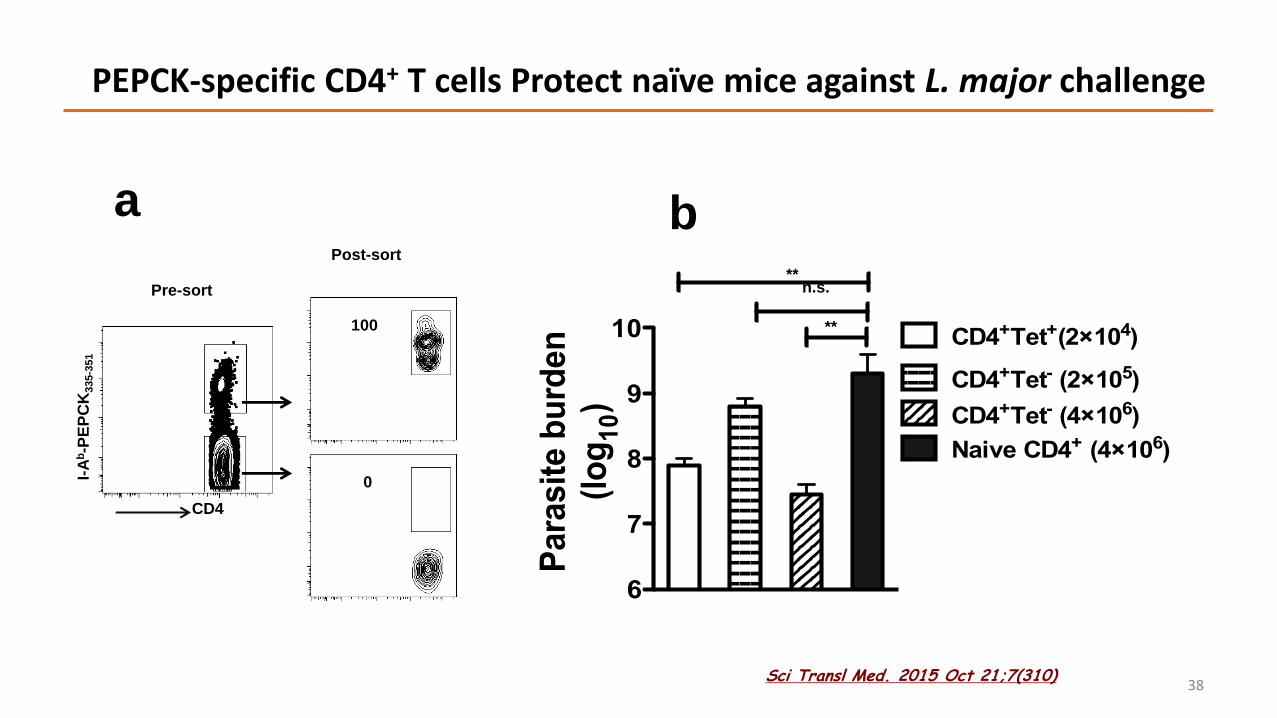
10 CD4+Tet+(2×104)
CD4+Tet- (2×105)
CD4+Tet- (4×106)
Naive CD4+ (4×106)
Para
sit
e b
urd
en
(lo
g1
0)
**n.s.
**
a
Pre-sort
I-A
b-P
EP
CK
335-3
51
CD4
Post-sort
b
100
0
PEPCK-specific CD4+ T cells Protect naïve mice against L. major challenge
38Sci Transl Med. 2015 Oct 21;7(310)
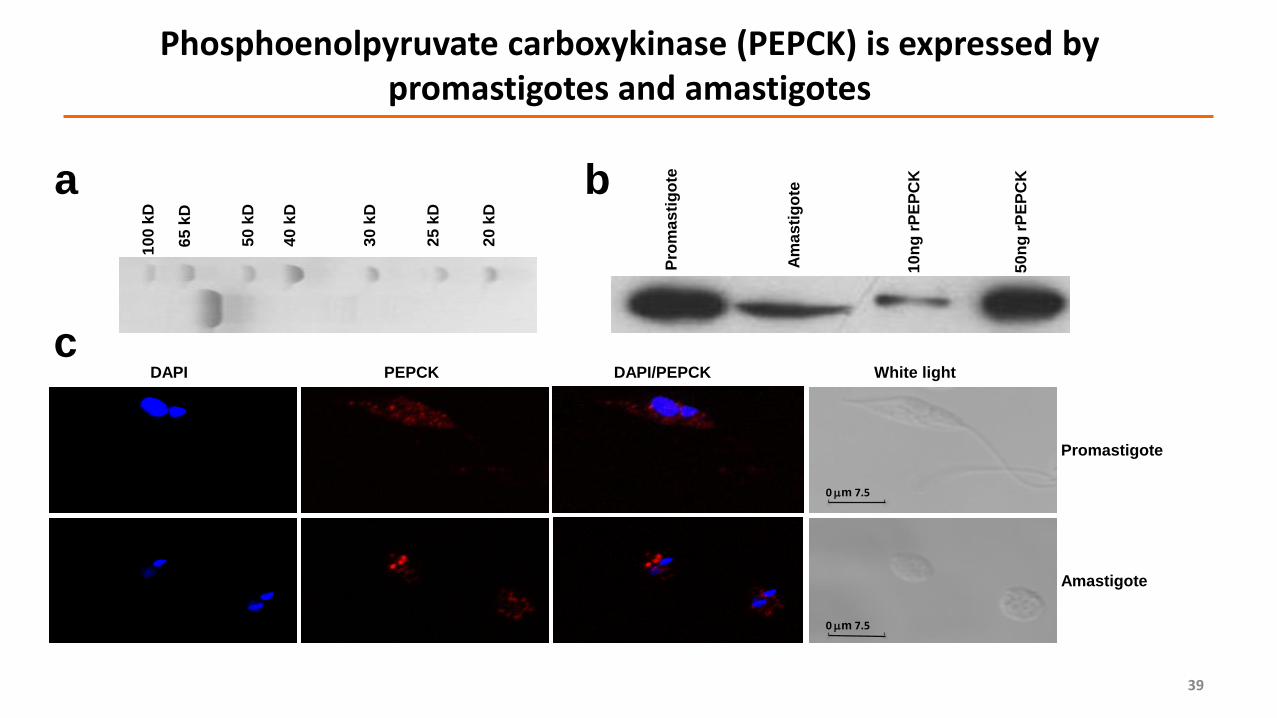
Pro
ma
sti
go
te
Am
as
tig
ote
10
ng
rP
EP
CK
50
ng
rP
EP
CKa
DAPI PEPCK DAPI/PEPCK White light
c
0 mm 7.5
0 mm 7.5
Promastigote
Amastigote
10
0 k
D
65
kD
50
kD
40
kD
30
kD
25
kD
20
kD
b
Phosphoenolpyruvate carboxykinase (PEPCK) is expressed by promastigotes and amastigotes
39
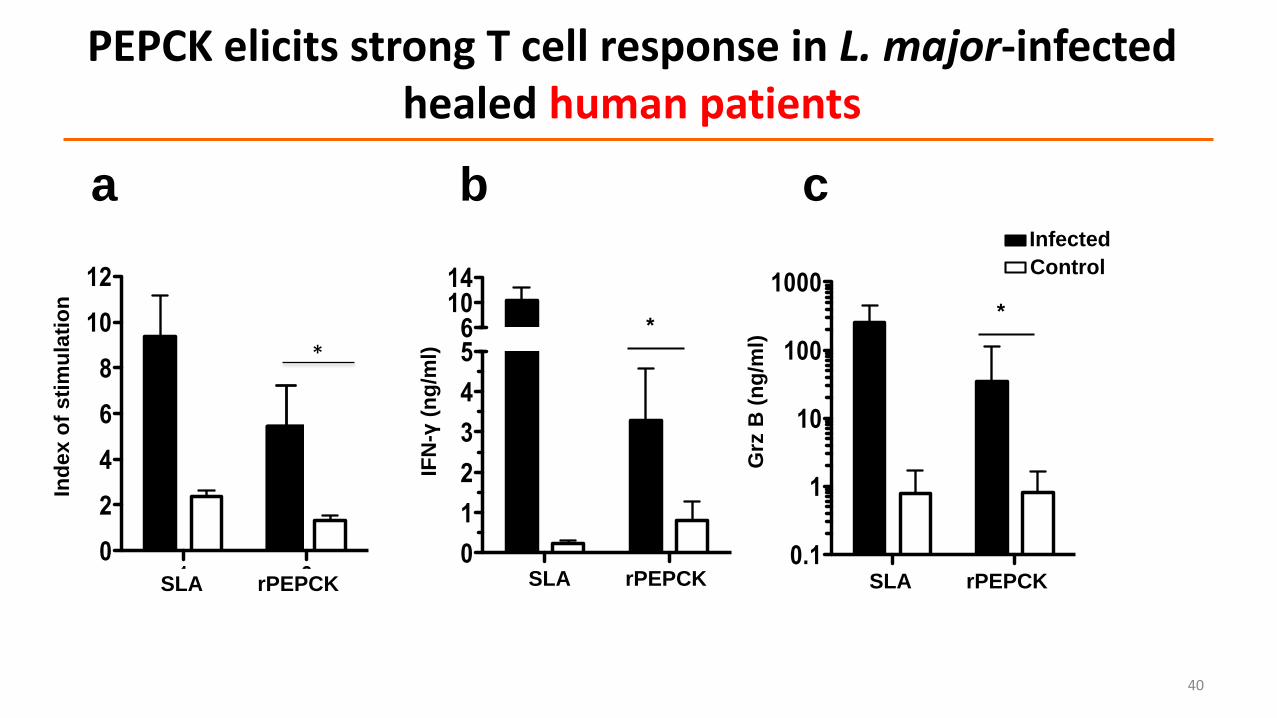
PEPCK elicits strong T cell response in L. major-infected healed human patients
Control
Infected
40
1 20
1
2
3
4
56
1014
a b c
1 20
2
4
6
8
10
12
SLA rPEPCK
Ind
ex o
f sti
mu
lati
on
SLA rPEPCK
IFN
-γ(n
g/m
l)
*
1 20.1
1
10
100
1000
SLA rPEPCK
Grz
B (
ng
/ml)
*
1 20
2
4
6
8
10
12*

Challenge:L. major
L. donovani
Immune
response
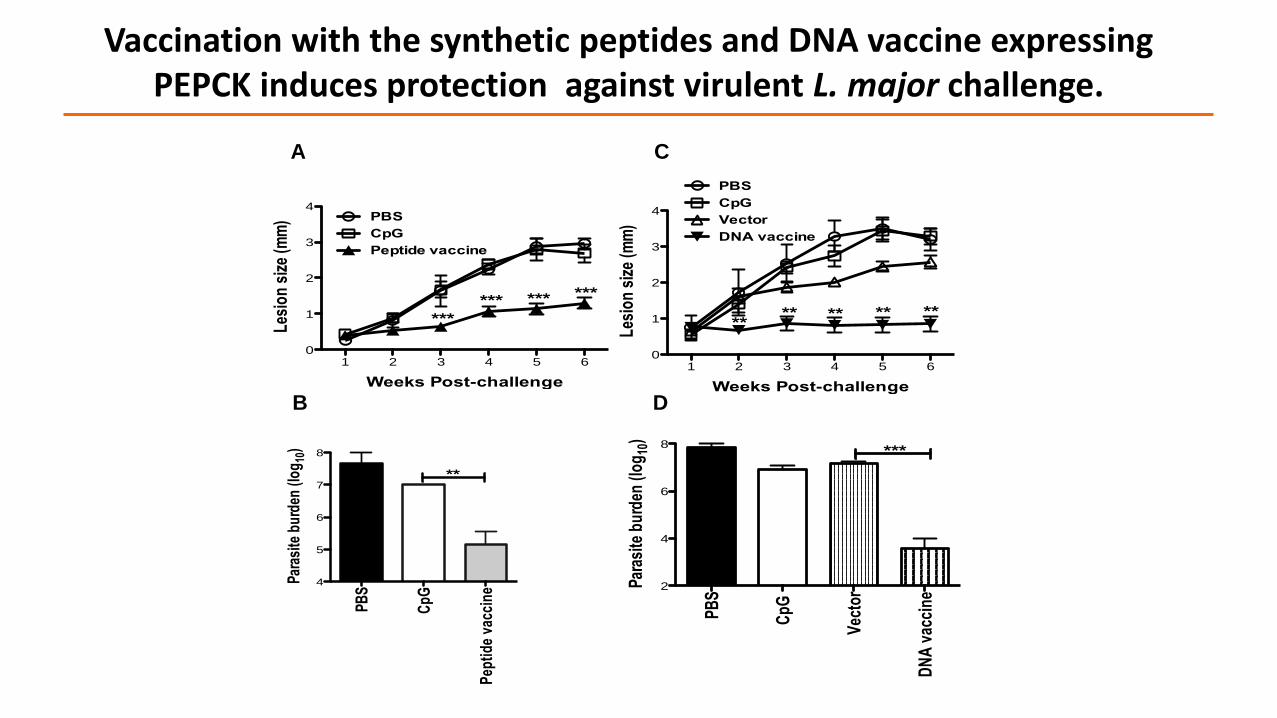
Peptide Vaccine:
1) PBS
2) CpG control
3) Peptide + CpG
Does Vaccination with PEPCK Protect Against Virulent Challenge?
DNA Vaccine:
1) PBS
2) CpG control
3) DNA+ CpG
4) Vector + CpG
Protein Vaccine:
1) PBS
2) CpG control
3) Protein + CpG
4) Protein only
0 1 2 3 4 5 6 8 10wks
12
Challenge:L. major
1 2 3 4 5 60
1
2
3
4PBS
CpG
Peptide vaccine
****** *** ***
Weeks Post-challenge
Les
ion
siz
e (m
m)
B
A
D
C
PB
S
CpG
Vec
tor
DN
A v
acci
ne
2
4
6
8***
Par
asite
bu
rden
(lo
g10
)
PB
S
CpG
Pep
tide
vac
cin
e
4
5
6
7
8
**
Par
asite
bu
rden
(lo
g10
)
1 2 3 4 5 60
1
2
3
4
PBS
CpG
Vector
DNA vaccine
**** ** ** **
Weeks Post-challenge
Les
ion
siz
e (m
m)
Vaccination with the synthetic peptides and DNA vaccine expressing PEPCK induces protection against virulent L. major challenge.
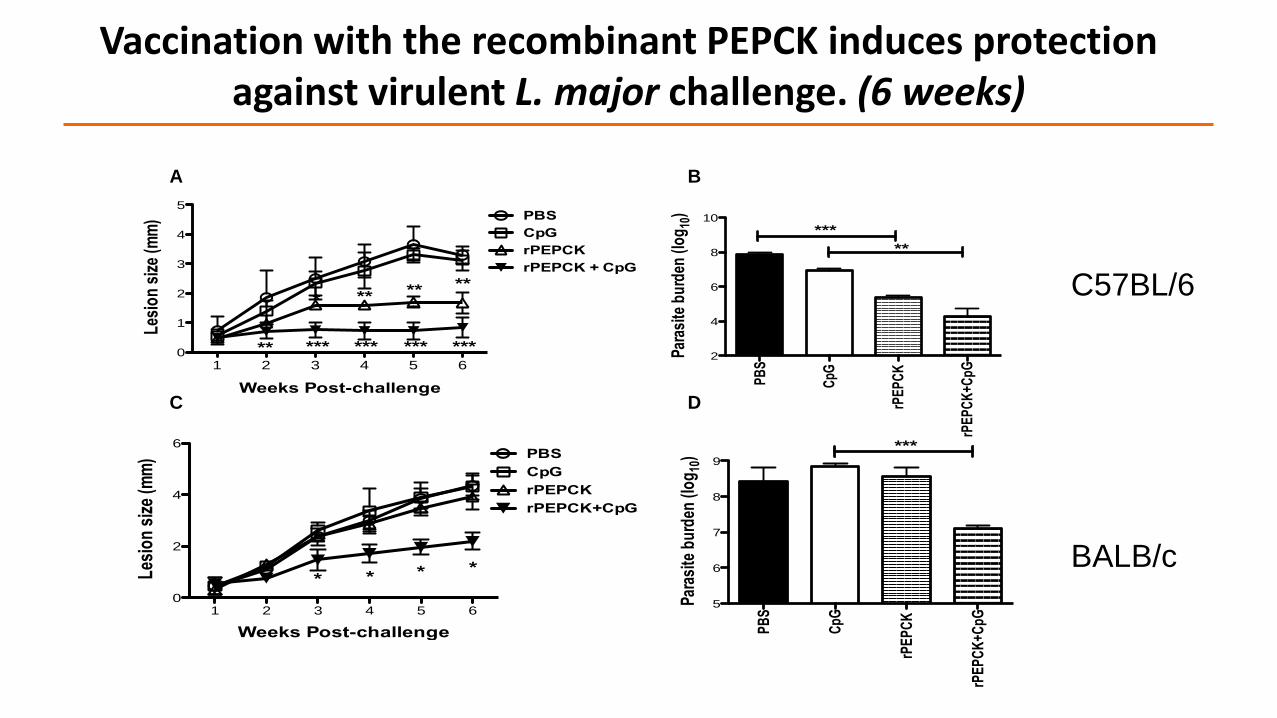
C57BL/6
PB
S
CpG
rPE
PC
K
rPE
PC
K+C
pG
2
4
6
8
10
***
**
Par
asite
bu
rden
(lo
g10
)
BALB/c
1 2 3 4 5 60
1
2
3
4
5PBS
CpG
rPEPCK
rPEPCK + CpG
* ** ** **
*** *** *** *****
Weeks Post-challenge
Les
ion
siz
e (m
m)
PB
S
CpG
rPE
PC
K
rPE
PC
K+C
pG
5
6
7
8
9
Par
asite
bu
rden
(lo
g10
)
***
1 2 3 4 5 60
2
4
6PBS
CpG
rPEPCK
rPEPCK+CpG
***
*
Weeks Post-challenge
Les
ion
siz
e (m
m)
A B
C D
Vaccination with the recombinant PEPCK induces protection against virulent L. major challenge. (6 weeks)
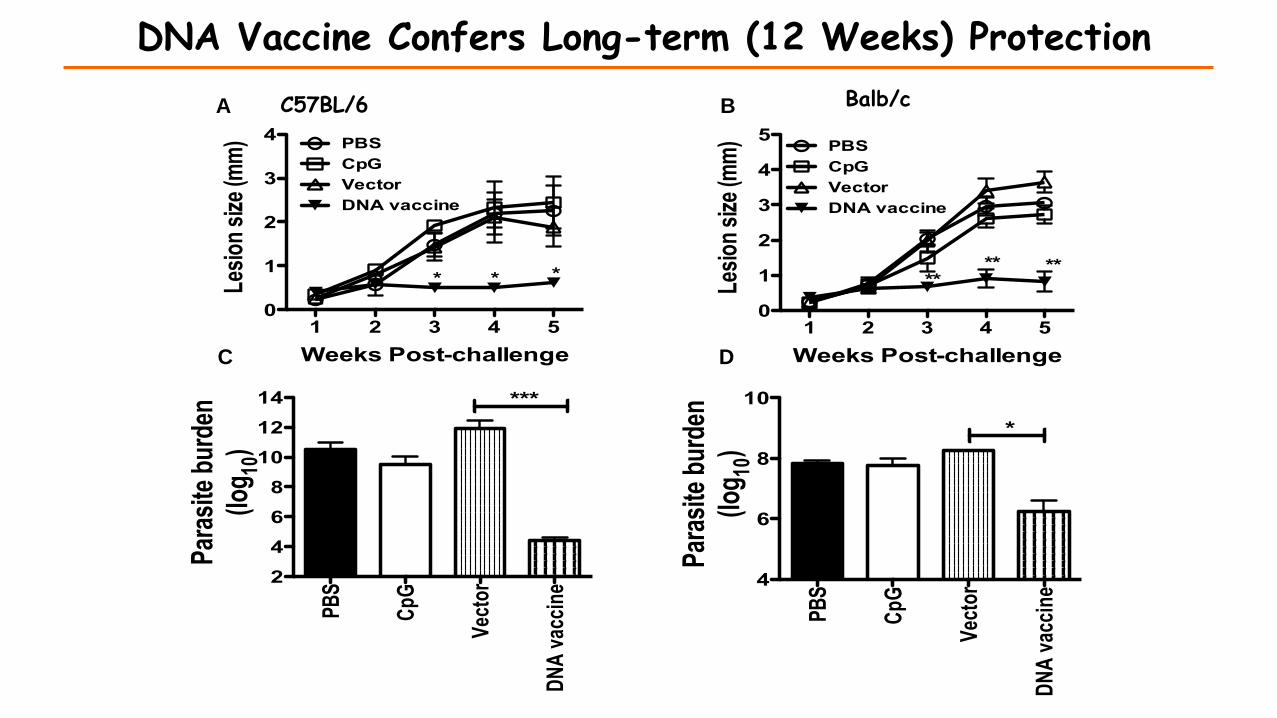
DNA Vaccine Confers Long-term (12 Weeks) Protection
Balb/c
1 2 3 4 50
1
2
3
4
5PBS
CpG
Vector
DNA vaccine
**** **
Weeks Post-challenge
Lesi
on s
ize
(mm
)
PB
S
CpG
Vec
tor
DN
A v
acci
ne
4
6
8
10
*
Par
asite
bur
den
(log 10
)
C57BL/6
1 2 3 4 50
1
2
3
4PBS
CpG
Vector
DNA vaccine
* * *
Weeks Post-challenge
Lesi
on s
ize
(mm
)
PB
S
CpG
Vec
tor
DN
A v
acci
ne2
4
6
8
10
12
14 ***
Par
asite
bur
den
(log 10
)
C
A
D
B
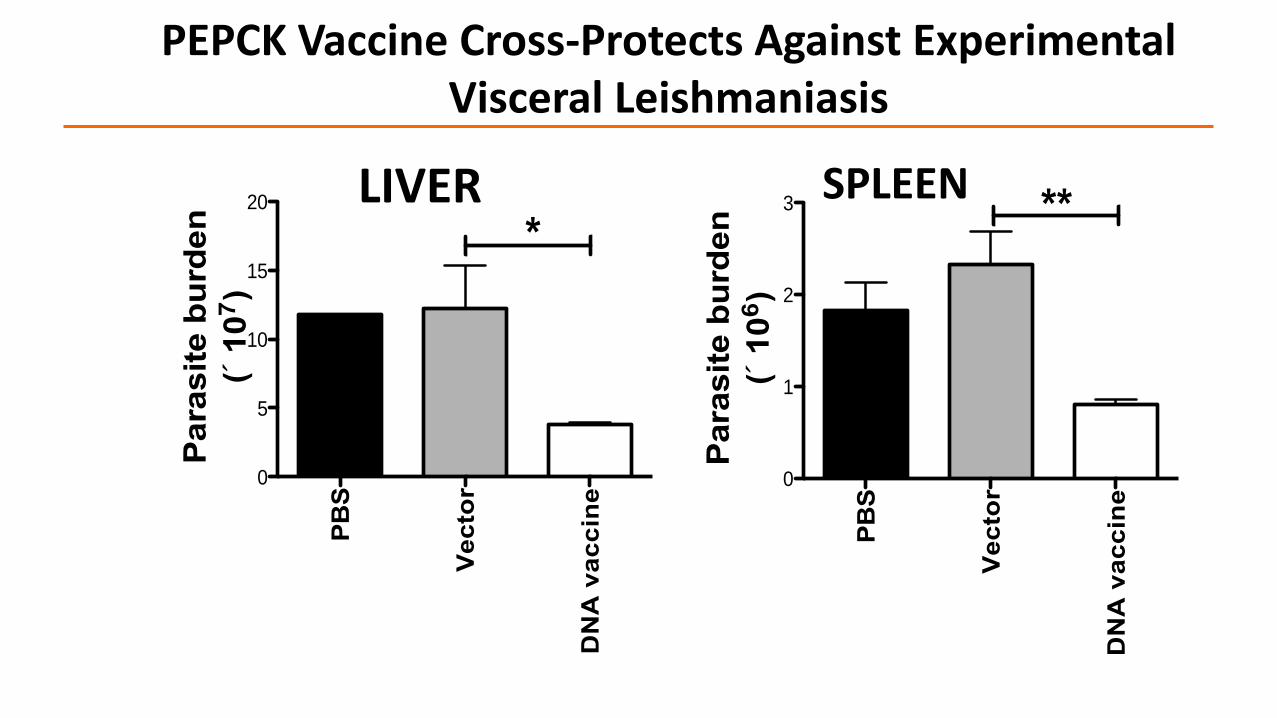
PEPCK Vaccine Cross-Protects Against Experimental Visceral Leishmaniasis
PB
S
Ve
cto
r
DN
A v
ac
cin
e
0
5
10
15
20
*P
ara
sit
e b
urd
en
(´10
7)
PB
S
Ve
cto
r
DN
A v
ac
cin
e
0
1
2
3 **
Para
sit
e b
urd
en
(´10
6)
LIVER SPLEEN

Naturally processed Leishmania major peptides were eluted and identified from MHCII molecules on L. major -infected BMDCs
PEPCK-specific T cells are immunodominant, polyfunctional cytokine producers andprotect against L. major
Vaccination with PEPCK peptides, PEPCK DNA or rPEPCK induces cross-protectionagainst Leishmania challenges
L. major, L. donovani, L. infantum (preliminary evidence)
Summary:
46

Implications for Vaccine Designs and Vaccination Strategies
• Conventional methods of immunogen prediction may be misleading
In silico predictions, early versus memory immune responses
• Approach aimed at identifying antigens that are really relevant
Reverse immunology
• May be relevant in other parasitic infections where concomitant immunity is critical for resistance
Malaria, toxoplasmosis

Next Step:
• Vaccination studies in NHP, Dogs
• Clinical Trials?
• Collaborators and funding wanted!!!

Acknowledgements
Project Leader:
Dr. Zhirong Mou
Uzonna Lab membersJintao LiForough Khadem
Manitoba centre for Protemics and System BiologyDr. John WilkinsPeyman Ezzati
Third Military Medical University, ChinaChuanmin Hu; Weijing Yi; Shufeng Wang
Flow cytometry Core FacilityChristine Zhang
Cancer Care ManitobaLudger Klewes
Collaborators:
Funding:
Pasteur Institute of Tunis, TunisiaThouraya BoussoffaraHechmi Louzir
University of Toyama , JapanHiroyuki KishiHiroshi Hamana
McGill UniversityMomar Ndao
Sichuan University, ChinaJianping Chen
Pasteur Institute of Iran, IranSima Rafati
University of IowaChristine Petersen
49