a cost-benefit assessment of two multi-species … 29(1)/001 thalassas 29(1).pdf · crecimiento y...
TRANSCRIPT

A COST-BENEFIT ASSESSMENT OF TWO MULTI-SPECIES ALGAE DIETS FOR JUVENILES OF
Mytilus galloprovincialis
(1) Université Abdelmalek Essaâdi, Faculté des Sciences, Laboratoire de Biologie Appliquée et Pathologie,BP. 2121, Mhannech 2, 93002 Tétouan, Morocco
(2) University of Vigo, Department of Biochemistry, Genetics and Immunology,ECIMAT-Faculty of Marine Sciences, Illa de Toralla, 36331 Vigo, Spain
*Corresponding author: Pablo PresaUniversity of Vigo. Dep. Biochemistry, Genetics and Immunology, 36310 Vigo, Spain
Tel. & Fax: +34-986-812567. E-mail: [email protected]
Thalassas, 29(1) · January 2013: 9-16An International Journal of Marine Sciences
Key words: Chaetoceros gracilis, cost-benefi t assessment, multi-species algae diets, Mytilus galloprovincialis, Phaeodactylum tricornutum.
Palabras clave: Chaetoceros gracilis, costes de producción, dietas microalgales multiespecífi cas, Mytilus galloprovincialis, Phaeodactylum tricornutum.
A. AGHZAR(1), M. MIÑAMBRES(2), P. ALVAREZ(2) & P. PRESA(2)*
9Thalassas, 29(1) · January 2013
ABSTRACT
The optimisation of productivity in mussel hatchery requires the assessment of multi-species algae diets to enhance growth and survival during retention time. The present study monitored shell length in Mytilus galloprovincialis juveniles using diets of four micro-algae, Tetraselmis suecica, Isochrysis galbana, Phaeodactylum tricornutum and Chaetoceros gracilis. The experiment lasted 116 days to assess shell length response, used a full-sib mussel progeny to minimise familiar effects on growth variance, and enforced a mini-raft suspension system to assure equal food deliverance to all juveniles under treatment. Shell-length increment in the 26% C. gracilis-based diet was not different from the 26% P. tricornutum-based diet except in two samplings. None differences between diets were observed neither in the growth trend nor in survival (99%). Present data indicate that multi-species algae diets incorporating C. gracilis have a slight non-significant positive effect on growth of M. galloprovincialis juveniles and that such growth gain is not compensated by its higher production cost (4 times) as compared to diets composed by cheaper and easy-culturing species such as P. tricornutum. The global cost required to fill a whole raft (ca 200 ropes) using two-month old hatchery-produced mussel juveniles (One month retention time after fixation i.e. juveniles 2,000 μm in length) fed P. tricornutum was less (1,892 €/raft) than that fed C. gracilis (7,551 €/raft) or its equivalent from rocky scrapping (2,000 €/raft).
RESUMEN (Evaluación de costes y rendimiento de dos dietas microalgales multiespecíficas para juveniles de Mytilus galloprovincialis)
La optimización de la producción de semilla de mejillón en criadero requiere la evaluación de dietas multiespecificas para mejorar el crecimiento y la supervivencia durante la fase de retención. En este trabajo se efectúa el seguimiento de la longitud valvar de juveniles de Mytilus galloprovincialis alimentados con dietas compuestas de cuatro cepas microalgales, i.e. Tetraselmis suecica, Isochrysis galbana, Phaeodactylum tricornutum y Chaetoceros gracilis. El experimento duró 116 días para valorar la respuesta de crecimiento, se empleó una progenie de hermanos completos para minimizar la varianza de crecimiento debida a efectos familiares, y se utilizó un sistema experimental de minibateas en suspensión que aseguró la distribución homogénea de alimento entre los juveniles tratados. El incremento en longitud valvar del grupo sometido a la dieta que contenía 26% de C. gracilis sólo difirió del de la dieta con 26% de P. tricornutum en dos muestreos. No se observaron diferencias entre dietas, ni en la curva de crecimiento ni en la supervivencia juvenil (99%). Estos resultados indican que las dietas multialgales que incorporan C. gracilis provocan un efecto positivo en el crecimiento de juveniles de mejillón pero no es significativo al com-pararlo con dietas similares. Además este efecto positivo de C. gracilis sobre el crecimiento no está compensado por un mayor rendimiento del stock cultivado, pues su coste de producción es 4 veces mayor que el de dietas multialgales más baratas y fáciles de cultivar, tal como las basadas en la especie P. tricornutum. El coste estimado para inocular una batea de cultivo con 200 cuerdas, con juveniles de 2 meses de vida (2.000 μm de longitud), producidos y alimentados en criadero con P. tricornutum (1 mes de retención tras la fijación) es de 1.892 €/batea, i.e. cuatro veces menos que los alimentados con C. gracilis (7.551 €/batea), y menor que su equivalente procedente de la compra de semilla en el mercado (2.000 €/batea).

A. AGHZAR, M. MIÑAMBRES, P. ALVAREZ & P. PRESA
INTRODUCTION
Live microalgae are believed to provide the most important nutrients for bivalves in the wild because their shortage trends to prolong the planktonic larval phase, increasing cohort mortality due to predation (Rumrill, 1990), and to negatively affect spat recruitment in artificial rope collectors. This phenomenon has been observed in several bivalves. For example, Mytilus californianus larvae have shown limited growth and delayed development under unsuitable food supply (Paulay et al., 1985; Pechenik et al., 1990). Mytilus edulis veligers have shown limited growth due to food supply scarcity in the embayment during summer (Fotel et al., 1999). With the advancement on bivalve reproduction techniques and juveniles pre-fattening in nurseries, the culture of live microalgae has become a key requisite for massive production of juveniles (Page & Hubbard, 1987). Microalgae are used to feed larvae, post-larvae and juveniles as well as in broodstock conditioning, and their production represents 30% - 40% of hatchery operation costs (Coutteau & Sorgeloos, 1992; Helm et al., 2004). Therefore, optimisation of microalgae diets is a commonly sought parameter in bivalve hatchery production for larvae and juveniles (Liu et al., 2009). Bivalve larvae and juveniles are often fed multispecies algal diets because they exhibit better balancing of the essential nutritional compounds with particular reference to polyunsaturated fatty acids, and assure higher growth and survival than single species diets (Enright et al., 1986; Laing & Millican, 1986; De Pauw & Persoone, 1988; Nevejan et al., 2007).
Previous studies have shown growth enhancement of juveniles of some bivalves fed fresh microalgae (Ponis et al., 2003; Cragg 2006; Liu et al., 2009) belonging to several genera such as Tetraselmis, Isochrysis, Phaeodactylum, Chaetoceros, Pavlova, Dunaliella, Thalassiosira and Skeletonema (Helm et al., 2004). The food value of T. suecica is considered low possibly because of the presence of a rigid cell wall which might make it difficult to digest (Rico-Mora, 1987). However, T. suecica is an appreciated species because of its high ratio of protein content/cell volume as compared to that of many other species used in aquaculture. I. galbana has shown some technical problems in large-scale cultures (Coeroli et al., 1984), has no advantage over P. tricornutum, and is a poorer food than C. gracilis. However, I. galbana is an excellent food for many filter-feeders (Sukenik & Wahnon, 1991), especially at larval stages (Liu et al., 2009). P. tricornutum is not generally considered a good food source although it is easy to produce and has been successfully used for growing oyster larvae (Epifanio et al., 1981) and for feeding juveniles of clams and mussels (Foster-Smith,
1975). A higher growth has been observed in bivalves and copepods fed Chaetoceros sp. than fed Phaeodactylum, the latter microalgae having a reduced nutritional value of 12% of lipids against 19% of the former (Helm et al., 2004; Liu et al., 2009; Puello-Cruz et al., 2009). However, Chaetoceros sp. is more difficult and expensive to maintain in large volumes than Phaeodactylum. C. gracilis is one of the easier Chaetoceros species for culture and is widely used in larger volumes around the world (Lambade & Mohamed, 2002).
The present study aimed at assessing shell-length growth in M. galloprovincialis juveniles in nursery using multispecific diets of the above cited microalgae species, in order to address if the expected higher juvenile growth fed C. gracilis (Medium difficulty cultivation and high energetic efficiency, rich in the PUFA 20:5ω3) compensates its higher production costs against the cheaper species P. tricornutum (Low cultivation difficulty and medium-low energetic efficiency but rich in 22:6ω3). To properly estimate the cost-benefit ratio between diets, several growing parameters were fixed, such as the use of a full-sib progeny of M. galloprovincialis to minimise familiar effects of mixed progenies on growth variance, the implementation of an in vitro mini-raft suspension system that minimises differences in food availability by all juveniles under treatment, or an experimental time of 116 days to properly assess shell length response under different diets.
MATERIALS AND METHODS
On October 15, 2007 a full-sib progeny of M. galloprovincialis was generated in ECIMAT Marine Station (University of Vigo) from wild genitors collected at Ría de Vigo (NW Spain). A random sample of 11,000 3.5-month-old juveniles from this progeny was reared under two experimental microalgae diets and a 50-μm filtered seawater control, lasting from January 26to May 29 in 2008. Five PVC tanks (two for each experimental diet and one for the control) with a dimension of 40 cm length x 30 cm width x 32 cm depth (Fig. 1a), equipped with small growing ropes, were used for the trial. The progeny had been allowed to settle on 13 ropes of 20 cm length at an initial density of approximately 850 individuals per rope (Fig. 1b). The 13 inoculated ropes (six per diet (three per tank), and one for control) were placed into experimental tanks using horizontal PVC sticks (mini-rafts, Fig. 1b). There were no significant differences in initial shell length or weight between the juveniles randomly distributed among ropes (Table 1).
Cultured microalgae consisted of Isochrysis galbana (Class Prymnesiophyceae; cell of strain # CCMP1323), Tetraselmis suecica (Class Prasinophyceae; cell of
10 Thalassas, 29(1) · January 2013

Fig 1
A COST-BENEFIT ASSESSMENT OF TWO MULTI-SPECIES ALGAE DIETS FOR JUVENILES OF Mytilus galloprovincialis
strain # CCMP904), Phaeodactylum tricornutum (Class Bacillariophyceae; Ecimat collection cell of strain # CCMP633) and Chaetoceros gracilis (Class Coscinodiscophyceae; cell of strain # CCMP1317). These strains were grown in six litres flasks in a batch system at 20±1ºC and a continuous lighting intensity of 6,000 luxes from three cool daylight fluorescent tubes. Flasks were bubbled and enriched with Walne’s medium, and pH was maintained between 7.4 and 8.3 by unmetered, continuous CO2 injection. Collection of cells for feeding was made at the exponential growth stage, and cell number per microalgae culture was estimated daily using a Neubauer counting chamber and a microscope.
The two experimental multispecies diets conformed as follows: Diet-A was composed by I. galbana and T. suecica (37% or 60 mg of the equivalent wet weight for each species) and P. tricornutum (26% or 43 mg of the equivalent wet weight); Diet-B was mostly composed as Diet-A andbut contained C. gracilis (26% or 43 mg of the equivalent wet weight) instead of P. tricornutum; instead of C. gracilis (26% or 43 mg of the equivalent wet weight); Diet-C orwas the control dietgroup and received a constant supply of 50 μm filtered seawater. The average number of cells per millilitre at the exponential culturing phase was 12,000,000 for I. galbana, 2,300,000 for T. suecica, 15,000,000 for P. tricornutum, and 3,000,000 for C. gracilisTo achieve a food equivalent of 20% of the average body wet weight of mussels, the amount of live microalgae delivered to each experimental tank contained 163 mg of wet weight day-1
For daily diet adjustment we used the cell organic weight established for C. gracilis (30 μg per 106 cells) and
P. tricornutum (23 μg per 106 cells) (Helm et al., 2004) and applied the expression (See Miñambres et al., 2011 for details):
VD = (C* Vc)/D)*fc where,
VD = deliverable volume of C. gracilisVc=Vt*0.25= deliverable volume of P. tricornutum (25%)C= No. cell/ml P. tricornutum as estimated in daily counting D= No. cell/ml C. gracilis as estimated in daily countingfc=23/30=0.77= conversion factor from P. tricornutum to C. gracilis
Final microalgae formulae was diluted to 60 L with 50 μm filtered seawater for better dosing and both diets were continuously pumped at 0.5-0.6 mL s-1. The tank for the control (Diet-C) received a constant supply of 50 μm filtered seawater (the same used to prepare Diet-A and Diet-B). All tanks received a water inflow rate of 40 mL s-1 (complete volume replacement every 24 min). Diet-C received an average phytoplankton amount from sea water of 2.0 mg DW day-1 in the course of the experiment. Experiments were conducted at 18±2ºC and tanks were aerated during the experiment to provide oxygen and to prevent microalgae cell sedimentation. Tanks were cleaned and disinfected three times a week to remove faeces and adherent particles and juveniles were rinsed with a gently spray of seawater. The absence of pseudofaeces and the presence of well defined faeces at the bottom of the tank indicated an efficient consumption of the whole diet delivered. Growth was calculated in terms of shell length increase (μm day-1). Individual shell length was measured every 15 days on a
11Thalassas, 29(1) · January 2013
b a
Figure 1:PVC tanks used for mini-raft simulation in the diet trial (Panel a); 20 cm-length culture ropes,
each containing 850 juveniles from a full-sib progeny (Panel b)

A. AGHZAR, M. MIÑAMBRES, P. ALVAREZ & P. PRESA
subsample of 50 individuals randomly taken from each mini-raft feeding. Shell-length measurements were taken with an image analysis system (Nis elements BR 3.0) connected to a magnifying glass (Nikon SMZ 1500). Three months after the beginning of the experiment, shell size of mussels in the experimental diets was large enough to replace the optical measurement system by a digital calliper with 0.01 mm accuracy (Mitutoyo IP67).
Normality length and weight per diet and sample was explored with Kolmogorov-Smirnov test. Shell-length measurements taken on serial replicates within diet were compared with a Mann-Whitney test. Comparison of average shell length between diets at each sampling as well as between temporal samplings within diet, were performed with the Kruskall-Wallis test, the ANOVA analysis and the ad hoc Scheffée test, all of them implemented in SPSS 17.0. Mortality day-1 was calculated after the number of dead animals (shells) found at the bottom of each experimental tank by the time of shell length control ca. once a week.
The production costs per rope were calculated from the
experimental data and scaled up for industrial implication. In mussel aquaculture,initial 4.5 mm juveniles taken from collector ropes or from rocky scrapping are fixed to growth ropes suspended from rafts normally placed in protected areas such as estuaries. After 3 to 5 months growing in the sea juveniles are unfolded into 2-3 ropes of about 50 kg mussels each. After an additional year of sea growth we obtain 2-3 harvesting ropes of about 150 kg mussels each (30-35 individuals per kg). Discounting 10% mortality in the first growing period we finally require
11,500 juveniles per rope and 2,310,000 juveniles per 200-rope raft. The cost of juveniles produced in nursery for 116 and 30 days of retention was considered for each multispecies diet. In addition, ten kg of juveniles of 5-15 mm length are required to fill a growth rope. Therefore, a 200- rope raft requires 2,000 kg such juveniles. The cost of mussel juveniles from rock-scrapping (1 €/kg) required to fill a raft (2,000€/raft) was also compared to the cost of similar-sized juveniles produced in nursery for 116.
RESULTS
Evolution of shell length within diet
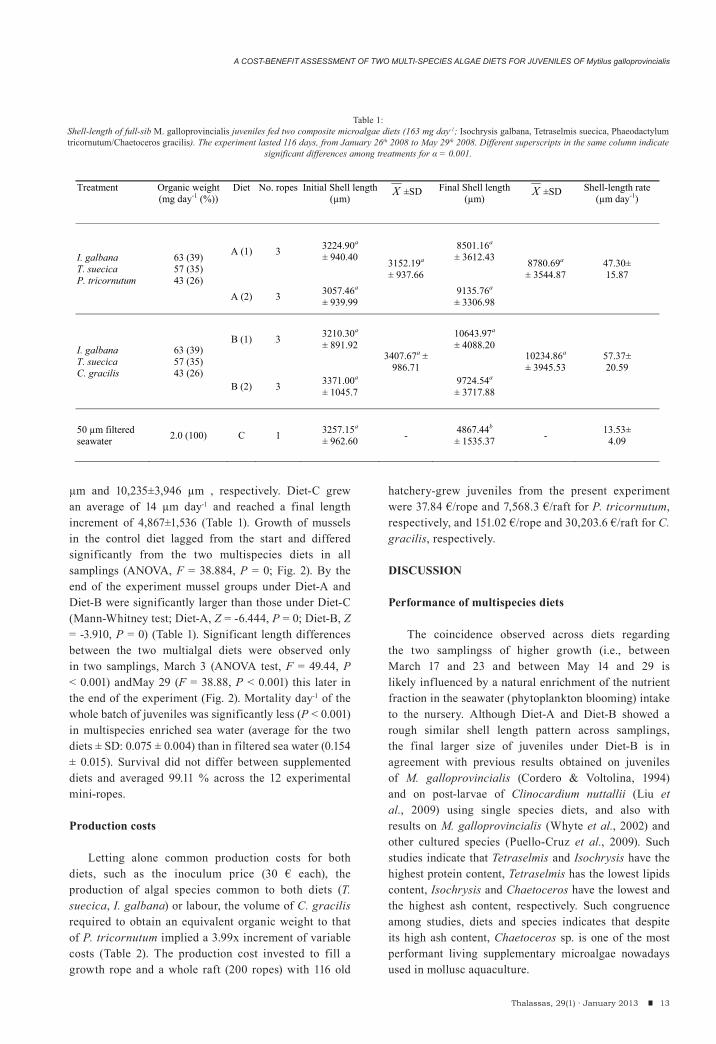
Shell length adjusted to a normal distribution in all serial samplings per diet. No significant weight or length differences were observed between replicates within diet by the end of the trial, i.e. Diet-A (Mann-Whitney test, Z = -2.276, P = 0.023) and Diet-B (Mann-Whitney test, Z = -1.517, P = 0.129) (Table 1). Average shell length differed significantly within both diets between 2008 samplings on March 10 and 23 (Scheffée test; Diet-A, P = < 0.001, Diet-B, P < 0.03), as well as between samplings of April 20 and May 29 (Scheffée test, Diet-A, P < 0.001, Diet-B, P = < 0.001) (Fig. 2). Significant pairwise length differences within Diet-C were observed between samplings March 17 and 23 as well as between samplings May 14 and 29 (Scheffée test, Control Diet, P < 0.001).
Evolution of shell length between diets
Diet-A and Diet-B grew an average of 47 μm day-1 and 57 μm day-1 and reached a final length of 8,781±3,545
12 Thalassas, 29(1) · January 2013
B
A
C
Figure 2:In vitro evolution of shell-length of a 3.5 months-old full-sib progeny during a 116 days retention period. Sampling of juveniles from each multispecies diet was taken at intervals of 1-week (first 53 days) and 2-3 weeks (last 66 days). Diet-A (diamonds), differentially fed 26% Phaeodactylum tricornutum; Diet-B
(squares), differentially fed 26% Chaetoceros gracilis; Diet-C (triangles), control diet fed 50-μm filtered seawater.
A COST-BENEFIT ASSESSMENT OF TWO MULTI-SPECIES ALGAE DIETS FOR JUVENILES OF Mytilus galloprovincialis
μm and 10,235±3,946 μm , respectively. Diet-C grew an average of 14 μm day-1 and reached a final length increment of 4,867±1,536 (Table 1). Growth of mussels in the control diet lagged from the start and differed significantly from the two multispecies diets in all samplings (ANOVA, F = 38.884, P = 0; Fig. 2). By the end of the experiment mussel groups under Diet-A and Diet-B were significantly larger than those under Diet-C (Mann-Whitney test; Diet-A, Z = -6.444, P = 0; Diet-B, Z = -3.910, P = 0) (Table 1). Significant length differences between the two multialgal diets were observed only in two samplings, March 3 (ANOVA test, F = 49.44, P < 0.001) andMay 29 (F = 38.88, P < 0.001) this later in the end of the experiment (Fig. 2). Mortality day-1 of the whole batch of juveniles was significantly less (P < 0.001) in multispecies enriched sea water (average for the two diets ± SD: 0.075 ± 0.004) than in filtered sea water (0.154 ± 0.015). Survival did not differ between supplemented diets and averaged 99.11 % across the 12 experimental mini-ropes.
Production costs
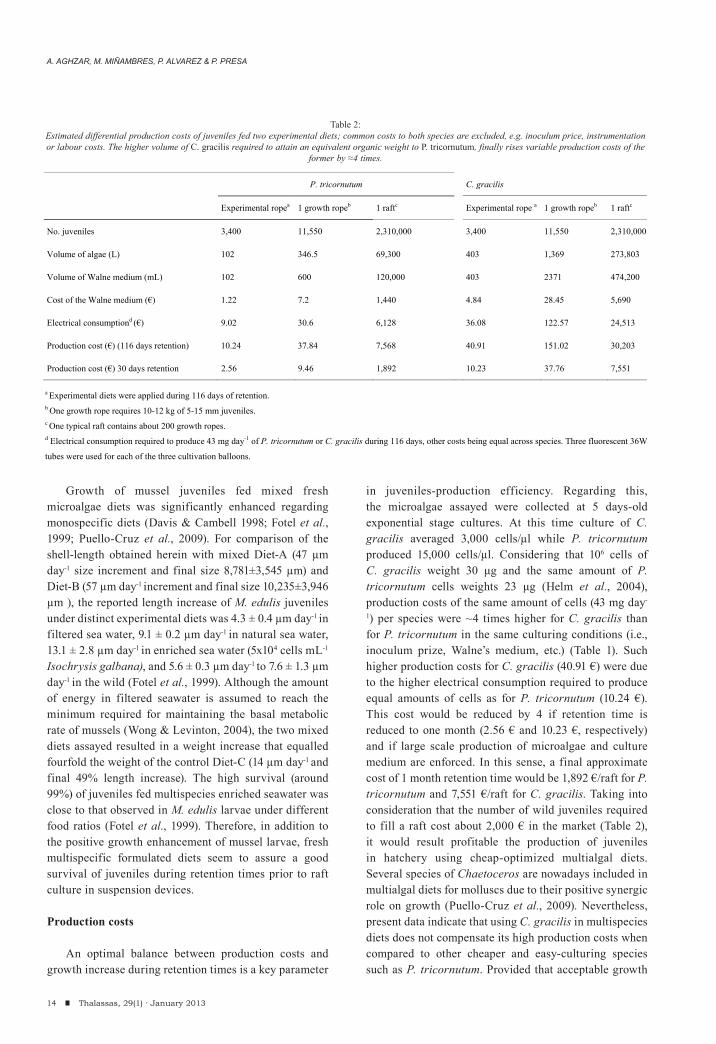
Letting alone common production costs for both diets, such as the inoculum price (30 € each), the production of algal species common to both diets (T. suecica, I. galbana) or labour, the volume of C. gracilis required to obtain an equivalent organic weight to that of P. tricornutum implied a 3.99x increment of variable costs (Table 2). The production cost invested to fill a growth rope and a whole raft (200 ropes) with 116 old
hatchery-grew juveniles from the present experiment were 37.84 €/rope and 7,568.3 €/raft for P. tricornutum, respectively, and 151.02 €/rope and 30,203.6 €/raft for C. gracilis, respectively.
DISCUSSION
Performance of multispecies diets
The coincidence observed across diets regarding the two samplingss of higher growth (i.e., between March 17 and 23 and between May 14 and 29 is likely influenced by a natural enrichment of the nutrient fraction in the seawater (phytoplankton blooming) intake to the nursery. Although Diet-A and Diet-B showed a rough similar shell length pattern across samplings, the final larger size of juveniles under Diet-B is in agreement with previous results obtained on juveniles of M. galloprovincialis (Cordero & Voltolina, 1994) and on post-larvae of Clinocardium nuttallii (Liu et al., 2009) using single species diets, and also with results on M. galloprovincialis (Whyte et al., 2002) and other cultured species (Puello-Cruz et al., 2009). Such studies indicate that Tetraselmis and Isochrysis have the highest protein content, Tetraselmis has the lowest lipids content, Isochrysis and Chaetoceros have the lowest and the highest ash content, respectively. Such congruence among studies, diets and species indicates that despite its high ash content, Chaetoceros sp. is one of the most performant living supplementary microalgae nowadays used in mollusc aquaculture.
13Thalassas, 29(1) · January 2013
Treatment Organic weight (mg day-1 (%))
Diet
No. ropes Initial Shell length (μm)
X ±SD Final Shell length (μm)
X ±SD Shell-length rate (μm day-1)
I. galbana T. suecica P. tricornutum
63 (39) 57 (35) 43 (26)
A (1) 3 3224.90a ± 940.40 3152.19a
± 937.66
8501.16a ± 3612.43 8780.69a
± 3544.87 47.30± 15.87
A (2) 3 3057.46a ± 939.99
9135.76a ± 3306.98
I. galbana T. suecica C. gracilis
63 (39) 57 (35) 43 (26)
B (1) 3 3210.30a ± 891.92
3407.67a ± 986.71
10643.97a ± 4088.20
10234.86a ± 3945.53
57.37± 20.59
B (2) 3 3371.00a ± 1045.7
9724.54a ± 3717.88
50 μm filtered seawater 2.0 (100) C 1 3257.15a
± 962.60 - 4867.44b ± 1535.37 - 13.53±
4.09
Table 1:Shell-length of full-sib M. galloprovincialis juveniles fed two composite microalgae diets (163 mg day-1; Isochrysis galbana, Tetraselmis suecica, Phaeodactylum tricornutum/Chaetoceros gracilis). The experiment lasted 116 days, from January 26th 2008 to May 29th 2008. Different superscripts in the same column indicate
significant differences among treatments for α = 0.001.
A. AGHZAR, M. MIÑAMBRES, P. ALVAREZ & P. PRESA
Growth of mussel juveniles fed mixed fresh microalgae diets was significantly enhanced regarding monospecific diets (Davis & Cambell 1998; Fotel et al., 1999; Puello-Cruz et al., 2009). For comparison of the shell-length obtained herein with mixed Diet-A (47 μm day-1 size increment and final size 8,781±3,545 μm) and Diet-B (57 μm day-1 increment and final size 10,235±3,946 μm ), the reported length increase of M. edulis juveniles under distinct experimental diets was 4.3 ± 0.4 μm day-1 in filtered sea water, 9.1 ± 0.2 μm day-1 in natural sea water, 13.1 ± 2.8 μm day-1 in enriched sea water (5x104 cells mL-1 Isochrysis galbana), and 5.6 ± 0.3 μm day-1 to 7.6 ± 1.3 μm day-1 in the wild (Fotel et al., 1999). Although the amount of energy in filtered seawater is assumed to reach the minimum required for maintaining the basal metabolic rate of mussels (Wong & Levinton, 2004), the two mixed diets assayed resulted in a weight increase that equalled fourfold the weight of the control Diet-C (14 μm day-1 and final 49% length increase). The high survival (around 99%) of juveniles fed multispecies enriched seawater was close to that observed in M. edulis larvae under different food ratios (Fotel et al., 1999). Therefore, in addition to the positive growth enhancement of mussel larvae, fresh multispecific formulated diets seem to assure a good survival of juveniles during retention times prior to raft culture in suspension devices.
Production costs
An optimal balance between production costs and growth increase during retention times is a key parameter
in juveniles-production efficiency. Regarding this, the microalgae assayed were collected at 5 days-old exponential stage cultures. At this time culture of C. gracilis averaged 3,000 cells/μl while P. tricornutum produced 15,000 cells/μl. Considering that 106 cells of C. gracilis weight 30 μg and the same amount of P. tricornutum cells weights 23 μg (Helm et al., 2004), production costs of the same amount of cells (43 mg day-
1) per species were ~4 times higher for C. gracilis than for P. tricornutum in the same culturing conditions (i.e., inoculum prize, Walne’s medium, etc.) (Table 1). Such higher production costs for C. gracilis (40.91 €) were due to the higher electrical consumption required to produce equal amounts of cells as for P. tricornutum (10.24 €). This cost would be reduced by 4 if retention time is reduced to one month (2.56 € and 10.23 €, respectively) and if large scale production of microalgae and culture medium are enforced. In this sense, a final approximate cost of 1 month retention time would be 1,892 €/raft for P. tricornutum and 7,551 €/raft for C. gracilis. Taking into consideration that the number of wild juveniles required to fill a raft cost about 2,000 € in the market (Table 2), it would result profitable the production of juveniles in hatchery using cheap-optimized multialgal diets. Several species of Chaetoceros are nowadays included in multialgal diets for molluscs due to their positive synergic role on growth (Puello-Cruz et al., 2009). Nevertheless, present data indicate that using C. gracilis in multispecies diets does not compensate its high production costs when compared to other cheaper and easy-culturing species such as P. tricornutum. Provided that acceptable growth
14 Thalassas, 29(1) · January 2013
P. tricornutum C. gracilis
Experimental ropea 1 growth ropeb 1 raftc Experimental rope a 1 growth ropeb 1 raftc
No. juveniles 3,400 11,550 2,310,000 3,400 11,550 2,310,000
Volume of algae (L) 102 346.5 69,300 403 1,369 273,803
Volume of Walne medium (mL) 102 600 120,000 403 2371 474,200
Cost of the Walne medium (€) 1.22 7.2 1,440 4.84 28.45 5,690
Electrical consumptiond (€) 9.02 30.6 6,128 36.08 122.57 24,513
Production cost (€) (116 days retention) 10.24 37.84 7,568 40.91 151.02 30,203
Production cost (€) 30 days retention 2.56 9.46 1,892 10.23 37.76 7,551
a Experimental diets were applied during 116 days of retention.
b One growth rope requires 10-12 kg of 5-15 mm juveniles. c One typical raft contains about 200 growth ropes. d Electrical consumption required to produce 43 mg day-1 of P. tricornutum or C. gracilis during 116 days, other costs being equal across species. Three fluorescent 36W
tubes were used for each of the three cultivation balloons.
Table 2:Estimated differential production costs of juveniles fed two experimental diets; common costs to both species are excluded, e.g. inoculum price, instrumentation or labour costs. The higher volume of C. gracilis required to attain an equivalent organic weight to P. tricornutum, finally rises variable production costs of the
former by ≈4 times.

A COST-BENEFIT ASSESSMENT OF TWO MULTI-SPECIES ALGAE DIETS FOR JUVENILES OF Mytilus galloprovincialis
and survival seem to be guaranteed during hatchery retention times, advances in juveniles culturing using performant live microalgae would come from reducing it cost-benefit production ratio.
ACKNOWLEDGEMENTS
This research was funded by Xunta de Galicia through grant program Ayudas para la Consolidación de Unidades Competitivas de Investigación INCITE07PXI310152ES. A. Aghzar has been supported by AECID (Ministerio Español de Asuntos Exteriores y Cooperación) through a research scholarship II-A from MAEC-AECID #0000215094 (2007-2008).
REFERENCES
Brown MR, Jeffrey SW, Garland CD (1989). Nutritional aspects of microalgae used in mariculture; a literature review, CSIRO Marine Laboratories Report, 205: 1-43.
Coeroli M, De Gaillande D, Landret JP (1984). Recent innovations in cultivation of molluscs in French Polynesia, Aquaculture, 39: 45-67.
Cordero B, Voltolina D (1994). Growth of Mytilus galloprovincialis fed with four microalgae and two feeding regimes, Journal of the World Aquaculture Society, 25(3): 471-476.
Coutteau P, Sorgeloos P (1992). The use of algal substitutes and the requirement for live algae in the hatchery and nursery rearing of bivalve molluscs: an international survey, Journal of Shellfish Research, 11(2): 467-476.
Cragg SM (2006). Development, physiology, behaviour and ecology of scallop larvae. In: SE Shumway, GJ Parsons, (eds.), Scallops: Biology, Ecology and Aquaculture, Elsevier B.V. Amsterdam, pp. 45-105.
Davis JP, Cambell CR (1998). The use of a Schizochytrium-based HUFA enriched dry feed for culturing juvenile mussels (Mytilus galloprovincialis) and the comparative routine costs of producing live algae in a commercial bivalve hatchery. In: H Grizel, P Kestemont (eds.), Aquaculture and water: fish culture, shellfish culture and water usage, Oostende Belgium European Aquaculture Society Spec. Publ. 26: 65-66.
De Pauw N, Persoone G (1988). Micro-algae for aquaculture. In: MA Borowitzka, LJ Borowitzka (eds.), Micro-algal biotechnology, New York, Cambridge U. Press, p 197-221.
Enright CT, Newkirk GF, Craigie JS, Castell JD (1986). Evaluation of phytoplankton as diets for juvenile Ostrea edulis L, Journal of Experimental Marine Biology and Ecology, 96: 1-13.
Epifanio CE, Valenti CC, Turk CL (1981). A comparison of Phaeodactylum tricornutum and Thalassiosira pseudonana as foods for the oyster, Crassostrea virginica, Aquaculture, 23: 347-353.
Foster-Smith RL (1975). Some mechanisms for the control of pumping activity in bivalves, Marine Behavior and Physiology, 4: 41-60.
Fotel FL, Jensen NJ, Wittrup L, Hansen BW (1999). In situ laboratory growth by a population of blue mussel larvae (Mytilus edulis L) from a Danish embayment, Knebel Vig, Journal of Experimental Marine Biology and Ecology, 233: 213-230.
Helm MM, Bourne N, Lovatelli A (2004). Hatchery culture of bivalves: A Practical Manual, FAO Fisheries Technical Paper 471, Rome.
Laing I, Millican PF (1986). Relative growth and efficiency of Ostrea edulis L. spat fed various algal diets, Aquaculture, 54: 245-262.
Lambade SB, Mohamed KS (2002). Laboratory - scale high density culture of the marine diatom Chaetoceros sp, Indian Journal of Fisheries, 49(1): 13-21.
Liu W, Pearce CM, Alabi AO, Gurney-Smith H (2009). Effects of microalgal diets on the growth and survival of larvae and post-larvae of the basket cockle, Clinocardium nuttallii, Aquaculture, 293: 248-254.
Miñambres M, Pérez M, Alvarez P, Presa P (2011). Cálculo de parámetros elementales para el cultivo de microalgas en criadero y su aplicación en el diseño de raciones alimentarias. In: Métodos y Técnicas de Investigación Marina, JM Estévez, C Olabarria, S Pérez, E Rolán, G Rosón (eds.), Chapter XIV: pp. 159-172.
Nevejan N, Davis J, Little K, Kiliona A (2007). Use of a formulated diet for mussel spat Mytilus galloprovincialis (Lamarck 1819) in a commercial hatchery, Journal of Shellfish Research, 26(2): 357-363.
Page H, Hubbard DM (1987). Temporal and spatial patterns of growth in mussels Mytilus edulis on an offshore platform: relationships to water temperature and food availability, Journal of Experimental Marine Biology and Ecology, 111: 159-179.
Paulay G, Boring L, Strathmann RR (1985). Food limited growth and development of larvae: experiments with natural sea water, Journal of Experimental Marine Biology and Ecology, 93: 1-10.
Pechenik JA, Eyster LS, Widdows J, Bayne BL (1990). The influence of food concentration and temperature on growth and morphological differentiation of blue mussel Mytilus edulis L. larvae, Journal of Experimental Marine Biology and Ecology, 136: 47-64.
Ponis E, Robert R, Parisi G (2003). Nutritional value of fresh and concentrated algal diets for larval and juvenile Pacific oysters (Crassostrea gigas), Aquaculture, 221: 491-505.
Puello-Cruz AC, Mezo-Villalobos S, González-Rodríguez B, Voltolina D (2009). Culture of the calanoid copepod Pseudodiaptomus euryhalinus (Johnson 1939) with different microalgal diets, Aquaculture, 290: 317-319.
Rico-Mora R (1987). Efecto interactivo de la temperatura y de la concentración de microalgas en la fisiología alimenticia y
15Thalassas, 29(1) · January 2013

A. AGHZAR, M. MIÑAMBRES, P. ALVAREZ & P. PRESA
16 Thalassas, 29(1) · January 2013
la energía potencial para el crecimiento de Modiolus capax (Conrard) (Bivalvia:Mytilidae). Thesis Dissertation, Centro de Investigación Científica y de Educación Superior de Ensenada, Departamento de Ecología Marina, Ensenada, México.
Rumrill SS (1990). Natural mortality of marine invertebrate larvae, Ophelia, 32: 163-198.
Sukenik A, Wahnon R (1991). Biochemical quality of marine unicellular algae with special emphasis on lipid composition I: Isochrysis galbana, Aquaculture, 97: 61-72.
Whyte JNC, Sherry K, Ginther N, Peribere G (2002). Effects of a Schizochytrium-based diet in the growth and nutritional condition of the mussel, Mytilus galloprovincialis. In: Aquaculture Canada 2002 Abstracts. http://www.aquacu lt u reassocia t ion .ca /ac02/abs t r ac t s /mussel .htm#Whyte.
Wong WH, Levinton JS (2004). Culture of the blue mussel Mytilus edulis (Linnaeus, 1758) fed both phytoplankton and zooplankton: a microcosm experiment, Aquaculture Research, 35: 965-969.
(Received: December 9, 2011; Accepted: April 30, 2012)