[6oi ionic regulation in the palaemonid prawn...
TRANSCRIPT

[6oi
IONIC REGULATION IN THE PALAEMONID PRAWNPALAEMON (=LEANDER) SERRATUS
BY G. PARRY*
Zoology Department, University of Cambridge
{Received 20 February 1954)
I. INTRODUCTIONIt has now come to be accepted that osmotic regulation of body fluids of inverte-brates is secondary to ionic regulation (Pantin, 1931), and a number of papersdealing with the relation between the two in Crustacea has appeared in the last twodecades (Krogh, 1939 for summary; Robertson, 1939, 1949, 1953; Webb, 1941).The greater part of this work has been concerned with animals whose naturalenvironment is sea water, and which can tolerate dilutions of this medium. Thestudy of osmoregulation of palaemonid prawns (Panikkar, 1941) has establishedthis group as one which is normally hypotonic to a sea-water environment—a character which is held in common with a number of other decapods, viz. Meta-penaeus (Panikkar & Viswanathan, 1948), Crangon (Broekema, 1942), grapsoidcrabs (Pearse, 1932; Schwabe, 1933; Baumberger & Olmsted, 1928; Jones, 1941;Edmonds, 1935; Conklin & Krogh, 1938) and ocypodid crabs (Pearse, 1932). Allof these animals show an osmotic independence of the medium, and although notall have been investigated for ionic regulation, it is to be expected from Pantin'sthesis that they will all show well-developed ionic regulation. The role played bythe antennal gland in maintaining this regulation has been studied in only a fewexamples, viz. Eriocheir (Scholles, 1933), Maia (Bialaszewicz, 1932), Cancer(Robertson, 1939), Carcinus (Nagel, 1934; Bethe, von Hoist & Huf, 1935; Webb,1941), Cambarus (Lienemann, 1938), Potamobius (Herrmann, 1931).
The palaemonid prawns are convenient for a study of this hypotonicity, sincedifferent species show varying degrees of osmotic independence. Panikkar (1941)investigated the osmotic behaviour of three species, but there appears to be nopublished data for the concentration of inorganic ions in the body fluids. Panikkarestablished the hypotonicity of the blood and urine of Palaemon ( = Leander)serratus (Pennant), P. elegans (Rathke) = Leander squiUa (L.) and Palaemonetesvarians (Leach), when living in sea water, and he also demonstrated their hyper-tonicity to the medium in brackish water of varying dilutions. Of the group,P. varians is the most adaptable to changes in the medium, Palaemon serratus theleast. Individual specimens of Palaemonetes varians can survive salinities of theorder 0-5 % NaCl without much change in the internal medium, and a completelyfresh-water variety of this species is common in some rivers of southern Europe(Boas, 1898).
• Now at Bedford College. University of London.JHB. 31, 4 40

602 G. PARRY
Osmotic behaviour of palaemonids
The hypotonicity of the blood to sea water when the animals are living in thatmedium was demonstrated by Panikkar (1941) by means of vapour-pressuremeasurements. Palaemonetes varians has a blood equivalent to 23 % NaCl(A 1-38° C) , Palaemon serratus 2-6-2-8% NaCl (A 1-56-1-68° C.) and P. elegans2-6 % NaCl (A 1-56° C.) (calculated from the relationship A/o-6 = % NaCl, Ramsay,1949). Palaemonetes varians is almost completely homoiosmotic at about 2 % NaCland varies only by about o-8— i-o % NaCl for external salinities from o to 5 %. Thusit can tolerate, with small internal changes, a range of salinity from water which isnearly fresh to 5 2 % NaCl. This behaviour is less well developed in Palaemonserratus, which has a much more limited tolerance to varying salinity in the environ-ment. When the environment is diluted to 2-5 % NaCl (which is about isotonicwith the blood) the concentration of the blood is maintained; but in greater dilutionsthere is a steady fall in blood concentration. P. elegans is very similar to P. serratus,but the homoiosmotic behaviour is a little better developed, so that it falls into anintermediate position between the other two species. When any one of theseprawns is transferred to very dilute media, the blood osmotic pressure falls for14-24 hr., but after this there is a slight compensating rise, and a steady state isthereafter maintained.
The urine of all three species is nearly isotonic with the blood in all conditionsof external salinity. Different parts of the excretory system do not seem to haveany salt- or water-regulating mechanism, as the osmotic pressure of fluids fromdifferent parts of the gland in P. serratus do not show any significant differences.During the moult there is a slight rise in the blood osmotic pressure; this isfollowed by the absorption of water and a return of the blood osmotic pressure toa normal level a few days later.
In an attempt to elucidate further the regulation of the blood in prawns, chemicalanalyses were made of the blood and urine of P. serratus. This species was chosenbecause it is larger than the others available and yielded quantities of fluid whichit was possible to analyse by ordinary semi-microchemical methods. This speciesis the least homoiosmotic of the four which are easily available in Britain, so thatany regulation of inorganic ions apparent in this species may be expected in greaterdegree in the others.
II. METHODSBlood was collected from P. serratus by inserting a glass cannula into the dorsalside of the abdomen, just in front of the telson, and applying gentle suction. Inthis way blood was collected from the extensive sinuses of the abdomen. Ifinsufficient blood was collected, a second cannula was inserted into the heart orpericardium from the back of the thorax and the blood gently sucked out. A ratherlarge specimen would provide 0-2-0-3 ml. blood, but samples as small as 002 ml.could be used for some analyses. Urine samples were collected by a glass cannulafrom the 'nephroperitoneal' or 'epigastric' sac (a dorsal backwardly projectingextension of the bladder), or from the excretory opening of the gland. To collect

Ionic regulation in the palaemonid pravm 603
Irom the sac, a small cut was made in the ridge of the rostrum and the pointedcannula pushed into the sac from an antero-dorsal position; to collect from theexcretory pores, the animal was laid on its back beneath a binocular microscope,a sharply tapered cannula fitted into one of the openings of the gland, and the urinedrawn up. The former procedure was less satisfactory, since samples could occasion-ally be contaminated with blood from one of the anterior arteries. For this reason,most of the later collections were made from the excretory pores. Volumes of01-001 ml. were obtained.
The volume of samples, when necessary for the calculation of ion concentrations,was measured by weighing an identical quantity of distilled water in the collectingcannula. The results are thus expressed primarily as mg./ml. blood. Samples couldnot be weighed directly as the blood coagulates very rapidly. The concentrationscan be expressed in other terms of concentration by using the experimentallydetermined estimates of water content and specific gravity.
The blood and urine samples were taken from both males and females, and wereanalysed without removal of corpuscles. Both these procedures will add to thevariability of the results, since Panikkar (1941) found a difference of o-i % NaClconcentration in the blood of ovigerous females compared with males. The removalof corpuscles by centrifugation was found to be impracticable with such smallsamples, since manipulation would reduce their volume still further and so leadto increased errors in the analyses used. It was hoped that the concentrationdifferences apparent after sojourn in different salinities would be sufficiently largeto make these variations relatively insignificant.
The procedures for chemical analysis were based on those of Robertson & Webb(1939), since these are suited to the samples available. There were some modifica-tions introduced for the rather smaller quantities of fluid analysed. These methodshave been reviewed in a previous paper (Parry, 1953). Chloride determinations inthis investigation were all done by Conway's microdiffusion technique (1950). Inalmost all the analyses samples of standard sea water (Copenhagen) were treatedin exactly the same way as the blood or urine samples; the final calculations ofthe ion concentration could thus be made by a comparison of titration volumes,precipitate weights, etc. In this way almost all personal errors due to slightlydifferent temperatures, or times for reaction, are minimized. In many cases thevolumes of samples were also determined and could be used as a check of theaccuracy of the method. The results were calculated by comparison with the ionicconcentrations in sea water given by Sverdrup, Johnson & Fleming (1942, p. 173).
III. EXPERIMENTAL RESULTSComposition of the blood of animals living in sea water
The concentrations of inorganic ions found in the blood of animals living in theirnormal sea-water environment (sea water from Plymouth) is shown in Table 1.The results are expressed here as mg./ml. blood, since the volume of the sampleswas measured at the time of the analysis. These values have been converted to-
40-2

604 G.PARRY
m.equiv./l.; they may also be converted to m.equiv./kg. water from the estimateof the water content of the blood.
The water content of the blood was determined by drying weighed samples atno° C. to a constant weight, and gave an estimated mean of 877 mg./ml. blood(nine observations). The original values for ion concentration as mg./ml. blood canbe expressed in terms of molality by using this figure.
The specific gravity of the blood was determined by weighing a sample of bloodin its collecting tube, and then weighing an identical volume of distilled water inthe washed tube. The mean of nine such determinations was 1-030.
Table 1. Concentration of some inorganic ions in the blood ofPalaemon serratus living in sea water
Ion
SodiumPotassiumCalciumMagnesiumChlorideSulphate
mg./ml. blood
9-09 ± 0 2 6 (15}0-290 ±0-021 (8)O'50O±OpOI2 (9)O'307±o-os8 (13)
15-20 ±0-13 (20)o-948±o-i73 (13)
m.equiv./l.
3947-7
25225-2
43O5-2
mg./ml. blood o/
mg./ml. sea water '"
7970
" 522
749
Bracketed figures are the number of determinations.
The total mg. ions/kg, water analysed in blood is about 5 % less than thatcalculated from Panikkar's (1941) measurements of the osmotic pressure:
By analysis, 856 mg. ions/1, blood = 876 mg. ions/kg, water.From O.P. (Ai*56°C.), 914 mg. ions/1. blood= 948 mg. ions/kg, water.
This may indicate that either the estimate of A is too high, or that the analyses givea result which is about 5 % too low. It seems possible that the estimate of anions issome 20 m.equiv. too low, as there are only 436 m.equiv. anions/1. to balance452 cations. If the anions totalled about 450, the total mg. ions/1, would be about900. (This difference of 5 % is well within the error of determinations of the con-centration of individual ions. Panikkar (1941), while he gives the range of hisvapour-pressure determinations, does not give any standard errors of the means.)A determination of the total base in blood (by the method of Hald, 1933, 1934)gave a value of 446 m.equiv./l. (nine observations). This compares fairly well withthe sum of the cations for the individual analyses (452 m.equiv.), indicating that itis the figure for the sum of the anions which is too low. The deficit must be entirelydue to a low figure for chloride, since sulphate is relatively much less important inthis numerical sense. The chloride analyses in question were mostly done duringJune, July and August when Panikkar (1941) records that the osmotic pressure islower than in winter.
The concentration of proteins in the blood has not been measured directly, but
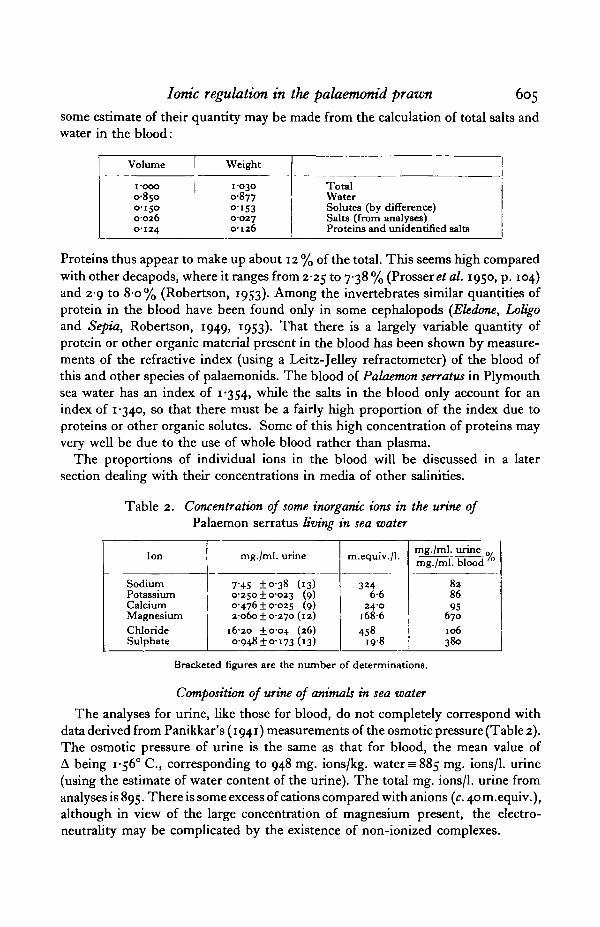
Ionic regulation in the palaemonid prawn 605
some estimate of their quantity may be made from the calculation of total salts andwater in the blood:
Volume
i-ooo0-8500-1500-0260-124
Weight
1-0300-877O-IS30-0270-126
TotalWaterSolutes (by difference)Salts (from analyses)Proteins and unidentified salts
Proteins thus appear to make up about 12 % of the total. This seems high comparedwith other decapods, where it ranges from 2-25 to 7-38 % (Prosser et al. 1950, p. 104)and 2-9 to 8-o% (Robertson, 1953). Among the invertebrates similar quantities ofprotein in the blood have been found only in some cephalopods (Eledone, Loligoand Sepia, Robertson, 1949, 1953). That there is a largely variable quantity ofprotein or other organic material present in the blood has been shown by measure-ments of the refractive index (using a Leitz-Jelley refractometer) of the blood ofthis and other species of palaemonids. The blood of Palaemon serratus in Plymouthsea water has an index of 1-354, while the salts in the blood only account for anindex of 1-340, so that there must be a fairly high proportion of the index due toproteins or other organic solutes. Some of this high concentration of proteins mayvery well be due to the use of whole blood rather than plasma.
The proportions of individual ions in the blood will be discussed in a latersection dealing with their concentrations in media of other salinities.
Table 2. Concentration of some inorganic ions in the urine ofPalaemon serratus living in sea water
Ion
SodiumPotassiumCalciumMagnesium
ChlorideSulphate
mg./ml. urine
7-45 ±0-38 (13)0-250 ±0-023 (9)0476 ±0025 (9)2-o6o± 0-270 (12)
16-20 ±0-04 (26)0-94810-173(13)
m.equiv./l-
3246-6
24-01686458
19-8
mg./ml. urine 0 /
mg./mf. blood / o
828695670
106380
Bracketed figures are the number of determinations.
Composition of urine of animals in sea water
The analyses for urine, like those for blood, do not completely correspond withdata derived from Panikkar's (1941) measurements of the osmotic pressure (Table 2).The osmotic pressure of urine is the same as that for blood, the mean value ofA being 1-56° C, corresponding to 948 mg. ions/kg, water = 885 mg. ions/1, urine(using the estimate of water content of the urine). The total mg. ions/1, urine fromanalyses is 895. There is some excess of cations compared with anions (c. 4om.equiv.),although in view of the large concentration of magnesium present, the electro-neutrality may be complicated by the existence of non-ionized complexes.

606 G. PARRY
The water content of the urine (determined as for blood) is 933 mg. water/ml,blood (nine determinations).
The specific gravity, determined in the way described for blood, is 0-977 (ninedeterminations). This appears surprisingly low. Determinations of the specificgravity of the urine were variable, but consistently less than unity. Nine sampleswere used. If the ammonium ion is present as an excretory product, this couldaccount for a reduction in the specific gravity, if suitably large quantities werepresent. Of the total constituents of urine, there are 912 mg./g. water, whichleaves 65 mg. for all solutes. Analysis of the inorganic ions, sodium, potassium,calcium, magnesium, chloride and sulphate, account for 27 mg. of this, so thatabout 38 mg. remain for other ions and excretory products. If all this 38 mg. wereattributed to the ammonium ion, there would be a concentration of c. 4 % ammonia.With this quantity of ammonia present in pure water, the specific gravity of sucha solution would be 0-983. The concentration of ammonia is likely to be a variablequantity depending upon the amount of food eaten and the rate of urine flow.
The concentrations of individual ions will be discussed below.
Composition of blood and urine of animals living in hypo- andhypertonic media
Animals were kept in the laboratory in certain dilutions and concentrations ofsea water in order to accentuate the differences between the internal and externalenvironments. The media chosen were 50 and 120% of Plymouth sea water. Thefirst was made simply by diluting Plymouth sea water with the same quantity ofdistilled water; the second by concentrating Plymouth sea water to twice its usualconcentration, correcting this as to pH by adding drops of sodium bicarbonate,and diluting with the appropriate quantity of ordinary sea water. In this way thebalance of ions should have been fairly similar to that in normal sea water, althoughsome calcium appears to come out of solution with the initial evaporation. Thissediment was kept in contact with the solution, and a large part of it appeared todissolve again when the final medium of 120% sea water was prepared. In boththese media the animals were able to survive indefinitely at 18-200 C. Experimentalanimals were first acclimatized for about 1 week in an intermediate salinity, andthen left for at least a further week in the final salinity.
The analyses were made in exactly the same way as for animals living in ordinarysea water. The results are shown in Table 3 for blood and Table 4 for urine. Thevalues for blood and urine of animals in sea water are added for comparison.
The correspondence of the observed mg. ions/1, blood and urine and the valuesof A for blood and urine in all three salinities is shown in Table 5. Although thecalculated and observed mg. ions/kg, water are not identical, the discrepancies arenot unduly large in relation to the general variability of the individual analyses,except in the case of urine of animals living in 120% sea water. In this medium theurine contains such large quantities of magnesium and sulphate that the activity ofthe inorganic salts in solution may well be reduced, leading to a consequent
Iortic regulation in the palaemonid prawn 607
reduction in A. (The freezing-point depression of a magnesium sulphate solutionis only 58-5 % of an isomolar sodium chloride solution.)*
The control of individual ions
The relative proportions of ions in the blood and in the medium in which theanimals are kept indicate the conservation, excretion or exclusion of those ions.
Table 3. Comparison of the composition of the blood of Palaemon serratus living in50, 100 and 120% sea water
(Concentrations expressed as m.equiv./l.)
Medium as . „ ,.% sea water | Sodium
50 ' 257±i8100 1 394±i2120 473 ± 8
Potassium
6-7 ±0-977±o-6
IO-2±I-3
Calcium
28-3 ±1-425-2 ±o-6299 ± 1 8
Magnesium
22-2 ± 3 825-2 ±4-84i-2±3-4
Chloride
3°°±i743° ± 4494 ±39
Sulphate
2-3±o-85 - 2 ± I O6-4 ± 1 - 2
Table 4. Comparison of the composition of the urine of Palaemon serratus living in50, 100 and 120% sea water
(Concentrations expressed as m.equiv./l.)
Medium as \ „% sea water j sod""™
50 206 ±18100 • 324116120 , 432 + 11
Potassium
7-4 ±0-96-6 ±o-68-5 ± i - 5
Calcium
25-o±i-324-0 ± 1-3293 ± i - 8
Magnesium | Chloride
67-1 ±12-6 ' 3 3 O ± I - II 6 8 - 6 ± 2 2 - I ' 45812-5184-5 ±22-2 1 622 ±2-5
Sulphate
3-6±i-oi9-8±3-6320 ±9-3
Table 5. Calculated and observed values of mg. ions/kg. water in bloodand urine of Palaemon serratus
Mediumas %
sea water
5O
1 0 0
1 2 0
Sample
BloodUrineBloodUrineBloodUrine
A°C.(Panikkar)
1-03°3•S6
i-S6[•7»t[•72
mg. ions/kg,(from
A°C.)«
6*3623944944
10151015
mg. ions/1,observed
59°-1591-3855-59189
1016-01185-4
mg. ions/kg,(from
sp.gr.)t
608580882901
10461160
Differencea s %
2 4696-64'53 0
1 4 3
• Calculated from data in Sverdrup et at. (1942, p. 67).t Calculated from observed mg. ions/1, sample, assuming that specific gravity of blood is 1-03 and
specific gravity of urine is 0-98.% By extrapolation of Panikkar's graph (1941).
Since the analyses in this investigation were made on whole blood and not ona dialysed plasma, a part of the differences observed in ion concentration can beattributed to the contents of the corpuscles, a part to the effects of a Donnanequilibrium, and a part to a binding of ions to the proteins present in the plasma.This last factor is likely to be of importance only with regard to the level of calciumin the blood. The differences in ion concentration in Palaemon serratus are for the
* I am indebted to Mr P. C. Croghan for this information (personal communication).

608 G. PARRY
most part considerably greater than one would expect from the Donnan equilibrium,even when the large standard error of the figures is taken into account. It wouldappear that the ionic differences between the blood of the animal and the mediumare significant ones, and that they must be maintained by the animal in some waywhich cannot be attributed merely to the presence of proteins in the blood.
In animals living in Plymouth sea water, the sodium, potassium and chloride ofthe blood are all about 70-80% of their external concentrations. This might beexpected for sodium and chloride, since the total blood concentration is about 80 %of the medium (A blood = 1-56° C ; A sea water = 1-98° C). In many marineinvertebrates which have been examined (Robertson, 1953; Prosser et al. 1950) theblood potassium is higher than in the medium; but this does not seem to be thecase in P. serratus. The figure for potassium concentration has a high standarderror (c. 7 %), but even when this is considered, the potassium concentration iscomparatively low.
Of the other ions calcium shows some evidence of conservation, although in theabsence of any data about the binding of this ion to proteins in the blood it isdifficult to estimate the level of ionic calcium. A calcium content of 115-120% ofthe medium could be maintained by the effect of the Donnan equilibrium andcalcium-protein complexes (Robertson, 1949).
In sea water the concentration of magnesium and sulphate in the blood isremarkably low. The analyses of urine from animals in this medium show that theconcentration of these two ions is very much greater than it is in the blood, whilethat of other ions is not so markedly different.
The relative concentrations of ions in the medium, blood and urine in all threeexperimental media are shown in Fig. 1. The ion concentration of the medium iscalculated from data in Sverdrup et al. (1942, p. 173).
Sodium is at a constantly lower level in the blood than in the medium, except in50% sea water, where it is very similar in concentration. While the osmoticpressures of blood and medium are similar in 80% sea water, the sodium con-centrations are similar at a greater dilution of the medium. The sodium concentra-tion of the urine is consistently less than that of the blood and maintains about thesame proportion except at the highest salinity where the curve becomes a littlesteeper, and the concentrations in blood and urine approximate more closely.Perhaps this might be interpreted as an attempt to excrete a larger quantity ofsodium in response to the increased salinity of the environment, although themagnitude of the change is not very great. The urine, although it maintains a con-stantly lower level of sodium than the blood, can hardly be considered as the mainsource of the control of sodium shown by the animal.
The chloride concentration in the three fluids shows a similar pattern. In theblood, chloride is constantly lower than in the medium, except at the lowestsalinity where it approximates to that of the medium. In contrast to sodium,however, the urine concentration is consistently higher than that of the blood,although this difference is scarcely sufficient to account for the control of chloride,even in the highest salinity, where the difference between blood and urine con-

Ionic regulation in the palaemonid prawn 609
centrations is at its greatest. It appears from the graph that there is a markedincrease in the excretion of chloride in 120% sea water.
The concentrations of potassium and calcium in the blood and urine showrelatively only small changes with changes in the external salinity. The standarderror is large in relation to the observed differences, so that the changes cannot be
Potassium
600
300
100 100
50 100Medium (% sea water)
120 50 100Medium <% tea water)
120
Fig. 1. Relative concentrations of ions in sea water ( ), blood (©—©) and urine (A—A)-Abscissae: concentration of medium as a percentage of Plymouth sea water. Ordinates: con-centration of ions expressed as m.equiv./l.
regarded as very significant. For potassium, the blood concentration is lower thanthat of the medium, except in 50 % sea water, when it is rather higher in the blood,the isotonic point being at about 60% sea water. It is kept relatively constantbetween 50 and 100% sea water but tends to rise a little in 120% sea water. Theurine concentration follows a similar curve, which for the most part lies below thatfor blood. In general, the blood and urine concentrations are very similar, so thatthe antennal gland cannot be very important in regulating potassium.

610 G. P A R R Y
Calcium is also relatively constant in blood and urine in spite of changes in theexternal concentration. The slightly lower level in the urine is scarcely significantin relation to the standard errors and is certainly not sufficient to account for theregulation of the ion.
The concentration of magnesium and sulphate in blood and urine with differentconcentrations of the medium are maintained always at very different levels.Magnesium in the blood is remarkably low and relatively constant in all threesalinities. It is obvious from the high urine concentrations that this efficientregulation of the blood magnesium could be brought about by its excretion in theurine. The urine concentration rises steeply as that of the outside medium rises,although less steeply as 120% sea water is approached. In the same conditions theblood concentration rises a little as though the maximum possible rate of excretionof the ion was being approached. Sulphate in the blood is similarly very low. In50 % sea water the urine concentration is also low but rises steeply as the externalconcentration increases. The low excretion in 50% sea water where the blood levelis still much below that outside, may indicate that sulphate is excluded from thebody fluids in low external concentrations. This exclusion appears inadequate athigher concentrations, so that the antennal gland plays an increasingly importantrole.
IV. DISCUSSIONThe principal ions of Palaemon serratus blood are much less concentrated than inother decapod crustaceans for which data are available. Following the lower osmoticpressure of the blood of animals in sea water, the concentrations of sodium,potassium and chloride are all 70-80 % of the outside concentration. Calcium, onthe other hand, is maintained at a level slightly higher than that of the medium,perhaps indicating the degree of binding to proteins in the blood. Conversely,magnesium and sulphate are proportionately much lower than in other marinedecapods. This could be an adaptation to an estuarine invasion, especially in thecase of sulphate, since an efficient means of regulating this ion is of relativelygreater importance in waters which may possibly contain a greater proportion thansea water (Howes, 1937). The low value of the ratio of magnesium to other ions inthe blood places this prawn with the most active crustaceans (Robertson, 1953)-Slow-moving forms such as spider crabs have a ratio (Na + K/Ca + Mg) about80-100% of that of sea water; most active forms have a value less than 50% thatof sea water. P. serratus may be compared in this respect with Homarus andNephrops, with which it has indeed many similarities of behaviour and activity.
Without some indication of the quantity of urine produced in different circum-stances it is not possible to deduce completely the function of the antennal gland.The volume of urine produced in different salinities has been measured in a relatedpalaemonid species and will be the subject of a further paper. While the inorganicanalyses of urine do not necessarily demonstrate the importance of excretion inmaintaining the osmotic control of the animal, they do indicate how it maintainsionic control, and make possible some deductions as to the mechanism of urineproduction.

Ionic regulation in the palaemonid prawn 611
The analyses of sodium, chloride, potassium and calcium in urine are, on thewhole, very similar to those of blood, and their concentrations would not beincompatible with the conception of the urine as an ultrafiltrate of the blood. Themuch higher concentrations of magnesium and sulphate in urine could result fromsome active excretion of these ions after the formation of an ultrafiltrate. The lowerconcentration of sodium, and the higher concentration of chloride in the final fluidproduced, might result from a necessity to balance the fluid ionically after theinflux of magnesium and sulphate (and the ammonia which is presumed to bepresent). The comparatively constant proportion of sodium and chloride in bloodand urine, independent of the external salinity, suggests that the ultrafiltration isunaffected (except perhaps in its rate) by the external conditions or the internalconditions resulting therefrom.
By analogy with the vertebrate glomerulus we can imagine that the ultrafiltrateis produced in the end-sac of the antennal gland, and that the excretion of additionalmagnesium, sulphate and perhaps ammonia into this ultrafiltrate takes place alongthe length of the nephridial canal, together with the reabsorption or secretion ofsmall quantities of other ions in order to preserve electro-neutrality. The antennalglands of palaemonid prawns lack any segment such as the tubule of fresh-watercrayfishes which might perform a salt-absorbing or water-regulating function, andthere is certainly no experimental evidence that this takes place. The bladder andepigastric sac do not appear to play any part in the chemical elaboration of the urine.The histology of the gland (Grobben, 1880; Weldon, 1889; Marchal, 1892; Allen,1892; Cu6not, 1895; Patwardhan, 1937) and its blood supply are in accord withthis conception of the function of the antennal gland.
Some part of the ionic regulation of the animal can thus be said to be performedby the antennal glands, but the analyses do not tell us how the osmotic regulationis brought about. Presumably this must be effected extra-renally.
V. SUMMARY
1. Analyses have been made of the blood and urine of Palaemon serraius for theinorganic ions Na, K, Ca, Mg, Cl, SO4, the animals being kept in 50, 100 and 120%sea water.
2. When the animal is in 100 % sea water the concentrations of ions in the blood,expressed as percentages of their concentrations in the medium (to the nearest 5 %)are as follows: Na, K and Cl, 85%; Ca, 105%; Mg, 20%; SO4, 10%.
3. When the animal is in 50% sea water the corresponding figures are: Na andCl, 105%; K, 120%; Ca, 200%; Mg, 20%; SO4, 10%.
4. When the animal is in 120% sea water the corresponding figures are: Na,K and Cl, 85%; Ca, 115%; Mg, 30%; SO4, 20%.
5. The concentrations of Na, K and Ca in the urine are always slightly (^ 20 %)less than their concentrations in the blood. The concentration of Cl is slightlygreater in the urine than in the blood (10-20 %) and the concentrations of Mg andSO4 are very much greater, by factors of up to 7 times. The relative concentrations

612 G. PARRY
of ions in blood and urine do not change substantially with changes in the externalmedium.
6. The antennal gland, although it plays no part in purely osmotic regulation,is no doubt partly responsible for maintaining the low blood concentrations ofMg and SO4.
I should like to thank Prof. Sir James Gray and Prof. H. Munro Fox for thehospitality of their departments during the course of this work. My acknowledge-ments are also due to Dr J. A. Ramsay and Dr J. D. Robertson for advice andcriticism, and to Prof. H. Munro Fox for reading the manuscript. For a part ofthe time engaged on this research I was in receipt of a Science Scholarship fromthe University of New Zealand, and latterly of a research grant from the ZoologyDepartment of Cambridge, for both of which I should like to record my appreciation.
REFERENCESALLEN, E. J. (1892). Nephridia and body-cavity of some decapod Crustacea. Quart. J. Micr. Sci.
34.75-BAUMBERGER, J. P. & OLMSTED, J. M. D. (1928). Changes in the osmotic pressure and water content
of crabs during the moult cycle. Phytiol. Z06I. 1, 531.BETHE, A., VON HOLST, E. & HUF, E. (1935). Die Bedeutung des mechanischen Innendrucka filr die
Anpassung gepanzerter Seetiere an Aenderungen des osmotischen Assendruclcs. Pflug- Arch.ges. Phytiol. 335, 330.
BiALASZEWicz, K. (1932). Sur la regulation de la composition minerale de rhemolymphe chez lecrabe. Arch. int. Phytiol. 35, 98.
BOAS, J. E. V. (1898). Kleinere carcinologische Mitteilungen (a). Zool. Jb. Abt. 1, 4, 793.BROEKEMA, M. M. M. (1942). Seasonal movements and the osmotic behaviour of the shrimp,
Crangon crangon. Arch. nierL Zool. 6.CONKLIN, E. G. & KKOOH, A. (1938). Osmotic behaviour of Briocheir in concentrated sea water.
Z. vergl. Phytiol. 36, 239.CONWAY, E. J. (1950). Microdiffusion Analytit and Volumetric Error. London: Crosby Lockwood.CUBNOT, L. (1895). Etudes physiologiques sur les Crustac^es Decapodes. Arch. Biol., Parit, 13, 245.EDMONDS, E. (1935). The relations between the internal fluid of marine invertebrates and the water
of the environment, with special reference to the Australian Crustacea. Proc. Litm. Soc. N.S.W.60, 233.
GROBBEN, C. (1880). Die Antennendruse der Crustaceen. Arb. zool. Inst. Wien, 3, 93.HALD, P. M. (1933). The determination of the bases of serum and whole blood. J. Biol. Chem.
103, 471-HALD, P. M. (1934). A note on the gravimetric determination of total base in serum and blood.
J. Biol. Chem. 105, 675.HERRMANN, F. (1931). Ueber den Wasserhaushalt des Flusskrebses. Z. vergl. Phytiol. 14, 479.HOWES, N. H. (1937). The ecology of a saline lagoon in South-East Essex. J. Linn. Soc. Land.
40, 383.JONES, L. L. (1941). Osmotic pressure relations of nine species of crabs of the Pacific coasts of North
America. J. Cell. Comp. Physiol. 18, 79.KROGH, A. (1939). Osmotic Regulation in Aquatic Animals. Cambridge University Press.LIENEMANN, L. J. (1938). The green glands as a mechanism for osmotic and ionic regulation in the
crayfish (Cambarus). J. Cell. Comp. Physiol. n , 149.MAHCHAL, P. (1892). R^cherches anatomiques et physiologiques sur l'appareil excreteur des
crustaceea Decapodes. Arch. Zool. exp. gen. 10, ser. 2, 75.NAGEL, H. (1934). Die Aufgaben der Excretionesorgane und der Kienen bei des Osmoregulation
von Cardnut maenas. Z. vergl. Phytiol. a i , 468.PANIKKAR, N. K. (1941). Osmoregulation in some palaemonid prawns. J. Mar. Biol. Ass. U.K.
25, 317-PANIKKAR, N. K. & VISWANATHAN, R. (1948). Active regulation in Metapenaeus monoceros. Nature,
Lond,, 161, 138.

Ionic regulation in the palaemonid prazim 613PANTIN, C. F. A. (193 I) . The origin of the composition of the body fluids in animals. Biol. Rev.
6, 459-PARRY, G. (1953). Osmotic and ionic regulation in the isopod crustacean, Ligia oceamca. J. Exp.
Biol. 30, 567.PATWARDHAN, S. S. (1937). Palaemon. Indian Zool. Mem. 6, 1.PEARSE, A. S. (1932). Freezing-points of blood of certain littoral and estuarine animals. Pap.
Tortugas Lab. 28, 93.PROSSER, C. L. et al. (1950). Comparative Animal Physiology. Philadelphia: W. B. Saunders and Co.RAMSAY, J. A. (1949). A new method of freezing-point determination for small quantities. J. Exp.
Biol. lib, 57.ROBERTSON, J. D. (1939). The inorganic composition of body fluids of three marine invertebrates.
J. Exp. Biol. 16, 387.ROBERTSON, J. D. (1949). Ionic regulation in some marine invertebrates. J. Exp. Biol. 16, 387.ROBERTSON, J. D. (1953). Further studies on the ionic regulation in marine invertebrates. J. Exp.
Biol. 30, 277.ROBERTSON, J. D. & WEBB, D. A. (1939). Microestimation of sodium, potassium, calcium, magnesium,
chloride and sulphate in sea water and the body fluids of marine animals. J. Exp. Biol. 16, 155.SCHOIXES, W. (1933). Ueber die Mineralregulation wasserlebender Evertebraten. Z. vergl. Physiol.
19. 522.SCHWABE, E. (1933). Uber die Osmoregulation verschliedner Krebse. Z. vergl. Physiol. 19, 183.SVERDRUP, H. V., JOHNSON, M. W. & FLEMING, R. H. (1942). The Oceans. New York: Prentice-Hall.WEBB, D. A. (1941). Ionic regulation in Carcinus maenas. Proc. Roy. Soc. B, 129, 107.WELDON, W. E. R. (1889). The coelom and nephridia of P. serratus. J. Mar. Biol. Ass. U.K. 1, 162.